a second transcriptionally active dna-binding site for the wilms
TRANSCRIPT

Proc. Natl. Acad. Sci. USAVol. 90, pp. 88%-8900, October 1993Medical Sciences
A second transcriptionally active DNA-binding site for the Wilmstumor gene product, WT1
(zinc-finger protein/suppressor/gene regulation)
ZHAO-YI WANG, QING-QING Qiu, KEVIN T. ENGER, AND THOMAS F. DEUEL*Departments of Medicine and Biochemistry and Molecular Biophysics, Washington University School of Medicine and The Jewish Hospital of St. Louis, 216South Kingshighway, St. Louis, MO 63110
Communicated by David M. Kipnis, June 28, 1993
ABSTRACT The putative Wilms tumor suppressor gene,wtl, encodes a zinc-finger protein that binds to the DNAsequence 5'-GCGGGGGCG-3'. We previously reported thatWT1 has separable domains that function either to activate orsuppress tanscription. We now have identified a second WT1binding sequence (5'-TCCTCCTCCTCCTCTCC-3') 3' to thetranscription initiation site of the platelet-derived growth fac-tor A-chain gene by DNase I footprinting and gel mobility shiftassays. WT1 requires both 5' and 3' binding sites for tran-scriptional suppression; however, WT1 functions as a tran-scriptional activator when it binds to either the 5' or 3' sitealone. This second WT1 binding sequence functions equaflywell as the previously identified 5'-GCGGGGGCG-3' sequencewhen analyzed in transient transfection assays. A core DNAsequence recognized by WT1 was defined by using relatedsynthetic oligonucleotides. We also identified sequences similarto the WT1 binding site within the promoter regions of fiveother growth-related genes and demonstrated that each ofthesesequences also binds WT1 in gel mobility shift assays. Theseresults thus identify a second WT1 binding site and suggest thatadditional growth-related genes may be tran riptionafly in-fluenced by WT1.
Wilms tumor (WT) is an embryonal cell malignancy of thekidney resulting from the continued proliferation of embry-onic kidney blastemal cells. By comparing overlapping chro-mosomal deletions in different WT cells, a potential tumorsuppressor gene, wtl, has been identified and cloned (1, 2).wtl encodes a protein that contains four zinc-finger domainsof the cysteine-histidine type that bind to the DNA sequence5'-GCGGGGGCG-3' (3), a recognition element common tothe early growth response (EGR) family of zinc-finger pro-teins (4). The N-terminal domain ofWT1 is rich in proline andglutamine residues and appears to be essential for the func-tion of WT1 on transcriptional regulation (5, 6). Recently,several growth-related genes, including EGR-1, insulin-likegrowth factor II, and the platelet-derived growth factor(PDGF) A-chain gene, have been shown to be transcription-ally suppressed by WT1 in cotransfection assays (5, 7, 8),supporting the idea that the putative tumor suppressor ac-tivity ofWT1 may function through suppression of transcrip-tion of positive regulators of cell growth. However, recentlywe made the observation that WT1 also activates transcrip-tion and that separable domains within WT1 are required forsuppression and activation, respectively (6), suggesting thatWT1 has both positive and negative transcriptional regula-tory potential in vivo. More recently, an activation activity ofWT1 also was observed by Maheswaran et al. (9), who alsofound that WT1 physically and functionally interacts withanother tumor suppressor, p53.
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.
The potential of WT1 to suppress transcription of thePDGF A-chain gene may be ofbiological importance becausethe PDGF A-chain gene encodes a potent mitogen andchemoattractant (10) that is highly expressed in severaltransformed cell lines and in some WTs (10-12). To under-stand mechanisms by which the PDGF A-chain gene isupregulated in these tumor cell lines, we identified andanalyzed the promoter region of the PDGF A-chain gene anddemonstrated that a highly G+C-rich region proximal to theTATA box is required for its optimum transcription (13-15).We (8) also reported that this region interacts with WT1 andconfers the striking suppression of PDGF A-chain genepromoter activity by WT1. Recently, we observed that aregion 3' to the transcription initiation site is also required formaximum suppression of PDGF A-chain promoter activityby WT1 (6), suggesting that another WT1 binding site mayexist 3' to the initiation site of transcription.Here we identify and characterize a second functional WT1
binding sequence. We also have identified similar sequenceswithin the promoters of five other growth-related genes thatbind WT1. The results thus establish another functional WT1binding site and identify a number of growth-related genesthat may serve as targets for WT1.
MATERIALS AND METHODSPlamid Construction. Expression vectors containing the
full-length protein coding regions or the zinc-finger domain ofthe human wtl genes, the cytomegalovirus (CMV) immediate-early promoter, and the simian virus 40 polyadenylylationsignal were constructed as described (5). The chimeric PDGFA-chain minimal promoter plasmids were constructed as fol-lows. An oligonucleotide was synthesized according to theDNA sequence of the PDGF A-chain promoter (-60 to +3)and cloned into the HindIII and Xba I sites ofthe pCAT-Basicvector (Promega); the resulting vector was designated aspACAT. Oligonucleotides containing the three typical EGR-1binding sites (5'-CGCCCCCGCCCCCGCGCCGGCGGCGC-CCCCGCCCCCGCGCCA-3') and three contiguous "TCC"motifs (see Results) (5'-TCCTCCTCCTCCTCTCCTCCT-CCTCCTCTCCTCCTCCTCCTCTCC-3') with HindIlI orXba I sites at both ends were also synthesized and cloned intothe HindIII site upstream of the transcription start site, down-stream in the Xba I site ofpACAT, or into both the upstreamand downstream sites of the transcriptional start site. p-60PDGF-CAT was constructed as described (15). Oligonucleo-tides containing the same EGR-1 binding sites and TCC motifwere also cloned into the HindIII site upstream of p-60PDGF-CAT, designated as p3XE PDGF-CAT and p3XTPDGF-CAT.
Abbreviations: PDGF, platelet-derived growth factor; CMV, cyto-megalovirus; EGR, early growth response gene; CAT, chloramphen-icol acetyltransferase; WT, Wilms tumor.*To whom reprint requests should be addressed.
8896

Proc. Natl. Acad. Sci. USA 90 (1993) 8897
DNase I Footprintfng and Gel Mobility Shift Assays. TheApa I-Sma I DNA fragment (-158 to +138, relative to thetranscription start site) of the PDGF A-chain gene wasend-labeled with dATP, dCTP, and dGTP, and avian myelo-blastosis virus reverse transcriptase. The DNase I footprint-ing assay was done as described (15).For gel mobility shift assays, 20 or 100 ng of the purified
WT1 and EGR-1 (3) were combined with binding buffer (10mM Tris-HCl, pH 7.9/50 mM MgCl2/1 mM dithiothreitol/1mM EDTA/5% glycerol/5% sucrose/l mM phenylmethyl-sulfonyl fluoride) and 2 ,ug of poly(dI-dC) (Boehringer Mann-heim). After 5 min at room temperature, 1 ng (=1 x 10W cpm)of 32P-labeled oligonucleotide probe was added, the incuba-tion was continued for 15 min, loading buffer was added, andDNA-protein complexes were separated from unboundDNA in nondenaturing 7.5% acrylamide gels by electropho-resis at 100 V for 3-4 hr. The running buffer consisted of 45mM Tris-borate (pH 8.3), 45 mM boric acid, and 1 mMEDTA. The gels were dried and analyzed by autoradiogra-phy. The oligonucleotides were end-labeled with T4 polynu-cleotide kinase and [y-32P]ATP (NEN). Unlabeled competi-tor DNAs, when appropriate, were added at the same time asthe labeled DNAs.DNA Transfections and Chloramphenicol Acetyltransferase
(CAT) Assays. NIH 3T3 fibroblast cells were maintained inDulbecco's modified Eagle's medium with l1o0 (vol/vol)fetal calf serum. The cells were plated at a density of 1 x 106cells per 100-mm dish 24 hr prior to transfection and trans-fected by calcium phosphate coprecipitation (16) with re-porter plasmid (5 ug) or (-galactosidase expression plasmid(1 ug) to establish transfection efficiency. The total amountof the expression vector added in each transfection wasadjusted to 20 pg by addition of CMV vector alone. Forty-eight hours after transfection, cell extracts were prepared,aliquots were normalized for transfection efficiency by assayof 13-galactosidase activity (17), and CAT activity was deter-mined (18). After autoradiographic exposure, the TLC plateswere assayed by scintillation spectrometry, and the percentconversion values were calculated.
RESULTSTo identify binding sequences that may recognize WT1within the DNA sequences downstream of the initiation siteof transcription of the PDGF A-chain, we used DNase Ifootprinting assays with a 292-bp 32P-end-labeled DNA frag-ment containing nucleotides -154 to +138 (relative to tran-scription start site +1) and purified WT1. It was found thatWT1 protects a region ofDNA downstream of the transcrip-tion start site (Fig. 1A) that mapped within the T+C richregion (+50 to +70) including the DNA sequence 5'-CTACTCCTCCTCCTCCTCTCC-3' (known as the TCC mo-tif; ref. 19) (Fig. 1B). WT1 also protects an extensive regionbetween residues -95 and -48 (Fig. 1B), a region thatincludes the previously identified two overlapping WT1 bind-ing sequences (5'-GCGGGGGCG-3') that are found from -69to -55 within the PDGF A-chain promoter (8). Two otherpartially protected sequences, which correspond to se-quences 5'-GCCCGCCCA-3' and 5'-GCCGCCGGCCG-GC-3' were also identified, but these sites did not appear tobe functional in previous cotransfection assays (6).To confirm the results ofDNase I footprinting, gel mobility
shift assays were performed with a synthetic oligonucleotidethat encompasses the TCC motifand purified WT1. A distinctprotein-DNA complex was found (Fig. 2A), indicating thatthis DNA element recognizes WT1. Since EGR-1 shares asimilar zinc-finger domain with WT1 (1), in a control exper-iment, purified EGR-1 also was tested and shown to form acomplex with the end-labeled probe in a gel mobility shiftassay. The specificity of WT1 for this DNA element was
A 123C
nW!:Ug
t.. N.* N .* ..^i '' |
,: .: ..:
! w *
-a.__ :.t W
*__ X,
456
] IH
I
B GGGCCCTGCCGGGTCCGCACGAACCCCTAGCGCTTCCGAGGTGCGGGT-95 I
CCCAGGCCCGGAATCCGGGGAGGCGGGGGGGGGGGGGCGGGGGCGGGl-4a
GGCGGGGGAGGGGCGCGGCGGCGGCGCTATAA:CCCTCTCCCCGCCGCC
GGCCGGCTCOCACACGCGCGCCCTGCGGAGCCCGCCCAACTCCGGCGAG*50 II *70
CCGGCCTGCGCCTACTCCTCCTCCTCCTCTCCCGGCGGCGGCTGCGGCGG
AGGCGCCG
FIG. 1. (A) DNase Ifootprinting ofthe putative WT1 binding sitesin the PDGF A-chain promoter. The probe was exposed to DNase Iwithout WT1 (lanes 1 and 4) or with 500 ng (lanes 2 and 5) or 1000ng (lanes 3 and 6) of purified WT1. An autoradiograph is shown of theDNase I footprinting result obtained with the noncoding strand of the292-bp Apa I-Sma I probe. (B) The sequence of the minimal PDGFA-chain promoter is shown. The transcription start site is depicted bya bent arrow (+1), the TATA box is boxed, the results of the DNaseI footprinting are indicated by overlines (I and II).
examined in competition assays with unlabeled probe andoligonucleotides that contain the Spl and typical EGR-1binding consensus sequence (Fig. 2C). The unlabeled TCCmotif itself and the EGR-1 binding consensus sequenceelement compete effectively for WT1, whereas the oligonu-cleotide containing the Spl binding consensus sequence failsto compete (Fig. 2B), suggesting that WT1 recognizes theTCC motif sequence in a sequence-specific manner.To determine the DNA sequences that are required for the
binding of WT1, oligonucleotides of different lengths weresynthesized, end-labeled, and directly tested in gel mobilityshift assays with purified WT1 (Fig. 3). The results establishthat the TCTCCC sequence at the 3' end is required foroptimum WT1 binding; shorter oligonucleotides result inweakly detectable or undetectable complexes in gel retarda-tion assays. Base pair substitutions were also tested; dinu-cleotide substitutions within the TCC motif strongly reducethe binding ofWT1 (Fig. 3; M4-M8), and single substitutionsof T in the TCC sequence markedly reduce the effectivenessof the oligonucleotides in recognizing WT1 (Fig. 3; M9).These data indicate that the 5'-TCCTCCTCCTCCTC-TCCC-3' sequences are required for optimum WT1 bindingand suggest that this nucleotide sequence has only a limitedtolerance for change to retain recognition of WT1.To determine if this second WT1 binding sequence is
transcriptionally functional, we cloned oligonucleotides con-taining three contiguous TCC motifs upstream of a "basal"PDGF A-chain promoter (-60 to +386)-driven CAT reportergene (p-60 PDGF-CAT), designated p3XT PDGF-CAT (Fig.
Medical Sciences: Wang et al.

8898 Medical Sciences: Wang et al.
A Wit EGR-illOngsOng lO
UIMbeld EOR-i SpiB probe .t site
0 1lOx 5OslOx SOxlOx 50X TCCM1 M2 M3 M4 M5 M6 M7 M8 M9
.#IJIL.-,
TCC:Ml:M2:M3:M4:M5:M6:MTM8:MS
TACTCCTCCTCCTCCTCTCCCGTACTCCTCCTCCTCCGTACTCCTCCTCCGTACTCCTCCGTAAACCTCCTCCTCCTCTCCCGTACTAAATCCTCCTCCTCTCCCGTACTCCTAATCCTCCTCTCCCGTACTCCTCCTAATCCTCTCCCGTACTCCTCCTCCTAATCTCCCGTACACCACCACCTCCTCTCCCG
Freeprobe
Ree___probe
C Probe: 5 - T AC T CC T CC TCCT CCT CTCCCG- 33 -ATGAGGAGGAGGAGGAGAGGGC- 5'
Spi site: 5- ATTCGATCGGGGCGGGGCGAGC -3'3- TAAGCTAGCCCCGCCCCGCTCG-55
EGR-1 site: 5 - GGCCCTCGCCCCCGCGCCGGG - 33- CCGGGAGCGGGGGCGCGGCCC -5
FIG. 2. Gel mobility shift and competition assays. (A) DNA-binding activities ofWT1 and EGR-1. The oligonucleotide probe wasend-labeled and mixed with 10 ng and 50 ng of purified WT1 andEGR-1 proteins. The DNA/protein mixture was fractionated on a7.5% native polyacrylamide gel, dried, and exposed to x-ray film. (B)Competition assays of WT1-DNA complexes with unlabeled probeand oligonucleotides corresponding to the Spl binding sequence andthe EGR-1 binding sequence. Numbers (lOx and 50x) indicate themolar excess of competitor to labeled probe. (C) DNA sequences ofprobe and competitors.
4A). p60 PDGF-CAT lacks most of the 5' G+C-rich regionimmediately upstream of the TATA box, including the twooverlapping WT1 binding sites (-55 to -69) (Fig. 4A). p3XTPDGF-CAT was cotransfected into NIH 3T3 cells withCMV-WT1, an expression vector that contains the full-lengthWT1 cDNA under control of the CMV early promoter. CATactivity in lysates from p3XT PDGF-CAT-transfected cells isreduced by 90%6 in the presence ofWT1 (Fig. 4B). However,in the control experiment, CAT activity is enhanced 3- to4-fold when p-60 PDGF-CAT is cotransfected with the WT1expression vector (Fig. 4C), a result that is consistent with
FIG. 3. Gel mobility shift assay. Purified WT1 was incubated with0.2 nM end-labeled TCC motif and different double-stranded oligo-nucleotides substituted within or deleted from the TCC motif andelectrophoresed on nondenaturing 7.5% polyacrylamide gels. Thesequences ofthe DNA probes are shown at the right with the mutatedsites underlined.
our previous report (6) that WT1 transcriptionally activatesthe PDGF A-chain promoter when the upstream 5'-GCGGGGGCG-3' overlapping sequences (-55 to -69) areremoved. As an additional control, three contiguous typicalEGR-1 binding sites (5'-GCGGGGGCG-3') were cloned atthe same position (designated as p3XE PDGF-CAT) andshown to function equally as well as the TCC motif incotransfection assays (Fig. 4B). These results indicate thatthe TCC motif functions essentially equally well as thepreviously identified WT1 binding sequence (5'-GCGGGGGCG-3'). In control experiments, CMV-WTZF,the control expression vector that contains only the zinc-finger domains, has essentially no effect when cotransfectedwith either of the reporter constructs (Fig. 4B).
Recently, we (6) established that WT1 contains domainsthat function independently to suppress or activate transcrip-tion of the PDGF A-chain gene. We (6) and others (7) alsoobserved that WT1 requires binding sequences positionedboth 5' and 3' of the transcription initiation site for transcrip-tional suppression. To further demonstrate that the TCCmotif functions equally well as the 5'-GCGGGGGCG-3'binding site, chimeric reporter constructs were made with thepreviously described WT1 binding sequence 5'-GCGGGGGCG-3' and the TCC motif in different combina-tions at both of the 5' and 3' positions (Fig. 5) in pACAT, aiinimal PDGF A-chain promoter (residues -60 to +3)
driving the CAT reporter gene construct. pACAT includesessentially only the TATA box and the transcription start site
A
p-60 PDGF-CAT
p3XE PDGF-CAT 3X
p3XT PDGF-CAT m
C p-60 PDGF-CAT
BITATAI r CAT l
-60 +388
n nI ITATA1 Ir CAT-60 +388
XTC ITATA|r CAT-60 +388
p3XT PDGF-CAT
CMV-WT1 CMV-WTZF
ig 0 2.5 5 15 20 20
p3XE PDGF-CATI~~~~~~~~~~~~~~~~~~~~~
CMV-WT1 CMV-WTZF
0 2.5 5 15 20 20
*0
1 2 3 4 5 6Activity 1.0 0.9 0.35 0.1 0.05 1.2
iC.)
0
5;
LiN
H
..
1 2 3 41.0 3.5 3.8 1.2
7 8 9 10 11 121.0 0.8 0.3 0.08 0.05 1.4
FIG. 4. Transcriptional function of the TCC motif. (A) Reporter plasmids used in transient transfectionassays contain the PDGF A-chain promoter (-60 to + 388) or the PDGF promoter with three TCC motifs or withthree EGR-1 binding sites (5'-GCGGGGGCG-3') fused upstream (p3XE PDGF-CAT and p3XT PDGF-CAT,respectively). (B and C) Calcium phosphate-mediated transfections were performed in murine NIH 3T3fibroblasts. Each dish of cells was transfected with the indicated reporter vector (5 ,ug), the indicated expressionplasmid [in B, 0-20 Mg was added; in C, 20 Mg ofCMV, 5 gg (1:1, weight ratio ofreporter to expression plasmid)or 20 Mg (1:4) of CMV-WT1, or 20 Ag of CMV-WTZF was added], and a P-galactosidase expression vector (1mg) as an internal control for transfection efficiency. The total amount of vector in each transfection mixturewas kept constant at 20 ,ug by addition of CMV vector alone. Forty-eight hours after transfection, cell extractswere prepared and aliquots were normalized for transfection efficiency via assay of 3-galactosidase activity todetermine the CAT activity. After autoradiographic exposure, the TLC plates were counted and percentconversion values were calculated. Results are expressed relative to the CAT activities in mock transfectantswhich were arbitrarily set at 1.0.
Z .-,. .1 ... :... I.",
r;
Proc. Natl. Acad Sci. USA 90 (1993)
AL.IIA .. 1L.. ...k-.410. *11.1VW, W, "
:
Proc. Natl. Acad. Sci. USA 90 (1993) 8899
of the PDGF A-chain gene (6). These constructs were co-transfected into NIH 3T3 cells with the WT1 expressionvector. In a control experiment, WT1 did not influence thelow but reproducible activity of the PDGF A-chain minimalpromoter. When reporter plasmids containing either of theWT1 binding sequences (5'-GCGGGGGCG-3' or TCC motif)placed 5' or 3' of the transcriptional start site were cotrans-fected with CMV-WT1, an 3-fold increase of CAT activitywas repeatedly observed. Thus, the 5'-GCGGGGGCG-3' andTCC motifappear to functionally influence promoter activityin an essentially identical manner. Furthermore, when eitherof the WT1 binding sites was fused into either the 5' or 3'position in any combination with the other, cotransfectionwith CMV-WT1 efficiently suppressed transcriptional activ-ity of the PDGF A-chain promoter =90%o (Fig. 5), indicatingthat either the 5'-GCGGGGGCG-3' or the TCC motif func-tions equally well at these sites.We also noted a number of potential TCC motifs in the 5'
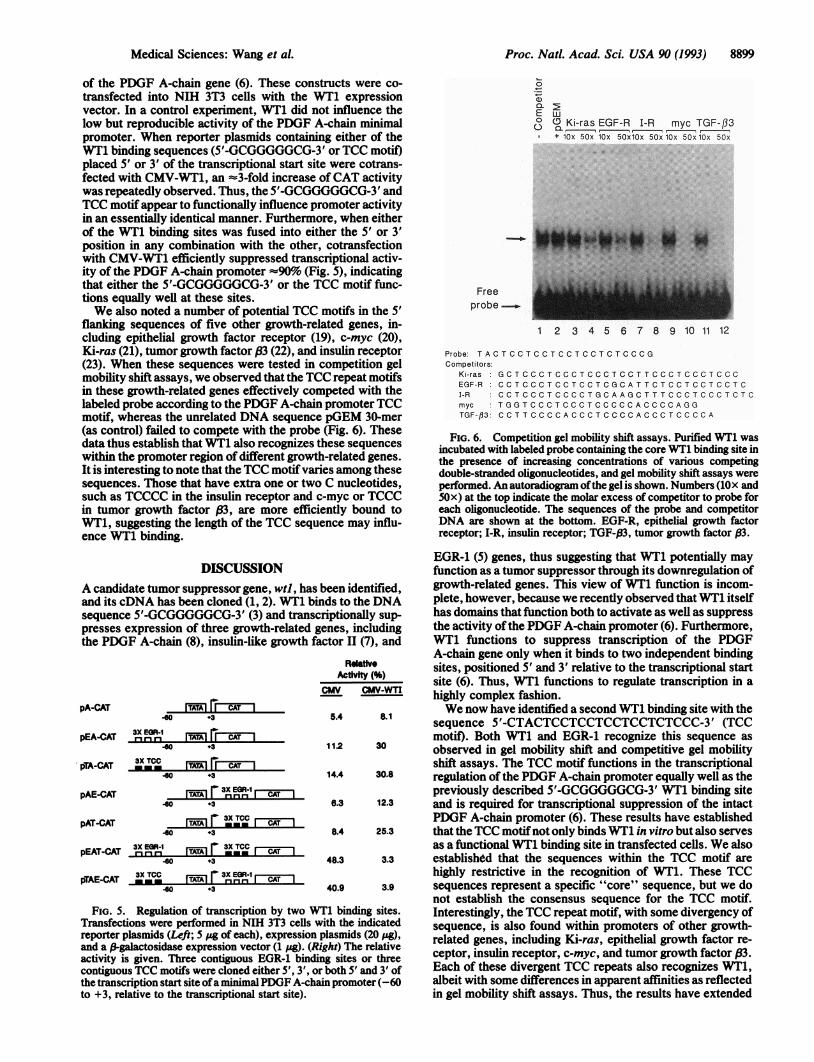
flanking sequences of five other growth-related genes, in-cluding epithelial growth factor receptor (19), c-myc (20),Ki-ras (21), tumor growth factor (3 (22), and insulin receptor(23). When these sequences were tested in competition gelmobility shift assays, we observed that the TCC repeat motifsin these growth-related genes effectively competed with thelabeled probe according to the PDGF A-chain promoter TCCmotif, whereas the unrelated DNA sequence pGEM 30-mer(as control) failed to compete with the probe (Fig. 6). Thesedata thus establish that WT1 also recognizes these sequenceswithin the promoter region of different growth-related genes.It is interesting to note that the TCC motifvaries among thesesequences. Those that have extra one or two C nucleotides,such as TCCCC in the insulin receptor and c-myc or TCCCin tumor growth factor (83, are more efficiently bound toWT1, suggesting the length of the TCC sequence may influ-ence WT1 binding.
0
a)
E0
w(DKi-ras EGF-R I-R myc TGF-f33+ lOx 50x lOx 50xlOx 50x lOx 50x1Ox 50x
Freeprobe
1 2 3 4 5 6 7 8 9 10 11 12
Probe: TACTCCTCCTCCTCCTCTCCCGCompetitors
Ki-ras :GCTCCCTCCCTCCCTCCTTCCCTCCCTCCCEGF-R CCTCCCTCCTCCTCGCATTCTCCTCCTCCTCI-R CCTCCCTCCCCTGCAAGCTTTCCCTCCCTCTCmyc TGGTCCCTCCCTCCCCCACCCCAGGTGF-fl3 CCTTCCCCACCCTCCCCACCCTCCCCA
FIG. 6. Competition gel mobility shift assays. Purified WT1 wasincubated with labeled probe containing the core WT1 binding site inthe presence of increasing concentrations of various competingdouble-stranded oligonucleotides, and gel mobility shift assays wereperformed. An autoradiogram ofthe gel is shown. Numbers (lOx and50x) at the top indicate the molar excess of competitor to probe foreach oligonucleotide. The sequences of the probe and competitorDNA are shown at the bottom. EGF-R, epithelial growth factorreceptor; I-R, insulin receptor; TGF-P3, tumor growth factor ,B3.
DISCUSSIONA candidate tumor suppressor gene, wtl, hasand its cDNA has been cloned (1, 2). WT1 bisequence 5'-GCGGGGGCG-3' (3) and trans4presses expression of three growth-related Ithe PDGF A-chain (8), insulin-like growth f;
pA-CAT TAA3Ir CAT I-80 +3
3X EpEA-CAT 3i E-uR-i ITATAI I CAT I.60 +3
pTA-CAT 3X TCC rTATAI ri CAT I-60 +3
pAE-CAT
pAT-CAT
ryA-A 3X EGR-1I[T^!^, r lnnrnr CAT 140 +3
--r 3X TCC
ITATAI mmr I CAT I40 +3
3X E0R-1 3X TCCpEAT-CAT -r-i" ITATAI r 3XTC I CAT.60 +3
PTAE-CAT 3X TCC rVAI r 3X EGR-1 CAT40 +3
FIG. 5. Regulation of transcription by two UTransfections were performed in NIH 3T3 cells N
reporter plasmids (Left; 5 Mg of each), expressionand a ,-galactosidase expression vector (1 pg). (Ractivity is given. Three contiguous EGR-1 bindicontiguous TCC motifs were cloned either 5', 3', othe transcription start site ofa minimal PDGF A-ch;to +3, relative to the transcriptional start site).
EGR-1 (5) genes, thus suggesting that WT1 potentially mayfunction as a tumor suppressor through its downregulation of
,been identified, growth-related genes. This view of WT1 function is incom-inds to the DNA plete, however, because we recently observed that WT1 itselfcriptionally sup- has domains that function both to activate as well as suppressgenes, including the activity ofthe PDGF A-chain promoter (6). Furthermore,actor 11(7), and WT1 functions to suppress transcription of the PDGF
A-chain gene only when it binds to two independent bindingRAtiityV sites, positioned 5' and 3' relative to the transcriptional start
Activity ( )w site (6). Thus, WT1 functions to regulate transcription in aCM CM-WTI highly complex fashion.
We now have identified a second WT1 binding site with thesequence 5'-CTACTCCTCCTCCTCCTCTCCC-3' (TCCmotif). Both WT1 and EGR-1 recognize this sequence as
11.2 30 observed in gel mobility shift and competitive gel mobilityshift assays. The TCC motif functions in the transcriptional
14.4 30.8 regulation of the PDGF A-chain promoter equally well as thepreviously described 5'-GCGGGGGCG-3' WT1 binding site
6.3 12.3 and is required for transcriptional suppression of the intactPDGF A-chain promoter (6). These results have established
8.4 25.3 that theTCC motifnot only binds WT1 in vitro but also servesas a functional WT1 binding site in transfected cells. We also
48.3 3.3 established that the sequences within the TCC motif arehighly restrictive in the recognition of WT1. These TCC
40.9 3.9 sequences represent a specific "core" sequence, but we donot establish the consensus sequence for the TCC motif.
VTl binding sites. Interestingly, the TCC repeat motif, with some divergency ofwith the indicated sequence, is also found within promoters of other growth-Xplasmids (20 .g), related genes, including Ki-ras, epithelial growth factor re-
ing sites or three ceptor, insulin receptor, c-myc, and tumor growth factor (3.srboth 5' and 3' of Each of these divergent TCC repeats also recognizes WT1,ain promoter (-60 albeit with some differences in apparent affinities as reflected
in gel mobility shift assays. Thus, the results have extended
Medical Sciences: Wang et aL

8900 Medical Sciences: Wang et al.
the number of growth-related genes to which WT1 may bindand perhaps that may transcriptionally influence expression.
Alternatively spliced forms ofWT1 have been described inWT (24). One of the alternatively spliced WT1 gene productsencodes three amino acids (KTS) in between zinc fingers 3and 4 of WT1 that prevent WT1 from recognizing the se-quence 5'-GCGGGGGCG-3'. This form ofWT1 (WT+KTS)is the predominant form of human and murine WT1 mRNAin all cells that express WT1 (24). However, becauseWT+KTS fails to recognize 5'-GCGGGGGCG-3', the func-tion of this product of the alternatively spliced mRNAs wasunknown. Recently, however, it has been found that anothersequence (5'-CTCCCTCCCAGGAGAGGGAGGAT-3') rec-ognizes both WT1 and WT1+KTS using the whole genomePCR method (25), suggesting that WT1 actually recognizedthe T+C-rich sequence. Previously, a sequence (CTC-CCCAG) was also cloned as a recognition site of WT1 (3).However, these sequences were not identified in the pro-moter of any gene, and thus their functional significance isunknown. It thus may be of considerable interest to note thatthis sequence is similar to the TCC repeat motif that we havenow described, both in the sense and in the antisense strands,suggesting that WT+KTS may recognize the TCC motiffound in PDGF A-chain gene.The results we describe have identified and characterized
another WT1 binding element, the TCC repeat motif, thatfunctions equally with the previously described 5'-GCGGGGGCG-3' sequence. This TCC motif is found in fiveother growth-related gene promoters that also bind WT1.Thus, WT1 appears to have the potential to function in ahighly versatile manner to influence transcription of multiplegrowth-related genes.
We thank Dr. Frank Rauscher III (Wistar Institute, Philadelphia)for the generous gifts ofpurified WT1 and EGR-1 and the expressionvectors CMV-WT1 and CMV-WTZF. This work was supported bygrants HL31102, HL14147, and CA49712 from the National Insti-tutes of Health, and by a grant from Monsanto.
1. Call, K. M., Glaser, T., Ito, C. Y., Buckler, A. J., Pelletier, J.,Haber, D. A., Rose, E. A., Kral, A., Yeger, H., Lewis, W. H.,Jones, C. & O'Housman, D. E. (1990) Cell 60, 509-520.
2. Gessler, M., Poustka, A., Gavenee, W., Neve, R. L., Orkin,S. H. & Bruns, G. A. P. (1990) Nature (London) 343, 774-778.
3. Rauscher, F. J., III, Morris, J. F., Tournay, D. E., Cook,D. M. & Curran, T. (1990) Science 250, 1259-1262.
4. Sukhatme, V. P. (1990) J. Am. Soc. Nephrol. 1, 859-866.5. Madden, S. L., Cook, D. M., Morris, T. F., Gashler, A.,
Sukhatme, V. P. & Rauscher, F. J., III (1991) Science 253,1550-1553.
6. Wang, Z.-Y., Qiu, Q.-Q. & Deuel, T. F. (1993) J. Biol. Chem.268, 9172-9175.
7. Drummond, I. A., Madden, S. L., Rohwer-Nutter, P., Bell,G. I., Sukhatme, V. P. & Rauscher, F. J., III (1992) Science257, 674-678.
8. Wang, Z.-Y., Madden, S. L., Deuel, T. F. & Rauscher, F. J.,III (1992) J. Biol. Chem. 267, 21999-22002.
9. Maheswaran, S., Park, S., Bernard, A., Morris, J. F.,Rauscher, F. J., III, Hill, D. E. & Haber, D. A. (1993) Proc.Natl. Acad. Sci. USA 90, 5100-5104.
10. Deuel, T. F. (1987) Annu. Rev. Cell Biol. 3, 443-492.11. Raines, E. W., Bowen-Pope, D. F. & Ross, R. (1990) in
Handbook of Experimental Pharmacology: Peptide GrowthFactors and Their Receptors I, eds. Sporn, M. B. & Roberts,A. B. (Springer, New York), Vol. 95, pp. 173-262.
12. Fraizer, G. E., Bowen-Pope, D. F. & Vogel, A. M. (1987) J.Cell. Physiol. 133, 169-174.
13. Wang, Z.-Y., Lin, X.-H., Nobuyoshi, M. & Deuel, T. F. (1992)J. Biol. Chem. 267, 13669-13674.
14. Wang, Z.-Y., Lin, X.-H., Qiu, Q.-Q. & Deuel, T. F. (1992) J.Biol. Chem. 267, 17022-17031.
15. Lin, X.-H., Wang, Z.-Y., Gu, L.-J. & Deuel, T. F. (1992) J.Biol. Chem. 267, 25614-25619.
16. Graham, F. L. & Van der Eb, A. T. (1973) J. Virol. 52,456-467.
17. Spaete, R. R. & Mocarski, E. S. (1985) J. Virol. 56, 135-143.18. Gorman, G. M., Moffat, L. F. & Howard, B. H. (1982) Mol.
Cell. Biol. 2, 1044-1051.19. Johnson, A. C., Jinno, Y. & Merlino, G. T. (1988) Mol. Cell.
Biol. 8, 4174-4184.20. Kinniburgh, A. T. (1989) Nucleic Acids Res. 17, 7771-7778.21. Hoffman, E. K., Trusko, S. P., Murphy, M. & George, D. L.
(1990) Proc. Natl. Acad. Sci. USA 87, 2705-2709.22. Latyatis, R., Renher, F., Williams, T., Sporn, M. B. & Rob-
erts, A. B. (1991) Nucleic Acids Res. 19, 6419-6425.23. Seino, S., Seino, M., Nishi, S. & Bell, G. (1989) Proc. Natl.
Acad. Sci. USA 86, 114-118.24. Haber, D. A., Sohn, R. L., Buckler, A. J., Pelietier, J., Call,
K. M. & Housman, D. E. (1991) Proc. Natl. Acad. Sci. USA88, 9618-9622.
25. Bickmore, W. A., Ogheen, K., Little, M. H., Seawright, A.,Van Heyninger, V. & Hastie, N. D. (1992) Science 257, 235-237.
Proc. Natl. Acad. Sci. USA 90 (1993)