a review of phylogeography and the integration between ... · luciano beheregaray molecular ecology...
TRANSCRIPT

Luciano Beheregaray
Molecular Ecology Lab
School of Biological Sciences
A review of Phylogeography
and the integration between
biologists and Earth scientists

Outline
• Earth sciences and Phylogeography
• Perspectives
• A review of twenty years of Phylogeography
• The phylogeographer as a powerful naturalist
• Case Studies:- young radiation & ecological speciation in Brazil- history and conservation rescue in the Galápagos- comparative biogeography in Wallacea

www.bio.mq.edu.au/molecularecology (soon to be moved to Flinders)

Molecular Ecology Group for Marine Research
Our focus:
Understanding genetic and ecologic
connectivity in the sea

The Phylogeographer:
A Powerful Naturalist
• A nature admirer’s, an explorer
• A population biologist
• Integrative in essence:
- from genes to geosciences- from generations to millennia

Phylogeography, the Study of Population History
Phylogeography is concerned about patterns and processesat the tips of the Tree of Life (TOL)

Let’s zoom in the TOL

modified from Sunnucks (2000) TREE
Three ‘time slices’ in genetics
Dark = most informative
Genotypic
Allelic
Time (generations)Data type:
Phylogenetic

Work in progress… a review on:
2 - Identify achievements, trends and challenges for the field

Articles and citations in phylogeography*
0
5000
10000
15000
20000
25000
0
200
400
600
800
1000
1200
1987 1988 1989 1990 1991 1992 1993 1994 1995 1996 1997 1998 1999 2000 2001 2002 2003 2004 2005 2006 2007 2008 2009 2010
Nu
mb
er o
f ci
tati
on
s
Nu
mb
er o
f art
icle
s
Publication year
Articles
Citations
* 2010 (projections as in Nov 1st)

Distribution of articles in phylogeography by taxonomic group
0
5
10
15
20
25
Mammals Fishes Plants
(terrestrial)
Invertebrates
(terrestrial)
Herps Invertebrates
(aquatic)
Birds Microorganisms Plants
(aquatic)
Fungi
% o
f a
rtic
les

Genetic markers in phylogeography: temporal trends

• mtDNA still reigns supreme (81% of all studies or 75% alone) • recently augmented by ever growing DNA barcode data…
• recent escalation of multilocus surveys (e.g. introns and microsatellites) improves inferences about population history:
- estimates of divergence time and Ne- accommodate for coalescence stochasticity
Some food for thought on genetic markers
• multilocus surveys can synergistically combine the power of POPULATION GENETICS with phylogeography:
- identify population units, hybridization and migrants (& their rates)- brings ecology (niche) and behaviour into phylogeography

• problems with multilocus (nuclear) markers:
- technical development can be difficult- coalescence tends to be less ‘convenient’ (much longer time)- rates of evolution more variable and unpredictable- population screening can be difficult and expensive (diploidy and
recombination)
Some food for thought on genetic markers
• Genomics provides solutions to many such problems and brings ‘new’ markers into play (exon capture, functional SNPs, etc)
Pattern? mtDNA (in animals)!
Processes? multilocus endeavour!

0.77
0.15
0.0240.06
Northern Hemisphere
Southern Hemisphere
Northern & Southern
Global
0.65
0.18
0.17 Terrestrial
Freshwater
Marine
The global geography of Phylogeography

0
5
10
15
20
25
30
35
Europe North
America
Asia Africa Australia South
America
Antarctica
Continent
% o
f art
icle
sThe global geography of Phylogeography

Mittermeier, Myers & Mittermeier
University of Chicago Press (2000)
Country Ranking based on endemism and
species diversity
Ranking based on papers in
phylogeography
Brazil 1 15
Indonesia 2 40
Colombia 3 65
Australia 4 5
Mexico 5 27
Madagascar 6 54

Conservation Biology is about prioritizing resources
RARITY RESTRICTED DISTRIBUTION CHARISMA
ECONOMICS ECOLOGICAL IMPORTANCE
EVOLUTIONARILY DISTINCT POPULATIONS
(as revealed by phylogeography)
Some food for thought on conservation & phylogeography
Historically isolated populations are not only genetically distinct
but also tend to be locally (ecologically) adapted
Comparative Phylogeography can help a lot here!

Genomics has jumped forwarded and moved into the wild!
Some food for thought on genomics & phylogeography
PROBLEM: Technological and analytical challenges can increase
(even more) the gap between phylogeographers from the
developed and developing world
Ecological Genomics & Phylogeography:
„Functional Phylogeography‟ – an integration to inform on the
origins and evolutionary trajectories of neutral and adaptive
genetic variation at landscape level
Functional (ecological) Genomics & Natural History:
„Mechanistic natural history‟ (Americ. Soc. Naturalists, 2002)

0.68
0.25
0.08
Single taxon
Multiple taxa
Comparative phylogeography
0.69
0.11
0.02
0.18 Quaternary (0 - 2 My)
Tertiary (2 - 66 My)
Quaternary & Tertiary
Undetermined
Nature of the study and timescale in Phylogeography

- Geology: Geomorphology PalaeontologyMineralogy and petrology Volcanology Geochemistry Sedimentology
- Geophysics- Hydrology:
Oceanography Limnology
- Glaciology- Atmospheric sciences:
Meteorology Climatology
Phylogeography is a historical discipline and relies on information from Earth Sciences

Most earth scientists have generally ignored anything different than physical records
Phylogeographers use other data sets (in addition to genetics) but…
…they have often inefficiently, simplistically (and sometimes incorrectly) explored and interpreted data about Earth’s history
Earth Sciences and Phylogeography:

Earth scientistsTaxonomists
Phylogeographers &
population geneticists
An integration to discover, document and understand
historical dimensions of biodiversity
Beheregaray & Caccone (2007) Journal of Biology

CASE STUDY 1:
When the branches of the Tree of Life are not well
formed
Time (generations)

O. argentinensis
• marine and estuarine
• estuarine speciationBeheregaray & Levy (2000) Copeia
A very young radiation?
O. perugiae species group
• freshwater lakes
• morphological divergence
Dyer & Malabarba (2003) Ichthyol. Explor. Freshwaters

75 km
ArgentinaUruguay
Southern Brazil


10 km
Freshwater group
+ +
- -
- -
80,000 ya
16,000 ya
6,000 ya
Beheregaray et al. (2002) Proc. Royal Soc. London B

500 km
Marine-estuarine group
+ +
marine pops
+ +
estuarine pops
+ +
Beheregaray & Sunnucks (2001) Molecular Ecology

• colonization of new habitats
• divergent natural selection
10 km
“ecological speciation with gene flow”
Isolation by adaptation in estuaries
Beheregaray & Sunnucks (2001) Molecular Ecology

mtDNA network – all coastal Odontesthes + riverine perugiae
marine sp
estuarine sp
O. mirinensis
O. sp A
O. piquava
O. ledae
O. perugiae
O. bicudo
Beheregaray (in prep)

Microsatellite tree (Dce genetic distance)
0.01
O. perugiae
O. bicudo PLEISTOCENE
O. sp A HOLOCENE 1
O. mirinensis HOLOCENE 1
O. piquava HOLOCENE 1
O. ledae HOLOCENE 2
O. sp C100
100
100
100
84
71
O. sp B
O. argentinensis
riverine
marine
estuarine
coastal
lakes
?
Beheregaray (in prep)
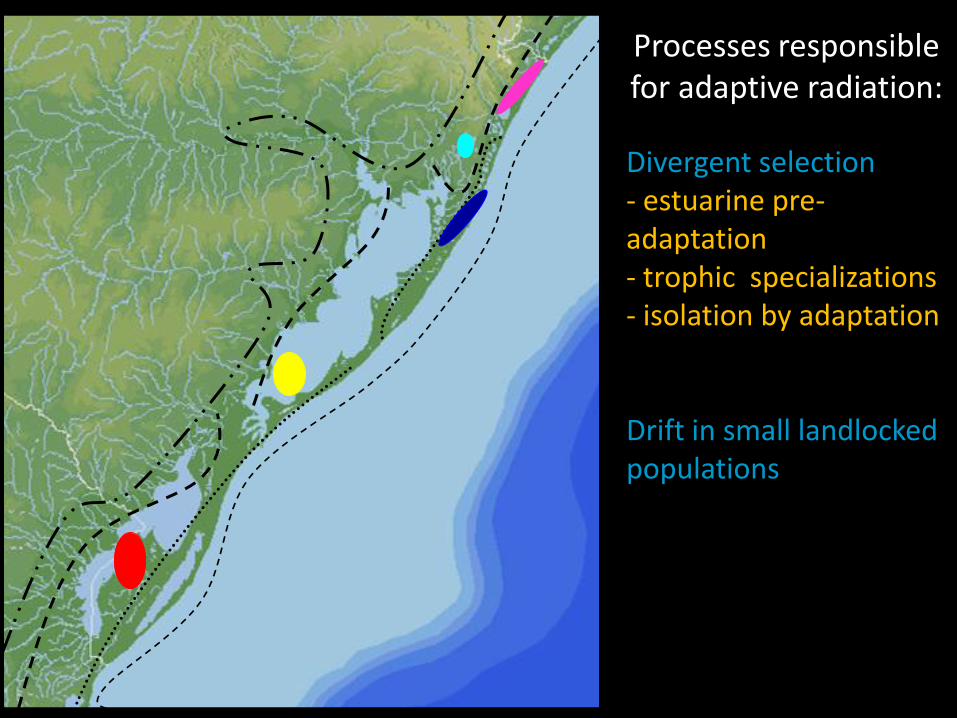
Processes responsible for adaptive radiation:
Divergent selection- estuarine pre-adaptation- trophic specializations - isolation by adaptation
Drift in small landlocked populations

CASE STUDY 2:
Population history and conservation rescue on
islands
Time (generations)
with Jeff Powell and Gisella Caccone (Yale)


male-biased dispersal
(Rasmann et al. 1997)
hybridization
(Grant & Grant 1996)
Biogeographic prediction:
“colonization and decreasing phylogeographic structure in E-W direction”
multiple colonization (Wright 1983)

DomedSaddleback
Giant Galápagos tortoises – Geochelone lineage

3 species finished off by zoo collectors and scientific expeditions!
Giant Galápagos tortoises – Geochelone lineage
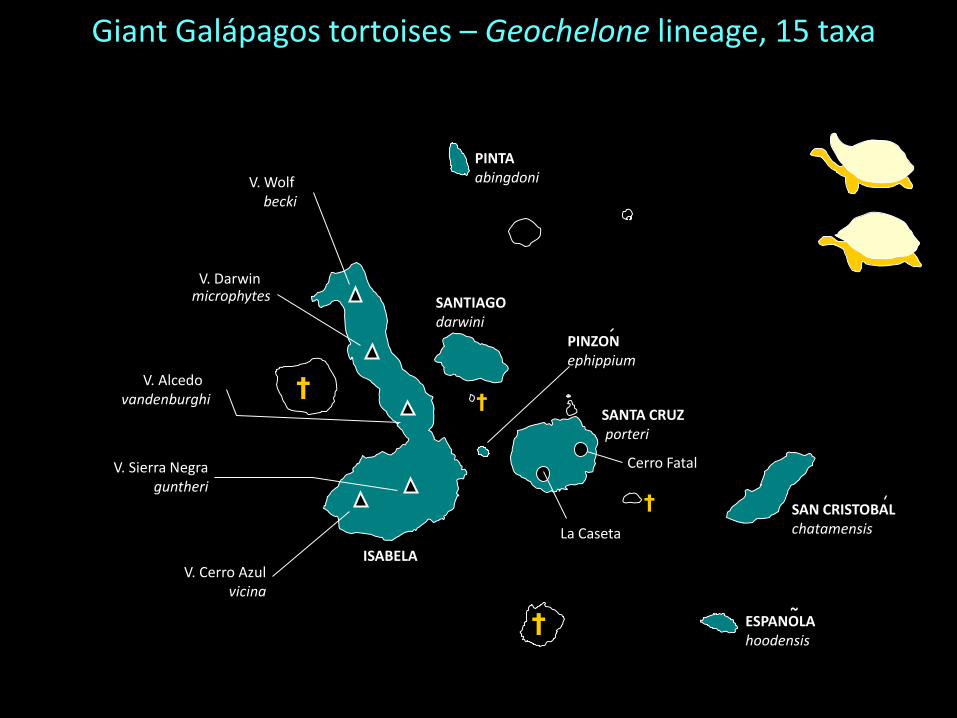
PINZONephippium
SANTIAGO darwini
SAN CRISTOBALchatamensis
Cerro Fatal
La Caseta
PINTA abingdoni
ESPANOLA hoodensis
ISABELA
V. Wolfbecki
V. Darwinmicrophytes
V. Alcedovandenburghi
SANTA CRUZ porteri
V. Cerro Azul vicina
V. Sierra Negraguntheri
~†
†
†
†
Giant Galápagos tortoises – Geochelone lineage, 15 taxa

Caccone et al. (2002) Evolution
Galápagos
Mainland South America
Bayesian tree4750 bp of mtDNA
Galapagos taxa evolved in situ
Caccone et al. (1999) PNAS

Younger islands (<1.2 My) Older Islands (~ 3.0 – 1.5 My)
Archipelago-wide phylogeography of giant tortoises (n = 802)

Younger islands (<1.2 My) Older Islands (~ 3.0 – 1.5 My)
Beheregaray et al. (2004) PNAS
Archipelago-wide phylogeography of giant tortoises (n = 802)

Older Islands
(~ 3.0 – 1.5 Ma)
• 100% lineages: island endemics
• highly divergent
• deep history:
inter-island divergences
of ~ 2 – 0.8 Ma
PINTA
ESPANOLA
SANTA CRUZ (La Caseta and Cerro Fatal)
SAN CRISTOBAL
PINZON
29 steps
~
16 steps48 steps
30 steps
20 steps
Beheregaray et al. (2004) PNAS

Younger Islands
(< 1.2 Ma)
• volcano populations are not
monophyletic
• recent migration events
• shallow history:
expansions / colonizations
of ~ 0.6 – 0.1 Ma
25 steps
ISABELA
V. Darwin
V. Alcedo
V. Cerro Azul
V. Sierra Negra
SANTIAGO
V. Wolf (northern ISABELA)
Clade B
Clade A
Beheregaray et al. (2004) PNAS
• Inferences supported
by microsatellite analysis
Ciofi et al. (2006) Genetics

biogeographic consensus!
Main findings…
established knowledge for further studies on:
- demographic history
- conservation rescue
identification of evolutionary events
chronologically associated with volcano
evolution

V. Wolf beckih = 0.69
V. Darwin microphyesh = 0.50
V. Sierra Negra
guntheri
h = 0.83
V. Cerro Azul
vicina
h = 0.87
Diversity of maternal lineages on Isabela Island
V. Alcedo
vandenburghi
h = 0.17

• largest extant pop in the archipelago (~ 4,000 – 6,000)
• Alcedo was probably never exploited by whalers or pirates
Why reduced genetic variability in
Alcedo tortoises?
Geist et al. (1994) Bulletin of Volcanology
- 3.4 Km3 of rhyolitic tephra
- K-Ar dating: ~ 100 (±20) kya
Alcedo experienced a catastrophic volcano
eruption!

Used larger sample and multilocus data to:
• test for past population contraction
• date the population contraction

A = 7.3 (± 0.5)
A = 9.1 (± 0.6)
P = 0.01
P = 0.68
P = 0.51A = 8.9 (± 0.6)
Massive bottleneck in Alcedo tortoises
P = 0.01
P = 0.68
P = 0.51
Beheregaray et al. (2003) Science

DNA dating: ~ 88 kya (110-70 kya)
K-Ar dating: ~ 100 (±20) kya
Estimating the age of the population bottleneck
61
6161
61
62 63
64 65
• 61 is an „old‟ maternal lineage
Genes have recorded a prehistoric volcano eruption in the Galapagos!
Beheregaray et al. (2003) Science

Phylogeography and Conservation Rescue

A NEW and highly endangered species of giant tortoises
PINTA
ESPANOLA
SANTA CRUZ (La Caseta and Cerro Fatal)
SAN CRISTOBAL
PINZON
29 steps
~
16 steps48 steps
30 steps
20 steps
Beheregaray et al. (2003) Conservation Genetics
Russello et al. (2005) Biology Letters
The Cerro Fatal species (~ 70 left)

Extinct in the wild (IUCN 1996)
The story of a conservation icon:
„Lonesome George‟ (Geochelone abingdoni)

www.firstscience.com/home/blogs/George.html
The story of a conservation icon:
„Lonesome George‟ (Geochelone abingdoni)

The sexual problems of a conservation icon…
„Is he gay, impotent or just bored?‟
New Scientist

„Alien‟ matrilines found on Volcano Wolf
The fortunate phylogeographic twist…
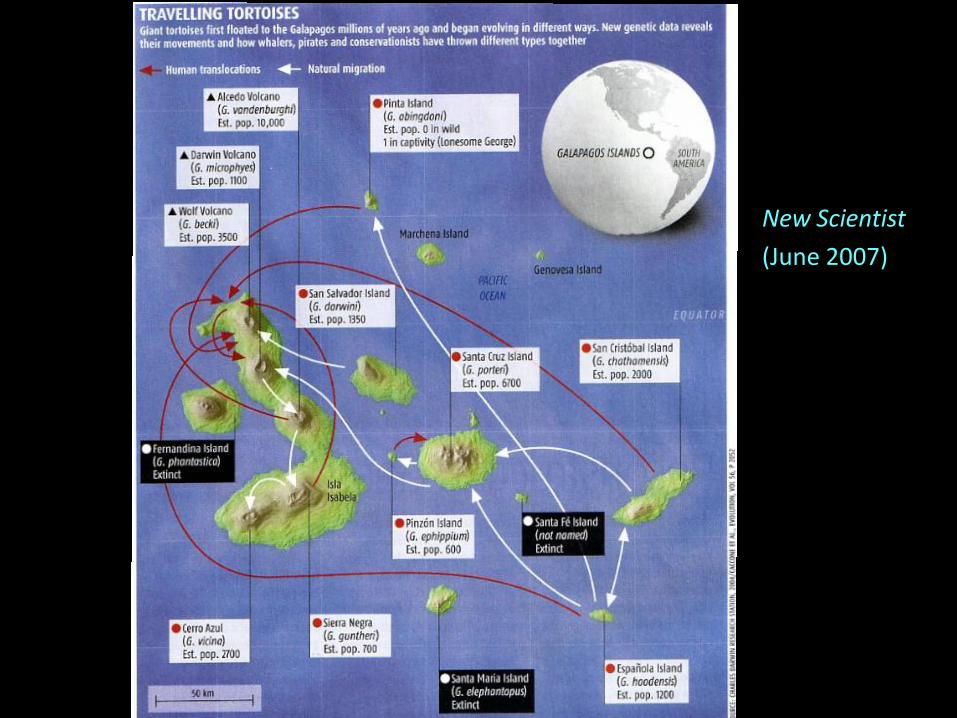
New Scientist
(June 2007)

-7 Pinta museum specimens:
CAS (San Diego)
- 700 bp mtDNA
- 10 microsatellite loci
- all archipelago populations
- + 27 aliens from Wolf
Building an extinct nuclear DNA database…

Russello, Beheregaray et al. (2007) Current Biology
Lonesome George is not alone among Galápagos tortoises!

CASE STUDY 1:
Comparative population history in Wallacea: tying
geology with genetics
Time (generations)

0 500 1000 2000
kilometres
Equator
DISTRIBUTION
MAP
Charaxes
CethosiaDelias
FIELD WORK:
100+ sites,
13 countries
Solomon
Islands
Vanuatu
New
Caldeonia
Papua
New Guinea
Indonesia
Philippines
Malaysia
China
TaiwanMyanmar
Thailand
Vietnam
Laos
Cambodia
Brunei
Australia

020 1090 304060 507080
Millions of years
Oligocene Pli
oc
en
e
PleistoceneMioceneEocenePaleogeneCretaceous
Cethosia
Chronogram based on
Bayesian topology
C. mindanensis1
C. mindanensis2C. hypsea3
C. hypseabatu
C. penthesileapak
C. methypsea
C. hypseafruh
C. hypsea6
C. hypsea7
C. gabinia
C. penthesileatim
C. penthesilealom
C. luzonica4
C. luzonica3C. luzonica6
C. cyane3C. cyanenwC. cyane1
C. tambora3
C. tambora5
C. tambora1C. picta
C. biblis 1
C. lamarckiC. moestaC. nietneri
C. obscura1
C. obscura7C. obscuraantC. obscura3
C. obscura5
C. vasilia1C. vasilia4C. cydippeobian
C. cydippe4C. cydippe2
C. myrinanw
C. cydippecydippeC. cydippeanton
C. myrinamyrinaC. myrinaribbei
Acraea
ActinoteC. lechenaulti
Heliconius1
Vagrans
Argynnis
Heliconius2
WA
LL
AC
EA
WALLACEAN
DIVERSIFICATION
Muller & Beheregaray (2010) MPE

Millions of years
010 535 152030 25
Charaxes
Chronogram based on
Bayesian topology
Oligocene Pliocene PleistoceneMioceneEocene
C. borneensis
C. bernardus
C. repetitus
C. fervens
C. plateni
C. marmax
C. aristogiton
C. bajula
C. psaphon
C. durnfordi
C. bupalus
C. kahrubra
C. sangana
C. antonius
C. amycus
C. latona
C. affinis
C. setan
C. musashi
C. eurialus
C. madensis
C. orilus
C. ocellatus
C. marki
C. mars
C. nitebis
C. elwesi
C. distanti
C. harmodius
C. solon
C. castor
Polyura
Euxanthe
Archaeoprepona
Prothoe
Anaea
CalinagaW
AL
LA
CE
AN
DIV
ER
SIF
ICA
TIO
N
WA
LL
AC
EA
Muller, Wahlberg & Beheregaray (2010) Biol J Linnean
Soc

5 045 101525303540
Millions of years
Oligocene PleistoceneMioceneEocene
20
Pliocene
Delias
Chronogram based on
Bayesian topology
D. elongatusD. clathrataD. roepkeiD. naisD. albertisiD. discusD. meekiD. niepeltiD. iltisD. bakeriD. nigrinaD. eximiaD. leuciasD. harpalyceD. dixeyiD. kummeriD. messalinaD. madetesD. descombesiD. ellipsisD. diaphanaD. caeneusD. periboeaD. mysisD. salviniD. bagoeD. eileenaeD. hypareteD. cunningputiD. chimbuD. heroniD. flavissisimaD. aroaeD. stresemanniD. schassmanniD. geraldinaD. microstichaD. gilliardiD. eichhorniD. abrophoraD. aganippeD. totilaD. ladasD. woodiD. blancaD. ninusD. benasuD. subnubilaD. surprisaD. belladonnaD. enniaD. chrysomelaenaD. apetalaD. laknekeiD. hippodomiaD. baracasaD. agostinaD. narsesD. lemoultiD. yagashitaiD. nysaD. brandtiD. schoenigiD. viduaD. themisD. kuehniCeporaLeuciacriaAporiaPrioneris1Prioneris2Eurema
WALLACEAN
DIVERSIFICATION
WA
LL
AC
EA
Muller & Beheregaray (in review) J Biogeog

CethosiaO
rie
nta
lW
alla
ce
a
vasilia
bernsteini
hypsea
cyane penthesilea
obianabiblis
luzonica picta
myrinaobscura lamarcki
vasilialechenaulti

Orie
nta
lCharaxes
Walla
ce
a
amycus distantiantonius
harmodius latona
marmaxbajula kahrubra aristogiton schaderi
mars eurialus markisetan
ocellatus sulaensis nitebis orilus madensis

Long term isolation of Wallacean taxa was promoted by the progressive fragmentation of a micro-continent
Expanding Wallacea Theory

A Few Perspectives and ChallengesDo we have ‘virtual phylogeographers’ out there or is it just my imagination?
• computationally intensive, model driven researchers• use lots of markers and coalescence theory
• appear more interested in the theory than study system• good natural history experience?
• observation of Nature is the source of our hypotheses• never dump the biology! • without that the parameters that go into models are just guesswork,
and the interpretation may be equally blind• look for better ways to synergistically combine natural history and
computational biology
Statistically rigorous phylogeography is important, but what really matters are the quality of research questions

• Intellectual maturation of Phylogeography?
Recent developments in DNA technology, theory and statistics AND syntheses of comparative information across different regions of the globe
• Create strategies for developing in situ capacity
A Few Perspectives and Challenges
• A lot of the natural history has not been done yet: many more descriptive phylogeographic surveys are needed in the developing world

Acknowledgements
• MELFU PhD students and postdocs (current and alumni)
• Louis Bernatchez (Laval U, Canada)
• Paul Sunnucks (Monash U, Australia)
• Jeff Powell and Gisella Caccone (Yale U, USA)
• Luciana Moller (Flinders U, Australia)
Financial Support