a novel small protein of bacillus subtilis involved in spore germination...
TRANSCRIPT

A Novel Small Protein of Bacillus subtilis Involvedin Spore Germination and Spore Coat Assembly
Takeko KODAMA,1;2 Takeshi MATSUBAYASHI,2 Tadayoshi YANAGIHARA,2
Hiroyuki KOMOTO,2 Katsutoshi ARA,1 Katsuya OZAKI,1 Ritsuko KUWANA,3
Daisuke IMAMURA,3 Hiromu TAKAMATSU,3 Kazuhito WATABE,3 and Junichi SEKIGUCHI2;y
1Biological Science Laboratories, Kao Corporation, 2606 Akabane, Ichikai, Haga, Tochigi 321-3497, Japan2Department of Bioscience and Textile Technology, Interdisciplinary Graduate School of Science and Technology,Shinshu University, 3-15-1 Tokida, Ueda, Nagano 386-8567, Japan3Faculty of Pharmaceutical Sciences, Setsunan University, Hirakata, Osaka 573-0101, Japan
Received January 13, 2011; Accepted March 15, 2011; Online Publication, June 13, 2011
[doi:10.1271/bbb.110029]
Two small genes named sscA (previously yhzE) andorf-62, located in the prsA-yhaK intergenic region of theBacillus subtilis genome, were transcribed by SigK andGerE in the mother cells during the later stages ofsporulation. The SscA-FLAG fusion protein was pro-duced from T5 of sporulation and incorporated intomature spores. sscA mutant spores exhibited poorgermination, and Tricine–SDS–PAGE analysis showedthat the coat protein profile of the mutant differed fromthat of the wild type. Bands corresponding to proteins at59, 36, 5, and 3 kDa were reduced in the sscA nullmutant. Western blot analysis of anti-CotB and anti-CotG antibodies showed reductions of the proteins at59 kDa and 36 kDa in the sscA mutant spores. Theseproteins correspond to CotB and CotG. By immunoblotanalysis of an anti-CotH antibody, we also observed thatCotH was markedly reduced in the sscA mutant spores.It appears that SscA is a novel spore protein involved inthe assembly of several components of the spore coat,including CotB, CotG, and CotH, and is associated withspore germination.
Key words: Bacillus subtilis; small protein; spore coat;germination; sscA
Bacillus subtilis, a nonpathogenic, spore-formingbacterium normally found in soil, is the best charac-terized of the Gram-positive bacteria. Its genomesequence was reported by Kunst et al. in 1997.1) Atthat time, approximately 4,100 genes were thought tocomprise the B. subtilis genome. These protein-codinggenes constitute 87% of the genome, a percentage that isvery similar to other bacterial genomes, such as that ofEscherichia coli.2) Recent updates to the sequence andannotation repository are available.3) In the last decade,a wealth of additional genes containing small regulatoryRNAs (sRNAs or ncRNAs for non-coding RNAs) havebeen discovered in both Gram-positive and Gram-negative bacteria.4–7) In addition, small ORFs arealso of interest for their functions in microbial physiol-ogy.8–12) Several genes for small ORFs that are tran-scribed in response to sporulation have provided newbiological insights.9,13–15) A 40-amino-acid peptide,
MciZ, acts to block Z-ring formation during sporulationby inhibiting the polymerization of FtsZ in the mothercell.9) ytrH was found to be a novel sporulation geneinvolved in cortex formation and stability.13) Schmalischet al.14) reported that six paralogous genes were newlyidentified small proteins under sporulation control.Mutants of these genes had no significant deleteriouseffects on the production of heat-resistant spores, but amultiple mutant lacking six paralogous genes showedsporulation efficiency slightly higher than the wild type.Sporulation-specific yjcZ encodes a 49-aa small poly-peptide15) (Fig. 7). YjcZ is a conserved protein inBacillus, but yjcZ mutant spores showed resistance toheat and lysozyme similar to those of the wild type.15)
Endospores of B. subtilis are encased in a thickprotein shell known as the spore coat. The coat’scomplex structure comprises an inner coat (IC) and anouter coat (OC), and includes more than 70 spore-specific proteins.16,17) The coat plays several roles in thespore’s resistance to various chemicals and to exogenouslytic enzymes that can degrade the spore cortex. It alsoregulates spore germination.17–19) The functions of mostindividual coat proteins are not known, although a fewhave been found to be morphogenic and to be involvedin overall spore coat assembly. These include CotE,SafA (YrbA), SpoIVA, and SpoVID.17,18) The coatprotein SpoIVA plays a primary role, as it appears toattach the coat to the outer of the forespore membranesand to be required for the formation of the cortex.20–22)
SpoVID, which assembles under the control of SpoIVA,is also required for attachment of the coat to theforespore.21,23) SafA is required for proper coat assem-bly, and interaction between SafA and SpoVID isessential for the localization and function of SafA.24,25)
CotE directs the assembly of a large subset of coatproteins, including most or all of the outer coatproteins.21,26–28) CotH, one of the so-called CotE-controlled proteins, directs the assembly of an importantsubset of coat proteins, including CotG. CotG in turncontrols the deposition of CotB.29,30)
In this study, we identified a sporulation-specific gene,sscA (small spore coat assembly protein, previouslyknown as yhzE), that is located in the prsA-yhaK
y To whom correspondence should be addressed. Tel/Fax: +81-268-21-5344; E-mail: [email protected]
Biosci. Biotechnol. Biochem., 75 (6), 1119–1128, 2011

intergenic region on the B. subtilis chromosome. Anadditional ORF, also found to be under sporulation control,was located downstream of the sscA gene, and we namedthis orf-62. Both SscA and ORF-62 have sequencesimilarity to YjcZ (Fig. 7). We report that spores of thesscA null mutant showed partial lack of assembly ofseveral coat components, including CotB, CotG, andCotH, and impairment of response during germination.
Materials and Methods
Bacterial strains, plasmids, and culture conditions. The B. subtilis
and Escherichia coli strains used in this study are listed in Table 1. The
B. subtilis strains were grown in Luria-Bertani (LB) medium (1%
tryptone, 0.5% yeast extract, and 1% NaCl) and DS medium.31)
Antibiotics were added to media at the following concentrations:
0.3 mg/mL of erythromycin, 50 mg/mL of spectinomycin, 5mg/mL of
chloramphenicol, and 10mg/mL of kanamycin. They were cultured in
LB medium at 30 �C for 15 h, and 1mL of culture broth was added to
200mL of DS medium in a Sakaguchi flask. The cells were cultured
for sporulation at 37 �C. The E. coli strains were used as the hosts for
plasmid construction and were grown in LB medium supplemented
with 50mg/mL of ampicillin.
Construction of sscA mutants. The primers used in this study are
listed in Table 2. The 50 and 30 regions of sscA (fragments 1 and 2)
were amplified using B. subtilis 168 chromosomal DNA and two pairs
of primers (61up+379F and 61up+379R for fragment 1, and
61dn+410F and 61dn+410R for fragment 2). The spectinomycin
resistance gene was amplified using pBlueSPR32) and primers PB-
M13-20 and PB-M13Rev (fragment 3). The 50-end sequences of
primers 61up+379R and 61dn+410F are complementary to those of
primers PB-M13-20 and PB-M13Rev respectively. Fragment 4 was
amplified using a mixture of fragments 1, 2, and 3 as templates, and
two primers (61up+379F and 61dn+410R). Fragment 4 was inserted
into B. subtilis 168 by double crossover recombination. After selection
for spectinomycin resistance (Spcr), the transformants were confirmed
by PCR analysis using primers 7 and 8. After confirmation of the
nucleotide sequences of the DNA regions used in homologous
recombination (fragments 1 and 2), the resulting mutant (sscA::spc)
was named �sscA. �62 (orf-62::spc) was constructed similarly to
�sscA. The 50 and 30 regions of cotB (fragments 1 and 2) were
amplified using B. subtilis 168 chromosomal DNA and two pairs of
primers (CotB�19F and CotB+506Rem for fragment 1, and
CotB+521Fem and CotB+1140R for fragment 2). The erythromycin
resistance gene was amplified using pMutin433) and primers Em-F5 and
Em-R5 (fragment 3). The 50-end sequences of primers CotB+506Rem
and CotB+521Fem are complementary to those of primers Em-F5 and
Em-R5 respectively. Fragment 4 was amplified using a mixture of
fragments 1, 2, and 3 as templates and two primers (CotB�19F and
CotB+1140R). Fragment 4 was inserted into B. subtilis 168 by double
crossover recombination. After selection for erythromycin resistance
(Emr), the transformants were confirmed by PCR analysis using
primers 7 and 8. The resulting �cotB mutant (cotB::erm) was
generated. �cotE, �cotG, �cotH, and �gerE mutants were con-
structed similarly to �cotB. The nucleotide sequences within the
�cotE, �cotG, and �gerE mutants used for homologous recombina-
tion (fragments 1 and 2) were confirmed. The region of sscA containing
the SigK consensus sequence was prepared by PCR with two primers
(sscA�141F-SmaI and sscA+244R-XbaI). The PCR fragment was
digested with SmaI and XbaI, and then ligated to the corresponding
pDHAFBLZ site34) to generate pDHAFBLZsscA. Then the nucleotide
sequence of the cloned DNA region pDHAFBLZsscA was confirmed.
This plasmid (linearized with PstI) was inserted into the amyE locus in
the B. subtilis 168 chromosome by means of double crossover
recombination. After selection for Cmr, the transformants were
confirmed by PCR analysis using primers pDHLZF1 and amyE+
2160R, and then C61 (sscA::spc, amyE::PsscA-sscA) was generated.
Table 1. Bacterial Strains and Plasmids Used in This Study
Strains Genotype Source or reference
Bacillus subtilis
168 trpC2 Laboratory stock
1S86 trpC2 spoIIA1 BGSCa
1S60 leuB8 tal-1 spoIIG41 BGSC
SpoIIIG�1 trpC2 spoIIIG�1 BGSC
1S38 trpC2 spoIIIC94 BGSC
�sscA trpC2 sscA::spc This study
�62 trpC2 orf-62::spc This study
CsscA trpC2 sscA::spc, amyE::PsscA-sscA This study
�cotB trpC2 cotB::erm This study
�cotE trpC2 cotE::erm This study
�cotG trpC2 cotG::erm This study
�cotH trpC2 cotH::erm This study
�gerE trpC2 gerE::erm This study
SscA-FLAG trpC2 sscA�pMUTsscA-FLAG (erm) This study
168 (pDGPsscA-FLAG) trpC2 harboring pDGPsscA-FLAG This study
Escherichia coli
JM109 recA1 endA1 gyrA96 thi-1 hsdR17 Laboratory stock
supE44 relA1�(lac-proAB)/F0
(traD36 proABþ lacIq, lacZ�M15)
C600 supE44 hsdR17 thi-1 thr-1 leuB6 lacY1 tonA21 Laboratory stock
Plasmids
pDHAFBLZ cat bla lacZ lacI 34
pDHAFBLZ61 cat bla lacZ lacI PsscA-sscA This study
pMUTIN4 erm bla lacZ lacI Pspac 33
pMUTsscA-FLAG erm bla lacZ lacI PspacPsscA-sscA This study
pDG148 kan bla Pspac 35
pDGPsscA-FLAG kan bla Pspac-PsscA-sscA-flag This study
pMALEH6 bla lacI tac promoter malE His6 36
pMCOTB1 bla lacI tac promoter malE cotB His6 This study
aBGSC, Bacillus Genetic Stock Center
1120 T. KODAMA et al.

Construction of an SscA-FLAG fusion strain. To construct an SscA-
FLAG fusion strain, sscA with a flag sequence at its 30-terminus was
constructed as follows: The region from �296 bp to þ84 bp of the sscA
sequence containing the P1 promoter was amplified by PCR with two
primers (61�246F and 61þ132FLAGR). The reverse primer contained
the flag tag, and SscA was fused with the FLAG sequence at its
C-terminal. The fragment was digested with HindIII and BamHI, and
ligated into pMUTIN433) to generate pMUTsscA-FLAG. The plasmid
from E. coli C600 (pMUTsscA-FLAG) was used in the transformation
of B. subtilis 168 to obtain the SscA-FLAG strain through Campbell-
type integration. Therefore the SscA-FLAG strain had a 1� FLAG
epitope tag (DYKDDDDK) at the C terminus of SscA and also intact
SscA in the chromosome.
Table 2. Primers Used in This Study
Primer Sequence (50 ! 30)a Restriction site
61up+379F GGCAGTATATTGATCGCC
61up+379R CACTGGCCGTCGTTTTACTGACTAGACGAATGCCCT
61dn+410F CATGGTCATAGCTGTTTCCGCTTATCATCGTAGGTGC
61dn+410R GCCTTGTCAGAGAGATTC
61UP�Fow CTTCCCAGTACTCTTTGA
61DN�Rev TATGGGACAGTTCAGAGA
62up+434F TGTAAAGCTCGCCTTTTG
62up+434R CACTGGCCGTCGTTTTACCAAACGATTAGGCGCTAGT
62dn+416F CATGGTCATAGCTGTTTCCAATCTCTCTGACAAGGCA
62dn+416R ATGCTGATTTCACTCGTC
62DN�Rev ATGTCCTGCCAATCATCT
PB-M13-20 GTAAAACGACGGCCAGTG
PB-M13Rev GGAAACAGCTATGACCATG
PB-SK-Fow TCCACTAGTTCTAGAGCG
PB-T7-Rev TTCGCCCTATAGTGAGTC
CotB�19F GTGACTAGGAGGAATTTG
CotB+506Rem CTCTTAAGTTTGCTTCTAAGTCTTCTTAAATCATCAGCTTCC
CotB+521Fem GGAAAGTTACACGTTACTAAAGGGGTCTGACTCATAAATGGG
CotB+1140R AAATTTACGTTTCCAGTG
CotE�509F ATGAAAAGCATGAAGCGC
CotE�2Rem GGAAAGTTACACGTTACTAAAGGGCCCTGTATTCAGACATTC
CotE+529Fem CTCTTAAGTTTGCTTCTAAGTCTTGGAGATCCTGAAGAATAA
CotE+1040R TCATGGATGCCTTTGCCG
CotE�559F CTGAAGCGCAAATCAATG
CotE+1118R CCCGTTGTTAAATCAGAC
CotG�402F AAGGGTCTTTATACTCCG
CotG+118Rem CTCTTAAGTTTGCTTCTAAGTCTTGCGACTTTTTATGACTGC
CotG+503Fem GGAAAGTTACACGTTACTAAAGGGACTACAAAAGACACGATG
CotG+1022R ATCACTGACATGAACGAC
CotG�514F CAAACTCTGCCTTTGGAG
CotG+1022R ATCACTGACATGAACGAC
CotH�3F TGGATGAAGAATCAATCC
CotH+521Rem CTCTTAAGTTTGCTTCTAAGTCTTTCATATCCAAGCTCCAGC
CotH+534Fem GGAAAGTTACACGTTACTAAAGGGGACTGAGGAAGATGATTT
CotH+1040R TACTCGCAGATCACATCC
GerE�420F AGGAGGTGCGGATTTCTT
GerE+78Rem CTCTTAAGTTTGCTTCTAAGTCTTATCTTGAACGAGCAATTC
GerE+121Fem GGAAAGTTACACGTTACTAAAGGGAAAACCGTTCGAAACCAT
GerE+636R ACCGATTGTCATATCTCC
GerE�535F ATGCCGTTTCAAGAATGG
GerE+760R GATGTATCCTGACAACAC
Em-F5 AAGACTTAGAAGCAAACTTAAGAG
Em-R5 CCCTTTAGTAACGTGTAACTTTCC
61�141F-SmaI ATGACCCGGGAGTGATAGCTGCTATTGC SmaI
61+244R-XbaI ATGCTCTAGACATCTTAAATAGCACCTC XbaI
pDHLZF1 CCTCAGGAAGATCGCACT
amyE+2160R TTAATCATCCTTGCAGGG
61�246F GCATAAGCTTCATGTTTGTGTAAAGCTC HindIII
61+132FLAGR GCCGGGATCCTTACTTGTCGTCATCGTCCTTGTAGTC BamHI
GTAGATGTAAGCTGCACC
61�141F-salI ATACGTCGACAGTGATAGCTGCTATTGC SalI
61+132FLAGR2 GCCGGCATGCTTACTTGTCGTCATCGTCCTTGTAGTC SphI
GTAGTAGTAAGCTGCACC
61-Fe GCGCGAATTCCTGATGACATACTTATATATTG EcoRI
61-Rb gtaatacgactcactatagggcGGATCCGTAGATGTAAGCTGCACCT BamHI
62-Fe GCGCGAATTCCTATTTAAGATGGGTGAAGTA EcoRI
62-Rb gtaatacgactcactatagggcGGATCCACAGCCAAGAAGCGCCG BamHI
COTB3 AATGGATCCGAGCAAGAGGAGAATGAAAT BamHI
COTB726R TCACTCGAGTCTTCCTGGTTTTGTTCAT XhoI
aRestriction sites (italics), PCR ligation regions (underlined), FLAG epitope tag coding sequences (double underlined), and T7 RNA polymerase
recognition sequences (lowercase) are indicated.
Small Spore Protein Involved in Coat Assembly 1121

Construction of pDGPsscA-FLAG. The region of sscA containing
the SigK consensus sequence was prepared by PCR with two primers,
61�141F-SalI and 61þ132FLAGR2. The PCR fragment was digested
with SalI and SphI and then ligated to the corresponding pDG148 site35)
to generate pDGPsscA-FLAG. After confirmation of the nucleotide
sequence of the cloned DNA region in pDGPsscA-FLAG, the 168 strain
was transformed using pDGPsscA-FLAG from E. coli C600. This
resulted in a kanamycin-resistant strain, 168 (pDGPsscA-FLAG).
Construction of pMCOTB1. To construct a plasmid, DNA frag-
ments encoding CotB were PCR-amplified from B. subtilis 168
genomic DNA using primers COTB3 and COTB726R. The PCR
products were digested with restriction enzymes, and were inserted
into BamHI/XhoI-digested pMALEH6 (this contained a 6� His-tag)
to generate recombinant plasmid pMCOTB1 (Table 1).36)
RNA preparation and Northern blot analysis. Cells were grown in
DS medium at 37 �C, and an aliquot was collected by centrifugation.
Preparation of total RNA was performed as described previously.37,38)
Aliquots (10mg) of the total RNA were electrophoresed and blotted on
a positively charged Hybond-N+membrane (GE Healthcare, Japan).
Hybridizations were performed with digoxigenin-labeled RNA probes
(10 ng) following the manufacturer’s instructions, with minor mod-
ifications (Roche Diagnostics, Japan). Templates for sscA and orf-62
specific probes were constructed by PCR. sscA and orf-62 fragments
were amplified from B. subtilis 168 chromosomal DNA using primers
61-Fe and 61-Rb, 62-Fe and 62-Rb respectively. The 50-end sequences
of 61-Rb and 62-Rb contained the T7 RNA polymerase recognition
site. Hybridization and detection were performed using a DIG
luminescent detection kit (Roche Diagnostics), following the manu-
facturer’s instructions. The primers used in this study are listed in
Table 2.
Preparation of spores, spore resistance, and spore germination.
B. subtilis strains were cultured in DSM at 37 �C until T48 (48 h after
the onset of sporulation). The spores were washed until all cell debris
and vegetative cells had been removed completely.39,40) The cells were
grown in DS medium at 37 �C for 24 h after the end of exponential
growth, and spore resistance was assayed as follows: Cultures were
heated at 80 �C for 30min, treated with lysozyme (final concentration,
0.25mg/mL) at 37 �C for 10min or treated with 10% v/v chloroform
at room temperature for 10min as described previously,41) diluted in
distilled water, plated on LB agar, and incubated overnight at 37 �C.
The numbers of survivors were determined by counting colonies. The
spores were heat-activated at 80 �C for 20min before germination,
unless otherwise noted, and then diluted with 10mM Tris–HCl buffer
(pH 8.4). Germination was initiated by the addition of L-alanine at
a final concentration of 10mM or AGFK, L-asparagine, D-glucose,
D-fructose, and KCl, at final concentrations of 10, 1, 1, and 10mM
respectively. Germination was monitored by measurement of the
decrease in the OD580 of the mixture (initial OD580 ¼ 0:3, approx-
imately 4� 107 spores) at 37 �C for up to 180min.
Measurement of dipicolinic acid. The amounts of dipicolinic acid
released from the spores during germination were determined.39) The
supernatant (1mL) was used to measure released dipicolinic acid, as
described by Nicholson and Setlow.41)
Solubilization of proteins from sporulating cells and mature spores.
The B. subtilis strains were cultured as described above. To prepare
protein-containing extracts from sporulating cells, we collected cells (at
approximately 2 OD600) from 1mL of culture medium by centrifuga-
tion, dissolved the resulting pellets in 1mL of wash buffer (25mM Tris–
HCl (pH 7.2)), and collected the cells by centrifugation again. Pellets
were suspended in 0.1mL of lysozyme solution (0.4mg/mL of
lysozyme, 25mM Tris–HCl pH 8.0, 50mM glucose, and 10mM EDTA)
containing phenylmethane sulfonyl fluoride (PMSF, 1mM), incubated
for 5min at 37 �C, solubilized in a Tricine-loading buffer (1X Novex�
Tricine SDS sample buffer, Invitrogen, Japan, and 1X NuPAGE�
reducing agent, Invitrogen), and heated at 85 �C for 2min. To prepare
proteins from mature spores, spores (approximately 10 OD600) were
harvested from 5mL of culture medium at 24 h after the cessation of
exponential growth (T24) and washed with 1mL of 25mM Tris–HCl
(pH 7.2) buffer. After centrifugation, the pellets were suspended in
0.1mL of lysozyme solution containing 1mM PMSF, incubated for
5min at 37 �C, and then washed with a wash buffer (25mM Tris–HCl
(pH 7.2), 0.5M NaCl). The spore proteins were solubilized in Tricine-
loading buffer and heated at 85 �C for 2min. The resulting samples
were analyzed by Tricine–SDS–PAGE, as described below.
Tricine–SDS–PAGE and Western blot analysis. The protein samples
were analyzed by Tricine–SDS–PAGE (10–20% Tricine gel, Invitro-
gen). For the details of the procedure, refer to the Novex Pre-Cast Gel
Electrophoresis Guide, available at www.invitrogen.com. For Western
blot analysis, proteins on Tricine–SDS–PAGE gels were transferred to
polyvinylidene difluoride membranes (Immobilon; 0.45 mm pore size;
Milipore, Tokyo). Proteins were detected using an ECL Plus Western
blotting detection system (GE Healthcare). We used an anti-FLAG M2
monoclonal antibody produced in mice (Sigma-Aldrich Japan, Tokyo),
an anti-CotB, -CotG,36) -CotH,36) and -YaaH36) polyclonal antibody
produced in rabbit as primary antibody, and horseradish-peroxidase-
conjugated donkey anti-mouse IgG (against anti-FLAG M2 antibody)
or donkey anti-rabbit (against anti-CotB, -CotG, -CotH, or -YaaH
antibody) as secondary antibody (GE Healthcare). Antisera were
diluted to 1/5,000 with 20mM Tris–HCl (pH 7.6) buffer containing
0.8% NaCl and 0.3% Tween 20.
Purification of recombinant MalE-CotB from E. coli. E. coli cells
were transformed with recombinant plasmids pMCOTB1. The result-
ing transformants were grown at 37 �C for 3 h in 200mL of LB
supplemented with ampicillin (50 mg/mL). The cultures were then
supplemented with 1mM isopropyl-�-D-thiogalactopyranoside and
incubated for an additional 3 h at 37 �C. His-tagged recombinant
protein MalE-CotB was purified by affinity chromatography using
Ni-NTA agarose beads (Qiagen, Hilden, Germany), and was further
purified by electro-elution from the resulting SDS–PAGE, as previ-
ously described.42)
Preparation of antisera against MalE-CotB. We prepared antisera
against MalE-CotB as follows: The purified proteins (1mL of 0.2
mg/mL) and 16mg of killed Mycobacterium tuberculosis cells (BD
Difco, Japan) were mixed with 2mL of complete Freund’s adjuvant
(BD Difco). Three mL of each emulsion was injected into healthy
rabbits. After 2 weeks, the rabbits were injected with solutions of the
purified proteins in incomplete Freund’s adjuvant (BD Difco). At 2
weeks after the second immunization, antiserum was isolated from the
rabbits as previously described.42)
Results
Transcriptional analysis of sscA and orf-62sscA has been predicted within the yhaK-prsA inter-
genic region of the B. subtilis chromosome.3) We foundan additional small ORF encoding a 28-aa polypeptidein the region between sscA and yhaK, and named it orf-62 (Fig. 1A). By Northern blot, we determined the sizesand times of appearance of the sscA and orf-62 mRNAsduring growth and sporulation. Two sizes of transcripts,0.35 kb and 0.15 kb, were detected, beginning in the T4
cells, by the sscA-specific probe (Fig. 1B). orf-62mRNA of 0.35 kb and 0.15 kb were detected beginningin the T4 and T5 cells respectively (Fig. 1C). No sscA ororf-62 mRNAs were detected at T6 within the sporula-tion sigma factor-deficient mutants or the gerE-deficientmutant (Fig. 1B and C). Our results indicate that theexpression of sscA and orf-62 was dependent on bothSigK RNA polymerase and GerE in the mother-cellcompartment. The nucleotide sequences upstream ofsscA and orf-62 showed similarity to the consensussequence, the �35 ((c/a)AC(c/a)) and �10 (CATA---TA), of the SigK RNA polymerase-dependent promoter
1122 T. KODAMA et al.

(Fig. 1A). Two putative GerE-binding sites were presentin the upstream regions of the sscA and orf-62 promoters(Fig. 1A). Both the 0.35 kb and the 0.15 kb mRNAshown in Fig. 1B were probably derived from the sscApromoter (P1). The smaller transcript ended at termi-nator T1, and the larger one was read through ending atterminator T2. The 0.15-kb mRNA was detected only inthe orf-62 mutant (Fig. 1B, lane 14). Since the mutantwas constructed by replacement of orf-62 containing theT2 terminator with the spectinomycin resistance gene,the sscA transcript from P1 to T2 was not detected in themutant. The orf-62 mRNAs (0.35 kb and 0.15 kb) andthe probe for orf-62 are shown in Fig. 1C. Thesetranscripts were probably derived from the sscA pro-moter (P1) and the orf-62 promoter (P2), and bothtranscripts ended at terminator T2. The 0.15-kb mRNAwas detected only in the sscA mutant (Fig. 1C, lane 13).Since this mutant was constructed by replacement ofsscA containing the P1 promoter with the spectinomycinresistance gene, the orf-62 transcript from P1 to T2 wasnot detected in the mutant. These results indicate thatsscA and orf-62 were transcribed from distinct promot-ers, P1 and P2, and that the transcripts ended at
terminators T1 and T2. These genes were also tran-scribed as operons from the P1 promoter to terminatorT2 (Fig. 1A, top), but the P2 promoter was much weakerthan the P1 promoter (Fig. 1B and C).
Properties of mutant sporesWe characterized the sscA and orf-62 mutant cells.
The vegetative growth of the mutant cells in DS mediumwas the same as that of the wild type (data not shown).Mature spores prepared from 24 h-cultures at 37 �C inDS medium exhibited resistance to heat, chloroform,and lysozyme (data not shown). During germination inbuffer containing the germination stimulant L-alanine,the sscA mutant spores exhibited a smaller decrease inA580 than the wild-type spores (Fig. 2A). The absorb-ance of the sscA mutant spores placed in AGFK toenhance germination exhibited a slower decrease until90min, but was similar to the wild-type spores after120min (Fig. 2B). sscA is followed by the orf-62 gene,and the two genes might be co-transcribed from the P1promoter during sporulation (Fig. 1). Hence we tested todetermine whether orf-62 has any role in germination.We found no differences in germination between
A
B
C
Fig. 1. Transcriptional Analysis of sscA and orf-62.A, Arrangement of genes around the prsA and yhaK loci on the B. subtilis genome. At the top in A are sizes of transcripts estimated by
prospective regions of promoters for sscA (P1) and orf-62 (P2) and terminators (T1, T2). At the bottom are nucleotide and deduced amino acidsequences of sscA and orf-62. The deduced amino acid sequences of SscA and ORF-62 are marked below the nucleotide sequences. Asterisksindicate stop codons. The nucleotide sequence of each gene promoter, the putative �35 and �10 regions, and the transcriptional start site (þ1)are shown within the box. These sequences are similar to the consensus sequences for SigK RNA polymerase, �35 ((c/a)AC(c/a)) and �10
(CATA---TA). Arrows indicate putative GerE-binding sites (consensus sequence, RWWTRGGY---YY; R, purine; W, A or T; Y, pyrimidine).Potential ribosomal binding sites (RBSs) are underlined. Dotted inverted arrows are potential stem-loop structures acting as transcriptionalterminators. B and C, Analysis of sscA and orf-62 mRNAs respectively by Northern hybridization. Total RNA was prepared from sporulatingcells, and mRNAs were detected by Northern blotting. T = harvesting times of cells, i.e., hours after the end of the exponential phase of growth.Lanes 1 to 7, 168; lane 8, spoIIAC (SigF�); lane 9, spoIIGAB (SigE�); lane 10, spoIIIG (SigG�); lane 11, spoIVCB (SigK�); lane 12, �gerE;lane 13, �sscA; lane 14, �62. Approximate positions of transcripts and sizes of RNAs are shown by arrowheads.
Small Spore Protein Involved in Coat Assembly 1123

L-alanine and AGFK, used as stimulants (Fig. 2A). Inaddition, the CsscA strain containing the sscA gene inthe amyE locus was constructed from germination-deficient �sscA. CsscA showed full recovery fromgermination deficiency. This indicates that the decreasein germination efficiency was due to disruption of sscA.
We also measured the amounts of dipicolinic acid inthe sscA mutant spores during germination in thepresence of L-alanine and AGFK (Fig. 3). The level ofdipicolinic acid released from sscA mutant spores in thepresence of L-alanine was low through the incubationperiod, and less than 50% of that released from the168spores at 180min (Fig. 3A). When AGFK was used asthe stimulant, the sscA mutant spores released dipico-linic acid at slightly lower levels, and eventually insimilar quantities to the 168 spores (Fig. 3B). Usingphase-contrast microscopy, we confirmed that almostall of the wild-type, orf-62 mutant, and CsscA sporesbecame dark (full germination) after 180min of incu-bation with L-alanine. In contrast, only some sscAmutant spores became phase gray or dark (partialgermination) under the same conditions (data notshown). This suggests that sscA is required for theprogress of spore germination stimulated by L-alanine,and that it also responds to AGFK during the earlygermination period.
Detection of the SscA-FLAG tag fusion in sporulatingcells and mature spores
To investigate the synthesis and location of the SscAprotein in sporulating cells, we constructed the SscA-FLAG strain, which expressed the SscA-FLAG fusion
protein as a single copy from the original promoter andribosomal binding site. This strain is assumed to produceboth the SscA-FLAG fusion protein and the intact SscAprotein, as described in ‘‘Materials and Methods.’’ Wetried to detect SscA-FLAG using FLAG-tag specificantiserum and Western blot, but we could not detect thefusion protein (data not shown). This failure might havebeen due to the low level of expression of sscA or todegradation of the FLAG protein. We constructedplasmid pDGPsscA-FLAG, in which SscA-FLAG wasexpressed from the original promoter and ribosomalbinding site. We overproduced it in E. coli and B. sub-tilis. SscA-FLAG from cultures of the 168 (pDGPsscA-FLAG) strain at T3 to T6 and T24 were analyzed byWestern blot with the anti-FLAG antibody. SscA-FLAGwas produced from T5, and the expression pattern wasconsistent with the results of Northern blot analysis, asdescribed above. We also found that SscA-FLAGaccumulated in mature spores of 168 (pDGPsscA-FLAG) at T24 (Fig. 4). A band with a molecular massof 4 kDa, corresponding to SscA-FLAG, was detected
A
B
Fig. 2. Spore Germination of B. subtilis 168,�sscA,�62, and CsscA.Spore germination was monitored at 580 nm after the addition of
L-alanine (A) or AGFK (B) as germination stimulant. Spores of 0.3OD580 from each strain were used for germination. The efficiency ofgermination is expressed as relative absorbance. Hollow circles,168; solid circles, �sscA; hollow triangles, �62; solid triangles,CsscA. Results represent averages of three individual experiments.Error bars correspond to standard error of the means (SEM).
A
B
Fig. 3. Dipicolinic Acid (DPA) Released from B. subtilis 168 and�sscA Spores during Germination.
Released DPA was measured after the addition of L-alanine (A) orAGFK (B) as stimulant. Hollow circles, 168; solid circles, �sscA.Spores of 0.3 OD580 from each strain were used in measurement ofDPA. Results represent averages of three individual experiments.Error bars correspond to standard error of the means (SEM).
Fig. 4. Western Blot of SscA-FLAG.Western blot was carried out to detect SscA-FLAG in the 168
(pDGPsscA-FLAG) strain with the anti-FLAG antibody. T, harvest-ing times of cells, i.e., hours after the end of the exponential phase ofgrowth. Proteins from cells (0.3 OD600 units) were prepared from168 (pDGPsscA-FLAG) (lanes 1–5) and 168 (pDG148) (lanes6–10), as described in ‘‘Materials and Methods.’’ The arrowheadindicates the position of SscA-FLAG.
1124 T. KODAMA et al.
in the protein extract from mature spores of 168(pDGPsscA-FLAG) but not in that from the 168(pDG148) spores (Fig. 4). This suggests that SscA is aspore protein that is synthesized during the later stagesof sporulation and is incorporated into the spores.
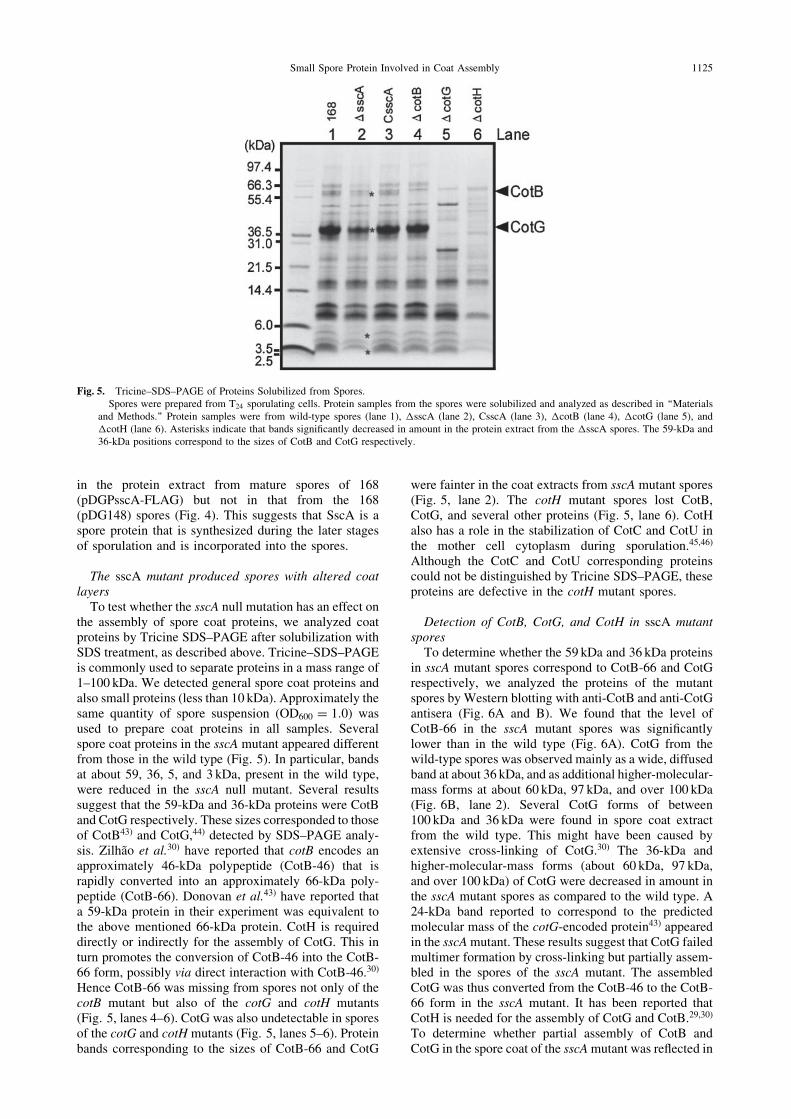
The sscA mutant produced spores with altered coatlayers
To test whether the sscA null mutation has an effect onthe assembly of spore coat proteins, we analyzed coatproteins by Tricine SDS–PAGE after solubilization withSDS treatment, as described above. Tricine–SDS–PAGEis commonly used to separate proteins in a mass range of1–100 kDa. We detected general spore coat proteins andalso small proteins (less than 10 kDa). Approximately thesame quantity of spore suspension (OD600 ¼ 1:0) wasused to prepare coat proteins in all samples. Severalspore coat proteins in the sscA mutant appeared differentfrom those in the wild type (Fig. 5). In particular, bandsat about 59, 36, 5, and 3 kDa, present in the wild type,were reduced in the sscA null mutant. Several resultssuggest that the 59-kDa and 36-kDa proteins were CotBand CotG respectively. These sizes corresponded to thoseof CotB43) and CotG,44) detected by SDS–PAGE analy-sis. Zilhao et al.30) have reported that cotB encodes anapproximately 46-kDa polypeptide (CotB-46) that israpidly converted into an approximately 66-kDa poly-peptide (CotB-66). Donovan et al.43) have reported thata 59-kDa protein in their experiment was equivalent tothe above mentioned 66-kDa protein. CotH is requireddirectly or indirectly for the assembly of CotG. This inturn promotes the conversion of CotB-46 into the CotB-66 form, possibly via direct interaction with CotB-46.30)
Hence CotB-66 was missing from spores not only of thecotB mutant but also of the cotG and cotH mutants(Fig. 5, lanes 4–6). CotG was also undetectable in sporesof the cotG and cotH mutants (Fig. 5, lanes 5–6). Proteinbands corresponding to the sizes of CotB-66 and CotG
were fainter in the coat extracts from sscA mutant spores(Fig. 5, lane 2). The cotH mutant spores lost CotB,CotG, and several other proteins (Fig. 5, lane 6). CotHalso has a role in the stabilization of CotC and CotU inthe mother cell cytoplasm during sporulation.45,46)
Although the CotC and CotU corresponding proteinscould not be distinguished by Tricine SDS–PAGE, theseproteins are defective in the cotH mutant spores.
Detection of CotB, CotG, and CotH in sscA mutantsporesTo determine whether the 59 kDa and 36 kDa proteins
in sscA mutant spores correspond to CotB-66 and CotGrespectively, we analyzed the proteins of the mutantspores by Western blotting with anti-CotB and anti-CotGantisera (Fig. 6A and B). We found that the level ofCotB-66 in the sscA mutant spores was significantlylower than in the wild type (Fig. 6A). CotG from thewild-type spores was observed mainly as a wide, diffusedband at about 36 kDa, and as additional higher-molecular-mass forms at about 60 kDa, 97 kDa, and over 100 kDa(Fig. 6B, lane 2). Several CotG forms of between100 kDa and 36 kDa were found in spore coat extractfrom the wild type. This might have been caused byextensive cross-linking of CotG.30) The 36-kDa andhigher-molecular-mass forms (about 60 kDa, 97 kDa,and over 100 kDa) of CotG were decreased in amount inthe sscA mutant spores as compared to the wild type. A24-kDa band reported to correspond to the predictedmolecular mass of the cotG-encoded protein43) appearedin the sscAmutant. These results suggest that CotG failedmultimer formation by cross-linking but partially assem-bled in the spores of the sscA mutant. The assembledCotG was thus converted from the CotB-46 to the CotB-66 form in the sscA mutant. It has been reported thatCotH is needed for the assembly of CotG and CotB.29,30)
To determine whether partial assembly of CotB andCotG in the spore coat of the sscAmutant was reflected in
Fig. 5. Tricine–SDS–PAGE of Proteins Solubilized from Spores.Spores were prepared from T24 sporulating cells. Protein samples from the spores were solubilized and analyzed as described in ‘‘Materials
and Methods.’’ Protein samples were from wild-type spores (lane 1), �sscA (lane 2), CsscA (lane 3), �cotB (lane 4), �cotG (lane 5), and�cotH (lane 6). Asterisks indicate that bands significantly decreased in amount in the protein extract from the �sscA spores. The 59-kDa and36-kDa positions correspond to the sizes of CotB and CotG respectively.
Small Spore Protein Involved in Coat Assembly 1125

the CotH defect, we carried out immunoblot analysisusing an anti-CotH antibody for samples of the samefractions. We observed that the amount of CotH in thesscA mutant spores was markedly reduced (Fig. 6C).This suggests that CotH failed in the assembly of thecoat in the sscA mutant, and thus might have causedmisassembly of both CotB and CotG. CotB, CotG, andCotH are transcribed by a SigK RNA polymerase, suchas SscA. YaaH is controlled by SigE, and the lattergoverns expression of genes in the mother cell duringearly sporulation.47) YaaH is used as a loading control forprotein content evaluation in spores.48) We examined todetermine whether YaaH was assembled in the spores ofthe sscA mutant by immunoblot analysis using an anti-YaaH antibody. The level of YaaH in the sscA mutantspores was similar to that in the wild type (Fig. 6D). Thisindicates that SscA is involved in the assembly of CotB,CotG, and CotH, either directly or indirectly.
Discussion
As shown in Fig. 7, SscA paralogs are found in eightprotein-coding genes within the B. subtilis genome.Typical ribosome binding sites are present upstream ofthe translation initiation codon (AUG) (Fig. 7A). Henceit is possible that the SscA paralogs are indeed translatedinto proteins. These small proteins have a highlyconserved sequence composed of hydrophobic aminoacid residues in the mid to C-terminal regions (Fig. 7B).Though the N-terminal region of the SscA paralogs iscomposed mainly of tyrosine and glysine residues, the
primary sequences and the numbers of amino acidresidues are different. The C-terminal extension se-quences following the hydrophobic region are alsodifferent. These distinct amino acid sequences suggestthat the SscA paralogs are composed of two or threedomains. Interestingly, orthologs of SscA are found onlyin the genus Bacillus, not in Clostridium. sscA, orf-62,and yjcZ are all transcribed during sporulation by SigKRNA polymerase and GerE in the mother-cell compart-ment. We found that some other paralogous genes(yoyG, yczM, and ykzV) have distinctive promoterssimilar to the SigK consensus sequence (data notshown). Most of the homologous genes might betranscribed at the same time in the mother cell. Thus,it is likely that the ORFs play similar roles insporulation. We found that spores of the sscA mutanthad low efficiency of response to L-alanine and AGFK,which stimulate spore germination, but orf-62 mutantspores exhibited germination similarly to the wild type.The level of transcription of orf-62 from P2 promoterwas markedly lower than that of sscA from P1 promoter.It is possible that the low amount of ORF-62 in the wild-type spores hardly affected germination. Partial assem-bly of CotB, CotG, and CotH in the spores of the sscAmutant suggests that other SscA paralogs are involved inthe synthesis or assembly of some spore coat proteins. Amultiple mutant lacking SscA paralogs might exhibit asignificant phenotypic defect more obviously than anindividual mutant. Our data suggest that these paralogshave a role in spore formation (data not shown).SscA-FLAG was detected in wild-type spores by
A B
C D
Fig. 6. Immunoblotting of CotB, CotG, CotH, and YaaH in �sscA Spores.Protein samples from spores (T24) were solubilized and analyzed as described in ‘‘Materials and Methods.’’ Immunoblotting was performed
with anti-CotB (A), anti-CotG (B), anti-CotH (C), and anti-YaaH (D) antisera. Arrowheads show the positions of proteins.
1126 T. KODAMA et al.

Western blot analysis (Fig. 3). Furthermore SscA wasexpressed in the mother-cell compartment by SigK andGerE, and thus SscA might be localized in the spore coatlayer. SscA and Orf-62, composed of 28 amino acidresidues, were the smallest proteins among the SscAparalogs. At present, CotC, composed of 66 amino acidresidues with a deduced molecular mass of 8.8 kDa, isrecognized as the smallest spore coat protein.45) Ourresults suggest that the 28 aa sequence of SscA issufficient for coat localization and that SscA is thesmallest spore coat protein of B. subtilis. We expect thatdetailed analysis of the function and localization ofSscA paralogs will provide clue to their role in thelocalization of the spore coat.
Tricine–SDS–PAGE showed that the wild-type sporeswere comprised of some proteins smaller than 6 kDa,and that the sscA mutant spores had decreased levels of5- and 3-kDa proteins (Fig. 5). We have two hypothesesrelating to the 5- and 3-kDa proteins in the sscA mutantspores. First, the 3-kDa protein in the 168 and someother spores might be SscA, since the deduced molecu-lar mass of SscA corresponded to the protein. Second,the 3-kDa one might be a protein not yet identified.
Since SscA is remarkably small in size, it is unlikelythat it plays a role as a receptor for germination-stimulating factors or participates directly in the signaltransduction pathway for spore germination. CotH isrequired for the proper assembly of spore coat proteinsand germination.29) The mutant spores lost manyproteins by complete depletion of CotH (Fig. 5, lane 6).In contrast, the sscA mutant spores lost few proteins(Fig. 5, lane 2). Western blot indicated that the activityof CotH is not completely lost but is limited in theabsence of SscA (Fig. 6C, lane 3). The defects ingermination of the sscA mutant spores are probablydue to abnormal spore coat assembly under the low levelof CotH. Because all of the SscA paralogs are very smallproteins, we imagine they are incorporated into smallspaces of the developing spore coat for stabilizationof certain coat proteins. Identification of the proteinsthat are assembled in spores depending on SscA is
important in the attempt to understand the moleculardetails of spore morphogenesis. It would be interestingto determine whether SscA operates to synthesize orassemble coat proteins. Our findings here should con-tribute to various applications in industrial, medical,and environmental engineering of spore-surface displaysystems.
Acknowledgments
We would like to thank Dr. Masakuni Serizawa andDr. Tatsuya Fukushima for technical advice, and Ms.Nozomi Hayashi for technical assistance. This researchwas carried out as a part of the Project for theDevelopment of a Technological Infrastructure forIndustrial Bioprocesses through R and D on NewIndustrial Science and Technology Frontiers of theMinistry of Economy, Trade, and Industry (METI), andwas supported by the New Energy and IndustrialTechnology Development Organization (NEDO). Itwas also supported by Grants-in-Aid for ScientificResearch on Priority Areas, Genome Biology(no. 12206005), by the Global COE Program, and byScientific Research (B) (no. 16380059) and (A)(no. 22248008 to J. S.) of the Ministry of Education,Culture, Sports, Science, and Technology of Japan.
References
1) Kunst F, Ogasawara N, Moszer I, Albertini AM, Alloni G,
Azevedo V, Bertero MG, Bessieres P, Bolotin A, Borchert S,
Borriss R, Boursier L, Brans A, Braun M, Brignell SC, Bron S,
Brouillet S, Bruschi CV, Caldwell B, Capuano V, Carter NM,
Choi SK, Codani JJ, Connerton IF, Cummings NJ, Daniel RA,
Denizot F, Devine KM, Dusterhoft A, Ehrlich SD, Emmerson
PT, Entian KD, Errington J, Fabret C, Ferrari E, Foulger D,
Fritz C, Fujita M, Fujita Y, Fuma S, Galizzi A, Galleron N,
Ghim S-Y, Glaser P, Goffeau A, Golightly EJ, Grandi G,
Guiseppi G, Guy BJ, Haga K, Haiech J, Harwood CR, Henaut
A, Hilbert H, Holsappel S, Hosono S, Hullo M-F, Itaya M, Jones
L, Joris B, Karamata D, Kasahara Y, Klaerr-Blanchard M, Klein
C, Kobayashi Y, Koetter P, Koningstein G, Krogh S, Kumano
M, Kurita K, Lapidus A, Lardinois S, Lauber J, Lazarevic V,
A
B
Fig. 7. SscA Paralogs.A, Predicted ORFs in the intergenic regions of B. subtilis. The start codon and stop codon of each ORF are boxed. Potential RBSs upstream of
ORFs are underlined. yjcZ was annotated by Kuwana et al.15) The other ‘‘y-genes’’ were annotated on the basis of the B. subtilis genomesequence.3) B, Protein alignment of SscA paralogs. Identical residues are indicated by asterisks.
Small Spore Protein Involved in Coat Assembly 1127

Lee S-M, Levine A, Liu H, Masuda S, Mauel C, Medigue C,
Medina N, Mellado RP, Mizuno M, Moestl D, Nakai S, Noback
M, Noone D, O’Reilly M, Ogawa K, Ogiwara A, Oudega B,
Park S-H, Parro V, Pohl TM, Portetelle D, Porwollik S, Prescott
AM, Presecan E, Pujic P, Purnelle B, Rapoport G, Rey M,
Reynolds S, Rieger M, Rivolta C, Rocha E, Roche B, Rose M,
Sadaie Y, Sato T, Scanlan E, Schleich S, Schroeter R, Scoffone
F, Sekiguchi J, Sekowska A, Seror J, Serror P, Shin B-S, Soldo
B, Sorokin A, Tacconi E, Takagi T, Takahashi H, Takemaru K,
Takeuchi M, Tamakoshi A, Tanaka T, Terpstra P, Tognoni A,
Tosato V, Uchiyama S, Vandenbol M, Vannier F, Vassarotti A,
Viari A, Wambutt R, Wedler E, Wedler H, Weitzenegger T,
Winters P, Wipat A, Yamamoto H, Yamane K, Yasumoto K,
Yata K, Yoshida K, Yoshikawa H-F, Zumstein E, Yoshikawa H,
and Danchin A, Nature, 390, 249–256 (1997).
2) Blattner FR, Plunkett G III, Bloch CA, Perna NT, Burland V,
Riley M, Collado-Vides J, Glasner JD, Rode CK, Mayhew GF,
Gregor J, Davis NW, Kirkpatrick HA, Goeden MA, Rose DJ,
Mau B, and Shao Y, Science, 277, 1453–1462 (1997).
3) Barbe V, Cruveiller S, Kunst F, Lenoble P, Meurice G,
Sekowska A, Vallenet D, Wang T, Moszer I, Medigue C, and
Danchin A, Microbiology, 155, 1758–1775 (2009).
4) Silvaggi JM, Perkins JB, and Losick R, J. Bacteriol., 188, 532–
541 (2006).
5) Johansson J and Cossart P, Trends Microbiol., 11, 280–285
(2003).
6) Wilderman PJ, Sowa NA, FitzGerald DJ, FitzGerald PC,
Gottesman S, Ochsner UA, and Vasil ML, Proc. Natl. Acad.
Sci. USA, 101, 9792–9797 (2004).
7) Borden JR, Jones SW, Indurthi D, Chen Y, and Papoutsakis ET,
Metab. Eng., 12, 268–281 (2010).
8) Burkholder WF, Kurtser I, and Grossman AD, Cell, 104, 269–
279 (2001).
9) Handler AA, Lim JE, and Losick R, Mol. Microbiol., 68, 588–
599 (2008).
10) Hemm MR, Paul BJ, Schneider TD, Storz G, and Rudd KE,
Mol. Microbiol., 70, 1487–1501 (2008).
11) Hemm MR, Paul BJ, Miranda-Rıos J, Zhang A, Soltanzad N,
and Storz G, J. Bacteriol., 192, 46–58 (2010).
12) Kim W, Silby MW, Purvine SO, Nicoll JS, Hixson KK, Monroe
M, Nicora CD, Lipton MS, and Levy SB, PLoS One, 4, e8455
(2009).
13) Silvaggi JM, Popham DL, Driks A, Eichenberger P, and Losick
R, J. Bacteriol., 186, 8089–8095 (2004).
14) Schmalisch M, Maiques E, Nikolov L, Camp AH, Chevreux B,
Muffler A, Rodriguez S, Perkins J, and Losick R, J. Bacteriol.,
192, 5402–5412 (2010).
15) Kuwana R, Yamamura S, Ikejiri H, Kobayashi K, Ogasawara N,
Asai K, Sadaie Y, Takamatsu H, and Watabe K, Microbiology,
149, 3011–3021 (2003).
16) Kuwana R, Kasahara Y, Fujibayashi M, Takamatsu H,
Ogasawara N, and Watabe K, Microbiology, 148, 3971–3982
(2002).
17) Henriques AO and Moran CP Jr, Annu. Rev. Microbiol., 61,
555–588 (2006).
18) Takamatsu H and Watabe K, Cell. Mol. Life Sci., 59, 434–444
(2002).
19) Setlow P, Curr. Opin. Microbiol., 6, 550–556 (2003).
20) Roels S, Driks A, and Losick R, J. Bacteriol., 174, 575–585
(1992).
21) Driks A, Roels S, Beall B, Moran CP Jr, and Losick R, Genes
Dev., 8, 234–244 (1994).
22) Catalano FA, Meador-Parton J, Popham DL, and Driks A,
J. Bacteriol., 183, 1645–1654 (2001).
23) Beall B, Driks A, Losick R, and Moran CP Jr, J. Bacteriol., 17,
1705–1716 (1993).
24) Takamatsu H, Kodama T, Nakayama T, and Watabe K,
J. Bacteriol., 181, 4986–4994 (1999).
25) Ozin AJ, Henriques AO, Yi H, and Moran CP Jr, J. Bacteriol.,
182, 1828–1833 (2000).
26) Zheng LB, Donovan WP, Fitz-James PC, and Losick R, Genes
Dev., 2, 1047–1054 (1988).
27) Little S and Driks A, Mol. Microbiol., 42, 1107–1120 (2001).
28) Bauer T, Little S, Stover AG, and Driks A, J. Bacteriol., 181,
7043–7051 (1999).
29) Naclerio G, Baccigalupi L, Zilhao R, De Felice M, and Ricca E,
J. Bacteriol., 178, 4375–4380 (1996).
30) Zilhao R, Serrano M, Isticato R, Ricca E, Moran CP Jr, and
Henriques AO, J. Bacteriol., 186, 1110–1119 (2004).
31) Schaeffer P, Millet J, and Aubert J-P, Proc. Natl. Acad. Sci.
USA, 54, 704–711 (1965).
32) Kodama T, Endo K, Ara K, Ozaki K, Kakeshita H, Yamane K,
and Sekiguchi J, J. Biosci. Bioeng., 103, 13–21 (2007).
33) Vagner V, Dervyn E, and Ehrlich SD,Microbiology, 144, 3097–
3104 (1998).
34) Yamamoto H, Mori M, and Sekiguchi J, Microbiology, 145,
2171–2178 (1999).
35) Stragier P, Bonamy C, and Karmazyn-Campelli C, Cell, 52,
697–704 (1988).
36) Kuwana R, Ikejiri H, Yamamura S, Takamatsu H, and Watabe
K, Microbiology, 150, 163–170 (2004).
37) Igo MM and Losick R, J. Mol. Biol., 191, 615–624 (1986).
38) Kodama T, Endo K, Ara K, Ozaki K, Kakeshita H, Yamane K,
and Sekiguchi J, J. Biosci. Bioeng., 103, 13–21 (2007).
39) Ishikawa S, Yamane K, and Sekiguchi J, J. Bacteriol., 180,
1375–1380 (1998).
40) Sekiguchi J, Fukushima T, and Ishikawa S, ‘‘Functional
Analysis of Bacterial Genes: A Practical Manual,’’ eds.
Shumann W, Ehrlich SD, and Ogasawara N, John Wiley &
Sons, Chichester, pp. 211–213 (2001).
41) Nicholson WL and Setlow P, ‘‘Molecular Biological Methods
for Bacillus,’’ eds. Harwood CR and Cutting SM, John Wiley &
Sons, pp. 391–450 (1990).
42) Takamatsu H, Chikahiro Y, Kodama T, Koide H, Kozuka S,
Tochikubo K, and Watabe K, J. Bacteriol., 180, 2968–2974
(1998).
43) Donovan W, Zheng LB, Sandman K, and Losick R, J. Mol.
Biol., 196, 1–10 (1987).
44) Sacco M, Ricca E, Losick R, and Cutting S, J. Bacteriol., 177,
372–377 (1995).
45) Isticato R, Esposito G, Zilhao R, Nolasco S, Cangiano G, De
Felice M, Henriques AO, and Ricca E, J. Bacteriol., 186, 1129–
1135 (2004).
46) Isticato R, Pelosi A, Zilhao R, Baccigalupi L, Henriques AO,
De Felice M, and Ricca E, J. Bacteriol., 190, 1267–1275
(2008).
47) Kodama T, Takamatsu H, Asai K, Kobayashi K, Ogasawara N,
and Watabe K, J. Bacteriol., 181, 4584–4591 (1999).
48) Kuwana R, Takamatsu H, and Watabe K, J. Biochem., 142,
681–699 (2007).
1128 T. KODAMA et al.