a comparative study of germinal center: fowls and mammals
TRANSCRIPT

A comparative study of germinal center: fowlsand mammals
Masahiro Yasudaa, c, Yasuho Tauraa, Yuichi Yokomizob,Shigeo Ekinoc, *
aDepartment of Veterinary Hospital, Faculty of Agriculture, Yamaguchi University, Yamaguchi, JapanbDepartment of Biological Product, National Institute of Animal Health, Ibaraki, JapancDepartment of Anatomy, Kumamoto University School of Medicine, Kumamoto, Japan
Received 3 October 1997
Abstract
The mammalian germinal center is organized into a dark zone containing proliferatingcentroblasts and a light zone ®lled with nondividing B cells (called centrocytes), folliculardendritic cells and a few scattered T cells. We clari®ed these two zones in the chicken
germinal center with immunohistology. Proliferating cells and immunoglobulin negativecells were detected in the circumference ring of the chicken germinal center. The centralpart of the chicken germinal center contained B cells expressing immunoglobulin, follicular
dendritic cells and a few T cells. Most of the B cells in the central part of the chickengerminal center did not enter into the S phase. These results suggest that the chickengerminal center is also organized into the dark zone (the circumference ring of germinalcenter) and the light zone (the central part of germinal center). # 1998 Elsevier Science Ltd.
All rights reserved.
Keywords: Germinal center; Dark zone; Light zone; Chicken; Comparative study
Re sumeÂ
Le centre germinatif mammifeÁ re est organise en une zone sombre contenant descentroblastes en prolife ration et une zone claire remplie de cellules B appele es centrocytes
ne se divisant pas, de cellules folliculaires dendritiques et quelques cellules T disse mine es.Nous avons e tudie ces deux zones dans le centre germinatif du poulet parimmunohistologie. Les cellules en prolife ration et les cellules IgG-ont e te de tecte es dans le
Comparative Immunology, Microbiology
& Infectious Diseases 21 (1998) 179±189
0911-6044/98/$19.00 # 1998 Elsevier Science Ltd. All rights reserved.
PII: S0147 -9571 (98)00007 -1
C OMPARATIVE
I MMUNOLOGY
M ICROBIOLOGY &
I NFECTIOUS
D ISEASESPERGAMON
* Author for correspondence. Tel.: 81-96-373-5047; Fax: 81-96-373-5048; e-mail: ekino@kaiju.
medic.kumamoto-u.ac.jp

cercle pe riphe rique du centre germinatif du poulet. La partie centrale du centre germinatifdu poulet contenait des cellules B exprimant des immunoglobulines, des cellules folliculaires
dendritiques et quelques cellules T. La plupart des cellules B de la partie centrale du centregerminatif du poulet ne prolife rait pas. Ces re sultats suggeÁ rent que le centre germinatif dupoulet est aussi organise dans la zone sombre (le cercle per riphe rique du centre germinatif)
et la zone claire (la partie centrale du centre germinatif). # 1998 Elsevier Science Ltd. Allrights reserved.
Mots-cleÂfs: Centre germinatif; Zone sombre; Zone claire; Poulet; EÂ tude comparative
1. Introduction
The germinal center (GC) is a histologically de®ned area in peripheral lymphoid
organs, such as lymph nodes and spleen, that develops following antigenic
stimulation [1±3]. GCs contain proliferating B cells, a few T cells, and are able to
trap and retain immune complexes on the surface of follicular dendritic cells
(FDCs) [1±3]. GCs with these features are found in both mammals and birds
(homoiothermic animals) [1, 4]. However, GCs have not been found in any classes
of poikilothermic animals. Homoiothermic animals which have the ability to form
GCs, display the features of anamnestic antibody (Ab) response: the a�nity and
heterogeneity of Ab. It is known that memory B cells, plasma blasts, a�nity
maturation, isotype switching, and further diversi®cation of B cells are generated
in GCs [1±4]. The microenvironments of GCs induce gene modi®cations such as
somatic hypermutation, gene conversion or secondary immunoglobulin (Ig) gene
rearrangement in GC B cells [5±8]. Mutated B cells are thought to be selected by
antigens retained on FDCs and di�erentiate to memory B cells or plasma blasts.
Immune complexes on FDCs are predominantly formed with IgG or IgA [9]. T
cells in GCs are thought to be necessary for further di�erentiation of GC B cells.
Especially, T cells or cytokines are supposed to be prerequisite to the escape of
apoptosis and isotype switching in GC [10].
The mammalian GC is histologically and functionally classi®ed into two zones:
the dark zone (DZ) and the light zone (LZ) [1±3, 10] (Table 1). The DZ is
constituted of proliferating B cells (called centroblasts), which are surface Ig (sIg)
negative. It is proposed that the mutational process occurs in proliferating
centroblasts in the DZ [10]. The LZ is composed of a few helper T cells, FDCs,
and non-dividing B cells which express Ig. In the LZ, mutated B cells are selected
on the basis of their capacity for activation by antigens held on FDCs, escape
from apoptosis, and di�erentiate into memory B cells or plasma blasts. The role
of helper T cells in the LZ in the selection of mutated B cells and di�erentiation
events remains to be determined in the mammalian GC.
On the other hand, the distinction between the DZ and the LZ was not clear in
chicken splenic GCs. In this report, GCs were studied with immunohistology.
Proliferating cells were detected by utilizing in vivo incorporation of
M. Yasuda et al. / Comp. Immun. Microbiol. Infect. Dis. 21 (1998) 179±189180

bromodeoxyuridine (BrdUrd) and anti-BrdUrd monoclonal antibody (mAb).Surface and cytoplasm Ig were detected with anti-m and anti-light chain mAbs. Tcells were detected with anti-CD3, anti-T cell receptor gd (TCR gd) and anti-TCRab mAbs. FDCs were detected by anti-g mAb, because antigens on FDCs alwaysform immune complexes with IgG [9].
2. Materials and methods
2.1. Chicken
Fertile eggs of H.B15 (B15) strain (Kumamoto University of Medicine, Japan)were used. Eggs were incubated and hatched in our own facilities. The chickenswere housed together under conventional conditions and were given free access tofood and water.
2.2. Immunization
Chickens were immunized with 0.1 mg of 4-hydroxy-3-iodo-5-nitrophenyl acetylcoupled bovine serum albumin (NIP-BSA) by intravenous injection via the wingvein 10 weeks after hatching. Spleen was obtained 7 days after intravenousimmunization and frozen for immunoperoxidase staining.
2.3. Monoclonal antibodies (mAbs)
mAbs speci®c for chicken Bu1a antigen (B cell speci®c), light-chain, m-chain, g-chain, CD3, TCR gd and TCR Vb1 were used. L22 (anti-Bu1a) and CVI-ChIgG-47.3 (anti-g chain) were described previously [12]. CVI-ChIgL-47.5 (anti-light
Table 1
Characteristics of the dark zone and the light zone
Zone
Constituent and characteristics of
cells in GC Function
Dark zone
1. B cells (centroblasts): loss of sIg and
proliferation.
1. B cells proliferate and diversify with gene
modi®cations (point mutation and gene
conversion).
Light zone
1. B cells (centrocyte): Ig expression. 1. FDCs hold antigens in the form of immune
complexes.
2. T cells. 2. Centrocytes are selected on the basis of their
capacity to be activated by antigens held on
FDCs.
3. FDCs. 3. Helper T cells in the light zone would
contribute the selection, isotype switching and
further di�erentiation of B cells.
M. Yasuda et al. / Comp. Immun. Microbiol. Infect. Dis. 21 (1998) 179±189 181

Fig. 1 Caption on facing page
M. Yasuda et al. / Comp. Immun. Microbiol. Infect. Dis. 21 (1998) 179±189182

chain) and CVI-ChIgM-59.7 (anti-m chain) were generated and characterized atthe Central Veterinary Institute [13, 14]. CT3 (anti-CD3), TCR1 (anti-TCR gd)and TCR2 (anti-TCR Vb1) were kind gifts from Dr. M. D. Cooper [15].
2.4. Immunoperoxidase staining of tissue sections
Cryostat sections of spleen were stained with mAbs using an indirectimmunoperoxidase technique as described [12]. Vectastain1 ABC kit (Vector Lab.Inc., Burlingame, CA) was used as the second-stage antibody.
2.5. BrdUrd incorporation in vivo
For in vivo labeling, BrdUrd (Sigma Chemicals Co., St Louis, MO) wasintravenously injected at the dose of 5 mg/chicken. Spleen was taken 3 h after theintravenous injection of BrdUrd. BrdUrd is incorporated into DNA-synthesizingcells and can be detected by anti-BrdUrd mAb (Becton Dickinson, MountainView, CA). BrdUrd+ cells in cryostat sections of spleen were studied by indirectimmunoperoxidase staining, as described [16].
3. Results
3.1. General architecture of chicken spleen
Splenic tissue is organized into red and white pulp (Fig. 1). The white pulpfollows and surrounds the vascular tree of the spleen. Lymphocytes, reticular cellsand macrophages make the sheath along arteries and arterioles (PALS: peri-arteriolar lymphoid sheath), and ellipsoids (PELS: peri-ellipsoidal lymphoidsheath) (Fig. 1) [11, 17, 18]. Ellipsoids (Shweigger±Seidel sheaths) develop in thecircumference of penicillary arterioles which are the terminal portion of the
Fig. 1. Immunohistological pictures of frozen sections of chicken spleen (a) and (b) and the scheme of
architecture of chicken spleen (c). (a),(b): Spleen at 7 days after primary immunization with NIP-BSA.
Frozen sections were stained with anti-Bu1 (B cell) (a) or anti-CD3 (b). (a): A germinal center (GC) is
observed as a B cell follicle at the bifurcation of arterioles (ar). Ellipsoids (el) are circumscribed by a
few layers of B cells (PELS: peri-ellipsoidal lymphoid sheath). (b): GC is surrounded at its base by a T-
cell area (PALS: peri-arteriolar lymphoid sheath). GC contains some T cells (arrows). (c): Spleen
contains distinct regions of lymphoid and erythroid cells known as the white and red pulps,
respectively. The splenic parenchyma can be reached via the splenic artery. After further rami®cations
into trabecular arteries, blood vessels enter the white pulp as central arterioles which extend for the
length of these cylindrical lymphoid zones. Eventually, central arterioles branch to form thin penicillary
arterioles that open into complex venous sinuses. The penicillary arterioles are surrounded by a few
layers of spindle-shaped reticular cells which trap antigens in the circulation. Such accumulations of
reticular cells commonly have the shape of an ellipsoid; hence the term ellipsoid is used for these
localized arrangements of reticular cells. Magni®cation: (a) and (b) �830.
M. Yasuda et al. / Comp. Immun. Microbiol. Infect. Dis. 21 (1998) 179±189 183

arteriolar branches [11]. A few layers of spindle-shaped reticular cells constituteellipsoids and tarps antigens in circulation [11, 17]. B cells (Bu1+ cells) formaggregates around ellipsoids and make PELS (Fig. 1(a) and (c)). PELS is de®nedas the B-cell area. A GC was observed as a B-cell follicle at the bifurcation ofarterioles in the spleens of immunized chickens (Fig. 1(a)). The chicken GC wascircumscribed by a thin capsule of collagenous ®bers, but not by the lymphocytecorona. Since PALS was predominantly composed of T cells (CD3+ cells)(Fig. 1(b)), PALS is called the T-cell area. The GC was adjacent to the T-cell areaand contained a few T cells (Fig. 1(b)). In chicken spleen, clear marginal zoneswhich form B-cell areas in mammals were not observed in the periphery of PALS,whereas scattered B cells were found in PALS (Fig. 1(a) and 2(c)).
3.2. Distribution of BrdUrd+ cells in the chicken GC
In mammals, a majority of BrdUrd+ cells are observed in the DZ [1±4]. Asshown in Fig. 2(a), a majority of BrdUrd+ cells accumulated within a GC in thespleen. Most of BrdUrd+ cells were located in the circumference ring of chickenGC (Fig. 2(b)). These results suggest that the circumference ring of chicken GCwould be equivalent to the DZ of mammalian GC.
3.3. Distribution of Ig positive cells in the chicken GC
In mammals, proliferating centroblasts in the DZ are known to be Ig negative(Table 1). As shown in Fig. 2(d) and (f), cells in the circumference ring of chickenGC expressed neither IgM nor light chain of Ig. The Ig negative zone wascomparable to the BrdUrd+ zone. Taken together, the results presented hereindicate that the circumference ring of chicken GC is equivalent to the DZ ofmammalian GC (Table 1). It is likely that the central part of the chicken GCmight be the LZ. Ig or IgM secreting plasma cells were found along the outside ofthe GC capsule, but not within the GC (Fig. 2(c) and (e)). Most of the plasmacells were distributed near the arterioles attached to the GC. A few layers of Igpositive cells surrounded ellipsoids and formed PELS. In PALS, scattered Igpositive cells were observed (Fig. 2(c)).
3.4. Distribution of IgG bearing FDCs
It is known that IgG forms immune complexes with antigens and is retained byFDCs in GCs [9]. IgG is observed as dendritic appearances of aggregates in GCs.As shown in Fig. 3(a), IgG was distributed as dendritic appearances in the centralpart of chicken GC. This indicates that FDCs retaining antigen±antibodycomplexed are located in the central part of the chicken GC. In the mammalianGC, FDCs are known to be localized in the LZ [1±4]. These results suggest thatthe central part of chicken GC is equivalent to the LZ of mammalian GC (Table 1and Fig. 4).
M. Yasuda et al. / Comp. Immun. Microbiol. Infect. Dis. 21 (1998) 179±189184

Fig. 2. Distribution of BrdUrd+ cells (a) and (b), light chain positive cells (c) and (d), and IgM positive
cells (e) and (f) in spleen. (a): The sample was obtained 3 h after the intravenous administration of
BrdUrd. The frozen section was stained with anti-BrdUrd mAb using indirect immunoperoxidase
staining. The accumulation of BrdUrd+ cells is observed within GC. (b): Most of BrdUrd+ cells are
located in the circumference ring of chicken GC. (c), (e): A majority of light chain or IgM positive cells
are distributed in PELS and GC. Scattered light chain positive cells is observed in PALS (c). Ig or IgM
secreting plasma cells are found along the outside of GC capsule, but not within the GC. Most of the
plasma cells are distributed near the arterioles attached to the GC. (d) and (f): The circumference ring of
GC is light chain and IgM negative. Magni®cation: (a) �710; (b), (d) and (f) �1220; (c) and (e) �660.
M. Yasuda et al. / Comp. Immun. Microbiol. Infect. Dis. 21 (1998) 179±189 185

Fig. 3. Distribution of IgG bearing FDCs (a) and (b), TCR2+ cells (c) and (d) and TCR1+ cells (e)
and (f). (a) and (b): IgG is observed as dendritic appearances of aggregates in the central part of GC.
The circumference ring of GC is IgG negative. (c): TCR2+ cells are distributed in PALS and surround
the GC. (d): Four TCR2+ cells are observed within the GC (small arrows). Two of them are found in
the border between the peripheral ring and the central part of GC. Another two TCR2+ cells locate in
the circumference ring of GC. (e): TCR1+ cells are infrequent in spleen. (f): One TCR1+ cell is
observed with the GC (a small arrow). TCR1+ cell locates in the border between the circumference
ring and the central part of GC. Magni®cation: (a) �820; (b), (d) and (f) �1220; (c) and (e) �660.
M. Yasuda et al. / Comp. Immun. Microbiol. Infect. Dis. 21 (1998) 179±189186
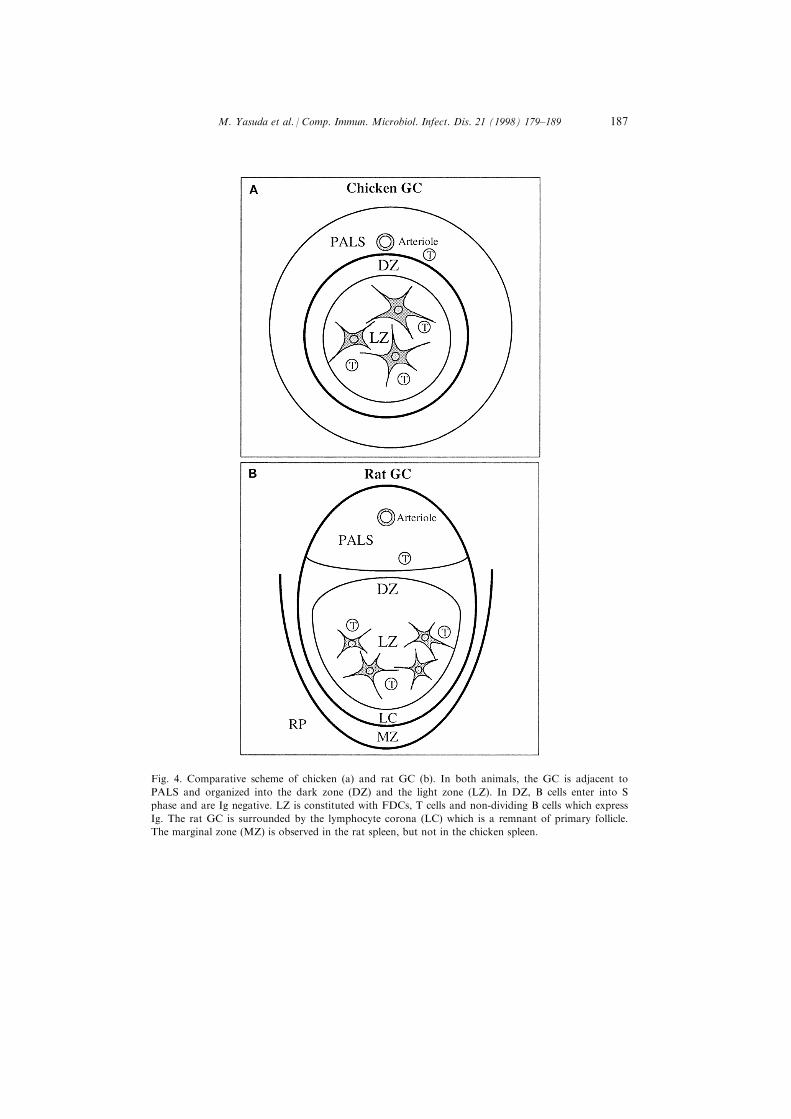
Fig. 4. Comparative scheme of chicken (a) and rat GC (b). In both animals, the GC is adjacent to
PALS and organized into the dark zone (DZ) and the light zone (LZ). In DZ, B cells enter into S
phase and are Ig negative. LZ is constituted with FDCs, T cells and non-dividing B cells which express
Ig. The rat GC is surrounded by the lymphocyte corona (LC) which is a remnant of primary follicle.
The marginal zone (MZ) is observed in the rat spleen, but not in the chicken spleen.
M. Yasuda et al. / Comp. Immun. Microbiol. Infect. Dis. 21 (1998) 179±189 187

3.5. Distribution of T cells in the chicken GC
Several CD3+ cells were observed in the chicken GC (Fig. 1(b)). They werefound in the border between the DZ and the LZ (Fig. 1(b), 3(d) and (f)). Most ofthe CD3+ cells were located in the LZ rather than the DZ. A majority of T cellsin PALS were TCR2+ cells (Fig. 3(c)). TCR1+ cells were in a minority in thechicken spleen (Fig. 3(e)). In addition, TCR2+ cells were dominant in comparisonwith TCR1+ cells in the chicken GC (Fig. 3(d) and (f)).
4. Discussion
It is suggested that the features of chicken GC are very similar to those ofmammalian GC, i.e. the distribution of immune complexes on the surface of adendritic cell type, T-cell dependence of the development of GCs and thedistribution of some T cells [1, 4]. In contrast to the situation in mammals, thechicken GC in the spleen are surrounded by a thin layer of ®brous tissue but notby the lymphocyte corona which is a part of the primary follicle. Furthermore,discrimination between the DZ and the LZ was not made in the chicken GC. Ourimmunohistological studies clarify that the chicken GC is also organized into theDZ and the LZ and is histologically equivalent to the mammalian GC (Table 1and Fig. 4).
The chicken GC is surrounded by a capsule of connective tissue, makingpossible a clear separation of a single GC [5]. We obtained single GCs induced byimmunization and studied Ig variable regions from single GCs [5]. Sincefunctional Ig genes and the germline template donor sequences for geneconversion are well characterized in chickens, we analyzed the genealogicalrelationship of light chain and heavy chain sequences derived from the single GCs.This report demonstrates the ongoing post-bursal diversi®cation of B-cells insplenic GCs by templated gene conversion as well as untemplated pointmutations [5]. Recently, it is reported that the microenvironment of mouse GCinduces secondary Ig gene rearrangement as well as somatic hypermutation inrearranged Ig variable regions [7, 8]. These results suggest that the chicken GC isfunctionally comparable to the mammalian GC.
Acknowledgements
This work is supported in part by a Grant-in-Aid (Bio media Program) fromthe Japanese Ministry of Agriculture, Forestry and Fisheries, the Ministry ofEducation, and The Osaka Foundation for Promotion of Clinical Immunology.
M. Yasuda et al. / Comp. Immun. Microbiol. Infect. Dis. 21 (1998) 179±189188

References
[1] Kroese FGM, Timens W, Nieuwenhuis P. Germinal center reaction and B lymphocytes: mor-
phology and function. Current Topics in Pathology 1990;84:103±48.
[2] Liu Y-J, Zhang J, Lane PJL, Chan EY-T, MacLennan ICM. Sites of speci®c B cell activation in
primary and secondary responses to T cell-dependent and T cell-independent antigens. Eur. J.
Immunol. 1991;21:2951±62.
[3] Kelsoe G. In situ studies of the germinal center reaction. Adv. Immunol. 1995;60:267±88.
[4] Nahm MH, Kroese FGM, Ho�mann JW. The evolution of immune memory and germinal centers.
Immunology Today 1992;13:438±41.
[5] Arakawa H, Furusawa S, Ekino S, Yamagishi H. Immunoglobulin gene hyperconversion ongoing
in chicken splenic germinal centers. The EMBO Journal 1996;15:2540±1546.
[6] Liu Y-J. Reuse of B lymphocytes in germinal centers. Science 1997;278:238±9.
[7] Papavasiliou F, Casellas R, Suh H, Qin X-F, Besmer E, Pelanda R, Nemazee D, Rajewsky K,
Nussenzweig MC. V(D)J Recombination in Mature B cells: A mechanism for altering antibody
responses. Science 1997;278:298±301.
[8] Han S, Dillion S, Zheng B, Shimoda M, Schlissel MS, Kelsoe G. V(D)J recombinase activity in s
subset of germinal center B lymphocytes. Science 1997;278:301±5.
[9] Heinen E, Coulie P, van Snick J, Braun M, Cormann N, Moeremans M, Kinet-Denoel C, Simar
LJ. Retention of immune complexes by murine lymph node or spleen follicular dendritic cells:
Role of antibody isotype. Scand. J. Immunol. 1986;24:327±34.
[10] Liu Y-J, Johnson GD, Gordon J, MacLennan ICM. Germinal centres in T-cell-dependent anti-
body responses. Immunology Today 1992;13:17±21.
[11] White RG, Henderson DG, Eslami MB, Nielsen KH. Localization of a protein antigen in the
chicken spleen: E�ect of various manipulative procedures on the morphogenesis of the germinal
centre. Immunology 1975;28:1±21.
[12] Ekino S, Riwar B, Kroese FGM, Schwander EH, Koch G, Nieuwenhuis P. Role of environmental
antigen in the development of IgG+ cells in the bursa of Fabricius. J. Immunol. 1995;155:4551±8.
[13] Jeurissen SHM, Janse EM, Ekino S, Nieuwenhuis P, Koch G, De Boer GF. Monoclonal anti-
bodies as probes for de®ning cellular subsets in the bone marrow, thymus, bursa of Fabricius, and
spleen of the chicken. Vet. Immunol. Imunopathol. 1998;19:225±38.
[14] Bianchi AT, Scholten JW, Jongenelen IM, Koch G. The use of monoclonal antibodies in an
enzyme immunospot assay to detect isotype-speci®c antibody-secreting cells in pigs and chickens.
Vet. Immunol. Immunopathol. 1990;24:125±34.
[15] Bucy RP, Chen C-HH, Cihak J, Losch U, Cooper MD. Avian T cells expressing gd receptors loca-
lize in the splenic sinusoids and the intestinal epithelium. J. Immunol. 1998;141:2200±5.
[16] Ekino S. Role of environmental antigens in B cell proliferation in the bursa of Fabricius at deona-
tal stage. Eur. J. Immunol. 1993;23:772±5.
[17] Hoshi H. On the nature of the periellipsoidal lymphoid tissue of chick spleen. Tohoku J. Exp.
Med. 1972;106:285±305.
[18] Jeurissen SHM, Vervelde L, Janse EM. Structure and function of lymphoid tissues of the chicken.
Poultry Science Rev. 1994;5:183±207.
M. Yasuda et al. / Comp. Immun. Microbiol. Infect. Dis. 21 (1998) 179±189 189