8.metabolismo 2014 15 - pro-thor.compro-thor.com/wp-content/uploads/metabolismo.pdf · proteína...
TRANSCRIPT

04/12/14
1
ì METABOLISMO
CONCEITOS
METABOLISMO
CATABOLISMO ANABOLISMO
Conjunto de reações químicas que ocorrem no interior da célula, de degradação e de biossíntese, promovidas por catalisadores eficientes e específicos – enzimas – cuja aDvidade é influenciada pela concentração de substrato, pH e temperatura. Inclui o catabolismo e o anabolismo.

04/12/14
2
CONCEITOS
CATABOLISMO
ANABOLISMO
Degradação de moléculas para obtenção de energia que é armazenada sob a forma de energia química.
ADvidade de biossíntese => síntese de moléculas complexas a parDr de moléculas simples, com consumo de energia.
2. Desdobramento de ATP
A energia libertada no desdobramento do ATP pode ser uDlizada durante o anabolismo para sinteDzar outras moléculas e para fornecer energia, como o transporte aDvo e a contração muscular.
1. Produção de ATP A energia libertada durante o catabolismo pode ser uDlizada para sinteDzar ATP.
adenosina Pi Pi Pi
adenosina Pi Pi Pi
CATABOLISMO A
NABOLISM
O

04/12/14
3
ENZIMAS
ENZIMAS Moléculas proteicas catalisadoras (aumentam a velocidade das reações químicas sem se alterarem a elas próprias). São específicas para a reação catalisada e para as moléculas que fazem parte desta reação. A conformação proteica é modificada (desnaturação), quando submeDda a valores extremos de temperatura e pH => as enzimas perdem a sua capacidade catalisadora.
ENZIMAS
Região da proteína que parDcipa diretamente na conversão do substrato no produto.
LOCAL ATIVO
SUBSTRATOS E PRODUTOS
As moléculas que reagem são chamadas substratos e as que se formam, produtos.
04/12/14
4
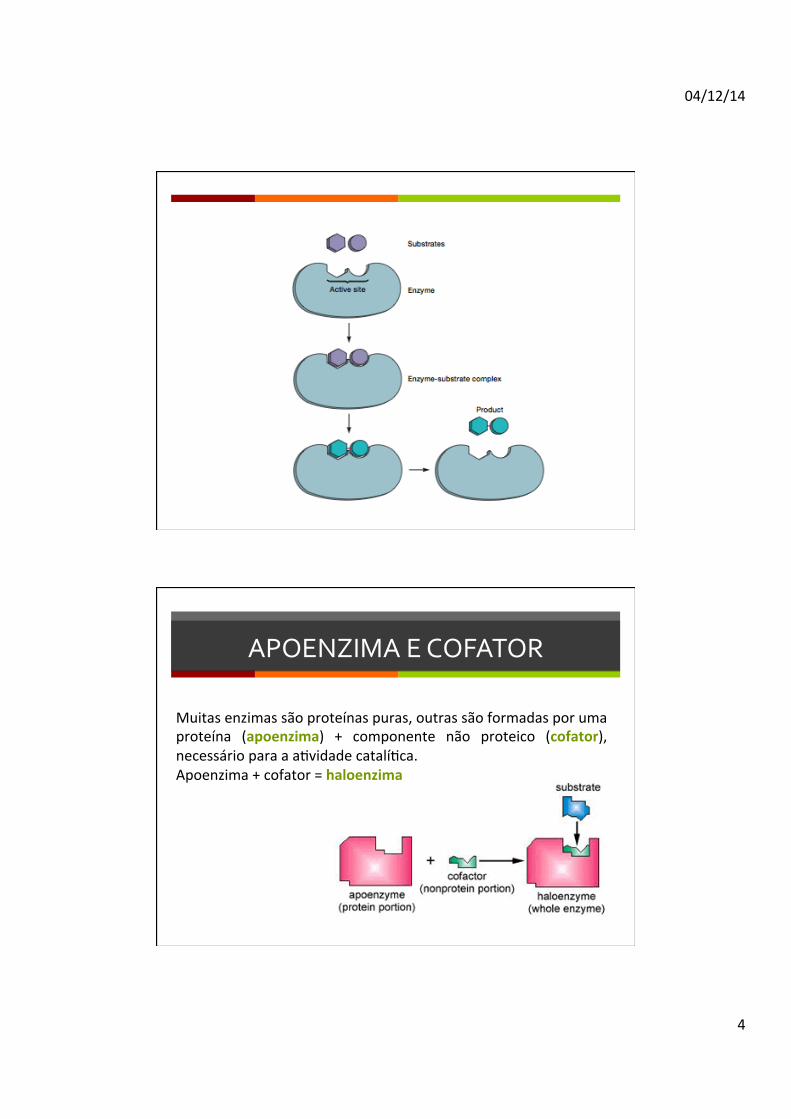
APOENZIMA E COFATOR
Muitas enzimas são proteínas puras, outras são formadas por uma proteína (apoenzima) + componente não proteico (cofator), necessário para a aDvidade catalíDca. Apoenzima + cofator = haloenzima

04/12/14
5
FONTES DE ENERGIA E DE CARBONO E RECEPTORES FINAIS DE ELECTRÕES
FONTES'DE'CARBONO,'ENERGIA'E'ELECTRÕES'FONTES'DE'CARBONO' '''''''''''AUTOTRÓFICOS' CO2'como'principal'ou'única'fonte'de'carbono.'''''''''''HETEROTRÓFICOS' Moléculas'orgânicas.'FONTES'DE'ENERGIA' '''''''''''FOTOTRÓFICOS' Luz'''''''''''QUIMIOTRÓFICOS' Oxidação'de'compostos'orgânicos'ou'
inorgânicos'FONTES'DE'ELECTRÕES' '''''''''''LITOTRÓFICOS' Moléculas'inorgânicas'reduzidas''''''''''''ORGANOTRÓFICOS' Moléculas'orgânicas'!

04/12/14
6
FONTES DE ENERGIA E DE CARBONO E RECEPTORES FINAIS DE ELECTRÕES
TIPOS&PRINCIPAIS&DE&MICRORGANISMOS&(NUTRIÇÃO)&
CLASSIFICAÇÃO*FONTES*DE*ENERGIA*/*ELECTRÕES*/*CARBONO*
EXEMPLOS*
FOTOLITOTRÓFICOS&AUTOTRÓFICOS&
Luz*/*Moléculas*inorgânicas*/*CO2*
Algas,*cianobactérias,*bactérias*verdes*e*púrpuras*sulfurosas.*
FOTOORGANOTRÓFICOS&HETEROTRÓFICOS&
Luz*/*Moléculas*orgânicas*/*Moléculas*orgânicas*
Bactérias*verdes*e*púrpuras*não*sulfurosas.*
QUIMIOLITROTRÓFICOS&AUTOTRÓFICOS&
Fonte*inorgânica*de*energia*química*/*Moléculas*inorgânicas*/*CO2*
Bactérias*oxidantes*do*enxofre,*do*férro*e*nitrificantes.*
QUIMIOORGANOTRÓFICOS&HETEROTRÓFICOS&
Fonte*orgânica*de*energia*química*/*Moléculas*orgânicas*/*Moléculas*orgânicas*
Protozoários,*Fungos*e*a*maioria*das*bactérias*não*fotossintéticas*(incluindo*as*patogénicas).*
!
04/12/14
7
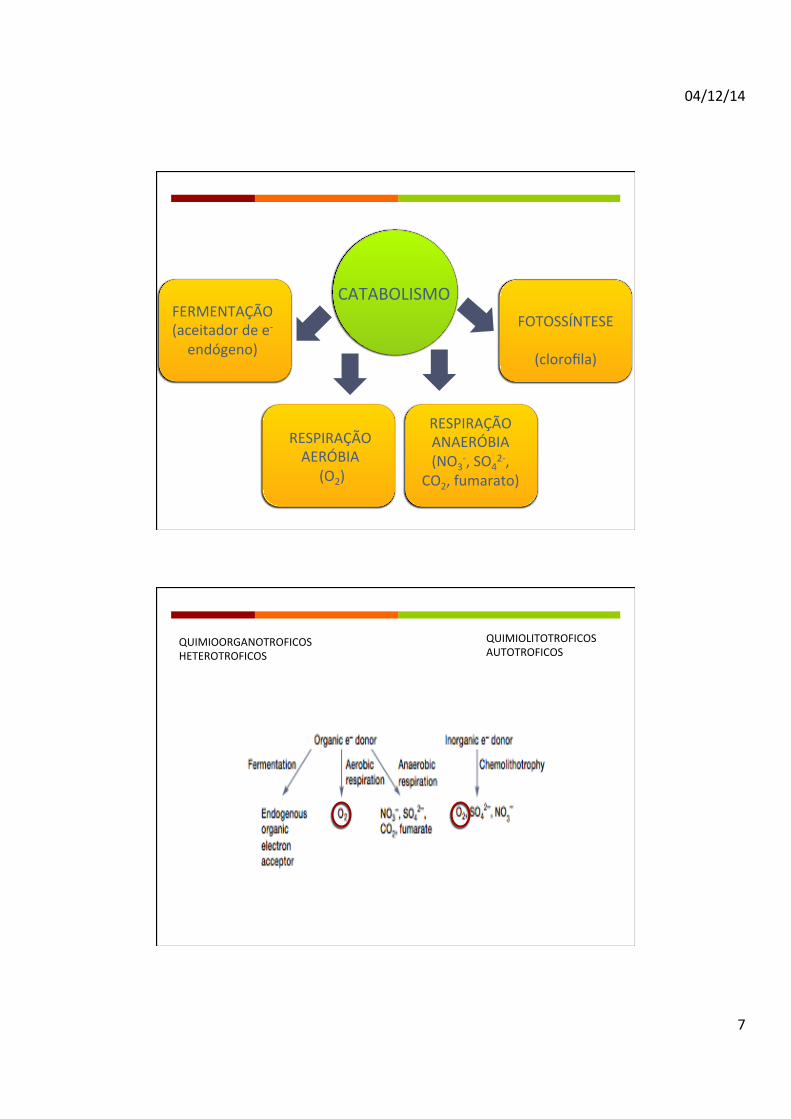
CATABOLISMO
RESPIRAÇÃO ANAERÓBIA (NO3
-‐, SO42-‐,
CO2, fumarato)
RESPIRAÇÃO AERÓBIA (O2)
FERMENTAÇÃO (aceitador de e-‐ endógeno)
FOTOSSÍNTESE
(clorofila)
QUIMIOORGANOTROFICOS HETEROTROFICOS
QUIMIOLITOTROFICOS AUTOTROFICOS

04/12/14
8
RESPIRAÇÃO AERÓBIA: os dadores de e-‐ podem ser orgânicos ou inorgânicos e o receptor de e-‐ é o O2.
RESPIRAÇÃO ANAERÓBIA: os dadores de e-‐ podem ser orgânicos ou inorgânicos e os receptores de e-‐ são compostos inorgânicos oxidados como o nitrato, o sulfato, o ião ferroso ou o CO2
FERMENTAÇÃO: dador de e-‐ orgânico. o substrato energéDco é oxidado e degradado sem a uDlização de uma fonte exógena ou externa como receptor de e-‐. Geralmente são usados intermediários produzidos nas vias metabólicas, como o piruvato, que vão funcionar como aceitadores de e-‐.
FOTOSSÍNTESE: a clorofila funciona como dador e receptor de e-‐.
AS TRÊS ETAPAS DO CATABOLISMO AERÓBIO
FASE 1:
As moléculas de nutrientes maiores (proteínas, polissacáridos e lípidos) são hidrolisados em moléculas mais pequenas (aminoácidos, monossacáridos, ácidos gordos e glicerol). As reações químicas que ocorrem durante esta fase não libertam muita energia.

04/12/14
9
AS TRÊS ETAPAS DO CATABOLISMO AERÓBIO
FASE 2:
Pode ocorrer aeróbica ou anaerobiamente e é normalmente produzido algum ATP e moléculas NADH e FADH2. São formados AceDl coenzima A (descarboxilação oxidaDva, catabolismo dos ácidos gordos), piruvato (glicólise) e intermediários do ciclo de Krebs (catabolismo dos aa).
AS TRÊS ETAPAS DO CATABOLISMO AERÓBIO
FASE 3:
Os nutrientes carbonados são usados no ciclo de Krebs e as moléculas são completamente oxidadas a CO2 com produção de ATP, NADH e FADH2. O ciclo trabalha aerobicamente e é responsável pela libertação de grande quanDdade de energia – a maioria do ATP resulta da oxidação do NADH e FADH2 na cadeia transportadora de electrões. O oxigénio é o aceitador final de electrões.

04/12/14
10
OBTENÇÃO DE ENERGIA NOS MICRORGANISMOS QUIMIOORGANOTRÓFICOS HETEROTRÓFICOS
FONTE DE CARBONO: moléculas orgânicas
FONTE DE ENERGIA:
DADORES DE ELECTRÕES:
RECEPTORES DE ELECTRÕES:
MECANISMOS:
moléculas orgânicas
O2 : respiração aeróbia; composto inorgânico (n itrato, sulfato, CO2): respiração anaeróbia; compostos orgânicos endógenos (fermentação)
Glicólise / Via das pentoses-‐fosfato/Via de Entner-‐Doudoroff Fermentação Respiração (aeróbia ou anaeróbia)
moléculas orgânicas

04/12/14
11
TRANSFORMAÇÃO DA GLICOSE EM PIRUVATO
VIA GLICOLÍTICA (GLICÓLISE)
VIA DAS PENTOSES FOSFATO
VIA DE ENTNER-‐DOUDOROFF
TRANSFORMAÇÃO DA GLICOSE EM PIRUVATO
A maioria das bactérias usa a glicólise e a via das pentoses-‐fosfato. Algumas usam a via Entner-‐Doudoroff – Pseudomonas e outras
Gram-‐. Muito poucas Gram+ têm esta via, o Enterococcus faecalis é uma rara exceção.

04/12/14
12
GLICÓLISE
Glucose
Glucose-6-P
Frutose-6-P
Frutose-1,6-P
Dihidroxicetona Gliceraldeído-3-P
1,3-diPGlicerato
3-P-Glicerato
2-P-Glicerato
Fosfoenolpiruvato
Piruvato

04/12/14
13
Glucose
Glucose-6-P
Frutose-6-P
Frutose-1,6-P
DihidroxicetonaP Gliceraldeído-3-P
1,3-diPGlicerato
3-P-Glicerato
2-P-Glicerato
Fosfoenolpiruvato
Piruvato

04/12/14
14
VIA DAS PENTOSES-‐FOSFATO
Uma segunda via para degradação da glicose que pode ser usada em vez da glicólise ou da via Entner-‐Doudoroff. Pode ocorrer anaeróbica ou aerobicamente. Começa com a oxidação da glucose-‐6P em 6-‐fosfogloconato, seguida da oxidação deste em ribulose-‐5P e CO2. Durante estas oxidações é produzido NADPH. A ribulose-‐5P é depois converDda numa mistura de açucares-‐P com 3 a 7C (Xilulose-‐5P, Ribose-‐5P, Sedoheptulose-‐7P, etc.).
VIA DAS PENTOSES-‐FOSFATO
• balanço final é: 3 moléculas de glucose-‐6P são converDdas em 2 moléculas de frutose-‐6P, 1 molécula de Gliceraldeído-‐3P, 3 moléculas de CO2 e 6 NADPH.
Estas moléculas intermédias podem ser usadas de 2 formas: -‐ frutose-‐6P pode reconverter-‐se em glucose-‐6P -‐ Gliceraldeído-‐3P é transformado em ácido piruvico (glicólise) ou
é reconverDdo em glucose-‐6P, podendo também retornar à via pentoses-‐fosfato.

04/12/14
15
Açucares com 3 a 7 C
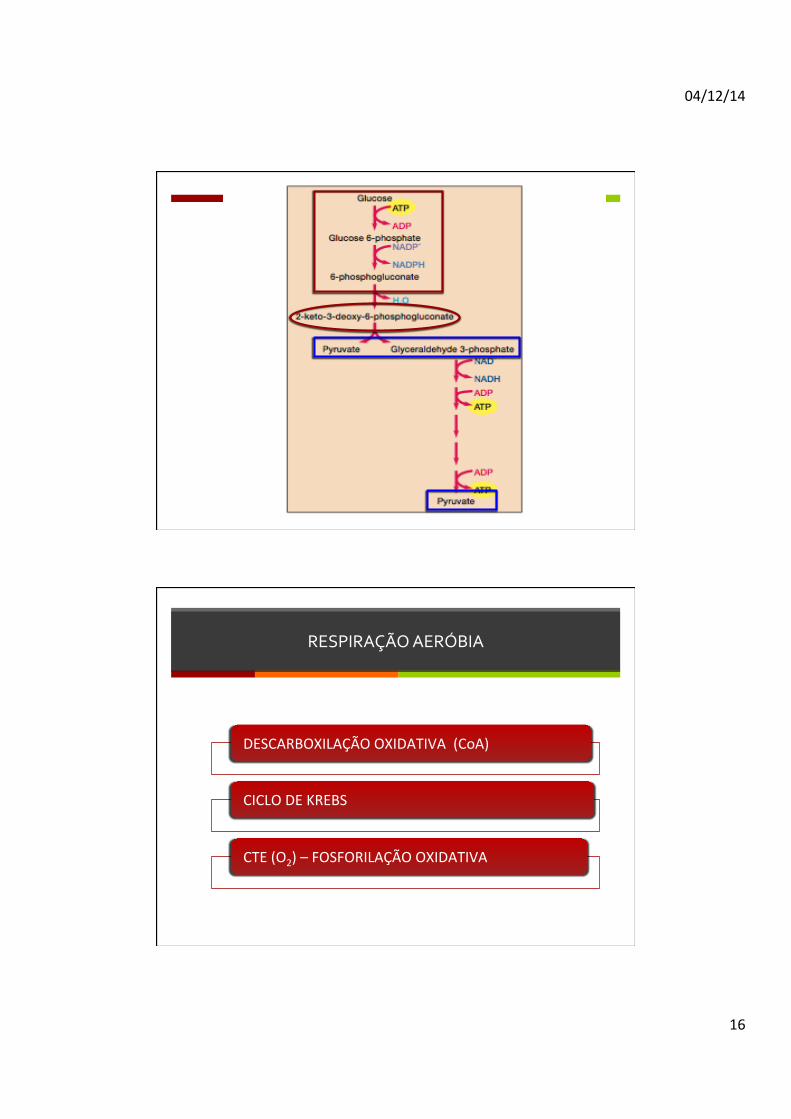
VIA DE ENTNER-‐DOUDOROFF
Apesar da glicólise ser a via mais comum para transformação de hexoses em piruvato, a via Entner-‐Doudoroff é uma via alternaDva com papel semelhante. Começa com as mesmas reações da via pentoses-‐fosfato, com a formação de Glucose-‐6P e 6-‐Fosfogloconato. Esta molécula, em vez de ser novamente oxidada, é desidratada para a forma 2-‐Keto-‐3-‐deoxi-‐fosfogluconato (KDPG). O KDPG é depois clivado em piruvato e gliceraldeído-‐3P que será também converDdo em piruvato. Através desta via são produzidos 1 ATP, 1 NADH e 1 NADPH por molécula de glicose metabolizada.
04/12/14
16
RESPIRAÇÃO AERÓBIA
DESCARBOXILAÇÃO OXIDATIVA (CoA)
CICLO DE KREBS
CTE (O2) – FOSFORILAÇÃO OXIDATIVA

04/12/14
17
DESCARBOXILAÇÃO OXIDATIVA
CICLO DE KREBS
3x
OxaloacetadoCitrato

04/12/14
18
Citrato
Malato
Cis-aconitato
Isocitrato
α-CetoglutaratoSuccinil CoA
Succinato
Fumarato
+
Oxaloacetado
+
CoACoA
Piruvato
AcetilCoA+ CoA
M Castel-‐Branco / C Viegas

04/12/14
19
CADEIA TRANSPORTADORA DE ELECTRÕES
FUNÇÕES: • Receber os electrões de um substrato dador e entregá-‐los ao O2; • Conservar parte da energia que é libertada durante a
transferência dos e-‐ para a síntese de ATP.
EUCARIOTAS: CTE inserida na membrana interna das mitocôndrias PROCARIOTAS: CTE localizada na MC
CADEIA TRANSPORTADORA DE ELECTRÕES
FOSFORILAÇÃO OXIDATIVA: síntese de ATP ligada ao transporte de electrões e entrada de O2.
Formação de 3 ATP por NADH Formação de 2 ATP por FAD

04/12/14
20
RESPIRAÇÃO AERÓBIA: BALANÇO ENERGÉTICO (EUCARIOTAS)
GLICÓLISE: Glicose -‐> Gliceraldeído 3P: -‐ 2 ATP 2x Gliceraldeído 3P -‐> 2x Piruvato: 3 ATP x 2 NADH = + 6 ATP 4 ATP = + 4 ATP + 8 ATP
OXIDAÇÃO DO PIRUVATO: 2x Piruvato -‐> 2x AceDl-‐CoA: 3 ATP x 2 NADH = + 6 ATP Ciclo de Krebs (x2): 3 ATP x 3 NADH = +18 ATP 2 ATP x 1 FAD = + 4 ATP 1 GTP = + 2 ATP + 30 ATP Embora as CTE bacterianas sejam parecidas com as mitocondriais dos eucariotas, são diferentes e menos eficientes na produção de ATP.
RESPIRAÇÃO ANAERÓBIA
Muitas bactérias têm cadeias transportadoras de electrões que podem funcionar com outros aceitadores exógenos para além do O2 – este processo de obtenção de energia chama-‐se RESPIRAÇÃO ANAERÓBICA. Os principais aceitadores de electrões são: Nitrato (NO3
-‐) => anaeróbias facultafvas (Pseudomonas, Bacillus) Sulfato (SO4
2-‐) – SULFITOREDUTORES (anaeróbios estritos) (Clostridium) CO2 -‐ METANOGENICOS (anaeróbios estritos) (arqueobactérias metanogénicas)

04/12/14
21
RESPIRAÇÃO ANAERÓBIA: NITRATO
BACTÉRIAS QUE USAM O NITRATO (ANAERÓBIAS FACULTATIVAS): Algumas bactérias (ex. Pseudomonas e Bacillus) usam o nitrato (NO3
-‐) como o aceitador de electrões no final da cadeia transportadora de electrões e ainda assim, produzem ATP => o metabolismo é Dpicamente respiratório com o O2 como aceitador final de e-‐ mas algumas espécies podem usar o nitrato como aceitador alternaDvo permiDndo o crescimento em condições de anaerobiose.
O nitrato pode ser reduzido a nitrito (NO2
-‐) pela nitrato redutase:
RESPIRAÇÃO ANAERÓBIA: NITRATO
A redução do nitrato a nitrito não é uma forma muito eficiente de produção de ATP porque a uma molécula de nitrato aceita apenas 2 electrões e, assim, são necessárias grandes quanDdades de nitrato para o crescimento. Para além deste, existe ainda o inconveniente do nitrito ser tóxico – por este moDvo o nitrato sofre frequentemente mais reduções até gás nitrogénio (N2) – processo chamado DESNITRIFICAÇÃO – desta forma, cada nitrato recebe 5 electrões e o produto final não é tóxico:

04/12/14
22
RESPIRAÇÃO ANAERÓBIA: NITRATO
A desnitrificação é levada a cabo por alguns membros dos géneros: Pseudomonas e Bacillus (por exemplo) – usam esta via como uma alternaDva à respiração aeróbica, podendo ser considerados ANAERÓBIOS FACULTATIVOS. Se o O2 está presente estas bactérias usam a respiração (a síntese da enzima nitrato redutase é reprimida pelo O2).
RESPIRAÇÃO ANAERÓBIA: SULFATO e CO2
Dois outros grandes grupos de bactérias são ANAERÓBIOS ESTRITOS: METANOGÉNICOS: os que usam CO2 como aceitador final de electrões, reduzindo-‐o a metano (ex: arqueobactérias metanogénicas); SULFITOREDUTORES: os que usam sulfato como aceitador final de electrões, reduzindo-‐o a sulfito (ex: Clostridium sulfitoredutores).

04/12/14
23
RESPIRAÇÃO ANAERÓBIA
A respiração anaeróbia não é tão eficiente na síntese de ATP como a aeróbia – não é produzido tanto ATP por fosforilação oxidaDva, tendo como aceitadores finais de electrões o nitrato, o sulfato ou o CO2. No entanto, a respiração anaeróbica é úfl porque é mais eficiente do que a fermentação e permite a produção de ATP por transporte de electrões e fosforilação oxidafva, na ausência de O2.
FERMENTAÇÃO
FERMENTAÇÃO: Oxidação da glucose por uma via FERMENTATIVA => na ausência de
um receptor externo de electrões.

04/12/14
24
FERMENTAÇÃO
Na ausência de respiração (aeróbica ou anaeróbica), o NADH não é oxidado pela cadeia transportadora de electrões porque não existe nenhum aceitador externo disponível. Assim, o NADH que é produzido na glicólise terá que ser novamente oxidado a NAD+ (se o NADH não fosse regenerado a glicólise teriam que parar, porque este existe em quanDdades limitadas na célula).
FERMENTAÇÃO
Muitos microrganismos resolvem este problema da seguinte forma: abrandam ou interrompem a afvidade da enzima piruvato desidrogenase (piruvato -‐> aceflCoA) e usam o piruvato (ou um derivado deste) com receptor (aceitador) de electrões e H+ para a reoxidação do NADH, num processo fermentafvo.

04/12/14
25
FERMENTAÇÃO
Em suma, nas fermentações microbianas: O NADH é oxidado a NAD+;
O aceitador de electrões é o piruvato ou um derivado deste; Não é necessário O2.

04/12/14
26
FERMENTAÇÃO
Existem muitos Dpos de fermentação que muitas vezes são caracterísDcas de determinados Dpos de microrganismos. As fermentações microbianas mais comuns são: FERMENTAÇÃO LÁCTICA: com redução do piruvato a lactato. Bactérias ácido-‐lácfcas (Streptococcus, Lactobacillus), Bacillus. FERMENTAÇÃO ALCOÓLICA: fermentação dos açucares a ETANOL + CO2. LEVEDURAS (e algumas bactérias, algas e protozoários);
FERMENTAÇÃO ALCOÓLICA
Oxidação da Glucose, com formação de 2 moléculas de CO2 e etanol.
Leveduras e outros microrganismos.
Regeneração do NAD+: na fermentação alcoólica o NADH cede os seus electrões ao acetaldeído que se reduz, originando etanol.

04/12/14
27
FERMENTAÇÃO ALCOÓLICA: BALANÇO ENERGÉTICO
Na FERMENTAÇÃO ALCOÓLICA o balanço energéDco é: 2 ATP
+ 2 etanol e CO2
OBS: para a levedura o produto essencial é o ATP mas para o tecnologista é o etanol (enólogo) e o CO2 (padeiro)
04/12/14
28
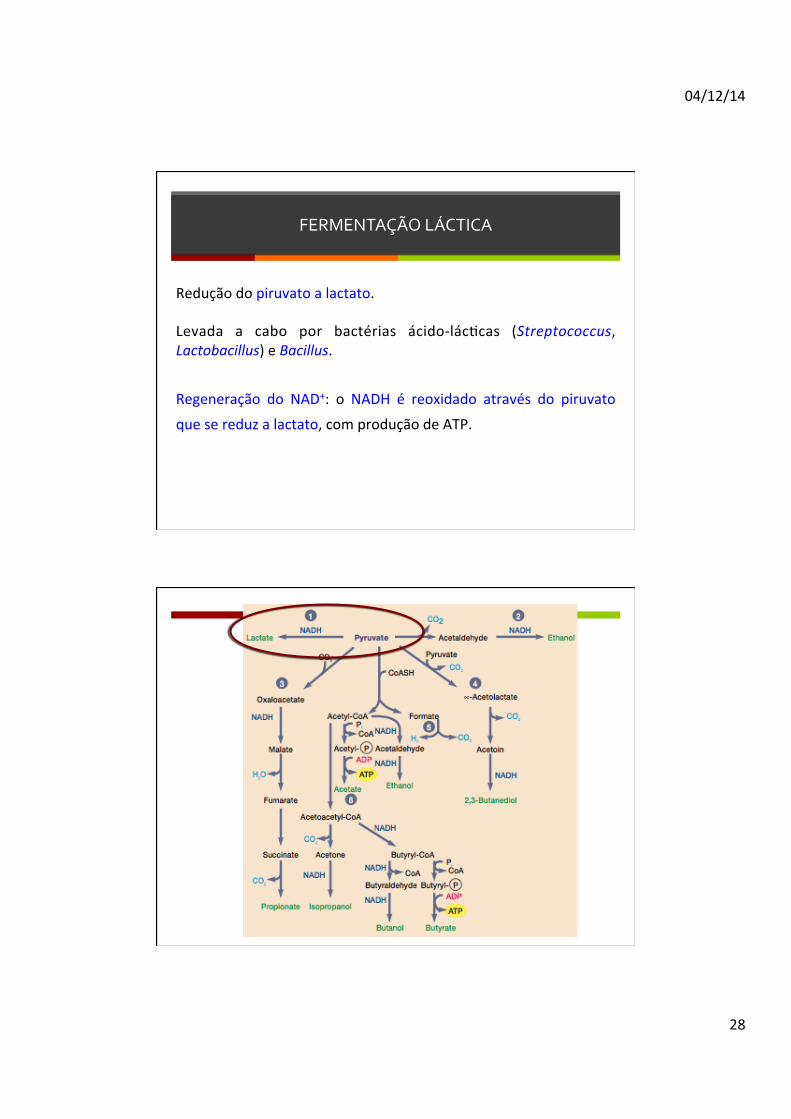
FERMENTAÇÃO LÁCTICA
Redução do piruvato a lactato. Levada a cabo por bactérias ácido-‐lácDcas (Streptococcus, Lactobacillus) e Bacillus. Regeneração do NAD+: o NADH é reoxidado através do piruvato
que se reduz a lactato, com produção de ATP.

04/12/14
29
OUTRAS FERMENTAÇÕES
OUTRAS: com formação de propionato (bactérias ácido-‐propionicas), de 2,3-‐Butanediol (Enterobacter, Serra8a, Bacillus), de H2 e CO2 (bactérias entéricas – Escherichia, Enterobacter,
Salmonella, Proteus), de acetato (Clostridium)...
OUTRAS FERMENTAÇÕES
Clostridium: Um grande nº de espécies do género Clostridium fermentam os
açucares com produção de ácido burrico. Algumas produzem acetona e butanol. Outros produzem acetato ...

04/12/14
30
OUTRAS FERMENTAÇÕES
Bactérias entéricas – Escherichia, Enterobacter, Salmonella, Proteus) :
Fermentação de açucares com produção de H2 e CO2.

04/12/14
31
OUTRAS FERMENTAÇÕES
Enterobacter, Serra6a, Bacillus: Fermentação de açucares com produção 2,3-‐Butanediol.

04/12/14
32
OUTRAS FERMENTAÇÕES
Bactérias ácido-‐propionicas (queijo do Dpo Emmental): Fermentação de açucares com formação de propionato.

04/12/14
33
OBTENÇÃO DE ENERGIA NOS MICRORGANISMOS QUIMIOLITOTRÓFICOS AUTOTRÓFICOS
FONTE DE CARBONO: CO2 (converDdo em glucose através do ciclo de Calvin)
FONTE DE ENERGIA:
DADORES DE ELECTRÕES:
RECEPTORES DE ELECTRÕES:
moléculas inorgânicas (H2, NH3, Fe2+, NO2-‐, SO3
2-‐)
O2
moléculas inorgânicas (H2, NH3, Fe2+, NO2
-‐, SO32-‐)
Exemplos: bactérias nitrificantes (usam o nitrito) e oxidantes do enxofre (usam compostos de enxofre reduzidos)– papel importante na reciclagem de compostos.

04/12/14
34
OBTENÇÃO DE ENERGIA NOS MICRORGANISMOS QUIMIOLITOTRÓFICOS AUTOTRÓFICOS
As bactérias quimiolitotróficas autotróficas obtêm energia pela oxidação de substâncias inorgânicas reduzidas.
Os e-‐ cedidos por estas moléculas são transferidos na CTE até ao aceitador final (O2).
OBS: estes sistemas de transporte são menos eficientes, formando-‐se menos ATP. Estas bactérias, como usam substâncias inorgânicas como dadores de e-‐ têm que recorrer ao CO2 como fonte de carbono.
OBTENÇÃO DE ENERGIA NOS MICRORGANISMOS FOTOLITOTRÓFICOS AUTOTRÓFICOS
FONTE DE CARBONO: CO2
FONTE DE ENERGIA:
DADORES DE ELECTRÕES:
RECEPTORES DE ELECTRÕES:
MECANISMO:
Luz
clorofila
clorofila
fotossíntese

04/12/14
35
ANABOLISMO uso da energia na biossíntese
Durante um processo de biossíntese, os microrganismos começa com percursores (moléculas inorgânicas e monómeros) e constrói moléculas cada vez mais complexas até ao fabrico de organelos e células.

04/12/14
36
Síntese de PEPTIDOGLICANO
PepDdoglicano: molécula formada por longas cadeias de polissacáridos com: ácido N-‐aceDlmurâmico (NAM) + N-‐aceDlglucosamida (NAG), alternados. + cadeias polipep{dicas adicionadas aos grupos NAM. O processo de biossíntese desta estrutura ocorre em ambos os lados da MC e parDcipam 2 transportadores: Uridina difosfato (UDP) Bactoprenol: álcool que se junta ao NAM e transporta os componentes do pepDdoglicano através da MC hidrofóbica.
Síntese de PEPTIDOGLICANO
A síntese de pepDdoglicano ocorre em 8 passos: (1) UDP, derivados do NAM e da NAG são sintefzados no
citoplasma;
(2) Adição sequencial de aa ao UDP-‐NAM para formar uma cadeia pentapeprdica com 2 D-‐Alaninas terminais adicionadas como um dipep{deo (necessidade de ATP); (3) O NAM-‐Pentapeprdeo é transferido do UDP para um bactoprenol-‐P da supertcie da membrana;

04/12/14
37
Síntese de PEPTIDOGLICANO
(4) UDP-‐NAG adiciona NAG ao NAM-‐pentapeprdeo, para formar a unidade estrutural do pepDdoglicano; (5) A unidade de pepfdoglicano NAM-‐NAG é transportada através da membrana para a outra supertcie pelo bactoprenol bifosfato;

04/12/14
38
Síntese de PEPTIDOGLICANO
(6) A unidade de pepfdoglicano é junta à extremidade da cadeia de pepfdoglicano que está a ser sintefzada; (7) O transportador bactoprenol volta para a supertcie interna da MC, libertando um fosfato durante este processo -‐> transforma-‐se em bactoprenol fosfato que pode ligar-‐se a outro NAM-‐pentapep{deo; (8) Finalmente são formadas ligações cruzadas entre cadeias de pepfdoglicanos por transpepfdação.

04/12/14
39