2006-2011 mission kearney foundation of soil science
TRANSCRIPT

2006-2011 Mission Kearney Foundation of Soil Science: Understanding and Managing Soil-Ecosystem
Functions Across Spatial and Temporal Scales Final Report: 2007033, 1/1/2009-12/31/2009
1University of California, Davis
2Michigan State University
3University of Texas, Austin
*Principal Investigator
For more information contact Dr. Valerie Eviner ([email protected])
Spatial and Temporal Dependence of Plant Effects on Soils across Multiple Scales: Building Mechanistic Understanding to Enhance Management Options
Valerie Eviner*1, Kevin Rice1, Carolyn Malmstrom2, Mary Cadenasso*1 and Christine Hawkes3
Project Background
A comprehensive understanding of vegetation impacts on soil properties and processes, and how
these vary over spatial and temporal scales, is a critical tool for ecosystem management.
Management-induced shifts in plant community composition can have marked impacts on almost
every soil process and property (reviewed in Wardle 2002, Eviner and Chapin 2003).
Furthermore, manipulations of vegetation composition are one of the most effective approaches
for managing multiple soil properties and functions (e.g. resistance to erosion, the capacity to
absorb and retain water, storage and cycling of carbon (C) and nutrients) (Lavelle 2000, Eviner
and Chapin 2001, Drinkwater 1999, Rhoades 1997, Coleman et al. 2001). Documenting the
multiple effects of plant species on soils provides an “ecological toolbox” (Sarrantonio 1994,
Ingels et al. 1998, Eviner and Chapin 2001), allowing managers to select species based on the
multiple services they provide, and to understand the consequences of management-induced
shifts in vegetation. This toolbox can be invaluable in sustainable agriculture, restoration, and
bioremediation.
Existing species toolboxes are based on the average effect of species on soil properties and
processes. Scientific focus on broad-scale generalizations provides a “one size fits all” solution
which is at odds with a manager’s need to provide key soil functions under variable conditions
across space and through time. The effects of a given plant species or community on soils can
vary greatly depending on landscape context (e.g. environmental conditions, management
practices, time, neighboring communities, interaction with other organisms). Ecological
management needs a framework that allows for decision making based on site-specific
conditions—shifting the focus from the average towards incorporating spatial and temporal
variability.

Spatial and Temporal Dependence of Plant Effects on Soils Across Multiple Scales: Building Mechanistic Understanding to Enhance Management Options—Eviner
2
Project Objectives Addressed
This project focuses on understanding the ecosystem effects of the prevalent species in
California grasslands, particularly focusing on:
- “Annuals”- invasive annual species that have dominated California grasslands for the
past two to three centuries and are now naturalized (e.g. Avena sp., Bromus sp.,
Lolium multiflorum)
- “Natives”- native grassland species, including short-lived species (e.g. Vulpia
microstachys, Bromus carinatus, Lupinus bicolor), and long-lived perennial
bunchgrasses (e.g. Nassella pulchra, Elymus glaucus, Leymus triticoides)
- Invasive “Weeds”- more recent annual grass invasions that are noxious weeds and are
currently displacing the previous invasive “annuals” in many areas. These species
Aegilops triuncialis, Taeniatherum caput-medusae) can decrease range productivity
50-80%.
Within and across these groups of species, our objectives included:
1. Determine the impacts of California grassland species on multiple soil processes and
properties. To what extent are species effects on different soil processes related or independent?
Are there likely tradeoffs and synergies in using plant species to provide multiple ecosystem
services?
Soil properties and processes include:
1. Carbon
- plant net primary productivity (aboveground and belowground)
- soil C by depth
- soil organic matter by depth
2. Soil cohesion (an indicator of surface erosion control)
3. Nitrogen
- total soil N
- net N mineralization and nitrification rates
- seasonal resin available N
- N leaching loss (on a subset of plots)
4. Soil water
- infiltration
- water holding capacity
- leaching loss (on a subset of plots)
- soil water content and/or potential
- potential of hydraulic lift
2. Determine how plant species effects on multiple soil processes and properties change over
space and time.
a. Spatial variation
- by soil depth
- across sites (differing in soil, aspect, microclimate, management history,
etc.)

Spatial and Temporal Dependence of Plant Effects on Soils Across Multiple Scales: Building Mechanistic Understanding to Enhance Management Options—Eviner
3
b. Temporal variation
- daily (hydraulic lift)
- seasonal
- annual (fluctuations due to weather, etc.)
- with time since establishment of a community (trajectory of change over
years)
c. Environmental variation- manipulation of key variables that aid in
understanding fluctuations across space and time
- amount and timing of grazing/clipping
- N additions
- precipitation manipulations
Approach and Procedures
A suite of experimental and observational plots were studied to assess the impacts of vegetation
composition on multiple soil processes and properties. These include:
1. Experimental plantings of a native grass, dominant annual grasses (naturalized exotics)
and a mix of the two groups. At Hopland Research and Extension Center’s lysimeter facility,
we planted lysimeter tanks (38cm dia x 70 cm depth) with: Nassella pulchra (native perennial
bunchgrass), two dominant annual exotic grasses (Aegilops triuncialis, Taeniatherum caput-
medusae), and a mix of the native and exotics (8 replicates of each).
2. Experimental plantings of monocultures and mixtures of natives, exotic annual grasses,
and newer invasive annual weeds. On the campus agricultural fields, we planted 1.5 x 1.5 m
plots with:
Monocultures of the following species:
Exotic annuals (naturalized):
- Avena fatua
- Bromus hordeaceus
- Lolium multiflorum
- Trifolium subterraneum
Invasive weeds:
- Aegilops triuncialis
- Taeniatherum caput-medusae
Natives:
- Bromus carinatus
- Elymus glaucus
- Leymus triticoides
- Lotus purshianus
- Lupinus bicolor
- Nassella pulchra
- Vulpia microstachys

Spatial and Temporal Dependence of Plant Effects on Soils Across Multiple Scales: Building Mechanistic Understanding to Enhance Management Options—Eviner
4
In addition, we planted each of the above groups as monotypic mixes, and in all 2- and 3-way
combinations. These group plantings were exposed to clipping treatments (none, fall, spring) and
N fertilization (none, 45 kg N/ha/yr). (All plots- monocultures and mixtures will be exposed to
precipitation manipulations—but these treatments were delayed during this project’s duration,
due to rain shelter damage in a storm). There are 8 replicates of each treatment.
3. Observational studies across the landscape of areas restored with native species vs. adjacent
unrestored areas (dominated by annual exotic grasses). Across Yolo County, we sampled 11
native grassland restoration sites of varying ages, and adjacent unrestored areas.
4. Observational studies across the landscape of areas dominated by invasive weeds vs. those
dominated by naturalized annual exotics. Across the Willow Slough Watershed in Winters,
CA, we sampled 30 sites that were invaded by Aegilops and/or Taeniatherum, and adjacent
sites dominated by long-term annual exotics (e.g. Avena, Bromus, Lolium).
Measurements
1. Carbon
- Net primary productivity (aboveground and belowground) - clipping from
aboveground rings, and root cores down to 30 cm (experimental plots) and 100
cm (observational plots)
- Soil C by depth- Incremental soil sampling (0-15 cm, 15-30, 30-45, 45-60, 60-
90 depths) for bulk density and soil %C
- Soil organic matter by depth- combustion method
2. Soil cohesion (an indicator of surface erosion control)- using a torsional sheer vein tester on
the soil surface (and for experimental approach #1, with depth)
3. Nitrogen
- total soil N- %N and bulk density by depth (see depth increments for C)
- net N mineralization and nitrification rates- week-long aerobic incubations (0-
15 cm depth for all plots, additional samples at 15-30 cm depths in
experimental plots)
- seasonal resin available N- in experimental approach #2, placed at 5-10cm
depth
- N leaching loss- in plots from experimental approach #1—lysimeter leachate was
collected after each rainfall and analyzed for N
4. Soil water
- infiltration- disk infiltrometer for experimental approach #1, double-ring
infiltrometer for others
- water holding capacity- lab assay for soil at same depth increments as for C

Spatial and Temporal Dependence of Plant Effects on Soils Across Multiple Scales: Building Mechanistic Understanding to Enhance Management Options—Eviner
5
(above)
- leaching loss in plots from experimental approach #1—lysimeter leachate
was collected after each rainfall
- soil water content and/or potential- in experimental approach #2, TDR was
used to determine water content, psychrometers to determine water potential.
Gravimetric methods were used for other experimental approaches
- potential of hydraulic lift- in experimental approach #2- psychrometers at 30 cm
and 80 cm depths
Results
1. Experimental plantings of a native grass, dominant annual grasses (naturalized exotics)
and a mix of the two groups.
a. Soil Carbon
Compared with the native Nassella pulchra, exotic annuals had higher percent soil organic matter
(SOM) in the top 15 cm, and lower SOM at the 15-30 cm depth, leading to no difference in total
SOM content in the top 30 cm of soil (Figure 1). The mix of exotics and natives was not
significantly different from the exotics alone. Soil %C in the top 15 cm mirrors the results from
SOM (data not shown).

Spatial and Temporal Dependence of Plant Effects on Soils Across Multiple Scales: Building Mechanistic Understanding to Enhance Management Options—Eviner
6
Figure 1. Soil organic matter after 10 years of lysimeters planted to exotic grasses, the native Nassella pulchra, and a mix of exotics and the native. Percent soil organic matter in the top 15 cm (top panel) and 15-30 cm depth (middle panel), and SOM content from 0-30 cm (bottom panel).

Spatial and Temporal Dependence of Plant Effects on Soils Across Multiple Scales: Building Mechanistic Understanding to Enhance Management Options—Eviner
7
b. Soil cohesion
Vegetation type had no significant effect on soil cohesion at any soil depth (Figure 2).
Figure 2. Soil cohesion, by depth, after 10 years of soils being affected by the planting of exotic grasses, the native Nassella pulchra, and a mix of exotics and the natives.
c. Nitrogen
Soil percent nitrogen was higher for exotics than for natives, or natives + exotic mixes in the top
15 cm of soil (Figure 3). We are currently analyzing deeper soil depths.
Rates of net N mineralization were not significant (p=0.06), but soils in Nassella plots tended to
have higher rates than the mixtures and exotics (Figure 4). Net nitrification rates did significantly
differ (p=0.003), with Nassella having higher rates than exotics or the mix (Figure 4). We are
currently analyzing more samples to determine how these vegetation effects vary by depth, and
season, and to see how they relate to seasonal leaching loss patterns of N.
Figure 3. Soil %N in the top 15 cm of soil.

Spatial and Temporal Dependence of Plant Effects on Soils Across Multiple Scales: Building Mechanistic Understanding to Enhance Management Options—Eviner
8
Figure 4. Rates of net N mineralization (top panel) and nitrification (bottom panel) in top 15 cm of soil.

Spatial and Temporal Dependence of Plant Effects on Soils Across Multiple Scales: Building Mechanistic Understanding to Enhance Management Options—Eviner
9
d. Water
Early in the growing season, water leaching losses were consistently lower in the native Nassella
plots than in the exotics, and even the unvegetated plots. There were no, or only small
differences in water leached from vegetation treatments over the rest of the growing season
(Figure 5). Infiltration rates were also slower for Nassella plots than for the exotics at the soil
surface, but not over the rest of the soil depth profile (Figure 6).
2. Experimental plantings of monocultures and mixtures of natives, exotic annual grasses,
and newer invasive annual weeds. a. Carbon- Samples are currently being processed for aboveground and belowground
biomass, and soil C with depth .
b. Soil cohesion- Soil cohesion is currently being analyzed, and there were no significant
differences in vegetation type on soil compaction with depth (data not shown).
c. Nitrogen- Seasonal plant N, net mineralization and nitrification rates, and resin-
available N are currently being analyzed.
d. Water- Infiltration rates differed across species, and were relatively high in Lolium
plots and low in Aegilops and Elymus plots. There were no significant differences across broad
groups (weeds vs. annuals vs. natives) or their mixtures (Figure 7).
Figure 5. Amount and timing of leachates from lysimeters planted with different vegetation types. Top panel is 2005-6 growing season bottom panel is 2006-7 growing season
Figure 6.
Spatial and Temporal Dependence of Plant Effects on Soils Across Multiple Scales: Building Mechanistic Understanding to Enhance Management Options—Eviner
10
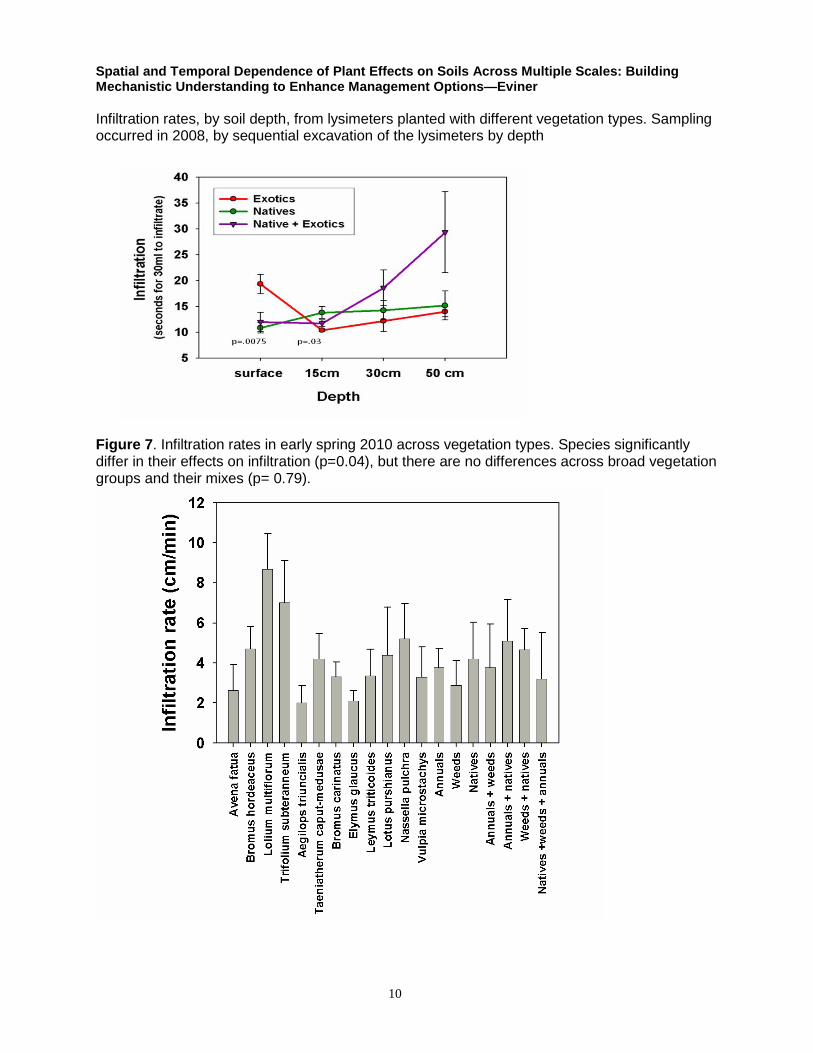
Infiltration rates, by soil depth, from lysimeters planted with different vegetation types. Sampling occurred in 2008, by sequential excavation of the lysimeters by depth
Figure 7. Infiltration rates in early spring 2010 across vegetation types. Species significantly differ in their effects on infiltration (p=0.04), but there are no differences across broad vegetation groups and their mixes (p= 0.79).
Spatial and Temporal Dependence of Plant Effects on Soils Across Multiple Scales: Building Mechanistic Understanding to Enhance Management Options—Eviner
11
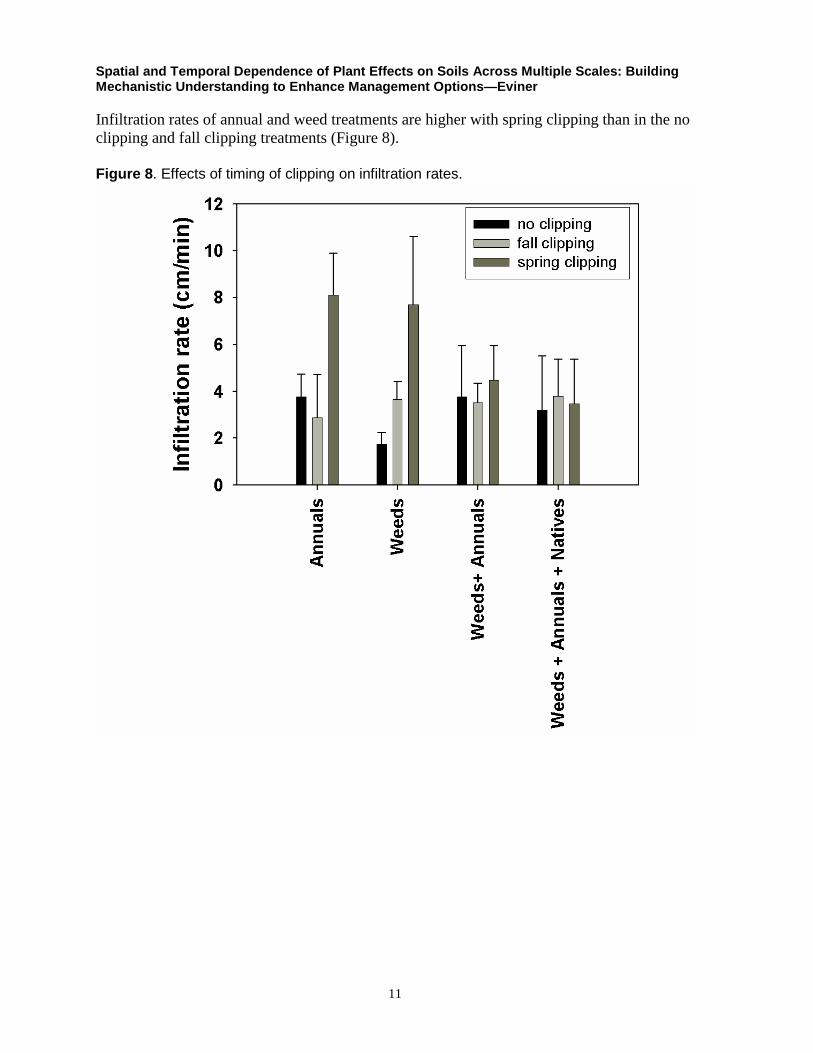
Infiltration rates of annual and weed treatments are higher with spring clipping than in the no
clipping and fall clipping treatments (Figure 8).
Figure 8. Effects of timing of clipping on infiltration rates.

Spatial and Temporal Dependence of Plant Effects on Soils Across Multiple Scales: Building Mechanistic Understanding to Enhance Management Options—Eviner
12
Potential for hydraulic lift was assessed only in the native plots, since these species are the most
active during the dry summers. Psychrometer readings at 30 vs. 80 cm depths were compared at
different times of the day, indicating significant increases in water potential as soon as plants are
exposed to shade (1830 to 1945 hours), with significant drawdown at dawn, particularly at the 30
cm depth (Figure 9). Currently, in the summer of 2010, we are assessing how these daily
moisture fluctuations impact N cycling rates across vegetation treatments.
Figure 9. % change in psychrometer readings between daily time points indicate likely hydraulic lift in native plots.
Spatial and Temporal Dependence of Plant Effects on Soils Across Multiple Scales: Building Mechanistic Understanding to Enhance Management Options—Eviner
13
3. Observational studies across the landscape of areas restored with native species vs. adjacent
unrestored areas (dominated by annual exotic grasses).
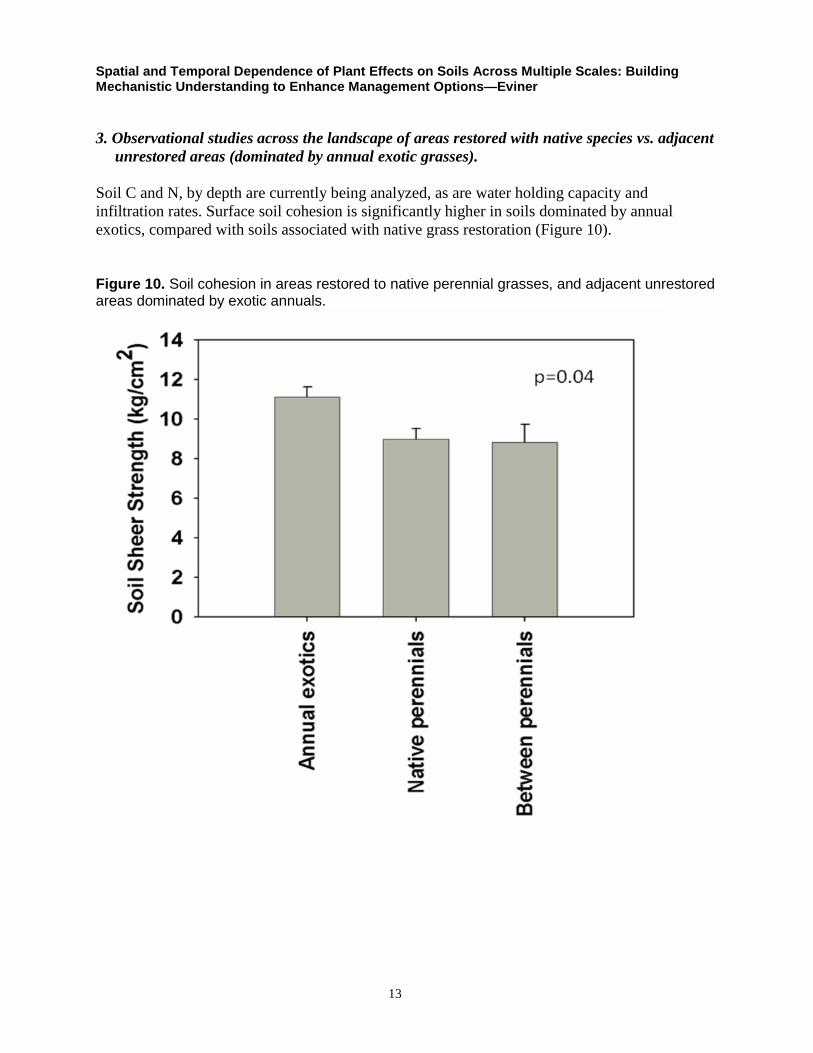
Soil C and N, by depth are currently being analyzed, as are water holding capacity and
infiltration rates. Surface soil cohesion is significantly higher in soils dominated by annual
exotics, compared with soils associated with native grass restoration (Figure 10).
Figure 10. Soil cohesion in areas restored to native perennial grasses, and adjacent unrestored areas dominated by exotic annuals.
Spatial and Temporal Dependence of Plant Effects on Soils Across Multiple Scales: Building Mechanistic Understanding to Enhance Management Options—Eviner
14
4. Observational studies across the landscape of areas dominated by invasive weeds vs. those
dominated by naturalized annual exotics.
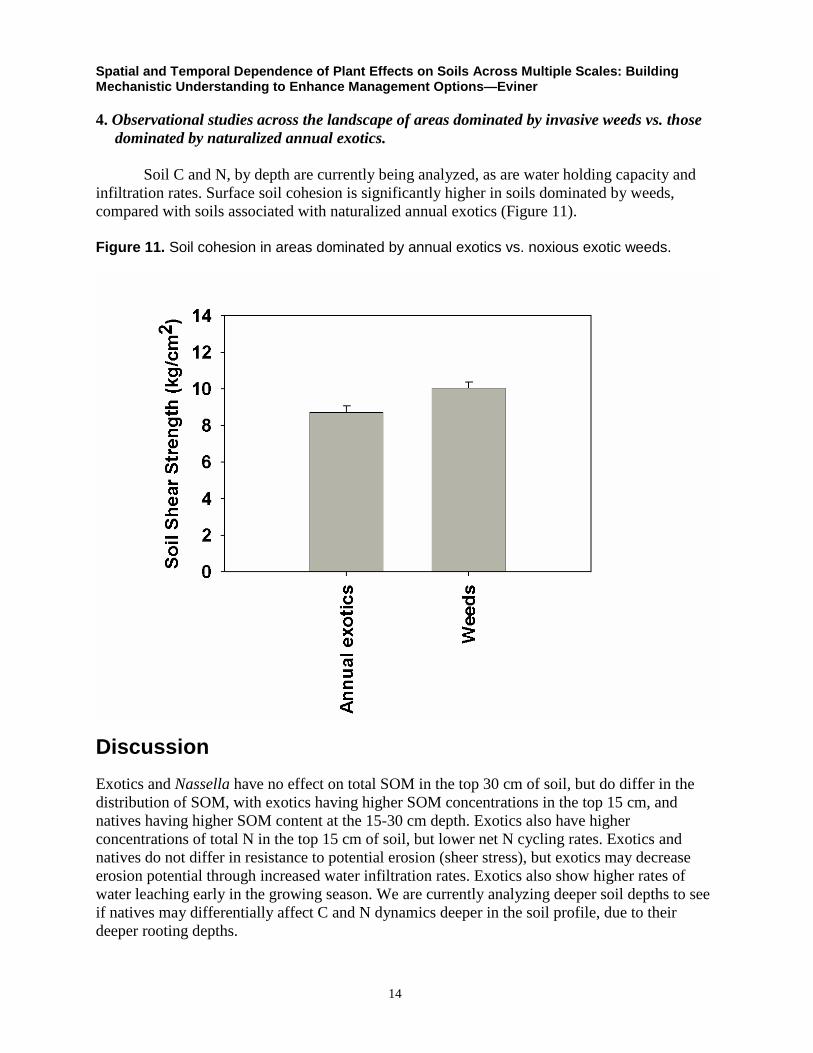
Soil C and N, by depth are currently being analyzed, as are water holding capacity and
infiltration rates. Surface soil cohesion is significantly higher in soils dominated by weeds,
compared with soils associated with naturalized annual exotics (Figure 11).
Figure 11. Soil cohesion in areas dominated by annual exotics vs. noxious exotic weeds.
Discussion
Exotics and Nassella have no effect on total SOM in the top 30 cm of soil, but do differ in the
distribution of SOM, with exotics having higher SOM concentrations in the top 15 cm, and
natives having higher SOM content at the 15-30 cm depth. Exotics also have higher
concentrations of total N in the top 15 cm of soil, but lower net N cycling rates. Exotics and
natives do not differ in resistance to potential erosion (sheer stress), but exotics may decrease
erosion potential through increased water infiltration rates. Exotics also show higher rates of
water leaching early in the growing season. We are currently analyzing deeper soil depths to see
if natives may differentially affect C and N dynamics deeper in the soil profile, due to their
deeper rooting depths.

Spatial and Temporal Dependence of Plant Effects on Soils Across Multiple Scales: Building Mechanistic Understanding to Enhance Management Options—Eviner
15
Did not see some of these effects in experimental plots in Davis—likely due to the shorter
duration of the plantings (3 vs. 10 years), but also potentially due to differences in climate and
soils between sites.
While most of this work is on-going, the conclusions we can draw thus far highlight that
species effects on multiple soil processes and properties are independent, so that there are some
benefits and detriments associated with all vegetation groups. For example, while native
grassland species increase deeper soil carbon, they delay water leaching at the start of the
growing season, and have lower soil surface cohesion than the exotic annuals that have displaced
them. While the noxious invasives, Aegilops triuncialis and Taeniatherum caput-medusae
greatly decrease rangeland productivity, they also may decrease surface soil erosion.
Another key conclusion is that the impacts of species can vary depending on time since
establishment, site, and grazing patterns. Many other studies have demonstrated that the
ecosystem effects of a given species are not constant. Both the values and relative ranking of
species effects on various soil properties and processes change across sites (Lovett and Rueth
1999, Scheffer et al. 2001, Kalburtji and Mamolas 2000, Verchot et al. 2001, Bridgham and
Richardson 2003). These variations in the soil effects of a given species can be as great as the
variation across different species (Bridgham and Richardson 2003, Eviner et al. 2006, Eviner and
Hawkes 2008). As we wrap-up analyses of samples that we have collected in this project, we
will assess the relative importance of the following mechanisms in determining the context-
dependence of plant species effects on soils:
1. Plant traits change in response to changes in environmental conditions and time, and
these shifts in traits directly alter soil processes (can use current predictive frameworks
linking multiple traits with soil processes, just shifting the values of species traits). For
example, in California’s grasslands, seasonal changes in root C inputs and the chemistry of
remaining litter are partial drivers of seasonal changes in plant species effects (Eviner et al.
2006).
2. The relative importance of traits in determining soil processes changes with
environmental conditions and time (shifts in which traits are used to predict soil effects).
For example, in California’s grasslands, species effects on net N mineralization are driven by
species effects on soil temperature and litter C in the fall, by root C inputs in the winter, and
by litter chemistry and species effects on soil moisture in the spring (Eviner et al. 2006).
3. There is a fundamental change in how a trait affects a soil process over environmental
gradients and time (a shift in the relationship between a trait and a soil process). For
example, the relationship between decomposition and litter lignin differs at low versus high
actual evapotranspiration (AET) (Meetenmeyer 1978), while the relationship between litter
quality and decomposition varies with nutrient availability (Hobbie 2000, Vesterdal 1999).
Relevance to Kearney Mission/Land Management and Policies
Understanding the interaction of environmental heterogeneity and plant community impacts on
soil processes is a critical tool for management of ecosystem services, agricultural productivity

Spatial and Temporal Dependence of Plant Effects on Soils Across Multiple Scales: Building Mechanistic Understanding to Enhance Management Options—Eviner
16
and control of noxious weeds. In this research, we address Kearney’s current mission by
developing a mechanistic understanding of plants impacts on multiple soil processes, and how
these impacts change across time (ranging from daily to decadal) and space (ranging from soil
depth profiles to broad landscape patterns). Our data will enable us to develop an ecological
toolbox.
References:
Bridgham, S. D. and C. J. Richardson. 2003. Endogenous versus exogenous nutrient control over
decomposition and mineralization in North Carolina peatlands. Biogeochemistry. 65:151-178.
Coleman, J., K. Hench, K. Garbutt, A. Sexstone, G. Bissonnette, and J. Skousen. 2001.
Treatment of domestic wastewater by three plant species in constructed wetlands.
Drinkwater, L. E. 1999. Using plant species composition to restore soil quality and ecosystem
function. In: Olesen, JE, MJ Gooding and J Kopke (eds) International Workshop on
Designing and Testing Crop Rotation for Organic Farming. Sponsored by the Danish
Institute of Agricultural Sciences. Borris Landbrugsskole. Danish Research Centre for
Organic Farming, Tjele. pp. 37-46.
Eviner, V. and F. Chapin III. 2001. Plant species provide vital ecosystem functions for
sustainable agriculture, rangeland management, and restoration. California Agriculture.
55:54-59.
Eviner V.T. and F.S. Chapin III 2003. Functional matrix: A conceptual framework for predicting
multiple plant effects on ecosystem processes. Annual Review Ecology and Systematics,
34:455-485.
Eviner V.T., Chapin III F.S. and Vaughn C.E. 2006. Seasonal variations in species effects on N
and P cycling. Ecology, 87:974-986.
Eviner, V.T. and C.V. Hawkes. 2008. Embracing variability in the application of plant-soil
interactions to the restoration of communities and ecosystems. Restoration Ecology 16:713-
729.
Hobbie, S. E. 2000. Interactions between litter lignin and soil nitrogen availability during leaf
litter decomposition in a Hawaiian Montane forest. Ecosystems. 3:484-494.
Kalburtji, K. L. and A. P. Mamolos. 2000. Maize, soybean and sunflower litter dynamics in two
physicochemically different soils. Nutrient Cycling in Agroecosystems. 57:195-206
Lavelle, P. 2000. Ecological challenges for soil science. Soil Science. 165:73-86.
Lovett, G. M. and H. Rueth. 1999. Soil nitrogen transformations in beech and maple stands along
a nitrogen deposition gradient. Ecological Applications. 9:1330-1344.
Meentemeyer, V. 1978. Macroclimate and lignin control of litter decomposition rates. Ecology.
59:465-472.
Rhoades, C. C., T. M. Nissen, and J. S. Kettler. 1997. Soil nitrogen dynamics in alley cropping
and no-till systems on Ultisols of the Georgia Piedmont, USA. Agroforestry Systems. 39:31-
44.
Sarrantonio, M. 1994. Northeast Cover Crop Handbook. Rodale Institute, Emmaus, PA.

Spatial and Temporal Dependence of Plant Effects on Soils Across Multiple Scales: Building Mechanistic Understanding to Enhance Management Options—Eviner
17
Scheffer, R. A., R. S. P. van Logtestijn, and J. T. A. Verhoeven. 2001. Decomposition of Carex
and Sphagnum litter in two mesotrophic fens differing in dominant plant species. Oikos.
92:44-54.
Verchot, L. V., Z. Holmes, L. Mulon, P. M. Groffman, and G. M. Lovett. 2001. Gross vs. net
rates of N mineralization and nitrification as indicators of functional differences between
forest types. Soil Biology & Biochemistry. 33:1889-1901.
Vesterdal, L. 1999. Influence of soil type on mass loss and nutrient release from decomposing
foliage litter of beech and Norway spruce. Canadian Journal of Forest Research-Revue
Canadienne de Recherche Forestiere. 29:95-105.
Wardle, D. 2002. Communities and ecosystems: linking the aboveground and belowground
components: Princeton University Press Princeton.392.
This research was funded by the Kearney Foundation of Soil Science: Understanding and Managing Soil-Ecosystem Functions Across Spatial and Temporal Scales, 2006-2011 Mission (http://kearney.ucdavis.edu). The Kearney Foundation is an endowed research program created to encourage and support research in the fields of soil, plant nutrition, and water science within the Division of Agriculture and Natural Resources of the University of California.