17β-estradiol (βe2) protects human retinal müller cell against oxidative stress in vitro:...
TRANSCRIPT

17b-Estradiol (bE2) Protects Human Retinal M€uller CellAgainst Oxidative Stress In Vitro: Evaluation of ItsEffects on Gene Expression by cDNA MicroarrayCHAO LI,1 YUHONG TANG,2 FENG LI,1 SEAN TURNER,2 KONG LI,1,3 XIAOHONG ZHOU,1
MICHAEL CENTOLA,2 XIAORONG YAN,1 AND WEI CAO1*1Department of Ophthalmology, University of Oklahoma Health Science Center, Dean A. McGee Eye Institute,608 Stanton L. Young Blvd, Oklahoma City, Oklahoma2Microarray Research Facility, Oklahoma Medical Research Foundation, Oklahoma City, Oklahoma3Department of Histoembryology of Dalian Medical University, Dalian, Liaoning, People’s Republic of China
KEY WORDSestrogen; M€uller cells; microarray; gene expression; real-time PCR; hydrogen peroxide; apoptosis; protection
ABSTRACT17b-estradiol (bE2) is an effective neuroprotectant againsthydrogen peroxide (H2O2)-induced retinal neuronal cell deathand light-induced photoreceptor degeneration. M€uller cells arethe principal macroglia responsible for supporting retinal neu-ronal survival, information processing and removing metabolicwaste. However, the role of bE2 on human M€uller cells isunclear. In this study, the effects of bE2 on human M€uller cellsurvival and gene expression were examined. Our data re-vealed that bE2 is able to increase human M€uller cell viabilityafter exposure to H2O2 through inhibition of apoptosis. Micro-array analysis revealed significant changes in the expression of69 genes (total of 21,324 genes screened) in cultured humanM€uller cells 6 h after bE2 treatment. Four of the bE2-respon-sive genes [thrombospondin 1 (TSP1), mitogen-activated pro-tein kinase kinase kinase 3 (MAP3K3), large conductance cal-cium-activated potassium channel b2 subunit (KCNMB2), andSRY (sex-determining region Y)-box 11 (SOX11)] were vali-dated by both real-time qRT-PCR and semi-quantitative RT-PCR. Interestingly, exposure of human M€uller cells to bE2 in-creased pigment epithelium-derived factor (PEDF) gene expres-sion as measured by both RT-PCR and real time qRT-PCR. Ourdata demonstrate, for the first time, that bE2 protects culturedhuman M€uller cells against H2O2-induced cell death throughthe inhibition of apoptosis. This protective effect may operatethrough regulation of genes, such as TSP1, MAP3K3, SOX11,TSP1, and PEDF, and may in turn exert an important role inprotecting retinal neurons. VVC 2005Wiley-Liss, Inc.
INTRODUCTION
17b-Estradiol (bE2) is a steroid hormone synthesizedin the ovary, placenta, adrenal gland, testes, and periph-eral tissues. In addition, estrogen is not restricted tofemales because the male sex hormone testosterone (andother 19-carbon steroids) can be converted chemically tobE2 in various tissues, including the brain, by an aroma-tase P-450 enzyme (Schumacher et al., 2002). Severallines of evidence suggest that estrogen has neurotrophicand neuroprotective properties (Garcia-Segura et al.,2001; Wise, 2002). bE2 exerts its neuroprotective effect
by, both nuclear estrogen receptor-dependent and -inde-pendent activities (Amentea et al., 2005). We have pre-viously demonstrated that (1) bE2 protects significantlyagainst H2O2-induced retinal neuron degeneration throughactivation of the phosphoinositide 3-kinase/Akt signalingpathway, and (2) bE2 treatments results in phosphorylationof insulin receptor b-subunit by an unidentified mechanism(Cao et al., 2003). It has also been shown that systemicadministration of bE2 significantly protected structure andfunction of rat retinas against light-induced photoreceptorcell degeneration and inhibited photoreceptor apoptosis(Yu et al., 2004).
M€uller cells radially traverse the retina from its innerborder to the distal end of the outer nuclear layer contact-ing and ensheathing every type of neuronal cell body.M€uller cells are responsible for the structural stabilizationof the retina and the maintenance of the layered arrange-ment of retinal neurons (Willbold et al., 1997), and playcrucial roles in supporting neuronal survival and informa-tion processing (Reichenbach et al., 1993; Newman andReichenbach, 1996) by providing trophic substances toneurons and removing metabolic waste (Poitry, et al.,2000). M€uller cells metabolize glucose to lactate, which istaken up by photoreceptors as an energy source (Poitry-Yamate et al., 1995), and regulate the extracellular home-ostasis of ions, pH, and water (Newman, 1993; Newmanet al., 1984). Selective M€uller cell death results in retinaldysplasia, photoreceptor apoptosis, and retinal degenera-tion (Dubois-Dauphin et al., 2000).
It is well established that steroids synthesized by theneural system, mainly by glial cells, can act directly onmembrane receptors for neurotransmitters (Sierra, 2004).
Grant sponsor: National Center for Research Sources (COBRE program); Grantnumber: P20 RR17703; Grant sponsor: National Institutes of Health; Grant num-ber: EY014427; Grant number: EY12190; National Institutes of Health; Grantnumber: P20 RR16478-04; Grant number: P20 RR020143; Grant number: P20RR15577; Grant number: NIH 01700172; Grant sponsor: OCAST; Grant number:HR04-110F.
*Correspondence to: Wei Cao, Department of Ophthalmology, University of Okla-homa Health Sciences Center, Dean A. McGee Eye Institute, 608 Stanton L. YoungBlvd, Oklahoma City, OK 73104. E-mail: [email protected]
Received 29 April 2005; Accepted 13 September 2005
DOI 10.1002/glia.20291
Published online 8 December 2005 in Wiley InterScience (www.interscience.wiley.com).
VVC 2005 Wiley-Liss, Inc.
GLIA 53:392–400 (2006)

More recently, several lines of investigation have shownthat estrogen exerts an anti-inflammatory activity onprimary cultures of rat microglia (Vegeto et al., 2001),enhances the uptake of amyloid b-protein by microgliaderived from the human cortex (Li et al., 2000),increases the expression of ApoE in astrocytes andmicroglia (Stone et al., 1997), and reduces activation ofnuclear factor jB (NFjB) induced by amyloid A b andlipopolysaccharide in cultured astrocytes (Dodel et al.,1999). In addition, because estradiol increases glutamateuptake in astrocytes derived from Alzheimer’s patients,it may contribute to the potential protective effectagainst this neurodegenerative disease (Liang et al.,2002). Perhaps the most important mechanism involvedin estrogen-mediated neuroprotection through glia is therelease of factors that promote neuronal survival. How-ever, the effects of bE2 on M€uller cells, the principalglial cells in the retina, are yet to be explored.
Hydrogen peroxide (H2O2), a by-product of oxidativestress, can trigger apoptosis in various types of cells,including glial cells (Clement et al., 1998; Kitamuraet al., 1999; Cao et al., 1999). To understand the role ofbE2 in the protection of M€uller cells, the transcriptionalresponse of human M€uller cells to bE2 treatment wasstudied using microarray technology, and the effects ofbE2 on protecting human M€uller cells against H2O2-induced cell death were also investigated.
MATERIALS AND METHODSHuman M€uller Cell Culture
Human M€uller cells (Institute of Ophthalmology, Lon-don, UK) were grown in Dulbecco’s modified Eagle’smedium (DMEM; Invitrogen, Carlsbad, CA), which didnot contain estrogenic compound phenol red. The me-dium was supplemented with 2 mM glutamine, 10 IU/mlpenicillin, 10 lg/ml streptomycin, and 10% heat-inacti-vated fetal calf serum (FCS; Invitrogen). The isolationand characterization of these cells is described elsewhere
(Limb et al., 2002). Cells were grown to confluence in anincubator with a humidified atmosphere of 5% CO2, 95%air at 37�C and were trypsinized and seeded into 6-wellflat-bottomed plates (Falcon, Fort Worth, TX) containing3 ml of the same medium. After 48 h of incubation, eachwell contained �105–106 cells, and various treatmentswere conducted with cells at this stage.
MTT Assay
The MTT assay was performed as described previously(Cao et al., 1999).
Flowcytometric Detection of M€uller Cell Apoptosis
Cultured human M€uller cells were treated with PBS,H2O2 (200 lM), bE2 (100 nM) 1 H2O2 (200 lM), bE2
(100 nM), respectively. Stocks of 10 lM bE2 and 20 mMH2O2 were prepared in DMEM and added to the 3 mlcell culture as needed. M€uller cells were collected at 6,12, 18, and 24 h after treatment. For bE2 (100 nM) 1H2O2 (200 lM) treatment, H2O2 was added to the cul-ture 30 minutes after addition of bE2. Cells were col-lected after trypsin digestion, and apoptosis was deter-mined by flowcytometry using Annexin V-FITC kit(Beckman Coulter, Fullerton, CA) according to the man-ufacturer’s instructions. Briefly, human M€uller cells(105/ml) were washed with phosphate-buffered saline(PBS) and resuspended in binding buffer before staining,in darkness, with 2 ll annexin V (0.5 lg/ml) and 10 llpropidium iodide (0.6 lg/ml) for 10 min at room tem-perature. After staining, cells were analyzed immedi-ately using a FACScan flowcytometer (Beckman Coulter)with simultaneous monitoring of green fluorescence(530 nm, 30-nm bandpass filter) for annexin V-FITC andred fluorescence (long-pass emission filter that transmitslight >650 nm) for propidium iodide. A total of 30,000events were collected and analyzed.
Microarray Analysis of mRNA of bE2-TreatedM€uller Cells
Microarray analysis was performed as described pre-viously (Yu et al., 2005). The transcriptional response ofM€uller cells to bE2 treatment was examined at six timepoints (0, 0.5, 1, 2, 3 and 6 h) after bE2 treatment. Thespecifications of the oligo arrays were the same as pre-viously described (Jarvis et al., 2004). Two biologic repli-cates were used for each treatment. RNA isolation fromcultured human M€uller cells was carried out by the Tri-zol method according to the manufacturer’s instructions(Invitrogen). RNA was quantified spectrophotometricallyand RNA quality was assessed by the fluorescence inten-sity ratio of 28s: 18s rRNA resolved in 1.2% denaturing-agarose gel. cDNA labeling, hybridization, and dataquantification of the microarray experiments were thesame as previously described (Jarvis et al., 2004). Selec-
Abbreviations used:
bE2 17b-estradiolH2O2 hydrogen peroxidePBS phosphate buffered salineSDS sodium dodecyl sulfateMTT 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium
bromideSD standard deviationANOVA analysis of variance between groupsPEDF pigment epithelium-derived factorTSP1 thrombospondin 1MAP3K3 mitogen-activated protein kinase/extracellular-
regulated kinase kinase kinase-3KCNMB2 large conductance calcium-activated potassium
channel b2 subunitSOX11 SRY (sex-determining region Y)-box 11RT-PCR reverse transcriptase-polymerase chain reactionreal-time
qRT-PCR real-time quantitative RT-PCRVEGF vascular endothelial growth factorGAPDH glyceraldehyde-3-phosphate dehydrogenaseBK channel large conductance Ca21- and voltage-dependent
potassium channel.
393EFFECT OF ESTROGEN ON RETINAL M €ULLER CELLS
tion of ‘‘hypervariable’’ genes as shown previously (Doz-morov et al., 2004). Genes determined to be hypervari-able were tested against their 0-h controls. Genes withat least two time points of expression level, above/below2 times of the 0-h controls, are classified as up/downre-gulated accordingly. For each gene, the ratios against0-h control for each of the five time points were used forWard’s clustering analysis, using Spotfire Decision Site8.1 (Spotfire, Somerville, MA).
Real-Time Quantitative Reverse Transcription(qRT)-PCR
Real-time quantitative reverse transcription-polymer-ase chain reaction (qRT-PCR) was performed as pre-viously described (Yu et al., 2005; Zhou et al., 2005).Four genes were chosen for confirmation on the basis oftheir functional importance. Five biologic replicates wereused for each group, and RNAs were harvested from cul-tured human M€uller cells under similar treatments asused for the microarray study. Fold changes (mean 6 SD)were calculated from five independent replicate groups.
Semi-quantitative Reverse Transcription(RT)-PCR Analysis
Semi-quantitative RT-PCR was performed as previ-ously described (Yu et al., 2005; Zhou et al., 2005).Briefly, first-strand cDNA was synthesized as describedfor real-time qRT-PCR. The same primer pairs (Table 1)used for real-time qRT-PCR were also used for semi-quantitative RT-PCR.
Statistical Analysis
For bE2 and/or H2O2 treatments, percentage (mean 6SD) of cell viability and apoptosis to the control wereobtained from 3–5 biologic replicates. Differences be-tween treatments or treatment vs. control were analyzedby one-way analysis of variance (ANOVA), followed by aBonferroni multiple-comparison post hoc test. P-valuesof <0.05 were considered significant.
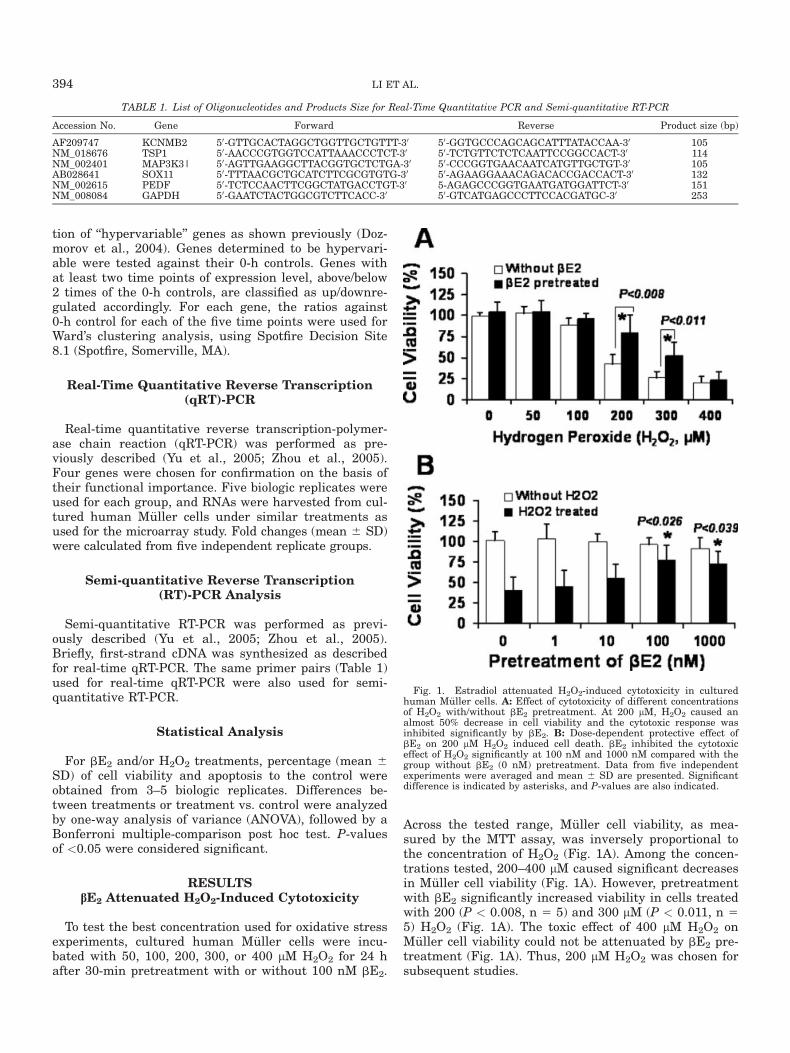
RESULTSbE2 Attenuated H2O2-Induced Cytotoxicity
To test the best concentration used for oxidative stressexperiments, cultured human M€uller cells were incu-bated with 50, 100, 200, 300, or 400 lM H2O2 for 24 hafter 30-min pretreatment with or without 100 nM bE2.
Across the tested range, M€uller cell viability, as mea-sured by the MTT assay, was inversely proportional tothe concentration of H2O2 (Fig. 1A). Among the concen-trations tested, 200–400 lM caused significant decreasesin M€uller cell viability (Fig. 1A). However, pretreatmentwith bE2 significantly increased viability in cells treatedwith 200 (P < 0.008, n 5 5) and 300 lM (P < 0.011, n 55) H2O2 (Fig. 1A). The toxic effect of 400 lM H2O2 onM€uller cell viability could not be attenuated by bE2 pre-treatment (Fig. 1A). Thus, 200 lM H2O2 was chosen forsubsequent studies.
TABLE 1. List of Oligonucleotides and Products Size for Real-Time Quantitative PCR and Semi-quantitative RT-PCR
Accession No. Gene Forward Reverse Product size (bp)
AF209747 KCNMB2 50-GTTGCACTAGGCTGGTTGCTGTTT-30 50-GGTGCCCAGCAGCATTTATACCAA-30 105NM_018676 TSP1 50-AACCCGTGGTCCATTAAACCCTCT-30 50-TCTGTTCTCTCAATTCCGGCCACT-30 114NM_002401 MAP3K3| 50-AGTTGAAGGCTTACGGTGCTCTGA-30 50-CCCGGTGAACAATCATGTTGCTGT-30 105AB028641 SOX11 50-TTTAACGCTGCATCTTCGCGTGTG-30 50-AGAAGGAAACAGACACCGACCACT-30 132NM_002615 PEDF 50-TCTCCAACTTCGGCTATGACCTGT-30 5-AGAGCCCGGTGAATGATGGATTCT-30 151NM_008084 GAPDH 50-GAATCTACTGGCGTCTTCACC-30 50-GTCATGAGCCCTTCCACGATGC-30 253
Fig. 1. Estradiol attenuated H2O2-induced cytotoxicity in culturedhuman M€uller cells. A: Effect of cytotoxicity of different concentrationsof H2O2 with/without bE2 pretreatment. At 200 lM, H2O2 caused analmost 50% decrease in cell viability and the cytotoxic response wasinhibited significantly by bE2. B: Dose-dependent protective effect ofbE2 on 200 lM H2O2 induced cell death. bE2 inhibited the cytotoxiceffect of H2O2 significantly at 100 nM and 1000 nM compared with thegroup without bE2 (0 nM) pretreatment. Data from five independentexperiments were averaged and mean 6 SD are presented. Significantdifference is indicated by asterisks, and P-values are also indicated.
394 LI ET AL.

We then determined the effect of pretreatment with in-creasing concentrations of bE2 on H2O2-induced cell death.As shown in Figure 1B, treatment with various concentra-tions of bE2 alone did not affect cell viability. However,when combined with 200 lM H2O2 treatment, there was adose-dependent protective effect of bE2 treatment (Fig. 1B).Cell viability increased from 40% without to 45%, 60%, 75%and 70% with bE2 pre-treatment at the concentrations of1, 10, 100 and 1000 nM, respectively (Fig. 1B). Pretreat-ment with 100 and 1000 nM bE2 resulted in significantincreases in cell viability when compared cells that werenot treated with bE2. Pre-treatment with 100 nM bE2 pro-vided the best protective effect and was thus used for sub-sequent studies.
Dynamics of bE2 in Protecting AgainstH2O2-Induced Cultured Human
M€uller Cell Apoptosis
To examine further whether the effect of bE2 on in-creasing cell viability is through inhibition of apoptosis,we assessed apoptosis in human M€uller cells treatedwith PBS (control), 100 nM bE2, 100 nM bE2 for 30 minbefore 200 lM H2O2, and 200 lM H2O2 alone. Represen-tative flowcytometry images of 12-h treatment areshown for both control (Fig. 2A) and H2O2 treated sam-ples (Fig. 2B). (Fig. 2A, C1) and E1 (Fig. 2B, E1) repre-sent the portion of cells that underwent apoptosis. Asshown in Figure 2, H2O2 treatment greatly increasedthe apoptosis in the cultured human M€uller cells. Whendata from three independent experiments were compiledand analyzed quantitatively (Fig. 2C), we found thatpretreatment with 100 nM bE2 significantly decreasedthe percentage apoptotic cells treated for 12 h with 200lM H2O2. Treatment with bE2 alone did not signifi-cantly affect the percentage apoptotic cells. When trea-ted with H2O2 alone, the percentage apoptotic M€ullercells peaked at 12 h (18%) and decreased to 10% at 18–24 h after treatments. Pre-treatment of cultured humanM€uller cells with 100 nM bE2 for 30 min reduced cellapoptosis induced by H2O2 significantly at 12 h (Fig. 2C,P < 0.01, n 5 3), 18 h (Fig. 2C, P < 0.01, n 5 3), and 24h (Fig. 2C, P < 0.05, n 5 3), but no significant differencewas observed at 6 h after initial treatment.
Identification of Genes Responsive to bE2
Treatment by Microarray Analysis
To identify genes that could be involved in the anti-apoptotic effects of bE2, cultured human M€uller cellswere exposed to 100 nM bE2 for different periods of time(0, 0.5, 1, 2, 3, and 6 h) and changes in gene expressionwere analyzed by microarray analysis. Because the effectsof treatment on transcription occurred earlier than actualphysiological changes, gene expression was studied at ear-lier time points (Fig. 2C).
Using a selection procedure established previously(Dozmorov et al., 2004), genes that responded to bE2
treatment were selected as ‘‘hypervariable’’ genes. From
the 21,324 genes on the array, 69 genes were selected as‘‘hypervariable genes’’. Detailed information about thesegenes is presented in Table 2. Among these 69 genes, 33were upregulated, 12 genes were downregulated, and 24genes were of mixed patterns. Thirty-four genes thathad known annotation information belonged to variouscategories, including genes involved in the formation ofextracellular matrix, signal transduction, cell growth,transcription, apoptosis, and intracellular transport.
Using a hierarchical clustering program in Spotfire De-cision Site 8.1, the clustering patterns of the 69 hyper-variable genes were determined according to Ward’s hier-archical clustering algorithm (cite a reference here). Foreach gene, ratios against 0-h control from five treatmentswere used in clustering analysis. Thus, genes that clus-tered together have very similar dynamic patterns. A‘‘heat map’’ of changes of these genes and their descrip-tions are presented in Figure 3. Known and unknowngenes are spread throughout the heat map. Significantlymore genes were upregulated by bE2.
Fig. 2. Protective effect of bE2 on H2O2-induced apoptosis in cul-tured M€uller cells sampled at four time points: 6, 12, 18, and 24 h aftertreatment. Flowcytometry images of M€uller cells 12 h after treatment:control group treated with PBS only (A); samples treated with H2O2
(B). C1: E1, portion of cells that underwent apoptosis. C2: E2, portionof cells that died of secondary necrosis. C3: E3, live cells. C4: E4, necro-tic cells (C). Bar graph presentation of cultured human M€uller cellapoptotic rate under four different treatment conditions at four timepoints. Data for three biologic replicates are obtained and mean 6 SDof apoptotic rate are presented. Significant difference is indicated byasterisks; P-values are also presented.
395EFFECT OF ESTROGEN ON RETINAL M €ULLER CELLS

TABLE 2. Hypervariable Genes Responsive to bE2 Treatment in Cultured Human M€uller Cells*
Accession No. Gene symbol Description Cluster order Cluster
NM_018242 FLJ10847 Hypothetical protein FLJ10847 1 1NM_017851 FLJ20509 Hypothetical protein FLJ20509 2 1NM_006410 HTATIP2 HIV-1 Tat interactive protein 2, 30 kD 3 1NM_001099 ACPP Acid phosphatase, prostate 4 1NM_003368 USP1 Ubiquitin-specific protease 1 5 1BC016919 Homo sapiens, clone MGC:21130
IMAGE:4400601, mRNA, complete cds6 1
NM_033089 C20orf99 Chromosome 20 open reading frame 99 7 1NM_001536 HRMT1L2 HMT1 hnRNP methyltransferase-like 2
(S. cerevisiae)8 1
AK056512 FLJ22625 Hypothetical protein FLJ22625 9 1AK026853 Homo sapiens cDNA: FLJ23200 fis,
clone KAIA3887110 1
NM_014254 TMEM5 Transmembrane protein 5 11 1NM_000856 GUCY1A3 Guanylate cyclase 1, soluble, a3 12 1AK024242 Homo sapiens cDNA FLJ14180 fis,
clone NT2RP200379913 2
NM_021080 DAB1 Disabled homologue 1 (Drosophila) 14 2AK055980 Homo sapiens cDNA FLJ31418 fis,
clone NT2NE200035315 2
NM_014385 SIGLEC7 Sialic acid-binding Ig-like lectin 7 16 2NM_013994 DDR1 Discoidin domain receptor family, member 1 17 2NM_012119 CCRK Cell cycle-related kinase 18 2AL137279 Homo sapiens mRNA- cDNA DKFZp434O1214
(from clone DKFZp434O1214)19 2
BC018537 Homo sapiens, Similar to RIKEN cDNA1300003P13 gene, clone MGC:16810IMAGE:4340152, mRNA, complete c
20 2
NM_004979 KCND1 Potassium voltage-gated channel,Shal-related subfamily, member 1
21 3
NM_003007 SEMG1 Semenogelin I 22 3NM_002763 PROX1 Prospero-related homeobox 1 23 3AK001558 Homo sapiens cDNA FLJ10696 fis,
clone NT2RP300048424 3
AB011106 KIAA0534 KIAA0534 protein 25 4AK055482 Homo sapiens cDNA FLJ30920 fis,
clone FEBRA200642726 4
AK054610 Homo sapiens cDNA FLJ30048 fis,clone ADRGL1000018
27 4
NM_002401 MAP3K3 Mitogen-activated protein kinase kinase kinase 3 28 5NM_032342 MGC12992 Hypothetical protein MGC12992 29 5AF017336 Homo sapiens genomic clone X17/P1-68
encoding RNA which may be differentiallyexpressed in individual
30 5
NM_002835 PTPN12 Protein tyrosine phosphatase, nonreceptor type 12 31 5AK054702 B29 B29 protein 32 5NM_000716 C4BPB Complement component 4-binding protein, b 33 5AK023586 Homo sapiens cDNA FLJ13524 fis, clone PLACE1006002 34 5AB028641 SOX11 SRY (sex determining region Y)-box 11 35 5AK001045 Homo sapiens cDNA FLJ10183 fis, clone
HEMBA1004276, highly similar toHomo sapiens AP-4 adaptor comp
36 6
NM_032855 HSH2 Hematopoietic SH2 protein 37 7NM_017433 MYO3A Myosin IIIA 38 7NM_018156 FLJ10619 Hypothetical protein FLJ10619 39 7NM_017886 FLJ20574 Hypothetical protein FLJ20574 40 7NM_017830 OCIA Ovarian carcinoma immunoreactive antigen 41 7NM_003243 TGFBR3 Transforming growth factor, beta receptor III
(b-glycan, 300kD)42 7
NM_015972 LOC51082 RNA polymerase I 16-kDa subunit 43 7AK022418 Homo sapiens cDNA FLJ12356 fis,
clone MAMMA100233944 7
NM_003024 ITSN1 Intersectin 1 (SH3 domain protein) 45 7AL360145 Homo sapiens mRNA full length insert
cDNA clone EUROIMAGE 83955146 7
NM_001122 ADFP Adipose differentiation-related protein 47 7NM_017558 DKFZp434L0850 Hypothetical protein DKFZp434L0850 48 7NM_014100 PRO1770 PRO1770 protein 49 7AF209747 KCNMB2 Potassium large conductance
calcium-activated channel, subfamily M, b member 250 7
NM_018676 LOC55901 TMTSP for transmembrane moleculewith thrombospondin module
51 8
NM_020356 C20orf32 Chromosome 20 open reading frame 32 52 8AB007861 KIAA0401 KIAA0401 protein 53 9NM_018099 FLJ10462 Hypothetical protein FLJ10462 54 9AK054684 Homo sapiens cDNA FLJ30122 fis,
clone BRACE100008755 9
BC011816 Homo sapiens, clone IMAGE:3346747,mRNA, partial cds
56 9
396 LI ET AL.

Confirmation of Candidate Genes bySemi-quantitative RT-PCR and
Real-Time qRT-PCR
Ten of the 69 hypervariable genes were selected forconfirmation based on their functional importance andrelevance with the research project according to existingknowledge. The expression patterns of four genes wereconfirmed by semi-quantitative and quantitative RT-PCR:thrombospondin 1 (TSP1), mitogen-activated protein kinasekinase kinase-3 (MAP3K3), large conductance calcium-acti-vated potassium channel b2 subunit (KCNMB2), and SRY(sex-determining region Y)-box 11 (SOX11). The expressionof three genes, KCNMB2, MAP3K3 and SOX11, all havevery similar patterns, starting with a slight upregulation at0.5–1 h, followed by a 2–4-fold increase in expression overthe 0-h control at 3 or 6 h after treatment (Figs. 4A, 5A,and 6A). The real-time qRT-PCR showed that the expres-sion of TSP1 significantly decreased significantly comparedto the control at 1 h, then returned to the control level at3 h (Fig. 7A). The dynamical trend for each gene observedby real-time qRT-PCR was also confirmed by semi-quantita-tive RT-PCR (Figs. 4B, 5B, 6B, and 7B). The quantitativecomparison of fold change between microarray and real-time quantitative PCR showed that these 4 genes displayedconsistent trends by both microarray and real-time quanti-tative PCR (Table 3).
bE2 Regulates Expression of PigmentEpithelium-Derived Factor (PEDF) in Cultured
Human M€uller Cells
Although the PEDF gene was not present on the arraychip used in this study, it was previously shown to pro-tect cultured retinal neurons against H2O2-induced apo-ptosis (Cao et al., 1999) and enhance M€uller cell devel-opment (Jablonski et al., 2001). Expression of this genein M€uller cells was, therefore, examined in addition tothe selected genes from our microarray analysis. Usingboth quantitative and semi-quantitative RT-PCR, we ob-
served a significant, �3.5-fold increase (n 5 5, P < 0.05)in expression of PEDF compared to the 0-h control at3 h after bE2 treatment (FIG. 8A). A similar trend wasobserved by semi-quantitative RT-PCR (Fig. 8B). The
TABLE 2. (continued).
Accession No. Gene symbol Description Cluster order Cluster
NM_014251 SLC25A13 Solute carrier family 25, member 13 (citrin) 57 9NM_005274 GNG5 Guanine nucleotide binding protein (G protein), g 5 58 9NM_032317 MGC12943 Hypothetical protein MGC12943 59 9NM_000965 RARB Retinoic acid receptor, b 60 9AB028960 KIAA1037 KIAA1037 protein 61 9AK022164 Homo sapiens cDNA FLJ12102 fis,
clone HEMBB100268462 10
AL110125 Homo sapiens, clone IMAGE:4663822,mRNA, partial cds
63 10
NM_057159 EDG2 Endothelial differentiation, lysophosphatidicacid G-protein-coupled receptor, 2
64 10
AK055219 Homo sapiens cDNA FLJ30657 fis, clone DFNES2000335 65 10NM_032814 FLJ14627 Hypothetical protein FLJ14627 66 10NM_000217 KCNA1 Potassium voltage-gated channel, shaker-related
subfamily, member 1 (episodic ataxia with myokymia)67 10
AK056055 Homo sapiens cDNA FLJ31493 fis, clone NT2NE2004490 68 10AK001033 Homo sapiens cDNA FLJ10171 fis, clone HEMBA1003807 69 10
*Genes are in the same order as presented in Fig. 3.
Fig. 3. Heat map representation of the changes of five time pointsagainst 0-h control for all 69 selected hypervariable genes. These geneswere clustered with Ward’s hierarchical clustering method. The colorschema is shown beneath the heat map. Gene descriptions are given atthe right side.
397EFFECT OF ESTROGEN ON RETINAL M €ULLER CELLS

expression pattern of PEDF is very similar to the pat-terns for KCNMB2, MAP3K3, and SOX11, as seen inFigures 4–6.
DISCUSSION
In the present study, we demonstrated for the firsttime that bE2 significantly inhibits H2O2-induced hu-man M€uller cell apoptosis. This result is significant be-cause M€uller cells play crucial roles in (1) supportingneuronal survival (Reichenbach et al., 1993; Newmanand Reichenbach, 1996) by providing trophic substancesto neurons; (2) removing metabolic waste (Poitry, et al.,2000); and (3) stabilizing the structures of the retinaand maintaining of the layered arrangement of retinalneurons (Willbold et al., 1997). Gene expression profileanalysis in this study showed that the protective effectof bE2 on human M€uller cells may operate through reg-ulation of gene expression.
Microarray technology was used to study the mechan-ism underlying the bE2-mediated protective effect onM€uller cells on a global scale. Our data indicated thatgenes functioning in different cell processes, including
formation of extracellular matrix, signal transduction,cell growth regulation, transcription regulation, apopto-sis, and intracellular transport are involved in the estro-gen-mediated transcriptional response. The altered ex-pressions of four genes in these pathways, including TSP1,MAP3K3, KCNMB2, and SOX11 confirmed by RT-PCR andreal-time qRT-PCR demonstrated that they are truly regu-lated by bE2 in M€uller cells.
The TSPs, a family of proteins that regulate tissuegenesis and remodeling, have potent anti-angiogenicproperties (Lawler and Detmar, 2004). It is known thatkeratocyte synthesis of TSP1 is upregulated after var-
Fig. 4. Bar graph presentation of fold change of MAP3K3 transcriptquantified by real-time qRT-PCR (A) and images of semi-quantitativeRT-PCR with GADPH as loading control (B). Data presented are mean6 SD from five biologic replicates. Significant difference is indicated byasterisk, and P-value is also indicated.
Fig. 5. Bar graph presentation of fold change of SOX11 transcriptquantified by real-time qRT-PCR (A) and images of semi-quantitativeRT-PCR with GADPH as loading control (B). Data presented are mean6 SD from five biologic replicates. Significant difference is indicated byasterisk, and P-value is also indicated.
Fig. 6. Bar graph presentation of fold change of KCNMB2 transcriptquantified by real-time qRT-PCR (A) and images of semi-quantitativeRT-PCR with GADPH as loading control (B). Data presented are mean6 SD from five biologic replicates. Significant difference is indicated byasterisk, and P-value is also indicated.
Fig. 7. Bar graph presentation of fold change of TSP1 transcriptquantified by real-time qRT-PCR (A) and images of semi-quantitativeRT-PCR with GADPH as loading control (B). Data presented are mean6 SD from five biologic replicates. Significant difference is indicated byasterisk, and P-values is also indicated.
TABLE 3. Fold Change of Expression Across Different Time Points:Comparison of Microarray and qRT-PCR (Microarray/qRT-PCR)
AccessionNo.
Genesymbol
Fold change against 0-h control
0 0.5 h 1 h 2 h 3 h 6 h
AF209747 KCNMB2 1 1.18/1.35 1.26/1.21 1.13 3.47 3.74/3.65NM_018676 TSP1 1 0.45 0.48/0.34 0.91 1.4/0.95 1.59NM_002401 MAP3K3 1 1.08 0.92/1.29 0.88 3.99/3.76 1.21AB028641 SOX11 1 1.44/1.011 1.86 1.33 3.91/3.26 2.89/3.41
398 LI ET AL.

ious stromal insults in vivo (Hiscott et al., 1999) andthat TSP1 can induce apoptosis of endothelial and can-cer cells via its receptor CD36 (Li et al., 2003). Exposureof human M€uller cells to hypoxia resulted in an elevatedsecretion of TSP1 (Eichler et al., 2004a), which may becaused by increased H2O2 production during hypoxia.Downregulation of TSP1 expression by bE2 suggeststhat bE2 may decrease the apoptotic rate of M€uller cellsby turning down genes involved in this process.
MAP3Ks were originally identified as components ofsignaling cascades in which a MAP3K phosphorylatesand activates a mitogen-activated protein kinase kinase,which in turn activates a mitogen-activated protein kinasecascade (Zhao and Lee, 1999). It has been reported thatelevated expression of MAP3K3 in cells leads toincreased NFjB activity and increased expression of cellsurvival factors, ultimately contributeing to their resis-tance to apoptosis (Samanta et al., 2004). It is clear inthis study that bE2 significantly upregulated MAP3K3gene expression in cultured human M€uller cells, whichmay increase the resistance to apoptosis caused by oxi-dative stress.
Large conductance calcium-activated potassium chan-nel b2 subunit (KCNMB2) has been found to conferrapid and complete inactivation to large conductanceCa21- and voltage-dependent potassium (BK) channelcomplex (Bentrop et al., 2001). The BK channels havebeen implicated in the regulation of the proliferationrate of cultured M€uller cells (Puro et al., 1989). Theactivity of BK channels is necessary for elevated DNAsynthesis in M€uller cells (Kodal et al., 2000) and M€ullercell regulation of the extra-cellular homeostasis. Ourdata demonstrated that bE2 increases KCNMB2 geneexpression in cultured human M€uller cells. Since it inac-tivates the BK channel, the role of KCNMB2 in bE2-mediated protection against apoptosis in M€uller cellsremains to be elucidated.
SOX11 is a transcription factor expressed briefly inmany tissues that undergo inductive remodeling during
embryo development. Deletion of this gene is fatal becausethe absence of this gene product failed proper develop-ment of vital organs, such as the heart (Sock et al., 2004).This gene is also involved in neuronal differentiation, andits expression is upregulated during nerve regeneration(Tanabe et al., 2003). Highly specific over-expression ofthe SOX11 was found in human malignant gliomas (Wei-gle et al., 2005). Upregulation of SOX11 in M€uller cells inresponse to bE2 treatment indicates that this gene couldplay a role in the broad function of estrogen on the cellphysiology. The specific role of SOX11 in bE2-mediatedprotection in human M€uller cells should be further stu-died.
It is worth noting that the peak rate of apoptosisinduced in cultured human M€uller cells appears at 12 hafter H2O2 treatment. This time-dependent activation ofapoptosis in M€uller cells is similar to that observed incultured primary retinal neurons (Cao et al., 1999). Asexpected, the effect of H2O2 on M€uller cell apoptosis isdosage-dependent. Exposure of M€uller cells to 200–400lM H2O2 effectively induced cell apoptosis. This dose ishigher than that sufficient to effectively induce apopto-sis in cultured retinal neurons (60–100 lM) (Cao et al.,1999) and in cultured human retinal pigment epithelium(100–200 lM) (Yu et al., 2005). This suggests thatM€uller cells may be more resistant to oxidative stressthan neurons and retinal pigment epithelium.
PEDF is a potent and broadly acting neurotrophic fac-tor whose functions have been characterized in modelsof several diseases, such as inherited photoreceptor de-generation (Cayouette et al., 1999), H2O2-induced neuro-nal cell death (Cao et al., 1999), ischemic retinal injury(Ogata et al., 2001), and retinal light damage (Caoet al., 2001). PEDF also inhibits oxidative stress-inducedapoptosis and protects cultured retinal pericytes (Amanoet al., 2005). M€uller cells can express and release PEDF(Eichler et al., 2004b). PEDF plays a critical role duringthe final stages of M€uller cell morphogenesis (JablonskiMM et al., 2001). Our study on the gene expression ofPEDF in the bE2-treated M€uller showed that bE2 treat-ment leads to increased PEDF expression perhaps en-hancing M€uller cell survival and the maintenance offunction of cells. PEDF is a secretable neurotrophic sur-vival factor that can be released from M€uller cells. Tounderstand how the increased PEDF expression in M€ullercells by bE2 protects retinal neurons requires furtherinvestigation.
In conclusion, bE2 protects Human M€uller cells fromH2O2-induced cell death. This protective effect may func-tion through regulation of survival related genes, asdemonstrated by upregulating genes that are positivelyinvolved in cell regeneration and amplification, such asPEDF, SOX11, and MAP3K3, and by downregulatingapoptosis promoting genes, such as TSP1. To our knowl-edge, this is the first study to focus on the effects of estro-gen on human M€uller cell survival and gene expression.Although our study is a preliminary step in understand-ing the potential influences of estrogens on retinal M€ullercells, our data provide an important reference in clarify-ing the protective mechanism of estrogen in the retina.
Fig. 8. Bar graph presentation of fold change of PEDF transcriptquantified by real-time qRT-PCR (A) and images of semi-quantitativeRT-PCR with GADPH as loading control (B). Data presented aremean 6 SD from five biologic replicates. Significant difference is indi-cated by asterisk, and P-value is also indicated.
399EFFECT OF ESTROGEN ON RETINAL M €ULLER CELLS

ACKNOWLEDGMENTS
The authors thank Dr. G.A. Limb for generously pro-viding human M€uller cell line MIO-M1, and Sheng Lifor technique support. The authors also thank Drs.Michael H. Elliott and John Loftin Simpson for proof-reading this manuscript. The authors are grateful foran unrestricted grant from RPB to the Department ofOphthalmology.
REFERENCES
Amano S, Yamagishi S, Inagaki Y, Nakamura K, Takeuchi M, Inoue H,Imaizumi T. 2005. Pigment epithelium-derived factor inhibits oxida-tive stress-induced apoptosis and dysfunction of cultured retinal peri-cytes. Microvasc Res 69:45–55.
Amantea D, Russo R, Bagetta G, Corasaniti MT. 2005. From clinicalevidence to molecular mechanisms underlying neuroprotection af-forded by estrogens. Pharmacol Res 52:119–132.
Bentrop D, Beyermann M, Wissmann R, Fakler B. 2001. NMR struc-ture of the ‘‘ball-and-chain’’ domain of KCNMB2, the b2-subunit oflarge conductance Ca21- and voltage-activated potassium channels.J Biol Chem 276:42116–42121.
Cao W, Tombran-Tink J, Chen W, Mrazek D, Elias R, McGinnis JF.1999. Pigment epithelium-derived factor protects cultured retinalneurons against H2O2-induced cell death. J Neurosci Res 57:789–800.
Cao W, Tombran-Tink J, Elias R, Sezate S, Mrazek D, McGinnis JF.2001. In vivo protection of photoreceptors from light damage by pig-ment epithelium-derived factor Invest Ophthalmol Vis Sci 42:1646–1652.
Cao W, Rajala RV, Li F, Anderson RE, Wei N, Soliman CE, McGinnisJF. 2003. Neuroprotective effect of estrogen upon retinal neuronsin vitro. Adv Exp Med Biol 533:395–402.
Cayouette M, Smith SB, Becerra SP, Gravel C. 1999. Pigment epithe-lium-derived factor delays the death of photoreceptors in mousemodel of inherited retinal degenerations. Neurobiol Dis 6:523–532.
Clement MV, Ponton A, Pervaiz S. 1998. Apoptosis induced by H2O2 ismediated by decreased superoxide anion concentration and reductionof intracellular milieu. FEBS Lett 440:13–18.
Dodel RC, Du Y, Bales KR, Gao F, Paul SM. 1999. Sodium salicylateand 17beta-estradiol attenuate nuclear transcription factor NF-kap-paB translocation in cultured rat astroglial cultures after exposure toamyloid A beta (1–40) and lipopolysaccharides. J Neurochem 73:1453–1460.
Dozmorov I, Knowlton N, Tang Y, Shields A, Pathipvanich P, JarvisJN, Centola M. 2004. Hypervariable genes-experimental error or hid-den dynamics. Nucleic Acids Res 32:e147.
Dubois-Dauphin M, Poitry-Yamate C, de Bilbao F, Julliard AK, JourdanF, Donati G. 2000. Early postnatal M€uller cell death leads to retinalbut not optic nerve degeneration in NSE-Hu-Bcl-2 transgenic mice.Neuroscience 95:9–21.
Eichler W, Yafai Y, Wiedemann P, Reichenbach A. 2004a. Angiogenesis-related factors derived from retinal glial (M€uller) cells in hypoxia.NeuroReport 15:1633–1637.
Eichler W, Yafai Y, Keller T, Wiedemann P, Reichenbach A. 2004b.PEDF derived from glial Muller cells: a possible regulator of retinalangiogenesis. Exp Cell Res 299:68–78.
Garcia-Segura LM, Azcoitia L, DonCarlos LL. 2001. Neuroprotection byestradiol. Prog Neurobiol 63:29–60.
Hiscott P, Armstrong D, Batterbury M, Kaye S. 1999. Repair in avascu-lar tissues: fibrosis in the transparent structures of the eye andthrombospondin 1. Histol Histopathol 14:1309–1320.
Jablonski MM, Tombran-Tink J, Mrazek DA, Iannaccone A. 2001. Pig-ment epithelium-derived factor supports normal Muller cell develop-ment and glutamine synthetase expression after removal of the ret-inal pigment epithelium. Glia 35:14–25.
Jarvis JN, Dozmorov I, Jiang K, Chen Y, Frank MB, Cadwell C,Turner S, Centola M. 2004. Gene expression arrays reveal rapidreturn to normal homeostasis in immunologically-challenged tropho-blast-like JAR cells. J Reprod Immunol 61:99–113.
Kitamura Y, Ota T, Matsuoka Y, Tooyama I, Kimura H, Shimohama S,Nomura Y, Gebicke-Haerter PJ, Taniguchi T. 1999. Hydrogen peroxide-induced apoptosis mediated by p53 protein in glial cells. Glia 25:154–64.
Kodal H, Weick M, Moll V, Biedermann B, Reichenbach A, BringmannA. 2000. Involvement of calcium-activated potassium channels in theregulation of DNA synthesis in cultured M€uller glial cells. InvestOphthalmol Vis Sci 41:4262–4267.
Lawler J, Detmar M. 2004. Tumor progression: the effects of thrombos-pondin-1 and -2. Int J Biochem Cell Biol 36:1038–1045.
Li K, Yang M, Yuen PM, Chik KW, Li CK, Shing MM, Lam HK, FokTF. 2003. Thrombospondin-1 induces apoptosis in primary leukemiaand cell lines mediated by CD36 and Caspase-3. Int J Mol Med 12:995–1001.
Li R, Shen Y, Yang LB, Lue LF, Finch C, Rogers J. 2000. Estrogenenhances uptake of amyloid beta-protein by microglia derived fromthe human cortex. J Neurochem 75:1447–1454.
Liang Z, Valla J, Sefidvash-Hockley S, Rogers J, Li R. 2002. Effects ofestrogen treatment on glutamate uptake in cultured human astro-cytes derived from cortex of Alzheimer’s disease patients. J Neuro-chem 80:807–814.
Limb GA, Salt TE, Munroe PMG, Moss SE, Khaw PT. 2002. In vitrocharacterization of a spontaneously immortalized human M€uller cellline. Invest Ophthalmol Vis Sci 43:864–869.
Newman E, Reichenbach A. 1996. The M€uller cells: a functional ele-ment of the retina. Trends Neurosci 19:307–312.
Newman EA. 1993. Inward-rectifying potassium channels in retinalglial (M€uller) cells. J Neurosci 13:3333–3345.
Newman EA, Frambach DA, Odette LL. 1984. Control of extracellular potas-sium levels by retinal glial cell K1 siphoning. Science 225:1174–1175.
Ogata N, Wang L, Jo N, Tombran-Tink J, Takahashi K, Mrazek D, Mat-sumura M. 2001. Pigment epithelium derived factor as a neuroprotec-tive agent against ischemic retinal injury. Curr Eye Res 22:245–225.
Poitry-Yamate CL, Poitry S, Tsacopoulos M. 1995. Lactate released byM€uller glial cells is metabolized by photoreceptors from mammalianretina. J Neurosci 15:5179–5191.
Poitry S, Poitry-Yamate C, Ueberfeld J, MacLeish PR, Tsacopoulos M.2000. Mechanism of glutamate metabolic signaling in retinal glial(M€uller) cells. J Neurosci 20:1809–1821.
Puro DG, Roberge F, Chan CC. 1989. Retinal glial cell proliferation andion channels: a possible link. Invest Ophthalmol Vis Sci 30:521–529.
Reichenbach A, Stolzenburg JU, Eberhardt W, Chao TI, Dettmer D,Hertz L. 1993. What do retinal M€uller (glial) cells do for their ‘‘smallsiblings’’? J Chem Neuroanat 6:201–213.
Samanta AK, Huang HJ, Bast RC Jr, Liao WS. 2004. Overexpression ofMEKK3 confers resistance to apoptosis through activation of NFjB.J Biol Chem 279:7576–7583.
Schumacher M, Akwa Y, Guennoun R, Robert F, Labombarda F, Desar-naud F, Robel P, Nicola AF, Baulieu EE. 2002. Steroid synthesis andmetabolism in the nervous system: trophic and protective effects.J Neurocytol 29:307–326.
Sierra A. 2004. Neurosteroids: the StAR protein in the brain. J Neu-roendocrinol 16:787–793.
Sock E, Rettig SD, Enderich J, Bosl MR, Tamm ER, Wegner M. 2004. Genetargeting reveals a widespread role for the high-mobility-group transcrip-tion factor Sox11 in tissue remodeling. Mol Cell Biol 15:6635–6644.
Stone DJ, Rozovsky I, Morgan TE, Anderson CP, Hajian H, Finch CE.1997. Astrocytes and microglia respond to estrogen with increasedapoE mRNA in vivo and in vitro. Exp Neurol 143:313–318.
Tanabe K, Bonilla I, Winkles JA, Strittmatter SM. 2003. Fibroblastgrowth factor-inducible-14 is induced in axotomized neurons and pro-motes neurite outgrowth. J Neurosci 23:9675–9686.
Vegeto E, Bonincontro C, Pollio G, Sala A, Viappiani S, Nardi F, Brusa-delli A, Viviani B, Ciana P, Maggi A. 2001. Estrogen prevents thelipopolysaccharide-induced inflammatory response in microglia. J Neu-rosci 21:1809–1818.
Weigle B, Ebner R, Temme A, Schwind S, Schmitz M, Kiessling A, Rie-ger MA, Schackert G, Schackert HK, Rieber EP. 2005. Highly specificoverexpression of the transcription factor SOX11 in human malignantgliomas. Oncol Rep 13:139–144.
Willbold E, Berger J, Reinicke M, Wolburg H. 1997. On the role ofM€uller glia cells in histogenesis: only retinal spheroids, but not tec-tal, telencephalic and cerebellar speroids develop histotypical pat-terns. J Hirnforsch 38:383–396.
Wise P M. 2002. Estrogens and neuroprotection. Trends EndocrinolMetab 13:229–230.
Yu X, Rajala RV, McGinnis JF, Li F, Anderson RE, Yan X, Li S, EliasRV, Knapp RR, Zhou X, Cao W. 2004. Involvement of insulin/phos-phoinositide 3-kinase/Akt signal pathway in 17b-estradiol-mediatedneuroprotection. J Biol Chem 279:13086–13094.
Yu X, Tang Y, Li F, FrankMB, HuangH, Dozmorov I, Zhu Y, Centola M, CaoW. 2005. Protection against hydrogen peroxide-induced cell death in cul-tured human retinal pigment epithelial cells by 17b-estradiol: a differen-tial gene expression profile. Mech AgingDev 126:1135–1145.
Zhao Q, Lee FS. 1999. Mitogen-activated protein kinase/ERK kinasekinases 2 and 3 activate nuclear factor-jB through IjB kinase-a andIjB kinase-b. J Biol Chem 274:8355–8358.
Zhou X, Li F, Kong L, Tomita H, Li C, and Cao W. 2005. Involvement ofinflammation, degradation and apoptosis in a mouse model of glau-coma. J Bio Chem 280:31240–31248.
400 LI ET AL.