1 the capacity of ul49.5 proteins to inhibit tap is widely distributed
TRANSCRIPT

1
The capacity of UL49.5 proteins to inhibit TAP is widely distributed amongst members of 1
the genus Varicellovirus 2
3
Running title: Inhibition of TAP by varicellovirus UL49.5 proteins 4
5
Marieke C. Verweij1, Andrea D. Lipińska2, Danijela Koppers-Lalic1, Wouter F. van Leeuwen3, 6
Jeffrey I. Cohen4, Paul R. Kinchington5, Ilhem Messaoudi6, Krystyna Bieńkowska-Szewczyk2, 7
Maaike E. Ressing3, Frans A.M. Rijsewijk7, Emmanuel J.H.J. Wiertz3* 8
9
1Department of Medical Microbiology, Center of Infectious Diseases, Leiden University Medical 10
Center, Leiden, The Netherlands 11
2Department of Molecular Virology, Faculty of Biotechnology, University of Gdańsk, Gdańsk, 12
Poland 13
3Department of Medical Microbiology, University Medical Center Utrecht, Utrecht, The 14
Netherlands 15
4Laboratory of Clinical Infectious Diseases, National Institute of Allergy and Infectious Diseases, 16
National Institutes of Health, Bethesda, USA 17
5Department of Ophthalmology, University of Pittsburgh, Pittsburgh, USA 18
6Vaccine and Gene Therapy Institute, Division of Pathobiology and Immunology, Oregon Health 19
and Science University, Beaverton, USA 20
7Department of Microbiology, Institute of Basic Health Sciences, Federal University of Rio 21
Grande do Sul (UFRGS), Porto Alegre, Rio Grande do Sul (RS), Brazil 22
23
Copyright © 2010, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.J. Virol. doi:10.1128/JVI.01621-10 JVI Accepts, published online ahead of print on 15 December 2010
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

2
Footnote 1, current address 1
MCV, Vaccine and Gene Therapy Institute, Oregon Health and Science University, Beaverton, 2
USA; DKL, Department of Molecular Cell Biology, Leiden University Medical Center, The 3
Netherlands 4
5
Footnote 2, corresponding author 6
Dr. Emmanuel J.H.J. Wiertz, Department of Medical Microbiology, University Medical Center 7
Utrecht, Heidelberglaan 100, 3584 CX Utrecht, The Netherlands. Tel.: +31 88 7550862. Fax: +31 8
30 2541770. E-mail address: [email protected] 9
10
Word count 11
Abstract: 243 12
Text: 5342 13
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

3
Abstract 1
2
The life long infection by varicelloviruses is characterized by a fine balance between the host 3
immune response and immune evasion strategies employed by these viruses. Virus-derived 4
peptides are presented to cytotoxic T lymphocytes by MHC class I molecules. The Transporter 5
associated with Antigen Processing (TAP) transports the peptides from the cytosol into the ER, 6
where loading of MHC I molecules occurs. The varicelloviruses bovine herpesvirus 1 (BoHV-1), 7
pseudorabies virus, and equid herpesvirus 1 and 4 have been found to encode a UL49.5 protein 8
that inhibits TAP-mediated peptide transport. To investigate to what extent UL49.5-mediated 9
TAP inhibition is conserved within the family of Alphaherpesvirinae, the homologs of another 10
five varicelloviruses, one mardivirus and one iltovirus were studied. The UL49.5 proteins of 11
BoHV-5, bubaline herpesvirus 1, cervid herpesvirus 1 and felid herpesvirus 1 were identified as 12
potent TAP inhibitors. The lack of TAP inhibition observed for varicella-zoster virus and simian 13
varicellavirus UL49.5 is not because of the absence of viral co-factors that might assist in this 14
process, as cells infected with these viruses did not show reduced TAP function. The UL49.5 15
homologs of the mardivirus Marek’s disease virus 1 and the iltovirus infectious laryngotracheitis 16
virus did not block TAP, suggesting that the capacity to inhibit TAP via UL49.5 has been 17
acquired by varicelloviruses only. A phylogenetic analysis of viruses that inhibit TAP through 18
their UL49.5 proteins reveales an interesting hereditary pattern, pointing towards the presence of 19
this capacity in defined clades within the genus Varicellovirus. 20
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

4
Introduction 1
2
Herpesviruses cause a life long infection in their host. The anti-viral immune response of the host 3
is counteracted by immune evasion strategies employed by these viruses. CD8+ cytotoxic T 4
lymphocytes (CTL) play an important role in immunity against viruses, recognizing viral peptides 5
presented on major histocompatibility complex class I (MHC I) molecules at the cell surface. The 6
antigenic peptides originate from proteasomal degradation of virus-encoded proteins in the 7
cytosol. The peptides are transported into the endoplasmic reticulum (ER) by the Transporter 8
associated with Antigen Processing (TAP) in an ATP-dependent manner1,6,32. TAP proteins are 9
highly conserved amongst various species: for example, human, porcine, bovine and rodent TAP 10
1 and TAP2 demonstrate 70-80% amino acid identity15,37. The transporter is thought to be 11
involved in MHC I-mediated antigen presentation in many different species, including 12
Xenopus42. 13
14
The inhibition of peptide transport by TAP is often exploited by herpesviruses to prevent 15
elimination by CTLs. The first TAP inhibitor to be identified was the ICP47 protein of herpes 16
simplex virus 1 (HSV-1)4. ICP47 of HSV-2 was later on found to have the same function48. The 17
ICP47 proteins of these viruses prevent peptide transport by obstructing the peptide binding site 18
of the TAP complex4,5,12,23,47. The US6 protein of the human cytomegalovirus (HCMV) interferes 19
with ATP binding to TAP, thereby limiting its energy supply and, consequently, the transport of 20
peptides3,19,21,22,31. The Epstein-Barr virus (EBV) encodes the TAP inhibitor BNLF2a that blocks 21
both the binding of peptides and ATP to TAP24,25. A fourth class of TAP-inhibiting proteins, 22
encoded by the UL49.5 gene, has been identified in the varicelloviruses bovine herpesvirus 1 23
(BoHV-1), equid herpesvirus (EHV) 1 and 4, and suid herpesvirus 1 or pseudorabies virus (PRV) 24
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

5
all inhibit TAP function28,30. The mechanisms by which UL49.5 homologs inhibit TAP 1
demonstrate remarkable heterogeneity. All proteins block conformational changes within the 2
complex that are required for peptide transport. In addition, BoHV-1 UL49.5 also induces 3
degradation of TAP1 and TAP228,30. In contrast, EHV-1 and EHV-4 UL49.5 prevent ATP 4
binding to TAP30. 5
6
Homologs of UL49.5 proteins are encoded by all herpesviruses sequenced10,38. The UL49.5 genes 7
encode a type 1 transmembrane protein that is often N-glycosylated and therefore known as 8
glycoprotein N or gN. In several herpesviruses, UL49.5 has been demonstrated to be involved in 9
virion maturation and infectivity: UL49.5 forms a heterodimeric complex with glycoprotein M 10
(gM) and is necessary for proper glycosylation and maturation of the complex14,27,35,43,50. Thus, 11
for some viruses, UL49.5 possesses a dual role, functioning both as a molecular chaperone and as 12
an immune evasion protein. 13
The family of Herpesviridae has been classified into three subfamilies, the Alpha-, Beta-, and 14
Gammaherpesvirinae. Isolated expression of the UL49.5 homologs encoded by members of these 15
subfamilies, including HSV-1 and -2 (alphaherpesvirus, genus Simplexviruses), HCMV 16
(betaherpesvirus) and EBV (gammaherpesvirus), did not result in reduced TAP function28. The 17
TAP-inhibiting UL49.5 proteins of BoHV-1, EHV-1, EHV-4, and PRV all belong to the genus 18
Varicellovirus of the Alphaherpesvirinae. However, human herpesvirus 3 or varicella-zoster virus 19
(VZV) and canid herpesvirus 1 (CaHV-1), which are members of the same genus, UL49.5 exhibit 20
no or poor TAP inhibition, respectively, indicating that the TAP-inhibiting capacity of UL49.5 21
proteins is only found for a selection of varicelloviruses. 22
23
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

6
The Alphaherpesvirinae include two other genera, the Mardivirus and the Iltovirus. Gallid 1
herpesvirus 2 or Marek’s disease virus 1 (MDV-1) is a member of the mardiviruses. Serotype 1 of 2
MDV is oncogenic, inducing T cell tumors in infected poultry. MHC I downregulation has been 3
observed on MDV-1 infected chicken cells26,33 and in epithelial and infiltrating cells derived from 4
brain tissue of infected chickens18. To date, the responsible viral protein(s) have not been 5
identified. Gallid herpesvirus 1 or infectious laryngotracheitis virus (ILTV) is a member of the 6
iltoviruses. At present, it is unclear if MHC I-restricted antigen presentation is affected by ILTV. 7
8
In this study, immune evasion by UL49.5 homologs of various clades within the genus 9
Varicellovirus was assessed. Human and virus-specific host cell-lines stably expressing the 10
UL49.5 proteins were screened for MHC I downregulation and TAP inhibition. If TAP inhibition 11
was observed, the stability of TAP and ATP binding to the TAP complex were determined to 12
investigate the mechanism utlized by the UL49.5 homologs to inhibit TAP function. The UL49.5 13
proteins of BoHV-5, water buffalo herpesvirus or bubaline herpesvirus 1 (BuHV-1), red deer 14
herpesvirus or cervid herpesvirus 1 (CvHV-1), and feline rhinotracheitis virus or felid herpesvirus 15
1 (FeHV-1) were identified as potent TAP inhibitors. The UL49.5 homolog of cercopithecine 16
herpesvirus 9 or simian varicella virus (SVV), which is closely related to VZV, only slightly 17
reduced peptide transport by rhesus macaque TAP. UL49.5 proteins of the alphaherpesviruses 18
MDV-1 (mardivirus) and ILTV (ilthovirus) fail to inhibit TAP. These and previous findings are 19
discussed in the context of the phylogeny of these viruses. 20
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

7
Materials and Methods 1
2
UL49.5 constructs Purified viral DNA from the clinical isolates BoHV-5 Evi 88/959, BuHV-1 3
strain B6 (kind gift from M.J. Studdert, Faculty of Veterinary Science, University of Melbourne, 4
Australia)45, CvHV-1 strain D2839 (kindly provided by P. F. Nettleton, Moredun Research 5
Institute, Edinburgh, Great Britain), FeHV-1 strain B927 (kindly provided by R. de Groot, 6
Department of Infectious Diseases and Immunology, Faculty of Veterinary Medicine, Utrecht 7
University, The Netherlands)17, and vaccine strain MDV-1 CVI988 (J. van Oirschot, Department 8
of Virology, Central Veterinary Institute, Lelystad, The Netherlands)18 were used as a template 9
for polymerase chain reaction (PCR) amplification. PCR-reactions were performed using Pfu 10
(Invitrogen), Taq (Promega) or KOD DNA polymerase (Novagen-Merck) and specific primers 11
(Table 1) for amplification of the coding sequence of the UL49.5 genes. The sequences of the 12
primers were based on published sequences found in the NCBI database. PCR-generated products 13
were sequenced and inserted into the retroviral expression vectors pLZRS-IRES-GFP, behind the 14
HCMV IE1 promoter and upstream of an internal ribosome entry site (IRES) element, followed 15
by GFP. SVV UL49.5 (derived from the clinical isolate Delta)39 was amplified from pcDNA3.1 16
by PCR (primers Table 1) and cloned into pLZRS using GATEWAY technology (Invitrogen). 17
ILTV UL49.5 (derived from the clinical isolate A489) was recloned from a pcDNA3.1 vector (a 18
kind gift from W. Fuchs and T.C. Mettenleiter, Institute of Molecular Biology, Friedrich-Loeffler 19
Institut, Greifswald-Insel Riems, Germany)13,14 into pLZRS (primers Table 1). Information on the 20
pLZRS vector can be obtained at www.stanford.edu/group/nolan/retroviral_systems/retsys.html. 21
22
The pDONR209 plasmids containing the UL49.5 constructs of MDV1 strains RB1B (clinical 23
isolate) and CVI988 (vaccine strain) were kind gifts from J. Haas (Devision of Pathway 24
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

8
Medicine, University of Edinburgh, UK). The inserts were cloned into the lentiviral vector 1
pDEST-LV-IRES-GFP (K. Franken, Department of Immunohematology and Blood Transfusion, 2
Leiden University Medical Center, The Netherlands) upstream of an IRES element, which was 3
followed by eGFP using the GATEWAY technology. Sequences were of the resulting expression 4
vectors were verified. 5
6
Cell lines and retroviruses The human melanoma cell line Mel JuSo (MJS), MJS BoHV-1 7
UL49.5-IRES-GFP28, MJS IRES-GFP, the epithelial cell line Madin-Darby bovine kidney 8
(MDBK) cells (American Type Culture Collection, ATCC; commonly used to propagate BoHV-9
5, BuHV-1 and CvHV-1) and MDBK UL49.5-IRES-GFP30 were maintained in RPMI-1640 10
medium; the feline-derived epithelial cell line CRFK (provided by R. de Groot, Department of 11
Infectious Diseases and Immunology, Faculty of Veterinary Medicine, Utrecht University, The 12
Netherlands), human fibroblast cell line MRC-5 (a gift from G.M. Verjans, Rotterdam Eye 13
Hospital, The Netherlands), telomerized rhesus fibroblasts (TRF) (obtained from J.A. Nelson, 14
Vaccine and Gene Therapy Institute, Oregon Health and Science University, USA), and the avian 15
hepatoma cell line LMH were cultured in DMEM medium; the rhesus macaque-derived epithelial 16
cell line LLC-MK2 (kindly provided by A.D. Hislop, School of Cancer Sciences, University of 17
Birmingham, United Kingdom) were cultured in EMEM supplemented with non-essential amino 18
acids. Media were supplemented with 10% heat-inactivated fetal bovine serum (FBS), 2 mM L-19
glutamine (Invitrogen), 140 IU/ml penicillin and 140 mg/ml streptomycin. 20
Retroviruses were made using the Phoenix amphotropic packaging system as described before 21
(www.stanford.edu/group/nolan/retroviral_systems/retsys.html). The retroviruses were used to 22
transduce target cells, after which eGFP positive cells were selected using a FACSAria cell sorter 23
(Becton Dickinson). The following stable cell lines were generated: MJS expressing BoHV-5, 24
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

9
BuHV-1, CvHV-1, FeHV-1, SVV, SVV-HA, and ILTV UL49.5; 5; CRFK expressing FeHV-1; 1
LLC-MK2 expressing SVV UL49.5; and LMH expressing ILTV UL49.5. The generation of 2
recombinant retroviruses for MDBK cell line transductions was described before30. GP2-293 3
pantropic packaging cells were co-transfected with pZLRS-UL49.5-IRES-GFP and pVSV-G 4
construct (envelope vector) to obtain retroviruses used for the establishment of MDBK stably 5
expressing BoHV-5, BuHV-1, and CvHV-1 UL49. 6
7
Antibodies The following antibodies were used for flow cytometry: anti-human MHC I 8
complexes mAb W6/327, anti-human MHC II HLA-DR mAb L243 (ATCC), anti-feline MHC I 9
complexes mAb H58A (VMRD Inc.), anti-FeHV-1 MCA2490 (Serotec), anti-human transferrin 10
receptor mAb CD71 (Becton Dickinson), anti-human epidermal growth factor receptor mAb Ab-11
5 (Calbiochem), and anti-chicken MHC I complexes mAb F21-2 (SouthernBiotech). 12
For detection of UL49.5, rabbit polyclonal anti-sera were raised against a synthetic peptide 13
(RLMGASGPNKKESRGRG) derived from the C-terminal domain of BoHV-1 UL49.535. In 14
addition, we used rabbit anti-ILTV UL49.5 (kindly provided by W. Fuchs and T.C. 15
Mettenleiter)14, anti-TAP1 mAb 148.340, anti-TAP2 mAb 435.3 (kind gift from P. van Endert, 16
INSERM, U580, Université Paris Descartes, Paris, France), and rabbit anti-GFP49. Anti-actin 17
mAb AC-74 (Sigma-Aldrich) was used as a control. 18
19
Flow cytometry Surface expression levels of MHC I and MHC II molecules were determined by 20
flow cytometry. Cells were stained with the indicated primary antibodies and, after washing, with 21
the secondary goat anti-mouse allophycocyanin Ab (Leinco Technologies) or goat anti-mouse 22
phycoerythrin Ab (Jackson ImmunoResearch Laboratories) at 4°C. Stained cells were measured 23
using a FACSCalibur and LSR II (Becton Dickinson) and analyzed using CellQuest (Becton 24
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

10
Dickinson) or FlowJo (Tree Star) software. One representative experiment out of two 1
independent experiments is shown. 2
3
Peptide transport assay Cells were permeabilized using 2.5 units/ml Streptolysin-O (Murex 4
Diagnostics) at 37˚C for 10 min. Permeabilized cells were incubated with 4.5 µM of the 5
fluorescein-conjugated synthetic peptide CVNKTERAY (N-core glycosylation site underlined) in 6
the presence of 10 mM ATP or 0.125 M EDTA at 37˚C for 10 min. Addition of EDTA, which 7
binds divalent cations, abrogates the ATP-hydrolysizing function of TAP. Peptide translocation 8
was terminated by adding 1 ml of ice-cold lysis buffer (1% Triton X-100, 500 nM NaCl, 2 mM 9
MgCl2, 50 mM Tris HCl, pH 8.0). After lysis for 30 minutes at 4°C, cells were centrifuged at 10
16,000 g for 20 minutes at 4°C in order to obtain post-nuclear lysates. Glycosylated peptides 11
were isolated from these lysates by incubation with concanavalin A-Sepharose beads (GE 12
Healthcare) for 2 hr at 4°C. After washing of the beads, glycosylated peptides were eluted from 13
the beads with elution buffer (500 mM mannopyranoside, 10 mM EDTA, 50 mM Tris HCl pH 14
8.0) during a one hour incubation step at room temperature. Fluorescence was measured using a 15
Mithras LB 940 multilabel reader (Berthold Technologies) or a VersaFluorTM fluorometer (Bio-16
rad). Peptide transport is expressed as percentage of translocation, relative to the translocation 17
observed in control cells (set at 100%). One representative experiment out of two independent 18
experiments is shown. 19
20
Immunoblotting Cells were lysed in 1% Nonidet P-40 and proteins were separated by SDS-21
PAGE and subsequently transferred to PVDF membranes (GE Healthcare). UL49.5 proteins were 22
separated using 16.5%-tricine PAGE. The blots were incubated with the indicated antibodies, 23
followed by horseradish peroxidase (HRP)-conjugated secondary antibodies (DAKO and Jackson 24
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

11
ImmunoResearch Laboratories). Bound HRP-labeled antibodies were visualized using ECL Plus 1
(GE Healthcare). One representative experiment out of two independent experiments is shown. 2
3
Transient expression MJS and LMH cells were transiently transfected using the Amaxa 4
Nucleofector II device according to the manufacturer’s instructions. For MJS, program T-020 and 5
solution L were used and for LMH, program T-020 and solution T. Two days post transfection 6
cells were analyzed for MHC I expression. 7
8
Viruses and viral infections CRFK cells were infected with FeHV-1 (strain B927) at an m.o.i. of 9
10 in serum-free DMEM. After 1 hour of virus adsorption, DMEM containing 10% FBS was 10
added to make the final FBS concentration 5%. At 5 or 12 h.p.i cells were harvested. To restrict 11
viral gene expression to immediate early (IE) and early (E) genes, cells were treated with 300 12
µg/ml phosphonoacetic acid (PAA: Sigma) for 2 hr before addition of the virus and during 13
infection. The Dutch BoHV-1.1 field strain Lam was used to infect MRC-5 cells at an m.o.i. of 14
20. After 2 hr of virus adsorption, the inoculum was removed and cells were incubated in the 15
presence of complete RPMI-1640 medium for another 3 hr. A bac-derived Rhesus CMV44 was 16
used to infect TRF at an m.o.i. of 1. The cells were harvested 36 h.p.i. 17
For VZV infection we used the recombinant viruses VZV pOka expressing ORF66 N-terminally 18
tagged with eGFP11 and VZV-GFP34 that contains eGFP inserted between ORF65 and ORF66. 19
Trypsinized VZV-infected MRC-5 cells were used to infect confluent monolayers of MRC-5 at a 20
4:1 ratio (uninfected cells to infected cells). This yielded a 100% eGFP-positive population when 21
harvested 72-88 h.p.i. During infection MRC-5 cells were cultured in DMEM supplemented with 22
2% FBS. For SVV infections we used the recombinant SVV delta, in which eGFP was inserted 23
between US2 and US3 through homologous recombination36. A confluent monolayer of TRF 24
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

12
cells was infected in a 6:1 ratio with previously SVV-infected TRF cells, The cells were 1
harvested 72 h.p.i. and were 100% positive for eGFP. 2
In all experiments, mock-infected cells were treated under the same conditions as infected cells. 3
One representative experiment out of two independent experiments is shown. 4
5
ATP-agarose binding assay TAP binding to ATP-agarose was assayed as described28. In brief, 6
cells were solubilized in 1% Nonidet P-40 or in 1% (w/v) digitonin, 50 mM Tris-HCl (pH 7.5), 5 7
mM MgCl2, 150 mM NaCl, 5 mM iodoacetamide, and 1 mM AEBSF to preserve the interactions 8
between TAP1, TAP2, and UL49.5. Hydrated C-8 ATP-agarose (Fluka/Sigma) was added to the 9
post-nuclear supernatant and incubated at 4°C o/n. The supernatant was separated from the ATP-10
agarose pellet by 5 minutes centrifugation. The resulting pellet was washed three times with 0.1% 11
(w/v) digitonin, 50 mM Tris-HCl (pH 7.5), 5 mM MgCl2 and 150 mM NaCl. Proteins bound to 12
the ATP-agarose beads were eluted with 500 mM EDTA and SDS-PAGE sample buffer was 13
added to both the supernatant and the pellet. The samples were separated using SDS-PAGE and 14
analyzed by immunoblotting. One representative experiment out of two independent experiments 15
is shown. 16
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

13
Results 1
2
TAP inhibition by herpesviruses infecting ruminants 3
The sequences of the UL49.5 proteins encoded by BoHV-5, BuHV-1, and CvHV-1 are 78.9%, 4
80.0% and 74.0% identical to the BoHV-1 UL49.5, respectively (Fig. 1 and Table 2). To evaluate 5
if the capacity to inhibit TAP is conserved for these proteins, the BoHV-5, BuHV-1, and CvHV-1 6
UL49.5 proteins were expressed in bovine cells (MDBK) and human (MJS) cells. Expression of 7
the UL49.5 homologs was verified using an antibody specific for BoHV-1 UL49.5. This antibody 8
was capable of detecting the UL49.5 proteins of BoHV-1, BoHV-5 and CvHV-1 in MDBK and 9
MJS cells (Fig. 2A). BuHV-1 UL49.5 could not be detected, possibly due to differences within 10
the C-terminal region against which the antibody was raised. In the retroviral vector used to 11
establish the cell-lines, the UL49.5 homologs were placed upstream of an internal ribosomal entry 12
site (IRES) that is followed by a eGFP gene. Since the two proteins are encoded by one transcript, 13
expression of eGFP reflects expression of the UL49.5 protein. Detection of eGFP in cells 14
transduced with the BuHV-1 UL49.5-IRES-GFP retrovirus confirmed expression of the 15
corresponding transcript (Fig. 2A). 16
Analysis of MHC I cell surface expression revealed that the BoHV-5, BuHV-1, and CvHV-1 17
UL49.5 proteins cause a strong downregulation on MDBK and MJS cells (Fig. 2B, upper and 18
middle panels, respectively). The reduction was comparable to that induced by BoHV-1 UL49.5. 19
This downregulation was specific for MHC I, as MHC II expression was unaffected by all 20
UL49.5 homologs (Fig. 2B, lower panels). Next, TAP function was assessed in BoHV-5, BuHV-21
1, and CvHV-1 UL49.5-expressing MDBK and MJS cells. Transport of the reference peptide was 22
reduced by 70 to 90% in both human and bovine cells, indicating that the observed reduction of 23
MHC I results from TAP inhibition (Fig. 2C). 24
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

14
Previously, BoHV-1 UL49.5 was found to induce degradation of human and bovine TAP28,30. To 1
evaluate whether BoHV-5, BuHV-1, and CvHV-1 UL49.5 posses a similar capacity, TAP1 and 2
TAP2 levels were analyzed in MJS cells expressing these proteins (Fig. 2D). TAP levels were 3
strongly reduced in the presence of all three UL49.5 homologs tested, suggesting that UL49.5-4
induced degradation of the TAP complex is conserved for UL49.5 proteins of herpesviruses 5
infecting ruminants. 6
7
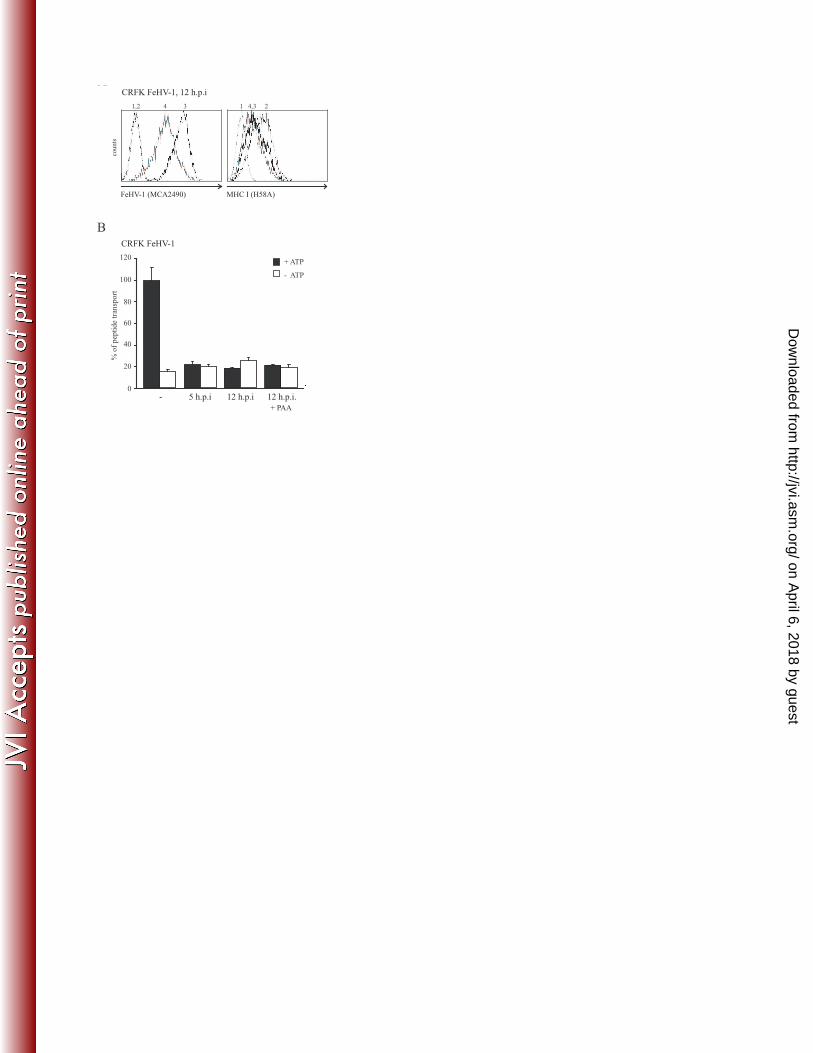
FeHV-1 UL49.5 inhibits TAP-mediated peptide transport 8
FeHV-1 has recently been found to reduce MHC I levels on infected cells41. To assess if this 9
downregulation resulted from the inhibition of peptide transport by TAP, TAP activity was 10
analyzed in feline CRFK cells infected with FeHV-1 for 5 and 12 hr. To evaluate the contribution 11
of FeHV-1 late proteins to TAP inhibition, the experiment was also performed in the presence of 12
PAA, which inhibits late viral protein synthesis. FeHV-1 infection of the cells was confirmed 13
using an antibody recognizing several FeHV-1 glycoproteins (Fig. 3A, left panel). As expected, 14
MHC I levels were downregulated on FeHV-1-infected cells (Fig. 3A, right panel) and, 15
additionally, TAP function was strongly reduced already at 5 h.p.i. (Fig. 3B). TAP was still 16
inhibited after 12 hr, irrespective of the presence of PAA, implying that an immediate early or 17
early gene is responsible for this effect, possibly UL49.5. 18
19
To study if FeHV-1 UL49.5 plays a role in the observed TAP inhibition, this homolog was 20
expressed in CRFK and MJS cells. FeHV-1 UL49.5 induced a strong downregulation of MHC I 21
in CRFK and a slightly weaker downregulation in MJS cells. The inhibition of MHC I expression 22
was specific, as MHC II expression was unaffected (Fig. 4A). TAP function was inhibited in both 23
FeHV-1 UL49.5-expressing cell lines, albeit less effective in the MJS cells (Fig. 4B). In the 24
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

15
absence of FeHV-1 UL49.5-specific antibodies, expression of UL49.5 gene was inferred from the 1
presence of eGFP that is encoded by the same transcript (Fig. 4C). The antibody used to detect 2
human TAP1 cross-reacted with feline TAP1 (Fig. 4C). Therefore, this antibody could be used to 3
investigate if FeHV-1 UL49.5 shares the capacity of UL49.5 of ruminant varicelloviruses to 4
degrade TAP. However, in the presence of FeHV-1 UL49.5 no degradation of TAP was observed 5
(Fig. 4C). 6
Preventing ATP-binding to TAP is a strategy often exploited by TAP inhibitors, including EHV-1 7
UL49.530. To investigate if FeHV-1 UL49.5 uses this strategy, the ATP-binding capacity of TAP 8
was assessed in lysates from untransduced cells and cells stably expressing FeHV-1 or EHV-1 9
UL49.5. In control CRFK cells, but also in FeHV-1 UL49.5-expressing CRFK cells, TAP1 was 10
detected in the pellet fraction, showing that TAP1 was bound to ATP (Fig. 4D; lane 1-4). 11
Correspondingly, in MJS cells expressing FeHV-1 UL49.5, TAP1 was found in the pellet 12
fractions (Fig. 4D; lanes 5-8). In contrast, in the presence of EHV-1 UL49.5, very little TAP1 was 13
found in the pellet fraction, confirming interference with ATP-binding to TAP by EHV-1 UL49.5 14
(Fig. 3D; compare lane 9 with 10). These results demonstrate that ATP-binding to TAP is not 15
affected by FeHV-1 UL49.5. 16
17
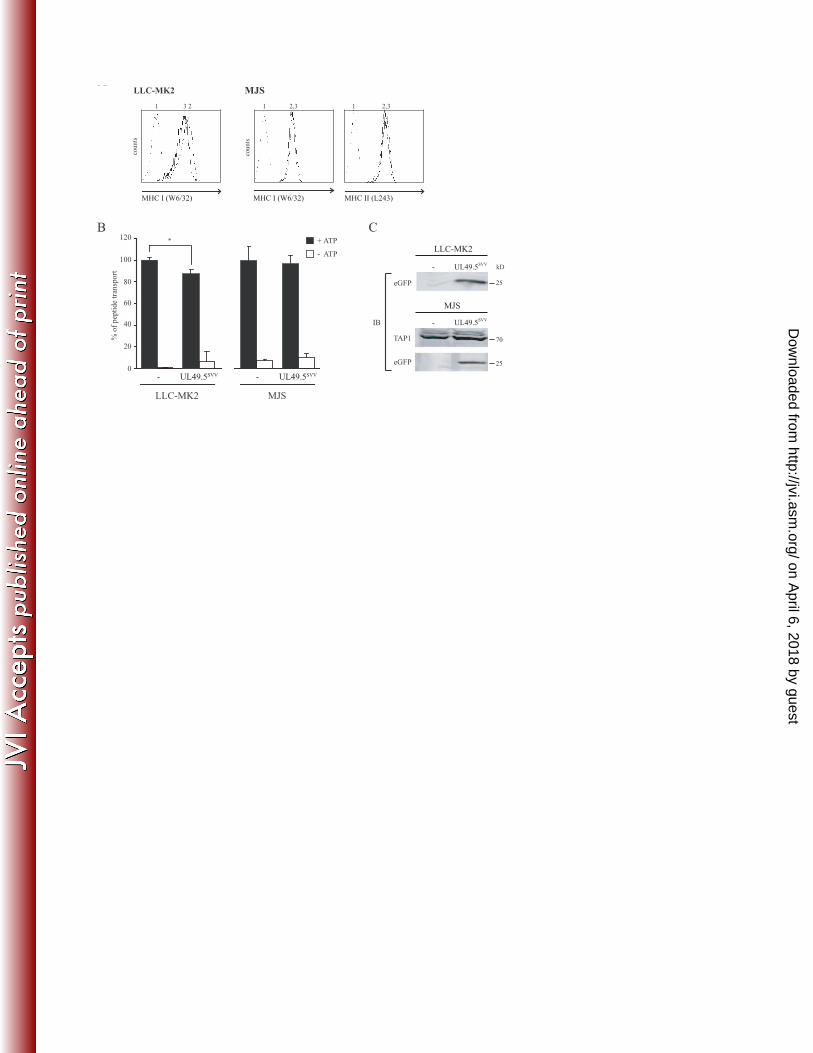
The effect of SVV UL49.5 on TAP-mediated peptide transport 18
The protein sequences of VZV and SVV UL49.5 display 63.2% sequence identity (Fig. 1 and 19
Table 2). Previously, we have shown that VZV UL49.5 does not interfere with TAP function30. 20
To investigate whether the UL49.5 protein of SVV blocks TAP, the viral protein was expressed 21
in a rhesus macaque-derived cell line (LLC-MK2) and in MJS cells. On LLC-MK2 cells 22
expressing SVV UL49.5, MHC I expression was slightly reduced (Fig. 5A). This was 23
accompanied by a 15% reduction of peptide transport in those cells (Fig. 5B). SVV UL49.5-24
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

16
induced downregulation of MHC I expression could not be detected on MJS (Fig. 5A). In 1
accordance, TAP inhibition was undetectable in SVV UL49.5-expressing MJS cells (Fig. 5B). 2
eGFP was expressed in both cell lines, indicating proper expression of the SVV UL49.5-encoding 3
transcript (Fig. 5C). Protein steady state levels of TAP were not affected by SVV UL49.5. All 4
together, these data indicate that SVV UL49.5 has a slight effect on TAP function in rhesus cells. 5
6
VZV infection does not result in TAP inhibition 7
Despite the fact that VZV UL49.5 can be found in association with the TAP complex, TAP 8
inhibition has not been observed30. A slight reduction in TAP function was abserved in the 9
presence of SVV UL49.5 (Fig. 5B). The UL49.5 proteins of VZV and SVV might require one or 10
more other viral gene product(s) to accomplish TAP inhibition. To investigate this possibility, 11
TAP function was assessed in cells infected with VZV or SVV. EGFP-expressing viruses allowed 12
us to determine the efficiency of infection by flow cytometry. For VZV, two different 13
recombinant viruses were used. In one virus, eGFP is inserted between ORF65 and ORF66 14
(VZV.GFP)34. In the second virus, eGFP is fused to the N-terminus of the ORF66 protein 15
(VZV.GFP-66)11. MRC-5 cells were infected with VZV.GFP and VZV.GFP-66 at a ratio of 4:1 16
(uninfected to VZV-infected cells) for 88 hr, after which the efficiency of infection was found to 17
be nearly 100% (Fig. 6A; upper panels). Next, cell surface levels of MHC I and transferrin 18
receptor (TfR) were determined on mock- and VZV-infected MRC-5 cells. A moderate 19
downregulation of MHC I was observed on VZV-infected cells, whereas TfR levels were 20
upregulated (Fig. 6A, middle and lower panels). These results are in line with previous 21
observations8,11. Subsequently, peptide transport was assessed in VZV.GFP-infected cells. As a 22
positive control, MRC-5 cells were infected with BoHV-1. Infection was verified by eGFP 23
expression (VZV) or by cell surface staining using a BoHV-1 gB-specific antibody (data not 24
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

17
shown). TAP function was not affected in VZV.GFP-infected MRC-5 cells (Fig. 6B). In contrast, 1
BoHV-1 infection resulted in a 40% reduction of peptide transport (Fig. 6B), which is in 2
accordance with previous observations29. The experiment was repeated and confirmed in MRC-5 3
cells infected with VZV.GFP-66 (data not shown). 4
The same experiments were performed with an SVV.eGFP recombinant (with eGFP inserted into 5
the US region)36. Infection of telomerized rhesus fibroblasts (TRF) was monitored via eGFP 6
expression using flow cytometry. Infection efficiency was found to be around 95% after 72 h.p.i. 7
(Fig. 6C; upper panel). Comparing MHC I and epidermal growth factor receptor (EGFR) cell 8
surface expression on mock and SVV-infected cells revealed a specific downregulation for MHC 9
I on SVV-infected cells (Fig. 6C, middle panel). TAP function was assessed in mock-, SVV- and, 10
as a positive control, Rhesus (Rh)CMV-infected cells (Fig. 6D). When compared to mock-11
infected cells, peptide transport appeared unaffected in SVV-infected cells, while infection with 12
RhCMV resulted in a 60% reduction. 13
These data imply that the VZV- and SVV-induced downregulation of MHC I does not result from 14
a block of TAP-mediated peptide transport. 15
16
UL49.5 of MDV-1 is not responsible for MHC I downregulation by this virus 17
MDV-1 is known to reduce MHC I cell surface expression levels on infected cells18,26. To assess 18
if MDV-1 UL49.5 is responsible for this phenomenon, we transiently expressed chicken 19
hepatoma cells (LMH) and MJS cells with plasmids coding for the UL49.5 homologs of the 20
MDV-1 strains RB1B and CVI988. BoHV-1 UL49.5 was used as a control. At 48 hr post 21
transfection, cell surface expression of MHC I and, for MJS, MHC II were analyzed. None of the 22
UL49.5 proteins was able to reduce MHC I cell surface expression on LMH cells (Fig. 7). In 23
contrast, on MJS cells, BoHV-1 UL49.5 induced a reduction of MHC I that was not observed for 24
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

18
either MDV-1 homolog. The reduction by BoHV-1 UL49.5 was specific, as cell surface 1
expression of MHC II was unaffected. 2
In addition, baculoviruses were used to express BoHV-1 and MDV-1 CVI988 UL49.5 in MJS 3
cells. While BoHV-1 UL49.5-expressing cells displayed evident MHC I downregulation, a 4
reduction of MHC I cell surface levels could not be detected in cells with MDV-1 UL49.5 5
(Supplementary Fig. 1A). Also, TAP function was not affected by MDV-1 UL49.5, whereas 6
BoHV-1 UL49.5 reduced peptide transport by 40% in the same assay (Supplementary Fig. 1B). 7
These two independent experiments indicate that MDV-1 UL49.5 does not affect the transport of 8
peptides via TAP. 9
10
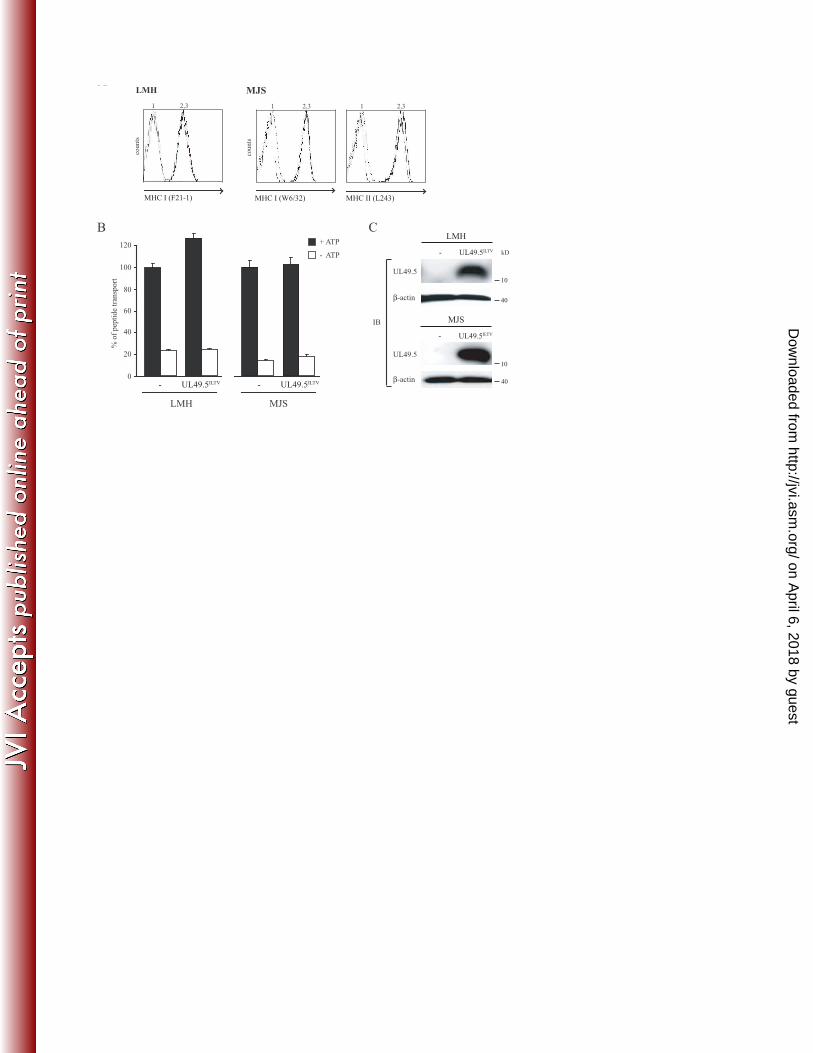
TAP function is not affected by ILTV UL49.5 11
To investigate whether ILTV UL49.5 interferes with peptide transport, the protein was expressed 12
in LMH or MJS cells by retroviral transduction. The analysis of MHC I cell surface levels on 13
UL49.5-expressing cells did not reveal ILTV UL49.5-induced downregulation (Fig. 8A). In 14
accordance, reduced TAP function was not detected in ILTV UL49.5-expressing cells (Fig. 8B). 15
Proper expression of ILTV UL49.5 was demonstrated using a specific antibody (Fig. 8C). All 16
together, these results indicate that ILTV UL49.5 does not function as a TAP inhibitor. 17
18
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

19
Discussion 1
2
Herpesviruses employ many strategies to avoid elimination by the host immune system20. A 3
strategy often exploited by these viruses is the inhibition of TAP-mediated peptide transport, 4
illustrating the key role of this process for immunity against these viruses. UL49.5 proteins of 5
varicelloviruses have been identified as a new class of TAP inhibitors that now includes the 6
UL49.5 homologs encoded by BoHV-1, BoHV-5, BuHV-1, CvHV-1, PRV, EHV-1, EHV-4 and 7
FeHV-128,30 and this study. SVV UL49.5 was found to have a minor effect on TAP-mediated 8
peptide transport, whereas VZV UL49.5 did not block TAP at all, despite its observed interaction 9
with the complex30. No TAP inhibition was found after infection with VZV or SVV, suggesting 10
that no other viral proteins contribute to TAP inhibition by these UL49.5 proteins, nor exhibit this 11
capacity themselves. Expression of the UL49.5 proteins encoded by alphaherpesviruses 12
belonging to the genera Mardivirus (MDV-1) and Iltovirus (ILTV) did not result in reduced 13
MHC I expression or inhibition of TAP function, suggesting that TAP inhibition is a property 14
unique to UL49.5 proteins encoded by varicelloviruses. 15
16
Within the genus Varicellovirus, clusters of closely related herpesviruses can be identified (Fig. 17
9). Varicelloviruses infecting ruminants, including BoHV-1, BoHV-5, BuHV-1, and CvHV-1, 18
evolved in close proximity46 and share a high degree of sequence identity (Table 2: the homology 19
between BoHV-1 and BoHV-5, BuHV-1 and CvHV-1 is 78.9%, 80.0% and 74.0%, respectively). 20
Here, we show that the UL49.5 proteins of these viruses affect TAP to the same extent and, most 21
probably, via the same mechanism. BoHV-1 UL49.5 renders the transporter in a translocation-22
incompetent state and induces the degradation of both TAP subunits. Correspondingly, the 23
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

20
UL49.5 proteins of BoHV-5, BuHV-1, and CvHV-1 also induce degradation of TAP. Thus, the 1
strategy to inhibit TAP seems conserved within the cluster of varicelloviruses of ruminants. 2
3
As displayed in figure 1, regions of homology between all UL49.5 proteins are limited and 4
scattered throughout the sequence. This makes it difficult to locate domains or individual amino 5
acid residues that might be responsible for TAP inhibition. So far, the only exception is the 6
UL49.5-mediated degradation of TAP, that appears to be related to defined amino acid residues 7
within the cytoplasmic tail of the UL49.5 protein28. Preliminary data suggest that the conserved 8
RGRG sequence at the C-terminus of BoHV-1, BoHV-5, BuHV-1, and CvHV-1 UL49.5 is 9
involved in the degradation process (M.C. Verweij, A.D. Lipińska, J. Koch (Georg-Speyer-Haus, 10
Frankfurt) and E.J.H.J. Wiertz, unpublished data). 11
12
CaHV-1 and FeHV-1, both infecting carnivores, are genetically and serologically related16. As 13
depicted in figure 9, these viruses are thought to have a common ancestor. The UL49.5 proteins 14
of these viruses share 53.5% sequence identity (Table 2). Earlier, we showed that CaHV-1 15
UL49.5 has a minor effect on TAP function in canine cells30. Surprisingly, in the present study, 16
FeHV-1 UL49.5 was identified as a potent inhibitor of feline TAP and, to a lesser extent, human 17
TAP. FeHV-1 and CaHV-1 are related to EHV-1 and EHV-4, herpesviruses infecting odd-toed 18
ungulates or Perrisodactyla (Fig. 9). The UL49.5 homologs encoded by these viruses were 19
previously shown to strongly inhibit TAP30. Hence, in this cluster of related varicelloviruses, 20
CaHV-1 appears to be the odd one out, coding for a UL49.5 protein that only moderately affects 21
TAP. This UL49.5 homolog might have lost the capacity to strongly inhibit TAP. 22
Previously, it has been shown that EHV-1 UL49.5 arrests the transporter in a translocation-23
incompetent state that is incompatible with ATP-binding30. In the presence of FeHV-1 UL49.5, 24
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

21
TAP was still capable of binding ATP. Thus, the mechanism through which FeHV-1 UL49.5 1
inhibits TAP differs from that utilized by the related EHV-1 and EHV-4 UL49.5 proteins. Similar 2
to FeHV-1 UL49.5, BoHV-1 and PRV UL49.5 do not interfere with ATP-binding to TAP30. 3
Presumably, differences in the capacity to inhibit ATP-binding result from slightly different 4
conformational changes in TAP that are induced by these proteins. 5
6
VZV and SVV belong to the evolutionary cluster of varicelloviruses that infect Old World 7
primates (Fig. 9). Despite an observed interaction between TAP and VZV UL49.5, this protein 8
does not inhibit TAP30. SVV UL49.5 moderately affects peptide transport in rhesus macaque 9
cells. Possibly, the UL49.5 proteins of these viruses have lost the capacity to (strongly) interfere 10
with TAP function or, alternatively, have never (fully) acquired this property. This somewhat 11
unique situation amongst varicelloviruses might be related to the evolutionary position of these 12
viruses, which seem to have separated from the other varicelloviruses at a relatively early stage 13
(Fig. 9). 14
Cells infected with VZV display reduced expression of MHC I at the cell surface2,8,11. The VZV 15
ORF66-encoded serine-threonine protein kinase has been shown to contribute to the observed 16
downregulation2,11, but additional proteins appear to be involved11. We did not observe reduced 17
TAP function in VZV-infected cells, indicating that VZV does not code for a TAP inhibitor. This 18
finding is rather surprising, as most of the alphaherpesviruses encode a TAP inhibitor20. The 19
isolated overexpression of the SVV UL49.5 protein resulted in a slightly reduced TAP activity, 20
however, this inhibition was not observed in SVV-infected cells. Therefore, it is unlikely that the 21
MHC I downregulation observed in SVV-infected cells is caused by inhibition of TAP by 22
UL49.5, or other SVV gene products. 23
24
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

22
All MDV serotypes, including MDV-1, MDV-2, and Meleagrid herpesvirus 1 or Turkey 1
herpesvirus (HVT) reduce MHC I expression on infected cells26. Highly virulent strains of MDV-2
1, such as RB1B, cause a stronger MHC I downregulation than less virulent or vaccine strains, 3
like CVI98818. Nevertheless, downregulation of MHC I was not detectable in chicken and human 4
cells expressing the UL49.5 proteins of RB1B and CVI988. In addition, MDV-1 CVI988 UL49.5 5
did not detectably affect TAP-mediated peptide transport in human cells. The UL49.5 protein of 6
ILTV did not reduce TAP-mediated peptide transport either. ILTV and the recently identified 7
Psittacid herpesvirus 1 (PsHV-1) are the only known iltoviruses38. These viruses have been 8
classified as members of the Alphaherpesvirinae10, but are distantly related to all other members 9
(Fig. 9). Similarly, MDV-1 branched away from the varicelloviruses and simplexviruses 10
relatively early in evolution (Fig. 9). Given their distant relatedness, ILTV and MDV-1 probably 11
evolved genes other than UL49.5 to circumvent MHC I-mediated antigen presentation. 12
13
In conclusion, TAP-inhibiting UL49.5 proteins have been identified uniquely in varicelloviruses, 14
now including BoHV-1, BoHV-5, BuHV-1, CvHV-1, PRV, EHV-1, EHV-4, and FeHV-1. These 15
UL49.5 homologs appear to have acquired a function in immune evasion, in addition to their 16
common function as a molecular chaperone for gM that seems to be conserved among all 17
herpesviruses. Possibly, this dual function relies on a hitherto unidentified structural similarity 18
between TAP and gM. 19
Despite the conserved function of the TAP-inhibiting UL49.5 proteins, their sequence identity 20
appears strikingly low. For example, the identity between the strong TAP-inhibitors BoHV-1 and 21
EHV-1 UL49.5 is only 30.2%. In contrast, the non-TAP inhibiting mardivirus MDV-1 UL49.5 22
shares 25.3% sequence identity with BoHV-1 UL49.5. The sequence identity between CaHV-1 23
and FeHV-1 UL49.5 is 53.5%, but the latter protein inhibits TAP much stronger than the former. 24
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

23
FeHV-1 UL49.5 displays functional resemblance to the BoHV-1, EHV-1, and PRV protein, with 1
which the sequence similarity is only around 30%. Thus, the homology between UL49.5 proteins 2
is not indicative of the capacity to inhibit TAP and, as suggested earlier, the different clades of 3
TAP-inhibiting homologs may have unique sequences that are mediating this function. 4
The UL49.5 homologs of the simplexviruses HSV-1 and HSV-2, the mardivirus MDV-1, the 5
iltovirus ILTV, the betaherpesvirus HCMV, and the gammaherpesvirus EHV have no effect on 6
TAP function28. However, within all three subfamilies other potent TAP inhibitors have been 7
identified (e.g. ICP47 in HSV-1 and -2, US6 in HCMV, and BNLF2a in EBV), making TAP 8
inhibition by herpesviruses a striking example of functional convergent evolution. 9
10
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

24
Acknowledgements 1
This work was supported by a grant from The Macropa Foundation, Leiden, The Netherlands (to 2
MCV), the START program of the Foundation for Polish Science (to ADL), the R01 grant 3
AG037042-01 from the National Institutes of Health (to I.M.) and the intramural research 4
program of the National Institute of Allergy and Infectious Diseases (to JIC). 5
We thank Femke Walraven-Berkhoff, Edwin Quinten, Guido de Roo and Menno van der Hoorn 6
for their technical support. 7
8
Abbreviations 9
CTL, cytotoxic T lymphocyte; Major histocampatibility complex I, MHC I; ER, endoplasmic 10
reticulum; TAP, transporter associated with antigen processing; PLC, peptide loading complex; 11
HSV, herpes simplex virus; HCMV, Human cytomegalovirus; EBV, Epstein-Barr virus; BoHV, 12
Bovine herpesvirus; EHV, Equid herpesvirus; PRV, Pseudorabies virus; VZV, Varicella-zoster 13
virus; CaHV, Canid herpesvirus; gM, glycoprotein M; BuHV-1, Bubaline herpesvirus 1; CvHV-14
1, Cervid herpesvirus 1; FeHV-1, Felid herpesvirus 1; SVV, Simian varicella virus; MDV-1, 15
Marek’s disease virus type 1; ILTV, Infectious laryngotracheitis virus; RhCMV, Rhesus CMV. 16
17
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

25
Reference List 1
2
1. Abele, R. and R. Tampe. 1999. Function of the transport complex TAP in cellular immune 3
recognition. Biochim. Biophys. Acta 1461:405-419. 4
2. Abendroth, A., I. Lin, B. Slobedman, H. Ploegh, and A. M. Arvin. 2001. Varicella-5
zoster virus retains major histocompatibility complex class I proteins in the Golgi 6
compartment of infected cells. J. Virol. 75:4878-4888. 7
3. Ahn, K., A. Gruhler, B. Galocha, T. R. Jones, E. J. Wiertz, H. L. Ploegh, P. A. 8
Peterson, Y. Yang, and K. Fruh. 1997. The ER-luminal domain of the HCMV 9
glycoprotein US6 inhibits peptide translocation by TAP. Immunity 6:613-621. 10
4. Ahn, K., T. H. Meyer, S. Uebel, P. Sempe, H. Djaballah, Y. Yang, P. A. Peterson, K. 11
Fruh, and R. Tampe. 1996. Molecular mechanism and species specificity of TAP 12
inhibition by herpes simplex virus ICP47. EMBO J. 15:3247-3255. 13
5. Aisenbrey, C., C. Sizun, J. Koch, M. Herget, R. Abele, B. Bechinger, and R. Tampe. 14
2006. Structure and dynamics of membrane-associated ICP47, a viral inhibitor of the MHC 15
I antigen-processing machinery. J. Biol. Chem. 281:30365-30372. 16
6. Amills, M., V. Ramiya, J. Norimine, and H. A. Lewin. 1998. The major 17
histocompatibility complex of ruminants. Rev. Sci. Tech. 17:108-120. 18
7. Barnstable, C. J., W. F. Bodmer, G. Brown, G. Galfre, C. Milstein, A. F. Williams, and 19
A. Ziegler. 1978. Production of monoclonal antibodies to group A erythrocytes, HLA and 20
other human cell surface antigens-new tools for genetic analysis. Cell 14:9-20. 21
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

26
8. Cohen, J. I. 1998. Infection of cells with varicella-zoster virus down-regulates surface 1
expression of class I major histocompatibility complex antigens. J. Infect. Dis. 177:1390-2
1393. 3
9. D'Arce, R. C., R. S. Almeida, T. C. Silva, A. C. Franco, F. Spilki, P. M. Roehe, and C. 4
W. Arns. 2002. Restriction endonuclease and monoclonal antibody analysis of Brazilian 5
isolates of bovine herpesviruses types 1 and 5. Vet. Microbiol. 88:315-324. 6
10. Davison, A. J., R. Eberle, B. Ehlers, G. S. Hayward, D. J. McGeoch, A. C. Minson, P. 7
E. Pellett, B. Roizman, M. J. Studdert, and E. Thiry. 2009. The order Herpesvirales. 8
Arch. Virol. 154:171-177. 9
11. Eisfeld, A. J., M. B. Yee, A. Erazo, A. Abendroth, and P. R. Kinchington. 2007. 10
Downregulation of class I major histocompatibility complex surface expression by 11
varicella-zoster virus involves open reading frame 66 protein kinase-dependent and -12
independent mechanisms. J. Virol. 81:9034-9049. 13
12. Fruh, K., K. Ahn, H. Djaballah, P. Sempe, P. M. van Endert, R. Tampe, P. A. 14
Peterson, and Y. Yang. 1995. A viral inhibitor of peptide transporters for antigen 15
presentation. Nature 375:415-418. 16
13. Fuchs, W. and T. C. Mettenleiter. 1996. DNA sequence and transcriptional analysis of the 17
UL1 to UL5 gene cluster of infectious laryngotracheitis virus. J. Gen. Virol. 77:2221-2229. 18
14. Fuchs, W. and T. C. Mettenleiter. 2005. The nonessential UL49.5 gene of infectious 19
laryngotracheitis virus encodes an O-glycosylated protein which forms a complex with the 20
non-glycosylated UL10 gene product. Virus Res. 112:108-114. 21
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

27
15. Garcia-Borges, C. N., B. Phanavanh, and M. D. Crew. 2006. Characterization of porcine 1
TAP genes: alternative splicing of TAP1. Immunogenetics 58:374-382. 2
16. Gaskell, R. and K. Willoughby. 1999. Herpesviruses of carnivores. Vet. Microbiol. 69:73-3
88. 4
17. Gaskell, R. M. and R. C. Povey. 1979. The dose response of cats to experimental infection 5
with feline viral rhinotracheitis virus. J. Comp Pathol. 89:179-191. 6
18. Gimeno, I. M., R. L. Witter, H. D. Hunt, L. F. Lee, S. M. Reddy, and U. Neumann. 7
2001. Marek's disease virus infection in the brain: virus replication, cellular infiltration, and 8
major histocompatibility complex antigen expression. Vet. Pathol. 38:491-503. 9
19. Halenius, A., F. Momburg, H. Reinhard, D. Bauer, M. Lobigs, and H. Hengel. 2006. 10
Physical and functional interactions of the cytomegalovirus US6 glycoprotein with the 11
transporter associated with antigen processing. J. Biol. Chem. 281:5383-5390. 12
20. Hansen, T. H. and M. Bouvier. 2009. MHC class I antigen presentation: learning from 13
viral evasion strategies. Nat. Rev. Immunol. 9:503-513. 14
21. Hengel, H., J. O. Koopmann, T. Flohr, W. Muranyi, E. Goulmy, G. J. Hammerling, U. 15
H. Koszinowski, and F. Momburg. 1997. A viral ER-resident glycoprotein inactivates the 16
MHC-encoded peptide transporter. Immunity 6:623-632. 17
22. Hewitt, E. W., S. S. Gupta, and P. J. Lehner. 2001. The human cytomegalovirus gene 18
product US6 inhibits ATP binding by TAP. EMBO J. 20:387-396. 19
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

28
23. Hill, A., P. Jugovic, I. York, G. Russ, J. Bennink, J. Yewdell, H. Ploegh, and D. 1
Johnson. 1995. Herpes simplex virus turns off the TAP to evade host immunity. Nature 2
375:411-415. 3
24. Hislop, A. D., M. E. Ressing, D. van Leeuwen, V. A. Pudney, D. Horst, D. Koppers-4
Lalic, N. P. Croft, J. J. Neefjes, A. B. Rickinson, and E. J. Wiertz. 2007. A CD8+ T cell 5
immune evasion protein specific to Epstein-Barr virus and its close relatives in Old World 6
primates. J. Exp. Med. 204:1863-1873. 7
25. Horst, D., D. van Leeuwen, N. P. Croft, M. A. Garstka, A. D. Hislop, E. Kremmer, A. 8
B. Rickinson, E. J. Wiertz, and M. E. Ressing. 2009. Specific targeting of the EBV lytic 9
phase protein BNLF2a to the transporter associated with antigen processing results in 10
impairment of HLA class I-restricted antigen presentation. J. Immunol. 182:2313-2324. 11
26. Hunt, H. D., B. Lupiani, M. M. Miller, I. Gimeno, L. F. Lee, and M. S. Parcells. 2001. 12
Marek's disease virus down-regulates surface expression of MHC (B Complex) Class I (BF) 13
glycoproteins during active but not latent infection of chicken cells. Virology 282:198-205. 14
27. Klupp, B. G., R. Nixdorf, and T. C. Mettenleiter. 2000. Pseudorabies virus glycoprotein 15
M inhibits membrane fusion. J. Virol. 74:6760-6768. 16
28. Koppers-Lalic, D., E. A. Reits, M. E. Ressing, A. D. Lipinska, R. Abele, J. Koch, R. M. 17
Marcondes, P. Admiraal, D. van Leeuwen, K. Bienkowska-Szewczyk, T. C. 18
Mettenleiter, F. A. Rijsewijk, R. Tampe, J. Neefjes, and E. J. Wiertz. 2005. 19
Varicelloviruses avoid T cell recognition by UL49.5-mediated inactivation of the 20
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

29
transporter associated with antigen processing. Proc. Natl. Acad. Sci. U. S. A 102:5144-1
5149. 2
29. Koppers-Lalic, D., M. Rychlowski, D. van Leeuwen, F. A. Rijsewijk, M. E. Ressing, J. 3
J. Neefjes, K. Bienkowska-Szewczyk, and E. J. Wiertz. 2003. Bovine herpesvirus 1 4
interferes with TAP-dependent peptide transport and intracellular trafficking of MHC class 5
I molecules in human cells. Arch. Virol. 148:2023-2037. 6
30. Koppers-Lalic, D., M. C. Verweij, A. D. Lipinska, Y. Wang, E. Quinten, E. A. Reits, J. 7
Koch, S. Loch, M. M. Rezende, F. Daus, K. Bienkowska-Szewczyk, N. Osterrieder, T. 8
C. Mettenleiter, M. H. Heemskerk, R. Tampe, J. J. Neefjes, S. I. Chowdhury, M. E. 9
Ressing, F. A. Rijsewijk, and E. J. Wiertz. 2008. Varicellovirus UL49.5 proteins 10
differentially affect the function of the transporter associated with antigen processing, TAP. 11
PLoS. Pathog. 4:e1000080. 12
31. Lehner, P. J., J. T. Karttunen, G. W. Wilkinson, and P. Cresswell. 1997. The human 13
cytomegalovirus US6 glycoprotein inhibits transporter associated with antigen processing-14
dependent peptide translocation. Proc. Natl. Acad. Sci. U. S. A. 94:6904-6909. 15
32. Lehner, P. J. and J. Trowsdale. 1998. Antigen presentation: coming out gracefully. Curr. 16
Biol. 8:R605-R608. 17
33. Levy, A. M., I. Davidson, S. C. Burgess, and H. E. Dan. 2003. Major histocompatibility 18
complex class I is downregulated in Marek's disease virus infected chicken embryo 19
fibroblasts and corrected by chicken interferon. Comp Immunol. Microbiol. Infect. Dis. 20
26:189-198. 21
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

30
34. Li, Q., M. A. Ali, and J. I. Cohen. 2006. Insulin degrading enzyme is a cellular receptor 1
mediating varicella-zoster virus infection and cell-to-cell spread. Cell 127:305-316. 2
35. Lipinska, A. D., D. Koppers-Lalic, M. Rychlowski, P. Admiraal, F. A. Rijsewijk, K. 3
Bienkowska-Szewczyk, and E. J. Wiertz. 2006. Bovine herpesvirus 1 UL49.5 protein 4
inhibits the transporter associated with antigen processing despite complex formation with 5
glycoprotein M. J. Virol. 80:5822-5832. 6
36. Mahalingam, R., M. Wellish, T. White, K. Soike, R. Cohrs, B. K. Kleinschmidt-7
DeMasters, and D. H. Gilden. 1998. Infectious simian varicella virus expressing the green 8
fluorescent protein. J. Neurovirol. 4:438-444. 9
37. McCluskey, J., J. Rossjohn, and A. W. Purcell. 2004. TAP genes and immunity. Curr. 10
Opin. Immunol. 16:651-659. 11
38. McGeoch, D. J., F. J. Rixon, and A. J. Davison. 2006. Topics in herpesvirus genomics 12
and evolution. Virus Res. 117:90-104. 13
39. Messaoudi, I., A. Barron, M. Wellish, F. Engelmann, A. Legasse, S. Planer, D. Gilden, 14
J. Nikolich-Zugich, and R. Mahalingam. 2009. Simian varicella virus infection of rhesus 15
macaques recapitulates essential features of varicella zoster virus infection in humans. 16
PLoS. Pathog. 5:e1000657. 17
40. Meyer, T. H., P. M. van Endert, S. Uebel, B. Ehring, and R. Tampe. 1994. Functional 18
expression and purification of the ABC transporter complex associated with antigen 19
processing (TAP) in insect cells. FEBS Lett. 351:443-447. 20
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

31
41. Montagnaro, S., M. Longo, M. Pacilio, P. Indovina, A. Roberti, M. L. De, G. Iovane, 1
and U. Pagnini. 2009. Feline herpesvirus-1 down-regulates MHC class I expression in an 2
homologous cell system. J. Cell Biochem. 106:179-185. 3
42. Ohta, Y., S. J. Powis, R. L. Lohr, M. Nonaka, L. D. Pasquier, and M. F. Flajnik. 2003. 4
Two highly divergent ancient allelic lineages of the transporter associated with antigen 5
processing (TAP) gene in Xenopus: further evidence for co-evolution among MHC class I 6
region genes. Eur. J. Immunol. 33:3017-3027. 7
43. Osterrieder, N., A. Neubauer, C. Brandmuller, B. Braun, O. R. Kaaden, and J. D. 8
Baines. 1996. The equine herpesvirus 1 glycoprotein gp21/22a, the herpes simplex virus 9
type 1 gM homolog, is involved in virus penetration and cell-to-cell spread of virions. J. 10
Virol. 70:4110-4115. 11
44. Powers, C. J. and K. Fruh. 2008. Signal peptide-dependent inhibition of MHC class I 12
heavy chain translation by rhesus cytomegalovirus. PLoS. Pathog. 4:e1000150. 13
45. St George, T. D. and M. Philpott. 1972. Isolation of infectious bovine rhinotracheitis virus 14
from the prepuce of water buffalo bulls in Australia. Aust. Vet. J. 48:126. 15
46. Thiry, J., V. Keuser, B. Muylkens, F. Meurens, S. Gogev, A. Vanderplasschen, and E. 16
Thiry. 2006. Ruminant alphaherpesviruses related to bovine herpesvirus 1. Vet. Res. 17
37:169-190. 18
47. Tomazin, R., A. B. Hill, P. Jugovic, I. York, P. van Endert, H. L. Ploegh, D. W. 19
Andrews, and D. C. Johnson. 1996. Stable binding of the herpes simplex virus ICP47 20
protein to the peptide binding site of TAP. EMBO J. 15:3256-3266. 21
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

32
48. Tomazin, R., N. E. van Schoot, K. Goldsmith, P. Jugovic, P. Sempe, K. Fruh, and D. C. 1
Johnson. 1998. Herpes simplex virus type 2 ICP47 inhibits human TAP but not mouse 2
TAP. J. Virol. 72:2560-2563. 3
49. van den Born, E., C. C. Posthuma, K. Knoops, and E. J. Snijder. 2007. An infectious 4
recombinant equine arteritis virus expressing green fluorescent protein from its replicase 5
gene. J. Gen. Virol. 88:1196-1205. 6
50. Wu, S. X., X. P. Zhu, and G. J. Letchworth. 1998. Bovine herpesvirus 1 glycoprotein M 7
forms a disulfide-linked heterodimer with the U(L)49.5 protein. J. Virol. 72:3029-3036. 8
9
10
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

33
Figure legends 1
2
Fig. 1 Alignment of the amino acid sequences of a selection of alphaherpesvirus UL49.5 3
proteins. The amino acid sequence alignment of UL49.5 homologs was performed using the 4
ClustalV of the MegAlign software from DNASTAR. BoHV-5 UL49.5: Bovine herpesvirus 5 5
(accession number NP_954898); BuHV-1 UL49.5: bubaline herpesvirus 1 (F.A.M. Rijsewijk 6
personal communication); BoHV-1 UL49.5: bovine herpesvirus 1 (NP_045309); CvHV-1 7
UL49.5: ceprid herpesvirus 1 (F.A.M. Rijsewijk personal communication); PRV/SuHV-1 8
UL49.5: pseudorabies virus (YP_068325); CaHV-1 UL49.5: canid herpesvirus 1 (patent: 9
EPO910406); FeHV-1 UL49.5: Felid herpesvirus 1 (YP_003331529); EHV-1 UL49.5: equid 10
herpesvirus 1 (YP_053055); EHV-4 UL49.5: equid herpesvirus 4 (NP_045227); VZV/HHV3 11
UL49.5: varicella-zoster virus (YP_068406); SVV UL49.5: simian varicella virus (NP_077423); 12
HSV-1 UL49.5: herpes simplex virus 1 (NP_044652); HSV-2 UL49.5: herpes simplex virus 2 13
(NP_044520); MDV-1/GaHV-2 UL49.5: Marek’s disease virus 1 strain CVI988 (NP_057812); 14
MDV-1 strain RB1B (YP_001033979.1) ILTV/GaHV-1 UL49.5: infectious laryngotracheitis 15
virus (YP_182341). The bars at the top of figure are proportional in height to the degree of 16
homology of amino acid conservation among the different viral proteins. 17
18
Fig. 2 The UL49.5 proteins of BoHV-5, BuHV-1, and CvHV-1 inhibit peptide transport and 19
mediate the degradation of TAP1 and TAP2. (A) Lysates derived from control and UL49.5-20
expressing MDBK and MJS cells were stained for GFP and UL49.5 by SDS-PAGE and 21
immunoblotting (IB) using specific antibodies. The β-actin signal was used as a loading control. 22
(B) Surface expression of MHC I (MDBK and MJS) and MHC II (MJS) molecules was assessed 23
by flow cytometry on untransduced cells (graph 2) and on cells expressing the UL49.5 homologs 24
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

34
of BoHV-1, BoHV-5, BuHV-1, and CvHV-1 (graph 3) using the indicated antibodies. Graph 1, 1
background staining in the presence of secondary antibody only. (C) Transport activity of TAP 2
was analyzed in MDBK and MJS expressing the UL49.5 homologs. Peptide transport was 3
evaluated in the presence of ATP (black) or EDTA (white bars). (D) Steady state levels of TAP1 4
and TAP2 in MJS cells were determined using specific antibodies. The β-actin signal was used as 5
a loading control. 6
7
Fig. 3 TAP is inhibited in FeHV-1-infected cells by an immediate-early or early protein. (A) 8
CRFK cells were mock-infected (graph 2) or infected with FeHV-1 for 12 hr in the absence 9
(graph 3) or presence (graph 4) of PAA. The expression of FeHV-1 glycoproteins and cell surface 10
expression of MHC I were assessed by flow cytometry using specific antibodies. Graph 1, 11
secondary antibody only. (B) TAP activity was determined in mock- and FeHV-1-infected cells 12
at 5 and 12 h.p.i. (in the absence and presence of PAA). The assay was performed in the presence 13
of ATP (black) or EDTA (white bars). 14
15
Fig. 4 FeHV-1 UL49.5 downregulates MHCI and strongly inhibits TAP-mediated peptide 16
transport. (A) Surface expression of MHC I (CRFK and MJS) and MHC II (MJS) molecules 17
was assessed by flow cytometry on untransduced cells (graph 2) and on cells expressing FeHV-1 18
UL49.5 (graph 3) using the indicated antibodies. Graph 1, background staining in the presence of 19
secondary antibody only. (B) Transport activity of TAP was analyzed in CRFK and MJS 20
expressing the FeHV-1 UL49.5. Peptide transport was evaluated in the presence of ATP (black) 21
or EDTA (white bars). (C) The steady state levels of TAP1 and GFP in FeHV-1 UL49.5-22
expressing cells were determined by SDS-PAGE and immunoblotting (IB) using specific 23
antibodies. The β-actin signal was used as a loading control. (D) Immunoblot analysis of ATP-24
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

35
bound (p) or unbound (s) TAP molecules found in CRFK or MJS cells expressing FeHV-1 1
UL49.5 or EHV-1 UL49.5 (control). 2
3
Fig. 5 SVV-encoded UL49.5 does not affect TAP function. Surface expression of MHC I 4
(LLC-MK2 and MJS) and MHC II (MJS) molecules was assessed by flow cytometry on 5
untransduced cells (graph 2) and SVV UL49.5-expressing cells (graph 3) using the indicated 6
antibodies. Graph 1, secondary antibody only. (B) Transport activity of TAP was analyzed in 7
LLC-MK2 and MJS expressing SVV UL49.5. Peptide transport was evaluated in the presence of 8
ATP (black) or EDTA (white bars). *A difference at p<0.05 was considered significant. (C) The 9
steady state levels of TAP1 and GFP in SVV UL49.5-expressing cells were determined by SDS-10
PAGE and immunoblotting (IB) using specific antibodies. 11
12
Fig. 6 VZV- and SVV-infection do not result in reduced TAP function. (A) MRC5 cells were 13
either mock-infected (graph 2), or infected with VZV.GFP or VZV.GFP-66 (graph 3) for 88 hr 14
and analyzed by flow cytometry to determine the expression of eGFP and the cell surface 15
expression of MHC I molecules and TfR using the indicated antibodies. Graph 1, secondary 16
antibody only. (B) MRC5 cells were mock- and VZV.GFP-infected for 72 hr, after which peptide 17
transport was assessed. As a control for TAP inhibition, cells were infected with BoHV-1 for 3 hr 18
at a m.o.i. of 20. The transport activity of TAP was analyzed in the presence of ATP (black) or 19
EDTA (white bars). (C) TRF cells were mock-infected (graph 2) or SVV-infected (graph 3) for 20
72 hr. eGFP expression and cell surface expression of MHC I and EGFR were determined via 21
flow cytometry. (D) TRF cells were mock- or SVV-infected and at 72 h.p.i. TAP activity was 22
determined. As a control for TAP inhibition, TAP activity was also determined in cells that were 23
infected with RhCMV for 36 hr at a m.o.i. of 1. 24
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

36
1
Fig. 7 The UL49.5 proteins of MDV-1 RB1B and CVI988 do not affect MHC I surface 2
expression. BoHV-1, MDV-1 RB1B and MDV-1 CVI988 UL49.5 were transiently expressed in 3
LMH and MJS cells. Cell surface expression levels of MHC I and MHC II molecules were 4
assessed by flow cytometry on cells transfected with a control plasmid (graph 2) or with the 5
UL49.5-expressing plasmids (graph 3) using specific antibodies. Graph 1, secondary antibody 6
only. 7
8
Fig. 8 ILTV-encoded UL49.5 does not affect TAP function. (A) Surface expression of MHC I 9
(LMH and MJS) and MHC II (MJS) molecules was assessed by flow cytometry on untransduced 10
cells (graph 2) and ILTV UL49.5-expressing cells (graph 3) using the indicated antibodies. Graph 11
1, secondary antibody only. (B) Transport activity of TAP was analyzed in LMH and MJS 12
expressing ILTV UL49.5. Peptide transport was evaluated in the presence of ATP (black) or 13
EDTA (white bars). (C) The steady state levels of UL49.5 (MJS and LMH) and TAP1 (MJS) 14
were determined using SDS-PAGE and immunoblotting (IB) using the indicated antibodies. The 15
β-actin signal was used as a loading control. 16
17
Fig. 9 Phylogenetic tree of a selection of alphaherpesviruses based on gB sequences. The 18
tree has been constructed using the Clustal W (slow/accurate, Gonnet) method of the alignment 19
program MegAlignTM 500 of the sequence analysis software of DNASTAR Inc. Genera and 20
species nomenclature were previously described by A. Davison et al.10. BoHV-5 gB: bovine 21
herpesvirus 5 (AAD46112.2); BuHV-1 gB: bubaline herpesvirus 1 (AAL88794.1); BoHV-1 gB: 22
bovine herpesvirus 1 (P12640.1); CvHV-1 gB: ceprid herpesvirus 1 (AAD46115.2); PRV/SuHV-23
1 gB: pseudorabies virus (ACT78489.1); CaHV-1 gB: canid herpesvirus 1 (AAK51052.1); 24
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

37
FeHV-1 gB: felid herpesvirus 1 (YP_003331552.2); EHV-1 gB: equid herpesvirus 1 1
(YP_053078.1); EHV-4 gB: equid herpesvirus 4 (P17472.1); VZV/HHV3 gB: varicella-zoster 2
virus (AAP32845.1); SVV gB: simian varicella virus (NC_002686.2); HSV-1 gB: herpes simplex 3
virus 1 (ABM66851.1); HSV-2 gB: herpes simplex virus 2 (AAA60540.1); MDV-1/GaHV-2 gB: 4
Marek’s disease virus 1 (BAA02866); ILTV/GaHV-1 gB: infectious laryngotracheitis virus 5
(CAA39573.1). OW: old world. 6
7
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

38
Table 1. PCR primers
Primer name a Primer sequence a
BoHV-5 UL49.5 Fw: 5’-GCCGGATCCGCTCCACGACGACCATGTCGCGC-3’
BuHV-1 UL49.5 Fw:5’-GCCGGATCCGACGACCATGTCGCGCTCGCT-3’
BoHV-5/BuHV-1
UL49.5
Rev: 5’-GCGGAATTCCGCTCAACCCCGCCCCCGCAC-3’
CvHV-1UL49.5 Fw: 5’-GCCGGATCCGAGCCGAGCACCATGGCGAGG-3’
CvHV-1UL49.5 Rev: 5’-GCGGAATTCCGGTCAGCCCCGCCCCCGCGA-3’
FeHV-1 UL49.5 Fw: 5’-CGGGATCCCACCATGGATCGTTTATCC-3'
FeHV-1 UL49.5 Rev: 5'-GCGGAATTCTTAGTGTGGCATGC-3'
SVV UL49.5 Fw:5’-GGGGACAAGTTTGTACAAAAAAGCAGGCT
GAATTCACCATGGCTTCAAATTGCTCTT-3’
SVV UL49.5 Rev: 5’-GGGGACCACTTTGTACAAGAAAGCTGGGTC
CTCGAG-TTACCATGTACTACGTAAGACGGATCG-3’
SVV-HA UL49.5 Rev: 5’-GGGGACCACTTTGTACAAGAAAGCTGGGTC
CTCGAG-TTAAGCGTAGTCTGGGACGTCGTATGG-3’
MDV-1 UL49.5 Fw: 5’-CGGGATCCCACCATGGGACTCATG-3’
MDV-1 UL49.5 Rev: 5'-CCGAATTCCTTACCACTCCTCTTTAAAC-3’
ILTV UL49.5 Fw: 5’-CGGGATCCCACCATGAGGCTGC-3’
ILTV UL49.5 Rev: 5’-GGAATTCCTACCATCGAGAACTAATGAC-3’
a = restriction sites used in bold
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

39
Table 2. Shown are the percentages of amino acid sequence identity between the various UL49.5
homologs described in this paper. The percentages were calculated using the ClustalV program
(parameters: PAM 250) of the MegAlign software from DNASTAR.
ILT
V
MD
V-1
(R
BIB
)
MD
V-1
(CV
I988
)
HS
V-2
HS
V-1
SV
V
VZ
V
EH
V-4
EH
V-1
FeH
V-1
CaH
V-1
PR
V
Cv
HV
-1
Bo
HV
-1
Bu
HV
-1
Bo
HV
-5
BoHV-5 18.9 22.1 22.1 23.0 20.9 26.4 24.1 31.6 32.6 28.4 30.1 30.5 72.6 78.9 94.7 100
BuHV-1 20.0 23.2 23.2 20.7 19.8 26.7 25.3 31.6 30.5 28.4 26.7 29.5 74.7 80.0 100
BoHV-1 18.8 23.1 23.2 21.8 20.9 25.3 25.3 29.2 30.2 30.5 27.9 33.3 74.0 100
CvHV-1 19.4 22.1 22.1 18.4 22.0 25.3 21.8 27.6 28.6 29.5 27.9 28.6 100
PRV 22.4 22.1 22.1 20.7 23.1 28.7 25.3 29.6 28.6 32.6 30.2 100
CaHV-1 15.1 26.7 26.7 14.0 15.1 26.7 29.1 46.5 44.2 53.5 100
FeHV-1 18.9 26.3 26.3 18.4 17.6 29.9 32.2 38.9 35.8 100
EHV-1 21.0 23.2 23.2 18.4 19.8 33.2 27.6 88.0 100
EHV-4 20.0 24.2 24.2 16.1 18.7 33.3 29.9 100
VZV 23.0 32.2 32.2 13.8 11.5 63.2 100
SVV 27.6 32.2 32.2 14.9 14.9 100
HSV-1 17.6 17.6 17.6 54.0 100
HSV-2 20.7 18.4 18.4 100
(CVI988) MDV-1 21.1 100 100
(RBIB) MDV-1 21.1 100
ILTV 100
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from


- BoHV-1 BoHV-5 BuHV-1 CvHV-1
120
100
80
60
40
20
0
tro
psnart e
di tpe
p fo
%
+ ATP
- ATP
MJS120
100
80
60
40
20
0
tro
psnart e
ditpe
p fo
%
MDBK
B
C
D
stn
uoc
1 3 2
MJS
stn
uoc
MDBK
stn
uoc
BoHV-1 BoHV-5 BuHV-1 CvHV-1
MHC I (W6/32)
MHC I (W6/32)
MHC II (L243)
1 3 2 1 3 2 1 3 2
1 2,3 1 3,2 1 2,3 1 2,3
1 3 2 1 3 2 1 3 2 1 3 2
UL49.5UL49.5
- BoHV-1 BoHV-5 BuHV-1 CvHV-1
MJS
UL49.5-ct
eGFP
β-actin
IB
- BoHV-1 BoHV-5 BuHV-1 CvHV-1 kD
AMDBK UL49.5
- BoHV-1 BoHV-5 BuHV-1 CvHV-1
UL49.5 UL49.5 UL49.5 UL49.5
- BoHV-1 BoHV-5 BuHV-1 CvHV-1
β-actin
IB
TAP1
TAP2
10
40
25
MJS UL49.5
UL49.5-ct
eGFP
β-actin
IB
kD
10
40
25
MJS UL49.5
kD
70
40
64
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from
120
100
80
60
40
20
0
% o
f p
epti
de
tran
spo
rt
+ ATP
- ATP
A
B
counts
1,2 4 3
- 5 h.p.i 12 h.p.i 12 h.p.i.
+ PAA
MHC I (H58A)
1 4,3 2
FeHV-1 (MCA2490)
CRFK FeHV-1, 12 h.p.i
CRFK FeHV-1
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

120
100
80
60
40
20
0
% o
f p
epti
de
tran
spo
rt
+ ATP
- ATP
MJS
A
B
counts
1 3 2
MJS CRFK
1 2,31 3 2
CRFK
- UL49.5FeHV-1 - UL49.5FeHV-1
- UL49.5FeHV-1
IB
C
eGFP
β-actin
CRFK MJS
counts
MHC I (W6/32) MHC II (L243)MHC I (H58A)
TAP1
- UL49.5FeHV-1
S P S P
UL49.5FeHV-1
fraction:
CRFK
-
1 2 3 4 5 6 7 8 9 10
S P S PS P
UL49.5FeHV-1
MJS
- UL49.5EHV-1
D
TAP1 (70 kD)
kD
70
40
25
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from
120
100
80
60
40
20
0
% o
f p
epti
de
tran
spo
rt
+ ATP
- ATP
MJS
A
B
counts
1 2,3
MJSLLC-MK2
1 2,31 3 2
LLC-MK2
- UL49.5SVV - UL49.5SVV
*
- UL49.5SVV
TAP1
eGFP
IB
C
eGFP
LLC-MK2
counts
MHC I (W6/32) MHC II (L243)MHC I (W6/32)
- UL49.5SVV
MJS
kD
25
25
70
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

120
100
80
60
40
20
0
tro
psnart e
d itpe
p fo
%
MRC5
A
B
stn
uoc
1 3 2
VZV.GFP
1 2,3
- VZV.GFP BoHV-1
MHC I (W6/32)
TfR (CD71)
stn
uoc
1 3 2
VZV.GFP-66
1 2 3
stn
uoc
stn
uoc
eGFP
stn
uoc
120
100
80
60
40
20
0
tro
psnart e
di tpe
p fo
%
TRF
C
D
stn
uoc
1 3 2
SVV.GFP
1 2,3
- SVV.GFP RhCMV
MHC I (W6/32)
EGFR (Ab-5)
stn
uoc
stn
uoc
eGFP
stn
uoc
+ ATP
- ATP
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

counts
MJS
counts
LMH
counts
BoHV-1
UL49.5
MDV-1 RB1B
UL49.5
MDV-1 CVI988
UL49.5
MHC I (W6/32)
MHC I (W6/32)
MHC II (L243)
3,21 3,
3,
3,21 21
3 21
,321
2 3,21
32, 32,1 1
1
MJS
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from
120
100
80
60
40
20
0
% o
f p
epti
de
tran
spo
rt
+ ATP
- ATP
MJS
A
B
counts
1 2,3
MJSLMH
1 2,31 2,3
LMH
- UL49.5ILTV - UL49.5ILTV
- UL49.5ILTV
UL49.5
β-actin
IB
C
counts
MHC I (W6/32) MHC II (L243)MHC I (F21-1)
UL49.5
β-actin
LMH
- UL49.5ILTV
MJS
kD
10
10
40
40
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

BoHV-5
BuHV-1
BoHV-1
CvHV-1
PRV (SuHV-1)
CaHV-1
FeHV-1
EHV-1
EHV-4
VZV (HHV-3)
SVV
HSV-1
HSV-2
MDV-1 (GaHV-2)
ILTV (GaHV-1)
Varicellovirus
Simplexvirus
Mardivirus
Iltovirus
Artiodactyl
Artiodactyl
Carnivore
Perissodactyl
OW primate
OW primate
Avian
Avian
UL49.5-mediated
TAP inhibition
+
+
+
+
+
+
±
±
+
+
-
-
-
-
-
on April 6, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from