1 comportamiento, ecología y selección...
TRANSCRIPT

3/16/15
1
Cronograma
Seminarios y TP's: Inicio 19 de marzo Primer seminario: 26/3 1. Mueller, J.C., F. Pulido and B. Kempenaers. Identification of a gene associated with avian migratory behaviour. Proceedings of the Royal Society B 278: 2848-2856. 2. Zuk, M, J.T. Rotenberry & R.M. Tinghitella. Silent night: adaptive disappearance of a sexual signal in a parasitized population of field crickets. Biology Letters 2: 521-524.
Ecología y Comportamiento Animal Departamento de Ecología, Genética y Evolución Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires Teórica #1: Comportamiento, ecología y selección natural

3/16/15
2
Breve historia del estudio del comportamiento (I)
Heráclito (480 AC, y otros filósofos estoicos) clasificaban a los seres animados en dos categorías: sin alma (todos los animales) y con alma (el hombre). Al no disponer de alma todos los animales, menos el hombre, actuaban irracionalmente por lo que no era posible estudiar su comportamiento. Aristóteles (384-322 AC) reconocía en todos los animales las capacidades de percepción, emociones, aprendizaje, memoria e inteligencia y situaba al hombre al mas alto nivel de su Scala Naturae (la capacidad intelectual del hombre no difería cualitativamente de la de las otras especies, los cambios eran graduales). En la edad media San Alberto Magno y Santo Tomas de Aquino eliminaron al hombre de la Scala Naturae de Aristóteles ya que lo consideraban único y con un comportamiento regido por la razón. Los animales actuaban por su ciego instinto natural (dicotomía instinto-razón). Esta dicotomía persiste más de 500 años.
Breve historia del estudio del comportamiento (II)
En el siglo XVII Descartes y sus seguidores aún planteaban la dicotomía instinto-razón, si bien la defensa de esta distinción era más moralista que científica. Salvo algunas excepciones (Hume, Voltaire, Rousseau) hasta la aparición de ”El Origen de las Especies" (Darwin 1859) la conducta dependía de dos conceptos mutuamente excluyentes: el instinto y la razón siendo esta última una prerrogativa exclusiva del hombre. Darwin (1859, 1871, 1872) plantea que la morfología, la fisiología y el comportamiento se integran en un conjunto de adaptaciones que le permiten a los individuos sobrevivir y reproducirse. Al descartar la idea de discontinuidad en la evolución (comunidad de descendencia), no precisa analizar al hombre en forma independiente (su comportamiento sería derivado del que tenían sus ancestros primates). Romanes (discípulo de Darwin) en su libro “Animal Intelligence” (1882) provee información comparativa que apoya la teoría de Darwin.

3/16/15
3
Breve historia del estudio del comportamiento (III)
A comienzos del siglo XX, el estudio del comportamiento animal pone el énfasis en los procesos de aprendizaje. Se destacan el fisiólogo ruso Ivan P. Pavlov (condicionamiento clásico) y el psicólogo norteamericano Edward L. Thorndike (condicionamiento instrumental). Entre 1930 y 1950 Konrad Lorenz desarrolla varios modelos teóricos sobre los aspectos motivacionales del comportamiento (teorías de "mecanismo desencadenante innato", "suma heterogénea de estímulos" y "energía de acción específica”). Además profundiza el estudio de un tipo especial de aprendizaje que denomina ”imprinting". Niko Timbergen es quien por primera vez plantea en forma explícita la necesidad de comprender el valor adaptativo del comportamiento de los animales estudiándolos en su ambiente natural. Su libro “El estudio del instinto” (1951) plantea explícitamente cómo los componentes “innatos” del comportamiento son el resultado del proceso de selección natural.
Breve historia del estudio del comportamiento (IV)
La teoría del instinto de los etólogos clásicos fue muy criticada por la escuela de psicólogos experimentales norteamericanos (controversia natura-nurtura). Principal crítica (Lehrman 1953) era que muchos de los ejemplos de comportamientos innatos se basaban en datos inadecuados en los que no se había eliminado (controlado) la posibilidad de aprendizaje. Esta escuela “behaviorista” (Watson, Skinner) ponía el énfasis en el efecto del aprendizaje sobre el comportamiento. Para ellos el aprendizaje era la formación de asociaciones entre estímulos y respuestas (entidades visibles). Ellos consideraban que no era posible hacer afirmaciones científicas acerca de procesos mentales (la introspección no era confiable y la mente de los animales inaccesible). Como reacción a esta escuela en la década del 50 surge la escuela de los psicólogos cognitivos (Chomsky, Broadbent) quienes planten que los animales tienen una vida mental que si puede ser investigada científicamente. Robert Hinde (zoólogo inglés discípulo de Timbergen) contribuye en forma importante a la unificación entre "etólogos clásicos" y "behavioristas" (1959, 1966).

3/16/15
4
Científicos influyentes en la etología “clásica”
Karl von Frisch (1886-1982)
Premio Nobel en fisiología o medicina 1973 "for their discoveries concerning organization and elicitation of individual and social behaviour patterns"
Comunicación en insectos sociales
Aspectos motivacionales del comportamiento
desarrollo, conducta como rasgo filogenético
Visión evolutiva y ecológica, valor adaptativo del
comportamiento, experimentos naturales
Ecología del Comportamiento
Konrad Lorenz (1903-1989)
Nikolaas Tinbergen 1907-1988
Desarrollo de la etología moderna Durante la primer mitad de la década del 60 la etología desarrolla una metodología adecuada pero sigue teniendo un enfoque naturalista, carente de un marco conceptual y teórico general. Desde mediados de los 60 hasta mediados de los 70 se produce una revolución conceptual que se centra en entender el comportamiento como el resultado de la selección natural actuando a nivel de los individuos. Las principales ideas desarrolladas en este periodo son: • El abandono de la idea de selección natural actuando a nivel de grupos en favor de la idea de selección natural actuando a nivel de individuos (Maynard Smith 1964, George G. Williams 1966). • El desarrollo de la teoría de selección parental (kin selection, Hamilton 1964, 1966). • La adopción de modelos matemáticos de optimización (teoría de consumo óptimo, Charnov 1973). • La aplicación de la teoría de juegos a problemas biológicos y desarrollo del concepto de Estrategias Evolutivamente Estables (Maynard Smith 1972, 1982). • Una revalorización de la importancia de los mecanismos de selección sexual para explicar los sistemas de apareamiento, la elección de pareja y el cuidado e inversión parental (Trivers 1972, 1974).

3/16/15
5
Desarrollo de la etología moderna
Esta revolución conceptual se plasma en la publicación del libro Sociobiología, la nueva síntesis (Edward O. Wilson, 1975) y posteriormente en la publicación de las distintas ediciones de An Introduction to Behavioural Ecology (1981, 1986, 1993, 2012) y Behavioural Ecology: An Evolutionary Approach (1978, 1984, 1991, 1997) por John R. Krebs y Nicholas B. Davies.
Ecología del comportamiento
La ecología del comportamiento es una rama de las ciencias biológicas que intenta responder preguntas sobre el valor adaptativo del comportamiento (consecuencias del comportamiento sobre el éxito reproductivo de los individuos). Los patrones comportamentales son analizados como el resultado de la selección natural (han evolucionado como resultado de presiones de selección del ambiente físico y social en el que los animales viven). En la actualidad, la mayoría de los estudios en comportamiento animal tienen un enfoque que contempla tratar de entender el valor adaptativo del comportamiento. Principales revistas: Animal Behaviour, Behavioral Ecology, Behavioral Ecology and Sociobiology, Ethology, Behaviour. Principales sociedades: International Society of Behavioral Ecology, Animal Behavior Society-Asociation for the Study of Animal Behaviour.

3/16/15
6
Distintos tipos de preguntas en comportamiento: causas próximas y causas últimas
Ernst Mayr (1961) distinguió entre los biólogos que estudian las causas próximas (mecanismos) y aquellos que estudian las causas últimas (evolución). Niko Timbergen (On aim and methods of ethology, 1963) planteó que cada una de estas categorías podía a su vez ser subdividida en otras dos. Las preguntas de las causas próximas, o de “cómo”, pueden responderse en términos de los mecanismos fisiológicos del comportamiento (neurales, hormonales) o de su ontogenia (efectos de la edad y experiencia). Las preguntas de las causas últimas, o de “por qué”, pueden responderse en términos del valor adaptativo del comportamiento o de su historia evolutiva.
Los cuatro por qué de Timbergen (1963) a) Mecanismos responsables del comportamiento b) Valor adaptativo o función del comportamiento c) Desarrollo u ontogenia del comportamiento. d) Origen evolutivo o filogenia del comportamiento.
Cuatro tipos de preguntas en comportamiento
Ontogenia Filogenia
Valor adaptativo Mecanismos
Causas próximas
Causas últimas
Algunos autores plantean un quinta forma en términos de los procesos cognitivos y emocionales subyacentes a ese comportamiento (motivación, conciencia, pensamiento) pero estos procesos son, en última instancia, procesos fisiológicos (causas próximas) aún no totalmente comprendidos.

3/16/15
7
Efecto Bruce Hilda Bruce 1959. También denominado bloqueo del embarazo. Tendencia de las hembras de algunas especies de roedores a interrumpir el embarazo luego de la exposición a olores de machos no familiares (ej. los progenitores de las crías). También presente en una especie de primates (Theropithecus gelada, Science 335: 1222-1225, 2012) y postulado en leones. ¿Por qué las hembras interrumpen el embarazo ante la presencia de un macho desconocido?
Efecto Bruce (mecanismo) Mecanismo estudiado en detalle en ratones. La orina de los machos contiene proteínas del tipo MHC (Complejo Mayor de Histocompatibilidad) clase I que se unen a receptores del órgano vomeronasal (estructura llena de mucus en el septum nasal). Estas señales químicas son específicas de cada macho y serían aprendidas por la hembra durante el apareamiento o inmediatamente después del apareamiento. La hormona vasopresina participa del acoplamiento entre la señal química y la respuesta fisiológica (en animales que tienen "knocked out" el receptor VP 1b la señal química no interrumpe el embarazo). Otras hormonas involucradas son noradrenalina (baja la receptividad al estímulo) y oxitocina (facilita el aprendizaje del olor del macho). La información va del órgano vomeronasal al bulbo olfatorio, de ahí a la amígdala (zona corticomedial del cerebro), tracto olfatorio accesorio y stria terminalis y finalmente al hipotálamo. El hipotálamo aumenta la secreción de dopamina que inhibe la prolactina. La ausencia de prolactina inhibe la secreción de progesterona por el cuerpo lúteo y eso resulta en la interrupción del embarazo.

3/16/15
8
Efecto Bruce (función) ¿Cuál es el posible valor adaptativo del efecto Bruce para machos y hembras? Machos Se beneficiarían por la interrupción del embarazo de la hembra ya que de esta forma pueden aparearse inmediatamente con ella (mayor probabilidad de dejar descendientes antes de ser desplazados por otro macho -o ser depredados-). Machos dominantes bloquean posibles embarazos por parte de machos subordinados (los dominantes dejan más marcas de orina que los subordinados). ¿Hembras? En muchas especies de roedores los machos matan a crías que no están relacionadas genéticamente con él (infanticidio). El efecto Bruce es más frecuente en especies de roedores con poliginia. En estas especies el riesgo de infanticidio es mayor. La hembra "salvaría" tiempo y recursos interrumpiendo el embarazo. La hembra podría controlar su embarazo buscando o evitando nuevos machos (elección de pareja post-apareamiento).
Causas próximas y últimas
Desarrollo Expresión de genes Interacciones genes-ambiente
Fisiología
Sistema motor Sistema nervioso Sistema endócrino
Comportamiento Exito reproductivo diferencial Cambios en la frecuencia génica en la siguiente generación

3/16/15
9
“Fitness” o aptitud biológica Darwiniana
Es una medida de la habilidad de un individuo para producir descendencia viable y contribuir a las generaciones futuras. Los individuos de una población varían en su fitness relativo y estas diferencias pueden deberse en mayor o menor medida a diferencias genéticas entre los individuos. También pueden deberse a diferencias del ambiente en que se desarrollaron esos individuos. Los individuos de una población que tienen mayor fitness van a contribuir desproporcionadamente a la siguiente generación. Si las diferencias en fitness tienen un componente genético, entonces, la composición genética de la siguiente generación va a ser modificada. Este proceso se conoce como selección natural.
Selección natural (Darwin 1859) • Los individuos de una misma especie difieren en su morfología, fisiología y/o comportamiento (variación).
• Algunas de estas diferencias son heredables, es decir, los hijos tienden a parecerse más a sus progenitores que a otros individuos de la población (herencia).
• Los organismos tienen una gran capacidad para reproducirse y, en general, producen muchos más individuos de los que llegan a la edad reproductiva. El número de individuos en una población permanece más o menos constante a través de tiempo. Por lo tanto, entre los individuos existe competencia por recursos escasos como alimentos, parejas, o lugares donde vivir.
• Como resultado de esta competencia, algunas variantes dejarán más descendientes que lleguen a la edad reproductiva que otras. Estas variantes van a heredar las características de sus padres y de esta forma tendrá lugar un cambio evolutivo por selección natural.
• Como consecuencia de este cambio evolutivo, los organismos van a adaptarse a su ambiente. Los individuos seleccionados serán aquellos mas eficientes para encontrar recursos escasos como alimentos, parejas o lugares donde vivir.

3/16/15
10
Selección natural (Endler 1986)
La selección natural puede ser definida como un proceso en el cual si una población tiene: a) Variación entre sus individuos en algún atributo o carácter: Variación b) Una relación consistente entre este carácter y la habilidad para aparearse, habilidad para fertilizar, fecundidad y/o supervivencia: diferencias en fitness o eficacia biológica Darwiniana c) Una relación consistente, para ese carácter, entre padres e hijos, la cual es al menos parcialmente independiente de efectos ambientales comunes: herencia Entonces: 1) La frecuencia de dicho carácter va a diferir entre clases de edad mas allá de lo esperado por la ontogenia. 2) Si la población no esta en equilibrio, entonces la distribución del carácter en el conjunto de los hijos va a ser diferente la distribución del carácter en el conjunto de los padres, más allá de lo esperado por las condiciones a y c.
Comportamiento, ecología, y evolución
El éxito de un individuo en sobrevivir y reproducirse depende (en gran parte) de su comportamiento. La selección natural va a favorecer (i.e. éxito reproductivo diferencial) a aquellos individuos que sean más eficientes buscando alimentos, evitando predadores, copulando, alimentando y defendiendo a sus hijos, etc. El grado de adaptación de estos comportamientos dependerá de las distintas alternativas comportamentales disponibles, las que a su vez van a depender de restricciones impuestas por la filogenia, la fisiología y/o la ecología. Adaptación: Cualquier carácter heredable (morfológico, fisiológico o comportamental) que posea un organismo y que ayude a su supervivencia o reproducción. Característica que ha sido modelada por selección natural u otro tipo de selección (i.e. sexual) para su función actual (Gould & Vrba 1982).

3/16/15
11
Puesta a prueba de hipótesis acerca del valor adaptativo de un determinado comportamiento
1) Comparación entre individuos de una especie (observaciones-experimentos naturales). 2) Experimentación (experimentos controlados). 3) Comparación entre especies (método comparado). Ej: ¿Cuál es el valor adaptativo de vivir en grupos?
Puesta a prueba de hipótesis acerca del valor adaptativo de un determinado comportamiento
Observaciones vs. Experimentos Asociación no implica causalidad. Experimento: manipulación por parte del experimentador de la variable independiente (aquella que se supone causa el efecto que queremos medir o variable dependiente). Validez interna vs. validez externa Validez interna: cuan adecuada es la metodología de estudio y sus resultados para responder la pregunta formulada. Validez externa: cuan extrapolables son mis estudios y resultados a otras poblaciones, especies o situaciones.
Principio de parsimonia Si dos hipótesis explican igualmente bien un fenómeno, siempre debe darse preferencia a aquella que sea mas simple (menor número de supuestos). Hipótesis alternativas

3/16/15
12
Medidas directas e indirectas utilizadas para estimar el “fitness” de un determinado comportamiento
Medidas directas del éxito reproductivo 1. Producción de gametas 2. Tasa de cópulas 3. Producción de huevos fertilizados 4. Producción de crías (nacidas) 5. Producción de crías (independientes). 6. Producción de crías durante una temporada reproductiva que alcanzan la
edad reproductiva. 7. Producción de crías durante toda la vida del individuo que alcanzan la edad
reproductiva (éxito reproductivo a lo largo de la vida). Medidas indirectas del éxito reproductivo 1. Eficiencia de locomoción u otra actividad. 2. Acceso a alimento. 3. Acceso a un territorio o pareja. 4. Probabilidad de sobrevivir.
Niveles a los que actúa la selección natural: ¿selección a nivel de individuos o grupos?
Podemos arreglar los niveles de selección a lo largo de un continuo que describe el grado de relación genética.
genes -> individuos -> parientes (kin) -> grupos -> especies Selección a nivel de genes planteada originalmente por Williams (1966) y Dawkins (1976). Genes actúan como replicadores y son preservados a través del tiempo. Los organismos son los vehículos que transportan los genes. Esta idea fue criticada argumentándose que la selección no puede actuar directamente sobre los genes sino sólo a través de sus “envases”. Hull (1980) introduce los conceptos de “replicador” (entidad que pasa con su estructura intacta directamente en la replicación), e “interactor” (entidad que es capaz de interaccionar como un todo con el medio ambiente de manera que se produce replicación diferencial de los distintos interactores).

3/16/15
13
Niveles a los que actúa la selección natural: ¿selección a nivel de individuos o grupos?
Selección de grupos en su forma más simple planteaba que la evolución de un carácter podía ser explicada “por el bien de la especie” (Wynne-Edwards 1962: autocontrol de la natalidad para no sobreexplotar los recursos o predador prudente). Argumentos de Wynne-Edwards (y otros biólogos de la época -Lorenz, Huxley) fueron refutados por Willams (Adaptation and Natural Selection, 1966). La selección a nivel de grupos (o especies) sería un proceso análogo al de selección individual (grupos tienen variabilidad, heredabilidad y diferencias en éxito reproductivo). Ocurriría cuando un grupo de individuos (o un grupo de especies estrechamente relacionadas) produce más grupos de descendientes (o especies) que otros grupos de individuos (o clados). Condiciones para que ocurra selección de grupos son mucho más restrictivas que para que ocurra selección individual.
Tamaño de puesta en Parus major
Histograma de frecuencias del tamaño de puesta (número de huevos puestos) en el Carbonero Común (Parus major) en Whytam woods Oxford. ¿Por qué algunos individuos ponen 5 o 6 huevos si podrían poner 11 o 12?
3/16/15
14
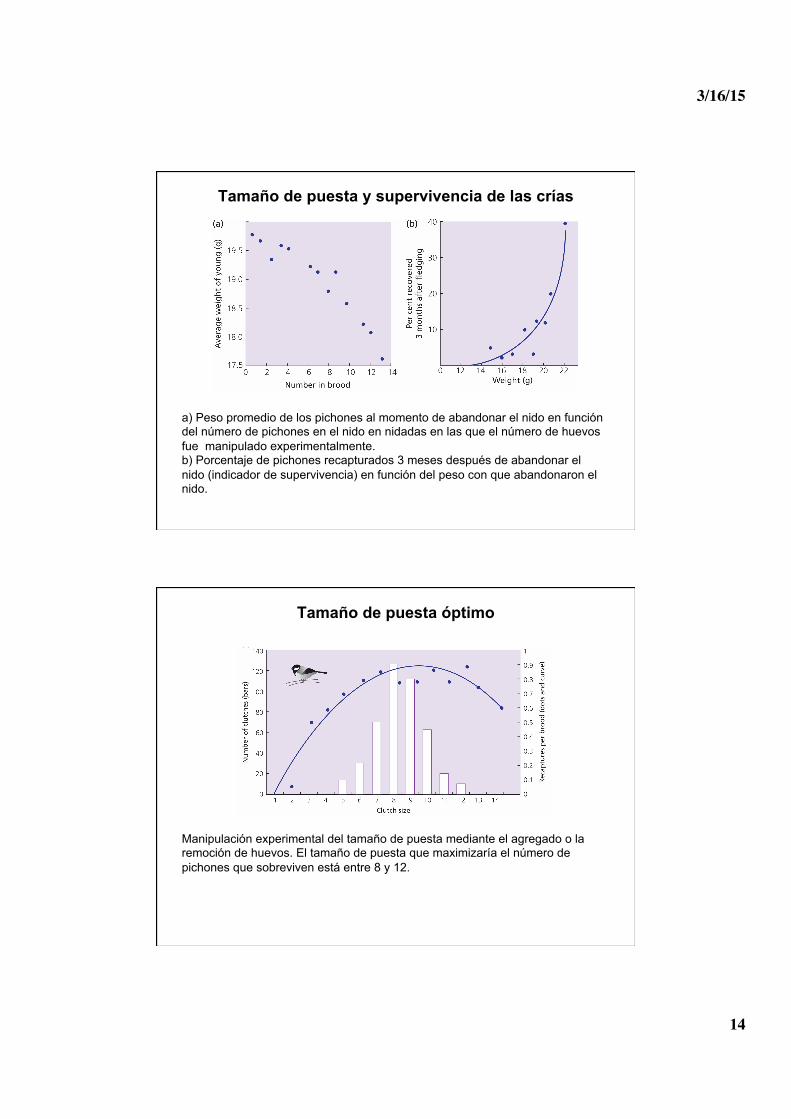
Tamaño de puesta y supervivencia de las crías
a) Peso promedio de los pichones al momento de abandonar el nido en función del número de pichones en el nido en nidadas en las que el número de huevos fue manipulado experimentalmente. b) Porcentaje de pichones recapturados 3 meses después de abandonar el nido (indicador de supervivencia) en función del peso con que abandonaron el nido.
Tamaño de puesta óptimo
Manipulación experimental del tamaño de puesta mediante el agregado o la remoción de huevos. El tamaño de puesta que maximizaría el número de pichones que sobreviven está entre 8 y 12.

3/16/15
15
Maximización del éxito reproductivo
Efecto de la mortalidad de los adultos sobre el tamaño de puesta óptimo. El número de descendientes presenta un máximo en b1 pero si se tiene en cuenta la mortalidad de los adultos el tamaño de puesta óptimo (aquel que maximiza el número de descendientes a lo largo de la vida) es b2.
Éxito reproductivo o a lo largo de la vida
Éxito reproductivo de hembras de Parus major. El éxito reproductivo fue medido como: supervivencia de la hembra hasta la siguiente temporada reproductiva x (número de crías que sobrevivieron hasta la próxima temporada reproductiva x 0.5). Los datos corresponden a 3 grupos experimentales en los que las hembras criaron 2 pichones adicionales pagando los costos totales (poner huevos y criar a los pichones), sólo el costo de poner huevos (free eggs), o sólo el costo de criar pichones (free chicks). El fitness de la hembra está expresado relativo al del grupo control (sin crías extras).

3/16/15
16
Los individuos pueden aumentar su “fitness” a través de distintas vías: Fitness directo: Componente del fitness obtenido a través de la reproducción personal (i.e. producción de hijos). Fitness indirecto: Componente del fitness obtenido a través de ayudar a la supervivencia de individuos emparentados que no son descendientes directos (i.e. ayudar a un hermano o a los padres a producir hijos extra). Fitness inclusivo: La suma del fitness directo e indirecto. Un indicador del grado de parentesco comúnmente utilizado es el coeficiente de parentesco (r). r = probabilidad que un gen presente en un individuo sea una copia idéntica por descendencia de un gen presente en otro individuo.
Fitness directo, indirecto, e inclusivo
Relación
Padre < > hijo
Hermanos completos
Medios hermanos
Hermanos gemelos
Hermanos mellizos
Abuelo < > nieto
Tío/a < > sobrino/a
Primos hermanos
Coeficientes de parentesco en organismos diploides
r
0.5
0.5
0.25
1
0.5
0.25
0.25
0.125

3/16/15
17
Coeficientes de parentesco en organismos haplodiploides
Relación
Madre < > hija
Padre > hija
Hija > padre
Madre > hijo
Hijo > madre
Hermanos
Hermanas
Hermano > hermana
Hermana > hermano
r
0.5
1
0.5
0.5
1
0.5
0.75
0.5
0.25
Regla de Hamilton
Un individuo ayuda a otro a producir un número b de descendientes extra pero incurre en un costo de c descendientes directos. De acuerdo a Hamilton el carácter altruista va a aumentar de frecuencia en la población si:
r1b - r2c > 0 donde r1 es el coeficiente de parentesco entre el altruista y la descendencia del beneficiario y r2 es el coeficiente de parentesco entre el altruista y su propia descendencia. La ecuación anterior puede convertirse en:
rb - c > 0 donde r es el coeficiente de parentesco entre el altruista y el beneficiario. Otra forma de la misma ecuación es:
b/c > 1/r

3/16/15
18
Eusocialidad en insectos “But with the working ant we have an insect differing greatly from its parents, yet absolutely sterile; so that it could never have transmitted successively acquired modifications of structure or instinct to its progeny. It may well be asked how is it possible to reconcile this case with the theory of natural selection?” Darwin C. (1859). The Origin of Species. “Pero en la hormiga obrera tenemos un insecto que difiere marcadamente de sus padres y es absolutamente estéril por lo que nunca podría haber transmitido satisfactoriamente a su progenie las modificaciones de estructura o instinto adquiridas. Uno bien podría preguntarse ¿cómo es posible reconciliar este caso con la teoría de la selección natural?”
Eusocialidad en insectos "Using this concept it soon becomes evident that family relationships in Hymenoptera are potentially very favourable to the evolution of reproductive altruism. If a female is fertilized by only one male all the sperm she receives is genetically identical. Thus, although the relationship of a mother to her daughters has the normal value of 1/2, the relationship between daughters is 3/4. Consider a species where the female consecutively provisions and oviposits in cell after cell so that she is still at work when the first of her female offspring ecloses, leaves the nest and mates. Our principle tells us that even if this new adult had a nest ready constructed and vacant for her use she would prefer, other things being equal, returning to her mother’s and provisioning a cell for the rearing of an extra sister to provisioning a cell for a daughter of her own. Hamilton, W.D. (1964). The Genetical Evolution of Social Behaviour. II. Journal of Theoretical Biology 7: 17-52.
rb - c > 0