拟南芥 arf2...
DESCRIPTION
拟南芥 ARF2 对生长素信号传导、细胞分裂、种子大小以及其它器官组织的影响. 报告人 : 王爱华. 一、背景 1 、胚不是决定种子大小的唯一因素。 2 、大型的 Cvi 种子主要是因为种皮和胚乳快速生长和延长发育。 3 、少量突变体基因影响种子大小。( miniature 1 , hairu , TRANSPARENT TESTA , AINTEGUMENTA ) 4 、 mnt/arf2 突变体证明了受精之前胚珠的发育程度决定最终种子大小。( megaintegumenta , AUXIN RESPONSE FACTOR 2 ). 二、结果 - PowerPoint PPT PresentationTRANSCRIPT

拟南芥拟南芥 ARF2ARF2对生长素信号对生长素信号传导、细胞分裂、种子大小传导、细胞分裂、种子大小以及其它器官组织的影响以及其它器官组织的影响
报告人 : 王爱华

一、背景一、背景11 、胚不是决定种子大小的唯一因素。、胚不是决定种子大小的唯一因素。22 、大型的、大型的 CviCvi 种子主要是因为种皮和胚乳快种子主要是因为种皮和胚乳快
速生长和延长发育。 速生长和延长发育。 33 、少量突变体基因影响种子大小。、少量突变体基因影响种子大小。
(( miniature miniature 11 ,, hairuhairu ,, TRANSPARENT TRANSPARENT TESTATESTA ,, AINTEGUMENTAAINTEGUMENTA ))
44 、、 mnt/arf2mnt/arf2 突变体证明了受精之前胚珠的突变体证明了受精之前胚珠的发育程度决定最终种子大小。发育程度决定最终种子大小。(( megaintegumentamegaintegumenta ,, AUXIN AUXIN RESPONSE FACTOR 2 RESPONSE FACTOR 2 ))

二、结果1 、筛选mnt突变体2 、大的mnt种子内腔贯穿发育全过程3 、 mnt突变体胚的增大归因于珠被中特大型细胞的分裂4 、 mnt突变体中影响种子重量的因素5 、 mnt突变体的花和植株生长发育6 、 mnt 是 ARF2 的等位基因7 、 ARF2 在生殖器官中的表达 8 、 arf2 突变体中促进细胞分裂的基因表达被延长

Fig. 1. Pleiotropic effects of the mnt mutation.
(A-D) Mature seeds (A,B) and embryos (C,D) from self-pollinated wild-type Col-3 (A,C) and mnt mutant plants (B,D). (E) Wild-type (left) and mnt (right) plants, showing the mnt stem phenotype. (F,G) Inflorescences from wild-type (F) and mnt plants (G); insets show flowers at similar stages.
返回

Fig. 2. Seed development in wild-type and mnt mutant plants.(A-J) Cleared seeds imaged with differential contrast optics; (A-E) wildtype and (F-J) mnt seeds at similar stages of embryogenesis. Scale bar: 100 μm. (K) CLSM images of Feulgen-stained seeds at 5 dap. Scale bars: 20 μm in embryo and chalazal endosperm; 25 μm in peripheral endosperm.
返回

Fig. 3. Ovule development in wild-type and mnt mutant plants.(A-F) Early ovule development in wild type (A,C,D) and mnt (B,E,F). (A,B) SEMs of stage 1-II ovules. (C,E) Sections of stage 2-III ovules showing megaspore mother cell (arrow) and initiating inner integument (*). (D,F) SEMs of later stage 2 ovules showing integuments beginning to extend over nucellus. (G,H) Sections of mature wild-type (G) and mnt (H) ovules. The arrow in (H) shows an extra cell layer. Scale bars: 10 μm in A-F; 20 μm in G,H. (I-K) Comparison of number of cells (I), total length of integument (J) and mean size of cells (K) in the abaxial oi2 and ii1’ layers of wild-type and mnt integuments. Error bars: s.e.m. n=4 wild type, 8 mnt.
返回

Fig. 4. Comparison of seeds generated by wild-type and mnt mutant plants. (A-D) Seeds from pollinations where all secondary shoots were allowed to set seed; (E-H) seeds from pollinations where secondary shoots were removed (see text). (A,E) [wild type × wild type]; (B,F) [wild type × mnt]; (C,G) [mnt × mnt]; (D,H) [mnt × wild type]. Scale bar: 250 μm. (I) Comparison of seed weights following the two treatments. Error bars: s.e.m.
返回

Fig. 5. Floral and vegetative phenotypes of mnt. (A) Alignment of buds from wild-type and mnt inflorescences, approximately stage 12–16. In each part of the figure the bottom row of buds shows the top row partially dissected. (B) Lengths of floral organs in stage 13 flowers. (C) Nail varnish casts of epidermis from carpel valve (left) and medial sepal (right) of wild-type and mnt stage 13 flowers. In each pair the wild type is on the left. (D) Transverse sections of inflorescence stem between nodes 2 and 3 from the base: (top) wild-type, (bottom) mnt. (E) Largest leaf from a wild-type (left) and mnt mutant (right) rosette. Scale bars: 1 mm in A; 50 μm in C; 200 μm in D, left; 100 μm in D, right. G, gynoecium (excluding stigma); P, petal; Se, medial sepal; St, long stamen.
返回

Fig. 6. Mapping and sequencing MNT. (A) mnt mutations were mapped to BAC MTG10 on chromosome V. Figures below the marker names show the number of recombination events in 1578 chromosomes scored. (B) Scoring of an allelism test between mnt mutants and Salk_108995 TDNA insertion mutants by floral phenotype. (C) Scoring of complementation by floral and seed phenotype. mnt + ARF2=T1 progeny of an mnt mutant transformed with the wild-type ARF2 gene and flanking genomic DNA. This construct restored wild-type floral (left) and seed phenotypes (right) to the mutant. (D) The ARF2 gene with the positions of the mnt/arf2-9 and Salk_108995/arf2-8 mutations marked. (E) Alignment of a fragment of wild-type ARF2 cDNA with the mutated region in mnt/arf2-9 showing a 4-bp deletion. (F) Alignment of the N-terminal portion of the wild-type ARF2 and mutant mnt/arf2-9 proteins, showing an early frameshift and stop codon. (G) The ARF2 protein marked with the position of the stop codon generated by the mnt/arf2-9 lesion. DBD, DNA binding domain; MR, variable middle region; III and IV, domains involved in dimerization with other ARFs or with Aux/IAAs (Liscum and Reed, 2002).
返回
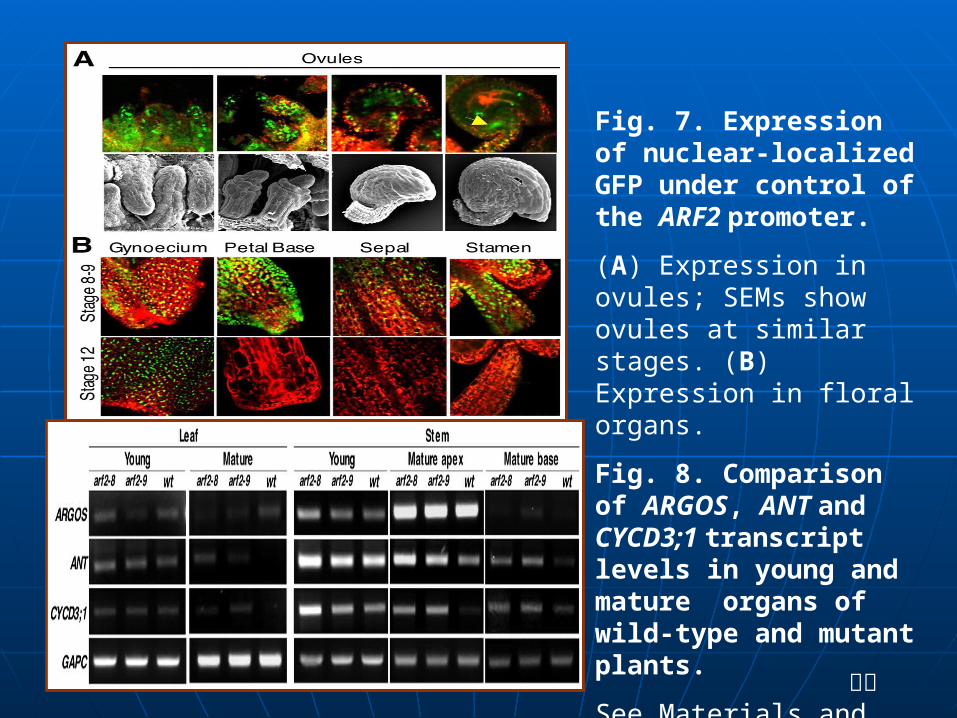
Fig. 7. Expression of nuclear-localized GFP under control of the ARF2 promoter.
(A) Expression in ovules; SEMs show ovules at similar stages. (B) Expression in floral organs.
Fig. 8. Comparison of ARGOS, ANT and CYCD3;1 transcript levels in young and mature organs of wild-type and mutant plants.
See Materials and methods for explanation of developmental staging.
返回

三、讨论三、讨论11 、、 arf2arf2 突变体突变体‘‘ integument-led’integument-led’ 理理
论 论 mnt/arf2-9mnt/arf2-9 的种皮比野生型包含更多的细胞,主要原的种皮比野生型包含更多的细胞,主要原
因是受精前珠被中特大细胞的作用因是受精前珠被中特大细胞的作用 (Fig. 3)(Fig. 3) 。 。 arf2arf2 突变体突变体对种子大小具有母体效应,根本上取决于珠被的膨大 ;即对种子大小具有母体效应,根本上取决于珠被的膨大 ;即使使 arf2arf2 突变体种子充实度保持不变,突变体种子充实度保持不变, arf2arf2 突变体的种子仍突变体的种子仍比野生型重比野生型重 21% (Fig. 4)21% (Fig. 4) 。。 arf2arf2 突变体种子增大是珠被突变体种子增大是珠被诱导的结果。 诱导的结果。
珠被诱导发育的一个理由是突变体种子大的种子腔内可珠被诱导发育的一个理由是突变体种子大的种子腔内可以容纳更多的胚乳生长发育以容纳更多的胚乳生长发育 (Fig. 2)(Fig. 2) 。另一可能大的种子。另一可能大的种子腔提供了大的可接触空间给带有种皮的胚乳,包含了更多腔提供了大的可接触空间给带有种皮的胚乳,包含了更多的营养物质。的营养物质。

2 、 ARF2 抑制细胞分裂 mnt/arf2-9 突变体造成成熟的植株有不同的表现型,有粗杆、大的玫瑰型叶片、花蕾不开 (Figs 1, 5) 。这些性状是与大型细胞的分裂和膨大有关。 ARF2 的表达贯穿植物生长 (Ulmasov et al., 1999b) 、生殖器官、花蕾胚珠的发育过程,这与突变体表现型基因的多效性是一致的。
我们发现 mnt/arf2-9 突变体促进许多组织细胞分裂,而对形态没有太大的影响,而两个 arf2 等位基因延长了茎杆和玫瑰状叶片中 CYCD3 ;1 的表达、延长细胞周期初始的 D-type cyclin 、控制器官生长和组织细胞分裂过程中的转录因子。我们认为 ARF2 对植株地上部分器官组织的细胞分裂有普遍抑制作用。
mnt/arf2-9 突变体中大型细胞分裂是由于植株从开始增殖转变到器官完全分化阶段的延长造成的。我们观察到突变体胚珠的珠被细胞分裂很快,更是长周期, ANT 和 CYCD3 ; 1 基因的表达在幼嫩时是增强的,在成熟的组织中更是持续不变的,这些结果支持了上面的观点。

3 、 ARF2 表达 对 ARF2 主要观点是 ARF2 与生长素调节基因的启动子中的AuxREs 结合,通过刺激或抑制基因转录来调节生长素信号传导。生长素影响 ARF 的功能是通过影响基因表达来实现的,影响 Aux/IAA 蛋白质生成量、短期活性的核蛋白 AuxREs ,都促进或退化植物对生长激素的响应。 Aux/IAA 和 ARFS 反应成为二聚物,在原生质体转染试验中抑制 ARF2 刺激基因表达的能力。 我们发现 arf2 突变体叶片和茎杆中 ANT 和 CYCD3 ; 1 基因表达被延长 (Fig. 8) ,作用机理已被发现。 ANT 和 CYCD3 ; 1 的表达在ARGOS 过表达的植物的成熟叶片中是相同的 (Hu et al., 2003) 。 ARGOS 被生长素所诱导,它的过量表达导致地上部分器官组织的膨大,根本原因是细胞分裂增多 (Hu et al., 2003) 。未发现arf2 突变体影响幼嫩的或成熟组织中 ARGOS 的表达,说明 ARF2 基因不会通过 ARGOS 的表达来调控 ANT 或 CYCD3 ; 1 的表达 (Fig. 8) 。
