第 六 章 分子生物学基本研究法 (下)基因功能研究技术
DESCRIPTION
第 六 章 分子生物学基本研究法 (下)基因功能研究技术. 随着越来越多的基因组序列相继被测定,人类对生物本质的认识已经发生了重大变化。 但是,海量序列信息也向我们提出了新的挑战: 如何通过生物化学、分子生物学等方法研究基因的功能,从而进一步了解 生物体内各种生理过程 ,了解生物体 生长发育的调节机制 ,了解 疾病的发生 、 发展规律 ,给出 控制、减缓甚至完全消除人类遗传疾病 ?. 转录组测序技术 原位杂交技术 基因芯片技术 为研究单个或多个基因在生物体某些特定发育阶段或在不同环境条件下的表达模式提供了强有力的手段。. - PowerPoint PPT PresentationTRANSCRIPT

第 六 章 分子生物学基本研究法
(下)基因功能研究技术

随着越来越多的基因组序列相继被测定,人类对生物本质的认识已经发生了重大变化。
但是,海量序列信息也向我们提出了新的挑战:
如何通过生物化学、分子生物学等方法研究基因的功能,从而进一步了解生物体内各种生理过程,了解生物体生长发育的调节机制,了解疾病的发生、发展规律,给出控制、减缓甚至完全消除人类遗传疾病?

转录组测序技术
原位杂交技术
基因芯片技术
为研究单个或多个基因在生物体某些特定发育阶段或在不同环境条件下的表达模式提供了强有力的手段。

基因定点突变( site-directed mutagenesis )技术
基因敲除( gene knock-out )技术
RNAi 技术
可以全部或部分抑制基因的表达,通过观察靶基因缺失后生物体的表型变化研究基因功能。

酵母单杂交
酵母双杂交技术
各类相互作用技术
研究蛋白质间相互作用、蛋白质 -DNA 相互作用。
科研人员还可以在活细胞内研究蛋白质之间的相互作用,为认识信号转导通路、蛋白质翻译后修饰加工等提供了丰富的技术支持。

6. 1 基因表达研究技术6. 1. 1 转录组学研究
转录组 (transcriptome) ,广义上指在某一特定生理
条件或环境下,一个细胞、组织或者生物体中所有 RN
A 的总和,包括信使 RNA ( mRNA )、核糖体 RNA
( rRNA )、转运 RNA ( tRNA )及非编码 RNA (n
on-coding RNA 或 siRNA) 。现常指细胞中转录出来
的所有 mRNA 的总和。

基因组-转录组-蛋白质组( genome - transcripto
me - proteome )是中心法则在组学框架下的主要表现形式。
通过特定生理条件下细胞内的 mRNA 丰度来描述基因表达水平并外推到最终蛋白质产物的丰度是目前基因表达研究的基本思路。

传统的转录组研究方法主要包括:表达序列标签( exp
ressed sequence tag , EST )测序技术,基因表达
系列分析技术( serial analysis of gene expression ,
SAGE )和基因芯片技术。
EST 测序数据是目前数量最多,涉及物种最广的转录
组数据,但测序读长较短(每个转录本测定 400bp -
500bp ),测序通量小,测序成本较高。

SAGE 测序法:将不同转录本 3’ 端第一个 CATG 位
点下游 14 bp 长的短标签序列来标识相应的转录本。
该法所采用的过短的序列标签使得序列唯一性降低,
即使改进过的 LongSAGE 用 21 bp 标签测序,仍然
有约一半的标签无法被准确注释到基因组上。

高通量测序技术 (high-throughput sequencing) ,又
名二代测序( second-generation sequencing )或
深度测序 (deep sequencing) ,可以一次性测序几十
万甚至几百万条,是传统测序技术的革命。
利用高通量测序技术对转录组进行测序分析被称为 RN
A-Seq ,对测序得到的大量原始读长( reads )进行
过滤、组装以及后续的生物信息学分析。

表 6-1 454 和 Solexa 高通量测序平台比较
454 Solexa
读取长度( bp ) 约 700 50- 150
单次测序数据量 700 Mb 600 Gb
测序周期 23小时 7- 14天
测序成本 较高 低
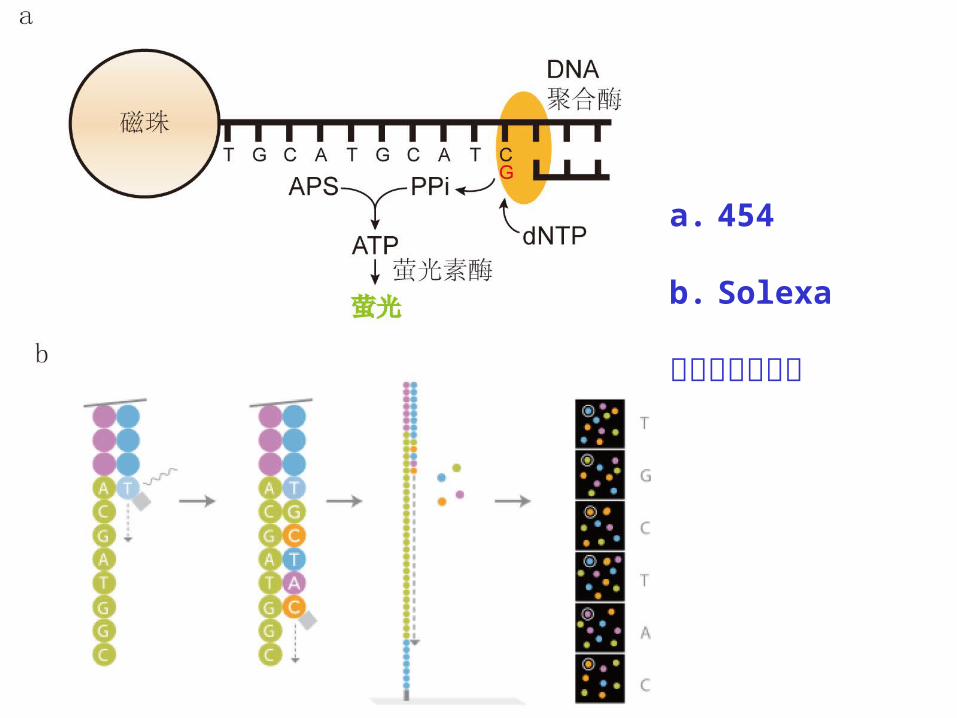
a. 454
b. Solexa
测序原理示意图

454 系统采用焦磷酸测序 (pyro-sequencing) :按 T 、
A 、 C 、 G 顺序加入单个 dNTP 与模板配对,释放一
分子焦磷酸 (PPi) ,在 ATP 硫酸化酶作用下 PPi 和腺
苷酰硫酸( adenosine-5’-phosphosulfate, APS )
结合形成 ATP ,在萤光素酶的催化下产生可见光。
454 则由于缺少终止反应的元件,相同碱基的连续掺
入常会带来“插入-缺失”类型的测序错误。

Solexa 测序系统采用萤光标记 dNTP ,其 3’ 羟基末
端可被化学切割,每个循环反应只允许掺入一个碱基,
由激光扫描反应板表面,读出这一轮反应新加的萤光
信号,从而判定碱基种类。之后,经过化学切割恢复
3’ 端粘性,进行下一轮聚合反应。
随着 Solexa 测序反应的进行,已有萤光信号会使新的
萤光难以准确分辨,因此该方法的测序读长较短,测
序错误主要是碱基替换。

图 6 - 2 DNA 序列的从头组装de novo assembly
在实际组装过程中,图中红色标示区域覆盖度过低,且读长缺乏相对位置信息的区域,其可信度较低,应当剔除,只保留两侧序列。

利用大量读长之间重叠覆盖和成对读长( pair-end re
ads )的相对位置关系,组装得到尽可能完整的转录
本。
以单位长度转录本上覆盖的读长数目( reads per kil
o-base gene per million bases , RPKM )作为衡
量基因表达水平的标准。

以棉花转录组学数据为例,分析不同组织或纤维不同
发育时期基因表达情况(表 6-2 )。 Solexa 测序得
到 26.86Gb 数据,经过从头组装总共获得了 42,773
条非重复序列,平均长度 1,054 碱基。每个不同组织
中分别有 23,265至 26,427 个独立转录本。

表 6-2 陆地棉 6 个组织的 RNA-Seq 数据分析
组织样品 读长数 碱基数Q201
(%) N50
独立基因总数 独立基因
长度 (nt)
开花后 0天胚珠 47,907,298 3593047350 98.22 827 26427 778
开花后 5天胚珠 53,022,210 3976665750 97.86 842 23520 786
花 48,049,786 3603733950 97.99 823 23265 775
叶 54,191,238 4064342850 97.51 820 25280 776
根 79,438,254 7149442860 91.68 786 23905 753
茎 49,713,024 4474172160 91.46 782 24088 746
总计 332321810 26861404920 1306 42773 1054
1Q20 ,测序准确率达到 99%。

转录组数据不但能用来分析不同组织中独立转录本数
量,还被用于分析特定转录本在某个组织中的表达强
度(表 6-3)。
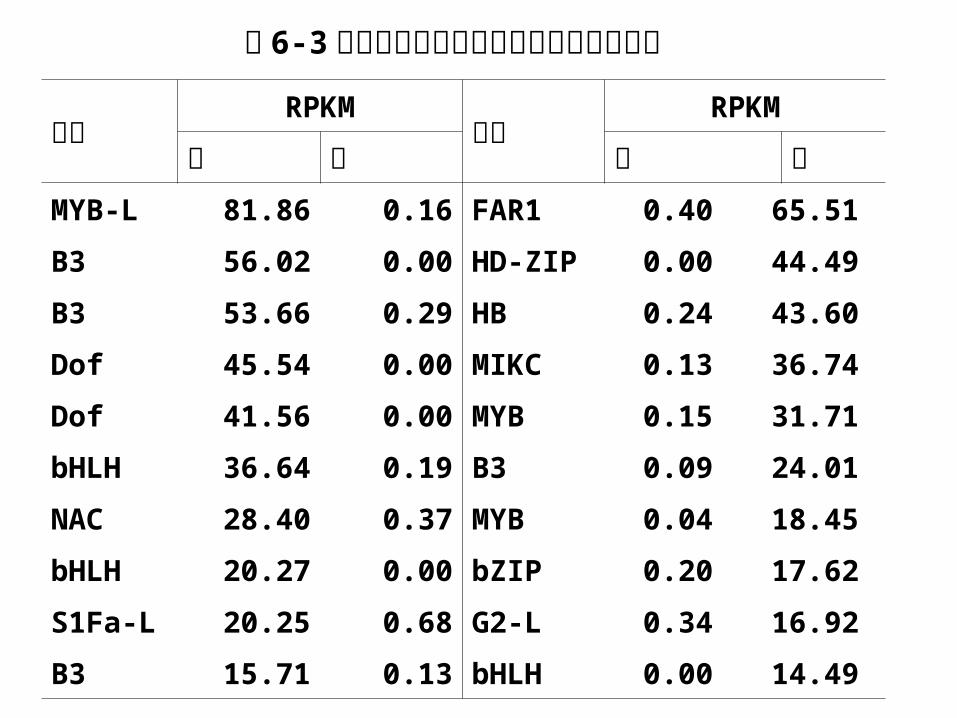
表 6-3棉花组织特异性转录因子表达强度分析
基因RPKM
基因RPKM
根 茎 根 茎MYB-L 81.86 0.16 FAR1 0.40 65.51
B3 56.02 0.00 HD-ZIP 0.00 44.49
B3 53.66 0.29 HB 0.24 43.60
Dof 45.54 0.00 MIKC 0.13 36.74
Dof 41.56 0.00 MYB 0.15 31.71
bHLH 36.64 0.19 B3 0.09 24.01
NAC 28.40 0.37 MYB 0.04 18.45
bHLH 20.27 0.00 bZIP 0.20 17.62
S1Fa-L 20.25 0.68 G2-L 0.34 16.92
B3 15.71 0.13 bHLH 0.00 14.49

6. 1. 2 RNA 的选择性剪接
RNA 的选择性剪接是指用不同的剪接方式从一个mRNA 前体产生不同的 mRNA剪接异构体的过程。包括: 平衡剪接、 5’选择性剪接、 3’选择性剪接、外显子遗漏型剪接及相互排斥性剪接。
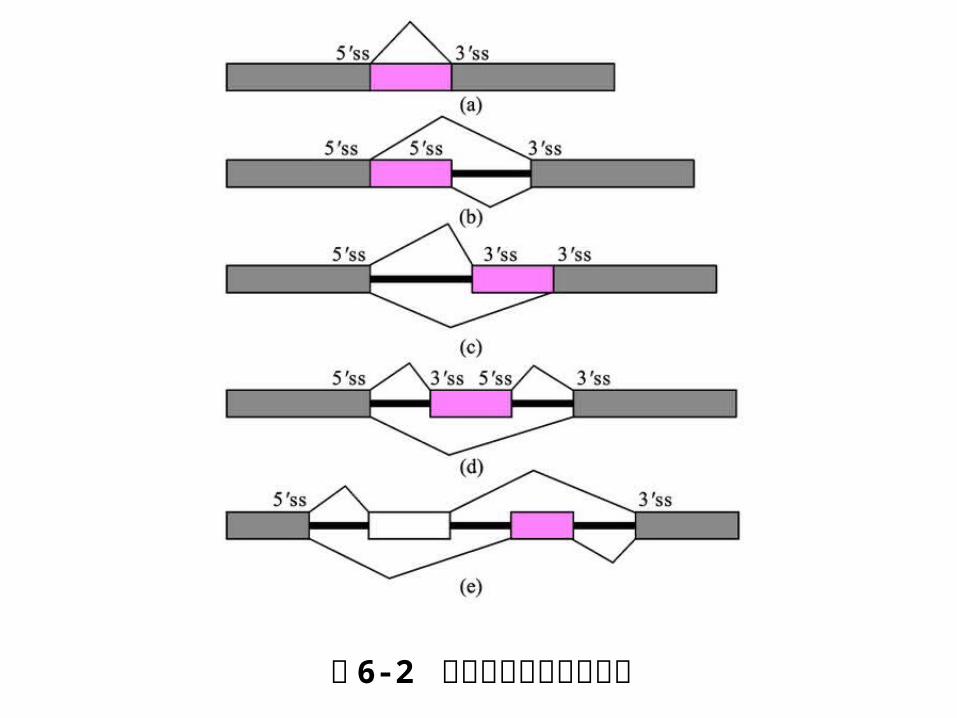
图 6-2 选择性剪切的不同类型

图 6-3 RT-PCR检测 5个拟南芥转录调控因子基因的选择性剪切

G. raimondii
基因组中,四种主要的选择性剪切事件为:

IR is the most frequent AS event in G. raimondii
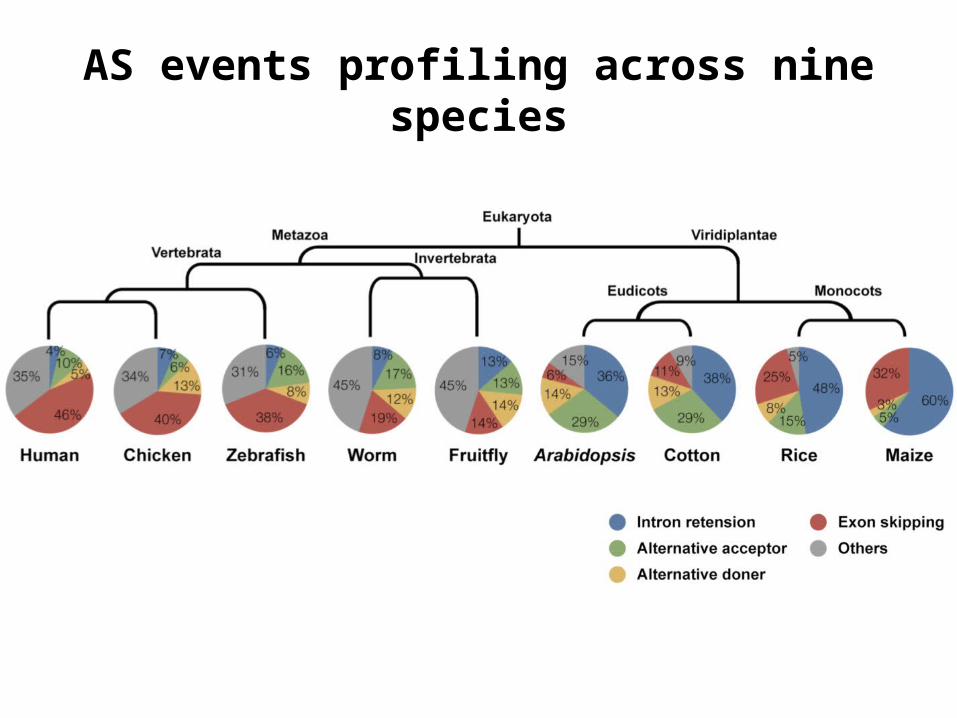
AS events profiling across nine species

图 6-4 果蝇( Drosophila melanogaster)的 Dscam基因可以通过可变剪接产生 38000多种可能的mRNA异
构体

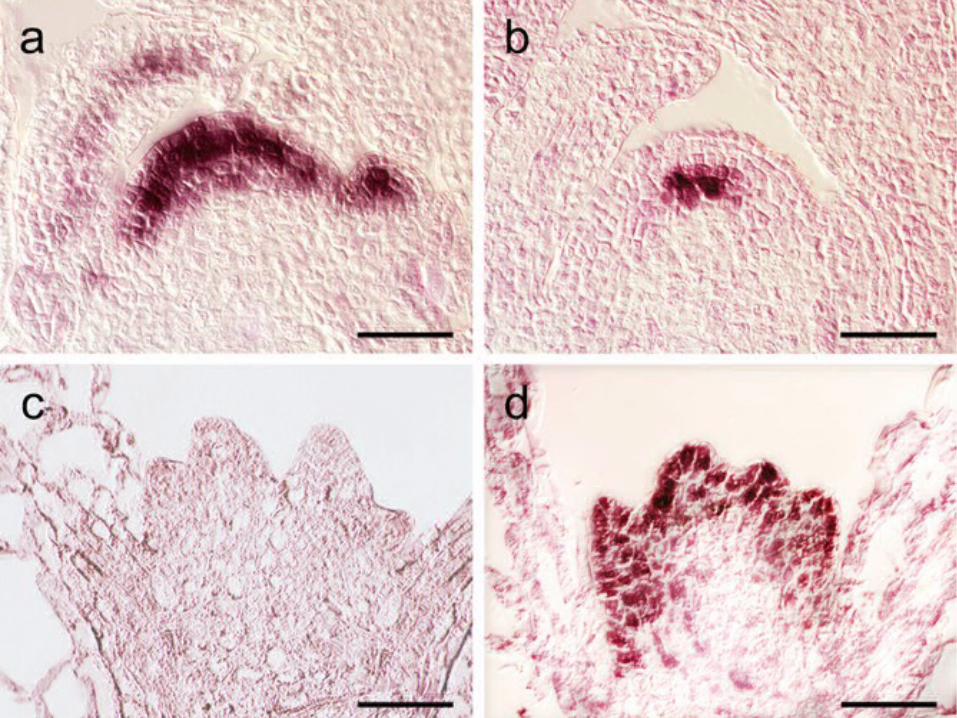
6. 1. 3 原位杂交技术原位杂交 ( In Situ Hybridization , ISH) 是用标记的核酸探针,经放射自显影或非放射检测体系,在组织、细胞、间期核及染色体上对核酸进行定位和相对定量研究的一种手段,分为 RNA 和染色体原位杂交两大类。

RNA 原位杂交用放射性或非放射性(如地高辛、生物素等)标记的特异性探针与被固定的组织切片反应,若细胞中存在与探针互补的 mRNA 分子,两者杂交产生双链 RNA ,可通过放射性标记或经酶促免疫显色,对该基因的表达产物做出定性定量分析。
用 mRNA 的互补链为探针进行杂交。
负对照, mRNA 的同义链 , 无杂交信号。

荧光原位杂交 (fluorescence in situ hybridization , FISH).
对寡核苷酸探针做特殊的修饰和标记,用原位杂交法与靶染色体或 DNA 上特定的序列结合,再通过与荧光素分子相耦联的单克隆抗体来确定该 DNA
序列在染色体上的位置。

人肌肉糖原磷酸酶基因在 11 号染色体上的 FISH 原位杂交结果。
6. 1. 4 基因定点突变 (site-directed mutagenesis).
通过改变基因特定位点核苷酸序列来改变所编码的氨基酸序列,用于研究某个(些)氨基酸残基对蛋白质的结构、催化活性以及结合配体能力的影响,也可用于改造DNA 调控元件特征序列、修饰表达载体、引入新的酶切位点等。

图 6-6 寡核苷酸介导的 DNA 突变技术

目前,主要采用两种 PCR 方法,重叠延伸技术和大引物诱变法,在基因序列中进行定点突变。


图 6-8 大引物诱变法示意图

6. 2 基因敲除技术6. 2. 1 基本原理
经典遗传学( Forward genetics )是从一个突变体的表型出发,研究其基因型,进而找出该基因的编码序列。
现代遗传学( Reverse genetics ,反向遗传学)首先从基因序列出发,推测其表现型,进而推导出该基因的功能。

基因敲除( gene knock-out )又称基因打靶,通过外源 DNA与染色体 DNA 之间的同源重组,进行精确的定点修饰和基因改造,具有专一性强、染色体 D
NA 可与目的片段共同稳定遗传等特点。
基因敲除分为完全基因敲除和条件型基因敲除(又称不完全基因敲除)两种。
完全基因敲除是指通过同源重组法完全消除细胞或者动植物个体中的靶基因活性,条件型基因敲除是指通过定位重组系统实现特定时间和空间的基因敲除。

图 6-11 用取代型
( a )或插入型( b )
载体进行完全基因敲除
实验 .

图 6-12 正负筛选法( PNS 法)筛选已发生同源重组的细胞

正向选择基因 neo 通常被插入载体靶 DNA 功能最关键的外显子中,通过同源重组法置换靶基因的功能区。
负向选择基因 HSV-tk则被置于目的片段外侧,含有该基因的重组细胞不能在选择培养基上生长。如果细胞中发生了随机重组,负向选择基因就可能被整合到基因组中,导致细胞死亡。

噬菌体的 Cre/Loxp系统、 Gin/Gix系统、酵母细胞的 FLP/FRT系统和 R/RS系统是现阶段常用的四种条件型定位重组系统,尤以 Cre/Loxp系统应用最为广泛。

图 6-13 条件型基因敲除策略

6. 2. 2 高等动物基因敲除技术真核生物基因敲除的技术路线主要包括构建重组基因载体,用电穿孔、显微注射等方法把重组 DNA 导入胚胎干细胞纯系中,使外源 DNA与胚胎干细胞基因组中相应部分发生同源重组,将重组载体中的 DNA
序列整合到内源基因组中并得以表达。
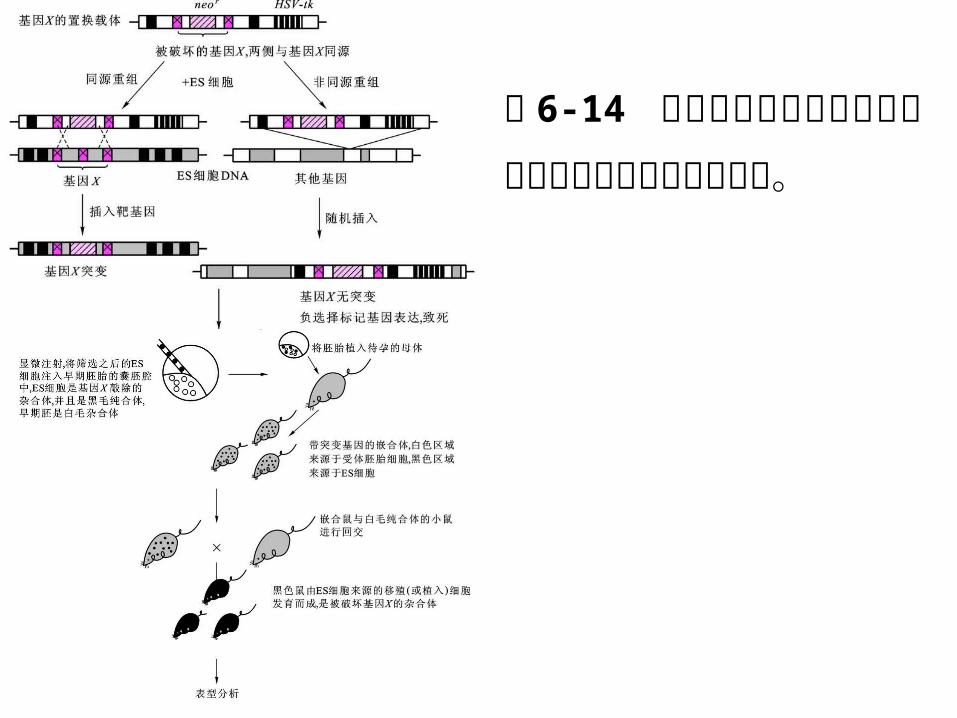
图 6-14 模式动物小鼠中完全基因敲除的主要技术策略与应用。

显微注射命中率较高,技术难度相对大些。电穿孔法命中率比显微注射低,操作使用方便。
胚胎干细胞( ES 细胞)分离和体外培养的成功奠定了哺乳动物基因敲除的技术基础。
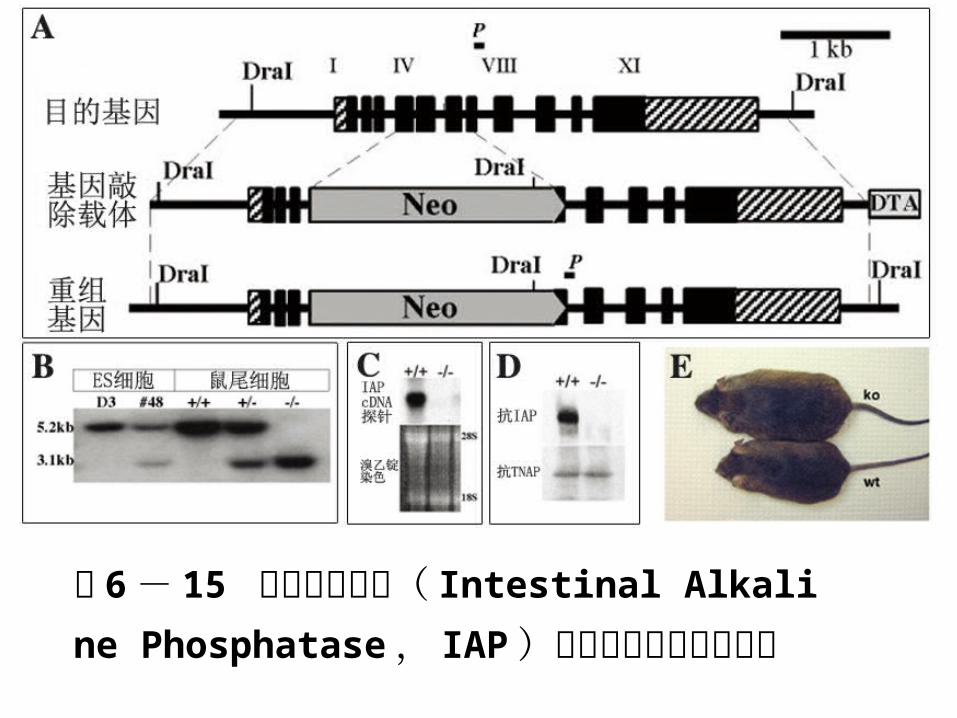
图 6 - 15 肠碱性磷酸酶( Intestinal Alkalin
e Phosphatase , IAP )基因敲除加速小鼠肥胖
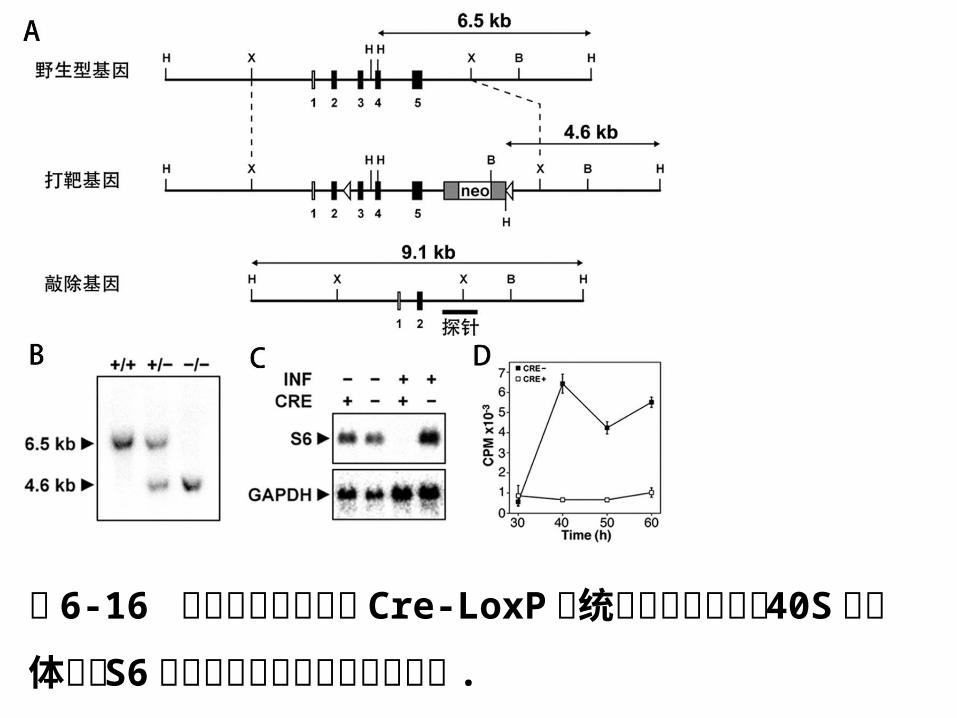
图 6-16 肝组织特异性表达 Cre-LoxP 系统条件型敲除小鼠 40S核糖体蛋白 S6 基因导致肝细胞分裂增殖受阻 .

A.构建带有 S6 基因的 LoxP打靶载体的 ES 细胞,
经过杂交筛选,获得纯合小鼠,与带有肝组织特异
性、受 INF-α诱导的 Mx-Cre 转基因小鼠杂交,删
除 neo 和外显子 3-5 ,得到肝组织特异性敲除 S6
基因的小鼠。
B. Southern杂交表明,纯合体小鼠内有 DNA 片段
缺失。

C. Northern杂交表明, INF-α诱导后 Cre 小鼠体
内 S6 基因不表达。
D. [ 3H]胸腺嘧啶能被 S6 基因未敲除小鼠吸收,
却不能掺入 S6 基因敲除小鼠肝组织,表明细胞不
能进入 S期,不能正常分裂增殖。

外显子 1 外显子 2
内含子 1
转录
剪切
翻译
GUS, 无启动子
外显子 1 外显子 2
基因陷阱载体
随机整合
内含子 1
外显子 1 外显子 2
随机插入内含子或外显子中
GUS
转录
剪切
翻译
(A) 基因捕获前
(B) 基因捕获后
图 6-17 基因捕获法原理A 、未插入基因捕获载体前,需要被敲除的靶基因转录翻译出活性蛋白质 ,未知; B 、插入基因捕获载体后,靶位点基因转录翻译出带有GUS 蛋白区段的融合蛋白,可用组化方法检测。

6. 2. 3 植物基因敲除技术
T-DNA 插入失活技术是目前在植物中使用最为广泛的基因敲除手段。
利用根癌农杆菌 T-DNA介导转化,将带有报告基因的 DNA 序列整合到基因组 DNA 上,如果这段 D
NA 插入到目的基因内部或附近,就会影响该基因的表达,从而使该基因“失活”。
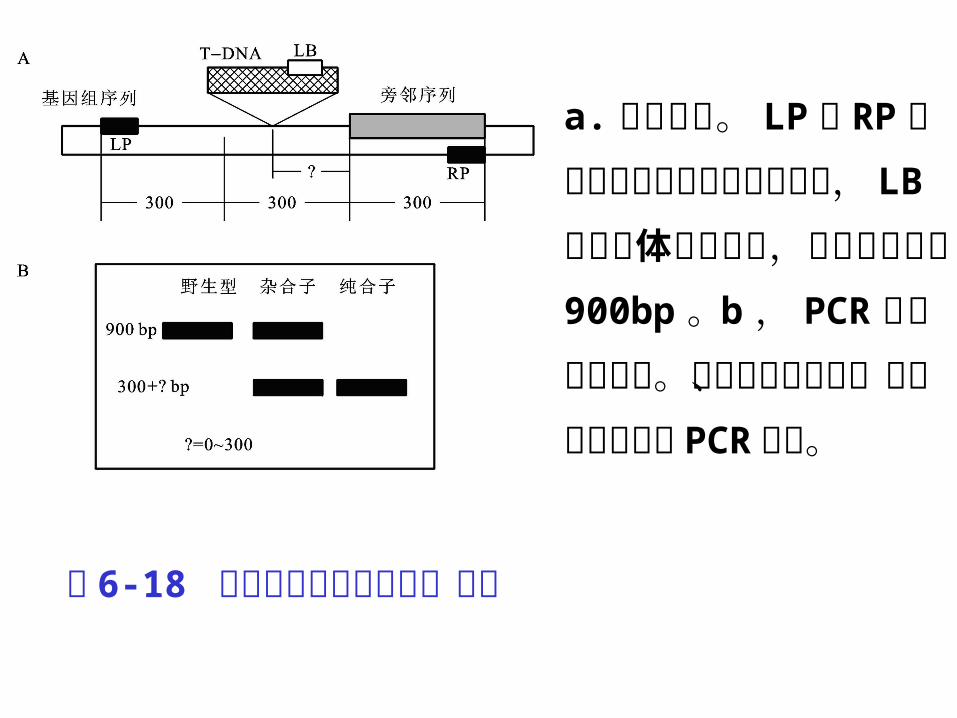
a.引物设计。 LP 和 R
P 分别代表 插 入基 因两端的引物, LB 是指载体上的引物,目的基因片段 900bp 。 b ,PCR 产物电泳结果。分别代表野生型、杂合子和纯合子 PCR 条带。
图 6-18 植物基因敲除及突变体筛选

Identifiation of bard1-3 homozygous lines

6. 2. 4 植物基因的定点整合技术

TALEN 介导的基因组定点修饰技术 • TALEN: transcription activator-like effector nucleases /类转录激活因子效应物核酸酶
TALE 重复序列
RVD12 13
FokIFokI C TALEN 结构图
重复单元
RVD 和对应碱基类型HD: C NG: T NI: A NN: G/A
• RVD: Repeat variable di-residue / 重复可变双残基 • NLS: Nuclear localization signal /核定位信号
• AD: Activation domain / 转录激活结构域
非特异性核酸酶
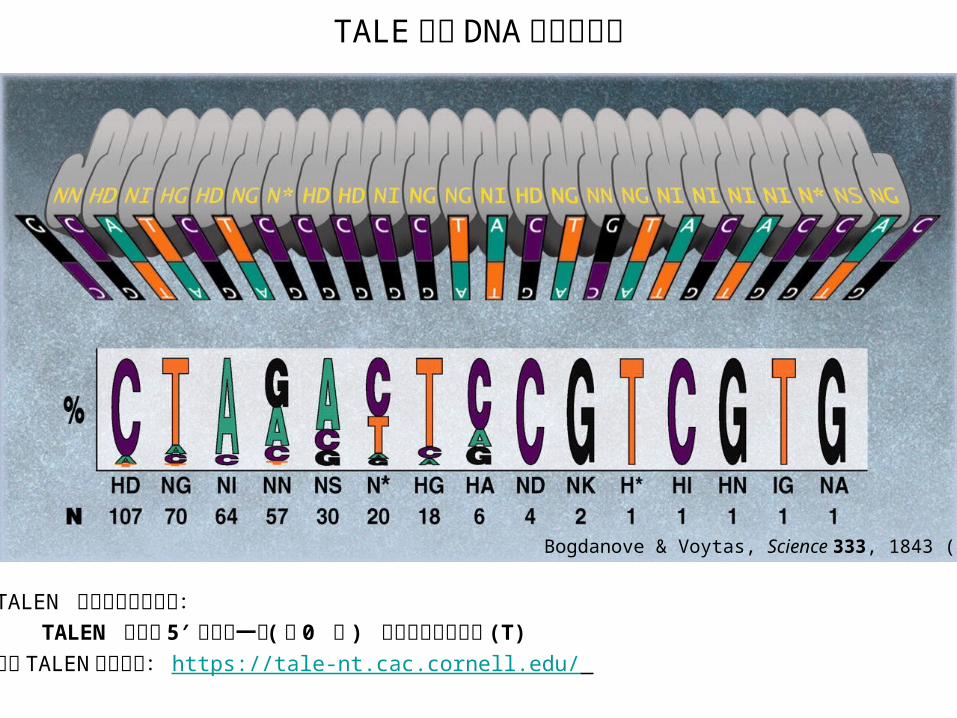
TALE 识别 DNA 序列示意图
Bogdanove & Voytas, Science 333, 1843 (2011)
• TALEN 靶位点的挑选原则: TALEN 靶位点 5′ 端的前一位 (第 0 位 ) 碱基应为胸腺嘧啶 (T)• 设计 TALEN 靶点网站: https://tale-nt.cac.cornell.edu/
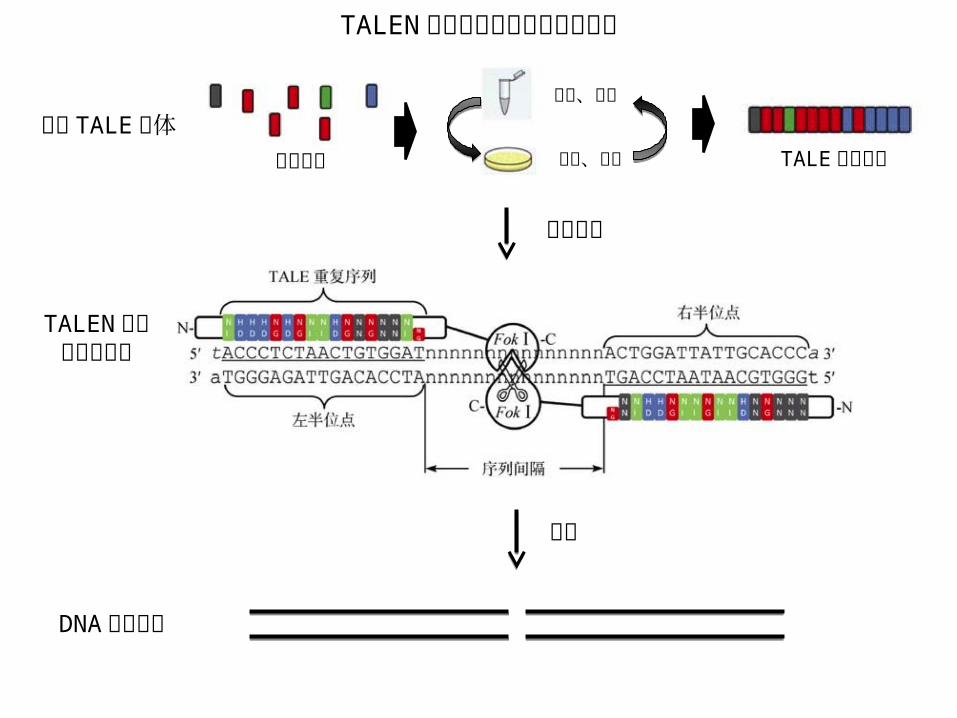
TALEN 介导的基因组定点修饰流程
TALEN 识别靶基因位点
DNA 双链断裂
构建 TALE 载体重复单元
酶切、连接
转化、克隆 TALE 重复序列
导入细胞
酶切

非同源末端连接( NHEJ )(简单高效,错误率高)
同源重组( HR )(效率低,保真度高)
DNA 双链断裂
细胞固有的DNA 修复机制
DNA 修复
>70% 产生基因突变
模版
酶切检测、筛选
改造靶基因序列
Knock out Knock in
TALEN 介导的基因组定点修饰流程

6. 3 蛋白质及 DNA 相互作用技术6. 3. 1 酵母单杂交系统( Yeast one-hybrid syste
m )
是上世纪 90年代中发展起来的研究 DNA- 蛋白质
之间相互作用的新技术,在酵母细胞内研究真核生
物中 DNA- 蛋白质之间的相互作用,并通过筛选 D
NA文库直接获得靶序列相互作用蛋白的编码基因。

真核生物转录调控因子具有组件式结构( modular )
特征,这些蛋白往往由两个或两个以上相互独立的结
构域,其中 DNA 结合结构域( binding domain , B
D )和转录激活结构域( activation domain , A
D )是转录激活因子发挥功能所必须的。
BD 能与特定基因启动区结合,但不能激活基因转录,
由不同转录调控因子的 BD 和 AD所形成的杂合蛋白
却能行使激活转录的功能。

图 6-19 酵母单杂交的基本原理示意图

将顺式作用元件与最基本启动子( minimal promot
er , Pmin )相连并将报告基因连到 Pmin 下游,将
待测转录因子 cDNA 与酵母转录激活结构域( trans
cription-activating domain, AD )融合表达载体导
入酵母细胞,该基因产物如果能够与顺式作用元件相
结合,就激活 Pmin启动子,报告基因表达。

图 6-20 从拟南芥 cDNA 文库中筛选与 顺 式元件 DRE 结合的 转 录 因 子 示意图。

将不同基因或基因片段分别克隆到 pYF503 中 (GAL4
AD 为 GAL4 激活结构域 , empty vector 表示无基因克隆到 pYF503 中 ), 转入酵母中表达 N 端带有 G
AL4 DNA 结合结构域的融合蛋白,观察报告基因表达与否。
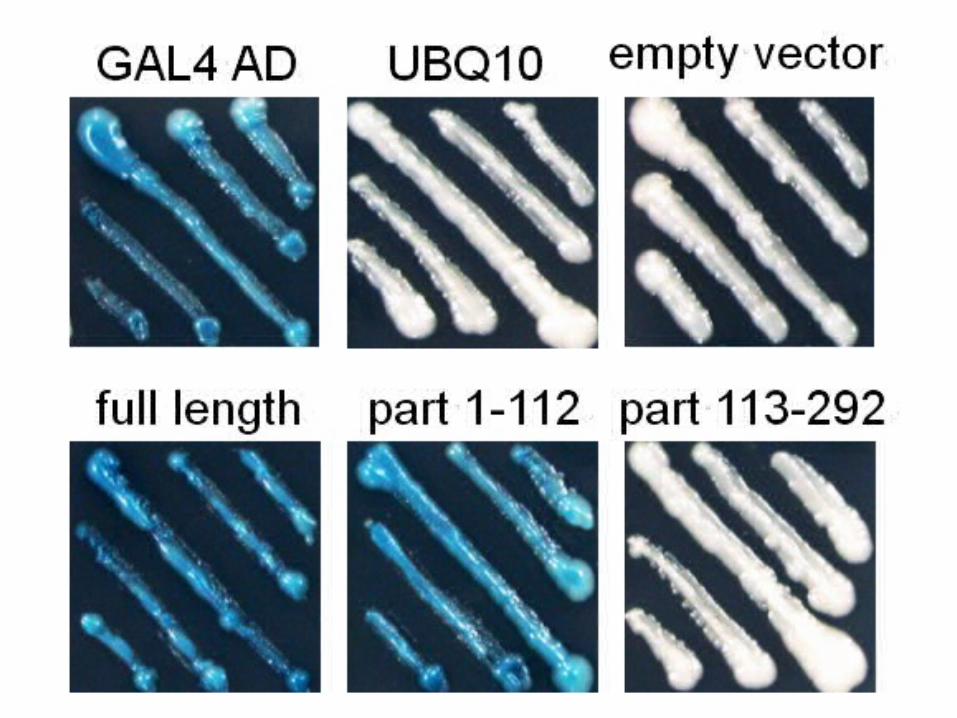

6. 3. 2 酵母双杂交系统( Yeast two-hybrid syste
m )
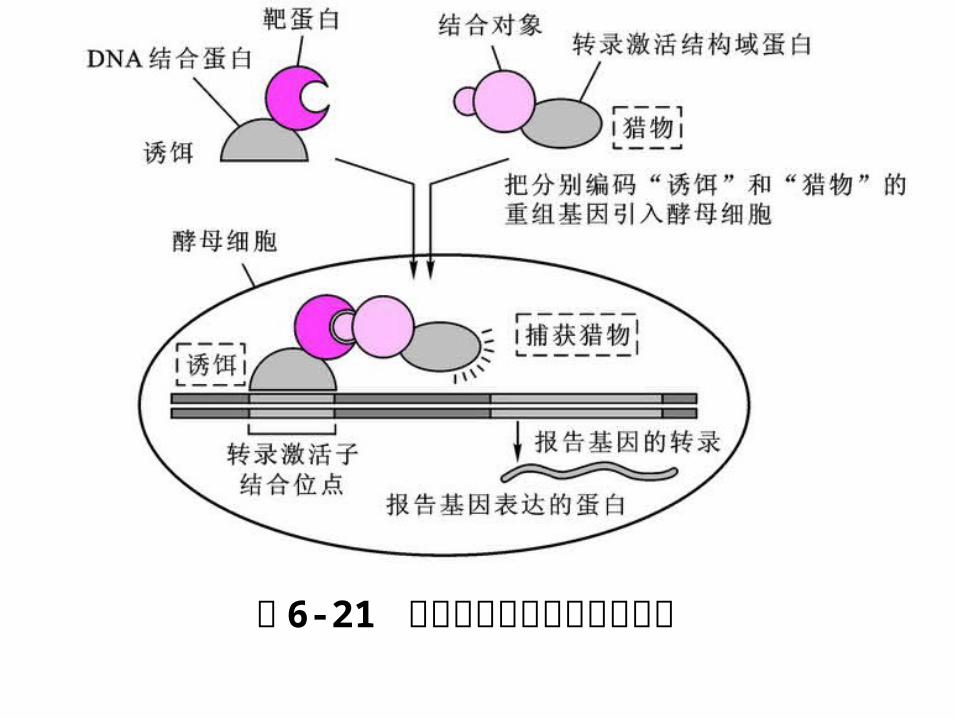
图 6-21 酵母双杂交技术原理示意图

6. 3. 3 体外蛋白质相互作用技术
1 、等离子表面共振技术
诱饵蛋白结合于固定在纳米厚度的金属膜上的葡聚糖
表面,当有蛋白质混合物经过时,如果有蛋白质同
“诱饵”蛋白发生相互作用,那么两者的结合使金属
膜表面的折射率上升,从而导致共振角度的改变。而
共振角度的改变与该处的蛋白质浓度成线性关系,由
此可以检测蛋白质之间的相互作用(图 6-22 )。

信号输出光源
棱镜
光感受器
自由的待分析物
被捕获的待分析物
液流通路流向
图 6-22 离子表面共振技术示意图
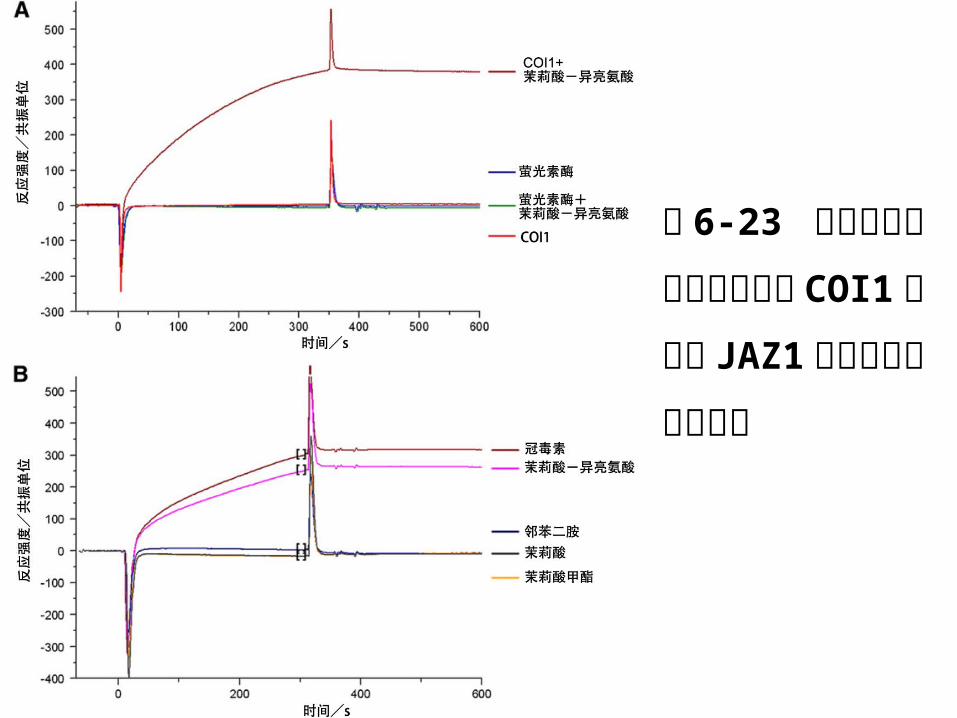
图 6-23 等离子表面共振技术研究 COI1 蛋白与JAZ1 蛋白之间的相互作用

研究人员首先在葡聚糖芯片表面固定 1000共振单位
的茉莉酸( JA )信号通路负调控因子 JAZ1 蛋白,
当体系中同时有茉莉酸-异亮氨酸( JA-Ile )及 CO
I 蛋白存在时,可以检测到最高达 380共振单位的 S
PR 反应信号。加入一种在结构和功能上与茉莉酸甲
脂( MeJA )非常相似的名为冠毒素 (COR) 的细菌
毒素,也能使 COI1 和 JAZ1发生相互作用。

2 、免疫共沉淀技术 (Co-IP)
将靶蛋白的抗体连接到固体基质上,加入可能与靶蛋
白发生相互作用的待筛选蛋白,用低离心力沉淀或微
膜过滤法在固体基质和抗体的共同作用下将蛋白复合
物沉淀到试管的底部或微膜上。

图 6-24 免疫共
沉淀示意图

载体构建
体外转录、翻译及产物混合温育
加 Myc或 HA抗体
过柱及 SDS - PAGE电泳分离
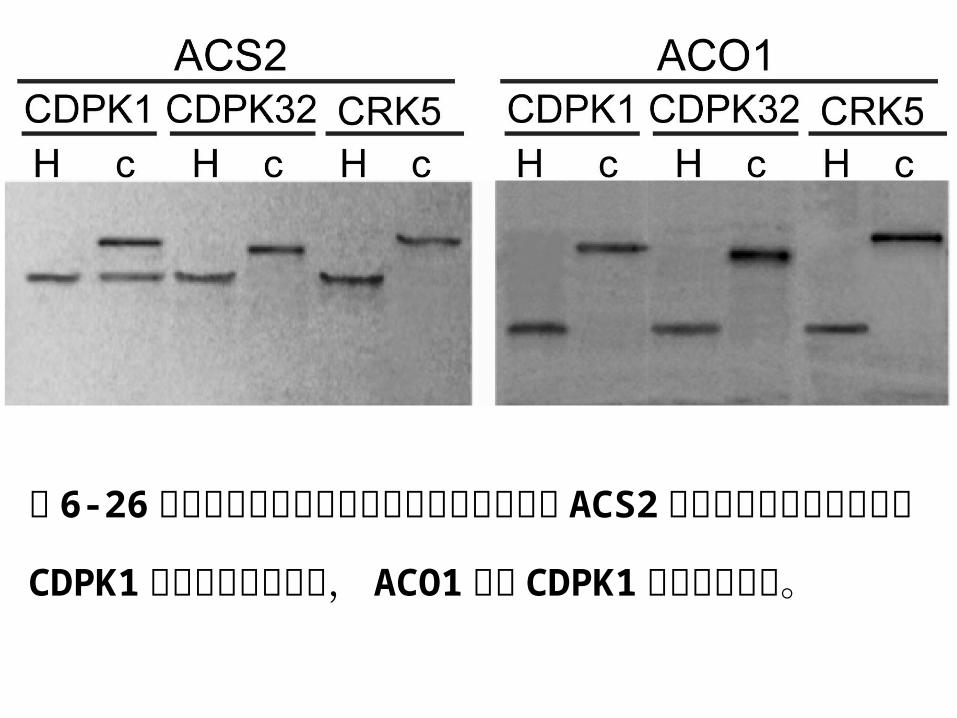
图 6-26免疫共沉淀实验表明棉花乙烯合成酶基因 ACS
2 与钙离子依赖性蛋白激酶 CDPK1 之间存在相互作用,
ACO1不与 CDPK1发生相互作用。

3 、 GST融合蛋白沉降技术
利用 GST 对谷胱甘肽偶联的琼脂糖球珠的亲和性,
从混合蛋白质样品中纯化得到相互作用蛋白。
GST沉降试验通常有两种应用:确定探针蛋白与未知
蛋白间的相互作用,确证探针蛋白与某个已知蛋白之
间的相互作用。
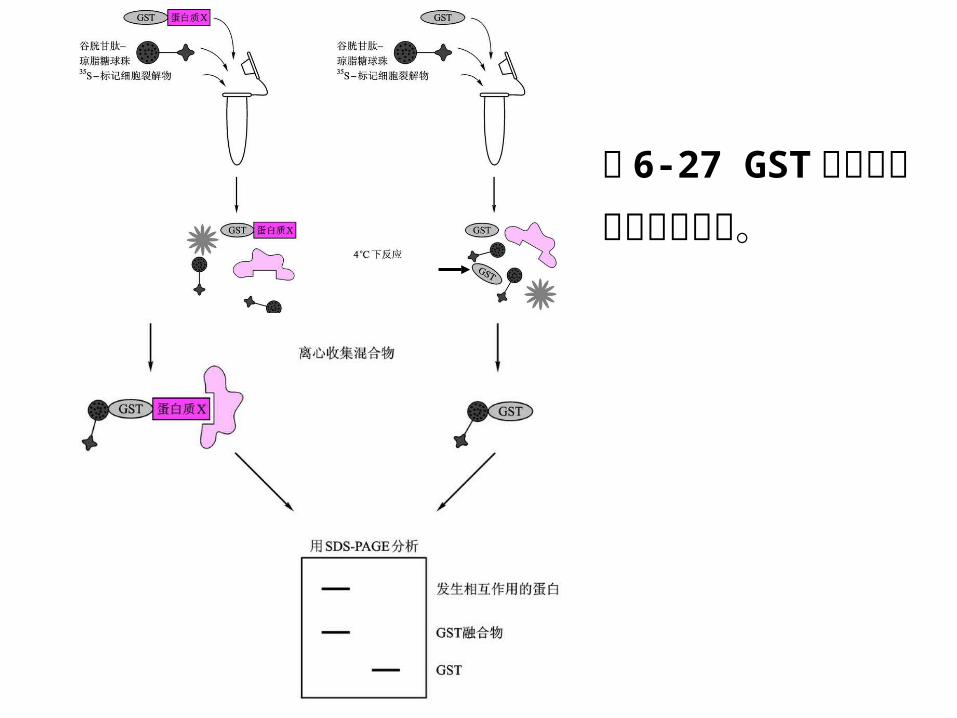
图 6-27 GST融合蛋白沉降技术流程。

图 6-28 体外融合蛋白沉
降研究表明,光敏素 B (p
hy B) 能与 PIF3发生强烈
的相互作用,缺失 N-37
位氨基酸的 phy B突变体
及光敏素 A (phy A) 只能
与 PIF3发生较弱的相互
作用。

6. 3. 4 染色质免疫共沉淀( ChIP: Chromatin Immu
no Precipitation )
在活细胞状态下固定蛋白质- DNA 复合物,并通过超声或酶处理将其随机切断为一定长度的染色质小片段,然后通过抗原抗体的特异性识别反应,沉淀该复合体,富集与目的蛋白相结合的 DNA 片段,通过对目的片段的纯化与检测,获得该蛋白质与 DNA 相互作用的信息,包括具体的 DNA 序列特征,位置,结合时间、亲和程度,对基因表达的影响力等。



• ChIP不仅可以检测体内转录因子与 DNA 的动态作用,还可以用来研究组蛋白的各种共价修饰与基因表达的关系。
• 定性或定量检测体内转录因子与 DNA 的动态作用。

6. 3. 5 RNAi ( RNA interference, RNA 干涉)技术及其应用
RNAi 技术利用双链小 RNA 高效、特异性降解细胞内同源mRNA 从而阻断靶基因表达,使细胞出现靶基因缺失的表型。 RNAi 的命名来源于安德鲁 · 法尔。

RNAi 作用机制示意图

双链 RNA 是 RNAi 的触发物,引发与之互补的单链 R
NA ( ssRNA, single-stranded RNA )的降解。
经过 Dicer (一种具有 RNAase III活性的核酸酶)的
加工,细胞中较长的双链 RNA ( 30 个核苷酸以上)
首先被降解形成 21~ 25 个核苷酸的小分子干扰核糖
核酸( siRNA , short interfering RNA ),并有效
地定位目标 mRNA 。

由 siRNA 中的反义链参与指导合成被称为 RNA诱
导的沉默复合体( RISC )的核蛋白体,再由 RIS
C介导切割目的 mRNA 分子中与 siRNA 反义链互
补的区域,从而实现干扰靶基因表达的功能。

6. 4 基因芯片及数据分析
基因芯片( DNA chip ),又称 DNA微阵列( DNA
microarray )技术是能同时监测大量靶基因表达的实
验手段,从而迅速准确地在基因组水平上阐述不同生
物组织或细胞中各种转录本的变化规律。

6-25 基因芯片技术流程图

基因芯片可用于进行基因诊断。先建立正常人特定组
织、器官的基因芯片,给出标准杂交信号图,用可疑
病人的 cDNA做探针与之杂交,确定哪些基因的表达
受抑制或激活。

通过两次平行杂交反应,将每个基因的表达水平做成对应散点图,只要绝大多数基因的杂交结果都在 45 度斜线附近,就说明实验结果是可靠的 。


a b
a. Upland cotton (Gossypium hirsutum Xuzhou142)
b. fuzzless-lintless mutant (fl) (Zhang and Pan, 1991).
+3w
t/-3wt
+5w
t/+3w
t+
10wt/+
3wt
+15w
t/+3w
t+
20wt/+
3wt
+10fl/+
3wt
+3fl/-3w
t
0wt/-3w
t
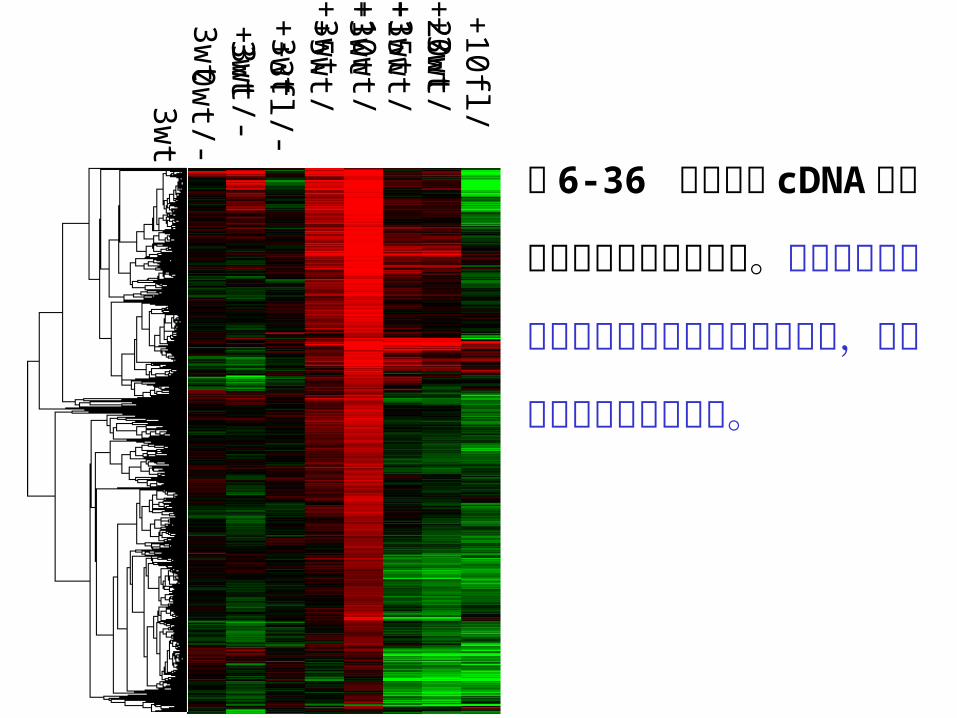
图 6-36 棉花胚珠 cDN
A 芯片差异表达基因聚
类分析。红色指示在纤
维细胞中特异性表达上
调的基因,绿色指示表
达下调的基因。

Table 1. Analysis of R-values
Correlation coefficients Microarray samples
Replicates (1,2)
Replicates (2,3)
Replicates (1,3)
+3wt/0wt 0.924 0.928 0.938
+3wt/+3wt 0.982 0.989 0.933
+3wt/+3fl 0.925 0.918 0.931
+5wt/+3wt 0.933 0.926 0.917
+10wt/+3wt 0.912 0.932 0.924
+15wt/+3wt 0.966 0.964 0.951
+20wt/+3wt 0.965 0.945 0.966
+10wt/+10fl 0.963 0.951 0.952

6. 5 利用酵母鉴定靶基因功能6. 5. 1 酵母基因转化与性状互补
利用 Yip (整合型质粒)可以对酵母基因组中的任意基因进行精确的敲除,通过孢子繁殖中的四分体分析技术进行观测和研究。

• 使带有致死突变位点和可能的外源功能互补基因的
酵母二倍体细胞在饥饿条件下形成四分体孢子。
• 将这四个孢子用酵母四分体分离系统分开。
• 如果引入的外源基因能互补突变位点,含有突变基
因的单倍体可以存活,否则就不能存活。

W1536 5B
elo2Δelo3Δ
KCS12
KCS6
KCS13
KCS2
棉花 KCS 基因互补酵母 elo2Δelo3Δ突变体。W1536 5B 为野生型单倍体细胞; elo2Δ
elo3Δ 为酵母自身的超长链脂肪酸合成酶双突变的单倍体,致死型; KCS12 , K
CS6 , KCS2 , KC
S13 为不同的棉花 K
CS 基因。

6. 5. 2 外源基因在酵母中的功能鉴定
• 将外源基因克隆于酵母表达载体上,转化野生型或突变酵母菌株,通过观察酵母的表型变化或分析细胞中化学成分的变化即可推测该基因的生物学功能。
• 在酵母细胞中表达外源基因与绿色荧光蛋白基因的融合蛋白后,可通过荧光显微镜观察荧光所在的亚细胞区域,从而了解该基因产物在细胞中的定位。
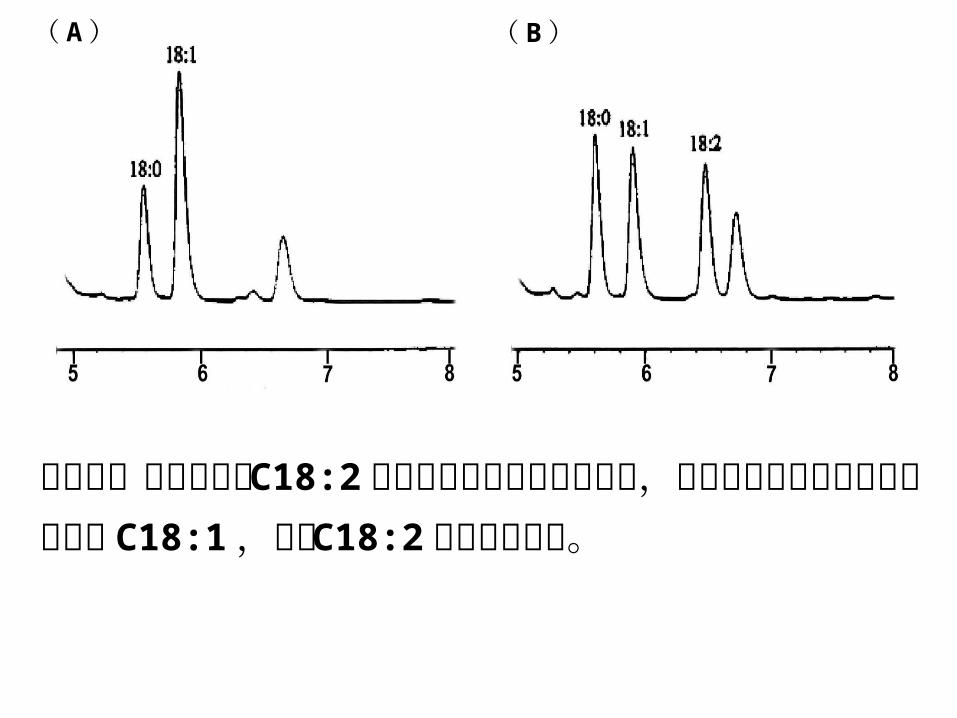
( A ) ( B )
将空载体转入无合成 C18:2 长链脂肪酸能力的酵母细胞,气相色谱证明细胞抽提物中只有 C18:1 ,没有 C
18:2不饱和脂肪酸。

• 野生型酵母细胞中 18碳不饱和脂肪酸主要是 C18:1 ,若将棉花中可能编码ω-6脂肪酸脱氢酶的基因转入野生型酵母细胞中,气质联动技术证明带有棉花基因的酵母细胞中出现 C18 : 2不饱和脂肪酸,证明该基因编码功能型 ω-6脂肪酸脱氢酶。

6. 6 其它分子生物学技术1 、 凝胶滞缓实验凝胶滞缓试验( gel retardation assay ),又叫作 D
NA迁移率变动试验( DNA mobility shift assay ),是用于体外研究 DNA 与蛋白质相互作用的一种特殊的凝胶电泳技术。

在凝胶电泳中,由于电场的作用,裸露的 DNA朝正电极移动的距离与其分子量的对数成反比。如果此时DNA分子与某种蛋白质相结合,那么,由于分子量增大,它在凝胶中的迁移作用便会受到阻滞,在特定电压和时间内朝正电极移动的距离也就相应缩短了。


Gel mobility shift assay using in vitro expressed AP2/EREBP protein and the conserved GCC or DRE cis-element
50 ng protein
50 ng protein
DRE: ATACTACCGACAT
DRE-2: ATACTGCCGACAT
mDRE: ATACTACTTATAT
GCC: TAAGAGCCGCCACT
mGCC: TAAGATCCTCCACT
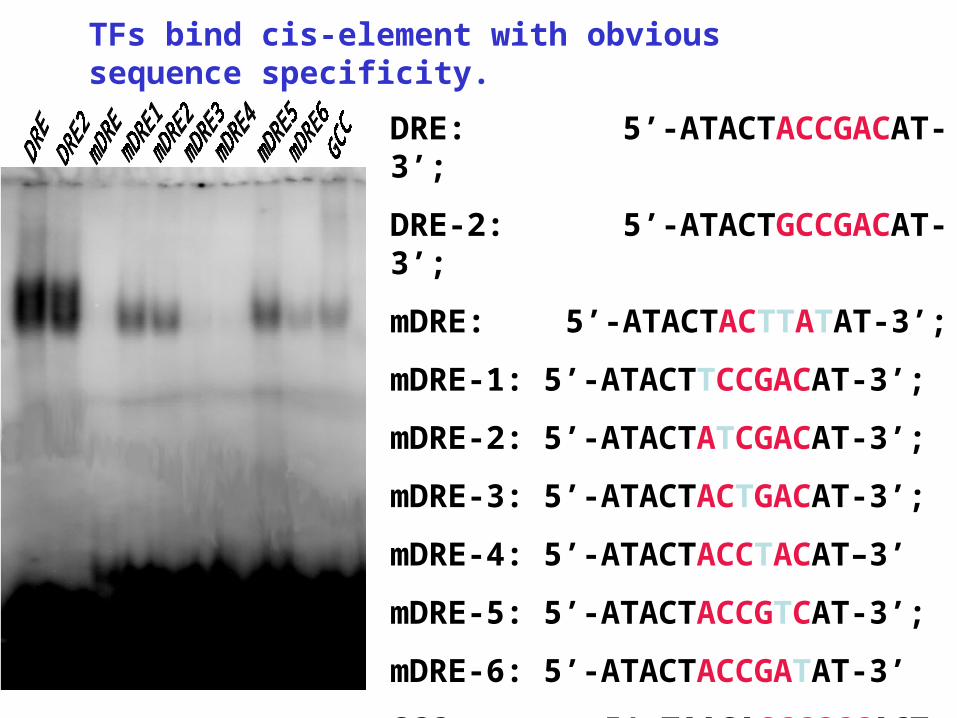
TFs bind cis-element with obvious sequence specificity.
DRE: 5’-ATACTACCGACAT-3’;
DRE-2: 5’-ATACTGCCGACAT-3’;
mDRE: 5’-ATACTACTTATAT-3’;
mDRE-1: 5’-ATACTTCCGACAT-3’;
mDRE-2: 5’-ATACTATCGACAT-3’;
mDRE-3: 5’-ATACTACTGACAT-3’;
mDRE-4: 5’-ATACTACCTACAT–3’
mDRE-5: 5’-ATACTACCGTCAT-3’;
mDRE-6: 5’-ATACTACCGATAT-3’
GCC: 5’-TAAGAGCCGCCACT-3’

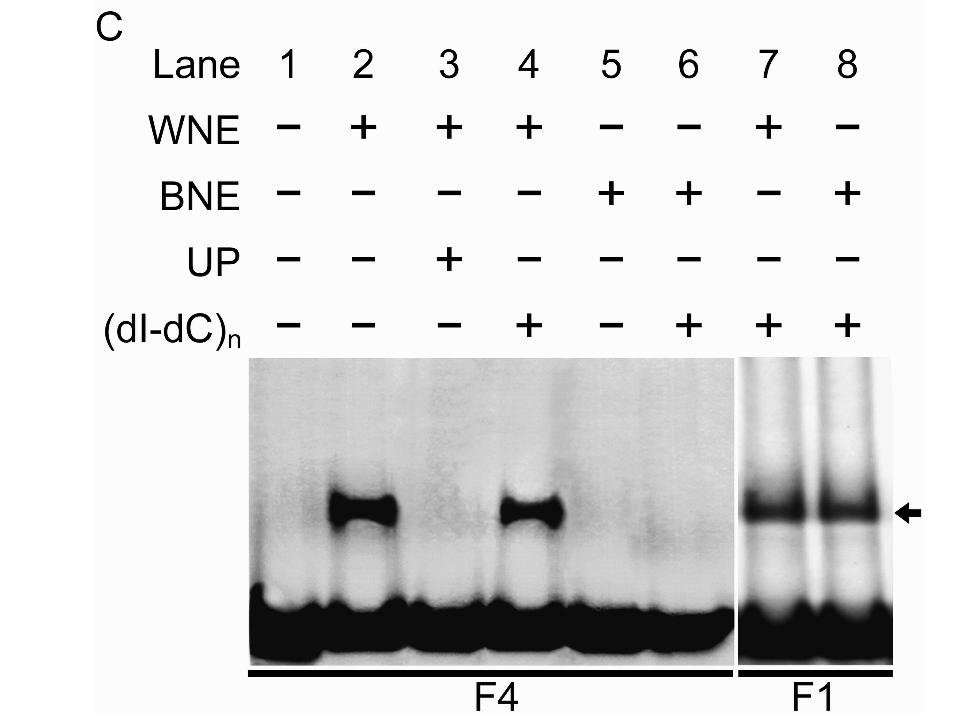

BARD1 Is Involved in the Formation of a Protein-DNA Complex with Specific Regionson the WUS Promoter.

2 、 蛋白质磷酸化分析技术
蛋白激酶催化 ATP或 GTP 的 γ 磷酸基团转移到底物蛋白的丝氨酸、苏氨酸或酪氨酸残基上,促使底物蛋白发生磷酸化,而蛋白质磷酸酯酶则能催化底物蛋白发生去磷酸化。此外,底物蛋白的组氨酸、赖氨酸、精氨酸、天冬氨酸、谷氨酸和半胱氨酸残基上也能发生可逆磷酸化。

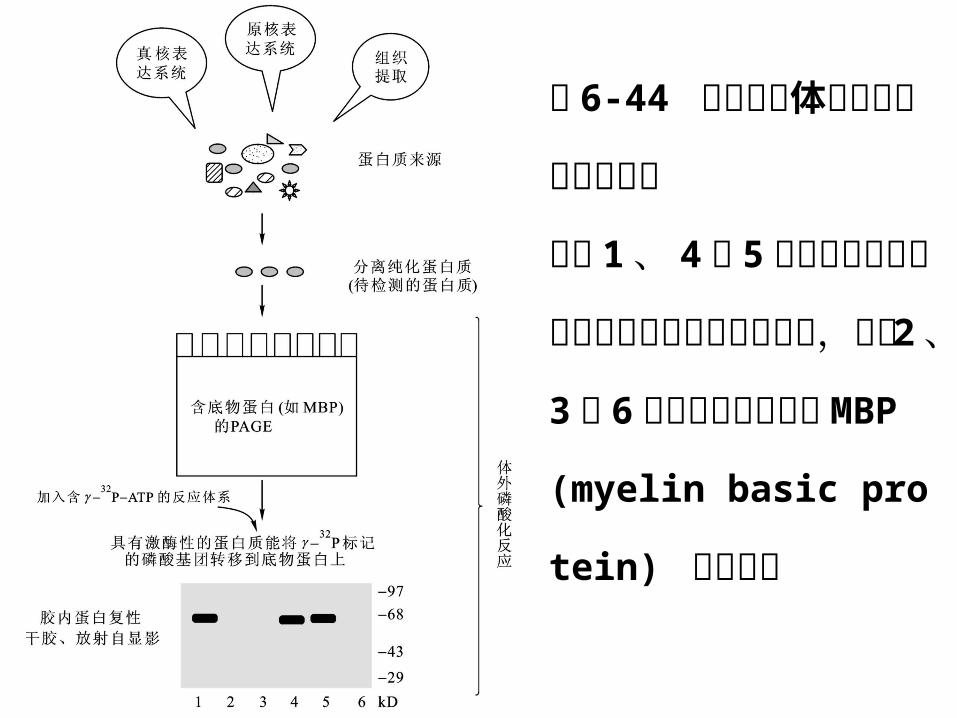
图 6-44 蛋白激酶体外
磷酸化活性分析。
泳道 1 、 4 和 5 中的蛋
白质具有使蛋白质磷酸
化的激酶活性,泳道 2 、
3 和 6 中的蛋白质不能
使 MBP (myelin basic
protein) 磷酸化。

3 、蛋白质免疫印迹实验( Western blotting )。
是在蛋白质凝胶电泳和固相免疫测定基础上发展起来
的一种技术,检测样品中是否存在蛋白抗原。
被测蛋白只能与标记的特异性抗体相结合,而这种结
合不改变该蛋白在凝胶电泳中的相对分子质量,被用
于测定抗原的相对丰度或与其它已知抗原的关系,也
是评价新抗体特异性的一种好方法。

图 6-46 免疫印迹
( Western Blottin
g )示意图

是检测样品中是否存在蛋白抗原的一种可靠方法。主要步骤:a 、蛋白样品的制备;b 、 SDS-PAGE 分离 ;
c 、转膜并封闭膜上未反应位点,减少非特异性吸附;d 、与相应非标记抗体(一抗)反应;e 、洗去多余一抗,加酶促偶联或放射性同位素标记的二抗,显色或放射自显影法检测凝胶中的蛋白成分。

4 、 细胞定位及染色技术
蛋白质在组织及细胞内的亚定位,一直是细胞生物学
和分子生物学研究的核心问题,因为了解特定蛋白质
的定位,才有可能了解其生物学功能。研究细胞定位
可采取多种方法,最常用的是荧光蛋白标记和免疫荧
光法。

最常用的可能是绿色荧光蛋白( GFP ),它由 238
个氨基酸组成,有两个吸收峰,最大吸收峰在 395 n
m ,另外一个在 475 nm ,前者由紫外光激发,后者
由蓝光激发。发射光也有两个峰值,分别是 509 nm
和 540 nm;呈绿色或黄绿色荧光。

实验中,将绿色荧光蛋白的编码序列与目的基因启动
子融合形成嵌合基因,转染植物或动物细胞,由于融
合基因与目的基因的表达模式相同(使用相同的表达
调控元件),通过荧光显微镜观察 GFP 在组织或亚细
胞中的分布就相应确定了目的蛋白在细胞中的定位。
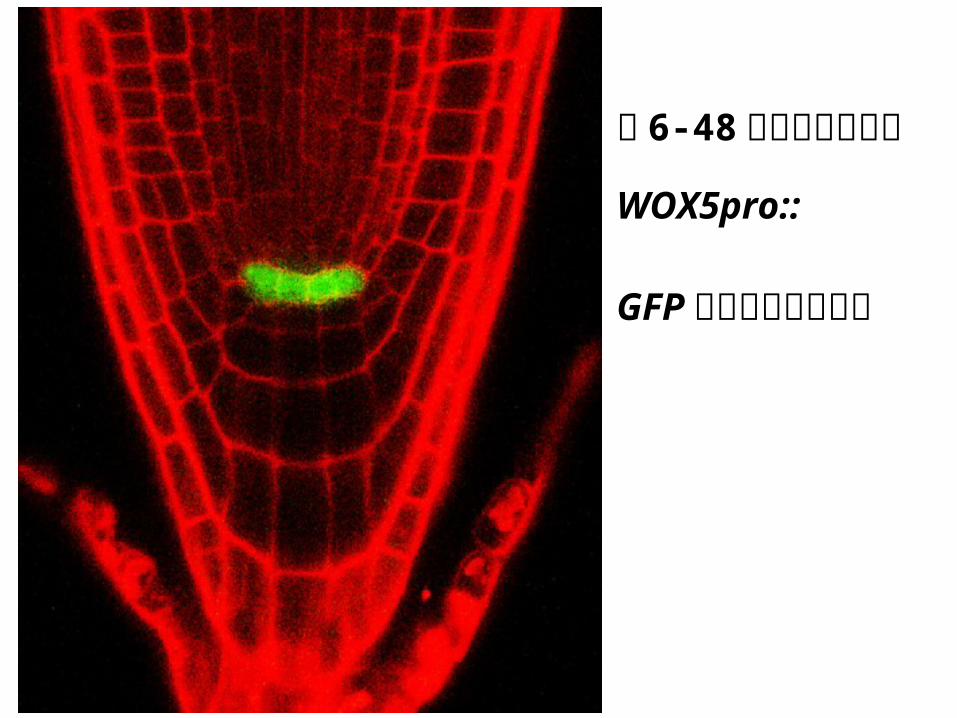
图 6-48野生型拟南
芥中 WOX5pro::
GFP融合基因的表
达。

免疫荧光技术是将免疫学方法(抗原抗体特异结合)
与荧光标记技术结合起来研究特异蛋白抗原在细胞内
定位的方法。
用针对特异蛋白抗原的荧光标记抗体作为分子探针检
测细胞或组织内的相应抗原,由于所形成的抗原抗体
复合物上含有荧光素,利用荧光显微镜观察标本,确
定荧光所在细胞或组织,从而对抗原蛋白进行定位。


1 、说出用 RNA-seq 技术进行转录组学分析的原理。2 、选择性剪接主要可能有那几种不同类型?3 、为什么可以用 RPKM 来衡量基因表达的水平?4 、 RNA 原位杂交的主要实验过程及应用。5 、基因定点突变的原理与实验过程。6 、基因芯片技术及应用。7 、写出蛋白质免疫印迹实验的原理和过程 .
8 、说出免疫共沉淀 (Co-IP) 实验的原理与过程。9 、说出经典遗传学( Forward genetics )与现代遗 传学(反向遗传学, Reverse genetics )的异同。

10 、什么是完全基因敲除和条件型基因敲除。
11 、说出动植物基因敲除实验的技术要点。
12 、为什么要用正负筛选法筛选发生同源重组细胞?
13 、酵母单、双杂交系统的基本原理与应用。
14 、蛋白质 Pull-down 实验原理与主要步骤。
15 、染色质免疫共沉淀 (ChiP) 技术原理与基本过程。
16 、 RNAi 技术原理。
17 、噬菌体展示技术的原理及应用?
18 、凝胶滞缓实验的原理与应用。

19 、蛋白质激酶功能研究的主要原理与方法。
20 、研究蛋白质细胞定位的主要方法?
21 、说出用荧光蛋白标记法进行细胞内蛋白质定位研
究的技术要点。
22 、说出用免疫荧光法定位抗原蛋白的技术要点。
23 、如何用外源基因在酵母中进行功能鉴定。
24 、如何通过等离子表面共振技术研究基因功能?25 、写出 TALEN 的英文全称及技术要点。

Thank You !