widespread occurrence of bacterial human virulence determinants
TRANSCRIPT

1
Widespread occurrence of bacterial human virulence determinants in soil and 2
freshwater environments 3
4
Ditte A. Søborga, Niels Bohse Hendriksena, Mogens Kilianb, and Niels Kroera# 5
6
a Department of Environmental Science, Aarhus University, Frederiksborgvej 399, DK-4000, 7 Roskilde, Denmark 8
b Department of Biomedicine, Aarhus University, Wilhelm Meyers Allé 4, DK-8000, Aarhus C, 9 Denmark 10
11
12
13
14
Running title: Virulence determinants in soil and freshwater environments 15
#Corresponding author. Mailing address: Department of Environmental Science, Aarhus 16 University, Frederiksborgvej 399, DK-4000 Roskilde, Denmark; E-mail [email protected]; Phone 17 +45 87 15 87 01 18
Copyright © 2013, American Society for Microbiology. All Rights Reserved.Appl. Environ. Microbiol. doi:10.1128/AEM.01633-13 AEM Accepts, published online ahead of print on 8 July 2013
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

2
ABSTRACT 19 The occurrence of 22 bacterial human virulence genes (encoding toxins, adhesins, secretion 20 systems, regulators of virulence, inflammatory mediators, and bacterial resistance) was 21 investigated by nested PCR in beech wood soil, roadside soil, organic agricultural soil, and 22 freshwater biofilm. The presence of clinically relevant bacterial groups known to posses the 23 virulence genes was tested by PCR of 16S and 23S rRNA genes. For each of the virulence genes 24 detected in the environments, sequencing and NCBI BLAST confirmed the identity of the PCR 25 products. The virulence genes showed widespread environmental occurrence as 17 different 26 genes were observed. Sixteen genes were detected in the beech wood soil, 14 in the roadside 27 and organic agricultural soils, while 11 were detected in the freshwater biofilm. All types of 28 virulence traits were represented in all environments, however, the frequency by which they 29 were detected was variable. A principal component analysis suggested that several factors 30 influenced the presence of the virulence genes, however, their distribution was most likely 31 related to the level of contamination by polycyclic aromatic hydrocarbons and pH. The 32 occurrence of the virulence genes in the environments did generally not appear to be the 33 result of the presence of clinically relevant bacteria, indicating an environmental origin of the 34 virulence genes. The widespread occurrence of the virulences traits, and a high degree of 35 sequence conservation between the environmental and clinical sequences, suggest that soil 36 and freshwater environments may constitute reservoirs of virulence determinants normally 37 associated with human disease. 38
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

3
INTRODUCTION 39 Bacterial pathogens continue to cause major threats to human health and welfare due to the 40 emergence of new pathogens, re-emergence of previously well-controlled infectious diseases, 41 and increasing prevalence of antimicrobial resistant bacteria (1-5). However, the origin of 42 many emerging human pathogens and their virulence genes remains to be elucidated (2,6). 43 Traditionally, it has been believed that the complex interactions between pathogenic 44 bacteria and their hosts are the primary driving forces for pathogens to develop mechanisms 45 to overcome host defense strategies (4). For this reason, most of our knowledge regarding 46 virulence factors and bacterial pathogenicity derives from studies of infections (6-8). 47 However, the origin of virulence genes may be environmental bacteria (8), and the acquisition 48 of virulence genes through horizontal gene transfer of mobile genetic elements in outer non-49 host (non-human) environments may play an important role for the evolution of bacterial 50 pathogens (4,9). Hence, studies on the occurrence and expression of virulence genes in non-51 host environments are important to obtain better knowledge of the origin and evolution of 52 bacterial pathogens (6,8). 53 Virulence factors are likely to have similar physiological functions in the environment as in 54 the host organism. This could be the case for e.g. iron acquisition systems or for structural 55 elements such as capsules or polysaccharides for which the role in virulence is a consequence 56 of the host’s mechanisms for detecting and eliminating pathogens (8,10). Other virulence 57 factors may have different functions in e.g. bacterial infection and in fitness to specific 58 environmental habitats (8,11). This dual role has been seen for an adhesion factor of Vibrio 59
cholera which mediates attachment to human epithelial cells required for intestinal 60 colonization as well as attachment to zooplankton enhancing its environmental survival (12). 61 Furthermore, it has been shown for E. coli, Pseudomonas aeruginosa, Legionella pneumophila, 62
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

4
and Listeria monocytogenes that virulence factors required for infecting humans are also 63 needed for infecting plants, invertebrates or insects (13-18). 64 If survival and adaptation of a bacterium in humans is similar to the survival and 65 adaptation of bacteria in the environment, then genes that typically are associated with 66 pathogenicity will be present in non-pathogens from the environment. For instance, Casas et 67
al. (19) have demonstrated that phage-encoded exotoxins are widespread in water, sediment 68 and soil. Additionally, sequence analysis revealed a high number of protein secretion systems 69 and virulence gene homologues in marine bacteria (6). These findings support the theory that 70 virulence genes are of environmental origin and that these traits may be used by naturally 71 occurring bacteria to survive and grow in the environment (20). 72 Here we report the widespread occurrence of several classes of human virulence 73 determinants, including toxins, adhesins, secretions systems, inflammatory mediators, 74 regulators of virulence, and bacterial resistance factors, in three different soils and a 75 freshwater biofilm. The study focuses on the environmental gene pool of pathogenic traits 76 rather than on the occurrence of specific pathogens in the environment. Previous studies have 77 either focused on the detection of clinically relevant bacteria in non-host environments (e.g. 78 (21,22) or on the environmental survival of known pathogens (e.g. (23). The widespread 79 occurrence of virulence genes in soil and freshwater suggests that these environments may 80 constitute reservoirs of virulence determinants normally associated with human disease. 81 82
MATERIALS AND METHODS 83
Sampling. Triplicate surface soil samples (0-10 cm depth) of approximately 1 kg were 84 collected on November 16th 2009 at three different localities: 1) an organic agricultural field 85 (Snubbekorsgård, Taastrup, Denmark), 2) a beech wood (Boserup Skov, Roskilde, Denmark), 86
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

5
and 3) an urban roadside (Jarmers Plads, Copenhagen, Denmark). Sampling was done using a 87 sterile shovel and samples were transfered to sterile plastic bags. Soil samples were passed 88 through a sterile 4 mm mesh sieve, homogenized by careful mixing with a sterile spoon, and 89 stored at 4°C until use. In addition to the soil samples, triplicate ~1 kg samples of pebbles 90 (diameter 2-4 cm) and 1 L of water were collected on December 1st 2009 from a lake (Lake 91 Lyngby, Lyngby, Denmark) at 50 cm depth using sterile gloves, plastic bags and glass bottles. 92 To dislodge biofilm, pebbles and water were sonicated for 30 min at room temperature using 93 a Branson 5210 ultrasonic bath (Branson, Germany). After removal of the pebbles, the water 94 was centrifuged at 3 000 g for 15 min at 20°C and the pelleted biofilm stored at 0°C until 95 further analysis. Sterile gloves were used during all handling of the samples. 96
Chemical and biological analyses. Soils were analyzed by Eurofins-Steins A/S (Odense, 97 Denmark) for pH, texture (grain sizes) and content of inorganic nutrients (phosphorus, total 98 nitrogen). Determination of water holding capacity and loss of ignition (carbon content) of the 99 soils were measured by standard procedures: 105°C overnight and 550°C for 2 h, respectively 100 (24) while the concentration of polycyclic aromatic hydrocarbons (PAHs) was measured 101 according to Hollender et al. (25). pH and concentration of dissolved organic carbon (26) 102 were determined in the water samples. 103 Total numbers of bacteria in the soils were determined by acridine orange (AO) direct 104 counting (27). Briefly, a 1:10 000 dilution of 1 g soil in 9 ml sterile Milli-Q water was filtered 105 onto 0.2 µm polycarbonate membranes (25 mm in diameter, Nucleopore®) and at least 400 106 bacteria per sample were counted. 107
Detection of virulence determinants. DNA was extracted using the PowerSoilTM DNA 108 Isolation Kit (MoBio Laboratories Inc., USA) according to the manufacturer’s instructions. Soil 109 DNA was extracted from triplicate 250 mg subsamples from each of the triplicate soil samples 110
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

6
per sampling locality, while biofilm DNA was extracted from triplicate 100 mg subsamples of 111 the triplicate biofilm samples. Thus, nine DNA extractions were prepared from each 112 environment. DNA was stored at -20°C until use. DNA concentration and purity were 113 evaluated spectrophotometrically by UV absorption at 260 nm (A260) and A260/A280 ratio, 114 respectively, using a NanoDropTM 2000C Spectrophotometer (Thermo Scientific, USA). 115 Detection of 22 different virulence genes (Table 1) was carried out by PCR. Primer sets 116 (Supplemental Table S1) were derived directly from the literature or in degenerated versions, 117 or they were designed de novo from available nucleotide sequences using Primer3 Version 118 0.4.0 (28). The specificity of all primer sets was verified by NCBI BLAST (29,30). 119 PCR was performed in 25 μl reaction mixtures containing 1.25 U AmpliTaq Gold® DNA 120 polymerase, 1 × PCR Gold buffer, 3.0 mM MgCl2 (all from Applied Biosystems, USA), 200 μM 121 dNTP (Thermo Scientific, USA), 0.4 µM of each primer and 1 µl DNA template (1-5 ng). The 122 PCR was performed with a Hybaid PCR Express Thermal Cycling (AH Diagnostics, Denmark) 123 and consisted of 1 cycle of denaturation at 95°C for 5 min, 35 cycles of denaturation at 95°C 124 for 45 s, annealing at the appropriate temperature for each primer set for 60 s, and extension 125 at 72°C for 90 s followed by a final extension at 72°C for 5 min after which the PCR products 126 were maintained at 4°C. Nested or semi-nested PCR was performed in a similar manner with 127 0.5 µl product of the first PCR as template. Two PCRs were run for each sample (i.e. 18 PCRs 128 per environment per primer set). PCR products were analyzed by 1.5% or 2.5% (nested PCR) 129 agarose gel electrophoresis. The presence of a virulence gene (i.e. an internal amplicon with 130 homology to a fragment of a targeted virulence gene) was only scored as positive if it was 131 documented in at least two replicate PCRs. For both conventional and nested PCR, 4-10 132 negative controls (no added DNA) were included for every 72 sample PCRs. In total this 133
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

7
summed up to more than 200 negative controls. In no case did any of these negative controls 134 give rise to false positives. 135 Bacteria known to possess the respective virulence genes (Table 1) were used as positive 136 controls. Generally, bacteria were cultured on Luria-Bertani agar and incubated overnight at 137 30°C. For H. pylori NCTC 11638, Columbia agar supplemented with 4% horse blood was used. 138 During the incubation period, a liquid film of brain heart infusion was maintained on the agar 139 surface. Incubation was performed under microaerobic conditions (10% [vol/vol] CO2, 85% 140 [vol/vol] N2, and 5% [vol/vol] O2) for 7 days. DNA was extracted by boiling (31). E. coli O157 141 and B. anthracis DNA was donated by Statens Serum Institute (Denmark). 142
Sequencing of PCR products. To confirm the identity of the PCR products, one PCR 143 product of each of the detected virulence gene fragments was sequenced. Products was run on 144 a 2% agarose gel, the specific bands cut out under UV light, and DNA extracted using the 145 QIAEX II Gel Extraction Kit (QIAGEN, Germany). Extracted DNA was amplified by PCR in 10 μl 146 reaction mixtures containing 1 µl BigDye® Terminator v1.1 Ready Reaction Mix, 0.75 X 147 Sequencing Buffer (Applied Biosystems, USA), forward or reverse primers at a concentration 148 of 2.5 µM, and 1 µl of the extracted DNA. The PCR conditions were 1 cycle of denaturation at 149 96°C for 5 min, 25 cycles of denaturation at 96°C for 30 s, annealing at 59°C for 15 s, and 150 extension at 60°C for 4 min after which the PCR products were maintained at 4°C. DNA was 151 precipitated with 75% isopropanol, and following centrifugation, the pellet was dried for 5 152 min in a vacuum oven and stored at -20°C. DNA was resuspended in Hi-DiTM Formamide 153 (Applied Biosystems, USA) and sequenced in one direction by Macrogen Inc. (Seoul, Korea) 154 using an ABI 3130xl Genetic Analyser (Applied Biosystems, USA). 155
Assessment of detection limit. To determine the detection limit of the nested PCR 156 procedure, bacteria known to possess the respective virulence associated genes (Table 1) 157
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

8
were added at densities of approximately 0, 10, 100, 1 000, and 10 000 cells per 250 mg soil 158 or 100 mg biofilm prior to DNA extraction. E. coli O157 and B. anthracis were not added as live 159 cells but as DNA. DNA extraction and PCR conditions were as described above. 160
Detection of clinically relevant bacteria in environmental samples. To assess whether 161 the origin of the observed virulence genes was environmental or due to the presence of 162 clinically relevant bacteria, bacterial groups from which the 22 virulence genes were 163 originally identified (Salmonella spp., E. coli, Helicobacter spp., Agrobacterium tumefaciens, 164 and B. cereus group bacteria; Supplemental Table S2), were targeted by nested PCR of 16S and 165 23S rRNA genes. PCR was performed as described for the detection of the virulence genes, 166 except that annealing temperatures were adjusted appropriately and 40 cycles were run for E. 167
coli and Helicobacter ssp. in the conventional PCR. The nine DNA extractions from each 168 environment, prepared for the detection of the virulence genes, were used as template DNA 169 and dublicate PCR was run for each extract. Three negative controls containing sterile Milli-Q 170 water instead of template DNA were set up for each of the primer sets. 171
Phylogenetic analysis. Alignments of positive PCR products were performed with the 172 Clustal software and sequence variations determined on basis of pairwise distances using 173 MEGA Version 5.01 (32). Sequences were manually edited and a few sequences of poor 174 quality omitted. Phylogenetic associations of the PCR products of each of the genes cytK2, 175
hblA, invA, stx-1, and wbdI were determined by including sequences from NCBI BLAST 176 searches (29,30) in the phylogenetic analyses. The analyses were performed with MEGA 177 Version 5.01 using the Minimum Evolution algorithm (complete deletion) (Fig. 3 and 178 Supplemental Fig. S1, A-D). 179
Statistical analyses. A principal component analysis (PCA), based on the frequency of 180 which the virulence genes were detected in the replicate environmental samples, was 181
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

9
performed using the software LatentiX Version 2.00 (Latent5 Aps, Denmark). One-way 182 ANOVA was applied to explore differences along the axes. Further, one-way ANOVA was 183 applied to determine differences between environmental characteristics of the soil 184 environments. 185
Accession numbers. Sequences longer than 200 bp were deposited in GenBank under the 186 following accession numbers: KC699845 to KC699851 for fimH BWS, focG BWS, wbdI BIO 187 1A.3, wbdI BWS 1B.2, wbdI BWS 3C.3, wbdI OAS 1C.2, and wbdI RS 1A.3, respectively. All 188 sequences, including those shorter than 200 bp, can be found in Supplemental Table S3. 189 190
RESULTS 191
Physico-chemical characteristics of environments. The physico-chemical characteristics of 192 the individual soil samples differed in a number of respects (Table 2). Most significant was the 193 difference in PAH content, with the roadside soil having a concentration > 25× higher than the 194 beech wood soil and > 100× higher than the organic agricultural soil (P < 0.0001 in both 195 cases). No significant differences in PAH content between the beech wood and organic 196 agricultural soils were observed (P > 0.5). Other characteristics that differed between the soils 197 were pH and phosphorus concentration. All other measured parameters were basically 198 similar, including the bacterial density. 199
Environmental distribution of virulence genes. The virulence genes showed 200 widespread environmental distribution as 77% of the investigated genes (17 out of 22) were 201 observed in the soil and freshwater habitats (Table 3). The highest number of genes [16] was 202 found in the beech wood soil, followed by the roadside and organic agricultural soils [14], and 203 the freshwater biofilm [11]. All the detected genes were present in the beech wood soil except 204
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

10
for the H. pylori vacuolating cytotoxin gene, vacA, which was only found in the organic 205 agricultural soil and the freshwater biofilm (Table 3). 206 Virulence genes of all groups of virulence determinants were found in at least one of the 207 investigated environments (Table 3). Among the group of toxins, five toxin genes were 208 observed. In the organic agricultural soil, all five genes were present, while four toxin genes 209 were detected in the other environments. The three undetected toxin genes were the E. coli 210 hemolysin A gene, hlyA, the B. cereus cytotoxin K1 gene, cytK1, and the B. anthracis edema 211 factor, cya (Table 3). All the adhesion group genes were found in the beech wood soil, while 212 the P fimbrial gene, papC, was found in all four environments. Two Salmonella genes involved 213 in the Type III secretion system, invA and spiA, were detected in all environments while a 214 third Salmonella secretion system gene complex, sipB-sipC, remained undetected. A fourth 215 gene, virD4, of A. tumefaciens was seen in two of the soil environments. Among the other 216 groups of virulence determinants (regulators of virulence, inflammatory mediators, and 217 bacterial resistance), the PhoQ sensor, phoQ, the wbdI gene of the E. coli O-antigen gene 218 cluster, the capsular biosynthesis protein, capA, and the polymyxin resistance gene, arnT, 219 were observed in all environments (Table 3). 220
Relative frequency of virulence genes. A semi-quantitative nested PCR approach (Fig. 1), 221 based on the number of positive to total number of PCRs, was applied to assess the relative 222 frequency of the investigated virulence determinants. As shown in Table 3, hblA, invA, phoQ, 223 and wbdI were relatively frequently observed in organic agricultural soil, beech wood soil, 224 roadside soil and freshwater biofilm, respectively, whereas e.g. cytK2 and spiA were relatively 225 infrequently observed in all the environments. The detection limit was approximately 400 226 gene copies (cells) g-1 soil (Fig. 1) and 1 000 gene copies (cells) g-1 biofilm (not shown). 227
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

11
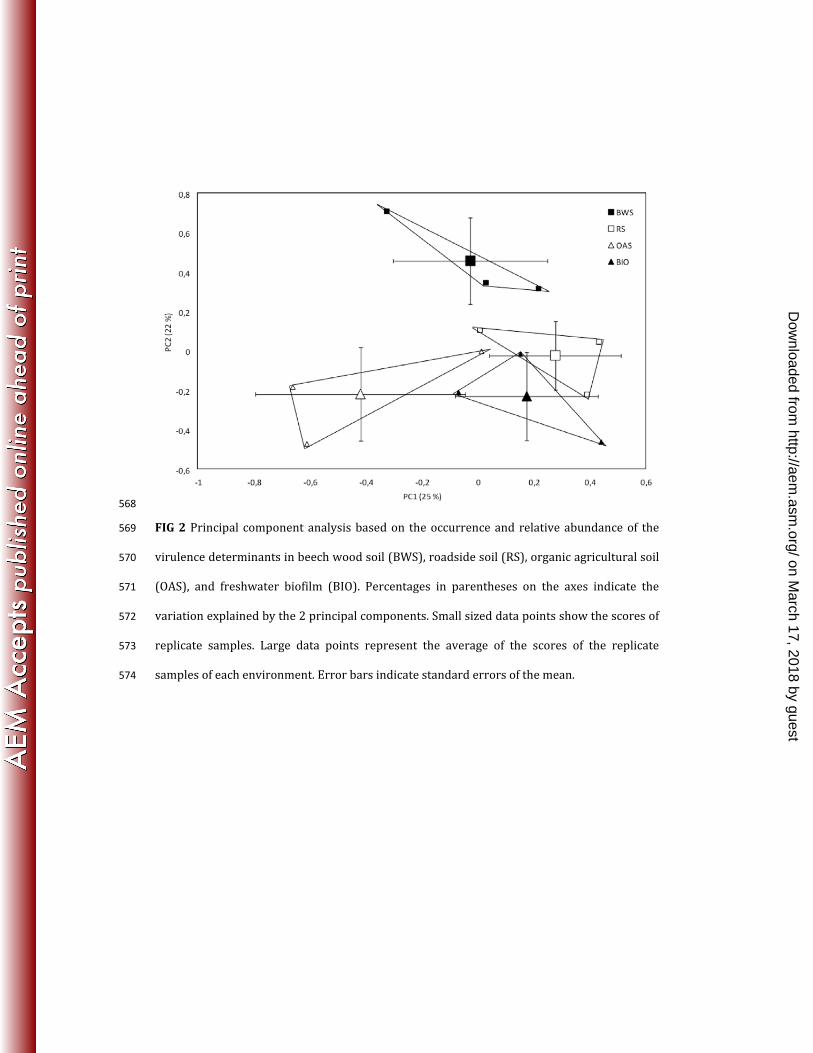
Differences between the four environments with respect to occurrence and relative 228 abundance of the virulence determinants were demonstrated by PCA (Fig. 2). Along the PC1 229 axis, the organic agricultural soil clustered separately from the roadside soil and the 230 freshwater biofilm (One-way ANOVA, P < 0.05). The distribution of the soil environments 231 along the PC1 axis, from the lowest concentration of PAHs (organic agricultural soil, beech 232 wood soil) to the highest concentration (roadside soil), suggested that PC1 explained 233 variation arising from differences in PAH concentration. Along the PC2 axis, the beech wood 234 soil was significantly different from all the other environments (One-way ANOVA, P < 0.05). A 235 possible explanation for the separation of beech wood soil from the other environments along 236 this axis could be differences in pH, as the beech wood soil had a low pH (5.3) compared to the 237 other environments (Table 2). 238
Sequence homology and phylogenetic analysis. The sequence homology between the 17 239 detected virulence gene fragments and the bacterial strains known to carry the corresponding 240 target genes was 90 - 100%. Furthermore, NCBI BLAST analyses demonstrated that the 241 environmental sequences were homologous to relevant sequences in the databases (highest 242 E-score ≤ 10-14 for best hit, stx-2). Alignment of sequences of all PCR products for cytK2, hblA, 243
invA, wbdI, and stx-1 showed a sequence variation of 2% for cyK2, 0 – 19% for hblA, 0 – 10% 244 for invA, 0 – 1% for wbdI, and 0% for stx1. Despite variations of up to 19%, as seen between 245 two hblA sequences, all sequences of the five genes were homologous to the appropriate 246 virulence associated genes (highest E-score ≤ 10-16 for best hit, stx-1). 247 Phylogenetic analysis of the partial cytK2, hblA, invA, wbdI, and stx-1 gene sequences 248 confirmed their close relatedness to genes of clinically relevant bacteria (Fig. 3 and 249 Supplemental Fig. S1 A-D). Thus for invA, 28 sequences representing all four environments 250 clustered closely together with invA of various serovars of Salmonella enterica (Fig. 3). Three 251
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

12
roadside soil sequences and one freshwater biofilm sequence, however, formed a separate 252 cluster. Compared to invA, the phylogenetic distances between the environmental and 253 database hblA sequences were relative large, and some of the environmental sequences 254 formed their own cluster or sub-cluster (Supplemental Fig. S1, A). For wbdI, the majority of 255 the environmental sequences fell within a cluster defined by several clinical E. coli serotype 256 O111; however, two sequences formed separate clusters (Supplemental Fig. S1, B). The ten 257
stx-1 and four cytK2 environmental sequences basically clustered together with the 258 corresponding clinical sequences (Supplemental Fig. S1, C and D). 259
Occurrence of clinical relevant bacteria. The occurrence of virulence genes in the 260 environments did generally not appear to be the result of the presence of clinically relevant 261 bacteria. Salmonella spp. and Helicobacter spp. were not detected in any of the environments. 262 Similarly, E. coli/Shigella were not detected in the beech wood and roadside soils, B. cereus 263 group bacteria were not detected in the freshwater biofilm, and A. tumefaciens not detected in 264 the roadside soil and the freshwater biofilm. B. cereus group bacteria, however, were 265 relatively frequent in the beech wood soil (17 out of 18 replicate PCR tubes; 94%), the organic 266 agricultural soil (94%), and the roadside soil (89%). Similarly, E. coli/Shigella were detected 267 in 55% of the replicate nested PCRs of the agricultural soil and in 89% of the replicate nested 268 PCRs of the freshwater biofilm, while A. tumefaciens was observed in the beech wood and 269 agricultural soils (11% and 22%, respectively). 270 271
DISCUSSION 272 Numerous bacterial virulence genes were found in three different soil environments and a 273 freshwater biofilm by use of a nested PCR approach, with DNA extracted directly from the 274 natural environments. The nested PCR approach theoretically made it possible to detect as 275
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

13
few as one bacterial cell carrying a specific gene in the reaction tube. However, to minimize 276 the influence of PCR inhibitors such as humic substances (33), it was necessary to dilute the 277 soil and biofilm extracts, resulting in a detection limit of approximately 400 genes copies g-1 278 soil (Fig. 1) or 1 000 gene copies g-1 biofilm. This is comparable to the detection limit of 100 - 279 1 000 gene copies g-1 soil found by Agersø et al. (34) who used nested PCR to study the 280 presence of the tetracycline resistant gene, tet(M), in farmland soil. Higher detection limits 281 have been reported in studies applying qPCR, e.g. 1.3 × 105 amoA gene copies g-1 dry soil (35), 282 and 104 narG gene copies g-1 soil (36). Thus, compared to qPCR, our approach was suitable for 283 the specific and sensitive detection of virulence genes present in low numbers. 284 Nested PCR has the limitation that only the presence or absence of a specific gene can be 285 assessed. However, we applied a semi-quantitative approach based on the fraction of positive 286 to total number of replicate PCRs. This was possible because the virulence genes were present 287 in numbers very close to the detection limit, hence, of stochastic reasons, not all the replicate 288 18 PCRs resulted in a positive amplification (Table 3). Since the number of positive to 289 negative PCR tubes thus followed the Poisson distribution, it was possible to determine the 290 relative abundance of the investigated virulence determinants by calculating the fraction of 291 positive reactions. 292 A common problem with sensitive methods such as nested PCR is the risk of obtaining false 293 positive results (37). To minimize the risk of detecting false positive virulence determinants, 294 the primers targeting the 22 virulence genes were subjected to homology search using NCBI 295 (29,30) and found to match the genes of interest. Annealing temperatures and primer lengths, 296 which are the main factors influencing the specificity of the amplification (38), were used as 297 described in the literature or carefully selected with respect to melting temperatures. 298 Nevertheless, several cases of unspecific PCR amplifications were observed, although only 299
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

14
when the specific gene of interest was absent (e.g. fimH, Fig. 1). In other words, in all positive 300 detections only a single band was observed after agarose gel electrophoresis. As a further 301 measure to eliminate false positives, sequencing of positive PCR products confirmed the 302 identity of the bands. Finally, numerous negative controls were included in all PCR 303 experiments. In no case was a negative control observed to be positive. 304 The study was based on DNA extracted directly from the natural environments. As a 305 consequence, it was difficult to assess if the 17 detected genes were present in naturally 306 occurring environmental bacteria, in bacteria spread to the environments from human or 307 animal sources (39-41), contained in free phage particles, or existed as free DNA. Clinically 308 relevant bacteria may survive for varying periods of time in the environment depending on 309 their ability to cope with the environmental conditions (42). Environmental survival of 310 potential human pathogens has for instance been seen for Vibrio spp., Aeromonas spp., E. coli, 311
Campylobactor spp., Arcobacter spp. (43), Salmonella spp., Clostridium perfringens, 312
Pleisiomonas shigelloides (44), and H. pylori (45). However, since we did not detect Salmonella 313 ssp. or Helicobacter ssp. in any of the environments and only found E. coli, B. cereus group 314 bacteria and A. tumefaciens in some of the environments, it is likely that the majority of 315 virulence genes were present amongst the natural bacterial (or phage) communities. This is 316 supported by the cases of separate clustering of sequences (Fig. 3 and Supplemental Fig. S1, 317 A-D), which suggests independent evolution. The presence of E. coli in the agricultural soil and 318 freshwater biofilm may be related to the fertilization of this soil with manure and discharge of 319 wastewater, respectively, and we cannot exclude that the virulence genes detected in these 320 environments were related to clinical strains. Bacilli and Agrobacterium are natural 321 components of soil microbial communities and it is, therefore, not surprising that these 322 bacteria were found in soil. 323
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

15
Differences in the occurrence of the virulence genes were observed between the four 324 environments with the highest number being present in the beech wood soil [16], followed by 325 the roadside and organic agricultural soils [14 each], and the freshwater biofilm [11]. It was 326 unexpected that the organic agricultural soil did not have the highest number of virulence 327 determinants as fertilization with manure would be expected to have positively influenced the 328 number of bacteria possessing these genes (46). Poor survival of the bacteria is probably the 329 reason why that was not the case. The lowest number of virulence genes was detected in the 330 freshwater biofilm. The presence of E. coli in this environment suggested the freshwater lake 331 to be influenced by human discharges and, as for the organic agricultural soil, a large number 332 of virulence genes were to be expected. It is difficult to assess if fewer pathogenic traits were 333 actually present in this environment, but the 2.5 × higher detection limit compared to the soil 334 environments is probably part of the reason for the relatively few observed genes. 335 A priori, we hypothesized that bacteria in outer non-host environments are exposed to 336 pressures selecting for survival traits that may confer virulence in host environments, and 337 that differences in the occurrence of virulence genes can be related to different environmental 338 pressures. The widespread distribution of the virulence determinants in the soil and 339 freshwater environments provided circumstantial evidence that environmental pressures 340 indeed select for traits that potentially confer virulence in host environments. Further proof 341 of the hypothesis was the high degree of homology (90 – 100%) between the environmental 342 and clinical virulence gene sequences and the fact that the large majority of the environmental 343 sequences clustered together with the clinical sequences (Fig. 3 and Supplemental Fig. S1, A-344 D). This high degree of sequence conservation suggests that the virulence factors are 345 housekeeping genes, essential for the survival and proliferation of the bacterial cells not only 346 in human host environments but also in outer environments. With respect to the second part 347
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

16
of the hypothesis, that differences in the occurrence of virulence genes can be related to 348 different environmental pressures, the principal component analysis indicated that about half 349 of the variation (46%) in the distribution of the virulence traits could be explained by 350 differences in PAH concentration (PC1) and pH (PC2) (Fig. 2). pH is known to affect microbial 351 communities by influencing the carbon and nutrient availability and by controlling the 352 biomass composition of fungi and bacteria (47). The mechanism behind the apparent effect of 353 pH on the presence of the virulence genes is, however, not evident. The rationale for including 354 contaminated roadside soil in the study was that “accessory” genes involved in e.g. 355 pathogenicity, antibiotic resistance, or degradation of xenobiotics are often clustered together 356 on mobile genetic elements such as plasmids and pathogenicity islands (PAIs) (48). Therefore, 357 we expected the added stresses in the PAH-contaminated roadside soil would have led to 358 selective conditions that promoted the existence of mobile genetic elements containing 359 virulence factors. Our data, however, did not provide evidence that this was the case. Firstly, 360 the high PAH concentration in the roadside soil did not result in more virulence traits 361 compared to the other soils and secondly, although plasmids were present in the roadside soil 362 their numbers did not seem to be higher than in the beech wood and organic agricultural soils 363 (unpublished data). Thus, similar to pH, the PAH concentration appeared to influence the 364 distribution of the virulence determinants but the potential underlying mechanism remains 365 elusive. 366 The variation explained by the 2 principal components was relatively low (Fig. 2). This 367 suggests that factors other than pH and PAH concentration affected the occurrence of 368 virulence genes in the four environments. Bacteria have developed different defence 369 mechanisms against predation including secretion of toxins, avoidance of lysozymal killing, 370 and intracellular multiplication. Virulence factors of Legionella, Mycobacterium, Pseudomonas, 371
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

17
and Vibrio have for instance been found to be important for the interaction with invertebrates 372 and mammalian hosts including humans (49). Therefore, another possible factor controlling 373 the presence of the virulence factors could be the interaction with bacterial predators such as 374 free-living protozoa and nematodes. 375 In conclusion, we demonstrated the occurrence of numerous bacterial human virulence 376 genes in soil and freshwater environments. A high degree of sequence conservation between 377 the clinical and environmental genes sequences was observed. Altogether, the data suggest 378 that outer environments constitute reservoirs of several groups of bacterial virulence 379 determinants that possibly play an essential role for the survival and adaptation of the 380 bacteria. Further studies are needed to investigate to which extent the virulence genes are 381 expressed by the environmental bacterial communities. 382
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

18
ACKNOWLEDGMENTS 383 The work was supported by a grant (645-08-0110) from The Danish Agency for Science, 384 Technology and Innovation to The Postgraduate School of Environmental Chemistry and 385 Toxicology (RECETO). Jan H. Christensen is thanked for valuable advice on the principal 386 component analysis. 387
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

19
REFERENCES 388 1. Wu, H. J., A. H. J. Wang, and M. P. Jennings. 2008. Discovery of virulence factors of pathogenic 389 bacteria. Curr. Opin. Chem. Biol. 12:93-101. 390 2. Waterfield, N. R., B. W. Wren, and R. H. ffrench-Constant. 2004. Invertebrates as a source of 391 emerging human pathogens. Nat. Rev. Microbiol. 2:833-841. 392 3. Yildiz, F. H. 2007. Processes controlling the transmission of bacterial pathogens in the 393 environment. Res. Microbiol. 158:195-202. 394 4. Wilson, B. A. and A. A. Salyers. 2003. Is the evolution of bacterial pathogens an out-of-body 395 experience? Trends Microbiol. 11:347-350. 396 5. Yang, X., H. Yang, G. Zhou, and G. P. Zhao. 2008. Infectious disease in the genomic era. Annu. 397 Rev. Genomics Hum. Genet. 9:21-48. 398 6. Persson, O. P., J. Pinhassi, L. Riemann, B. I. Marklund, M. Rhen, S. Normark, J. M. Gonzalez, 399 and A. Hagstrom. 2009. High abundance of virulence gene homologues in marine bacteria. 400 Environ. Microbiol. 11:1348-1357. 401 7. Casadevall, A. and L. A. Pirofski. 2000. Host-pathogen interactions: Basic concepts of 402 microbial commensalism, colonization, infection, and disease. Infect. Immun. 68:6511-6518. 403 8. Martinez, J. L. 2013. Bacterial pathogens: from natural ecosystems to human hosts. Environ. 404 Microbiol. 15:325–333. 405 9. Martinez, J. L. 2006. Role of Non-clinial Environments in the Selection of Virulence and 406 Antibiotic Resistance Determinants in Pathogenic bacteria. J. Biol. Sci. 6:1-8. 407 10. Casadevall, A. 2006. Cards of virulence and the global virulome for humans . Microbe 1:359-408 364. 409 11. Casadevall, A. 2008. Evolution of intracellular pathogens. Annu. Rev. Microbiol. 62:19-33. 410 12. Kirn, T. J., B. A. Jude, and R. K. Taylor. 2005. A colonization factor links Vibrio cholerae 411 environmental survival and human infection. Nature 438:863-866. 412 13. Brassinga, A. K., J. M. Kinchen, M. E. Cupp, S. R. Day, P. S. Hoffman, and C. D. Sifri. 2010. 413 Caenorhabditis is a metazoan host for Legionella. Cell Microbiol. 12:343-361. 414 14. Carruthers, M. D., B. H. Bellaire, and F. C. Minion. 2010. Exploring the response of 415 Escherichia coli O157:H7 EDL933 within Acanthamoeba castellanii by genome-wide 416 transcriptional profiling. FEMS Microbiol. Lett. 312:15-23. 417 15. Lainhart, W., G. Stolfa, and G. B. Koudelka. 2009. Shiga toxin as a bacterial defense against a 418 eukaryotic predator, Tetrahymena thermophila. J. Bacteriol. 191:5116-5122. 419 16. Lelong, E., A. Marchetti, M. Simon, J. L. Burns, D. C. van, T. Kohler, and P. Cosson. 2011. 420 Evolution of Pseudomonas aeruginosa virulence in infected patients revealed in a Dictyostelium 421 discoideum host model. Clin. Microbiol. Infect. 17:1415-1420. 422
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

20
17. Pushkareva, V. I. and S. A. Ermolaeva. 2010. Listeria monocytogenes virulence factor 423 Listeriolysin O favors bacterial growth in co-culture with the ciliate Tetrahymena pyriformis, 424 causes protozoan encystment and promotes bacterial survival inside cysts. BMC Microbiol. 425 10:26. 426 18. Rahme, L. G., F. M. Ausubel, H. Cao, E. Drenkard, B. C. Goumnerov, G. W. Lau, S. Mahajan-427 Miklos, J. Plotnikova, M. W. Tan, J. Tsongalis, C. L. Walendziewicz, and R. G. Tompkins. 428 2000. Plants and animals share functionally common bacterial virulence factors. Proc. Natl. 429 Acad. Sci. U.S.A 97:8815-8821. 430 19. Casas, V., J. Miyake, H. Balsley, J. Roark, S. Telles, S. Leeds, I. Zurita, M. Breitbart, D. 431 Bartlett, F. Azam, and F. Rohwer. 2006. Widespread occurrence of phage-encoded exotoxin 432 genes in terrestrial and aquatic environments in Southern California. FEMS Microbiol. Lett. 433 261:141-149. 434 20. Ivanova, N., A. Sorokin, I. Anderson, N. Galleron, B. Candelon, V. Kapatral, A. 435 Bhattacharyya, G. Reznik, N. Mikhailova, A. Lapidus, L. Chu, M. Mazur, E. Goltsman, N. 436 Larsen, M. D'Souza, T. Walunas, Y. Grechkin, G. Pusch, R. Haselkorn, M. Fonstein, S. D. 437 Ehrlich, R. Overbeek, and N. Kyrpides. 2003. Genome sequence of Bacillus cereus and 438 comparative analysis with Bacillus anthracis. Nature 423:87-91. 439 21. Garcia-Aljaro, C., E. Moreno, A. Andreu, G. Prats, and A. R. Blanch. 2009. Phylogroups, 440 virulence determinants and antimicrobial resistance in stx(2) gene-carrying Escherichia coli 441 isolated from aquatic environments. Res. Microbiol. 160:585-591. 442 22. Vezzulli, L., C. Pruzzo, A. Huq, and R. R. Colwell. 2010. Environmental reservoirs of Vibrio 443 cholerae and their role in cholera. Environ. Microbiol. Rep. 2:27-33. 444 23. Stauder, M., A. Huq, E. Pezzati, C. J. Grim, P. Ramoino, L. Pane, R. R. Colwell, C. Pruzzo, and 445 L. Vezzulli. 2012. Role of GbpA protein, an important virulence-related colonization factor, for 446 Vibrio cholerae's survival in the aquatic environment. Environ. Microbiol. Rep. 4:439-445. 447 24. Danish Standard. 1980. Determination of total residue and total fixed residue in water, sludge 448 and sediment. Danish Standard (DS 204:1980) 4 pp. 449 25. Hollender, J., B. Koch, C. Lutermann, and W. Dott. 2002. Efficiency of Different Methods and 450 Solvents for the Extraction of Polycyclic Aromatic Hydrocarbons from Soils. Int. J. Environ. 451 Anal. Chem. 83:21-32. 452 26. Kroer, N. 1993. Bacterial growth efficiency on natural dissolved organic matter. Limnol. 453 Oceanogr. 38:1282-1290. 454 27. Rasmussen, L. D., C. Zawadsky, S. J. Binnerup, G. Oregaard, S. J. Sorensen, and N. Kroer. 455 2008. Cultivation of hard-to-culture subsurface mercury-resistant bacteria and discovery of 456 new merA gene sequences. Appl. Environ. Microbiol. 74:3795-3803. 457 28. Rozen, S. and H. Skaletsky. 2000. Primer3 on the WWW for general users and for biologist 458 programmers. Methods Mol. Biol. 132:365-386. 459 29. Benson, D. A., I. Karsch-Mizrachi, K. Clark, D. J. Lipman, J. Ostell, and E. W. Sayers. 2012. 460 GenBank. Nucleic Acids Res. 40:D48-D53. 461
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

21
30. Sayers, E. W., T. Barrett, D. A. Benson, E. Bolton, S. H. Bryant, K. Canese, V. Chetvernin, D. 462 M. Church, M. Dicuccio, S. Federhen, M. Feolo, I. M. Fingerman, L. Y. Geer, W. Helmberg, Y. 463 Kapustin, S. Krasnov, D. Landsman, D. J. Lipman, Z. Lu, T. L. Madden, T. Madej, D. R. 464 Maglott, A. Marchler-Bauer, V. Miller, I. Karsch-Mizrachi, J. Ostell, A. Panchenko, L. Phan, 465 K. D. Pruitt, G. D. Schuler, E. Sequeira, S. T. Sherry, M. Shumway, K. Sirotkin, D. Slotta, A. 466 Souvorov, G. Starchenko, T. A. Tatusova, L. Wagner, Y. Wang, W. J. Wilbur, E. Yaschenko, 467 and J. Ye. 2012. Database resources of the National Center for Biotechnology Information. 468 Nucleic Acids Res. 40:D13-D25. 469 31. Hansen, B. M. and N. B. Hendriksen. 2001. Detection of enterotoxic Bacillus cereus and 470 Bacillus thuringiensis strains by PCR analysis. Appl. Environ. Microbiol. 67:185-189. 471 32. Tamura, K., D. Peterson, N. Peterson, G. Stecher, M. Nei, and S. Kumar. 2011. MEGA5: 472 molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, 473 and maximum parsimony methods. Mol. Biol. Evol. 28:2731-2739. 474 33. Braid, M. D., L. M. Daniels, and C. L. Kitts. 2003. Removal of PCR inhibitors from soil DNA by 475 chemical flocculation. J. Microbiol. Methods 52:389-393. 476 34. Agerso, Y., G. Sengelov, and L. B. Jensen. 2004. Development of a rapid method for direct 477 detection of tet(M) genes in soil from Danish farmland. Environ. Int. 30:117-122. 478 35. Okano, Y., K. R. Hristova, C. M. Leutenegger, L. E. Jackson, R. F. Denison, B. Gebreyesus, D. 479 Lebauer, and K. M. Scow. 2004. Application of real-time PCR to study effects of ammonium on 480 population size of ammonia-oxidizing bacteria in soil. Appl. Environ. Microbiol. 70:1008-1016. 481 36. Lopez-Gutierrez, J. C., S. Henry, S. Hallet, F. Martin-Laurent, G. Catroux, and L. Philippot. 482 2004. Quantification of a novel group of nitrate-reducing bacteria in the environment by real-483 time PCR. J. Microbiol. Methods 57:399-407. 484 37. Borst, A., A. T. Box, and A. C. Fluit. 2004. False-positive results and contamination in nucleic 485 acid amplification assays: suggestions for a prevent and destroy strategy. Eur. J. Clin. Microbiol. 486 Infect. Dis. 23:289-299. 487 38. Sachse, K. 2004. Specificity and performance of PCR detection assays for microbial pathogens. 488 Mol. Biotechnol. 26:61-80. 489 39. Beutin, L. 2006. Emerging enterohaemorrhagic Escherichia coli, causes and effects of the rise 490 of a human pathogen. J. Vet. Med. B. Infect. Dis. Vet. Public Health 53:299-305. 491 40. Chauret, C. 2011. Survival and control of Escherichia coli O157:H7 in foods, beverages, soil and 492 water. Virulence 2:593-601. 493 41. Mueller-Spitz, S. R., L. B. Stewart, J. V. Klump, and S. L. McLellan. 2010. Freshwater 494 suspended sediments and sewage are reservoirs for enterotoxin-positive Clostridium 495 perfringens. Appl. Environ. Microbiol. 76:5556-5562. 496 42. Guan, T. Y. and R. A. Holley. 2003. Pathogen survival in swine manure environments and 497 transmission of human enteric illness - a review. J. Environ. Qual. 32:383-392. 498 43. Maugeri, T. L., M. Carbone, M. T. Fera, G. P. Irrera, and C. Gugliandolo. 2004. Distribution of 499 potentially pathogenic bacteria as free living and plankton associated in a marine coastal zone. 500 J. Appl. Microbiol. 97:354-361. 501
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

22
44. Miller, W. A., M. A. Miller, I. A. Gardner, E. R. Atwill, B. A. Byrne, S. Jang, M. Harris, J. Ames, 502 D. Jessup, D. Paradies, K. Worcester, A. Melli, and P. A. Conrad. 2006. Salmonella spp., 503 Vibrio spp., Clostridium perfringens, and Plesiomonas shigelloides in marine and freshwater 504 invertebrates from coastal California ecosystems. Microb. Ecol. 52:198-206. 505 45. Carbone, M., T. L. Maugeri, C. Gugliandolo, C. E. La, C. Biondo, and M. T. Fera. 2005. 506 Occurrence of Helicobacter pylori DNA in the coastal environment of southern Italy (Straits of 507 Messina). J. Appl. Microbiol. 98:768-774. 508 46. Durso, L. M., G. P. Harhay, J. L. Bono, and T. P. Smith. 2011. Virulence-associated and 509 antibiotic resistance genes of microbial populations in cattle feces analyzed using a 510 metagenomic approach. J. Microbiol. Methods 84:278-282. 511 47. Rousk, J., P. C. Brookes, and E. Baath. 2009. Contrasting soil pH effects on fungal and 512 bacterial growth suggest functional redundancy in carbon mineralization. Appl. Environ. 513 Microbiol. 75:1589-1596. 514 48. Hacker, J. and J. B. Kaper. 2000. Pathogenicity islands and the evolution of microbes. Annu. 515 Rev. Microbiol 54:641-679. 516 49. Hilbi, H., S. S. Weber, C. Ragaz, Y. Nyfeler, and S. Urwyler. 2007. Environmental predators as 517 models for bacterial pathogenesis. Environ. Microbiol. 9:563-575. 518 519 520
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

23
TABLE 1 Targeted virulence genes 521
Virulence group Virulence factor Gene Bacterial strain known to carry target gene
Reference
Toxin B component of hemolysin BL
hblA Bacillus thuringiensis serovar tenebrionis
DSM 5526
Shiga-like toxin 1 stx-1 Escherichia coli O157 SSIa C45-89 Cytotoxin K2 cytK2 Bacillus thuringiensis serovar
kurstaki NRLL HD1
Shiga-like toxin 2 stx-2 Escherichia coli O157 SSIa C45-89 Vacuolating cytotoxin vacA Helicobacter pylori NCTC 11638 DSM 10242 Hemolysin A hlyA Escherichia coli CFT073 ATCC 700928 Cytotoxin K1 cytK1 Bacillus cereus NVH 391-98 DSM 22905T Edema factor cya Bacillus anthracis 2160 SSIa 2160 Adhesin P fimbriae papC Escherichia coli CFT073 ATCC 700928 papAH - - Type 1 fimbriae fimH - - F1C fimbriae focG - - Secretion system Type III, secretion
apparatus
invA Salmonella enterica subsp. enterica serovar Paratyphi A
ATCC 9150
Type III, secretion apparatus
spiA - -
Type IV (conjugative DNA transfer)
virD4 Agrobacterium tumefaciens ATCC 33970
Type III, translocators and effectors
sipB-sipC Salmonella enterica subsp. enterica serovar Paratyphi A
ATCC 9150
Regulator of virulence Regulator of virulence determinants
phoQ Salmonella enterica subsp. enterica serovar Paratyphi A
ATCC 9150
Inflammatory mediator Part of O-antigen 111 wbdI Escherichia coli O111:H25 SSIa C830-04 Part of O-antigen 157 rfbE Escherichia coli O157 SSIa C45-89 Bacterial resistance L-Ara4N transferase
arnT
Salmonella enterica subsp. enterica serovar Paratyphi A
ATCC 9150
L-Ara4N transferase yfbI Escherichia coli CFT073 ATCC 700928 Capsule biosynthesis
protein capA Bacillus anthracis 2160 SSIa 2160
a SSI-strains derived by Statens Serum Institute, 522 http://www.ssi.dk/Bestil/SSI%20Diagnostica/Produkter%20fra%20SSI%20Diagnostica/Bakteriesta523 mmer.aspx524
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

TABLE 2 Characteristics of soils and biofilm 525
Beech wood soil Roadside soil Organic
agricultural soil Freshwater
biofilm pH 5.3 7.3 7.0 7.4a < 2 µm 14.2 13.8 16.4 - 2 – 50 µm 25.9 21.2 23.2 -Texture % 50 – 250 µm 38.2 37.2 31.3 - 250 – 2000 µm 20.7 19.1 17.5 - > 2000 µm 0.9 7.5 11.7 -Total nitrogen % 0.27 0.36 0.23 -Phosphorus µg g-1 0.89 4.8 2.3 -DOC (watera) ppm - - - 11.1 ± 0.6Loss on ignition % 8.0 ± 0.3 8.1 ± 0.3 6.1 ± 1.7 -PAH, total µg kg-1 DW 94.1 ± 5.4 2537.4± 555.2 22.5 ± 4.3 -Water content % 26.4 ± 3.9 24.1 ± 1.7 21.1 ± 0.9 -WHC % 33.2 ± 2.6 32.4 ± 3.9 28.7 ± 1.4 -Number of bacteria × 109 g-1 2.1 ± 0.3 2.2 ± 0.2 2.2 ± 0.2 -Polycyclic aromatic hydrocarbons (PAHs), water holding capacity (WHC), dissolved organic carbon (DOC), dry weight (DW). 526 a Measured in water samples collected at the same time and location as the biofilm. 527
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from
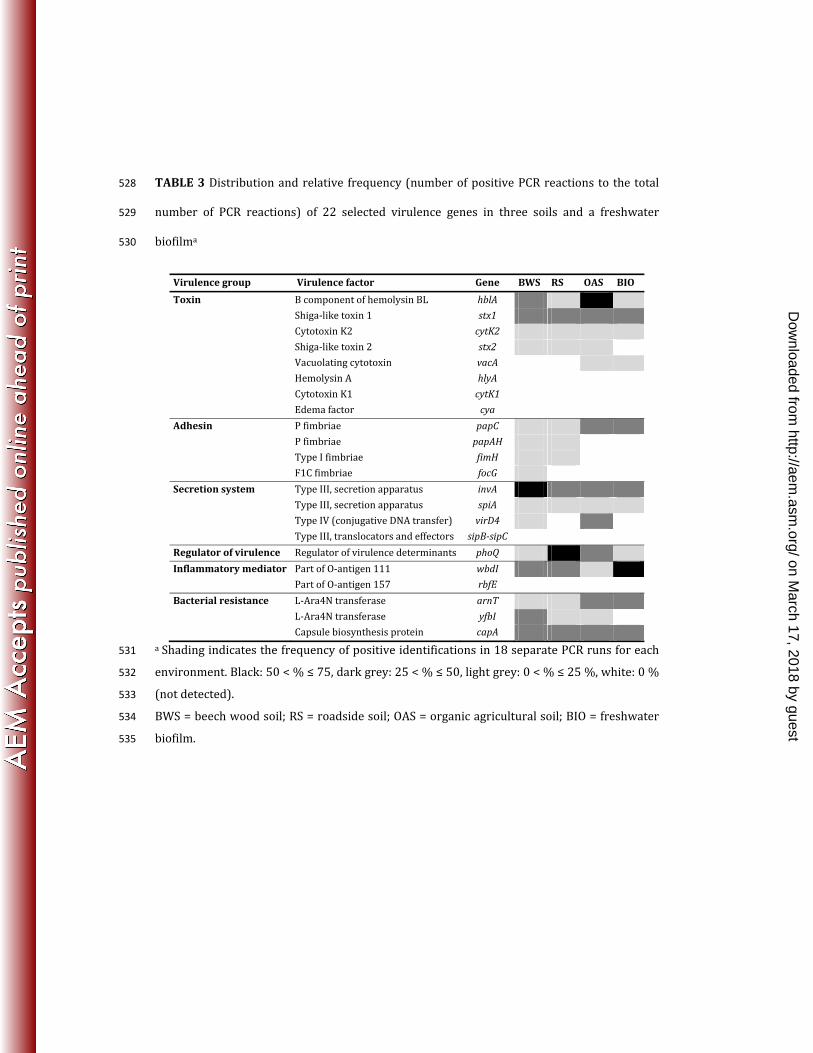
TABLE 3 Distribution and relative frequency (number of positive PCR reactions to the total 528 number of PCR reactions) of 22 selected virulence genes in three soils and a freshwater 529 biofilma 530
Virulence group Virulence factor Gene BWS RS OAS BIO
Toxin B component of hemolysin BL hblA Shiga-like toxin 1 stx1 Cytotoxin K2 cytK2 Shiga-like toxin 2 stx2 Vacuolating cytotoxin vacA Hemolysin A hlyA Cytotoxin K1 cytK1 Edema factor cya Adhesin P fimbriae papC P fimbriae papAH Type I fimbriae fimH F1C fimbriae focG Secretion system Type III, secretion apparatus invA Type III, secretion apparatus spiA Type IV (conjugative DNA transfer) virD4 Type III, translocators and effectors sipB-sipC Regulator of virulence Regulator of virulence determinants phoQ Inflammatory mediator Part of O-antigen 111 wbdI Part of O-antigen 157 rbfE Bacterial resistance L-Ara4N transferase arnT L-Ara4N transferase yfbI Capsule biosynthesis protein capA a Shading indicates the frequency of positive identifications in 18 separate PCR runs for each 531 environment. Black: 50 < % ≤ 75, dark grey: 25 < % ≤ 50, light grey: 0 < % ≤ 25 %, white: 0 % 532 (not detected). 533 BWS = beech wood soil; RS = roadside soil; OAS = organic agricultural soil; BIO = freshwater 534 biofilm. 535
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

26
FIGURE LEGENDS 536
FIG 1 Detection of the E. coli adhesion gene fimH in beech wood soil (BWS) (A) and organic 537 agricultural soil (OAS) (B), and estimation of the detection limit of fimH. fimH was targeted by 538 two parallel nested PCRs of triplicate DNA extractions of triplicate soil samples (i.e. 18 PCRs in 539 total per soil type). A: fimH was detected in 2 of 3 replicate beech wood soil samples and in 4 540 out of the total of 18 PCRs. B: fimH was not detected in any of the replicate soils samples of the 541 organic agricultural soil. The detection limit of fimH was estimated by adding a known 542 number of E. coli CFT073 carrying fimH to soil subsamples before DNA extraction. The 543 detection limit was estimated to be 400 bacteria g-1 soil. M is the molecular weight marker V 544 (pBR322-Hae III). 545
FIG 2 Principal component analysis based on the occurrence and relative abundance of the 546 virulence determinants in beech wood soil (BWS), roadside soil (RS), organic agricultural soil 547 (OAS), and freshwater biofilm (BIO). Percentages in parentheses on the axes indicate the 548 variation explained by the 2 principal components. Small sized data points show the scores of 549 replicate samples. Large data points represent the average of the scores of the replicate 550 samples of each environment. Error bars indicate standard errors of the mean. 551
FIG 3 Minimum evolution tree (unrooted) showing the phylogeny of the environmental invA 552 sequences (black triangles). Bootstrap values of 6 - 98. Numbers in parentheses indicate 553 numbers of identical invA sequences. The bacterial strain used as positive control is shown in 554 bold. 555
556
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

27
557
FIG 1 Detection of the E. coli adhesion gene fimH in beech wood soil (BWS) (A) and organic 558 agricultural soil (OAS) (B), and estimation of the detection limit of fimH. fimH was targeted by 559 two parallel nested PCRs of triplicate DNA extractions of triplicate soil samples (i.e. 18 PCRs in 560 total per soil type). A: fimH was detected in 2 of 3 replicate beech wood soil samples and in 4 561 out of the total of 18 PCRs. B: fimH was not detected in any of the replicate soils samples of the 562 organic agricultural soil. The detection limit of fimH was estimated by adding a known 563 number of E. coli CFT073 carrying fimH to soil subsamples before DNA extraction. The 564 detection limit was estimated to be 400 bacteria g-1 soil. M is the molecular weight marker V 565 (pBR322-Hae III). 566
567
B
A
E.c
oliC
FT0
73
M
DN
A1
DN
A2
DN
A3
DN
A2
DN
A1
DN
A3
DN
A1
DN
A2
DN
A3
E.c
oliC
FT0
73
OA
S1
OA
S3
OA
S2
Ne
g.co
ntr
olE
.col
iCF
T073
0E
.col
iCF
T073
/g
OA
S
40E
.col
iCF
T073
/g
OA
S
400
E.c
oliC
FT
073
/gO
AS
4,0
00E
.col
iCF
T07
3/
gO
AS
40,0
00
E.c
oliC
FT0
73/g
OA
S
Ne
g.co
ntr
ol
M M M
238 bp
238 bp
E.c
oliC
FT0
73
M
DN
A1
DN
A2
DN
A3
DN
A2
DN
A1
DN
A3
DN
A1
DN
A2
DN
A3
E.c
oliC
FT0
73
BW
S1
BW
S3
BW
S2
Neg
.con
trol
E.c
oliC
FT0
730
E.c
oliC
FT
073
/g
BW
S
40
E.c
oliC
FT
073
/g
BW
S
400
E.c
oliC
FT0
73/g
BW
S4
,00
0E
.col
iCF
T07
3/g
BW
S4
0,0
00E
.col
iCF
T073
/g
BW
S
Neg
.con
trol
M M M
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from
568
FIG 2 Principal component analysis based on the occurrence and relative abundance of the 569 virulence determinants in beech wood soil (BWS), roadside soil (RS), organic agricultural soil 570 (OAS), and freshwater biofilm (BIO). Percentages in parentheses on the axes indicate the 571 variation explained by the 2 principal components. Small sized data points show the scores of 572 replicate samples. Large data points represent the average of the scores of the replicate 573 samples of each environment. Error bars indicate standard errors of the mean. 574
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

29
575
FIG 3 Minimum evolution tree (unrooted) showing the phylogeny of the environmental invA 576 sequences (black triangles). Bootstrap values of 6 - 98. Numbers in parentheses indicate 577 numbers of identical invA sequences. The bacterial strain used as positive control is shown in 578 bold. 579
invA RS 1C.3
invA RS 3B.3
invA BIO 2A.3
invA RS 3A.3
EU311614.1 Salmonella enterica subsp. enterica serovar Abony
JN982041.1 Salmonella enterica subsp. enterica serovar Typhimurium OEHST5
CP000026.1 Salmonella enterica subsp. enterica serovar Paratyphi A ATCC 9150
DQ644627.1 Salmonella enterica subsp. houtenae ST-22
CP001127.1 Salmonella enterica subsp. enterica serovar Schwarzengrund CVM19633
FJ496648.1 Salmonella enterica subsp. salamae serovar Sofia
invA BWS 1B
EU348369.1 Salmonella enterica subsp. enterica serovar Senftenberg JXS-04*01
AM933172.1 Salmonella enterica subsp. enterica serovar Enteritidis P125109
invA BWS 3B.3 (11); invA OAS 3A.3 (5); invA RS 2C.3 (4); invA BIO 2A (6)
CP003416.1 Salmonella enterica subsp. enterica serovar Heidelberg B182
AM933173.1 Salmonella enterica subsp. enterica serovar Gallinarum 287/91
invA BWS 3C.3
EU348368.1 Salmonella enterica subsp. enterica serovar Pullorum 1794
CP003278.1 Salmonella enterica subsp. enterica serovar Typhi P-stx-12
CP001144.1 Salmonella enterica subsp. enterica serovar Dublin CT 02021853
CP000886.1 Salmonella enterica subsp. enterica serovar Paratyphi B SPB7
DQ644618.1 Salmonella enterica subsp. salamae CNM-169
DQ644631.1 Salmonella enterica subsp. indica CDC-1937
DQ644626.1 Salmonella enterica subsp. houtenae CNM-2556-03
CP003386.1 Salmonella enterica subsp. enterica serovar Typhimurium 798
CP001138.1 Salmonella enterica subsp. enterica serovar Agona SL483
CP001113.1 Salmonella enterica subsp. enterica serovar Newport SL254
EU348366.1 Salmonella enterica subsp. enterica serovar Gallinarum S9873
DQ644633.1 Salmonella bongori CNM-256
JN680859.1 Salmonella enterica subsp. enterica serovar Typhimurium EHO22-TU
JF951189.1 Salmonella enterica subsp. enterica serovar Paratyphi C XY25
GU183866.1 Salmonella sp. enrichment culture NEP50
DQ644617.1 Salmonella enterica subsp. salamae CNM-176
FR775234.1 Salmonella enterica subsp. enterica serovar Weltevreden 2007-60-3289-1
CP000857.1 Salmonella enterica subsp. enterica serovar Paratyphi C RKS4594
AE017220.1 Salmonella enterica subsp. enterica serovar Choleraesuis SC-B67
CP000880.1 Salmonella enterica subsp. arizonae serovar 62:z4z23:--
AY594271.1 Salmonella enterica group IIIa
DQ644625.1 Salmonella enterica subsp. diarizonae CNM-2667-02
AY594272.1 Salmonella enterica group IIIb
FR877557.1 Salmonella bongori NCTC 12419
DQ644629.1 Salmonella enterica subsp. indica CNM-186
DQ644616.1 Salmonella enterica subsp. salamae CNM-5936-02
AY594273.1 Salmonella bongori group V
AY594274.1 Salmonella enterica group VI
AM942759.1 Proteus mirabilis HI4320
CU468135.1 Erwinia tasmaniensis ET1/99
FR719187.1 Erwinia amylovora ATCC BAA-2158
CP001277.1 Candidatus Hamiltonella defensa 5AT (Acyrthosiphon pisum)
CP003034.1 Escherichia coli O7:K1 CE10
FN554766.1 Escherichia coli 042
CU928163.2 Escherichia coli UMN026
EU179217.1 Escherichia coli EC10
AY864672.1 Escherichia coli 36-5
CP003109.1 Escherichia coli O55:H7 RM12579
CP001368.1 Escherichia coli O157:H7 TW14359
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from