water transport, and transport across epithelia · membrane biogenesis and transport lecture 16...
TRANSCRIPT

Dale Sanders
16 March 2009
Module 0220502
Membrane Biogenesis and Transport
Lecture 16
Water Transport,and Transport across Epithelia

Aims:By the end of the lecture you should
understand…
How water moves through bilayers by solubility inlipid and through aquaporins;
How nutrients move across epithelia;
•
How absorptive epithelia move water acrossthemselves;
•
How secretory epithelia move water acrossthemselves.
•

Reading
Lodish et al. (2008) Molecular Cell Biology 6th ed pp. 444-446& 470-473
A good introduction to aquaporins and epithelial transport
Fujiyoshi et al. (2002) Structure and function of water channels.Curr. Opin. Struct. Biol. 12: 509-515
Murata et al (2000) Structural determinants of water permeationthrough aquaporin-1. Nature 407: 599-605

Water Transport
Osmosis is rapid, yet membranes have hydrophobic interior.
How do we reconcile these observations??
H-bonding props of water allow significant solubility in hydrophobic
bilayer:
Passive permeabilities of solutes through lipid bilayers can be
measured in an artificial system. E.g. for water
3H2O
1H2OLipid bilayer
Teflon chamber
Measure radioactivity appearing on trans side

For water, diffusive, passive permeability measured with 3H2O(Pw) 2.10-5 cm.s –1 – not very different from value expected onbasis of oil:water partition coefficient
slope = 1 codeine
butyric acid
H2O “corrected” for high diffusioncoefficient (small molecule)
H2O
1,2 propanediol
1,4 butanediol
acetamide
glycerol
urea
1,2 ethanediol
formamide
log (diffusionalpermeabilitycoefficient)
log (oil:water partition coefficient)
inorganicions
Results3
2
1
-1
-1
3 4

For many biological membranes, Pw has the value observed inbilayers:
Conclude: solvation then diffusion of water through the lipid phase isthe primary mechanism of transport
HOWEVER in many other membranes,
water ALSO crosses through specific
WATER CHANNELS
Evidence 1. Physical/chemical
A 2nd way to measure water permeability:
impose an osmotic gradient
H2O
= solute molecule

If water diffusion through lipid, “osmotic permeability” for H2O (Lp)should = Pw
in some cells, it doesn’t e.g.
Cell type + Hg2+ Lp Pw
10-5 cm s-1
Red blood cell - 20 2.0
+ 1.8 1.8
Kidney prox. tubule - 40 2.0
+ 3.2 1.0
• >10x discrepancy between Lp and Pw
• Discrepancy largely abolished by Hg2+

Hypothesis to explain discrepancy
A single file, multiple-occupancy water channel:
Lp is NET flow: 1 molecule entering on left knocks out 1 moleculeon right.
Pw is unidirectional flow: 3H2O entering on left could diffuse backand has many others to pass in pore – slows down the passage ofradiotracer
H2O

2. Molecular/Biochemical evidence
AQUAPORINS: members of the Major IntrinsicProtein family
MIP: lens epithelium
TIP: plant tonoplast
CHIP: Channel-forming intrinsic protein (red bloodcells)
Mr = 28,000 – A homotetramer with each monomerforming a channel

Evidence that CHIP and TIP are H2Ochannels:
Inject mRNA into Xenopus oocytes and look atswelling rate in response to hypo-osmotic solutions
• Swelling response also blocked by Hg2+
1.6
1.4
1.2
1.0
0 1 2 3 4 5
Re
lative
vo
lum
e
t (min)
TIP
CHIP
Water control
in hypotonic solution
Chrispeels & Agre (1994) Trend. Biochem. Sci. 19: 421-425
Control CHIP-expressing
30 s 1 min
2 min 3 min

The Hourglass Model
NPA
NPA1 2 3 4 5 6
repeat 1 repeat 2
N C
Connecting loop B Connecting loop E
Connecting loops B & E dip into membrane fromopposite sides to form aqueous pathway
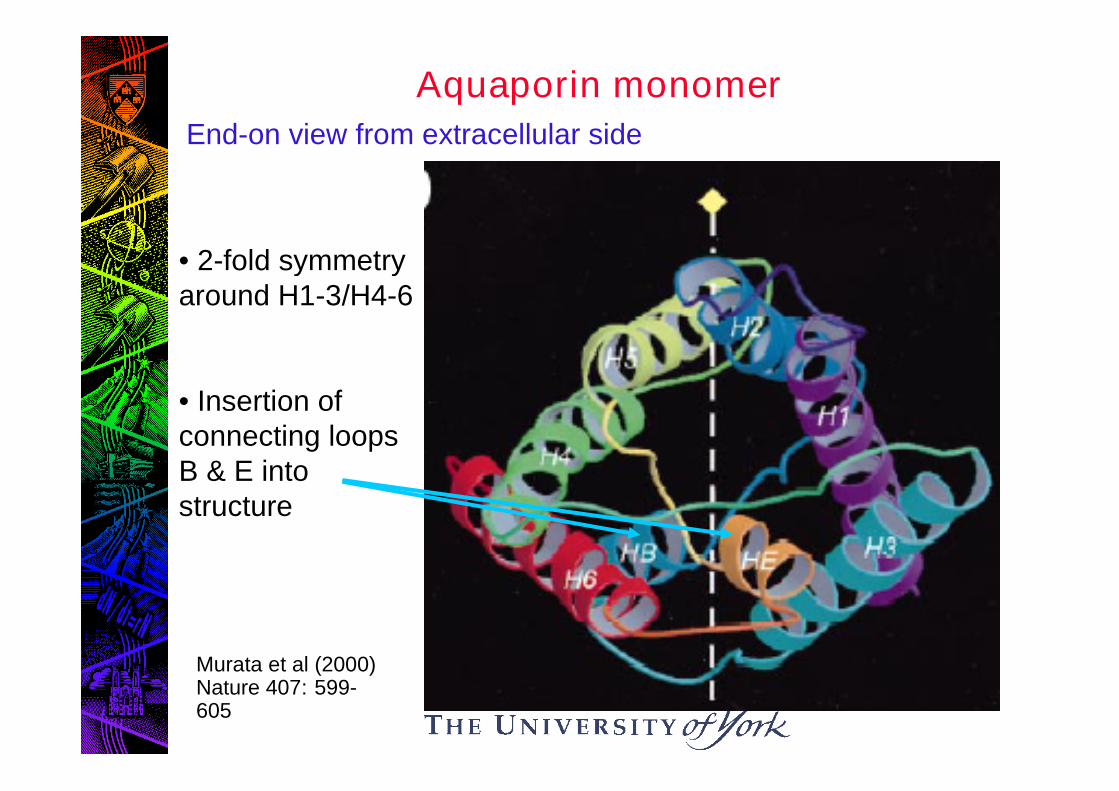
Aquaporin monomer
• 2-fold symmetryaround H1-3/H4-6
• Insertion ofconnecting loopsB & E intostructure
End-on view from extracellular side
Murata et al (2000)Nature 407: 599-605

Aquaporin monomer
Side view
Limits ofmembrane
Connectingloops B & Eenteringfromoppositesides
Murata et al (2000)Nature 407: 599-605

Space-filling models of thepore region of theaquaporin monomer
•Large number of hydrophobicresidues (yellow) line thepore
•Asn76 and Asn192 (red) are inthe NPA motifs: contributeto the tight constriction andH-bond with H2O
•Cys189 (green) is site of Hgbinding
Murata et al (2000)Nature 407: 599-605

Transport Across Epithelia
So far, we have considered cells as spatially uniform for transport:
A good model for most cells.
But an important exception:
Epithelial cells: asymmetrywith respect to transportbecause they must transportacross the tissue.
SS S
S
http://cellbio.utmb.edu/microanatomy/epithelia/00004493.jpg

Generalized representation of epithelial cells andsome definitions
Mucosal(lumenal)
side
Blood(serosal)
side
Intercellularspace
Brush border(apical) membrane
Basolateralmembrane
Junction(tight or leaky)

Epithelia can be broadly classified as either
1. Absorptive: Take up nutrients, salts, waterfrom mucosal side and deposit them onserosal side
2. Secretory: Transport water to mucosalside
Transepithelial transport involves co-ordinated activity of pumps, carriers andchannels.

1. Absorptive Epithelia
e.g. Small intestine, kidney tubules
(a) Solute transport, e.g: absorption of glucose in small intestine
S (BL) +M (BB)
M
phlorizin
–
cytochalasin
Na+
K+
glucose
ATPouabain
Intracellular
BB membrane BL membrane
SNa+
glucose
K+
RelativeelectrochemicalActivities:
Na+
glucose

A similar picture for other solutes eg amino acids
Evidence for this model:
1. Net transport of Na+ and glucose from M S impliescharge translocation:
A trans-epithelial potential difference of about -10 mV,mucosa negative, is measured…
Sensitive to inhibition of (Na+ + K+)-ATPase by ouabain
2. BB and BL membranes can be fractionated:
Na+ - coupled solute transport only at BB membrane,
ATPase activity only BL membrane
3. Selective use of inhibitors: they inhibit glucose transportin a side-specific manner.
collectingtubule
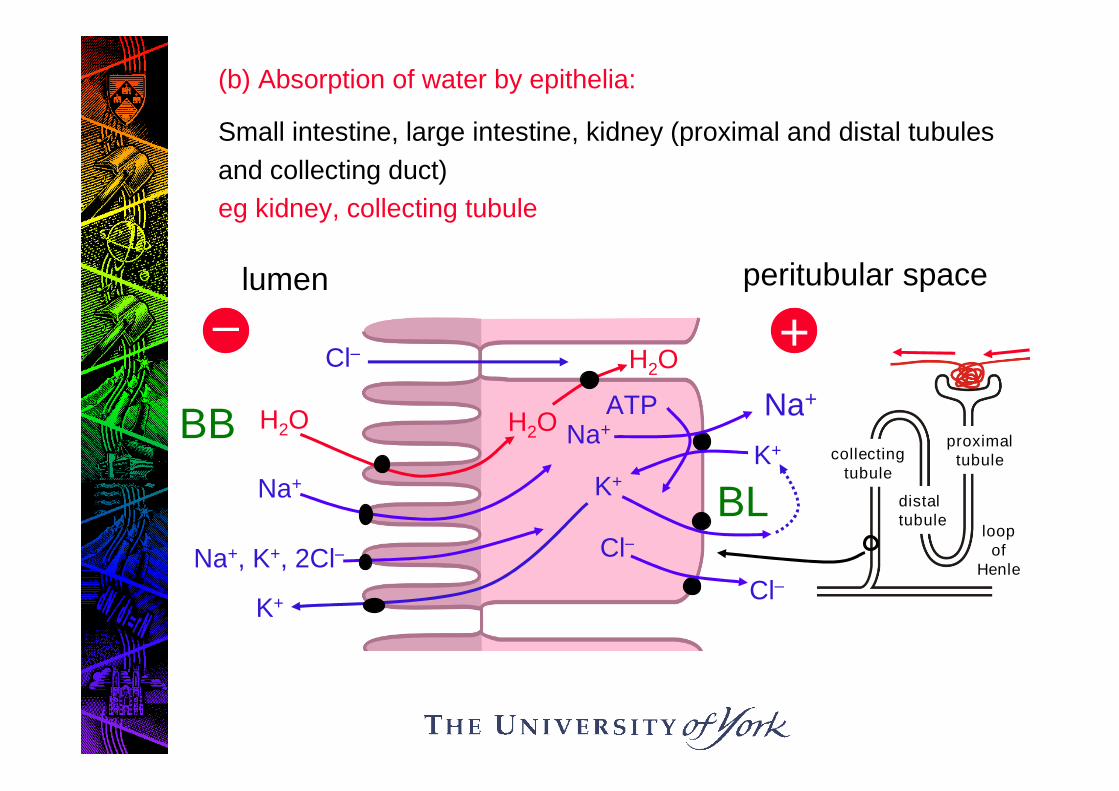
(b) Absorption of water by epithelia:
Small intestine, large intestine, kidney (proximal and distal tubules
and collecting duct)
eg kidney, collecting tubule
lumen
–
BB H2O
peritubular space
+
Na+
K+
ATP
Na+
Na+H2O
K+
K+
Cl–
Cl–
BL
H2O
Na+, K+, 2Cl–
Cl–
loopof
Henle
proximaltubule
distaltubule

1. (Na+ - K+)- ATPase generates electrochemical potentialdifference for Na+ across BL membrane
2. Na+ moves passively across BB membrane, and also drivesuptake of K+ and Cl- from lumen
3. K+ channels at both membranes allow recirculation of K+ andcontrol Δ.
4. Trans-epithelial Na+ transport sets up trans-epithelial Δ,which drives paracellular Cl- transport into cleft.
5. Cl- channels in BL membrane release Cl- on serosal side.
6. H2O absorbed from lumen in response to osmotic gradientgenerated by NaCl, especially in cleft.
Note that the whole mechanism of H2O transport driven ultimatelyby the (Na+/K+) – ATPase: sets up the ionic gradients.

2. Secretory Epitheliaeg salivary glands, sweat glands, lacrimal glands, exocrinepancreas, gastric mucosa, tracheal epithelium
serosalumen
–
H2O
Na+
Secretagogueseg ACh,catecholamines
cAMP
Na+
Ca2+
K+
Cl– Ca2+
BL
H2O
Na+, K+, 2Cl-
Cl–
ATP
BB
++
Ca2+-activated K+
channel maintainsintracellular Δnegative,sustaining thedriving force for Cl
-
release
CFTR

1. Secretagogues generate [Ca2+]i through opening of non-specific cation channels.
2. Ca2+ activates K+ channels at BL membrane causing K+
release.
3. K+ reabsorbed by Na+,K+,2Cl-
carrier at BL membrane, andby (Na+ + K+)-ATPase, which also removes incoming Na+.
4. Cl-release occurs through cAMP activated Cl
-channels at
BB membrane, resulting in formation of transepithelial Δ(lumen-negative)
5. Na+ moves paracellularly into lumen, driven by Δ.
6. H2O moves into lumen in response to osmotic gradient, setup by net movement of NaCI.

Summary
1. Water moves through bilayer membranes by H bonding inhydrophobic environment.
2. In some membranes, aquaporins greatly increase rate ofosmotically driven H20 flow.
3. Aquaporins are small (28 kDa) polypeptides with
6 t/m spans and NPA motifs on connecting loops
4. Solutes are moved across absorptive epithelia by Na+-coupling in BB membrane and non-coupled transporters(carriers) in BL membrane.
5. (Na++K+)-ATPase is exclusively in the BL membrane.
6. Flow of water across epithelia is driven by NaCI movement,with the membrane location of the Na+, K+, 2Cl
-carrier
determining direction of flow.