walsby's square bacterium: fine structure of...
TRANSCRIPT

Vol. 148, No. 1JOURNAL OF BACTERIOLOGY, Oct. 1981, p. 352-3600021-9193/81/100352-09$02.00/0
Walsby's Square Bacterium: Fine Structure of an OrthogonalProcaryote
WALTHER STOECKENIUS
Department of Biochemistry and Cardiovascular Research Institute, University ofCalifornia, San Francisco, California 94143
Received 28 April 1981/Accepted 16 June 1981
The "square" bacterium, first described by Walsby from brine collected at theRed Sea shore [A. E. Walsby, Nature (London) 283:69-71, 1980] was examinedby electron microscopy. The cells appeared as flat rectangular boxes in scanningelectron micrographs. In sections and freeze-fracture preparations, the edgeslooked more rounded. The thickness apparently remains constant as the cellsgrow and divide. Their sides were a few micrometers long, but the cells were only0.25 ,um thick. They showed typical procaryote structure, with a regular cell wall
and a gas vacuole fine structure similar to that of other halophilic procaryotes.The inner fracture faces of the cell membrane showed a much denser populationof intramembrane particles than the outer fracture faces, but no patches of purplemembrane, despite the presence of bacteriorhodospin-like pigment in the cellsuspension. Morphologically identical cells have been found in brine from BajaCalifornia, Mexico.
A new morphological type of procaryote hasrecently been described by A. E. Walsby (8).The organism was found in a natural salt pondof a sabkha bordering the western shore, Gulf ofEilat, between Nabq and Ophira and near thesouthern tip, Sinai peninsula. The site consistsof a roughly circular shallow depression about500 m in diameter which is separated from thesea by low sand dunes. Seawater seeps throughthe sand and collects in the depression, where itis concentrated by evaporation. We visited thesite in August 1980, when most of the depressionwas dry and only a crescent-shaped shallow poolremained at the northern end. It was 10 to 15 mwide, 150 m long, and not more than 20 to 50 cmdeep. Salt had crystallized in a thick crustaround the pool. Of 25 samples collected, 23contained the organism described by Walsby inaddition to other, mainly rod-shaped bacteriaand some unicellular algae with the morpholog-ical characteristics of Dunaliella sp.The "square" cells are flat rectangular boxes
with perfectly straight edges measuring a fewmicrometers on the side, and their height is nearthe limit of resolution of a light microscope. Thesmallest cells are always square, measuring 2 by2 ,um, whereas larger cells are often rectangular.They contain many gas vacuoles and also smallgranules which appear dark in the phase-con-trast image. Our observations essentially agreewith Walsby's description. He did not mentionthe dark granules, but a few can be seen in hispictures. They appear to be considerably more
abundant in our samples.Halophilic bacteria have recently attracted
increased interest mainly for two reasons. Theextreme halophiles belong to a group of procar-yotes so different from other bacteria that theyhave been classified as a new kingdom, the ar-chaebacteria (9), and some of them contain aphotosynthetic system not based on chlorophyll,but on bacteriorhodopsin, a chromoprotein re-sembling the visual pigments of animals (6).Since the new organism discovered by Walsby isobviously halophilic, we decided to study it moreclosely to determine whether it would showthese interesting features. We observed cellswith an electron microscope rather than a lightmicroscope to characterize them better morpho-logically. Attempts to grow the organism in thelaboratory are under way.
MATERIALS AND METHODSCells were sedimented by centrifugation of 6 liters
of brine. This yielded approximately 20 ml of cellsuspension (ca. 109 cells per ml) in which the squarebacterium was the dominant morphological species.The cells were fixed by the addition of a 40% formal-dehyde solution to a final concentration of 4% within3 days after collection. The cells remained in thissolution, and no visible change in their morphologywas seen. All further processing was carried out 3weeks later in San Francisco.The cells were prepared for sectioning and freeze-
fracture by standard techniques. No cryoprotectantwas used, and the freezing medium was Freon 22 (5).Before embedding and sectioning, cells were postfixed
352
on June 27, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

WALSBY'S SQUARE BACTERIUM STRUCTURE 353
in OS04 or in KMnO4 solution as described by Stoeck-enius and Rowen (7); no significant difference wasnoted in the effects of the two fixatives. For scanningelectron microscopy (SEM), the cells were allowed tosettle on a polylysine-coated cover slip, exposed to0804 vapor for 2 h, washed with distilled water, de-hydrated in ethanol, and after critical point drying,coated with gold. Preparations were viewed in a Sie-mens 101 or a Cambridge S 150 electron microscope.
RESULTSThe concentrated cell suspension obtained by
centrifugation had the reddish color character-istic of extreme halophiles and gave the typicalcarotenoid absorption spectrum. Flash spectros-copy showed a transient absorbance change in-dicative of the presence of bacteriorhodopsin (3)(Fig. 1). Light microscopy after centrifugationshowed that the gas vacuoles had collapsed, asexpected, and the dark granules were now muchmore prominent; no change in the shapes of thecells was detected (Fig. 2). Larger cells and es-pecially the frequently found sheets of cells wereoften bent so that only part of the cell was infocus when a high-power objective lens was usedand the plane of the cell was at a right angle tothe viewing direction. Scanning electron micros-copy confirmed the overall shapes of the cellsand the fact that they were the dominant mor-
300 400 500 600 700
Wavelength (nm)
FIG. 1. Absorption (A) spectrum of cells concen-
trated from Red Sea brine. The absorption maximaat 540, 505, and 475 nm and the shoulder at 450 nmare characteristic for carotenoids found in extremelyhalophilic bacteria. Insert, Difference spectrum forthe transient absorbance change 0.5 ms after anactinic flash at 500 nm. The minimum near 560 nmand maximum near 400 nm indicate the presence ofbacteriorhodopsin-like pigment.
FIG. 2. Phase-contrast micrographs of cells afterconcentration. The gas vacuoles were collapsed bythe centrifugation and are not visible. The dark gran-ules are prominent. In the lower micrograph, two ofthe square cells are seen in profile in the upper leftand lower right quadrant. Magnification, x1,600.
phological species (Fig. 3-5). It showed the cellsto be uniformly 0.25 ,um thick and to have amaz-ingly acute edges and comers. Bending aroundan axis parallel to the plane of the cells was evenmore pronounced than expected from light mi-croscopy and may have been enhanced by thepreparation technique (Fig. 3). The cells usuallyshowed a rather smooth surface. Where struc-ture was visible, it was irregular. There was littledifference from the background (Fig. 4). Thesheets of four or more adherent cells often seenin the light microscope were not observed in theelectron microscope. They apparently had notwithstood the preparation procedure. The sizeof the smallest cells was approximately 2.0 by2.0 by 0.25 ,um, but squares twice that size andrectangles with dimensions of 4.0 by 2.0 ,im withno cell wall septa were often seen (Fig. 5). Thethickness of the cells apparently remained con-stant. This suggests that the cells grow fromsmaller into larger squares of the same thicknessand then undergo two divisions in rapid succes-sion, with or without separation of the daughtercells after the first division. The larger sheets ofsquares seen in the light microscope may beexplained by assuming that the cells fail to sep-arate after the second division and then growand divide synchronously, or that division is
VOL. 148, 1981
74
on June 27, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
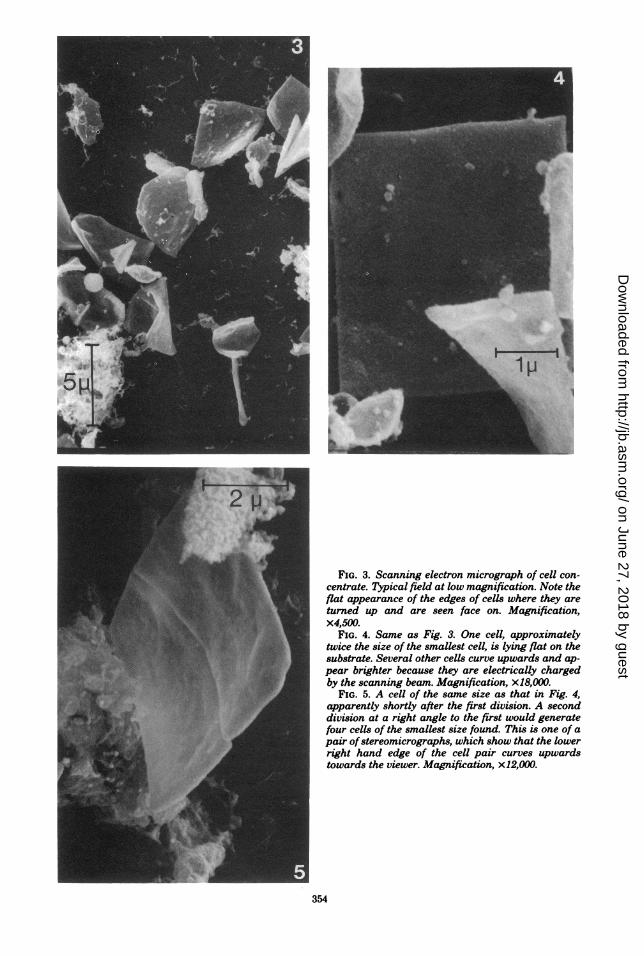
FIG. 3. Scanning electron micrograph of cell con-centrate. Typical field at low magnification. Note theflat appearance of the edges of cells where they areturned up and are seen face on. Magnification,x4,500.
FIG. 4. Same as Fig. 3. One cell, approximatelytwice the size of the smallest cell, is lying flat on thesubstrate. Several other cells curve upwards and ap-pear brighter because they are electrically chargedby the scanning beam. Magnification, x18,000.
FIG. 5. A cell of the same size as that in Fig. 4,apparently shortly after the first division. A seconddivision at a right angle to the first would generatefour cells of the smallest size found. This is one of apair ofstereomicrographs, which show that the lowerright hand edge of the cell pair curves upwardstowards the viewer. Magnification, x12,000.
354
on June 27, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

WALSBY'S SQUARE BACTERIUM STRUCTURE 355
delayed until a higher multiple of the standardcell size is reached. We have, however, failed tofind cells larger than 5 by 5 ,im in the scanningelectron microscope, and cell division lines maynot always be visible in the light microscope.
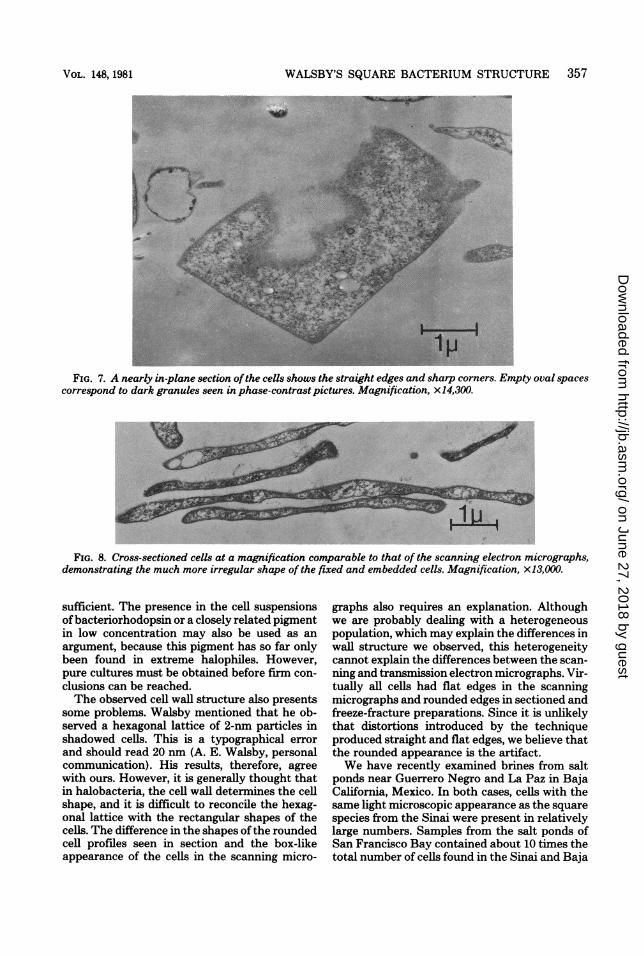
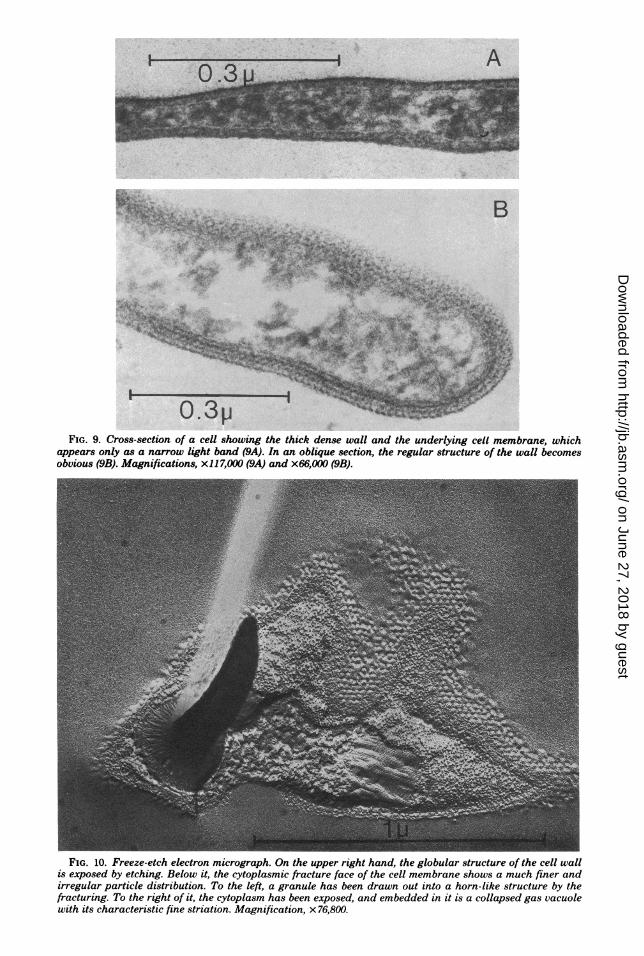
In the sectioned material, a given cell profilewas not always easy to identify. Sections in theplane of the square cells were necessarily rare,and if the cells were bent, they could not encom-pass a whole cell. Fortunately, the square cellsconstituted the majority of the material, and incross-section, they were easily recognized be-cause they were thinner than any of the othercells present. The long, slender profiles whichdominated in the sections must, therefore, be-long to the square cells (Fig. 6 and 8). A rare,nearly complete in-plane section is shown in Fig.7. The straight edges and sharp corners areobvious. The cytoplasm contains granules andfine strands of material, possibly ribosomes, andDNA similar to what is typically seen in otherprocaryotes, but rather less densely packed. Thedense granules seen in the light microscope haveapparently been dissolved and appear as ovalempty spaces up to 0.3 or 0.4 ,um in diameter. Incross-sections, their longer axis can be seen tolie in the plane of the cell. The collapsed gasvacuoles are also easily recognized because theyresemble unit membranes in cross-section butwith free ends (5, 7). Sections normal to theplane of the cells show their width to be lessuniform and their ends to be more rounded thanone would expect from the scanning electronmicroscopy images. We ascribe this to irregularshrinkage during embedding because the thick-ness of the cells was often considerably less thanthe rather constant 0.25 ,um seen in the scanningmicrographs, and the thinner the cells, the den-ser appeared the cytoplasm. The cells showed athick wall over the plasma membrane, whichappeared as a 3.0- to 4.0-nm-wide light line de-limiting the cytoplasm (Fig. 9A). The appear-ance of the wall varied considerably. It oftenshowed an inner, dense line and a broader bandof lighter material on the outside or vice versa.The width varied considerably from approxi-matley 15 to approximatley 25 nm in what ap-peared to be cross-sections. It clearly had aregular in-plane structure. Prominent periodici-ties of 20 nm were often seen in slightly obliquesections, and more oblique cuts gave rise tocomplex patterns (Fig. 9B). We made no at-tempts to analyze the patterns, because we sus-pected that the population of square cells con-sisted of more than one species and because allcells did not have the same wall structure.
Freeze-fracture preparations confirmed thegeneral shape of the cells and the presence of a
cell wall, cytoplasmic granules, and gas vacuoles.The varying thickness and rounded edges seenin cross fractures were similar to images seen insections and again suggested that some defor-mation had occurred. The granules were eitherbroken out of the fracture face, leaving round oroval depressions, or were drawn out into prom-inent, often bent, cone-shaped structures pro-truding from the fracture face (Fig. 10). Similardeformations have been shown to occur duringfreeze-fracture of latex spheres and a variety ofbiological structures, especially poly-fl-hydrox-ybutyrate granules in procaryotes (2, 4). In well-resolved replicas, the collapsed gas vacuolesshow the typical cross striation with a 2.0-nmperiodicity (5). In-plane fractures of the cellmembrane showed a dense population of parti-cles on the cytoplasmic leaflet and a much spar-ser particle distribution on the extemal leaflet,as do most cell membranes (Fig. 11). Eventhough flash spectroscopy indicated that bacte-riorhodopsin was present, we only very rarelyfound small, ordered arrays of intramembraneparticles, and they did not show the typicalappearance of the purple membrane patchesseen in other halobacteria (1). In slightly etchedpreparations, the cell wall was clearly visible andappeared approximately 15 nm wide. Its regularstructure was clearly seen in face view and con-sisted of round particles with a spacing of ap-proximately 23 nm (Fig. 11). The pattern ap-peared to be orthogonal or hexagonal, but etch-ing in the high salt concentration required isdifficult to obtain. We suspect that the cell pop-ulation is not homogeneous in morphology, butwe have not seen sufficient extended face viewsof the cell walls for an analysis. Fixation with0804 and subsequent washing with water didnot preserve the structure in heavy metal-shadowed preparations.
DISCUSSIONIf there were any lingering doubts left as to
whether or not the objects described by Walsbywere indeed bacterial cells, the fine structureobservations reported here should dispel them.Moreover, we have been able to grow the cellsin enrichment culture by adding peptone me-dium to the natural brine, but growth of purecolonies on agar has not been achieved so far.Their size, their lack of a nuclear membrane,their wall structure, and the occurrence of gasvacuoles clearly identify them as procaryotes.They are obviously halophilic; whether or notthey are extreme halophiles remains to be seen.Their apparently slow growth, red pigmentation,and regular cell wall structure are compatiblewith such a classification (5-7), but certainly not
VOL. 148, 1981
on June 27, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

A
FIG. 6. Transmission electron micrographs ofcross-sectioned cells. The cell shown in 6a is muchnarrower (approx. 0.1 Mim) and has a much densercytoplasm than the cell in 6b. (Note difference inmagnification.) The upper edge of the cell in 6b ap-pears squared off but the lower edge is rounded.Compared to the appearance in scanning electronmicrographs, the cells appear more irregular inshape. Magnifications, x54,000 (6a) and x28,600 (6b).
356
on June 27, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
WALSBY'S SQUARE BACTERIUM STRUCTURE
i':s'S:
FIG. 7. A nearly in-plane section ofthe cells shows the straight edges and sharp corners. Empty oval spacescorrespond to dark granules seen in phase-contrast pictures. Magnification, x 14,300.
*-
FIG. 8. Cross-sectioned cells at a magnification comparable to that of the scanning electron micrographs,demonstrating the much more irregular shape of the fixed and embedded cells. Magnification, x13,000.
sufficient. The presence in the cell suspensionsofbacteriorhodopsin or a closely related pigmentin low concentration may also be used as anargument, because this pigment has so far onlybeen found in extreme halophiles. However,pure cultures must be obtained before firm con-clusions can be reached.The observed cell wall structure also presents
some problems. Walsby mentioned that he ob-served a hexagonal lattice of 2-nm particles inshadowed cells. This is a typographical errorand should read 20 nm (A. E. Walsby, personalcommunication). His results, therefore, agreewith ours. However, it is generally thought thatin halobacteria, the cell wall determines the cellshape, and it is difficult to reconcile the hexag-onal lattice with the rectangular shapes of thecells. The difference in the shapes ofthe roundedcell profiles seen in section and the box-likeappearance of the cells in the scanning micro-
graphs also requires an explanation. Althoughwe are probably dealing with a heterogeneouspopulation, which may explain the differences inwall structure we observed, this heterogeneitycannot explain the differences between the scan-ning and transmission electron micrographs. Vir-tually all cells had flat edges in the scanningmicrographs and rounded edges in sectioned andfreeze-fracture preparations. Since it is unlikelythat distortions introduced by the techniqueproduced straight and flat edges, we believe thatthe rounded appearance is the artifact.We have recently examined brines from salt
ponds near Guerrero Negro and La Paz in BajaCalifornia, Mexico. In both cases, cells with thesame light microscopic appearance as the squarespecies from the Sinai were present in relativelylarge numbers. Samples from the salt ponds ofSan Francisco Bay contained about 10 times thetotal number of cells found in the Sinai and Baja
357VOL. 148, 1981
on June 27, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
A
FIG. 9. Cross-section of a cell showing the thick dense wall and the underlying cell membrane, whichappears only as a narrow light band (9A). In an oblique section, the regular structure of the wall becomesobvious (9B). Magnifications, x117,000 (9A) and x66,000 (9B).
FIG. 10. Freeze-etch electron micrograph. On the upper right hand, the globular structure of the cell wallis exposed by etching. Below it, the cytoplasmic fracture face of the cell membrane shows a much finer andirregular particle distribution. To the left, a granule has been drawn out into a horn-like structure by thefracturing. To the right of it, the cytoplasm has been exposed, and embedded in it is a collapsed gas vacuolewith its characteristic fine striation. Magnification, x 76,800.
on June 27, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

WALSBY'S SQUARE BACTERIUM STRUCTURE 359
FIG. 11. Freeze-fracture exposing, on the left, the external fracture face of the cell membrane. Next to it isthe cytoplasm with collapsed gas vacuoles and a smooth depression left by a granule removed by thefracturing. To the right is the cytoplasmic fracture face of the cell membrane. Magnification, x28,200.
California samples, but square cells were ex-tremely rare. It is, therefore, likely that they canbe found in other locations too and, so far, havenot been recognized as cells because of theirunusual shapes. Occasional particles with a sim-ilarly flat but triangular shape have been seen.These are also mentioned by Walsby, butwhether they are bacteria related to the squarespecies remains to be seen.
Since cells with this distinct morphology arefound in widely separated locations and morerelated species are likely to be found, the needfor a proper generic name arises; "square" bac-teria is obviously unsatisfactory. Traditionally,procaryotes have been named according to theirshape, and since the shape of these cells in thelight microscope and the scanning electron mi-croscope resembles that of a flat box, we tenta-tively suggest Arcula (F. L. n. arcula, a smallbox).
ACKNOWLEDGMENTSThis work was supported by National Aeronautics and
Space Administration grant NSG-7151. I thank R. A. Bogo-molni and R. H. Lozier for the flash spectroscopy data anddiscussion, Barbara J. Javor for help in collecting brines, andJ. A. Long for help with the scanning electron microscopy.Thelma Castellanos generously made available facilities for apreliminary inspection of samples at the Centro De Investi-gaciones Biologicas La Paz, B. C. S., Mexico.
LITERATURE CITED1. Blaurock, A. E., and W. Stoeckenius. 1971. Structure
of the purple membrane. Nature (London) New Biol.233:152-155.
2. Clark, Q. W., and C. Branton. 1968. Fracture faces infrozen outer segments from the guinea pig retina. Z.Zellforsch. 91:586-603.
3. Lozier, R. H., R. A. Bogomolni, and W. Stoeckenius.1975. Bacteriorhodopsin: a light-driven proton pump inHalobacterium halobium. Biophys. J. 15:955-962.
4. Sleytr, U. B., and Q. W. Robards. 1977. Plastic defor-mation during freeze cleavage. A review. J. Microsc.(Oxford) 110:1-25.
5. Stoeckenius, W., and W. H. Kunau. 1968. Further
VOL. 148, 1981
on June 27, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

360 STOECKENIUS
characterization of particulate fractions from lysed cellenvelopes of Halobacterium halobium and isolation ofgas vacuole membranes. J. Cell Biol. 38:337-357.
6. Stoeckenius, W., R. Lozier, and R. Bogomolni. 1979.Bacteriorhodopsin and the purple membrane of halo-bacteria. Biochim. Biophys. Acta 505:215-278.
7. Stoeckenius, W., and R. Rowen. 1967. A morphological
study ofHalobacterium halobium and its lysis in mediaof low salt concentration. J. Cell Biol. 34:365-393.
8. Walsby, A. E. 1980. A square bacterium. Nature (Lon-don) 283:69-71.
9. Woese, C. R., and G. E. Fox. 1977. Phylogenetic struc-ture of the prokaryotic domains: the primary kingdoms.Proc. Natl. Acad. Sci. U.S.A. 74:5088-5090.
J. BACTERIOL.
on June 27, 2018 by guesthttp://jb.asm
.org/D
ownloaded from