volatile products of the lipoxygenase pathway evolved from
TRANSCRIPT

Plant Physiol. (1993) 101: 13-24
Volatile Products of the Lipoxygenase Pathway Evolved from Phaseolus vulgaris (1.) Leaves lnoculated with
Pseudomonas syringae pv phaseolicola'
Kevan P. C. Croft, Friedrich Jiittner, and Alan J. Slusarenko*
lnstitut für Pflanzenbiologie, Zollikerstrasse 107, Zürich CH-8008, Switzerland
Activation of the "lipoxygenase pathway" in plants gives rise to a series of products derived from fatty acids. Analysis by gas chromatography-mass spectroscopy of volatile products produced by Phaseolus vulgaris (1.) cv Red Mexican leaves during a hyper- sensitive resistance response (HR) to the plant pathogenic bacte- rium Pseudomonas syringae pv phaseolicola showed evolution of several lipid-derived volatiles, including cis-3-hexenol and trans- 2-hexena1, which arise from the 13-hydroperoxide of linolenic acid. These compounds were not produced in detectable amounts by buffer-inoculated leaves, nor did they evolve to such a high degree during comparable stages of the susceptible response. The absence of frans-2,cis-6-nonadienal, a product expected from 9-hydroper- oxide of linolenic acid, suggests that lipid peroxidation during the HR proceeded primarily enzymically via bean lipoxygenase, which produces the 13-hydroperoxide, and not via autoxidative proc- esses. The effects of trans-2-hexena1, cis-3-hexeno1, and traumatic acid on P.s pv phaseolicola were investigated. trans-2-Hexenal appeared to be highly bactericidal at low concentrations, whereas cis-3-hexenol was bactericidal only at much higher concentrations. Traumatic acid appeared to have no effect on P.s. pv. phaseolicola at the concentrations tested. These results demonstrate that during plant defense responses against microbial attack, several lipid- derived compounds are produced by the plant, some of which possess antimicrobial activity and conceivably are involved in plant disease resistance. The time of production of these substances, in amounts that would be expected to be antibacterial in vitro, cor- related with a slowing down of the growth rate of bacteria in the leaves and was seen at a time before the accumulation of isofla- vonoid phytoalexins in the host.
In plants undergoing pathogen attack or other forms of biotic or abiotic stress or damage, various lipid-breakdown products, including c6 volatiles, may be formed from the polyunsaturated octadecanoid fatty acids linoleic acid and linolenic acid by sequential enzyme steps involving lipoxy- genase (EC 1.13.1 1.12), hydroperoxide lyase, and hydroper- oxide dehydrase, which form part of the so-called "lipoxy- genase pathway" (Vick and Zimmerman, 1984, 1987a, 198713). Hydroperoxide dehydrase is the single name now given to what were formerly known as hydroperoxide iso- merase and hydroperoxide cyclase (Vick and Zimmerman, 1987b). Hatanaka et al. (1987) surveyed a wide range of
K.P.C.C. was supported by a Fellowship from the Royal Society of London administered through the Science and Engineering Re- search Council (U.K.). F.J. and A.J.S. were supported by the Kanton of Ziirich.
* Corresponding author; fax 41-1-385-4204.
plants and found the c6 volatile-forming system was present in a11 the plants tested but with varying degrees of constitutive activity in different species. The enzymic breakdown of lin- olenic acid produces a range of products and intermediates, many of which contribute to characteristic odors of cut or damaged plant tissue (Hatanaka et al., 1987; Siedow, 1991). The biological activity of many of these substances is poorly understood (Vick and Zimmerman, 1987a; Siedow, 1991) but Vick and Zimmerman (1984) noted that some of the products had structures similar to those of the regulatory eicosanoids of animals and suggested that "octadecanoids" (i.e. Cls lipox- ygenase-pathway products derived from linoleic or linolenic acid) could fulfill similar roles in plants. Thus, it has been suggested that lipoxygenase activity might produce signal molecules important in host-pathogen interactions (Anderson 1989; Slusarenko et al., 1991; Farmer and Ryan, 1992). In this work, we explore the possibility that lipid-derived vola- tiles, produced ultimately as a result of lipoxygenase activity, may also have a more direct effect on the pathogens them- selves.
Depending on the source of the enzyme, lipoxygenase activity on linolenic acid will produce either 9- or 13-hydro- peroxylinolenic acid or a mixture of both. In bean, the 13- hydroperoxide is the major product of lipoxygenase activity (Matthew and Galliard, 1978). Figure 1 shows how a hydro- peroxide lyase can then catalyze the breakdown of 13-hydro- peroxylinolenic acid to a c6 product, cis-3-hexena1, and a Clz product, 12-0x0-cis-9-dodecenoic acid. In further reactions, cis-3-hexenal can form cis-3-hexenol, truns-2-hexeno1, truns- 3-hexenol, or truns-2-hexenal (Fig. 1). The CI2 product, 12- 0x0-cis-9-dodecenoic acid, is a precursor of the plant wound hormone traumatic acid (Strong and Kruitwagen, 1967; Vick and Zimmerman, 1987a; Anderson, 1989; Siedow, 1991). The 9-hydroperoxide of linolenic acid, when present, is con- verted in bean to truns-2,cis-6-nonadienal by a corresponding lyase activity (Matthew and Galliard, 1978).
truns-2-Hexenal (C6H100) is a six-carbon a,P-unsaturated aldehyde and is virtually immiscible with water (CRC Hand- book of Chemistry and Physics, 1989-1990). Although it has been suggested that truns-2-hexenal may be involved in the defense mechanism of plants to pathogens (Lyr and Basaniak, 1983; Vick and Zimmerman, 1987a) it has not until now been demonstrated that truns-2-hexenal was produced in host-
Abbreviations: cfu, colony-forming units; HR, hypersensitive re- sponse; NAG, nutrient agar and glycerol; NBG, nutrient broth and glycerol; TTC, 2,3,5-triphenyltetrazolium chloride.
13
Dow
nloaded from https://academ
ic.oup.com/plphys/article/101/1/13/6065904 by guest on 01 January 2022

14 Croft et al Plant Physiol. Vol. 101, 1993
COOH
LOX linolenic acid
O* 1 OOH
13-hydroperoxylinolenic acid 1 HLA
cis-3-hexenal 12.oxu-~i~9-dodecenoic acid
COOH
mm -3-hexenal ci.r-3-hexen-1-01
m t t s -3- hexenol iram-2-hexenal
12-oxo-iruw lO-dodecenoic acid
traumatic acid
rmm-2-hexenol
Figure 1. Biochemical pathway showing part of the lipoxygenase cascade from linolenic acid. LOX, Lipoxygenase; IF, isomerization factor; ADH, alcohol dehydrogenase.
pathogen interactions. trans-2-Hexenal was isolated from leaves of Ginkgo biloba and was reported to inhibit growth of the fungus Monilinia fructicola at a concentration of 300 ppm (Major et al., 1960). It was suggested that trans-2-hexenal was involved in the ginkgo tree's remarkable resistance to pests and diseases. In various studies, trans-2-hexenal has also been shown to possess antiprotozoal (Von Schildknecht and Rauch, 1961), antifungal, insecticidal, and acaricidal activity (Lyr and Basaniak, 1983). Recently, Zeringue and McCormick (1989) demonstrated that volatiles, including trans-2-hexenal, produced from wounded cotton leaves were able to inhibit growth of the saprophytic fungus Aspergillus flavus growing in liquid cultures. Matthew and Galliard (1978) proved that trans-2-hexenal and certain other products were formed from fatty acid hydroperoxides via the lipoxy- genase pathway during wounding in Phaseolus vulgaris and showed the lyase and cleavage enzymes were present con- stitutively. We previously demonstrated an increase in lipox- ygenase isozymes in beans undergoing a HR after inoculation with an avirulent race of the plant pathogenic bacterium Pseudomonas syringae pv phaseolicola but found no increase in activity in the susceptible response after inoculation with virulent bacteria (Croft et al., 1990). Fatty acid peroxidation
was shown to be occurring in vivo during the HR by moni- toring alkane gas production (ethane and pentane) from whole leaves (Croft, 1990; Croft et al., 1990). Ethane and pentane evolution has been widely used as a reliable index of in vivo lipid peroxidation (Konze and Elstner, 1978; Kunert and Tappel, 1983; Slater, 1984). Evidence for a role for lipoxygenase in plants during disease responses has accu- mulated in recent years (Keppler and Novacky, 1986, 1987; Ocampo et al., 1986; Yamamoto and Tani, 1986; Ohta et al., 1990), and elicitor-induced lipid peroxidation has been re- ported in bean (Rogers et al., 1988).
Substrates for lipoxygenase, such as linoleic and linolenic acids, are common constituents of plant membranes (Epton and Deverall, 1968; Hoppe and Heitefuss, 1974) but are not readily acted upon by the enzyme unless they are in the free acid form (Leshem, 1984). Polyunsaturated fatty acids are very susceptible to autoxidative processes in the free acid form but much less so when they are in situ in membranes (Galliard, 1978). Hoppe and Heitefuss (1974) established that linoleic and linolenic acids were among the most common fatty acids found in the lipid components of healthy P. vulgaris membranes, linolenic being more common than lin- oleic. Epton and Deverall (1968) calculated that linolenic acid
Dow
nloaded from https://academ
ic.oup.com/plphys/article/101/1/13/6065904 by guest on 01 January 2022

Antibacterial Activity of Lipid Breakdown Products 15
was present in P. vulgaris leaves at a level of around 320 pg g-' of leaf tissue.
The presence of suitable substrates, the evidence of in- creases in lipoxygenase activity as a widespread plant disease- resistance response, and the constitutive presence of the other necessary enzymes indicate that the production of lipid- breakdown substances may be an important factor in disease resistance. In view of the antifungal and antiprotozoal activity of trans-2-hexenal reported in the literature (Major et al., 1960; von Schildknecht and Rauch, 1961), it seemed feasible that trans-2-hexenal and perhaps other fatty acid-derived substances produced in the plant were involved in the resist- ance of P. vulgaris to infection with P.s. pv phaseolicola. Experiments were camed out to determine if any lipid- derived volatile substances were produced during the HR of P. vulgaris induced by P.s. pv phaseolicola and to test the effect in vitro of cis-3-hexenol and trans-2-hexenal on P.s. pv phaseolicola. Because traumatic acid is formed from the re- maining moiety of linolenic acid when C6 volatiles are pro- duced, the effect of this substance on P.s. pv phaseolicola was also investigated. The possible importance of volatile lipid- breakdown products in plant defense responses is discussed.
MATERIALS AND METHODS
Plant and Bacterial Material
Seeds of Phaseolus vulgaris (L.) cv Red Mexican U13 were lightly scarified using fine glasspaper, allowed to imbibe tap water overnight, with severa1 water changes, and planted individually in 2.5-inch plastic pots containing compost. The pots were transferred to growth cabinets with a 16-h photo- period at day and night temperatures of 25O and 2Z0C, respectively. The primary leaves were ready for vacuum infiltration approximately 8 to 9 d after sowing.
Pseudomonas syringae pv phaseolicola (Burkholder) isolate 31A (race 1) and isolate 1301A (race 3), donated by Dr. J.D. Taylor, Institute of Horticultural Research, Wellesbourne, UK, were obtained from the culture collection of the Department of Applied Biology, University of Hull, England. Working cultures were routinely maintained on nutrient agar (Oxoid) supplemented with 1% (v/v) glycerol (NAG plates). After streaking out onto NAG plates, cultures were grown at 25OC for 1 to 2 d to allow heavy bacterial growth.
Plant lnoculation Procedure
Bacteria for inoculation were grown in 5 mL of nutrient broth (Oxoid) containing 1% (v/v) glycerol (NBG) in a 25- mL Sterilin tube in an orbital shaking incubator at 150 cycles min-' at 25OC. The day before inoculation, cells growing in NBG were used to inoculate a 25-mL culture of King's B broth (20% [w/v] protease peptone No. 3 [Difco], 2% [w/v] KH2P04. 3H20, 1.5% [w/v] MgSO4-7H20). After approxi- mately 18 h of growth, cultures were centrifuged at 3000g for 10 min and the bacterial pellet resuspended gently in inoculation buffer (I buffer = 0.0014 M KH2PO4, 0.0025 M
Na2HP04, pH 7.00) and adjusted to an A600 of 0.06, which dilution plate counts showed to contain approximately 1 X 10' cfu mL-I. Plants were vacuum infiltrated as previously described (Croft et al., 1990). The underside of the plants
were dark green and water soaked after inoculation but regained normal appearance after 30 min in the growth cabinet. The inoculated leaf area was regularly 90% or more of the total. After inoculation, plants were returned to the growth chamber and placed under constant light.
Bacterial Growth in Vivo
Leaf discs (10, 1 cm in diameter) were cut from inoculated leaves at various times after inoculation and ground up in 5 mL of inoculation buffer with a mortar and pestle, and seria1 dilutions in inoculation buffer were plated out and incubated for 3 d at 25OC on nutrient agar (Oxoid) plates supplemented with 1% (v/v) glycerol to determine bacterial numbers. The data from three independent inoculations were pooled and each time point represents the data from three to six plants collectively .
Effect of trans-2-Hexenal on Bean Plants
Individual seedlings were placed in airtight polypropylene containers (13 cm high, enclosed volume 1 L). One proprie- tary cotton bud (Johnson & Johnson) was placed in the compost in each pot, and either 25 1 L of ethanol or 5, 10, or 25 pL of trans-2-hexenal dissolved in ethanol was pipetted onto it. The pots were sealed and replaced in the growth chamber. After 2 d, leaf tissue was extracted and analyzed for phytoalexins as described by Slusarenko et al. (1989). The experiments were performed twice with two seedlings each time.
Collection of Plant Volatiles and GC-MS Analysis
The method used was that of Jüttner (1988) modified for the analysis of emission products. The plant hypocotyl was cut with a scalpel approximately 1 to 2 cm up from soil level and the base wrapped in aluminum foi1 to prevent water loss from the cut surface. The plant was then placed in an air- tight glass container with two ports. The exit port was at- tached to a collection cartridge containing 150 mg of Tenax TA sorbent (40-60 mesh) and then by a Luer lock via copper tubing to a stainless steel pump and back via the inlet port to the glass vessel. Air in the enclosed system was circulated at a flow rate of 1.2 L min-' for 30 min over the Tenax sorbent. Two 40-W light bulbs were placed near to the exit port over the collection cartridge and Tenax bed to prevent water condensation. Once collection of volatiles was com- plete, the cartridge was sealed in an air-tight glass-stoppered vessel until it was analyzed by GC-MS (Hewlett-Packard 5970A).
To desorb the collected volatiles, the Tenax cartridge was placed in a movable heating block (40-W capacity) and fixed to a helium gas supply with a flow rate of 44 mL min-' while the Luer end of the column was fixed with a side-port needle and introduced through the GC injection port septum. The interna1 carrier gas supply for the GC was switched off and the gas supply passing over the Tenax collection cartridge allowed to replace it. The oven was cooled to O°C to trap the released volatiles and the Tenax cartridge was heated to 27OOC over 2 min and maintained at this temperature for 2
Dow
nloaded from https://academ
ic.oup.com/plphys/article/101/1/13/6065904 by guest on 01 January 2022

16 Croft et ai. Plant Physiol. Vol. 101, 1993
min. The collection cartridge needle was then removed from the septum, the interna1 carrier gas supply restored, and the temperature program started (4 min at OOC, 5OC min-' up to 15OOC). The GC column was a 30-m silica gel capillary column (0.32 mm o.d., DB 1301, J & W Scientific). Mass spectra were taken in the range 28.5 to 250 atomic mass units with two scans s-' and a scan threshold of 500. Compounds were quantified by integration of the fragment ions ( m / z 41 for both trans-2-hexenal and cis-3-hexenol), which were ex- tracted from chromatograms taken in either the scan or selected ion monitoring mode. The fragment ions m/z 41, 69, and 70 were used in the selected ion monitoring mode to determine trans-2,cis-6-nonadienal quantitatively. Under the analytical conditions used, the detection limit for this com- pound was 8 ng.
Preparation of Hexenal-Saturated Buffer and Quantification by GLC
Approximately 25 pL of trans-2-hexenal (Sigma) was added to 200 pL of inoculation buffer, vortexed, and allowed to equilibrate overnight at room temperature. The undissolved trans-2-hexenal was visible as an oily layer on the surface, and the saturated buffer below the trans-2-hexenal was re- moved using a syringe and needle. Quantification of trans-2- hexenal dissolved in the buffer was by GC according to the method of Galliard et al. (1986). Aliquots of up to 10 pL of the trans-2-hexenal-saturated buffer were analyzed on a Pye Unicam GCD gas chromatograph containing a 1.5 m X 4 mm column packed with 10% Carbowax, chromosorb W 80/100 mesh, acid washed, and dimethyldichlorosilane treated. The column temperature was llO°C, and the detection and injec- tion temperatures were 18OOC. The carrier gas was nitrogen at a flow rate of 30 mL min-'. trans-2-Hexenal is readily soluble in pentane (Matthew and Galliard, 1978), so a series of standard solutions was prepared using authentic trans-2- hexenal (Sigma) dissolved in pentane. Under the conditions described, the retention time for the trans-2-hexenal standard was approximately 2 min 20 s, no other peaks being detected. When aliquots from six different preparations of trans-2- hexenal-saturated buffer were analyzed by GC, the solutions were shown to contain on average 3.33 nL of trans-2-hexenal per pL of buffer injected (SE = k0.047 nL FL-'). After GC quantification, the trans-2-hexenal-saturated buffer was used in bacterial toxicity experiments. In the different experiments described in this work, it is appropriate to express the amount of trans-2-hexenal used either as a volume (nL) applied to Petri plates, as a concentration ( p ~ ) in shake-culture, or as a mass (pg) when liberated as a volatile from leaves. To assist in quantitative comparison between these different situations, the following equivalents should be borne in mind 1 nL of trans-2-hexenal = 0.841 pg, and 1 PM of trans-2-hexenal = 98 ng mL-'.
Activity on Bacterial Growth of frans-2-Hexenal Spotted on Filter Discs
Approximately 10 mL of an overnight culture of P.s. pv. phaseolicola, grown in NBG medium, was centrifuged at 4000g in a Denley bench centrifuge for 10 min, and the pellet
was resuspended in approximately 5 mL of 1 buffer. The optical density of the bacterial solution was adjusted to A600 = 0.06. Approximately 150 pL of bacterial suspension was pipetted onto a fresh, dry NAG plate and evenly distributed with a glass spreader. The plate was covered and incubated for 4 h at 25OC to allow the agar surface to dry. Plates seeded in this way showed heavy growth of bacteria within 1 d when incubated at 25OC.
To test the effect of trans-2-hexenal on growth of bacteria on NAG plates, sterile 14 mm X 2 mm filter discs (Whatman) were placed centrally on the agar surface of previously seeded NAG plates. Different amounts of trans-2-hexenal or trans- 2-hexenal-saturated I buffer were pipetted onto filter discs on Petri plates. trans-2-Hexenal was applied in a range from 75 to 1 pL. trans-2-Hexenal-saturated buffer was diluted to 50 pL with I buffer to give aliquots containing 167, 83.3, 16.5, 1.7, 0.33, or 0.03 nL of trans-2-hexenal. These aliquots were then spotted onto filter discs. Control plates were pre- pared by pipetting 50 pL of I buffer onto filter discs. Plates were then sealed with Nescofilm and incubated at 25OC overnight. The experiment was repeated four times.
Effect on Bacterial Crowth of frans-2-Hexenal in Agar Plugs
Sterile 9-cm Petri dishes containing NAG to a depth of 5 mm had 10-mm diameter wells cut in the agar with a cork borer. Various amounts of trans-2-hexenal-saturated buffer were brought to 105 pL with I buffer and pipetted into the wells, then mixed with 150 pL of 45OC NAG medium by drawing back and forth with a pipette. A control with no trans-2-hexenal was also prepared. Once the agar plugs had set, 300 pL of P.s. pv phaseolicola, adjusted with I buffer to A600 = 0.1, was mixed with 5 mL of 45OC molten top agarose (0.4% agarose, 10 mM MgS04) then poured onto the agar plate. Incubation was overnight at 25OC. Another agarose overlay including a chromogenic substrate was used to dis- tinguish between areas where bacteria had grown and where they had been killed, and was adapted from the TLC overlay method of Slusarenko et al. (1989). Molten agarose (5 mL) at 45OC was taken to 0.1% (w/v) with TTC (Sigma), mixed thoroughly, poured over the bacteria-seeded agarose surface, and incubated at room temperature. Color development be- gan almost immediately and was complete by the next day. Areas where bacterial growth was inhibited were colorless, whereas areas of bacterial growth were pink due to the reduction of TTC to formazans by bacterial dehydrogenases. The experiment was repeated six times.
Effect of trans-2-Hexenal on Bacterial Crowth in Liquid Cultures
Sterile 250-mL flasks containing 25 mL of NBG were inoculated with 2 mL of an overnight culture of P.s. pv phaseolicola and 1 pL (0.35 mM), 5 pL (1.73 mM), or 25 pL (8.65 mM) of trans-2-hexenal was added per flask. Growth was compared with that in a control flask containing no trans-2-hexenal. The flasks were incubated in an orbital shaker at 25OC, 150 rpm, and changes in the A600 of the flasks were measured over time. After 2.5 h of incubation, a
Dow
nloaded from https://academ
ic.oup.com/plphys/article/101/1/13/6065904 by guest on 01 January 2022

Antibacterial Activity of Lipid Breakdown Products 17
loopful of culture from each flask was streaked onto a NAG plate and the plates were incubated as before.
The experiment above was repeated using various quan- tities of trans-2-hexenal-saturated I buffer added to the bac- teria1 cultures. The final concentrations of trans-2-hexenal in the flasks were 173, 86.4, 57.6, 28.8, and 5.8 p ~ . The exper- iment was repeated twice.
EM of trans-2-Hexenal-Treated Bacteria Crowing in Liquid Culture
A 100-mL culture of P.s. pv phaseolicola with an AbO0 of 1.00 was made 2.2 mM with trans-2-hexenal and incubated at 25OC with shaking at 150 cycles min-I. The A600 was measured every 15 min for the next 1.5 h, by which time it had dropped to approximately 0.8. A control flask was treated with 25 pL of sterile distilled water. Bacteria were collected by centrifugation at 4000g for 10 min and resuspended in 2.5% (v/v) glutaraldehyde in 0.1 M phosphate buffer, pH 7.2, and left for 1 h at room temperature. The bacteria were washed 3 times in 0.1 M phosphate buffer, pH 7.2, at room temperature for 5 min and collected by centrifugation. The final resuspension was in postfixative 1% (w/v) osmium tetroxide in 0.1 M phosphate buffer, pH 7.2, and samples were left at room temperature for 30 min before being centrifuged and washed four times for 5 min each time in 0.1 M phosphate buffer, pH 7.2. Dehydration of the samples was carried out in a graded ethanol series, with 5 min in each solution, up to absolute ethanol. Samples were embedded in an araldite/epon mixture and polymerized at 7OoC for 24 h. Sections were cut using glass knives on an ultramicrotome and mounted onto uncoated 300-mesh copper grids. Staining followed, in 2% (w/v) uranyl acetate for 30 min and lead citrate for 5 min. Sections were viewed on a JEOL 100 c electron microscope at 80 kV.
Effect on Bacterial Crowth of cis-3-Hexenol Sealed in Agar Plugs
Sterile 6-cm Petri dishes were filled to a depth of 5 mm with NAG and allowed to dry, and 10-mm diameter wells were cut centrally. Because at the concentrations used the cis-3-hexenol (Sigma) was highly insoluble in the water- based agar, it was necessary to add ethanol and DMSO as solubilizing agents to final concentrations of 8% (v/v) and 36% (v/v), respectively. The cis-3-hexenol solution was taken to a total volume of 140 pL with water and then mixed with 140 pL of molten NAG at 45OC and pipetted into the central well of the agar plate and allowed to solidify. The volume of cis-3-hexenol added to the plates ranged from 1 to 15 pL. A control plate was prepared in the same way but with no cis- 3-hexenol. The bacterial overlay and subsequent chromo- genic overlay were used as described above for the trans-2- hexenal but were scaled down for the smaller size of the Petri dishes to 2.5 mL of each overlay solution. The experi- ment was repeated four times.
Effect of Traumatic Acid on Bacterial Crowth in Liquid Cultures
Sterile 250-mL flasks containing 25 mL of NBG were inoculated with P.s. pv phaseolicola as above and traumatic
acid (Sigma) was added to the flasks to final concentrations of 1.88, 0.94, and 0.47 mM. Because traumatic acid is very insoluble in aqueous solutions, it was first dissolved in DMSO and then added to the flasks; DMSO was also added to the control flask (no traumatic acid). The final concentration of DMSO in the culture medium was 2.9% (v/v), which had no apparent effect on bacterial growth in the control but kept the traumatic acid in solution. It was impossible to take traumatic acid to a concentration above 2 mM because of its insolubility and the undesirability of higher DMSO concen- trations. The experiment was repeated twice.
RESULTS
Production of Lipid-Derived Volatiles by lnoculated Plants
Plants inoculated with either race 1 or race 3 of P.s. pv phaseolicola produced a range of volatiles (Fig. 2), and plants undergoing an HR produced the greatest amounts. Between 15 and 24 h after inoculation the average amounts of truns- 2-hexenal and cis-3-hexenol produced by plants undergoing an HR were approximately 17 and 16 times higher, respec- tively, than the amounts produced by plants undergoing a suceptible response (Fig. 3, A and B). The identity of the volatiles was confirmed by GC-MS, and retention times and mass spectra were compared to those of authentic reference substances. The mass spectra of the substances putatively identified as fruns-2-hexenal and cis-3-hexenol are shown in Figure 4, A and B. At 24 h after inoculation during the HR, cis-3-hexenol (“leaf alcohol”) was the major volatile detected and the second most abundant substance was cis-3-hexena1, which is the precursor of cis-3-hexenol (Fig. 1). Direct and indirect pathways can lead to trans-2-hexenal production from cis-3-hexenal, and, indeed, a large peak of trans-2- hexenal was detectable, as well as a smaller peak of trans-2- hexenol, the alcohol that is formed from reduction of trans- 2-hexenal by alcohol dehydrogenase.
It is interesting that when plants showed good hypersen- sitive cell collapse (i.e. from 15-24 h after inoculation), the leaves emitted a distinctive odor due to volatile production, somewhat like dried tea. Indeed, cis-3-hexenol and trans-2- hexenal are mainly responsible for the “fresh” odor of green tea (Takeo, 1992). Buffer-inoculated plants, or plants undergoing a susceptible response at 24 h after inoculation, did not have this distinctive odor.
Effects of trans-2-Hexenal on Bean Plants
At the concentrations tested, bean plants showed loss of turgor by 48 h after beginning treatment. By this time, plants that received 25 pL L-’ trans-2-hexenal had collapsed com- pletely and dried out. At the lower concentrations, plants were flaccid and epinastic. TLC of ethanol extracts of leaf tissue showed that trans-2-hexenal-treated plants had suf- fered a reduction in photosynthetic pigments compared with the controls. No isoflavonoid phytoalexins were detected.
Effects of trans-2-Hexenal on Bacteria
As little as 1 pL of trans-2-hexenal spotted onto a filter disc prevented a11 bacterial growth on 9-cm diameter NAG plates
Dow
nloaded from https://academ
ic.oup.com/plphys/article/101/1/13/6065904 by guest on 01 January 2022

18
I I I I I I I 1
O 1
Croft et al.
O
O
1 O
I.= ................. ........... 8 ..........................................
Plant Physiol. Vol. 101, 1993
I I I I I I I 1
O
-
O
O
O -
-
I.= - ................. ........... 8 ..........................................
trans-2-hexenal
A
cir-3-hexenol IM
tmns-2-hexenol U L B
C
A
r 6
m - o & e . I n I I I a ] - O 6 12 10 24 30 36 42 40 54
Hours after inoculation
c - 25 u! I I I I I I 1 I I
O O
\
c O
- u)
4 o g 20
- Iu k O I I E o X o c o o ol a
r
I
I
l5 t O i ...................................o...
1 I I a O - - O 6 12 10 24 30 36 42 40 54
Hours after inoculation Figure 3. A, Liberation of trans-2-hexenal by inoculated plants and B, liberation of cis-3-hexenol. Each data point shows the amount liberated by a single plant over a 30-min sampling period. O, Plants inoculated with P.s. pv phaseolicola race 1 (incompatible combi- nation); O, plants inoculated with virulent lace 3 of the pathogen (compatible combination); O, buffer-inoculated plants. The dotted line indicates the maximum leve1 of volatile production seen in the compatible combination. The arrow shows the mean time at which HR cell collapse visible to the naked eye occurred. The data values for the 24-h sampling time were chosen for statistical analysis using a one-way analysis of variance. Mean trans-2-hexenal for HR = 1.9 pg, SE = 0.6, n = 5; for susceptible response, 0.2 pg, SE = 0.1, n = 5; with buffer, 0.0, SE = 0.0, n = 4. Mean cis-3-hexenol in the HR = 10.7 pg, SE = 3.3, n = 4; for susceptible response, 0.7 pg, S E = 0.3, n = 5; with buffer, 0.1 pg, SE = 0.1, n = 4. There were significant differences among the treatments in the amounts of both trans-2- hexenal and cis-3-hexenol 11.1 and FZ,,, = 8.00, respectively, both P < 0.01). Furthermore, Fisher’s tests of the LSDS showed that
Figure 2. Gas chromatograph of volatiles produced by P. vulgaris cv Red Mexican plants over a 30-min period at 24 h after inocula- tion. A, HR; B, susceptible response; C, buffer-inoculated control plants.
the significant differences were due to the HR being significantly different from the susceptible response and buffer treatments in both cases. There were no significant differences between the buffer treatment and the susceptible response.
Dow
nloaded from https://academ
ic.oup.com/plphys/article/101/1/13/6065904 by guest on 01 January 2022

Antibacterial Activity of Lipid Breakdown Products 19
A 12
1 839
29
,|
55
1.II ,i.ll
6
n
9a
70
——— u—
3
96ll
30 4O 50 CO TO 80 90mass/charge
B 9
11 e1c2 3|!•
» n
31
|L .1
41
67
55
1 1-ll, .,|||j .
8259,l| ,« WO
3 0 4 O S 0 6 0 7 0 8 0 9 0 X X )mass/charge
Figure 4. MS profiles for substances putatively identified as trans-2-hexenal (A) and c/s-3-hexenol (B).
previously seeded with P.s. pv phaseolicola. Control plates(no frans-2-hexenal) showed heavy bacterial growth. Smalleramounts of frans-2-hexenal applied as a saturated buffersolution to filter discs delayed or retarded bacterial growth(Table I). frans-2-Hexenal seemed to exert a bactericidal effectin amounts greater than 16.5 nL. The effect was uniformover the entire plate and not just in the immediate areasurrounding the filter disc, showing that the frans-2-hexenaIwas volatile enough to have a bactericidal effect some dis-tance away from the immediate point of application. Whenfrfltts-2-hexenal was applied as a buffer-saturated solutionsolidified in agar plugs, the volatility was reduced and distinctzones of inhibition of bacterial growth could be seen onseeded plates (Fig. 5). Even at the lowest concentration ofrratts-2-hexenal tested (plug "a" contained 55 nL of trans-2-hexenal) there was clear evidence of bactericidal activity. Thefrans-2-hexenal appears to have diffused out from the point
Table I. The effect of trans-2-hexenal-saturated buffer pipetted ontofilter discs on the growth of bacteria on NAC plates
++++, Heavy bacterial growth; +, weak bacterial growth.nL of trans-2-Hexenal in 50 jiL of
Buffer Added to Each PlateGrowth after 1 d ofIncubation at 25°C
0.030.331.7
16.583.3
167
Control3No growth
f
Buffer only (50 f«L).
Figure 5. Inhibition of bacterial growth on NAC plates by trans-2-hexenal sealed in agar plugs. The amount of (rans-2-hexenal in eachagar plug: a, 55 nL; b, 115 nL; c, 170 nL; d, 230 nL; e, 290 nL; f,345 nL; o, control (no trans-2-hexenal).
of application, killing bacteria as it went. The size of thecolorless area where bacterial growth was inhibited wasdependent on the concentration of bacterial inoculum usedto seed the plate. Concentrated inoculum gave rise to asmaller-diameter inhibition and more dilute inocula to alarger one. This would imply that as the rrans-2-hexenaldiffused through the agar it was effectively titrated out bythe bacteria that it killed. For comparison of results, it isimportant that the bacteria are fresh, the A600 is known, andthat the volume of bacteria added per plate is constant. Thisplate assay method is superior to the method in which trans-2-hexenal was spotted onto filter discs in that the trans-2-hexenal is localized by being set in agar and gives clear zonesof inhibition rather than having a diffuse effect over theentire plate.
When frans-2-hexenal was added to liquid cultures of P.s.pv phaseolicola to concentrations of 8.65 mM, 1.73 HIM, and360 tiM, bacterial growth ceased almost immediately in allflasks, and over the next hour the A600 of the culturesdropped, indicating lysis of the bacteria. Over the next 6 h,the control culture with no added fra«s-2-hexenal grew nor-mally. Loopfuls of culture from each flask were then spreadonto NAG plates and incubated as before. After 2 d ofincubation at 25°C, some bacteria were shown to have re-mained viable in those cultures that contained 1.73 and 0.36mM frans-2-hexenaI. At lower concentrations (173-5.8 /*M),prepared by adding frans-2-hexenal-saturated buffer to themedium, there appeared to be a direct relationship betweenthe amount of frans-2-hexenal present and the retardation ofbacterial growth (Fig. 6).
EM of frans-2-Hexenal-Treated Bacteria
Bacteria exposed to 2.2 mM frans-2-hexenal showed con-siderable disruption to membranes and cell constituents, andthe DNA appeared to have been precipitated (Fig. 7A). Fromthe appearance of the cells and the evidence of a decrease in
Dow
nloaded from https://academ
ic.oup.com/plphys/article/101/1/13/6065904 by guest on 01 January 2022

20 Croft et al. Plant Physiol. Vol. 101, 1993
0.4OD600
Time(h) Hexenal1.5 173
Figure 6. Growth of P.s. pv phaseolicola in liquid cultures in thepresence of trans-2-hexenal. The increase in A600 (OD6oo) is shownfor cultures treated with various amounts of trans-2-hexenal andthe control (no trans-2-hexenal) at selected time points during theincubation period. The scales for trans-2-hexenal concentration andtime are not linear.
A60o following the addition of frans-2-hexenal, we concludedthat cell lysis had occurred. The appearance of control bac-teria (no frons-2-hexenal treatment) is shown in Figure 7B.
Effect of c/s-3-Hexenol in Agar Plugs on Bacterial GrowthIn comparison with fr«ns-2-hexenal, ds-3-hexenol was
much less inhibitory to bacterial growth (Fig. 8). Like trans-2-hexenal, ds-3-hexenol was fairly insoluble in aqueous so-lutions. However, ds-3-hexenol caused inhibition only whenit was added to agar plugs in amounts greater than 1 juL.Thus, although ds-3-hexenol did possess some antibacterialactivity, it was approximately 20 times less effective thanfrans-2-hexenal.
Effect of Traumatic Acid on Bacterial Growth inShake Culture
At the concentrations of traumatic acid used in this exper-iment (1.88, 0.94, and 0.47 HIM), there was no apparentdifference in bacterial growth in comparison with the controlflask. It was impossible to experiment at higher concentra-tions because of the insolubility of traumatic acid, but com-pared with fra«s-2-hexenal, which is active at low concentra-tions, traumatic acid seemed to have no effect on growth ofP.s. pv phaseolicola.
Bacterial Growth in VivoFigure 9 shows that, under the conditions of the experi-
ment, bacteria in the compatible and incompatible combina-
Figure 7. A, Electron micrograph of trans-2-hexenal-treated cells ofP.s. pv phaseolicola sampled after 1.5 h incubation in the presenceof 2.2 mM trans-2-hexenal. Magnification = 30,000x. B, Electronmicrograph of control cells. Magnification = 30,000x.
ISfll 10*11
Figure 8. Inhibition of bacterial growth on NAG plates by c/s-3-hexenol sealed in agar plugs. The amount of ds-3-hexenol in eachagar plug is shown, and the control contains no cis-3-hexenol.
Dow
nloaded from https://academ
ic.oup.com/plphys/article/101/1/13/6065904 by guest on 01 January 2022
Antibacterial Activity of Lipid Breakdown Products 21
I I I I I I I I
1 os
1 os 1 O 6 12 18 24 30 36 42 48 54
Hours after inoculation
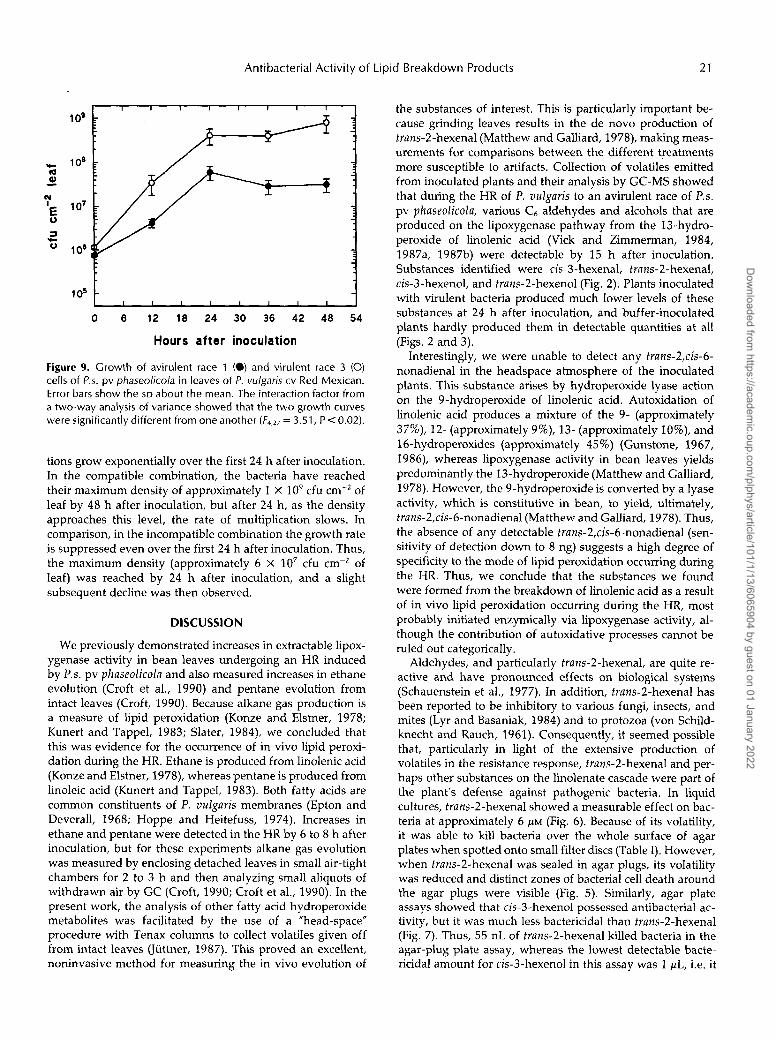
Figure 9. Growth of avirulent race 1 (O) and virulent race 3 (O) cells of P.s. pv phaseolicola in leaves of P. vulgaris cv Red Mexican. Error bars show the SD about the mean. The interaction factor from a two-way analysis of variance showed that the two growth curves were significantly different from one another (F4,*, = 3.51, P < 0.02).
tions grow exponentially over the first 24 h after inoculation. In the compatible combination, the bacteria have reached their maximum density of approximately 1 X 109 cfu cm-' of leaf by 48 h after inoculation, but after 24 h, as the density approaches this level, the rate of multiplication slows. In comparison, in the incompatible combination the growth rate is suppressed even over the first 24 h after inoculation. Thus, the maximum density (approximately 6 X 107 cfu cm-2 of leaf) was reached by 24 h after inoculation, and a slight subsequent decline was then observed.
DlSCUSSlON
We previously demonstrated increases in extractable lipox- ygenase activity in bean leaves undergoing an HR induced by P.s. pv phaseolicola and also measured increases in ethane evolution (Croft et al., 1990) and pentane evolution from intact leaves (Croft, 1990). Because alkane gas production is a measure of lipid peroxidation (Konze and Elstner, 1978; Kunert and Tappel, 1983; Slater, 1984), we concluded that this was evidence for the occurrence of in vivo lipid peroxi- dation during the HR. Ethane is produced from linolenic acid (Konze and Elstner, 1978), whereas pentane is produced from linoleic acid (Kunert and Tappel, 1983). Both fatty acids are common constituents of P. vulgaris membranes (Epton and Deverall, 1968; Hoppe and Heitefuss, 1974). Increases in ethane and pentane were detected in the HR by 6 to 8 h after inoculation, but for these experiments alkane gas evolution was measured by enclosing detached leaves in small air-tight chambers for 2 to 3 h and then analyzing small aliquots of withdrawn air by GC (Croft, 1990; Croft et al., 1990). In the present work, the analysis of other fatty acid hydroperoxide metabolites was facilitated by the use of a "head-space" procedure with Tenax columns to collect volatiles given off from intact leaves (Jüttner, 1987). This proved an excellent, noninvasive method for measuring the in vivo evolution of
the substances of interest. This is particularly important be- cause grinding leaves results in the de novo production of trans-2-hexenal (Matthew and Galliard, 1978), making meas- urements for comparisons between the different treatments more susceptible to artifacts. Collection of volatiles emitted from inoculated plants and their analysis by GC-MS showed that during the HR of P. vulgaris to an avirulent race of P.s. pv phaseolicola, various C6 aldehydes and alcohols that are produced on the lipoxygenase pathway from the 13-hydro- peroxide of linolenic acid (Vick and Zimmerman, 1984, 1987a, 1987b) were detectable by 15 h after inoculation. Substances identified were cis-3-hexenal, trans-2-hexena1, cis-3-hexeno1, and trans-2-hexenol (Fig. 2). Plants inoculated with virulent bacteria produced much lower levels of these substances at 24 h after inoculation, and buffer-inoculated plants hardly produced them in detectable quantities at a11 (Figs. 2 and 3).
Interestingly, we were unable to detect any trans-2,cis-6- nonadienal in the headspace atmosphere of the inoculated plants. This substance arises by hydroperoxide lyase action on the 9-hydroperoxide of linolenic acid. Autoxidation of linolenic acid produces a mixture of the 9- (approximately 37%), 12- (approximately 9%), 13- (approximately l O % ) , and 16-hydroperoxides (approximately 45%) (Gunstone, 1967, 1986), whereas lipoxygenase activity in bean leaves yields predominantly the 13-hydroperoxide (Matthew and Galliard, 1978). However, the 9-hydroperoxide is converted by a lyase activity, which is constitutive in bean, to yield, ultimately, trans-2,cis-6-nonadienal (Matthew and Galliard, 1978). Thus, the absence of any detectable trans-2,cis-6-nonadienal (sen- sitivity of detection down to 8 ng) suggests a high degree of specificity to the mode of lipid peroxidation occurring during the HR. Thus, we conclude that the substances we found were formed from the breakdown of linolenic acid as a result of in vivo lipid peroxidation occurring during the HR, most probably initiated enzymically via lipoxygenase activity, al- though the contribution of autoxidative processes cannot be ruled out categorically.
Aldehydes, and particularly trans-2-hexena1, are quite re- active and have pronounced effects on biological systems (Schauenstein et al., 1977). In addition, trans-2-hexenal has been reported to be inhibitory to various fungi, insects, and mites (Lyr and Basaniak, 1984) and to protozoa (von Schild- knecht and Rauch, 1961). Consequently, it seemed possible that, particularly in light of the extensive production of volatiles in the resistance response, trans-2-hexenal and per- haps other substances on the linolenate cascade were part of the plant's defense against pathogenic bacteria. In liquid cultures, trans-2-hexenal showed a measurable effect on bac- teria at approximately 6 PM (Fig. 6). Because of its volatility, it was able to kill bacteria over the whole surface of agar plates when spotted onto small filter discs (Table I). However, when trans-2-hexenal was sealed in agar plugs, its volatility was reduced and distinct zones of bacterial cell death around the agar plugs were visible (Fig. 5). Similarly, agar plate assays showed that cis-3-hexenol possessed antibacterial ac- tivity, but it was much less bactericidal than trans-2-hexenal (Fig. 7). Thus, 55 nL of trans-2-hexenal killed bacteria in the agar-plug plate assay, whereas the lowest detectable bacte- ricidal amount for cis-3-hexenol in this assay was 1 ML, i.e. it
Dow
nloaded from https://academ
ic.oup.com/plphys/article/101/1/13/6065904 by guest on 01 January 2022

22 Croft et al. Plant Physiol. Vol. 101, 1993
was approximately 20 times less effective than trans-2-hex- enal. Cis-3-hexenal, which readily isomerizes to trans-2-hex- enal (Hatanaka and Harada, 1973), was also evolved in substantial amounts in the HR (Fig. 2). However, it was not tested for bactericidal activity because it is relatively unstable and not commercially available.
Exactly how trans-2-hexenal exerts its antimicrobial activity is not known. The electron micrographs of trans-2-hexenal- treated bacterial cells (Fig. 7A) appear to show precipitation of DNA into strands and lysis of some cells, which is not seen in the untreated control cells (Fig. 7B). It has been shown that the antiviral activity of saturated aldehydes was negli- gible when compared with that of the corresponding alde- hydes with an a,p-unsaturated group (Schauenstein et al., 1977), and it was concluded that this was the active portion of the molecule. Antiviral activity of trans-2-hexenal was thought to be due to the effect of protein cross-linking and modifications of sulfhydryl and amino groups (Schauenstein et al., 1977). The sulfhydryl groups of proteins and of com- pounds like GSH are generally essential to their function. Detoxification of a,P-unsaturated aldehydes is possible by enzyme-catalyzed addition to GSH, oxidation to less toxic unsaturated acids, or reduction to alcohols (Schauenstein et al., 1977), although the alcohols, such as cis-3-hexenol, still possess some antimicrobial activity (Fig. 8).
Hydroperoxide lyase activity produces the CI2 and c6 moieties that are further metabolized to traumatic acid and the various c6 aldehydes and alcohols, respectively (Fig. 1). However, traumatic acid was not detected under the condi- tions used here for GC-MS. Traumatic acid, which is pro- duced from both linoleic acid and linolenic acids, is associated with wounding and cell proliferation in plants (Vick and Zimmerman, 1987a) and causes abscission in cotton buds (Strong and Kruitwagen, 1967). It appeared to have no effect on the rate of growth of bacterial cells in liquid culture at the concentrations tested, and we conclude that a direct antibac- teria1 role is unlikely. Other effects of traumatic acid on the plants themselves, e.g. the induction of phytoalexins, were not measured.
The actual quantity of volatiles produced in leaves is almost certainly greater than that measured in the head-space pro- cedure, which can only detect those that escape from the plant. Thus, certain amounts will react with cell contents, bacteria, etc. and will in effect be titrated out. A similar situation applies in the agar-plug assay, where an increase in the bacterial inoculum added to the overlay reduces the size of the zone of inhibition. Presumably, the bacterial cells bind with and thus titrate out the truns-2-hexenal. The data pre- sented here show the earliest time at which we can detect release of volatiles from diseased leaves and do not mean that biologically significant quantities of these active sub- stances are not being produced earlier.
Thus, we hypothesize that the lipid-derived C6-volatiles produced during HR cell death in plants may play an anti- pathogen role in plant defense responses, behaving like vol- atile phytoalexins. The early timing of production of the c6 aldehydes and alcohols may mean that they are important in the early plant resistance responses because they are detect- able in relatively high concentrations at between 15 and 24 h after inoculation. By 15 to 24 h after inoculation with
avirulent bacteria, the primary leaves showed extensive hy- persensitive cell collapse. At this stage, each pair of primary leaves was evolving approximately 2.2 pg of trans-2-hexenal and 10.3 pg of cis-3-hexenol per 30 min of collection time. This would be equivalent to approximately 40 pg of trans-2- hexenal and approximately 185 pg of cis-3-hexenol, respec- tively, over the 9-h period. With such large amounts of the volatile substances being lost from leaves, it seems fair to assume that the effective concentrations, particularly for frans-2-hexenal, at sources within the leaves might be in the range that would affect bacterial growth. Duniway (1973) showed that P. syringae induced stomatal closure in tobacco during an HR. Assuming this is also true in bean, it would further support the suggestion that toxic levels of volatiles might be reached in the intercellular spaces.
Earlier work with this pathosystem showed that numbers of both virulent and avirulent bacterial isolates increased in leaves for the first 24 h after inoculation and that around the time of HR cell collapse the rate of multiplication of the avirulent isolate began to decline (Lyon and Wood, 1975, 1976). Similar trends were shown for the growth of compat- ible P.s. pv syringae and the nonhost plant pathogenic bac- terium P. coronafaciens (HR-eliciting) in bean tissues (Daub and Hagedorn, 1980). Because the above studies employed different inoculation procedures, inoculum concentrations, and plant growth conditions, we carried out our own studies on the growth of virulent and avirulent isolates of P.s. pv phaseolicola in bean leaves.
In agreement with the earlier studies (Lyon and Wood, 1975, 1976; Daub and Hagedorn, 1980), our results also showed a divergence in growth rates between virulent and avirulent races over the first 48 h after inoculation. Growth of the avirulent race 1 cells appeared to be depressed quite early compared with virulent race 3 cells, and race 1 cells had reached their maximum leaf density by 24 h after inoc- ulation (Fig. 9). In the compatible combination the bacteria reached the maximum density usually observed for this pa- thosystem of approximately 1 x 109 cfu cm-2 of leaf (Young, 1973) by 48 h after inoculation, but after 24 h, as the density approaches this level, the rate of multiplication slows (Fig. 9). Thus, it seems possible that trans-2-hexenal and perhaps to a lesser extent the other lipid-derived volatiles may be in part responsible for the early suppression of bacterial multi- plication observed in the HR, either by killing a proportion of the bacteria directly or by suppressing the growth rate of the population as a whole. However, it must be stressed that, as with the accumulation of "conventional" isoflavonoid phy- toalexins in bean, a direct antimicrobial action in vivo has not yet been demonstrated, and, because the magnitude of the effect at the time of production is not large (Fig. 9), fatty acid-derived volatiles may not form a major component of the plant's defense repertoire. However, the well-described isoflavonoid phytoalexins of bean (e.g. phaseollin, coumes- trol, kievitone), begin to accumulate only at 24 h after infec- tion and reach maximal values at 48 h or more (Lyon and Wood, 1975, 1976). Thus, it seems unlikely that phytoalexins are responsible for the early suppression of bacterial growth in the incompatible combination, but they might contribute largely to the later stages of inhibition and are perhaps also
Dow
nloaded from https://academ
ic.oup.com/plphys/article/101/1/13/6065904 by guest on 01 January 2022

Antibacterial Activity of Lipid Breakdown Products 23
important in resisting colonization of necrotic tissues by secondary, opportunistic invaders.
Because truns-2-hexenal damaged the plants and at high concentrations caused necrosis, it is of course possible that it contributes to causing the host cell death seen in the HR. However, large amounts were produced only after visible necrosis had occurred, and it seems likely that #runs-2-hexenal is produced rather as a consequence of cell collapse. It does not appear to play a role in signal transduction because hexenal-treated plants did not accumulate phytoalexins.
It is interesting that no saturated C6 alcohols or aldehydes, which would derive from linoleic acid-13-hydroperoxide, were detected. Because inoculated plants evolved pentane (Croft, 1990), peroxidation of linoleic acid does occur during the HR; therefore, we conclude that in the HR any linoleic acid 13-hydroperoxide that forms must be metabolized via other pathways, perhaps those utilizing dehydrase (isomerase and cyclase) activities. Thus, the metabolites would not feed into the pathway leading to these C6 compounds. This ob- servation, together with the apparent lack of production of trans-2,cis-6-nonadienal (see above), suggests that the per- oxidation of fatty acids, which specifically accompanies the HR in bean, is under tightly regulated metabolic control.
ACKNOWLEDCMENTS
We thank John Friend for productive discussions and encourage- ment, Janice Halder (University of Hull, UK) for help with the EM, Paul Ward (University of Ziirich) for help with the statistical analysis, and Robert Dudler for critical reading of the manuscript.
Received July 6, 1992; accepted September 23, 1992. Copyright Clearance Center: 0032-0889/93/101/0013/12.
LITERATURE ClTED
Anderson JM (1989) Membrane derived fatty acids as precursors to second messengers. In WF Boss, DJ MorrP, eds, Second Messengers in Plant Growth and Development, Plant Biology Vol 6. Alan R Liss, New York, pp 181-212
CRC Handbook of Chemistry and Physics, Ed 70, Section C-308. CRC Press, Boca Raton, Florida
Croft KPC (1990) The hypersensitive response of bean inoculated with Pseudomonas syringae pv. phaseolicola race 1. PhD thesis. University of Hull, Hull, UK
Croft KPC, Voisey CR, Slusarenko AJ (1990) Mechanism of hyper- sensitive cell collapse: correlation of increased lipoxygenase activity with membrane damage in leaves of Phaseolus vulgaris (L.) cv. Red Mexican inoculated with avirulent race 1 cells of Pseudomonas syringae pv. phaseolicola. Physiol Mo1 Plant Pathol 36 49-62
Daub ME, Hagedorn DJ (1980) Growth kinetics of Pseudomonas syringae with susceptible and resistant bean tissues. Phytopathol-
Duniway JM (1973) Pathogen-induced changes in host water rela- tions. Phytopathology 63 458-466
Epton HAS, Devera11 BJ (1968) A biochemical difference between healthy bean leaves resistant and susceptible to the halo blight disease caused by Pseudomonas phaseolicola. Ann App Biol 61:
Farmer EF, Ryan CA (1992) Octadecanoid precursors of jasmonic acid activate the synthesis of wound-inducible proteinase inhibi- tors. Plant Cell4: 129-134
Galliard T (1978) Lipolytik and lipoxygenase enzymes in plants and their action in wounded tissues. In G Khal, ed, The Biochemistry of Wounded Plant Tissues. Walter de Gruyter, Berlin, New York,
Ogy 70 429-436
255-268
pp 155-201
Galliard T, Phillips DR, Reynolds J (1986) The formation of cis-3- nonenal, trans-2-nonenal and hexanal from linoleic acid hydro- peroxide isomers by a hydroperoxide cleavage enzyme system in cucumber fruits. Biochim Biophys Acta 441: 181-182
Gunstone FD (1967) An Introduction to the Chemistry and Biochem- istry of Fatty Acids and their Glycerides, Ed 2. Chapman & Hall, London, New York, pp 92-114
Gunstone FD (1986) Chemical properties 10.2. Autoxidation and photo-oxygenation. In FD Gunstone, JL Harwood, EB Padley, eds, The Lipid Handbook. Chapman & Hall, London, New York, pp 453-457
Hatanaka A, Harada T (1973) Formation of cis-3-hexena1, trans-2- hexenal and cis-3-hexenol in macerated Thea sinensis leaves. l'hy- tochemistry 12: 2341-2346
Hatanaka A, Kajiwara T, Sekiya J (1987) Biosynthetic pathway for C6-aldehyde formation from linolenic acid in green leaves. Chem Phys Lipids 44 341-361
Hoppe HH, Heitefuss R (1974) Permeability and membrane lipid metabolism of Phaseolus vulgaris infected with Uromyces phaseoli. I. Changes in the efflux of cell constituents. Physiol Plant Pathol 4: 5-9
Jiittner F (1988) Quantitative trace analysis of volatile organic com- pounds. Methods Enzymoll67: 600-616
Keppler LD, Novacky A (1986) Involvement of membrane lipid peroxidation in the development of a bacterially induced hyper- sensitive reaction. Phytopathology 7 6 104-107
Keppler LD, Novacky A (1987) The initiation of membrane lipid peroxidation during bacteria-induced hypersensitive reaction. Physiol Mo1 Plant Pathol 3 0 233-245
Konze JR, Elstner EF (1978) Ethane and ethylene formation by mitochondria as an indication of aerobic lipid degradation in response to wounding of plant tissue. Biochim Biophys Acta 528
Kunert K-J, Tappel AA (1983) The effect of vitamin C on in vivo lipid peroxidation in guinea pigs as measured by pentane and ethane evolution. Lipids 18 271-274
Leshem Y (1984) Caz+: calmodulin-induced and ethylene mediated phospholipid catabolism as a mode of plant senescence. In PA Siegenthaler, W Eichenberger, eds, Structure, Function and Metab- olism of Plant Lipids. Elsevier Science Publishers, Amsterdam, pp 181-188
Lyon F, Wood RKS (1975) The hypersensitive reaction and other responses of bean leaves to bacteria. Ann Bot 40 479-491
Lyon F, Wood RKS (1976) Alteration of response of bean leaves to compatible and incompatible bacteria. Ann Bot 41: 359-367
Lyr H, Basaniak L (1983) Alkenals, volatile defense substances in plants, their properties and activities. Acta Phytopathol Acad Sci Hung 1 8 3-12
Major RT, Marchini P, Sproston T (1960) lsolation from Ginkgo biloba (L.) of an inhibitor of fungus growth. J Biol Chem 235:
Matthew JA, Galliard T (1978) Enzymic formation of carbonyls from linoleic acid in leaves of Phaseolus vulgaris. Phytochemistry
Ocampo CA, Moerschbauer 8, Grambow HJ (1986) Increased li- poxygenase activity is involved in the hypersensitive response of wheat leaf cells infected with avirulent rust fungi or treated with funga1 elicitor. 2 Naturforsch 41c: 559-563
Ohta H, Shida K, Morita Y, Peng YL, Furusawa I, Shishiyama J, Aibara S, Morita Y (1990) The occurrence of lipid hydroperoxide decomposing-enzyme activities in rice and the relationship of such activities to the formation of antifungal substances. Plant Cell Physiol31: 1117-1122
Rogers KR, Albert F, Anderson AJ (1988) Lipid peroxidation is a consequence of elicitor activity. Plant Physiol86 547-553
Schauenstein E, Esterbauer H, Zollner H (1977) Aldehydes in biological systems-their natural occurrence and biological activi- ties. Translated by PH Gore. Pion Limited, London, pp 32-35 and
Siedow JN (199 1) Plant lipoxygenase: structure and function. Annu
Slater TF (1984) Overview of methods used for detecting lipid
2 13-221
3298-3299
17: 1043-1044
40-41
Rev Plant Physiol Plant Mo1 Biol42' 145-188
peroxidation. Methods Enzymol105 283-293
Dow
nloaded from https://academ
ic.oup.com/plphys/article/101/1/13/6065904 by guest on 01 January 2022

24 Croft et al. Plant Physiol. Vol. 101, 1993
Slusarenko AJ, Croft KPC, Voisey CR (1991) Biochemical and molecular events in the hypersensitive response of bean to Pseu- domonas syringae pv. phaseolicola. In CJ Smith, ed, Biochemistry and Molecular Biology of Host-Pathogen Interactions. Clarendon Press, Oxford, UK, pp 126-143
Slusarenko AJ, Longland AC, Whitehead IM (1989) A convenient, sensitive and rapid assay for antibacterial activity of phytoalexins. Bot Helv 92: 203-207
Strong FE, Kruitwagen E (1967) Traumatic acid: an accelerator of abscission in cotton explants. Nature 21: 1390-1391
Takeo T (1992) Green and semi-fermented teas. In KC Wilson, MN Clifford, eds, Tea: Cultivation to Consumption. Chapman & Hall, London, pp 413-457
Vick BA, Zimmerman DC (1984) Biosynthesis of jasmonic acid in plants. Plant Physiol 75: 458-461
Vick BA, Zimmerman DC (1987a) Oxidative systems for modifica-
tion of fatty acids: the lipoxygenase pathway. In PK Stumpf, ed, Lipids: Structure and Function. The Biochemistry of Plants, Vol9. Academic Press, New York, pp 53-90
Vick BA, Zimmerman DC (1987b) Pathways of fatty acid hydro- peroxide metabolism in spinach leaf chloroplasts. Plant Physiol85
Von Schildknecht H, Rauch G (1961) Die chemische Natur der "Luftonphytoncide" von Blattpflanzen insbesondere von Robinia pseudacacia. Z Naturforsch 16b 422-429
Yamamoto H, Tani T (1986) Possible involvement of lipoxygenase in the mechanism of resistance of oats to Puccinia coronata avenae. J Phytopathol116 329-337
Young JM (1973) Development of bacterial populations in vivo in relation to plant pathogenicity. NZ J Agric Res 17: 105-113
Zeringue HJ, McCormick SP (1989) Relationships between cotton leaf-derived volatiles and growth of Aspergillus flavus. J Am Oil Chem SOC 66 581-585
1073-1078
Dow
nloaded from https://academ
ic.oup.com/plphys/article/101/1/13/6065904 by guest on 01 January 2022