vol. 253, no. 12, issue of june 25, pp. 44084418, 1978 ... · turnover of plasma membrane...
TRANSCRIPT

Vol. 253, No. 12, Issue of June 25, pp. 44084418, 1978 Printed in U.S. A.
Turnover of Plasma Membrane Glycoproteins and Glycolipids of Hepatoma Tissue Culture Cells*
(Received for publication, December 8, 1977)
Heinz Baumann$ and Darrell Doyle
From the Department of Molecular Biology, Roswell Park Memorial Znstitute, Buffalo, New York 14263
Tritiated borohydride reduction of cells treated in situ with galactose oxidase and neuraminidase was used to label externally oriented glycoproteins and gly- colipids of the plasma membrane. The tritium was in- corporated into galactose and galactosamine residues of both glycoproteins and glycolipids of two rat hepa- toma cell types-monolayer H-35 cells and suspension cultures of HTC cells. The glycoproteins with accessible externally oriented galactose residues also were iodi- nated when cells in situ were subjected to lactoperoxi- dase-catalyzed iodination. Thus, tyrosine residues as well as galactose residues of the plasma membrane glycoproteins could be labeled in situ in these hepatoma cells. The label incorporated into the carbohydrate res- idues of externally oriented membrane glycoproteins was lost during subsequent cell culture in a biphasic manner. Two-thirds of the label incorporated into gly- coprotein was lost with a half-life of about 1 day. The rest of the label was lost more slowly corresponding to a half-life of 100 h. Iodinated membrane glycoproteins isolated by concanavalin A chromatography also showed biphasic turnover with nearly identical kinet- ics to that shown by the total galactose-labeled plasma membrane glycoproteins.
Contributing significantly but not solely to the early rapid phase of glycoprotein degradation was a rapidly turning-over trypsin-sensitive plasma membrane gly- coprotein(s) in each of the two different hepatoma cells. Fragments of these glycoproteins were found in serum- free medium and were similar to fragments released from cells treated with trypsin. Thus, some surface glycoproteins are degraded by membrane proteases or cell proteases released into the medium. Most of the other labeled glycoproteins were degraded at similar rates to small molecular weight material. The glycolip- ids of the plasma membrane also were degraded with a half-life of 100 h. These results suggest that the main pathway for plasma membrane degradation is by inter- iorization and degradation of structural membrane units.
We are using hepatoma tissue culture (HTC) cells, a sus- pension culture of epithelial cells derived originally from a minimal deviation rat hepatoma (1, 2) as a system to examine the mode of plasma membrane biogenesis in mammalian cells. The HTC cell has lost the regional specializations, such as bile canaliculi, tight and gap junctions, etc., that characterize the
* This investigation was supported by United States Public Health Service Research Grants CA17149 and GM19521. The costs of pub- lication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertise- merit” in accordance with 18 USC. Section 1734 solely to indicate this fact.
$ Postdoctoral Fellow of the Swiss National Science Foundation.
plasma membrane of rat liver hepatocytes. But we reasoned that a cell growing in suspension culture with a relatively undifferentiated and structurally homogeneous surface might provide an experimentally manipulatable model system to examine the synthesis, assembly, and turnover of the polypep- tides forming the plasma membrane. Thus far, we have shown that the plasma membrane of this cell has a set of externally disposed polypeptides which are accessible to lactoperoxidase- catalyzed iodination (3). There are at least 50 to 60 such externally oriented polypeptides in the membrane, as resolved by sodium dodecyl sulfate-polyacrylamide gel electrophoresis. Indeed, they constitute the bulk of the protein comprising the plasma membrane. Most and possibly all of these iodinated polypeptides are degraded or lost from the membrane at equivalent rates, implying that the plasma membrane of HTC cells turns over as a structural unit (4). Unit turnover of the major polypeptides forming the membrane places certain restrictions on the mode of biogenesis of this cell organelle (5).
The question arises of whether the mode of turnover of the polypeptides accessible in situ to iodination, which are the more abundant proteins in the membrane, is representative of the turnover of all the proteins in the membrane, including the less abundant polypeptides. The latter class might include glycoprotein receptors for specific hormones (6, 7) or for other small and large molecules in the serum (8-11). In this respect, we have examined previously the turnover of the sugar moie- ties of fucose-labeled plasma membrane glycoproteins of HTC cells (4). In contrast to the loss of incorporated iodide, fucose in these glycoproteins showed some heterogeneity in its turn- over, suggesting that either the fucose-labeled glycoproteins were a different class of membrane proteins in terms of turnover properties, or that the carbohydrate residues of these membrane glycoproteins were turning over independently of the amino acid backbone as labeled with iodide. However, the use of fucose as a marker for plasma membrane glycoproteins introduces complications because fucose-containing glycopro- teins are shared by the plasma membrane and by an internal membrane system (12, 13). In this paper then, we used triti- ated borohydride reduction of galactose oxidase-treated cells to label specifically externally disposed plasma membrane glycoproteins and glycolipids containing accessible galactose residues (14-17). This method of in situ labeling introduces tritium into both glycoproteins and glycolipids, allowing a comparison of the turnover properties of both of these surface membrane constituents. To ensure that the results obtained were not peculiar to only HTC cells which is a suspension cell culture of rat hepatoma origin, we also examined the fate of the incorporated tritium in a different but related epithelial cell, H-35, which was independently derived from a rat hepa- toma and which grows only in monolayer culture. The results of these labeling studies together with previous studies of the turnover of iodinated plasma membrane proteins indicate that
4408
by guest on June 5, 2018http://w
ww
.jbc.org/D
ownloaded from

Plasma Membrane Glycoproteins of Hepatoma Cells 4409
in both suspension cultures of HTC cells and in monolayer cultures of Reuber H-35 cells, there is some heterogeneity in
the turnover of the sugar moieties of some surface glycopro- teins. These glycoproteins may represent a class of membrane proteins which are removed from the surface and degraded by a different mechanism than that involved in the turnover of the more abundant plasma membrane proteins.
EXI’EItIMEN’I’AL 1’IWCEL)UIW.S
Cell Culture-A cloned line of hepatoma tissue culture (HTC) cells was grown in suspension culture in Eagle’s minimal essential medium containing 10% fetal calf serum as described previously by Tweto et al. (3). H-35 (Reuber) hepatoma cells were grown in monolayer in Dulbecco’s modified Eagle’s medium containing 5% fetal calf serum, 5% calf serum, and 25 mM 4-(2-hydroxyethyl)-I-piperazineethanesul- fonic acid (Hepes) buffer. The H-35’ cultures were kept under an atmosphere of 95% air and 5% COZ.
Radioactive Labeling-Externally oriented plasma membrane gly- coproteins and glycolipids were labeled with tritiated sodium boro- hydride after galactose oxidase treatment of the cell surface according to the procedure of Gahmberg and Hakomori (14). Dulbecco’s phos- phate-buffered salt solution was used as solvent throughout the labeling procedure. The density of HTC cells was kept for all chemical and enzymatic treatments at 5 X 10“ cells/ml; for each 25 cm” confluent monolayer of H-35 cells (33 x 10” cells), 2 ml of solution was used. Before oxidizing available galactose residue with galactose oxidase, the cells were treated with 1 mM NaBH, for 5 min at 20°C to eliminate any potentially reducible residues on the cell surface. Next, terminal sialic acids were removed by treatment of the cells with neuraminidase from Vibrio cholerae (Calbiochem) using a concentra- tion of 5 units/ml for 15 min at 37°C. The terminal galactose and galactosamine residues then were oxidized using highly purified ga- lactose oxidase (a generous gift of Dr. M. Ettinger, State University of New York at Buffalo, 3000 units/m& at a concentration of 6 units/ml for 15 min at 37’C. The aldehydes were reduced with NaB’H., (Amersham, 8 to 10 Ci/mmol) at a concentration of 1 mCi/ml for 5 min at 20°C. NaB”H4 was dissolved immediately before use in ice-cold 0.01 N NaOH.
HTC cells were enzymatically iodinated with laiI or ‘,“I (Amersham, carrier-free sodium salt) according to the procedure of Tweto et al. (3), which is a modification of the method of Phillips and Morrison (18) and Hubbard and Cohn (19). In some experiments Na”I was included in the labeling mixture. Neither iodination nor tritiated borohydride reduction, as described here, affects cell viability, as measured by exclusion of trypan blue. In all cases, the viability was 98% or greater.
Cell Extraction and Radioactivity Determinations+‘H]Borohy- dride-labeled and washed H-35 cells were scraped from the support in Dulbecco’s phosphate-buffered salt solution containing 1 mM phen- ylmethylsulfonyl fluoride (Sigma, 2 ml/25 cm’ confluent monolayer) and immediately precipitated with 10% trichloroacetic acid. Labeled and saline-washed HTC cells at a concentration of 5 x IO” cells/ml were precipitated with 10% trichloroacetic acid. The cells treated with trichloroacetic acid were sonicated, kept for 1 h at 4’C!, and centri- fuged. The supernatant fractions (acid-soluble) were counted directly in a Beckman Biogamma counter or a scintillation counter depending on label, or the supernatant fractions were extracted twice with an equal volume of diethyl ether. An aliquot of the aqueous phase resulting from the ether extraction was dried in a scintillation vial and the residue dissolved in 1 ml of 90% NCS solubilizer (Amersham) by incubation for 1 h at 55°C. The radioactivity was measured after addition of 10 ml of scintillation fluid in a Beckman liquid scintillation counter.
The acid-insoluble cell fraction was extracted twice with 10% trichloroacetic acid and then sonicated in chloroform/methanol (2:l). This mixture was kept for 1 h at 4°C. After centrifugation, the supernatant fraction (chloroform/methanol-soluble “lipids”) was transferred to a test tube. The residue was extracted once again with chloroform/methanol (2:l). The second supernatant was combined with the first and mixed thoroughly. An aliquot was removed and counted directly in the gamma counter or was dried in a scintillation vial, solubilized in NCS solubilizer, and counted in the scintillation counter. Precipitation with trichloroacetic acid itself does not alter the kind or amount of labeled glycolipids as judged by thin layer
’ The abbreviation used is: H-35 cells, Reuber hepatoma cells.
chromatographic analysis of cells extracted directly with chloro- form/methanol and cell treated first with trichloroacetic acid. Fur- ther, analysis of the chloroform/methanol-soluble fraction by thin layer chromatography and dodecyl sulfate-polyacrylamide electro- phoresis after desalting by LH-20 Sephadex chromatography shows this fraction to contain a mixture of labeled glycolipids with little or no material with molecular weight below 800 or greater than’12,OOO.
Identification of Labeled Products after Galactose Oxidase Treat- ment-HTC cells (2.5 x 10’) were labeled after neuraminidase and galactose oxidase treatment with NaB’H,. After precipitation with trichloroacetic acid, the insoluble material was extracted twice with chloroform/methanol (2:l). The combined extracts were desalted by chromatography over LH-20 Sephadex in chloroform:methanol:water (2:1:0.05). Aliquots of the chloroform/methanol-soluble fraction (“lip- ids”) and insoluble fraction (“proteins”), each containing 100,000 cpm, were methanolyzed and N-acetylated according to Chambers and Clamp (21). In the lipid sample 95%) and in the protein sample 92%’ of the radioactivity was liberated and appeared in the methylglycoside fraction. The carbohydrates were trimethylsililated and chromato- graphed on 3% SE30’in a gas-liquid chromatography (Hewlett Pack- ard; Ref. 22). The eluted carbohydrates were collected by passing the gas through glass capillaries cooled by dry ice. The capillaries were washed with scintillation fluid and the radioactivity was determined. Using I.-[S ,‘H]fucose and I)-[6 “Hlgalactose (Amersham) as internal standards throughout the analysis recoveries of 57 and 59% respec- tively, were found. For determination of the retention times, a stan- dard mixture of r,-fucose, I)-galactose, I)-glucose, N-acetylgalactosa- mine, N-acetylglucosamine, and N-acetylneuraminic acid were ana- lyzed.
Trypsinization-HTC cells were suspended in phosphate-buffered saline or Earle’s balanced salt solution at a concentration of 5 x 10” cells/ml. H-35 cells were covered with phosphate-buffered saline, 2 ml/25 cm’ monolayer. To the cells was added aliquots of a trypsin solution (1 mg of tosyl phenylalanyl chloromethyl ketone-treated trypsin, Worthington, 276 units/mg in 1 ml of 0.001 N HCl), yielding the concentrations indicated in the legend to the appropriate figure or table. The cells were digested for different times at various tem- peratures (see “Results”). After addition of 1 1nM phenylmethylsul- fonyl fluoride, the incubation mixture was centrifuged for 5 min at 406 x g,,;,,. The resulting supernatant fractions were centrifuged again for 60 min at 200,000 X g,,,,. The proteins were precipitated with 10% trichloroacetic acid, washed twice with 10% trichloroacetic acid and ethanol, and in the amount determined (20). The cells in all cases were greater than 95% viable after trypsin treatment.
Concanavalin A-Sepharose Chromatography-Externally labeled HTC cells were treated without prior fractionation by sonication in 1% sodium deoxycholate, 10 mM Tris-HCl, pH 8.0, containing 0.5 mM phenylmethylsulfonyl fluoride at 0°C using a concentration of 5 x IO” cells/ml. The resulting extract was centrifuged for 60 min at 200,000 x grX,;,“. The supernatant fraction contained 70%) of the total protein bound radioactive iodide or 9OR of the total galactose-labeled mate- rial. The deoxycholate-soluble fraction was applied at a flow rate of 10 ml/h to a column (0.8 x 4 cm) of concanavalin A-Sepharose (Pharmacia). The column was prewashed with 400 ml of the deoxy- cholate/Tris buffer. The bound proteins were eluted after incubation of the column for 16 h with 0.5 M a-methylmannoside in deoxycho- late/Tris buffer. The eluted proteins were precipitated with trichlo- roacetic acid, lo%, and were washed twice with ethanol. The recovery of galactose-labeled glycoprotein from the deoxycholate-soluble frac- tion ranged between 20 to 300/o, while between 5 and 10% of the total iodinated protein was bound to the lectin column.
Gel Electrophoresis-Electrophoretic separations of the labeled proteins were performed in 7.5% polyacrylamide slab gels in the presence of 0.1% dodecyl sulfate according to the procedure of La- emmli (23). The following molecular weight markers were used: myosin, 210 x 10”; ,&galactosidase, 130 x 1O’l; phosphorylase A, 94 x lo”; albumin, 68 x lo”; and ovalbumin, 43 X IO”. Fluorographic detections of the radioactivity in the gels were carried out as outlined by Bonner and Laskey (24). For densitometric analyses of the fluo- rograms, a densitometer from Corning (model 750) was used.
1tE.SIJLT.S
Labeling of Cell Surface-The incorporation of tritium into externally exposed plasma membrane galactosyl and N-ace- tylgalactosaminyl residues of glycoproteins and glycolipids by successive treatment of cells with galactose oxidase and NaE?‘H4 is a widely used technique to label specifically the
by guest on June 5, 2018http://w
ww
.jbc.org/D
ownloaded from

4410 Plasma Membrane Glycoproteins of Hepatoma Cells
TARTX I
Specific radioactivity of externally labeled glycoproteins of HTC and H-35 cells
HTC cells, IO” cells, or confluent monolayers of H-35 cells in 75 cm2 flasks were labeled with NaB”H.,. The following treatments were performed before labeling: none; reduction of cells with 1 tnM unla- beled NaBH,,; reduction with unlabeled NaBR followed by reaction with neuraminidase, galactose oxidase, or neuraminidase, plus galac- tose oxidase (for details, see “Experimental Procedures”). The specific radioactivity in the trichloroacetic acid, chloroform/methanol (2:1)- insoluble fractions are shown and represent the mean of two experi- ments each carried out in duplicate.
Specific activity Enzyme treatment
HTC H-35
cpm X IO- ‘/mg protein
Not oretreated with 3.6 4.2 Na’BH4
Pretreated with mM NaBH,
Pretreated with InM NaBHl
Pretreated with mM NaBHl
Pretreated with mM NaBHA
1 2.9 2.2
1 Neuraminidase 2.8 2.6
1 Galactose oxidase 7.3 7.9
1 Neuraminidase + ga- 44.4 119.9 lactose oxidase
cell surface (14-17). We applied this method of external la- beling to two rat hepatoma cell lines, HTC cells growing in suspension culture and H-35 cells growing in monolayer, in order to follow the turnover of the labeled surface membrane glycoproteins and glycolipids.
The radioactive marker is located almost exclusively at the
cell surface, as shown by electron microscopic autoradiogra- phy of thin sections of labeled HTC cells.” Further support for specific labeling of the external side of the plasma mem- brane is the fact that some of the label is incorporated into glycoproteins, which are completely removed from the surface by trypsin (see below).
The extent of tritium incorporation into surface glycopro- teins is dependent on the way the cells are treated before reduction with labeled NaBH4 (Table I). The reduction of pre-existing reducible groups by incubation of the cells in 1 mM unlabeled NaBH4 lowers the background incorporation almost 50%. The exposure of subterminal galactose residues by removal of terminal sialic acids with neuraminidase en- hances, as expected, the incorporation significantly. The latter treatment increases not only the overall extent of incorpora- tion, but also renders additional glycoproteins available for oxidation via galactose oxidase. Separation of the tritium- labeled glycoproteins on polyacrylamide gels reveals that sev- eral proteins are detectable only after desialylation (see, for example, Figs. 1 and 7). NaB”H4 by itself labels very few proteins. The extent of this “nonspecific” labeling as men- tioned is reduced by pretreating the cells with cold NaBH4. The electrophoretic patterns of the labeled glycoproteins from HTC and H-35 cells are surprisingly different, considering the similar rat hepatoma origin of the two lines (Fig. 1). For example, in H-35 cells, several glycoproteins with apparent molecular weights above 150,000 are labeled; these proteins are not labeled in HTC cells. The electrophoretogram of the labeled material from H-35 cells is dominated by a heavily labeled band moving ahead of the tracking dye. This band is present in the chloroform/methanol (2:l) extract from labeled cells and represents glycolipids. There is much less of this material in HTC cells.
The distribution of incorporated radioactivity after acid and organic solvent extraction of cells is shown for both H-35 and
’ H. Baumann, C. Porter, and D. Doyle, unpublished observations.
HTC cells in Table II. In both cell types, neuraminidase treatment increases the incorporation of tritium into lipids as well as proteins. As mentioned previously, there is much more glycolipid label in H-35 cells relative to HTC cells. Calculated on the basis of equal amounts of cell protein in the two cell types, the H-35 cells contain 13 times more glycolipid acces- sible to reduction with NaBH4 after galactose oxidase and neuraminidase treatment than HTC cells. We did not deter- mine the chemical nature of the labeled compounds present in the trichloroacetic acid-soluble fraction. Volatile radioactiv- ity, representing tritiated hydrogen and water, is also gener- ated during the reduction with borohydride. The amount of volatile radioactivity present after the reaction varies depend- ing on lot of NaB”H4 crystal used in the reaction. The HTC cells generally have more volatile radioactivity associated with them than do the H-35 cells, probably due to the fact that the HTC cells are labeled in suspension.
Carbohydrate analysis of HTC cells, labeled externally by the neuraminidase-galactose oxidase-NaB”H reduction pro- cedures shows that in the lipid fraction, 56% of the total
HTC H-35
MW x IO*
- 68
- 43
-BPB
123 4 5 6 7 8 FIG. 1. Sodium dodecyl sulfate-polyacrylamide gel electrophoresis
of labeled surface glycoproteins from HTC and H-35 cells. HTC cells were treated with NaBH4 (I), galactose oxidase (2), or neuraminidase plus galactose oxidase (3), and confluent monolayers of H-35 cells were untreated (6), treated with unlabeled NaBHr (7), or treated with neuraminidase plus galactose oxidase (8), before labeling with NaB”H4. Glycoproteins also were isolated by concanavalin A-Sepha- rose affinity chromatography from HTC cells treated with neuramin- idase, galactose oxidase, and NaB’H4 (4), or from HTC cells iodinated with ““I (5). Details are given under “Experimental Procedures.” BP& bromphenol blue tracking dye.
TABLE II
Extraction of the radioactivity incorporated into hepatoma cells by acid and organic solvents
Aliquots of 1.4 x IO7 HTC cells and confluent monolayers of H-35 cells in 25-cmL flasks were treated with neuraminidase and galactose oxidase and externally labeled with NaB’R. The labeled cells were extracted first with 10% trichloroacetic acid then with chloroform/ methanol (2:l) and the radioactivity in the two soluble fractions and the remaining insoluble fraction was determined as described under “Experimental Procedures.” The actual counts per min are shown and in parentheses is given the percentage of the total cell radioactiv- ity in each extract. The amount of volatile radioactivity represents the portion not recovered in the three fractions and is expressed as a per cent of the total.
HTC cells treated H-35 cells treated
Fraction with neuraminidase with neuraminidase + galactose oxidase
+ NaB’H, + galactose oxidase
+ NaB’H,
Total cells 793,000 (100%) 1,800,OOO (100%) Trichloroacetic acid-soluble 38,000(5%) 70,000(4%) Chloroform/methanol-solu- i58,OOO (20%) 960,000(53%)
ble lipids Acid and chloroform/meth- 473,000 (59%) 650,000(36%)
anol-insoluble proteins Volatile (14%) (7%)
by guest on June 5, 2018http://w
ww
.jbc.org/D
ownloaded from

Plasma Membrane Glycoproteins of Hepatoma Cells 4411
radioactivity was in galactose and 19% in N-acetyl galactosa- mine, while in the protein fraction, 86% of total radioactivity was in galactose and 2% was in N-acetylgalactosamine.
The data presented in Tables I and II and Fig. 1 show that the carbohydrate moieties of both plasma membrane proteins and gbycolipids of the two hepatoma tissue culture cells are accessible to external labeling via the galactose oxidase and reductive tritiation procedure. We next determined whether the membrane glycoproteins with galactose accessible to ga- lactose oxidase were also accessible to lactoperoxidase-cata- lyzed iodination. Previously, we had shown that the iodide incorporated into HTC cells via lactoperoxidase-catalyzed iodination is present almost exclusively in tyrosines of exter- nally oriented plasma membrane proteins (3).
Only a small percentage of the total iodide incorporated into hepatoma cell membrane proteins is in glycoprotein. Hence, in order to compare those giycoproteins accessible to iodination with those labeled by reduction of oxidized galac- tose residues, the glycosylated proteins must be separated from the nonglycosylated proteins of the membrane. A con- canavalin A-Sepharose affinity chromatographic separation of deoxycholate-solubilized HTC cells was done; the fraction bound to the lectin was analyzed for incorporated iodide and for tritiated galactose (Fig. 1, Slots 4 and 5). The electropho- retie patterns of the differentially labeled glycoproteins show a high degree of similarity, only the relative extent of labeling of the bands being different. The most striking difference is in a band with an apparent molecular weight in the range of 85,000. This band which is very prominent in the galactose- labeled sample, appears to be labeled poorly by iodination. Nevertheless, we consider that most of the glycoproteins accessible to labeling by reductive tritiation of their external galactose residues are also accessible to iodination. Hence, it is possible to compare the turnover of both the amino acid backbone and the carbohydrate side chain of these membrane glycoproteins.
Sensitivity of Externally Labeled Membrane Glycoproteins to Trypsin-The effect of different conditions of treatment with trypsin on the release of externally labeled glycoproteins from HTC cells and H-35 cells is shown in Table III. In both cell types, some of the label incorporated into glycoprotein is sensitive to trypsin, while some is relatively stable to trypsin. In HTC cells, even a low concentration of trypsin (5 to 10 pg/ml) will remove within 10 min much of the labeled glyco- protein which is ultimately protease-sensitive, whereas in H- 35 cells, the removal occurs more progressively with increasing trypsin concentration and time of incubation. A comparison of the trypsin-treated cells by dodecyl sulfate-polyacrylamide electrophoresis shows that the two hepatoma cells contain different glycoprotein bands which are most sensitive to tryp-
TAHLE: III Effect of trypsin on the amount of external label in hepatoma cells
HTC cells, IO’, or eight confluent monolayers of H-35 cells in 25- cm’ flasks were reduced with 1 mM NaBH.,, treated with neuramini- dase and galactose oxidase, and labeled with NaE?‘H., (see “Experi- mental Procedures”). Two aliquots of 10’ HTC cells each and two flasks of H-35 cells were treated with trypsin under the conditions indicated in the table. The specific radioactivity of the proteins from the treated cells was determined and the average of the duplicates is shown.
Trypsin con- centration
P&T/ml
2.5 10 50
Digestion conditions
“C min
25 5 37 10 37 30
Specific activity
HTC H-35
cpm X 10 ‘/mzpr-stein 47.6 124.4 41.3 110.0 33.4 74.7 30.6 61.9
A
MOBILITY
B
After Tryprinkation
No Trypsin
2.5~0 Trypsinhl
GSOlra Trypsinhl
MOBILITY FIG. 2. Effect of trypsin on the surface glycoproteins of HTC and
H-35 cells. HTC and H-35 cells, from the experiment of Table III were labeled with NaB”H* after treatment with neuraminidase and galactose oxidase. The labeled proteins were analyzed by polyacryl- amide-dodecyl sulfate electrophoresis. The densitometric traces of the fluorograms are shown. A, H-35 cells undigested and digested with 2.5 pg of trypsin/ml for 5 min at 25°C. B, HTC cells undigested and digested with trypsin: 2.5 pg/ml for 5 min at 25’C and 50 pg/ml for 30 min at 37°C.
sin (Fig. 2). That is, in each cell type, one band is completely sensitive to the protease. In HTC cells, a labeled glycoprotein with an apparent molecular weight of about 85,000 and in H- 35 one with a molecular weight of 220,000 are removed almost quantitively from the cell surface by the protease.
Endogenous Degradation of Externally Labeled Glycopro- teins-The loss of total externally oriented label introduced into HTC and H-35 cells via reductive tritiation occurs in a biphasic manner (Figs. 3 and 4). During the first 30 to 40 h after labeling of cells treated with neuraminidase and galac- tose oxidase, the radioactivity in the protein fraction disap- pears with a half life of 18 to 23 h. After 48 h of culture, however, the rate of degradation decreases and the half-life approaches a value of about 100 h. A complete replacement of the culture medium by fresh medium after 72 h does not affect the degradation rate. Even daily changes of the medium did not alter the course of degradation in labeled H-35 cells (data not shown).
by guest on June 5, 2018http://w
ww
.jbc.org/D
ownloaded from
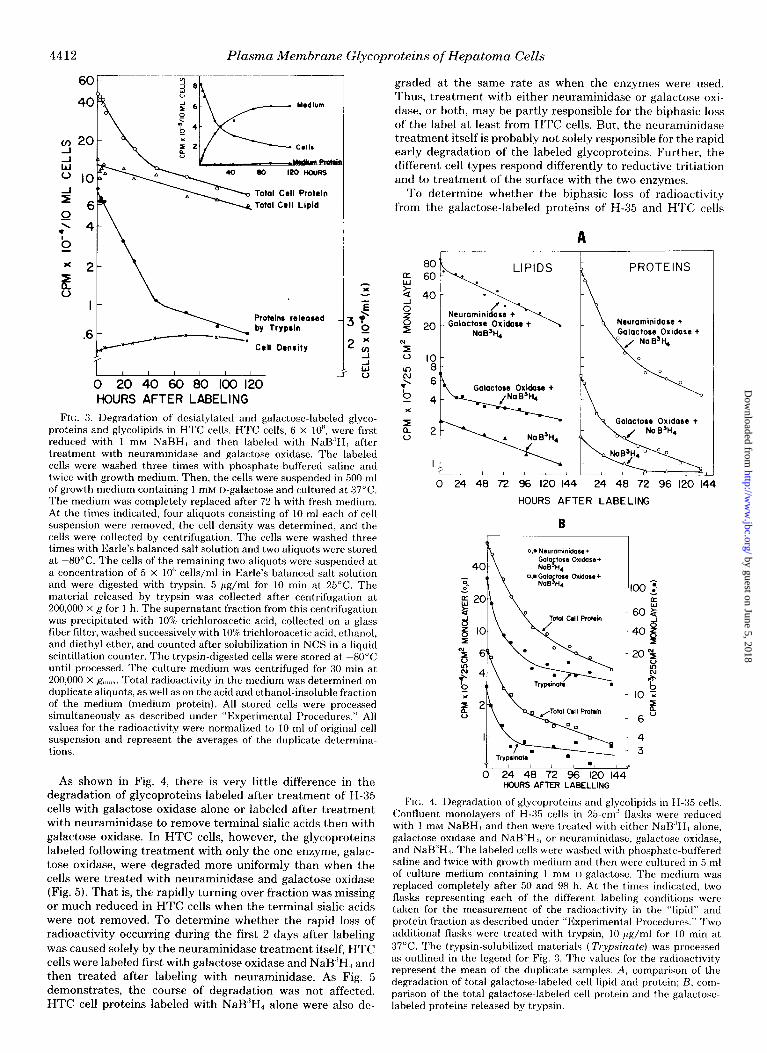
4412 Plasma Membrane Glycoproteins of Hepatoma Cells
i ba 0 20 40 60 80 100 120 HOURS AFTER LABELING
FIN:. 3. Degradation of desialylated and galactose-labeled glyco- proteins and glycolipids in HTC cells. HTC cells, 6 x IOH, were first reduced with 1 mM NaBH., and then labeled with NaB,‘H, after treatment with neuraminidase and galactose oxidase. The labeled cells were washed three times with phosphate-buffered saline and twice with growth medium. Then, the cells were suspended in 500 ml of growth medium containing 1 mM I)-galactose and cultured at 37°C. The medium was completely replaced after 72 h with fresh medium. At the times indicated, four aliquots consisting of 10 ml each of cell suspension were removed, the cell density was determined, and the cells were collected by centrifugation. The cells were washed three times with Earle’s balanced salt solution and two aliquots were stored at -80°C. The cells of the remaining two aliquots were suspended at a concentration of 5 x IO” cells/ml in Earle’s balanced salt solution and were digested with trvpsin, 5 pg/ml for 10 min at 25°C. The material released by trvpsin was collected after centrifugation at 200,000 x c: for 1 h. The supernatant fraction from this centrifugation was precipitated with 10% trichloroacetic acid, collected on a glass fiber filter, washed successive1.v with 10%) trichloroacetic acid, ethanol, and diethyl ether, and counted after solubilization in NCS in a liquid scintillation counter. The trypsin-digested cells were stored at -80°C until processed. The culture medium was centrifuged for 30 min at 200,000 x g-,,,;,,. Total radioactivity in the medium was determined on duplicate aliquots, as well as on the acid and ethanol-insoluble fraction of the medium (medium protein). All stored cells were processed simultaneously as described under “Experimental Procedures.” All values for the radioactivity were normalized to 10 ml of original cell suspension and represent the averages of the duplicate determina- tions.
As shown in Fig. 4, there is very little difference in the degradation of glycoproteins labeled after treatment of H-35 cells with galactose oxidase alone or labeled after treatment with neuraminidase to remove terminal sialic acids then with galactose oxidase. In HTC cells, however, the glycoproteins labeled following treatment with only the one enzyme, galac- tose oxidase, were degraded more uniformly than when the cells were treated with neuraminidase and galactose oxidase (Fig. 5). That is, the rapidly turning over fraction was missing or much reduced in HTC cells when the terminal sialic acids were not removed. To determine whether the rapid loss of radioactivity occurring during the first 2 days after labeling was caused solely by the neuraminidase treatment itself, HTC cells were labeled first with galactose oxidase and NaB”H, and then treated after labeling with neuraminidase. As Fig. 5 demonstrates, the course of degradation was not affected. HTC cell proteins labeled with NaB”H, alone were also de-
graded at the same rate as when the enzymes were used. Thus, treatment with either neuraminidase or galactose oxi- dase, or both, may be partly responsible for the biphasic loss of the label at least from HTC cells. But, the neuraminidase treatment itself is probably not solely responsible for the rapid early degradation of the labeled glycoproteins. Further, the different cell types respond differently to reductive tritiation and to treatment of the surface with the two enzymes.
To determine whether the biphasic loss of radioactivity from the galactose-labeled proteins of H-35 and HTC cells
A
LIPIDS
IO 8 c
PROTEINS
Neuraminidare +
Galactore Oxldase t
0 24 48 72 96 120 144 24 48 72 96 120 144
HOURS AFTER LABELING
B
R
0,. Nsurammid-aae+
40 Galapse Oxidass+ NaB Ii.
i i 24 I 48 I 72 1 96 I 720 / 144' I,
HOURS AFTER LABELLING
FIG. 4. Degradation of glycoproteins and glycolipids in H-35 cells. Confluent monolayers of H-35 cells in 25.cm’ flasks were reduced with 1 mM NaBH, and then were treated with either NaB.‘H, alone, galactose oxidase and NaB’H,, or neuraminidase, galactose oxidase, and NaB”H,. The labeled cells were washed with phosphate-buffered saline and twice with growth medium and then were cultured in 5 ml of culture medium containing 1 mM I)-galactose. The medium was replaced completely after 50 and 98 h. At the times indicated, two flasks representing each of the different labeling conditions were taken for the measurement of the radioactivity in the “lipid” and protein fraction as described under “Experimental Procedures.” Two additional flasks were treated with trypsin, 10 pg/ml for 10 min at 37°C. The trypsin-solubilized materials (T~y~s’innte) was processed as outlined in the legend for Fig. 3. The values for the radioactivity represent the mean of the duplicate samples. A, comparison of the degradation of total galactose-labeled cell lipid and protein; H, com- parison of the total galactose-labeled cell protein and the galactose- labeled proteins released by trypsin.
by guest on June 5, 2018http://w
ww
.jbc.org/D
ownloaded from

Plasma Membrane GLycoproteins of Hepatoma Cells 4413
PROTEIN
IO
Y d 6 u
$4 12
.'
4 2
Medwm total
Medwm Proteln
0 20 40 60 80 100 120 HOURS AFTER LABELLING
FIG. 5. Effect of neuraminidase on the degradation of galactose- labeled glycoproteins and glycolipids in HTC cells. HTC cells were fist reduced with 1 InM NaBH4. One aliquot of cells, 3 x lo’, were next treated with NaB3H4. A second aliquot, 6 x lo*, was treated with galactose oxidase before labeling with NaB3H4. After labeling, one- half of the cells which were treated with galactose oxidase and NaB3H4 were treated with neuraminidase. The differentially treated cells were washed three times with phosphate-buffered saline and
seen in Figs. 3 and 4 is due to the degradation of distinct proteins with different rates of turnover or to a change in the overall rate of protein degradation, the proteins were analyzed by dodecyl sulfate-polyacrylamide gel electrophoresis as a function of time the cells were in culture after labeling (Figs. 6-8). In HTC cells, there is a somewhat faster loss relative to the other labeled proteins of the glycoprotein band with an apparent molecular weight of 85,000 (Figs. 6,8A). In contrast, an HTC cell glycoprotein with an apparent molecular weight of 140,000 may be somewhat more stable relative to the other labeled surface glycoproteins. In H-35 cells, more rapid turn- over of any one protein band is not immediately obvious from the fluorogram of Fig. 7. However, comparison of the densi- tometric traces of the labeled glycoproteins after 0 and 72 h of cell culture reveals a more rapid disappearance of the band with apparent molecular weight of 220,000 (Fig. 8B). In both types of hepatoma cells, the dodecyl sulfate-polyacrylamide gel electrophoretic patterns of the glycoproteins from cells in culture for 72 h after external labeling resembles strikingly the pattern from cells after trypsinization (compare Figs. 8 and 2). That is, the same glycoprotein bands which are more sensitive to trypsin turn over faster in culture. Most of the other labeled glycoproteins in both cell types turn over with approximately equivalent rates (Figs. 6-8).
The amount of labeled surface glycoprotein which can be released by trypsin from HTC cells decreases with similar kinetics as the turnover of the total cell labeled protein (see Fig. 3). Similarly, in H-35 cells, the concentration of labeled surface glycoprotein available in situ to digestion by trypsin mimics the turnover of the total labeled protein (Fig. 4B). The fact that the rate of loss of trypsin-accessible surface glyco- proteins is not higher than the degradation rate of the total glycoprotein indicates that there is no large pool of interiorized labeled glycoproteins in either of these cell types. Further- more, more rapid loss of the trypsin-sensitive proteins cannot be solely responsible for the early rapid phase of surface
LIPID
Galoctose Oxldose +
24 , , , , , , * s
20 40 60 80 100 12(
!i
HOURS AFTER LABELLING
twice with growth medium. Each aliquot of cells then was suspended in 300 ml of growth medium, containing 1 mM D-gdactose. The radioactivity measurements of the cell and medium fractions were carried out as described in the legend to Fig. 3. The medium was obtained from cells labeled by the galactose oxidase and NaB3H4 procedure. The amount of the volatile radioactivity in the medium was estimated by drying the medium for 1 h at 100°C.
0 2 6 10 28 45 69 93 117
FIG. 6. Loss of galactose-labeled plasma membrane glycoproteins from HTC cells. HTC cells labeled via the neuraminidase, galactose oxidase, and NaB’H, procedure were obtained at the times indicated from the culture of the experiment described in Fig. 3. The cells were dissolved directly in dodecyl sulfate sample buffer. Equal amounts of radioactivity (30,000 cpm) were applied to the first four slots of the gel. Because of the amount of radioactivity lost during long term culture, equal amounts of protein had to be applied to the subsequent five slots of the gel. Numbers at the absczssa represent hours after labeling.
glycoprotein turnover. That is, the amount of protein sensitive to trypsin is not sufficient to account for the total amount of label lost from the surface in the early phase of turnover. This is especially true of HTC cells (see Fig. 3). The turnover of external membrane proteins of hepatoma cells not treated with neuraminidase is also rather uniform (Fig. 7)) suggesting again that most of the galactose-labeled surface glycoproteins turn over at the same or similar rates. The pattern of labeled glycoproteins resolved by dodecyl sulfate-polyacrylamide gel electrophoresis appears different between H-35 cells treated with both neuraminidase and galactose oxidase and those treated with galactose oxidase alone (Fig. 7); actually, when the latter cells are treated with neuraminidase after labeling, the pattern of labeled glycoproteins is very similar to that of cells treated with both enzymes before labeling. There is one
by guest on June 5, 2018http://w
ww
.jbc.org/D
ownloaded from

4414 Plasma Membrane Glycoproteins of Hepatoma Cells
Neuraminidase + MW Galactose Oxidase + NaB ‘H4 Galactose Oxidase +NaB3H4
BPB h
0 4 18 26 42 50 66 74 0 4 18 26 42 50 66 74
HOURS AFTER LABELLING FIG. 7. Loss of galactose-labeled plasma membrane glycoproteins
from H-35 cells. H-35 cells labeled either by neuraminidase, galactose radioactivity (30,000 cpm) were applied to the slots representing the
oxidase, and NaB”H4 or by galactose oxidase and NaB”H4 were 0-, 4-, I&, and 26-h samples from cells labeled by neuraminidase,
obtained at the times indicated from the experiment described in Fig. galactose oxidase, and NaB’H+ In all other slots, equal amounts of protein were used (200 pg). The left fluorogrum was exposed for 1
4. The monolayers were scraped from the flask, centrifuged, and week and the right one, for 1 month. dissolved in sodium dodecyl sulfate sample buffer. Equal amounts of
72 HOURS 72 HOURS
MOBILITY
B
difference, however, in that the M, = 220,000, trypsin-sensi- tive, more rapidly turning over surface glycoprotein of H-35 cells is much less accessible to borohydride labeling in cells not treated fist with neuraminidase.
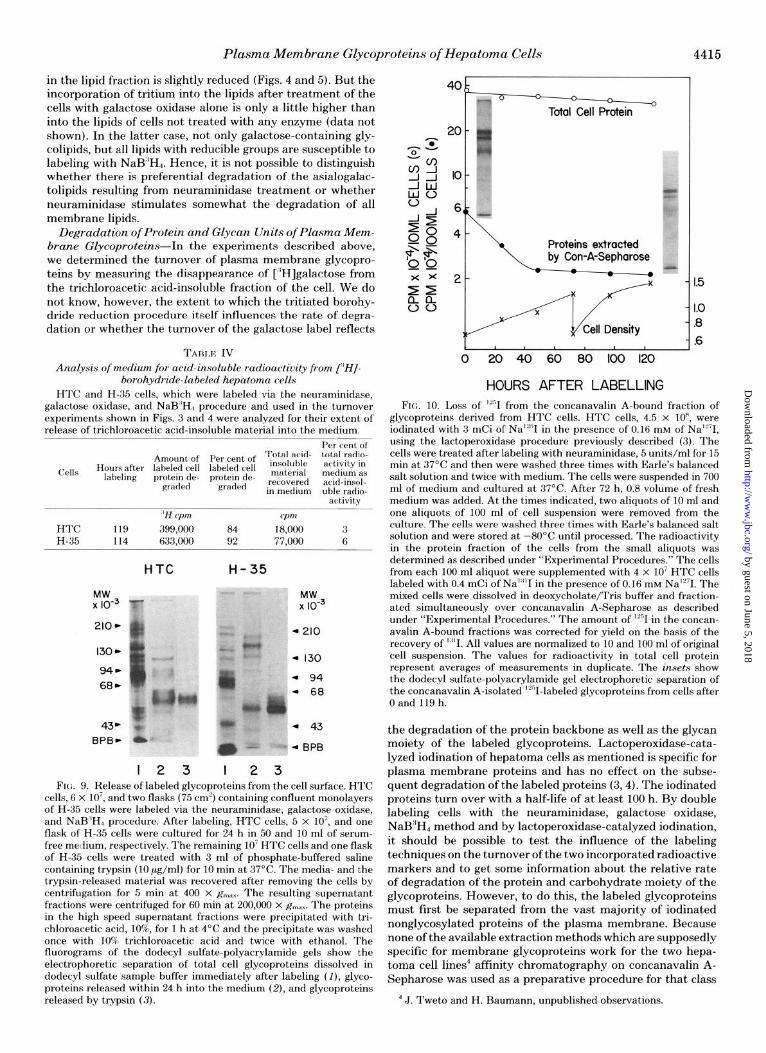
Most but not all of the products of degradation resulting from the turnover of surface-labeled glycoproteins (and gly- colipids) appear in the medium as small molecular weight material soluble in trichloroacetic acid. Only a small percent- age of the radioactivity appearing in the medium is acid- precipitable (Table IV). The acid-insoluble label in the me- dium may represent either “shed” proteins or larger proteo- lytic fragments which are not soluble in acid, or both. The acid-insoluble material released into serum-free medium from the labeled hepatoma cells during 24 h of culture next was examined and the results are shown in Fig. 9. The proteins found in the medium do not correspond to any present in the plasma membrane, suggesting that either shedding does not
MOBILITY
FIG. 8. Densitometric trace of galac- tose-labeled glycoproteins of HTC and H-35 cells in culture 0 and 72 h after labeling. In experiments similar to those described in Figs. 3 and 4, HTC (A) and H-35 (B) cells were labeled via the neur- aminidase, galactose oxidase, and NaB”Hd procedure and cultured for 72 h. The cells were dissolved in dodecyl sul- fate sample buffer 0 and 72 h after cul- ture and the labeled glycoproteins were separated on dodecyl sulfate-polyacryl- amide gels, yielding patterns identical to those in Figs. 6 and 7, respectively. The fluorograms were analyzed densitomet- rically.
occur or that the shed proteins are modified by the cells. It should be noted that one of the major labeled bands present in the medium co-migrates with a glycopeptide, which can be released by trypsin from the cell surface. This acid-insoluble peptide in fact is released by trypsin from the M, = 85,000 band of HTC cells.”
Degradation of Externally Labeled Glycolipids-In con- trast to the turnover of the surface glycoproteins, the glycolip- ids of the plasma membrane do not show a pronounced biphasic course of degradation (Figs. 3 to 5). In both HTC and H-35 cells, the degradation rates of glycolipids labeled via the neuraminidase, galactose oxidase, and NaB”H., method are the same as that observed for the glycoproteins during the second phase of turnover, about 100 h. When the cells were not treated with neuraminidase, the loss of the radioactivity
‘I H. Baumann and D. Doyle, unpublished observations.
by guest on June 5, 2018http://w
ww
.jbc.org/D
ownloaded from
Plasma Membrane Glycoproteins of Hepatoma Cells 4415
in the lipid fraction is slightly reduced (Figs. 4 and 5). But the incorporation of tritium into the lipids after treatment of the cells with galactose oxidase alone is only a little higher than into the lipids of cells not treated with any enzyme (data not shown). In the latter case, not only galactose-containing gly- colipids, but all lipids with reducible groups are susceptible to labeling with NaE?‘H,,. Hence, it is not possible to distinguish whether there is preferential degradation of the asialogalac- tolipids resulting from neuraminidase treatment or whether neuraminidase stimulates somewhat the degradation of all membrane lipids.
Degradation of Protein and Glycan Units of Plasma Mem- brane Glycoproteins--In the experiments described above, we determined the turnover of plasma membrane glycopro- teins by measuring the disappearance of [“Hlgalactose from the trichloroacetic acid-insoluble fraction of the cell. We do not know, however, the extent to which the tritiated borohy- dride reduction procedure itself influences the rate of degra- dation or whether the turnover of the galactose label reflects
TABLE Iv
Analysis of medium for acid-insoluble radioactivity from r’HJ- borohydride-labeled hepatoma cells
HTC and H-35 cells, which were labeled via the neuraminidase, galactose oxidase, and NaB”H, procedure and used in the turnover experiments shown in Figs. 3 and 4 were analyzed for their extent of release of trichloroacetic acid-insoluble material into the medium.
Prr rwlt of
Amount of Per cent of Total acid- total radio-
Hours after labeled cell labeled cell insoluble activity in Cells labeling protein de- protein de- material medium as
graded graded recovered acid-insol- in medium uble radio-
activitv
‘H cpm cpm
HTC 119 399,000 84 18,000 3 H-35 114 633.000 92 77.000 6
HTC H-35
130 -
94,
689
MW x 10-3
4 210
* 130
* 94 - 60
43’ BPB,
* 43
___ *I * BPB
123 I23 FIG. 9. Release of labeled glycoproteins from the cell surface. HTC
cells, 6 x lo’, and two flasks (75 cm’) containing confluent monolayers of H-35 ceils were labeled via the neuraminidase, galactose oxidase, and NaB”H,, procedure. After labeling, HTC cells, 5 x lo’, and one flask of H-35 cells were cultured for 24 h in 50 and 10 ml of serum- free medium, respectively. The remaining IO’ HTC cells and one flask of H-35 cells were treated with 3 ml of phosphate-buffered saline containing trypsin (10 pg/ml) for 10 min at 37°C. The media- and the trypsin-released material was recovered after removing the cells by centrifugation for 5 min at 400 x R.,.~. The resulting supernatant fractions were centrifuged for 60 min at 200,000 x R”,;,~. The proteins in the high speed supernatant fractions were precipitated with tri- chloroacetic acid, 100/o, for 1 h at 4°C and the precipitate was washed once with 10% trichloroacetic acid and twice with ethanol. The fluorograms of the dodecyl sulfate-polyacrylamide gels show the electrophoretic separation of total cell glycoproteins dissolved in dodecyl sulfate sample buffer immediately after labeling (I), glyco- proteins released within 24 h into the medium (Z), and glycoproteins released by trypsin (3).
4
t\ Proteins extracted by Con-A-Sepharose
I I
0 20 40 60 80 100 120
HOURS AFTER LABELLING
1.5
1.0 .8
.6
FIG. 10. Loss of “‘,I from the concanavalin A-bound fraction of glycoproteins derived from HTC cells. HTC cells, 4.5 x IO”, were iodinated with 3 mCi of Nat”1 in the presence of 0.16 mM of NaiL’I, using the lactoperoxidase procedure previously described (3). The cells were treated after labeling with neuraminidase, 5 units/ml for 15 min at 37’C and then were washed three times with Earle’s balanced salt solution and twice with medium. The cells were suspended in 700 ml of medium and cultured at 37°C. After 72 h, 0.8 volume of fresh medium was added. At the times indicated, two aliquots of 10 ml and one aliquots of 100 ml of cell suspension were removed from the culture. The cells were washed three times with Earle’s balanced salt solution and were stored at -80°C until processed. The radioactivity in the protein fraction of the cells from the small aliquots was determined as described under “Experimental Procedures.” The cells from each 100 ml aliquot were supplemented with 4 x lo7 HTC cells labeled with 0.4 mCi of Na”“1 in the presence of 0.16 tnM NalL71. The mixed cells were dissolved in deoxycholate/Tris buffer and fraction- ated simultaneously over concanavalin A-Sepharose as described under “Experimental Procedures.” The amount of “‘,I in the concan- avalin A-bound fractions was corrected for yield on the basis of the recovery of ’ “I. All values are normalized to 10 and 100 ml of original cell suspension. The values for radioactivity in total cell protein represent averages of measurements in duplicate. The rnsets show the dodecyl sulfate-polyacrylamide gel electrophoretic separation of the concanavalin A-isolated ‘L’I-labeled glycoproteins from cells after Oand119h.
the degradation of the protein backbone as well as the glycan moiety of the labeled glycoproteins. Lactoperoxidase-cata- lyzed iodination of hepatoma cells as mentioned is specific for plasma membrane proteins and has no effect on the subse- quent degradation of the labeled proteins (3,4). The iodinated proteins turn over with a half-life of at least 100 h. By double labeling cells with the neuraminidase, galactose oxidase, Nati’H4 method and by lactoperoxidase-catalyzed iodination, it should be possible to test the influence of the labeling techniques on the turnover of the two incorporated radioactive markers and to get some information about the relative rate of degradation of the protein and carbohydrate moiety of the glycoproteins. However, to do this, the labeled glycoproteins must first be separated from the vast majority of iodinated nonglycosylated proteins of the plasma membrane. Because none of the available extraction methods which are supposedly specific for membrane glycoproteins work for the two hepa- toma cell lines4 affinity chromatography on concanavalin A- Sepharose was used as a preparative procedure for that class
4 J. Tweto and H. Baumann, unpublished observations.
by guest on June 5, 2018http://w
ww
.jbc.org/D
ownloaded from

4416 Plasma Membrane Glycoproteins of Hepatoma Cells
020406060Klo HOURS AFTER LABELLIt’
FIG. 11. Degradation of “’ I- and “H-labeled proteins in HTC cells. HTC cells, 5 x IO’, were first iodinated with 5 mCi of Na”“1 in the presence of 0.16 mM Na’“71 and then, after reduction with unlabeled NaBH.,, the same cells were labeled via the neuraminidase, galactose oxidase, and NaB”H, procedure. The cells were washed three times with phosphate-buffered saline and twice with growth medium. The cells were- suspended in 700 ml of growth medium and cultured at 37°C. The medium was replaced completely after 69 h with fresh medium. At the times indicated, aliquots of 50 ml of cell suspension were removed from the culture. The cells were washed three times with Earle’s balanced salt solution and stored at -80°C until proc- essed. The 50-ml aliquots were mixed with unlabeled cells to bring the total cell number to about 10n cells. The cells were solubilized in deoxycholate/Tris buffer and all samples were chromatographed simultaneously on concanavalin A-Sepharose as described under “Ex- perimental Procedures.” The amounts of acid-insoluble “H and ““I were determined on the concanavalin A-bound fractions. The values are normalized to 50 ml of original cell suspension.
of cell glycoproteins that bind to the lectin (see Fig. 1). As shown previously and in Fig. 10, the total iodinated
plasma membrane proteins of HTC cells turn over very slowly following first order or pseudo-first order kinetics (4). In contrast, the ‘““I-labeled glycoproteins which bind to concan- avalin A turnover in the same biphasic manner as the galac- tose-labeled glycoproteins. The degradation rate of the iodi- nate glycoproteins in the second phase (after 40 h) is similar to, but not quite as fast as that of the tritium label introduced into glycoprotein galactose by borohydride reduction. The degradation rate in the second phase is more comparable to the degradation rate of the total iodinated proteins. Sodium dodecyl sulfate-polyacrylamide gel electrophoretic analysis of the concanavalin A-bound fraction from cells immediately and 119 h after iodination do not show any alterations of the patterns (Fig. 10). That is, in contrast, the somewhat hetero- geneous turnover of the “H label introduced into glycoprotein galactose, especially the turnover of the M,. = 85,000 surface glycoprotein (see Fig. BA), the degradation of iodinated sur- face concanavalin A-binding glycoproteins appears homoge- neous. It should be recalled, however, that the M,. = 85,000 glycoprotein is not as accessible to iodination as it is to reductive tritiation (see Fig. 1).
When HTC cells were labeled by both neuraminidase-ga- lactose oxidase-NaB”H4 reduction and lactoperoxidase-cata- lyzed iodination, the course of degradation of the tritium modified glycoproteins (Fig. 11) is as described for the same cells without iodination (Fig. 3). The glycoproteins which bind to concanavalin A lose the iodide and the cH]galactose labels at the same rate (Fig. 11). Furthermore, the biphasic degra- dation of the double-labeled glycoproteins correspond to those of single-labeled glycoproteins (Figs. 3, 10, and 11).
DISCU.SSION
The plasma membrane of hepatoma tissue culture cells is complex in composition containing between 60 and 100 poly-
peptides identifiable by having tyrosine residues accessible for lactoperoxidase-catalyzed iodination (3, 25). Actually, these externally oriented polypeptides comprise the bulk of the protein mass forming the membrane. In previous experiments (3, 4), we had shown that most, if not all, of these membrane polypeptides were degraded with equivalent rates, implying unit turnover of this complex cell organelle. Unit turnover of a large number of polypeptides, each present in the membrane in different molar amounts, imposes certain restrictions on the mechanism(s) of biogenesis of this cell organelle (5). Based primarily on considerations of unit turnover, we proposed and presented evidence for a model describing the pathway of biogenesis of the plasma membrane of hepatoma cells (5, 12, 13, 25). The mode of biogenesis involves synthesis and intra- cellular storage of preformed membrane particles identical in polypeptide composition to the units of plasma membrane lost from the surface via interiorization and degradation. However, our results suggesting unit turnover of membrane polypeptides were based mainly on the study of total iodinat- able polypeptides of the membrane. When we examined the turnover of less abundant plasma membrane glycoproteins using fucose as labeled precursor, there was some heteroge- neity in the rate constants of degradation of the fucose-con- taining membrane proteins (4). However, interpretation of these studies was complicated by the observation that in HTC cells the glycoproteins containing labeled fucose were not confined solely to the plasma membrane (12). Actually, the same glycoproteins were shared by the plasma membrane and by an internal membrane compartment and, consequently, any cross-contamination of one membrane fraction with the other would make the turnover experiments more difficult to interpret. For this reason, in the present manuscript, we used tritiated borohydride reduction in combination with in situ oxidation of cell with galactose oxidase to introduce label into externally oriented galactose residues of plasma membrane glycoproteins and glycolipids.
In both HTC and H-35 cells, there is a set of plasma membrane glycoproteins and glycolipids having galactose res- idues available for borohydride reduction after treatment of the cells with both galactose oxidase and neuraminidase. Most of the same glycoproteins also contain tyrosine residues which are accessible in situ to lactoperoxidase-catalyzed iodination. Indeed, we have compared those plasma membrane glycopro- teins accessible to iodination and able to bind to concanavalin A to those labeled by borohydride reduction of their galactose residues using a two-dimensional method of separation in- volving electrofocusing in the first dimension and dodecyl sulfate-polyacrylamide gel electrophoresis in the second (26, 27); all of the galactose-labeled glycoproteins resolvable by this technique are accessible to iodination.” These dual label experiments together with the electron microscopic autoradi- ographic localization of the incorporated tritium provide con- vincing evidence that the tritium incorporated into galactose and galactosamine is present in externally oriented glycopro- teins of the plasma membrane and not in intracellular glyco- proteins.
The externally oriented galactose-containing proteins that are susceptible to labeling by borohydride reduction differ somewhat between H-35 and HTC cells. However, some of the labeled glycoproteins are shared by the two cell types. These cell lines share a common origin in that they both were derived originally from minimal deviation hepatomas of Buf- falo rats. Differences in glycoprotein composition may have arisen during the time of subsequent culture of the cells or the differences may be involved in the different growth con- ditions of the two cell lines. The H-35 cells grow only in monolayer, while the HTC cells grow in suspension and will
by guest on June 5, 2018http://w
ww
.jbc.org/D
ownloaded from

Plasma Membrane Glycoproteins of Hepatoma Cells 4417
adhere at best only loosely to any solid support. We did not pursue the basis for the differences between these cells in glycoprotein and glycolipid composition here because we were more interested in following the fate of the incorporated label in the two cell types.
In both cell types the tritium incorporated into glycoprotein turns over in a “biphasic” manner. More than 70% of the label incorporated into externally oriented glycoprotein is lost from the membrane during the first 48 h of culture with a half-life of about 1 day. Thereafter, the labeled glycoproteins are turned over more slowly with a half-life of about 100 h, a rate similar to that found here for the galactose-labeled glycolipids of the membrane and for the total membrane proteins acces- sible for iodination (3, 4).
The reason for the biphasic kinetics is not clear, but may be due to a combination of different factors. One factor contrib- uting to the early rapid degradation,barticularly in HTC cells, may be the labeling procedure itself. That is, treatment of these cells with galactose oxidase and neuraminidase tends to make more pronounced the rapid component in the decay curve (compare Figs. 3 and 5). However, even in cells not subjected to the borohydride reduction, but instead labeled by iodination, the labeled glycoproteins still turn over with biphasic kinetics (see Fig. 10).
Another factor which may contribute significantly to the early rapid phase of glycoprotein turnover, but which cannot be solely responsible for it, is the preferential loss from the membrane of the trypsin-sensitive glycoproteins. For example, in HTC cells, almost 90% of the surface glycoprotein sensitive to trypsin is lost from the cell within 40 h of culture (Fig. 3). But, the glycoprotein sensitive to trypsin in situ represents only about 15% of the total label incorporated into surface glycoproteins. In H-35 cells, in contrast, more of the total label incorporated into surface glycoprotein is trypsin-sensitive.
It is also possible that the handling of the cells, such as washing, centrifugation, etc., necessary when the galactose and galactosamine residues are labeled, is partly responsible for the rapid phase of decay. We (4) and others (28) have noticed an initial rapid component to the turnover of plasma membrane proteins after iodination which was attributed to handling of the cells. However, the rapid phase of glycoprotein turnover is both more pronounced and lasts longer than the initial rapid loss of iodinated membrane proteins. Further- more, washing of the cells, centrifugation, changing of the medium, etc., during the slow phase of galactose-labeled gly- coprotein turnover does not alter appreciably the time course of degradation.
An important point about glycoprotein turnover that we wish to emphasize is that, although in both HTC cells and H- 35 cells the overall rate of glycoprotein degradation changes as a function of time in culture, most of the glycoproteins which are accessible to outside labeling have similar rates of degradation in both the rapid and slow phases of their turn- over. That is, the label is lost at about the same rate from every galactose-labeled glycoprotein resolved by dodecyl sul- fate-polyacrylamide gel electrophoresis, with two notable ex- ceptions.
In HTC cells, the glycan moiety of a M, = 85,000 glycopro- tein turns over faster and in H-35 cells a M, = 220,000 glycoprotein has a rate of degradation faster than the other labeled membrane proteins. These two proteins are also more sensitive to trypsin digestion than are the other proteins of the membrane. Indeed, the pattern of labeled glycoproteins present on the cell after trypsin treatment is very similar to the glycopeptide pattern after cells have been in culture for 72 h (compare Figs. 2 and 8). This result suggests that trypsin- like proteases present on the cell surface or present in the
medium may be partly responsible for the degradation of some externally disposed plasma membrane glycoproteins, particularly the M, = 85,000 glycoprotein of H-35 cells. Fur- ther evidence that such a degradative mode may function in these cells is the presence in the growth medium of the same or a very similar acid-insoluble proteolytic fragment as that released from each of these proteins by treatment of the cells with trypsin (Fig. 9).
The main pathway of plasma membrane glycoprotein deg- radation is still probably via interiorization followed b.v fusion of the interiorized membrane with a lysosome. The membrane proteins are then degraded to their constituent amino acids and sugars, which end up in the medium as small molecular weight material soluble in trichloroacetic acid (Figs. 3 and 5; Ref. 4). Very little plasma membrane protein is “shed” intact into the medium from these hepatoma cells.
Based on the very similar half-lives measured for the turn- over of galactose-labeled glycolipids (100 h), most of the proteins accessible to in situ iodination (at least 100 h) and the iodinated or galactose-labeled glycoproteins in the second phase of turnover (100 h), we propose that the interiorized unit of membrane contains both lipid and protein and that the entire membrane unit is degraded in the secondary lyso- some. As we show elsewhere (12), the membrane is then built up from presynthesized units containing both lipid and a complex array of proteins.
Finally, iodide which has been incorporat,ed into tyrosines of externally oriented membrane glycoproteins is lost via turnover at the same rate as is the galactose of the glycan moieties (Fig. 11). This result suggests that the carbohydrate side chains and the amino acid backbone of these proteins are degraded at the same rate or in unison. However, the iodinated glycoproteins by necessity were isolated based on their ability to bind to concanavalin A. It is possible, therefore, for the sugar moieties to be removed from these glycoproteins thus eliminating their affinity for the plant lectin. They could still, however, have an intact protein backbone. We are presently in the process of purifying some of the membrane proteins such that specific antibodies can be prepared and used to examine with more rigor the turnover of the carbohydrate and amino acid constituents of individual membrane glyco- proteins. These studies will be reported in subsequent com- munications.
Achno~ule&ment-We thank Esther Hou for her extremely valu- able assistance during iodination experiments.
1
2.
3.
4. 5.
6.
7.
8. 9.
10.
11.
12.
HEFEHENCES
Gelehrter, 1‘. 11. (1976) N. Engl. J. Med. 294, 522-526, 589-595, 646-65 1
Thompson, E:. B., Tomkins, G. M., and Curran, cJ. F. (1966) PRK. Natl. Acad. Sci. U. S. A. 56, 296-303
Tweto, J., Friedman, E., and Doyle, 11. (1976) Su~v~~mol. Struct. 4, 141-159
Tweto, J., and Doyle, I). (1976) J. Biol. Chem. 251, 872-882 Tweto, J., and Doyle, 1). (1977) in Cell Surfmx Reviews (I’oste,
G., and Nicholson, G. L., eds) Vol. 4, Elsevier, Amsterdam Gavin, J. H., Roth, J., Neville, I). M., Jr., De Meyts, I’., and Buell,
D. N. (1974) Proc. N&Z. Acad. Sci. U. S. A. 71, 84-88 Van Obberghen, E., De Meyts, I’., and Roth, J. (1976) J. Bid.
Chem. 251,6844-6851 Brown, M. S., and Goldstein, J. L. (1975) Cell 6, 307-316 Goldstein, J. L., Brown, M. S., and Stone, N. J. (1977) Cell 12,
629-64 1 Hudgin, H. L., Pricer, W. E., Jr., Ashwell, G., Stockert, K. J., and
Morell. A. G. (19741 J. Bid. Chem. 249. 5536-5543 Pricer, W. E., Jr., and Ashwell, G. (1976) J. Bid. Chem. 251,
7539-7544 Doyle, Il., Baumann, H., England, B., Friedman, E., Hou, E., and
by guest on June 5, 2018http://w
ww
.jbc.org/D
ownloaded from

4418 Plasma Membrane GLycoproteins of Hepatoma Cells
13. 14.
15.
16. 17. 18.
19. 20.
21.
Tweto, J. (1978) J. Biol. Chem. 253,965-973 England, B., and Doyle, D. (1977) J. Cell Biol. 75, 221a Gahmberg, C. G., and Hakomori, S. (1973) J. Biol. Chem. 248,
4311-4317 Juliano, R. L., and Stanley, P. (1975) Biochim. Biophys. Acta.
389,401-406 Critchley, D. R. (1975) Cell 3, 121-125
Hubbard, A. L., and Cohn, Z. A., (1972) J. Cell Biol. 55, 390-405 Lowrv. 0. H.. Rosebrounh, N. J.. Farr. A. L.. and Randall. R. J.
Steck, T. L., and Dawson, G. (1974) J. Biol. Chem. 219,2135-2142 Phillips, D. It., and Morrison, M. (1971) Biochemistry 10,
1766-1771
(195i) J. Biol. Chem. 193, 265-275 Chambers, R. E., and Clamp, J. R. (1971) Biochem. J. 125,
1009-1018 22. Laine, R. A., Essetman, W. J., and Sweeley, C. C. (1972) Methods
Enzymol. 28, 159-167 23. Laemmli, U. K. (1970) Nature 227, 680-685 24. Bonner, W. M., and Laskey, R. A. (1974) Eur. J. Biochem. 46,
83-88 25. Doyle, D., England, B., Friedman, E., Hou, E., and Tweto, J.
(1977) in Membrane Alterations as the Basis of Liver Injury (Popper, H., Bianchi, L., and Reutter, W., eds) MTP Press Ltd., Lancaster, England
26. O’Farrell, P. H. (1975) J. Biol. Chem. 250, 4007-4021 27. Ames, Ferro-Luzzi, G., and Nikaido, K., (1976) Biochemistry 15,
616-623 28. Hubbard, A. L., and Cohn, Z. A. (1975) J. Cell Biol. 64, 461-479
by guest on June 5, 2018http://w
ww
.jbc.org/D
ownloaded from

H Baumann and D Doyletissue culture cells.
Turnover of plasma membrane glycoproteins and glycolipids of hepatoma
1978, 253:4408-4418.J. Biol. Chem.
http://www.jbc.org/content/253/12/4408.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/253/12/4408.citation.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on June 5, 2018http://w
ww
.jbc.org/D
ownloaded from