virological and immunological studies of dengue … filevirological and immunological studies of...
TRANSCRIPT

VIROLOGICAL AND IMMUNOLOGICAL STUDIES
OF DENGUE VIRUS INFECTION
IN PIGTAIL MACAQUES (MACACA NEMESTRINA)
SUSANA WIDJAJA
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
2010

STATEMENT
Hereby I, Susana Widjaja, do declare that this dissertation entitled
“Virological and Immunological Studies of Dengue Virus Infection in Pigtail
Macaques (Macaca nemestrina)” is my own work and has not been submitted in
any form for another degree or diploma programs (course) to any university or
other institution. The content of the dissertation has been examined by the
advising committee and the external examiner.
Bogor, August 2010
Susana Widjaja
P067050051

ABSTRACT
SUSANA WIDJAJA. Virological and Immunological Studies of Dengue Virus
Infection in Pigtail Macaques (Macaca nemestrina). Supervised by DONDIN
SAJUTHI, JOKO PAMUNGKAS, DIAH ISKANDRIATI, and PATRICK J
BLAIR.
A non-human primate (NHP) model is essential for the study of dengue
hemorrhagic fever (DHF) pathogenesis and the evaluation of dengue (DEN)
vaccine and antiviral drug. Until now, it has been difficult to find an NHP DHF
pathogenesis model. Therefore, an evaluation of a DEN vaccine candidate is
performed in NHPs that show viremia after infected by DEN virus and the
vaccine efficacy is its capability to develop immunity that reduces viremia when
vaccinated NHPs are challenged by DEN virus. In this study, the potential of
pigtail macaque to serve as an animal model for DEN vaccine testing was
evaluated. Homologous sequential DEN challenges were conducted using primary
viral isolates from DEN patients in Indonesia. Two parameters, the ability to
support dengue viremia and to produce sufficient antibody responses were
measured. This study shows that primary infections of all four DEN serotypes
cause consistent, measurable viremia in pigtail macaques. The responses of IgM,
IgG and avidity antibody following primary and secondary DEN infections are
similar with antibody responses in human. The immunity produced by primary
infection is sufficient to protect against homologous virus. This species of
macaque therefore appears to be a suitable alternative model for testing DEN
vaccine candidates. Besides antibody, T lymphocyte also has an important role in
the protection and pathogenesis of DEN diseases. DEN specific T lymphocyte
measurements, ELISPOT and intracellular cytokine staining-flow cytometry (IC-
FC), were developed to support DEN studies in pigtail macaque. Peripheral blood
mononuclear cells (PBMC) collected before and after DEN infections were tested.
ELISPOT results show increase of DEN specific interferon-γ (IFN-γ) producing
cells as an individual response of pigtail to primary DEN-1, DEN-3 or DEN-4
infections. Using pools of PBMC taken from several animals, ELISPOT and
intracellular cytokine staining-flow cytometry (IC-FC) was run side by side to
quantify DEN specific lymphocytes following primary and secondary DEN-2
infections. ELISPOT revealed an increase of DEN specific IFN-γ producing cells
following primary infection and a significant increase after secondary infection.
Similarly, IC-FC also measured an increase of DEN specific producing IFN-γ
CD3+CD4+ and CD3+CD4- T lymphocytes. As such, ELISPOT and IC-FC can
be applied to measure DEN specific T lymphocytes in pigtail macaques.
Therefore, the application of these assays would be useful in elaborating adaptive
immunity induced by vaccine and the level of protection. Furthermore, the
development of pigtail as DHF model can be evaluated when further research on
the cross-reactive T lymphocyte and antibody responses during secondary
heterologous is conducted.
Keywords: M. nemestrina, dengue infections, viremia, antibody, T lymphocytes.

ABSTRAK
SUSANA WIDJAJA. Studi Virologi dan Imunologi Infeksi Virus Dengue Pada
Satwa Primata Beruk (Macaca nemestrina). Dibimbing oleh DONDIN SAJUTHI,
JOKO PAMUNGKAS, DIAH ISKANDRIATI, dan PATRICK J BLAIR.
Satwa primata sangat dibutuhkan untuk meneliti patogenesis demam
berdarah dengue (DBD) dan mengevaluasi vaksin dengue (DEN), juga obat
antivirus. Sampai saat ini sangat sulit mendapatkan model DBD pada satwa
primata. Jadi evaluasi kandidat vaksin DEN dilakukan pada satwa primata yang
memperlihatkan viremia setelah infeksi virus DEN dan vaksin yang efisien adalah
vaksin mampu menimbulkan kekebalan yang dapat mereduksi viremia pada
primata yang setelah divaksinasi kemudian diinfeksikan virus DEN. Untuk dapat
mengetahui potensi satwa primata beruk sebagai hewan model pada penelitian
vaksin DEN, beruk diinfeksikan berturutan dengan serotipe DEN yang sama.
Virus DEN yang digunakan berasal dari virus yang diisolasi dari pasien-pasien
DEN di Indonesia. Dua parameter yang diukur adalah viremia yang terjadi setelah
penyuntikan virus DEN dan antibodi sebagai respon beruk terhadap infeksi DEN
tersebut. Beruk memperlihatkan viremia yang konsisten setelah diinfeksikan
dengan virus DEN-1, DEN-2, DEN-3 dan DEN-4. Respon antibodi IgM, IgG dan
aviditas setelah infeksi primer dan sekunder menyerupai respon pada manusia.
Kekebalan yang terjadi setelah infeksi primer dapat melindungi beruk dari infeksi
sekunder homologus. Hasil ini menunjukkan bahwa beruk dapat digunakan untuk
evaluasi vaksin DEN dan menjadi hewan model alternatif untuk penelitian infeksi
DEN. Tidak hanya antibodi, limfosit T juga memiliki peran penting terhadap
proteksi dan patogenesa infeksi DEN. Pengukuran limfosit T spesifik DEN yang
memproduksi interferon-γ (IFN- γ) yaitu ELISPOT dan intracellular cytokine
staining-flow cytometry (IC-FC) dikembangkan untuk mendukung penelitian
DEN pada beruk. Pengujian dilakukan menggunakan peripheral blood
mononuclear cells (PBMC) yang diambil sebelum dan sesudah infeksi DEN.
Hasil ELISPOT memperlihatkan kenaikan jumlah limfosit T spesifik DEN
sebagai respon individu beruk terhadap infeksi DEN-1, DEN-3 dan DEN-4.
Dengan menyatukan PBMC dari beberapa beruk, ELISPOT dan IC-C dilakukan
secara bersamaan untuk mengukur jumlah limfosit T spesifik DEN setelah infeksi
DEN-2. ELISPOT memperlihatkan kenaikan limfosit T spesifik DEN yang
memproduksi IFN-γ setelah infeksi primer dan kenaikan yang lebih nyata sebagai
respon terhadap infeksi sekunder. Hasil IC-FC, pola kenaikan dari limfosit T
CD3+CD4+ dan CD3+CD4- spesifik DEN yang memproduksi IFN-γ sebagai
respon terhadap infeksi primer dan sekunder serupa dengan respon yang diukur
dengan ELISPOT. Jadi ELISPOT dan IC-FC dapat digunakan untuk mengukur
limfosit T spesifik DEN. Aplikasi kedua uji ini dapat digunakan untuk
mempelajari lebih rinci kekebalan adaptif yang didapat dari vaksinasi dan
kemampuan proteksinya. Demikian pula, pengembangan beruk sebagai model
DBD akan dapat dilakukan melalui penelitian respon reaksi silang dari limfosit T
dan antibodi pada infeksi sekunder heterologus.
Kata kunci: M. nemestrina, infeksi dengue, viremia, antibodi, limfosit T.

SUMMARY
SUSANA WIDJAJA. Virological and Immunological Studies of Dengue Virus
Infection in Pigtail Macaques (Macaca nemestrina). Supervised by DONDIN
SAJUTHI, JOKO PAMUNGKAS, DIAH ISKANDRIATI, and PATRICK J
BLAIR.
Dengue virus infections have caused a major public health problem in
tropical and sub-tropical countries. The geographical distribution, the frequency
of epidemic cycle and the number of cases have been increasing at an alarming
rate and highlighted the urgency of DEN vaccine (WHO 2005; Raviprakash et al.
2009). The clinical manifestations of DEN infections range from mild dengue
fever (DF) with high fever, headache, rash, and bone and muscle pain up to severe
dengue hemorrhagic fever (DHF) and dengue shock syndrome (DSS) with
evidence of thrombocytopenia, bleeding, plasma leakage and shock. The severe
manifestation cause a high mortality rate particularly in children (WHO 2005).
Since the 1980s, epidemiological data revealed that 85% of DHF and DSS were
heterologous secondary infections and immunopathological response to
heterologous secondary infection has been hyphothesized leading to DHF
pathogenesis (Halstead 1983). Antibody produced following primay DEN
infection confers the protection to homologous infection, however, heterologous
secondary infection may still occur. The pre-existing, non-neutralizing antibodies
binds DEN viruses and these complexes, then bind the target cells via the FcγRI
and FcγII, resulting in increased viral load, shortened incubation period and
increased disease severity (Fink et al. 2006). Meanwhile, DEN specific CD4+ and
CD8+ T lymphocytes are suggested to have a low binding affinity for the current
serotype, and consequently, inefficient to clear the infection (Fink et al. 2006).
The limitations of DEN study in human have hampered the understanding of these
two components of adaptive immunity, antibody and T lymphocyte, in the
pathogenesis of DHF.
Dengue vaccine evaluation in non-human primate (NHP) model is
required before the vaccine can be applied in human. Here, we explored the
posibility of pigtail macaque (Macaca nemestrina) as an animal model to evaluate
DEN vaccine. A total of seventeen Flavivirus-free pigtail macaques were
separated into four groups by DEN serotypes. Dengue-1 to dengue-4 viruses were
isolated from DEN patients in Indonesia. Aproximately 105 plaque forming unit
(PFU) DEN virus was injected subcutaneously into each individual in the lateral
chest area. Blood samples were obtained prior to virus injection as baseline
sample and daily for 10 days post-infection for virus detection and on 14, 28 and
87 days post-injection for anti-dengue antibody profile analysis. Consistent
viremia was detected by virus isolation (mosquito inoculation and C6/36 cell
culture) and RT PCR methods. Viremia was detected one or two days post-
infection in most of the animals. By RT-PCR and mosquito inoculation methods,
the least number of viremia days occurred with DEN-4 (5±1.4 and 3.3±1 days).
By isolation in C6/36, DEN-3 produced the least (4±0.8). DEN-2 resulted in the
longest average number of days viremia (7.8±0.5, 6.8±1 and 5.8±1 days as
measured by RT PCR, isolation in C6/36 and mosquito inoculation, respectively).
A challenge with homologous serotype six month after the first infection did not

result in any detectable viremia by virus isolation and only one to two days viral
RNA was detected in DEN-4 group. Prior in primary infection, IgM antibody was
detected, then followed by IgG antibody. During secondary infection, IgM was
not detected, whereas IgG increased rapidly. The avidity of IgG increased
overtime following primary infection and secondary infection. Similarly with IgG
and its avidity, high neutralizing antibody was generated following primary
infection and augmented in secondary infection. These antibody responses to
primary and secondary DEN infections were similar with antibody responses in
human. The predominat IgG subclass following primary and seconday infections
was IgG1. These data reveal that pigtail macaque is suitable for the study of DEN
infection. This animal can serve as an alternative model for evaluating DEN
vaccine, since the efficacy of a DEN vaccine is measured by its capability to
reduce viremia after vaccinated animals are challenged with live DEN virus.
To support DEN study in pigtail macaque, ELISPOT and intracellular
cytokine staining-flow cytometry (IC-FC) were established to enumerate DEN
specific T lymphocytes. The ELISPOT assay employs ELISA technique to trap
antigen- induced cytokine secretion around the cells by an immobilized anti-
cytokine antibody on polyvinylidene difluoride membrane, and then visualizes the
complexes by anti-cytokine conjugate and substrate. IC-FC uses brefeldin A to
trap cytokine intracellularly following antigen stimulation. Then, the cells are
permeabilized and specific anti-cytokine fluorescent antibodies can pass into the
cells and react with cytokines. Both assays measure functional T cells after
stimulation by DEN antigen, however, ELISPOT measures secreted cytokine
while IC-FC measures intracellular cytokine (Lecth and Scheibenbogen 2003).
Dengue antigens were generated from intra- and extra-cellular proteins of DEN
virus culture in Vero cells. The application of DEN antigen for in vitro stimulation
of T lymphocytes reduce the complexity of DEN specific T lymphocyte assays,
since the generation of antigen presenting cells or prior knowledge of antigenic
peptides is not required (Mangada et al. 2004). Homologous T cell responses were
observed. Peripheral blood mononuclear cells (PBMC) pre- and post DEN
infections had been isolated from heparinized blood collected during several
previous DEN studies and stored in LN2 until assayed. ELISPOT detected 0-40
DEN specific interferon-γ (IFN-γ) producing cells from PBMC before DEN
infection and 28-440 cells after DEN infections. Increase of DEN specific IFN-γ
producing cells was detected as an individual response of pigtail to DEN-1, DEN-
3 or DEN-4 infections. ELISPOT and IC-FC was run simultaneously to quantify
DEN specific lymphocytes following primary and secondary DEN-2 infections
using pools of PBMC taken from several animals. An increase of DEN-2 specific
IFN-γ cells following primary infection and a significant increase after secondary
infection were detected by ELISPOT. Similarly, increase CD3+CD4+ (T helper-
1) and CD3+CD4- (T cytotoxic) specific DEN after primary and secondary
infections were detected. These results show that both ELISPOT and IC-FC can
be used to measure DEN specific T lymphocytes.
Based on the susceptibility of pigtail macaque to the infections of all four
DEN serotypes, pigtail macaque is suitable model to study DEN infection and can
be used as an alternate NHP model to evaluate DEN vaccine or anti-viral.
Furthermore, the availability of DEN specific T lymphocyte measurements allow

more detail exploration on immunity induced by vaccination that protects the
pigtail from DEN challenge. As DHF is associated with heterologous secondary
infections, to evaluate whether pigtail macaque is suitable as the model, further
study on serotype cross-reactive antibody and T lymphocytes responses should be
investigated.

Copyright © 2010 Bogor Agricultural University
Copyright are protected by law,
1. It is prohibited to cite all or part of this thesis/ dissertation without
referring to and mentioning the source.
a. Citation only permitted for the sake of education, research,
scientific writing, report writing, critical writing or reviewing
scientific problem.
b. Citation doesn’t inflict the name and honor of Bogor Agricultural
University.
2. It is prohibited to republish and reproduce all or part of this thesis/
dissertation without the written permission from Bogor Agricultural
University.

VIROLOGICAL AND IMMUNOLOGICAL STUDIES OF
DENGUE VIRUS INFECTION IN PIGTAIL MACAQUES
(MACACA NEMESTRINA)
SUSANA WIDJAJA
Dissertation
submitted in partial fulfillment of the requirements
for the degree of Doctorate in the Primatology Major,
Graduate Program, Institut Pertanian Bogor.
GRADUATE PROGRAM
INSTITUT PERTANIAN BOGOR
BOGOR
2010

External examiners in private defense:
1 Dr. Irma Suparto, M.D., M.S.
2 Drh. Surachmi Setiyaningsih, Ph.D.
External examiners in public defense:
1 Bachti Alisjahbana, M.D., Ph.D
2 Tjahjani Mirawati Sudiro, M.D., Sp.M.K., Ph.D.

Title : Virological and Immunological Studies of Dengue Virus
Infection in Pigtail Macaques (Macaca nemestrina)
Name : Susana Widjaja
Student Number : P067050051
Approved by
Advisory Commitee
Prof. Drh. Dondin Sajuthi, M.St., Ph.D. Dr. Drh. Joko Pamungkas, M.Sc.
Major Advisor Co-Advisor
Dr. Drh. Diah Iskandriati Patrick J Blair, Ph.D.
Co-Advisor Co-Advisor
Acknowledged by
Chairman, Major Primatology Dean of Graduate School
Prof. drh. Dondin Sajuthi, M.St., Ph.D. Prof. Dr. Ir. Khairil A. Notodiputro, M.S.
Date of final examination: 8 October 2010 Date of graduation:

To Indonesian scientists in health research.
Let science be our first priority to achieve welfare for all Indonesians.

PREFACE
“Virological and Immunological Studies of Dengue Infection in Pigtail
Macaques (Macaca nemestrina)” consists of two research publications entitled
“Pigtail Macaque (Macaca nemestrina) and Dengue Virus Infectivity: a Potential
Model for Evaluating Dengue Vaccine Candidates” and “The Measurements of
Dengue Specific Interferon-γ Producing T Lymphocytes in Pigtail Macaques
(Macaca nemestrina)”. These two studies are intended to explore pigtail macaque
as a non-human primates (NHP) model for dengue research, therefore, more
diverse NHP species can be utilized. The urgency of available licensed dengue
vaccine draws attention to NHP requirements in the pre-clinical phase of vaccine
trial. And the lack of dengue hemorrhagic fever NHP model may be solved by
certain susceptible NHP species. Another purpose is to bring more opportunities
of pigtail macaque to be used in biomedical research. As pigtail macaque is
endemic NHP in Kalimantan and Sumatra islands, the use of Indonesian “natural
resource” in biomedical research without threatening the existence of the species
in its natural habitats in Indonesia, hopefully, can open a better chance for the
welfare of the people and NHP in Indonesia.

ACKNOWLEDGMENTS
I praise and thank God for His good hand is upon me in each step so I am
able to complete this dissertation. And this dissertation holds far more than the
culmination of research. It is also a result of great correlation with many brilliant,
generous, inspiring and lovely people.
My deepest gratitude goes to Prof. Kevin Porter, M.D., who had the
original idea and initial study of the pigtail macaque as an animal model for
dengue infection. Also, this dissertation would not be completed without
subsequent research and kind-hearted continual support from all the former Viral
Diseases program Directors: Charmagne G Beckett, M.D., Patrick J Blair, Ph.D.,
Timothy H Burgess, M.D., M.P.H., and Maya Williams, Ph.D.
My heartfelt gratitude also goes out to my supervisors, Prof. Drh. Dondin
Sajuthi, M.St., Dr. Drh. Joko Pamungkas, M.Sc., Dr. Drh. Diah Iskandriati, and
Patrick J Blair, Ph.D whose untiring effort, commitment, encouragement,
guidance and support helped me greatly in exploring the studies and writing the
dissertation.
My special thank to Gary T Brice, Ph.D., for tutoring the cellular
measurements, and for the long discussions that helped me sort out the technical
details of the work.
I am grateful to Prof. Dr. Ir. Sri Supraptini Mansjoer, Drh. Ikin Mansjoer,
M.Sc., Dr. Irma H. Suprapto, M.D., Dr. Erni Sulistiyawati, D.V.M, Dr. Ir. Dyah
Perwitasari and other lecturers in Primatology Major for teaching good research,
also giving continuous guidance and encouragement.
I acknowledge valuable direction and advice to finalize this dissertation
from the external examiners: Dr. Irma H. Suparto, M.D., M.S., Drh. Surachmi
Setiyaningsih, Ph.D., Bachti Alisjahbana, M.D., Ph.D., Tjahjani Mirawati Sudiro,
M.D., Sp.M.K., Ph.D.
My thank to the Primatology staff for assisting me with the administrative
tasks necessary for completing my doctoral program: Yanti and Yana.
I am in debt to my invaluable, supportive, forgiving, generous and loving
colleagues: Ratna Tan, Chairin Maroef, Imelda Winoto, Sri Hadiwidjaya, Ungke
Antonjaya, Sherly, Dasep, Deni, Haditya, Anton, Gustiani, Yuanita, Nurhidayah,

Nurhayati, Ester, Melinda, Santo, Mara, Anti, Ovi, Saraswati and other US
NAMRU-2 staff. Their incredible hard work and dedication to the US Navy and
scientific society inspire me for always doing high quality work. I am most
indebted to Herman Kosasih M.D. and Victor Sugiharto for abiding friendship,
careful review and discussion that graciously provided throughout all stages of
this dissertation fruition.
I am grateful to Sylvia, Tuah, Harri, and Suyanti for the friendship and
encouragement during and after the master degree program.
I greatly value the care and confidence from my best friends, Linda
Martini and Bimo Wicaksana, whose friendship have helped me keep moving on
and stay focus through the years.
Most importantly, none of this would have been possible without my
family; my sister and brothers: Susanti, Susanto and Sugiharto, my husband:
Herjadi, my children: Calista and Aldwin whose patience and love sustain me
through all my endeavours to complete this dissertation.
Jakarta, August 2010
Susana Widjaja

CURRICULUM VITAE
The author was born on the 3rd
of May in 1964 in Jakarta. She is the
second daughter of the four children from the late Bakri Widjaja and Betty
Gomulya. She was married with Laurentius Herjadi and has blessed with talented
daughter, Saphire Calista, and thoughtful son, Lotharius Aldwin.
She received Doctor of Veterinary Medicine from the Faculty of
Veterinary Medicine , Institut Pertanian Bogor in 1987. She entered the Graduate
Program at the Institut Pertanian Bogor for a master degree in Primatology Major
in 2003, then approved to continue directly to doctorate degree in 2005.
The author started to serve at the United States Naval Medical Research
Unit-2, Jakarta in December 1988. In this prestigious infectious diseases research
laboratory, she got the amazing opportunity to develop her skill and knowledge
from technical ability as a bench laboratory staff up to managerial flair as the
head of the Tissue Culture and Immunology Division of the Viral Diseases
Program. She was a member of Institutional Animal Care and Use Committee
since 2003. She received a visiting scientist scholarship in 2002-2003 and trained
for the measurements of dengue humoral and cellular immunity in Naval Medical
Research Center, Maryland. After completion of this training, she received
outstanding visiting scientist award. Over 20 years of faithful and exceptional
service, she was granted numerous awards and letters of recognition. She also
produced many scientific publications as the author or co-author together with
briliant US and Indonesian scientists. She proudly continued to serve at the US
NAMRU-2 until its unfortunate and sudden closure in 2010.

TABLE OF CONTENTS
page
LIST OF TABLES
LIST OF FIGURES
INTRODUCTION ........................................................................................
LITERATURE REVIEW .............................................................................
Dengue virus ................................................................................................
Dengue infections ........................................................................................
The roles of B and T memory lymphocytes in the pathogenesis of dengue
hemorrhagic fever .........................................................................................
Dengue vaccine and antiviral drug ...............................................................
Animal model for dengue infections ...........................................................
Dengue specific cytokine producing T lymphocyte measurements ............
GENERAL METHODOLOGY ...................................................................
PIGTAIL MACAQUE (MACACA NEMESTRINA) AND DENGUE
VIRUS INFECTIFITY: A POTENTIAL MODEL FOR EVALUATING
DENGUE VACCINE CANDIDATES ........................................................
THE MEASUREMENT OF DENGUE SPECIFIC INTERFERON-γ
PRODUCING T LYMPHOCYTES IN PIGTAIL MACAQUES
(MACACA NEMESTRINA) ..........................................................................
GENERAL DISCUSSION ...........................................................................
CONCLUDING REMARKS .......................................................................
REFERENCES .............................................................................................
APPENDIX ..................................................................................................
xiv
xv
1
4
4
6
7
7
9
10
12
16
33
48
51
52
55

LIST OF TABLES
page
1 Grading severity of dengue infection ..............................................
2 Dengue vaccine candidates in clinical and pre-clinical trials .........
3 Human T cell subsets .......................................................................
4 Homologous anti-DEN neutralizing antibody responses
after primary and secondary infections ............................................
5 DEN specific IFN-γ producing cells in pigtail macaques before
and after DEN infection .................................................................
6
8
11
26
41

LIST OF FIGURES
page
1 A schematic presentation of dengue polyprotein ...............................
2 The life cycle of dengue virus in the cell ...........................................
3 Outline of virus injection and blood collection for pigtail
susceptibility study .............................................................................
4 Outline of of virus injection and blood collection for the study of
cellular immunity specific to DEN measurements ............................
5 The length of viremia in pigtail challenged with DEN viruses ..........
6 IgM , IgG and avidity responses after primary and secondary
infections .............................................................................................
7 Anti-DEN IgG subclasses after primary infection with DEN-4 .........
8 Dengue-1, Dengue-3 and Degue-4 antigen optimization by
ELISPOT ...........................................................................................
9 Dengue-2 antigen optimization by ELISPOT and IC-FC ...................
10 ELISPOT and IC-FC results of DEN-2 primary and secondary
infections ............................................................................................
5
5
13
14
23
24
25
40
42
43

LIST OF APPENDICES
page
1 List of reagents for laboratory assays ...............................................
2 PCR cycle condition .........................................................................
3 List of reagents for ELISPOT and intracellular staining-flow
cytometry ...........................................................................................
55
58
59

INTRODUCTION
Dengue (DEN) virus infections have threatened more than one third of the
world population (WHO 2005). It has been estimated that there are 50-100 million
dengue fever (DF) cases annualy of which 2-4% result in severe forms of the
disease, dengue hemorrhagic fever (DHF) or dengue shock syndrome (DSS), a life
threatening disease particularly in children (WHO 2005). In Indonesia, more than
150 000 DHF and DSS cases with 1-2% mortality rate were reported in 2007 (Dit
Jen P2MPL 2008). Thus, dengue vaccine has become a priority of world health
research for effective prevention (Raviprakash et al. 2009).
Dengue virus consists of four distinct serotypes (DEN-1 to DEN-4) with up
to 30% dissimilarity among serotypes (Irie et al. 1989). While primary infection
confers protective immunity to the same serotype, heterologous secondary infection
has been hypothesized to be responsible for the immunopathogenesis of DHF or
DSS. Original antigenic sin theory has enlightened the role of B and T lymphocytes
(B and T cells) during heterologous secondary infections (Halstead et al. 1983).
Although the similarity between serotypes of primary and secondary infection result
in rapid expansion of pre-existing memory B and T cells, it generates low-avidity
antibodies and T cells to the infecting serotype. The antibodies bind, but do not
neutralize the virus. Instead, they augment virus entry to target cells through Fc
receptor (antibody dependent enhancement of infection, ADEI hypothesis)
(Halstead 2003). As consequent, increase of viral replication and increase of
infected cells result in more antigen presenting cells to stimulate T cells. Low
avidity T cells have less ability for viral clearance and produce predominantly pro-
inflamatory cytokines. Thus, altered T lymphocyte functions lead to DHF or DSS
(Rothman 2004).
An animal model of DEN infections will be invaluable to study the
pathogenesis of DHF or DSS, since study in humans has had many limitations
(Beckett et al. 2005; Raviprakash et al. 2009). Non-human primates (NHP)
commonly used in DEN research are rhesus (Macaca mullata) and cynomolgus (M.
fascicularis) macaques as they develop detectable viremia and antibodies following
DEN infections (Bente and Rico-Hesse 2006; Raviprakash et al. 2009). They are
used to test the efficacy of a DEN vaccine and antiviral drug which is evaluated by

2
their abilities to prevent, or to significantly reduce, viremia when animals are
challenged with live DEN virus. Until now, NHP as DHF animal model is still
difficult to find. As pigtail macaque (M. nemestrina) has been shown exceptional
suceptibility to human immunodeficiency virus (HIV) and simian
immunodeficiency virus (SIV) (Baroncelli et al. 2008), it may also be studied to see
whether it is better, compared to other non human primates, as the animal model
for DEN infections. This animal has never been reported as a model for DEN
infection (Raviprakash et al. 2009).
Compared with B cells and antibodies, T cells and their functions have been
limited to study. Conventional measurements of antigen specific T cells, such as H-
thymidine proliferation assay, Cr-release cytotoxic assay and secretion of cytokines
in bulk lymphocyte cultures are laborious and time consuming. Also, they produce
insensitive and inconsistent results (Hickling 1998, Gauduin et al. 2004). The
enzyme-linked immunospot (ELISPOT) and intracellular cytokine staining-flow
cytometry (IC-FC) assays measure T functional cells and employ the antigen
specific secretion of cytokines to detect specific T cells on a single cell level (Lecth
and Scheibenbogen 2003). These assays have become preferential, since they are
more straightforward and faster than conventional assays (Pahar et al. 2003). To
quantify DEN-specific T cells in cynomolgus macaques, Koraka et al. (2007 a,b)
employed ELISPOT and applied APC derived from autologous B cells to stimulate
interferon-γ (IFN-γ) producing T cells. An alternative technique for in vitro
stimulation of DEN specific T cells was an application DEN lysate antigen in bulk
human peripheral blood mononuclear cells (PBMC). Mangada et al. (2004) applied
these antigens and IC-FC assay to detect DEN specific IFN-γ producing T cells in
human.
In order to explore the possibility of pigtail macaque as a model for DEN
infection, DEN study in pigtail macaques was conducted. Virological and
immunological examinations were done thoroughly including virus isolation and
RT PCR for virerima detection, IgM, IgG, avidity IgG, subclass IgG and
neutralizing antibodies for the evaluation of humoral responses. Another study was
conducted for the development of ELISPOT and IC-FC assays, in order to enhance
DEN study in pigtail macaque. These assays used DEN antigens to stimulate T
cells. Our study shows that pigtail macaques support DEN replication resulting in

3
viremia and antibody responses that are similar with viremia and antibody responses
in human. Therefore, pigtail macaques are appropriate as animal model for vaccine
and antiviral evaluations. The ELISPOT and IC-FC revealed increase of DEN
specific IFN-γ Tcells after DEN infections.

LITERATURE REVIEW
Dengue virus. According to International Committee on Taxonomy of
Viruses (ICTV), a subgroup of Virology Division of the International Union of
Microbiology Societies, dengue virus belongs to the Flavivirus genus of the
Flaviviridae family (Calisher and Gould 2003). The virus particle is spherical, 40-
60 nm in diameter. Its icosahedral core consists of a capsid protein (C)
encapsulating a positive-sense, single-stranded RNA genome about 11 kilobases
(kb) in length. This RNA contains a 5’ cap (m7G5’ppp5’A) and functions as a
messenger RNA. The core is surronded by a lipid bilayer envelope with two viral
proteins, membrane (M) and envelope (E) protein (Lindenbach and Rice 2001,
2003).
The Flavivirus structure and replication is reviewed in detail by Lindenbach
and Rice (2001, 2003). The genome directs the synthesis of polyproteins.
Translation of one single open reading frame produces a large polyprotein that is
cleaved co- and post translationonally into three structural proteins: capsid (C),
precursor M (prM) and envelope (E) proteins, and seven non-structural (NS)
proteins: NS1, NS2A, NS2B, NS3, NS4A, NS4B and NS5. Figure 1 shows
schematic of DEN polyprotein. The C protein presumably mediates RNA
interaction, membrane association and also serves as signal peptide for ER
translocation of prM. The prM protein has chaperone-like activity protecting E
protein from undergoing rearrangement in the reduced pH environment of the early
secretory pathway. Then, the conversion of immature virus particles to mature
virions occurs in the secretory pathway with cleavage of prM into pr and M by the
Golgi resident furin or furin-like enzyme. The E glycoprotein is the major virion
surface proteins, which mediates binding and membrane fusion. The E protein is a
major target of humoral immunity. While stuctural proteins construct the viral
particle, non-strctural proteins support viral replication. An interaction of NS1 and
NS4A is required at a very early stage in RNA replication. Non-structural2A is
involved in coordinating the shift between RNA packaging and replication. Non-
structural2B is a co-factor for the serine protease of NS3. The carboxy terminal of
NS3 carries three enzymatic activities: a helicase to unwind double-stranded nucleic
acid during RNA replication, a NTPase to hydrolyse ATP to generate energy

5
neccesary during replication, and a RTPase to remove the terminal phosphate group
from the newly synthesized RNA for the formation of the viral cap structure at the
5’ end of genome. The NS5 methyl-tranferase (MTase) adds the cap (two methyl
groups) to the nucleotide. The RNA-dependent RNA polymerase (RdRp) produces
“copy-back” RNA.
Figure 1 A schematic presentation of dengue polyprotein. Dots represent enzyme
activity domains. Prot: protease, Hel: Helicasee/NTPase/RTPase, Mtase:
methyl-transferase, RdRp: RNA-dependent RNA polymerase.
(Lindenbach and Rice 2003).
Dengue virus enters into a host through the skin during mosquito feeding.
The replication of DEN virus begins when the virions infect a permissive host cells.
The primary target cells are mononuclear phagocytes and the entry is facilitated by
receptor mediated endocytosis. The best-characterized receptor that can mediated all
four serotypes of DEN virus is DC-SIGN. The virus is internalised into the
endosomal compartment where the acidic pH triggers a fusion of its envelope to the
endosomal membrane and deliver the viral genome into the cytoplasm. The viral
polyprotein is synthesized in association with the endoplasmic reticulum and is
processed into structural and non-structural protein by viral and cellular proteases.
Figure 2 The life cycle of dengue virus in the cell (Fink et at. 2006).

6
A viral replication complex is formed on the membrane of the endoplasmic
reticulum which facilitates replication of DEN genome. Newly synthesized viral
genomes are packed by core, envelope and membrane proteins along the secretory
pathway. Immature virus particles are transported by the secretory pathway to the
cell wall where furin cleaves the prM protein into M protein and the mature virion is
released. In a secondary infection, DEN virus binds to antibody from a previous
infection (antibody dependent enhancement of infection, ADEI) and is then
endocytosed by Fc receptor bearing cells, such as monocytes. The life cycle of
DEN virus was reviewed by Clyde et al. (2006) and Fink et al. (2006) (Fig 2).
Dengue infections. Incubation period usually varies from 3 to 14 days with
average 4 to 7 days (Gubler 1998). All four DEN infections in human may be
asymptomatic or may lead to undifferentiated fever, dengue fever (DF), dengue
hemorrhagic fever (DHF) or dengue shock syndrome (DSS) (WHO 2005).
Table 1 Grading the severity of dengue infection
Grade Symptoms Laboratory
DF Fever with two or more Leukopenia occasionally.
of the following sings: Trombocytopenia may be present.
headache, retro-orbital pain, No evidence of plasma loss
myalgia, arthralgia
DHF I Above signs plus Thrombocytopenia ≤ 100 000
positive tourniquet test. Hematocrit rise ≥ 20%
DHF II Above signs plus Thrombocytopenia ≤ 100 000
spontaneous bleeding Hematocrit rise ≥ 20%
DHF III* Above signs plus Thrombocytopenia ≤ 100 000
circulatory failure Hematocrit rise ≥ 20%
(weak pulse, hypotension,
restlessness)
DHF IV* Profound shock with Thrombocytopenia ≤ 100 000
undetectable blood Hematocrit rise ≥ 20%
pressure and pulse
* DHF grade III and IV are also called as dengue shock syndrome (DSS)
(WHO 2005)

7
The grades of DEN diseases are described in Table 1. Dengue fever is
characterized by the sudden onset of high fever (38-40oC) and a variety of non-
specific symptoms, including headache, retro-orbital pain, myalgia and arthralgia.
Dengue infection has an unpredictable course where most patients have a febrile
phase lasting 2 to 7 days and this is followed by a critical phase which is of
about 2 to 3 days duration. Usually during this defevercence phase, patient are at
risk of developing DHF/DSS. Symptoms and laboratory findings in DHF grade I
and II include trombocytopenia (less than 100 000) and a rise in hematocrit level
more than 20%. Spontaneous bleeding such as rash, bleeding from nose and gum or
melena distinguish DHF grade I and grade II. Weak pulse, hypotension or
undetectable blood pressure pulse indicate DHF grade III or IV.
The role of B and T memory lymphocytes in the pathogenesis of dengue
hemorrhagic fever. At the early phase of heterologous secondary infection, the
complexes of DEN virus and non-neutralizing antibody allow viral uptake via the
Fc portion of the antibody to FcγRI and FcγRII bearing cells (Littaua et al. 1990).
Consequently, a greater number of cells are infected resulting in increased viral
load, shortened incubation period and increased disease severity (Fink et al. 2006).
The preferential expansion of memory T cells with lower avidity for the infecting
serotype causes altered T cell functional responses (Mathew and Rothman 2008).
Cross-reactive CD8 (clusters of differentiation8) T cells with low binding affinity
for the current infection have less cytolitic activity. This may exacerbate the
infection and lead to significant immune-mediated tissue damage as more T cells
die and release cytokines (Mathew and Rothman 2008). In addition, low affinity
cross-reactive CD4 T cells also produce predominantly proinflamatory cytokines
and lyse bystander uninfected cells (Mathew and Rothman 2008, Rothman 2004).
Dengue vaccine and antiviral drug. Dengue vaccine has been expected as
an effective control for DEN infections. In spite of great efforts over the last seven
decades, a licenced vaccine has not been produced. All these efforts are
summarized in Table 2. Live attenuated vaccines (LAV) have led in the
development and clinical trials (Reviewed in Chaturvedi et al. 2005, Raviprakash et
al. 2009). However, these vaccines have been associated with clinical
complications due to their reactogenicity. As an attenuation to produce
adequate immunogenicity with minimal reactogenicity is the biggest handicap. To

8
overcome, recombinant LAV has been developed by mutation or deletion in the
viral genome (Reviewed in Raviprakash et al. 2009). However, formulating
monovalent combinations to attain tetravalent long-lasting protective immunity has
been a big problem because of serotype dominance and competition. Also, concern
regarding an application of replicating vaccine has been a long debate due to the
possibility of mutation or recombination that can initiate virulence. Therefore,
recombinant adenovirus vector and DNA shuffling technology offer an advantage of
expressing multiple antigens from a single vector and make multivalent vaccine
easy to produce (Raviprakash et al. 2009). Nevertheless, non-replicating vaccines
are not as effective as replicating vaccines, since they can not replicate in host cells
and mimick natural infection that induces adequate long lasting immunity
(Reviewed in Chaturvedi et al. 2005, Raviprakash et al. 2009). In addition,
genetically engineered vaccines based on particular components of DEN virus have
limitations for the immunity against other structural and non-structural components.
The utilization of more than one vaccine platform in a prime-boost strategy have
also been tried to discover an ideal DEN vaccine, which is tetravalent effective, safe
and globally affordable. The development of DEN vaccine still requires long-term
intensive studies.
Table 2 Dengue vaccine candidates in clinical and pre-clinical (NHP) trials
Replicating/ Vaccine* Monovalent (M)/ Status
Non-replicating Tetravalent (T)
Replicating LAV T Clinical trial
Replicating rLAV T Pre-clinical trial
Replicating YFV-DV T Clinical trial
Non-replicating Ad-vectored T Pre-clinical trial
Non-replicating PIV M Pre-clinical trial
Non-replicating Subunit M Clinical trial
Non-replicating DNA T Pre-clinical trial
Non-replicating VRP M Pre-clinical trial
*LAV: live attenuated vaccine, rLAV: recombinant LAV, YFV-DV: Yellow fever
virus-dengue virus chimera. Ad-vectored; Adeno vectored vaccine, PIV: purified
inactivated vaccine, Subunit: recombinant subunit protein vaccine, DNA: DNA
vaccine, VRP: venezuelan equine encephalitis replicon particle (Adapted from a
review by Raviprakash et al. 2009).

9
Compared with DEN vaccine, the development of antiviral to DEN virus
infection is still near the begining. There has been only few reports of DEN antiviral
drugs in NHP pre-clinical phase and their inhibition effects were not satisfactory.
Prophylactic ribavirin given one day before DEN infection was inefficient to inhibit
viremia in rhesus macaques (Malinoski et al. 1990). A recombinant human IFN-α
that was injected one day after onset of viremia could reduced viral burden and
improved viral clearance, however, further studies for more suppression is still
required (Ajariyakhajorn et al. 2005). Most of DEN antiviral drugs are still in
design or in vitro evaluation (Noble et al. 2010).
Either DEN vaccine or DEN antiviral should be evaluated in NHP, before
clinical evaluation. However, the lack of DHF animal model has hampered the
evaluation of DEN vaccine and antiviral capability to prevent DHF. Since the level
of viremia is associated with severity of disease, both vaccine and antiviral are
evaluated based on their capability to protect the animals from viremia
(Raviprakash et al. 2009, Nobel at al. 2010) .
Animal model for dengue infection. A total of 18 species from six families
of NHPs were experimentally infected by DEN virus (Reviewed by Bente and Rico-
Hesse 2006). The Old World monkeys, the Cercopithecidae family: Japanese
macaque (M. fuscata), rhesus macaques (Macaca mulatta), cynomolgus macaques
(M. fascicularis), green monkeys (Cercopithecus aethiops), patas monkeys
(Erythrocebus patas), yellow baboons (Papio cynocephalus) and mangabeys
(Cercocebus sp.); the New World monkeys: night monkeys (Aotus sp.), squirrel
monkeys (Saimiri scureus), saimiri monkeys (Saimiri orstedii), white face monkeys
(Cebus capucinus), cotton-top marmosets (Sanguinus oedipus) and marmosets
(Marikini geoffroyi), black spider monkeys (Ateles fusciceps), red spider monkeys
(A. geoffroyi), howler monkeys (Aluoatta palliata); and some Apes: chimpanzees
(Pan troglotdytes) and white handded gibbons (Hylobates lar) were used to study
DEN infection. Some of them were susceptible to DEN infections in terms of
detectable viremia and/or antibody response. However, none of these animals shows
clinical sign. Rhesus and cynomolgus macaques are the most common animal
model for the evaluation of DEN vaccine and antiviral.

10
In spite of some interesting findings and increasing demand of pigtail
macaque (M. nemestrina) in the studies of human immunodeficiency virus (HIV),
there has been no report of pigtail macaque as a model for DEN study. Similar with
human and rhesus, pigtail possesses dendritic cell-specific intercellular adhesion
molecule-3-grabbing non-integrin (DC-SIGN), a type II membrane protein with a
C-type lectin functions as a receptor binding domain and transmission factor for
several viral pathogens (Baribaud et al. 2001). Unlike rhesus and cynomolgus
macaques that have tripartite motif 5α (TRIM5α), pigtail has TRIM5 η or TRIM5 θ
factor which is incapable to inhibit the reverse transcription of viral replication. This
fact has been associated with the exceptional susceptibility of pigtail macaque to
HIV and simian immunodeficiency virus infections (Brennan et al. 2007).
Dengue specific cytokine producing T lymphocyte measurements. The T
helper (Th) and T cytotoxic (Tc) cells are the central of cellular adaptive immunity
(Janeway et al. 2001). The main function of Th is to initiate the responses of other
cells. They are divided into two functional classes: Th1 and Th2 cells. The function
of Th1 is to activate the microbicidal properties of macrophages and to induce
memory B cells to produce IgG antibodies that are effective at opsonizing
extracellular pathogens for uptake by phagocytic cells. T helper2 cells secrete
cytokines which activate naïve antigen specific B cells to produce IgM antibodies.
The Tc cells have ability to lyse target cells.
A naïve T cell must recognize a foreign peptide bound to a self major
histocompatibility molecule (MHC) which is expressed by professional antigen
presenting cell (APC) such as macrophage, dendritic cell and B cell in order to be
activated. Peptides from intracellular pathogens that multiply in the cytoplasm are
carried into the cell surface by MHC class I molecules and activated Tc cells to kill
the cells and produce cytokines. Pathogens that replicate in intracellular vesicles or
extracellular pathogens and proteins that are internalized into the intracellular
vesicles are degraded by proteases within the vesicles. These peptide fragments bind
to MHC class II molecules and they are delivered to the surface membrane of APC
to activate Th cells. The details of degradation, transportation and presentation of
antigens by MHC class I and II molecules were reviewed by Hickling (1998).
T cell subsets and the cytokine produced are shown in Table 3. The CD stands
for cluster of differentiation, a term for a cell surface molecule that is associated

11
with one or more functions on the cells. The CD4 is usually used as a marker for Th
cells, while CD8 is mostly a marker for Tc. Interferon-γ is the most frequent
cytokine used to determine specific Th1 or Tc responses, since it is produced by
much higher percentage of T cells.
Table 3 Human T cells
T cell subset Phenotype Functions
Th1 CD4+ Production of IL-2, IFN- γ and TNF α
Th2 CD4+ Production of IL-4, IL-5, IL-6, IL-10
and IL-13
T cytotoxic CD4+ or CD8+ Lyse target cells, production of IFN-γ
and TNF α
* IL: interleukin. TNF: tumor necroting factor (adapted from Hickling 1998).
Antigen specific T cells can be detected and enumerated after a short term in
vitro antigen stimulation followed by ELISPOT or IC-FC to detect T cells on a
single cell level (Lecth and Scheibenbogen 2003). The ELISPOT uses 96-well
membrane plate and coats the surface of the membrane with anti-cytokine antibody
to traps antigen induced cytokine secretion around the cells. Then, additional of
enzyme coupled second anti-cytokine antibody and substrate visualizes bound
cytokine. This is a sensitive assay that can count 10 cytokine secreting cells per one
million PBMC (Lecth and Scheibenbogen 2003). Whereas IC-FC uses brefeldin A
to trap cytokine intracellularly following antigen stimulation. Subsequently, the
cells are permeabilized, thus, specific anti-cytokine antibody conjugated with
fluorocrome can pass into the cells and react with cytokines (Lecth and
Scheibenbogen 2003). The advantage of IC-FC is its ability to phenotype the cells.
By applying anti CD-3 (CD-3 is a T cell marker), anti-CD4 and anti-CD8
monoclonal antibodies conjugated with different fluorocromes, the flow cytometer
quantify characterized T cells by the fluorocromes that are bound on and inside the
cells (Lecth and Scheibenbogen 2003).

GENERAL METHODOLOGY
Two studies were conducted for the development of pigtail macaque as
animal model in DEN research. The first study explored the possibility of pigtail
macaque as an animal model for DEN infection. It was conducted under approved
protocols by the Institutional Animal Care and Use Committee (IACUC), Naval
Medical Research Unit-2 (NAMRU-2) number 98AUC02. The second study was a
development of DEN specific cellular immunity measurements. This study utilized
samples collected during other DEN studies under protocols approved by the
IACUC of the NAMRU-2 or Animal Care and Use Committee of Primate Research
Center, Institut Pertanian Bogor. The approval numbers were 02AUC05 for DEN-1
and DEN-4, 99AUC01 for DEN-2, and P.09-08-IR for DEN-3.
Study of pigtail macaque susceptibility to DEN infection. Specific
pathogen free (tuberculosis, simian retrovirus, simian immunodeficiency virus,
simian T-lymphotropic virus, and Flavivirus) adult pigtail macaques were selected
and housed in mosquito-proof rooms at the NAMRU-2 AAALAC International-
accredited animal facility.
Figure 3 shows an outline of DEN injections and blood collections in this
study. The animals were separated into four groups assigned to receive either DEN-
1, DEN-2, DEN-3 or DEN-4 virus. Each group received two inoculations of virus.
The first inoculation, two animals in each group received live virus and another two
received phosphate buffered saline (PBS). The second inoculation, all animals in
the group received live virus. Each group (DEN virus serotype), therefore,
consisted of four examples of primary infection and two of secondary infection. The
NAMRU-2 veterinarian observed the animals daily for ten days post-inoculation,
and recorded pulse, respirations, rectal temperature, body weights, food and water
intake. Prior to virus injection, a 3 ml anti-coagulated blood sample was obtained
from the femoral vein of each monkey and used for baseline. Following DEN virus
injection, 1 ml blood samples were obtained daily for 10 days for detection the
presence of DEN virus. At 14 and 28 days post-inoculation, additional blood
samples were obtained for anti-dengue antibody profile analyses.

13
Figure 3 Outline of virus injections and blood collections for pigtail susceptibility
study.
Laboratory assays were done in the Viral Diseases Program, NAMRU-2. To
detect the presence of DEN virus, serum samples were analyzed by standard tissue
culture in C6/36 cells, mosquito inoculation and reverse transcriptase-polymerase
chain reaction (RT-PCR). The profiles of IgM, IgG, avidity IgG, subclasses IgG and
neutralizing antibody were analyzed. Reagents used for laboratory assays are listed
in Appendix 1. The cycles of RT-PCR and semi-nested PCR are in Appendix 2.
Detail procedures are described on page 20 to 22.
A development of DEN specific cellular immunity measurements.
Figure 4 shows the outline of DEN injections and blood collections for the
development of cellular immunity measurements. Peripheral blood mononuclear
cells were isolated from three ml heparinized blood by density gradient
centrifugation using Ficoll-Hypaque (Sigma Aldrich, St, Louis, Mo). The collected
PBMC were then, resuspended in 1 ml medium with 20% fetal bovine serum
(Invitrogen, Grand Island, NY) and 7.5% dimethyl sulfoxide (Sigma Aldrich, St.
Louis, Mo), and stored in liquid nitrogen.

14
Figure 4 Outline of DEN injections and blood collections for the study of
cellular immunity specific to DEN measurements.
To determine the response of pigtail to DEN infection, blood samples
collected before and one month after animal injected with DEN were used. Antigen
optimation was done using samples from uninfected animals that were taken for the
selection of Flavivirus-free animals and samples from infected animals that were
taken at the second month after challenged.
Dengue antigen was prepared from Vero cultures infected by either DEN-1
strain 16007, DEN-2 strain 16881, DEN-3 strain 16562 or DEN-4 strain 1036. Both
intra and extracellular DEN proteins were collected. Control antigen was prepared
similarly using uninfected Vero cells. The protein concentration in each antigen was
determined using bicinchoninic acid kit (Pierce, Rockford, IL). ELISA and western
blot were done to confirm the presence of dengue proteins.
A commercial ELISPOT assay specific for rhesus IFN- (Mabtech,
Stockholm, Sweden) was applied following the instruction from the manufacturer.
Phenotyping and intracellular cytokine detection was performed following
intracellular staining procedure from BD Biosciences and previously described
procedures (Pahar et al. 2003; Gauduin et al. 2004; Favre et al. 2009). All reagents
were optimized for pigtail PBMC. Reagents used for the cellular immunity
measurements were listed in Appendix 3. Positive control wells were stimulated
with 50 ng of phorbol 12-myristate 13-acetate (Sigma Aldrich, St. Louis, Mo) and
250 ng ionomycin (Sigma Aldrich, St. Louis, Mo). Wells with medium only

15
represented spontaneous IFN-γ producing cells. Detail assay procedures are
described on page 36 to 39.

Pigtail macaque (Macaca nemestrina) and Dengue Virus Infectivity:
a Potential Model for Evaluating Dengue Vaccine Candidates*
Susana Widjaja 1, Imelda Winoto
1, Jonathan Sturgis
1, Chairin N Maroef
1,
Erlin Listiyaningsih1, Ratna Tan
1, Joko Pamungkas
2,3, Diah Iskandriati
3,
Patrick J Blair1, Dondin Sajuthi
3,4** and Kevin Randall Porter
5
1Naval Medical Research Unit #2, Jalan Percetakan Negara 23, Jakarta 10560,
Indonesia;
2Department of Animal Diseases and Veterinary Public Health, School of
Veterinary Medicine, Institut Pertanian Bogor, Jalan Agatis, Bogor 16680,
Indonesia;
3Primate Research Center, Institut Pertanian Bogor, Jalan Lodaya II/5, Bogor
16151, Indonesia;
4Department of Clinic, Reproductive and Pathology, School of Veterinary Medicine,
Institut Pertanian Bogor, Jalan Agatis, Bogor 16680, Indonesia;
5Naval Medical Research Center, 503 Robert Grant Avenue, Silver Spring,
Maryland 20910, USA
**Corresponding author: Phone/Fax: +62-251-8314371,
E-mail: [email protected]
* Microbiology Indonesia. 2010. 2:1-6.

17
ABSTRACT
Pigtail macaque (Macaca nemestrina) has been shown to respond to
infectious disease agents, such as HIV, and is more sensitive compared to other
species of macaques such as rhesus (M. mulatta) and cynomolgus monkeys (M.
fascicularis). To evaluate pigtail macaque for the ability to support dengue (DEN)
viremia and serve potentially as an improved model for testing DEN vaccines, a
series of experiments were conducted using primary viral isolates from individuals
with DEN virus infections. This study shows that pigtail macaques develop
consistent, measurable viremia with all four DEN serotypes and produce immune
responses sufficient to protect against homologous virus. Anti-dengue antibodies
generated after infection are predominately IgG1. This species of macaque
therefore appears to be a suitable model for testing DEN virus vaccine candidates.
Keywords: dengue infection, Macaca nemestrina, viremia, antibody.
INTRODUCTION
Dengue fever (DF) and dengue hemorrhagic fever (DHF) are the most
important arthropod-borne viral diseases worldwide. An estimated 100 million DF
cases occur every year in dengue (DEN) endemic regions of the world (Halstead
1988). DHF, the more severe form of DEN infection, is associated with a mortality
of 1% to 5% and may be as high as 30% to 40% in untreated patients. The
tremendous public health impact of this disease emphasizes the need for an effective
preventive DEN vaccine.
There are many DEN vaccine candidates in clinical and pre-clinical trials.
Pre-clinical trials usually involve the evaluation of promising vaccine candidates in
non-human primates (NHP). Since DHF manifests only in humans, the model for
testing the efficacy of a DEN vaccine centers on the vaccine’s ability to prevent, or
significantly reduce, viremia after vaccinated animals are challenged with live DEN
virus. Rhesus macaques (Macaca mulatta) have been used mostly as a model for
pre-clinical trials, because this species of NHP supports DEN virus replication and
manifests consistent detectable viremia (Angsubhakorn et al. 1988, Eckels et al.
1994, Bray et al. 1996, Raviprakash et al. 2000, 2006, 2008, Durbin et al. 2001,
Putnak et al. 2003, 2005, Sun et al. 2005). Seventeen other species of NHP have
been used as models for dengue infections and vaccine trials (Bente and Rico-Hesse

18
2006). However, the susceptibility of the pigtail macaque (M. nemestrina) to
infection with DEN has not been tested.
Similar to rhesus, pigtail macaques possess dendritic cell-specific
intercellular adhesion molecule-3-grabbing non-integrin (DC-SIGN) that has similar
characteristics and functions as human DC-SIGN (Baribaud et al. 2001). This type
II membrane protein with a C-type lectin functions as a receptor binding domain for
dengue virus and several viral pathogens such as HIV-1 and influenza A/H5N1.
This fact supports the possibility of pigtail macaques as a model for DEN infection.
However, unlike other macaques, pigtails have been shown to be susceptible to HIV
infection. Pigtail macaques possess the cytoplasmic body protein TRIM5ø, which
is incapable of restricting HIV replication after viral entry to host cells. Other
macaques posses TRIM5α, which inhibits reverse transcription of retrovirus
(Stremiau et al. 2004, Brennan et al. 2007). The pigtail is the most potential
macaque model in which HIV can cause as AIDS-like syndrome in non-human
species (Agy et al. 1992, Baroncelli et al. 2008, Hatziioannou et al. 2009).
In this study, we evaluated the ability of dengue to replicate in pigtail
macaques. A series of experiments where pigtail macaques were inoculated with all
four serotypes of dengue virus were conducted. Viremia and anti-dengue antibody
responses were studied and revealed that this monkey species may serve as a
suitable model for evaluating experimental dengue vaccines.
MATERIALS AND METHODS
Animals. Thirty specific pathogen free (free of tuberculosis, simian
retrovirus, simian immunodeficiency virus, and simian T-lymphotropic virus)
pigtail macaques (M. nemestrina), between the ages of six months and one year,
were screened for anti-Flavivirus antibodies by hemaglutination inhibition assay,
IgG and IgM ELISA (Focus Diagnostics, Cypress, CA) and plaque reduction
neutralization tests (DEN and Japanese Encephalitis). The animals were obtained
from the Primate Research Center, Bogor, West Java. Seventeen flavivirus naïve
monkeys were selected and housed in mosquito-proof rooms at the Naval Medical
Research Unit #2 AAALAC International-accredited animal facility. The animals
ranged in weight from 2 kg to 3.5 kg and were pair caged with another animal
inoculated with the same dengue virus serotype. Animal care was administered
according to the Guide for the Care and Use of Laboratory Animals (NRC 1996).

19
The study was conducted under a protocol approved by the Institutional Animal
Care and Use Committee of the Naval Medical Research Unit #2 number
98AUC02.
Virus Inoculation and Blood Samples. The animals were separated into
four groups of four animals each. One monkey was used as an alternate to replace
any animal that needed to be excluded during the study for any reason. Each group
was assigned to receive either DEN-1, DEN-2, DEN-3 or DEN-4 virus. Each group
received two inoculations of virus. For the first inoculation, two animals in each
group received virus and two received phosphate buffered saline (PBS). For the
second inoculation, all animals in the group received live virus. Each group (DEN
virus serotype) therefore consisted of four examples of primary infection and two of
secondary infection. The extra monkey was later included in the DEN-3 group.
The DEN-1 virus used was isolated from a DEN fever patient hospitalized in
Jakarta, Indonesia and passaged five times in C6/36 cell culture. DEN-2, DEN-3
and DEN-4 isolates were derived from patients hospitalized in Palembang, Bandung
and Yogyakarta, Indonesia, respectively. The DEN-2 isolate was passaged five
times and the DEN-3 and DEN-4 isolates were passaged 4 times in C6/36 cell
culture. All isolates were obtained in 1998 with the exception of the DEN-4 isolate
that was obtained in 1996. Virus stocks were prepared from clarified cell culture
supernatant and stored at –70oC until used. The cells were used to confirm DEN
serotype and to rule out a possibility of other related viruses contamination by
indirect fluorescence assay using monoclonal antibodies to DEN-1 through DEN-4,
polyclonal antibodies to Flavivirus and Alphavirus. For each inoculation,
approximately 105 plaque-forming units (PFU) were administered subcutaneously in
the lateral chest area. Prior to inoculation, the site was shaved and cleaned with
70% alcohol.
Prior to virus injection, a 3 mL anti-coagulated blood sample was obtained
from the femoral vein of each monkey and used for collecting plasma and archiving
peripheral blood mononuclear cells (PBMC). Following the inoculation, 1 mL
blood samples were obtained daily for 10 days for virus isolation in C6/36 cells and
mosquito inoculation and virus detection by RT-PCR. The NAMRU-2 veterinarian
observed the animals daily for ten days post-inoculation recording pulse,
respirations, rectal temperature, body weights, food and water intake. At 14 and 28

20
days post-inoculation, additional blood samples were obtained for anti-dengue
antibody profile and antibody avidity analysis.
Dengue Virus Detection by RT-PCR. Qiamp Viral RNA mini kit
(QIAGEN Gmbh, Hilden, Germany) was used to extract viral RNA from 140 μL of
serum sample following manufacturer’s instruction. A total of 60 μL RNA was
obtained and 5 µL used in the RT-PCR reaction. The methods of Lanciotti were
used for the RT-PCR and semi-nested PCR (Lanciotti et al. 1997). PCR products
were resolved by electrophoresis using a 2% agarose gel and ethidium bromide
staining. Dengue viremic serum and negative serum were used as control positive
and negative.
Dengue Virus Isolation. Serum samples obtained post-infection for 10
days were analyzed for the presence of virus by standard tissue culture in C6/36
cells and by mosquito inoculation. Virus isolation in cell culture was performed as
described by Graham et al. (1999). For mosquito inoculation, Toxorhynchites
mosquitoes were used following the method of Yamamoto et al. (1987).
IgM and IgG Analysis. Anti-dengue IgM and IgG antibodies were detected
using a commercially available antibody capture ELISA kit (Focus Diagnostic,
Cypress, CA). Assays were performed following manufacturer’s procedures. A
numerical index was calculated by dividing the OD of sample with OD of the cutoff
control. A sample with an index greater than or equal to 1 was considered DEN
antibody positive. Serum samples drawn on day 0, day 14 and day 28 were tested
by IgM ELISA, while samples drawn at day 0, day 14, day 28 and day 87 were
tested by IgG ELISA.
Antibody Avidity Assay. The method of Gassmann et al. (1997) was used
with some modifications, to evaluate the avidity of anti-DEN IgG antibody. To
determine the appropriate urea concentrations to use in the test, high avidity and low
avidity positive control sera, diluted 1:100, were tested at different urea
concentrations that ranged from 6 M to 9 M in 0.5 M increments. Using the best
urea concentration, 6.5 M, the samples were tested in duplicate for IgG as indicated
above. After the initial sample incubation, one duplicate plate was incubated for 3
minutes with 6.5 M urea in PBS and then washed three times. The plates were
further processed according to the usual procedure. For samples giving an OD
value >2.0, the assay was repeated using two-fold serial dilutions starting at a 1:100

21
dilution. For samples giving an OD less than 0.6, samples were re-tested at two-
fold dilutions starting at 1:10.
For samples tested at a single 1:100 dilution, the avidity index was
calculated by dividing the OD from the urea treated sample by the OD of the
untreated control. For samples requiring serial dilutions, fine determinations of the
avidity indexes were calculated by dividing the dilution of the urea-treated curve
necessary for a defined OD by the respective dilution of the control curve at the
same OD. The defined OD was selected in the range 0.25–0.60 fold of the maximal
OD.
Antibody Subclass Analysis. The subclasses of anti-DEN IgG produced in
response to live virus infection were studied. The method of Shearer et al. (1999)
was used for this analysis. The distribution of IgG subclasses was examined with
the use of indirect ELISA. Briefly, dengue cell lysate antigen (DEN ag) and Vero-
76 cell lysate antigen (mock ag) in carbonate-bicarbonate buffer pH 9.6 were coated
onto five U8 Maxisorb microtiter plates (Nunc, Roskilde, Denmark). The plates
were incubated overnight at 4oC. The plates were washed with phosphate buffer
saline pH 7.4 containing 0.1% Tween 20 (PBS-T) six times and 100 μL of serum
(diluted 1/100 in PBS with 0.1% Tween 20 and 5% defatted milk powder) was
added into each well. After 1 hr serum incubation in 37oC, plates were washed.
Horseradish peroxidase labeled sheep anti human IgG1 to IgG4 (The Binding Site,
Birmingham, UK) at 1/50 dilution and horseradish peroxidase labeled goat anti
human IgG (Accurate, Westbury, USA) at 1/1000 dilution were added into each
plate, so plate 1 was incubated with anti-IgG1, plate 2 with anti-IgG2 and so on.
The plates were incubated 1 hour in 37oC. The 2.2’-azino-di[3-ethyl-benzthiazoline
sulfonate (6)] (ABTS) substrate (Kirkegaard and Perry, Gaithersburg, USA) was
added after the plates were washed. Following another one hour incubation at 37oC,
the absorbance was measured at 415 nm. The specific absorbance for each serum
sample was calculated as the mean A 415 nm DEN ag - the mean A 415 nm mock ag.
Checkerboard titrations of Den ag and the peroxidase labeled anti-Flavivirus 4G2
(Kikergard and Perry, Gaithersburg, USA) were performed to determine the optimal
dilution of DEN-1 to DEN-4 cell lysate ags.
Plaque Reduction Neutralization Test and Plaque Assay. Neutralizing
antibody titers at day 0, 14 and 28 were determined by standard plaque reduction

22
neutralization as performed by Morens et al. (1985) and the results expressed as the
reciprocal dilution that produces a 50% reduction in plaque count. Plaque assay
was performed to determine the virus titer during viremia by inoculating 1:10
diluted serum in PBS/BA into BHK12 Clone-15 suspension.
RESULTS
Clinical Signs and Symptoms. Following infection, the animals were
monitored daily for any signs and symptoms of DEN infection. There were no
abnormal changes in temperature or respirations following live virus injection. The
animals’ weights remained stable before, during and after viremia. No obvious
decreases in food consumption were noted.
DEN Viremia Following Infection. There were 16 episodes of
experimental primary infection among the animals. Figure 5 shows the days of
viremia for each DEN serotype according to the different virus detection methods.
Almost all of the animals became viremic within 1 to 2 days after DEN injections.
As expected, RT-PCR was the most sensitive method for detecting viremia.
However, consistent viremia was detected in each of the animals by all three
methods. DEN-2 resulted in the greatest average number of days viremia at
7.8±0.5, 6.8±1 and 5.8±1 days as measured by RT PCR, isolation in C6/36 and
mosquito inoculation, respectively. If going by RT-PCR and mosquito inoculation
methods, the least number of viremia days occurred with DEN-4 (5±1.4 and 3.3±1).
By isolation in C6/36, DEN-3 produced the least (4± 0.8).
Animals that were infected with each DEN serotype were re-challenged with
the homologous virus. None of the animals re-challenged with DEN-1, DEN-2 or
DEN-3 developed viremia. Two animals with secondary DEN-4 infections both
developed breakthrough viremia as detected by RT-PCR. DEN-4 was detected in
one animal on days 6 and 7 and in the other only on day 7.
Plaque assays were performed to quantify the amount of virus present. All
attempts failed to produce plaques and it was concluded that the level of viremia
was probably too low for detection by this method.
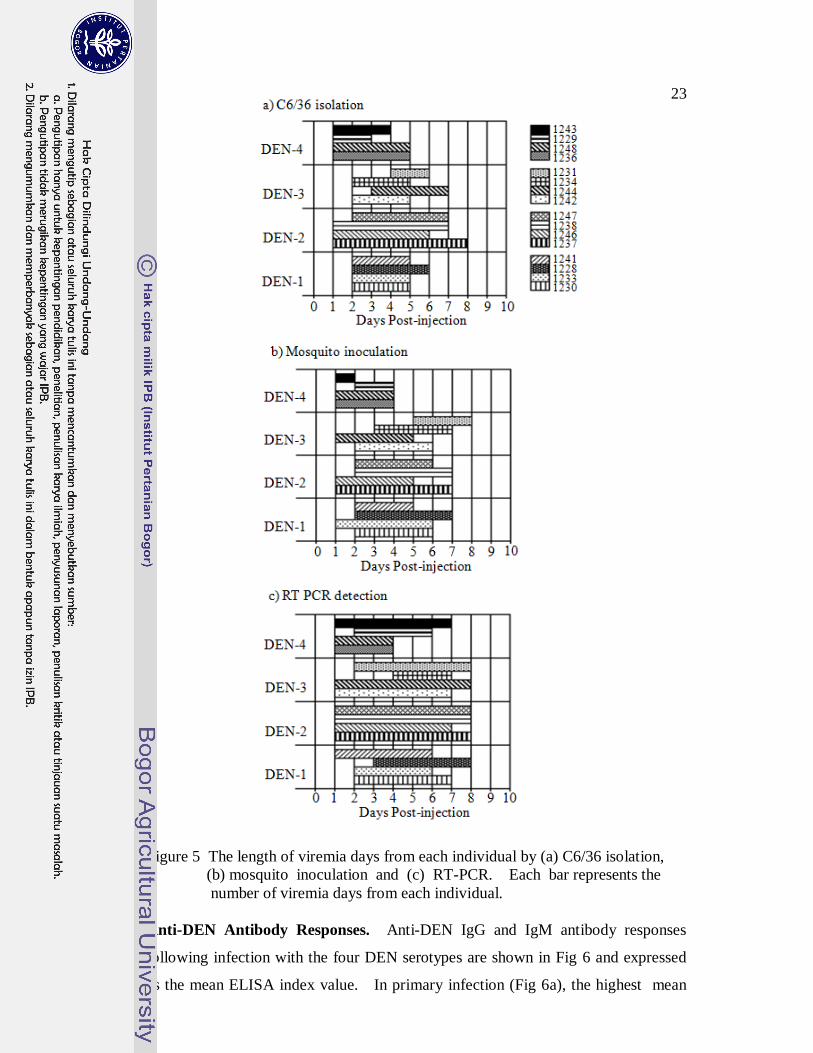
23
Figure 5 The length of viremia days from each individual by (a) C6/36 isolation,
(b) mosquito inoculation and (c) RT-PCR. Each bar represents the
number of viremia days from each individual.
Anti-DEN Antibody Responses. Anti-DEN IgG and IgM antibody responses
following infection with the four DEN serotypes are shown in Fig 6 and expressed
as the mean ELISA index value. In primary infection (Fig 6a), the highest mean
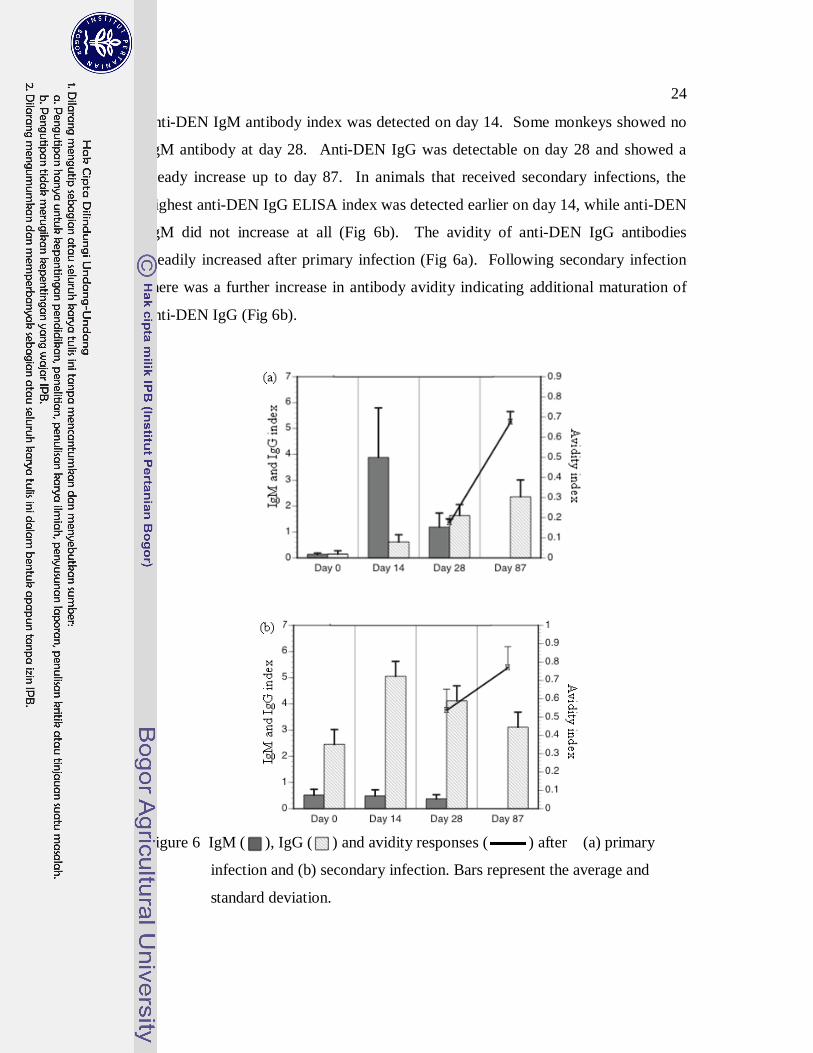
24
anti-DEN IgM antibody index was detected on day 14. Some monkeys showed no
IgM antibody at day 28. Anti-DEN IgG was detectable on day 28 and showed a
steady increase up to day 87. In animals that received secondary infections, the
highest anti-DEN IgG ELISA index was detected earlier on day 14, while anti-DEN
IgM did not increase at all (Fig 6b). The avidity of anti-DEN IgG antibodies
steadily increased after primary infection (Fig 6a). Following secondary infection
there was a further increase in antibody avidity indicating additional maturation of
anti-DEN IgG (Fig 6b).
Figure 6 IgM ( ), IgG ( ) and avidity responses ( ) after (a) primary
infection and (b) secondary infection. Bars represent the average and
standard deviation.
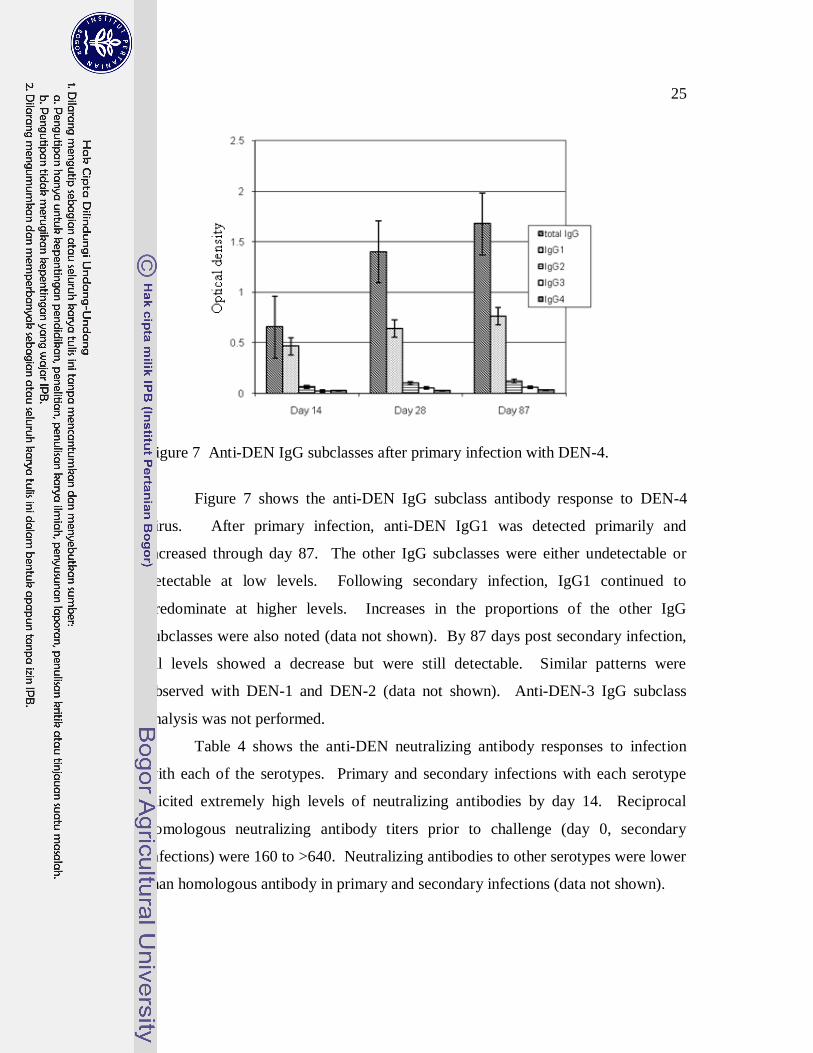
25
Figure 7 Anti-DEN IgG subclasses after primary infection with DEN-4.
Figure 7 shows the anti-DEN IgG subclass antibody response to DEN-4
virus. After primary infection, anti-DEN IgG1 was detected primarily and
increased through day 87. The other IgG subclasses were either undetectable or
detectable at low levels. Following secondary infection, IgG1 continued to
predominate at higher levels. Increases in the proportions of the other IgG
subclasses were also noted (data not shown). By 87 days post secondary infection,
all levels showed a decrease but were still detectable. Similar patterns were
observed with DEN-1 and DEN-2 (data not shown). Anti-DEN-3 IgG subclass
analysis was not performed.
Table 4 shows the anti-DEN neutralizing antibody responses to infection
with each of the serotypes. Primary and secondary infections with each serotype
elicited extremely high levels of neutralizing antibodies by day 14. Reciprocal
homologous neutralizing antibody titers prior to challenge (day 0, secondary
infections) were 160 to >640. Neutralizing antibodies to other serotypes were lower
than homologous antibody in primary and secondary infections (data not shown).
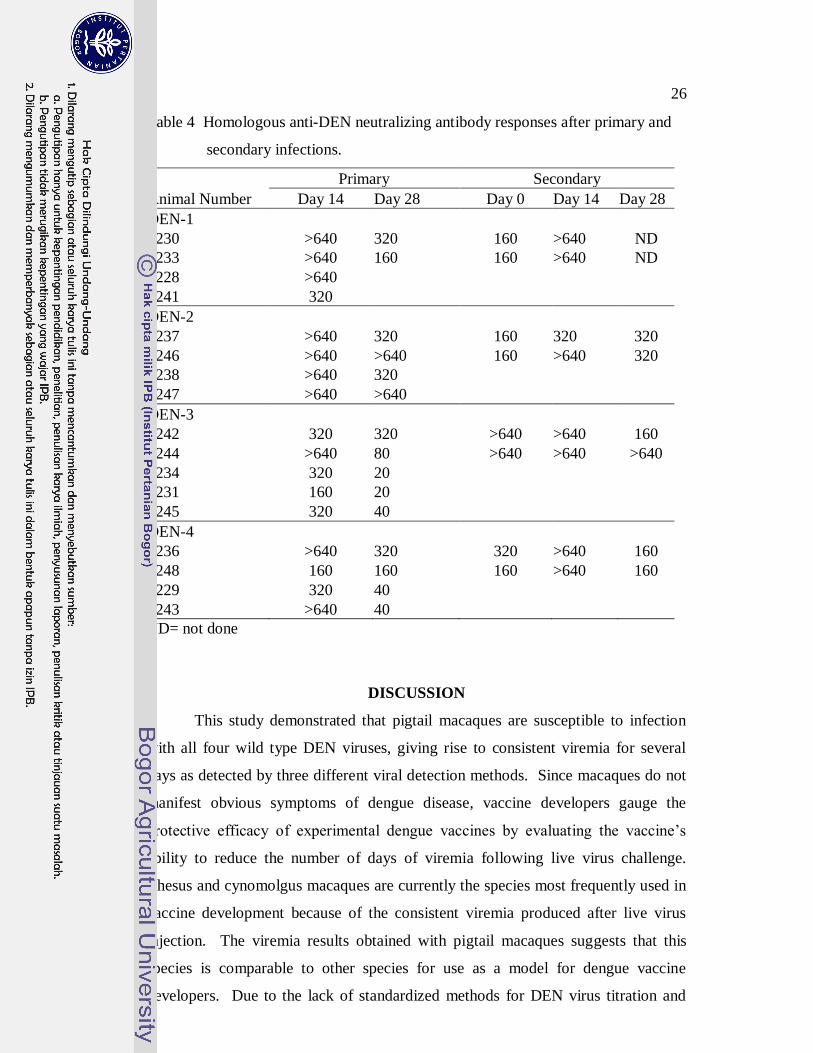
26
Table 4 Homologous anti-DEN neutralizing antibody responses after primary and
secondary infections.
Primary Secondary
Animal Number Day 14 Day 28 Day 0 Day 14 Day 28
DEN-1
1230 >640 320 160 >640 ND
1233 >640 160 160 >640 ND
1228 >640
1241 320
DEN-2
1237 >640 320 160 320 320
1246 >640 >640 160 >640 320
1238 >640 320
1247 >640 >640
DEN-3
1242 320 320 >640 >640 160
1244 >640 80 >640 >640 >640
1234 320 20
1231 160 20
1245 320 40
DEN-4
1236 >640 320 320 >640 160
1248 160 160 160 >640 160
1229 320 40
1243 >640 40
ND= not done
DISCUSSION
This study demonstrated that pigtail macaques are susceptible to infection
with all four wild type DEN viruses, giving rise to consistent viremia for several
days as detected by three different viral detection methods. Since macaques do not
manifest obvious symptoms of dengue disease, vaccine developers gauge the
protective efficacy of experimental dengue vaccines by evaluating the vaccine’s
ability to reduce the number of days of viremia following live virus challenge.
Rhesus and cynomolgus macaques are currently the species most frequently used in
vaccine development because of the consistent viremia produced after live virus
injection. The viremia results obtained with pigtail macaques suggests that this
species is comparable to other species for use as a model for dengue vaccine
developers. Due to the lack of standardized methods for DEN virus titration and

27
detection, making comparisons between different macaque species regarding their
ability to produce viremia following live dengue virus infection may be
inappropriate. Nevertheless, the days of viremia caused by DEN-1 and DEN-4
primary infections in this study were slightly longer than the days of viremia seen in
cynomolgus macaques infected with these serotypes (Koraka et al. 2007). In
addition, the overall length of viremia in pigtails was longer than that seen in earlier
rhesus monkey studies (Halstead et al. 1973, Freire et al. 2007). One prior study
with rhesus monkeys showed that several strains of DEN viruses failed to produce
viremia (Freire et al. 2007). The current study utilized only a single wild type
dengue viral isolate for each serotype. It remains to be determined if the viremia
characteristics seen with this set of viruses is characteristic of the viremia that
would occur if other viruses were used. While in theory, the same level of viremia
should occur with other viruses, further evaluation of the pigtail macaque with other
isolates is warranted.
Daily observations during the viremic period did not detect any overt signs
of clinical illness. The animals maintained their weight and food intake and no
obvious elevations in temperature were noted. This lack of clinical illness was
consistent with observations made in earlier studies of chimpanzees (Price et al.
1974, Scherer et al. 1978), owl monkeys (Schiavetta et al. 2003), rhesus monkeys
(Price et al. 1974, Freire et al. 2007) and cynomolgus monkeys (Price et al. 1974,
Koraka et al. 2007) as well as in other studies reviewed by Bente and Rico-Hesse
(2006).
The anti-DEN IgM and IgG antibody responses to each of the serotypes
were similar to primary and secondary responses seen in human dengue virus
infections (Innis 1997). Anti-dengue IgM was detected early and peaked within 2
weeks (2-4 weeks) after primary infection. Anti-DEN IgG was detected later and
increased slowly. Following secondary infection, anti-DEN IgM was low or
undetectable in some instances, as seen in dengue endemic areas where multiple
serotypes circulate. Anti-DEN IgG rapidly increased over the next 2 weeks.
Antibody avidity assay demonstrated the progressive increase in affinity in these
antibodies over time after primary infection. Secondary infection induced only a
slight increase in anti-DEN IgG antibody maturation. Avidity responses were also
similar with the response in human (US NAMRU-2 unpublished data).

28
There was no anti-pigtail IgG subclass available commercially, when IgG
subclass assay was set. Anti-human IgG subclasses were chosen, since these
reagents bound more anti-pigtail IgG compared with anti-macaque IgG subclasses
(data not shown). Anti-DEN IgG1 was primarily detected and predominant IgG
subclass following primary and secondary infections in pigtails. In humans, IgG1
and IgG3 were the predominant IgG subclasses throughout the course of illness
regardless of whether the illness was characterized as DF, DHF or DSS (Thein et al.
1993; Koraka et al. 2001). In this study, it is not further determined whether this
different due to different species response or less cross reaction of anti-human IgG3
than anti-human IgG2.
High-titer anti-DEN neutralizing antibodies were produced in response to
dengue virus infection in each animal and demonstrated cross-reactivity against the
DEN serotypes used in the plaque reduction neutralization assay. These antibodies
resulted in solid protection against challenge with the homologous dengue virus
serotype, although minimal break-through viremia occurred in two DEN 4 animals
as detected by RT-PCR. Because no live virus was detected in these animals, this
could have just represented residual RNA from non-viable virus.
The ability of pigtail monkeys to support the replication of all four
serotypes of dengue raises the question of whether this species is suitable for
evaluating the efficacy of experimental dengue vaccines and anti-DEN therapeutic
drugs. Further studies are planned to evaluate the efficacy of DNA-based vaccines
using the pigtail model.
ACKNOWLEDGMENT
This work was supported by the Naval Medical Research Center work unit
61102A.S13.S.S1415. The views expressed in this article are those of the authors
and do not necessarily reflect the official policy or position of the Department of the
Navy, Department of Defense, nor the US Government. Authors (SW, IW, JS,
CNM, EL, RT, PJB, KRP) as employees of the U.S. Government or military service
members, conducted the work as part of their official duties. Title 17 U.S.C. §105
provides that ‘Copyright protection under this title is not available for any work of
the United States Government.’ Title 17 U.S.C. §101 defines a US Government
work as a work prepared by a military service member or employee of the US
Government as part of that person’s official duties.

29
REFERENCES
Agy MB, Frumkin LR, Corey L, Coombs RW, Wolinsky SM, Koehler J, Morton
WR, Katze MG. 1992. Infection of Macaca nemestrina by human
immunodeficiency virus type-1. Science 257:103-6.
Angsubhakorn S, Yoksan S, Bhamarapravati N, Moe JB, Marchette NJ,
Pradermwong A, Sahaphong S. 1988. Dengue-4 vaccine: neurovirulence,
viraemia and immune responses in rhesus and cynomolgus monkeys. Trans R
Soc Trop Med Hyg 82:746-9.
Baribaud F, Pohlmann S, Sparwasser T, Kimata MT, Choi YK, Haggarty BS,
Ahmad N, Macfarlan T, Edwards TG, Leslie GJ, Arnason J, Reinhart TA,
Kimata JT, Littman DR, Hoxie JA, Doms RW. 2001. Functional and antigenic
characterization of human, rhesus macaque, pigtailed macaque and murine DC-
SIGN. J Virol 75:10281-9.
Baroncelli S, Negri DRM, Michelini Z, Cara A. 2008. Macaca mulata, fascicularis
and nemestrina in AIDS vaccine development. Expert Rev Vaccine 7:1419-34.
Bente DA, Rico-Hesse R. 2006. Models of dengue virus infection. Drug Discov
Today 3:97-103.
Bray M, Men R, Lai CJ. 1996. Monkeys immunized with intertypic chimeric
dengue viruses are protected against wild-type virus challenge. J Virol
70:4162-6.
Brennan G, Kozyrev Y, Kodama T, Hu S. 2007. Novel TRIM5 isoforms expressed
by Macaca nemestrina. J Virol 22:12210-7.
Durbin AP, Karron RA, Sun W, Vaughn DW, Reynolds MJ, Perreault JR, Thumar
B, Men R, Lai CJ, Elkins WR, Chanock RM, Murphy BR, Whitehead SS.
2001. Attenuation and immunogenicity in humans of a live dengue virus type-
4 vaccine candidate with a 30 nucleotide deletion in its 3'-untranslated region.
Am J Trop Med Hyg 65:405-13.
Eckels KH, Dubois DR, Summers PL, Schlesinger JJ, Shelly M, Cohen S, Zhang
YM, Lai CJ, Kurane I, Rothman A, Hasty S, Howard B. 1994. Immunization
of monkeys with baculovirus-dengue type-4 recombinants containing envelope
and nonstructural proteins: evidence of priming and partial protection. Am J
Trop Med Hyg 50:472-8.

30
Freire MS, Marchevvsky RS, Almeida LFC, Yamamura AMY, Caride EC,Brindeiro
PA, Motta MCA, Nogueira RMR, Kubelka CF, Bonaldo MC, Galler R. 2007.
Wild dengue virus types 1, 2 and 3 viremia in rhesus monkeys. Mem Inst
Oswaldo Cruz 102:203-8.
Gassmann C, Bauer G. 1997. Avidity determination of IgG directed against Tick-
Borne Encephalitis virus improves detection of current infections. J Med Virol
51:242-51.
Graham RR, Juffrie M, Tan R, Hayes CG, Laksono I, Ma'roef C, Erlin, Sutaryo,
Porter KR, Halstead SB. 1999. A prospective seroepidemiologic study on
dengue in children four to nine years of age in Yogyakarta, Indonesia I. studies
in 1995-1996. Am J Trop Med Hyg 61:412-9.
Halstead SB. 1988. Pathogenesis of dengue: challenges to molecular biology.
Science 239:476-81.
Halstead SB, Shotwell H, Casals J. 1973. Studies on the pathogenesis of dengue
infection in monkeys, I. Clinical laboratory responses to primary infection. J
Infect Dis 128:7-14.
Hatziioannou T, Ambrose Z, Chung NPY, Piatak M Jr., Yuan F, Trubey CM,
Coalter V, Kiser R, Schneider D, Smedley J, Pung R, Gathuka M, Estes JD,
Veazey RS, KewalRamani VN, Lifson JD, Bieniasz PD. 2009. A macaque
model of HIV-1 infection. Proc Nat Acad Sci USA 106:4425-9.
Innis BL. 1997. Antibody responses to dengue virus infections. In: Gubler DJ,
Kuno G, editors. Dengue and dengue hemorrhagic fever. New York: CAB
International. p 221-43.
Koraka P, Benton S, Amerongen G, Stittelaar KJ, Osterhaus ADME. 2007.
Characterization of humoral and cellular immune responses in cynomolgus
macaques upon primary and subsequent heterologous infections with dengue
virus. Microbes Infect 9:940-6.
Koraka P, Suharti C, Setiati E, MAiruhu TA, Van Gorp, Hack CE, Juffrie M,
Sutaryo J, Van Der Meer GM, Groen J, Oesterhaus ADME. 2001. Kinetic of
dengue virus-specific serum immunoglobulin classes and subclasses correlate
with clinical outcome of infection. J Clin Microbiol 39: 4332-8.
Lanciotti RS, Gubler DJ, Trent DW. 1997. Molecular evolution and phylogeny of
dengue-4 viruses. J Gen Virol 78:2279-84.

31
Morens DM, Halstead SB, Repik PM, Putvatana R, Raybourne N. 1985.
Simplified plaque reduction neutralization assay for dengue viruses by
semimicro methods in BHK-21 cells: comparison of the BHK suspension test
with standard plaque reduction neutralization. J Clin Microbiol 22:250-4.
National Research Council. 1996. The guide for the care and use of laboratory
animals. Washington DC: National Academic Press.
Price WH, Casals J, O'Leary W. 1974. Studies on the sequential immunization
against group B arboviruses in squirrel monkeys, cynomolgus monkeys, rhesus
monkeys, and chimpanzees. Am J Trop Med Hyg 23:118-30.
Putnak RJ, Coller BA, Voss G, Vaughn DW, Clements D, Peters I, Bignami G,
Houng HS, Chen RC, Barvir DA, Seriwatana J, Cayphas S, Garcon N, Gheysen
D, Kanesa-Thasan N, McDonnel M, Humphreys T, Eckels KH, Prieels JP,
Innis BL. 2005. An evaluation of dengue type-2 inactivated, recombinant
suunit, and line-attenuated vaccine candidates in the rhesus macaqe model.
Vaccine 23:4442-52.
Putnak R, Fuller J, VanderZanden L, Innis BL, Vaughn DW. 2003. Vaccination of
rhesus macaques against dengue-2 virus with a plasmid DNA vaccine encoding
the viral pre-membrane and envelope genes. Am J Trop Med Hyg 68:469-76.
Raviprakash K, Apt D, Brinkman A, Skinner C, Yang S, Dawes G, Ewing D, Wu
SJ, Bass S, Punnonen J, Porter K. 2006. A chimeric tetravalent dengue DNA
vaccine elicits neutralizing antibody to all four virus serotypes in rhesus
macaques. Virology 353:166-73.
Raviprakash K, Porter KR, Kochel TJ, Ewing D, Simmons M, Phillips I, Murphy
GS, Weiss WR, Hayes CG. 2000. Dengue virus type 1 DNA vaccine induces
protective immune responses in rhesus macaques. J Gen Virol 7:1659-67.
Raviprakash K, Wang D, Ewing D, Holman DH, Block K, Woraratanadharm J,
Chen L, Hayes C, Dong JY, Porter K. 2008. A tetravalent dengue vaccine
based on a complex adenovirus vector provides significant protection in rhesus
monkeys against all four serotypes of dengue virus. J Virol 82:6927-34.
Scherer WF, Russell PK, Rosen L, Casals J, Dickerman RW. 1978. Experimental
infection of chimpanzees with dengue viruses. Am J Trop Med Hyg 27:590-9.

32
Schiavetta AM, Harre JG, Wagner E, Simmons M, Raviprakash K. 2003. Variable
suceptibility of the owl monkey (Aotus nancymae) to four serotypes of dengue
virus. Comtemp Top Lab Anim Sci 42:12-20.
Shearer MH, Dark RD, Chodosh J, Kennedy RC. 1999. Comparison and
characterization of immunoglobulin G subclasses among primates. Clin Diagn
Lab Immunol 6:953-8.
Stremiau M, Owens CM, Kiessling M, Autissier P, Sodroski J. 2004. The
cytoplasmic body component TRIM5alpha retricts HIV-1 infection in Old
World monkeys. Nature 427:848-53.
Sun W, Nisalak A, Gettayacamin M, Eckels KH, Putnak JR, Vaughn DW, Innis BL,
Thomas SJ, Endy TP. 2005. Protection of rhesus monkeys against dengue
virus challenge after tetravalent live attenuated dengue virus vaccination. J
Infect Dis 193:1658-65.
Thein S, Aaskov J, Myint TT, Saw TT, Zaw A. 1993. Changes in levels of anti-
dengue virus IgG subclasses in patients with diseases of varying severity. J
Med Virol 40:102-6.
Yamamoto N, Kimura T, Ohyama A. 1987. Multiplication and distribution of type
2 dengue and Japanese encephalitis viruses in Toxorhynchites splendens after
intrathoracic inoculation. Arch Virol 97:37-47.

The Measurements of Dengue Specific Interferon-γ Producing T
Lymphocytes in Pigtail Macaques (Macaca nemestrina)
Susana Widjaja 1,2,*
, Dasep Purwaganda1, Victor A Sugiharto
1, Imelda L Winoto
1,
Gary T Brice1, Timothy H Burgess
1, Kevin R Porter
3, Charmagne G Beckett
3,
Maya Williams1, Joko Pamungkas
2,4,5, Diah Iskandriati
2,5, Dondin Sajuthi
2,6 and
Patrick J Blair1
1Naval Medical Research Unit #2, Jalan Percetakan Negara 23, Jakarta 10560,
Indonesia;
2Primatology Major, Graduate Program, Institut Pertanian Bogor, Jalan Lodaya
II/5, Bogor 16151, Indonesia;
3Infectious Diseases Directorate, Naval Medical Research Center, Silver Spring,
Maryland;
4Department of Animal Diseases and Veterinary Public Health, School of
Veterinary Medicine, Institut Pertanian Bogor, Jalan Agatis, Bogor 16680,
Indonesia;
5Primate Research Center, Institut Pertanian Bogor, Jalan Lodaya II/5, Bogor
16151, Indonesia;
6Department of Clinic, Reproductive and Pathology, School of Veterinary
Medicine, Institut Pertanian Bogor, Jalan Agatis, Bogor 16680, Indonesia;
*Corresponding author: Telephone: 62 21 8615126/ 6281310858650
E-mail: [email protected]

34
ABSTRACT
Pigtail macaques (Macaca nemestrina) have been reported susceptible to the
infections of all four dengue (DEN) serotypes and suitable for DEN vaccine
evaluation. To enhance DEN study in pigtail macaques, ELISPOT and intracellular
cytokine staining-flow cytometry (IC-FC) were developed to measure DEN specific
interferon-γ (IFN-γ) producing T lymphocytes. Peripheral blood mononuclear cells
(PBMC) collected before and after DEN infections were tested. ELISPOT results
show increase of DEN specific IFN-γ producing cells as an individual response of
pigtail to DEN-1, DEN-3 or DEN-4 infection. ELISPOT and intracellular cytokine
staining-flow cytometry (IC-FC) were run side by side to quantitate DEN specific
lymphocytes following primary and secondary DEN-2 infections using pools of
PBMC. ELISPOT revealed an increase of DEN specific IFN-γ producing cells
following primary infection and a significant increase after secondary infection. The
patterns of DEN specific IFN-γ producing CD3+CD4+ and CD3+CD4- T
lymphocytes by IC-FC were similar with the pattern of DEN specific IFN-γ
producing cells. Therefore, ELISPOT and IC-FC can be used to study DEN specific
IFN-γ producing T lymphocytes in pigtail.
Keywords: Dengue, T lymphocytes, pigtail macaque, IFN-γ
INTRODUCTION
Consisting of four serotypes that are antigenically related (dengue-1 to
dengue-4), dengue viruses (DEN) are estimated to infect up to 100 millions human
annually (WHO 2005). Dengue infections can result in asymptomatic infections or
cause a variety of clinical manifestations ranging from undifferentiated fever, mild
dengue fever and up to 2-4% of the total cases develop into the life threatening
dengue hemorrhagic fever (DHF) or dengue shock syndrome (DSS) (WHO 2005).
The facts that the majority of fatal cases were the results of heterologous secondary
dengue infections, in which there were high level of activated T lymphocytes and
cytokines, emphasize the role of pre-existing memory T cells in determining the
severity of DEN disease (Mathew and Rothman 2008). However, the involvement
of memory T cells to the pathogenesis of DHD or DSS has not been completely
understood indicating the importance to study it intensively to discover more ways
in combating DEN (Mathew and Rothman 2008, Thomas et al. 2009).

35
Dengue pathogenesis study in humans has been very much limited as it is
difficult to obtain well-characterized cases that have serial samples from prior to or
early infections until late convalescent to get the thorough observation of
immunologic events in the course of DEN infections (Mangada et al. 2004,
Beckett et al. 2005, Porter et al. 2005). As such, having a valid animal model will
be invaluable to the progress of DHF or DSS study (Mathew et al. 2008,
Raviprakash et al. 2009). A variety of non-human primates (NHP) had been shown
to experience viremia and developed DEN infections, but no clinical sign (Bente
and rico-Hesse 2006, Mathew et al. 2008, Raviprakash et al. 2009). Compared with
rhesus (Macaca mullata) and cynomolgus macaques (M. fascicularis), commonly
used in dengue research, pigtail macaque (M. nemestrina) has defective factor
tripartite motif 5 (TRIM5) that has been associated with the susceptibility of
pigtail macaque to human immunodeficiency virus type-2 (HIV-2) and simian
immunodeficiency virus infections (Otten et al. 1994, Brennan et al. 2007). In spite
of interesting finding in retrovirus and lentivirus research, pigtail has not been
studied widely as a model for DEN disease (Raviprakash et al. 2009, Widjaja et al.
2010).
Enzyme-linked immunosorbent spot (ELISPOT) and intracellular cytokine
flow cytometry (IC-FC) have been applied to quantify SIV specific T lymphocytes
in rhesus macaques (Pahar et al. 2003, Gauduin et al. 2004). The ELISPOT assay
employs ELISA technique to trap antigen induced cytokine secretion around the
cells by an immobilized antibody and then visualize them by enzyme-coupled
second antibody (Lecth and Scheibenbogen 2003). While IC-FC uses brefeldin A to
trap cytokine intracellularly following antigen stimulation. Subsequently, the cells
are permeabilized, thus, specific anti-cytokine antibody conjugated fluorochrome
can pass into the cells and react with cytokines (Lecth and Scheibenbogen 2003).
These assays have become the preferential techniques since they measure functional
antigen specific T lymphocytes. They are more straightforward and faster than
conventional assays such as lymphocyte proliferation and cytotoxic assays (Pahar et
al. 2003).
Koraka et al. (2007a,b) applied ELISPOT and used antigen presenting cells
(APC) derived from autologous B cells to enumerate DEN-specific T lymphocytes
as the response of dengue infections and vaccination in cynomolgus macaques. In

36
the present study, we developed ELISPOT and IC-FC assays for the quantification
of DEN specific T lymphocyte in pigtail macaques. DEN lysate antigens were
applied to obtain in vitro stimulation of T lymphocytes (Mangada et al. 2004). Our
results show that both ELISPOT and IC-FC quantify the increase of DEN specific
IFN-γ producing CD4 T lymphocyte as the response of pigtail to DEN infections.
MATERIALS AND METHODS
Sample collection. Sample collections were done during previous dengue
studies conducted at the Naval Medical Research Unit-2 AAALAC International-
accredited animal facility, Jakarta and at the Primate Research Center, Bogor
Agricultural University, West Java. The studies utilized specific pathogen free
(tuberculosis, simian retrovirus, simian immunodeficiency virus, simian T-
lymphotropic virus and Flavivirus) pigtail macaques (M. nemestrina). Animal care
was administered according to the standard operating procedures of Laboratory
Animal Facility, Naval Medical Research Unit-2 or Primate Research Center
following the Guide for the Care and Use of Laboratory Animals (NRC 1996). The
studies were conducted under protocols approved by the Institutional Animal Care
and Use Committee (IACUC) of the Naval Medical Research Unit-2 and/or Animal
Care and Use Committee (ACUC) of Primate Research Center. The approval
numbers are 02AUC05 for DEN-1 and DEN-4 study, 99AUC01 for DEN-2 study
and P.09-08-IR for DEN-3 study.
Peripheral blood mononuclear cells (PBMC) were isolated from three ml
heparinized blood by density gradient centrifugation using Ficoll-Hypaque (Sigma
Aldrich, St, Louis, Mo). The collected PBMC were then, resuspended in 1 ml
medium with 20% fetal bovine serum (Invitrogen, Grand Island, NY) and 7.5%
DMSO (Sigma Aldrich, St. Louis, Mo), and stored in liquid nitrogen.
To determine the response of pigtail to DEN infection, blood samples
collected before and one month after animal injected with DEN were used. Antigen
optimation was done using samples from uninfected animals that were taken for the
selection of Flavivirus-free animals and samples from infected animals that were
taken at the second month after challenge.
Preparation of viral antigen. Dengue-2 (DEN-2) antigen was prepared from
DEN-2 infected Vero cells. Briefly, six T-175 flasks confluent monolayer Vero-76
cells were inoculated with DEN-2 strain 16681. The multiplicity of infection was

37
0.1. When 50-75% cytopathic effect (CPE) was observed, the cultures were kept in -
70oC and then freeze-thawed at least three times to break down the cells. The
suspension was centrifuged at 4000xg for 30 minutes. The supernatant was
collected and centrifuged for 2 hours at 48000xg. The pellet was re-suspended with
PBS and layered on 10% glycerol in PBS. Another round of centrifugation was
done for 2 hours at 63000xg. The pellet was collected and re-suspended in 3 ml
medium (RPMI 1640 with10% FBS, 2% 200 mM L-glutamine and 1% antibiotic
antimycotic). Control antigen was prepared similarly using uninfected Vero cells.
Dengue-1, DEN-3 and DEN-4 antigens were also prepared similarly using DEN-1
strain 16007, Den-3 strain 16562 and Den-4 strain 1036.
The protein concentration in each antigen was determined using bicinchoninic
acid kit (Pierce, Rockford, IL). Elisa and western blot were done using anti-
Flavivirus and anti-DEN monoclonal antibodies (Clone 4G2 and 2H2), peroxidase
conjugated anti-mouse IgG (KPL, Gaithersburg, MD) to confirm the presence of
DEN proteins.
Stimulation of peripheral blood mononuclear cells. Cryopreserved PBMC
were quickly thawed in 37oC water bath and re-suspended with 1 ml warm medium
(RPMI 1640 with 10% fetal bovine serum, 2% 200 mM L-glutamine and 1%
antibiotic-antimycotic) containing 10 U benzonase (Merck KGaA, Darmstadt,
Germany). After completely thawed, the tube was tapped gently to mix, then added
with 4 ml medium. The cells were centrifuged at 800xg for 5 minutes and washed
twice with medium. The obtained pellet was re-suspended with 1 ml medium and
10 l suspension was taken and mixed with 190 l Guava ViaCount (Millipore,
Billerica, MA) to determine the number of viable cells using Guava PC (Millipore,
Billerica, MA). A total of 2x105
PBMC in 100 l medium was added to each
ELISPOT well and 1x106 PBMC for ICC. After 100 l of antigen was added to each
well, the plates were incubated at 37oC in a humidified 5% CO2 atmosphere. Serial
two-fold dilutions of antigens were used to determine the optimum concentration
for stimulation. Positive control wells were stimulated with 50 ng of phorbol 12-
myristate 13-acetate (Sigma Aldrich, St. Louis, Mo) and 250 ng ionomycin (Sigma
Aldrich, St. Louis, Mo). Wells with medium only represented spontaneous IFN-γ
producing cells.

38
ELISPOT assay. A commercial ELISPOT assay specific for rhesus IFN-
was applied (Mabtech, Stockholm, Sweden). The assay was performed following
the instruction from the manufacturer. Briefly, 100 l anti- IFN- monoclonal
antibody (GZ-4) at 15 g/ml concentration in PBS was coated onto multi-screen 96-
well immobilon polyvinylidene difluoride (PVDF) plate (Millipore, Bedford, MA)
at 4oC over night. The plate was washed with PBS six times in sterile condition. The
PBMC were transferred to the plate and stimulated with 100 l antigen for 20 hours.
After the stimulation, the plate was decanted and washed six times with 0.05%
Tween-20 (Sigma Aldrich, St. Louis, Mo) in PBS (PBST). Biotinylated anti-IFN-
monoclonal antibody (7-B6-1-biotin) at concentration 1 g/ml in PBS with 0.5%
FBS was added and incubated for 2 hours at room temperature. After washing,
alkaline phosphatase conjugated streptavidin at the concentration 1 g/ml was
added and the plate was incubated for another hour at room temperature. The spots
were developed using 5-bromo-4chloro-3 indolyl phosphatase/nitroblue tetrazolium
(BCIP/NBT) substrate (Sigma Aldrich, San Jose, CA). The reaction was stopped by
rinsing the wells with tap water. The spot were counted visually using Gel-Doc
(Biorad, modified spot reader). The mean number of DEN specific IFN-γ producing
cells/106 PBMC was then calculated. The mean number of DEN specific IFN-γ
producing cells in negative antigen wells was used as background.
Intracellular staining and flow cytometry. A total of 1x106
PBMC in 100 l
medium was transferred to each well of 96-well tissue culture U-bottom plate and
stimulated with 100 l antigen for 20 hours at 37oC, 5% CO2. Two microgram of
Golgi PlugTM
(BD Biosciences, San Jose, CA) was added to each well for the final
16 hours of incubation. Phenotyping and intracellular cytokine detection was
performed following intracellular staining procedure from BD Biosciences and
previously described procedures (Pahar et al. 2003, Gauduin et al. 2004, Favre et al.
2009). All reagents were optimized for pigtail PBMC. After stimulation period, the
cells were washed twice with Facs wash (PBS Dulbecco with 0.5% FBS). One
microliter of peridinin chlorophyll cychrome5.5 (PerCp Cy5.5) conjugated anti-
CD4 (clone L200, BD Biosciences, San Jose, CA) and allophycocyanin cychrome-7
(APC Cy-7) anti-CD3 (clone SP34, BD Biosciences, San Jose, CA) were used for
the surface staining to determine cell phenotype. Following 20 minutes incubation

39
on ice, the cells were washed twice. Cells were then fixed and permeabilized with
Cytofix/CytopermTM
(BD Biosciences, San Jose, CA) for 20 minutes on ice. After
washing with PermwashTM
(BD Biosciences, San Jose, CA), intracellular IFN-
production was detected by APC anti-IFN- (clone B27, BD Biosciences, San Jose,
CA). A total of 500,000 events from the lymphocyte gate were collected from each
well using FacsArray (BD Biosciences, San Jose, CA). The data was analyzed using
FlowJo version 4.3 software (Treestar, CA) and CelQuest version 5.2.1 (BD
Biosciences, San Jose, CA). Lymphocytes and monocytes were separated based on
forward and side light scatter. The CD4+ and CD4- cells were plotted from CD3+
lymphocytes. The percentage of DEN-specific IFN-γ producing CD4+ or CD4-
cells were calculated based on the number of CD4+ or CD4- cells.
RESULTS
Optimization of antigens for ELISPOT. Optimum concentration of each
DEN antigen for ELISPOT was determined using PBMC from primary DEN-1,
DEN-3 and DEN-4 infected pigtail macaques. Homologous antigen was used for in
vitro stimulation and PBMC from uninfected pigtail macaques were used as control
PBMCs. Figure 8 shows the highest DEN-1 and DEN-4 antigen concentration
generated the most DEN specific IFN-γ producing cells. The concentration of 20
ug/ml antigen was used for DEN-1 and DEN-4. Dengue-3 antigen gave the best
response at 10 ug/ml.

40
(a) DEN-1
(b) DEN-3
(c) DEN-4
Figure 8 Dengue-1, dengue-3 dan dengue-4 antigen optimization by ELISPOT.
The bars represent DEN specific IFN-γ cells per one million PBMC.
ELISPOT for the quantification of DEN-specific T cells in pigtail
macaques. Table 5 shows the numbers of DEN specific IFN-γ producing cells from
individual pigtail before and after primary DEN infection. The range of DEN

41
specific IFN-γ cells was zero to 40 cells per one million PBMC before DEN
infections and 28 to 40 cells after the infections. Most pigtails showed increasing
DEN specific IFN-γ producing cells significantly after primary infection, however,
one pigtail infected by DEN-1 and one by DEN-4 only showed two times raise. The
range of DEN specific IFN-γ producing cells from naive animals was zero to 10
cells.
Table 5 DEN specific IFN-γ producing cells in pigtail macaques before and after
DEN infection.
IFN-γ producing cells /106 PBMC
Infecting serotype Animal ID Before After infection
DEN-1 5164 10 60
1.3238 10 30
1.3245 10 80
5079 10 28
5080 0 53
DEN-3 1.3074 0 440
1.6387 0 125
1.1267 0 145
1.5713 5 305
DEN-4 5161 15 55
5166 40 240
5072 20 420
5158 20 425
1.3308 30 70
Uninfected macaques* 9172 10 0
1.5407 0 0
* stimulated with DEN-3 antigen
Dengue-2 antigen optimization by ELISPOT and IC-FC. Pools of PBMC
from DEN-2 infected and uninfected pigtail macaques were used to optimize DEN-
2 antigen by ELISPOT and IC-FC. The assays were run twice side by side.
ELISPOT results show the highest response at the concentration of 20
µg/ml (Fig 9a). Meanwhile, IC-FC show the least concentration of DEN-2 antigen
(5µg/ml) gave the best CD4+ and CD4- T lymphocyte responses (Fig 9b and 9c).

42
(a)
The higher the concentration of antigen, the less CD4+ and CD4- cells responded.
And high antigen concentration generated lots of CD4- T cell response in PBMC
from uninfected pigtail. For IC-FC, 5µg/ml antigen generated the optimum DEN
specific IFN-γ production responses of CD4+ and CD4- T cells.
Figure 9 Dengue-2 antigen optimization by ELISPOT and IC-FC. (a) ELISPOT,
(b) IFN-γ producing CD4+ T lymphocytes by IC-FC, (c) IFN- γ
producing CD4- T lymphocytes by IC-FC. The bars represent mean and
standard deviation.
ELISPOT and IC-FC assays to measure DEN-2 specific T cells after
primary and secondary DEN-2 infections. A comparison of ELISPOT and IC-FC
assays for the detection of DEN antigen specific cells was undertaken using pooled
PMBC from three DEN-2 infected and three uninfected pigtail macaques. Pigtail
macaques were re-injected with DEN-2 virus six months after the first injection. As
shown in Fig 10, DEN specific IFN-γ producing cells as detected by ELISPOT
increased as the response of primary infection. The quantity of these cells did not
(b) (c)
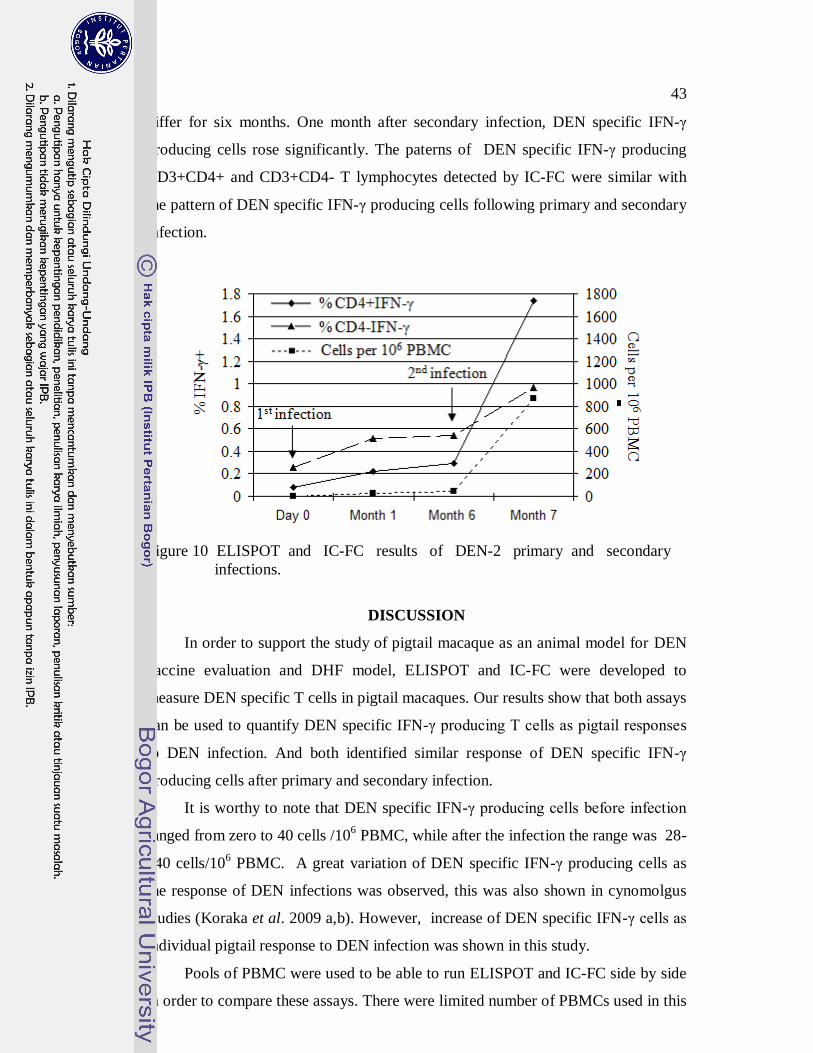
43
differ for six months. One month after secondary infection, DEN specific IFN-γ
producing cells rose significantly. The paterns of DEN specific IFN-γ producing
CD3+CD4+ and CD3+CD4- T lymphocytes detected by IC-FC were similar with
the pattern of DEN specific IFN-γ producing cells following primary and secondary
infection.
Figure 10 ELISPOT and IC-FC results of DEN-2 primary and secondary
infections.
DISCUSSION
In order to support the study of pigtail macaque as an animal model for DEN
vaccine evaluation and DHF model, ELISPOT and IC-FC were developed to
measure DEN specific T cells in pigtail macaques. Our results show that both assays
can be used to quantify DEN specific IFN-γ producing T cells as pigtail responses
to DEN infection. And both identified similar response of DEN specific IFN-γ
producing cells after primary and secondary infection.
It is worthy to note that DEN specific IFN-γ producing cells before infection
ranged from zero to 40 cells /106 PBMC, while after the infection the range was 28-
440 cells/106 PBMC. A great variation of DEN specific IFN-γ producing cells as
the response of DEN infections was observed, this was also shown in cynomolgus
studies (Koraka et al. 2009 a,b). However, increase of DEN specific IFN-γ cells as
individual pigtail response to DEN infection was shown in this study.
Pools of PBMC were used to be able to run ELISPOT and IC-FC side by side
in order to compare these assays. There were limited number of PBMCs used in this

44
study, since the samples that collected during previous DEN studies had been
planned for ELISPOT assay only. The optimation of DEN-2 antigen was re-run to
confirm the highest concentration of antigen (20µg/ml) gave the the optimum
induction in ELISPOT and the lowest antigen concentration (5µg/ml) was optimum
in IC-FC. The nature reaction may have influence to the requirement of optimum
antigen concentration in each assay.
Unlike other studies that used APC (Koraka et al. 2007a,b) or inactivated
dengue lysate antigens (Mangada et al. 2004) for in vitro stimulation, this study
used DEN antigens. It was because during our preliminary study that compared
several antigens to stimulate PBMC from human naturaly infected DEN revealed
that some antigens including DEN inactivated lysate antigens stimulated monocytes
to produce high interleukin-10 that seemed to hold back the functions of CD4+ T
cells (Widjaja et al. 2007). In this study, DEN-2 antigen could induce DEN specific
IFN-γ producing CD3+CD4+ T cells after primary and secondary infections.
Interestingly, antigen titration results and the results of primary and secondary
infections show DEN specific IFN-γ producing CD3+CD4- T cells, most probably
CD3CD8 cells, were more than DEN specific IFN-γ CD3CD4 T cells. However,
whether DEN antigen can induce CD3CD8 to produce IFN-γ could not be shown
clearly in this study, since anti-CD8 antibody was not applied to prove the evidence
of DEN specific IFN-γ producing CD3+CD8+ cells. On the contrary, Mori et al.
(1997) who used double immunocytochemistry to stain proliferated human cells
after DEN lysate antigen stimulation reported just opposite results where only a
few of CD3+CD8+ cells produced IFN-γ (10%) compared with CD3+CD4+ cells
(50-70%). It is interesting to know whether this different is due to the nature of
antigen or the response of species to DEN infection. Commonly, peptides were
applied to measure functional CD3+CD8+ cells specific to DEN (Bashyam et al.
2006, Wen et al. 2010). Simultaneous stimulation for CD4+ and CD8+ would
increase the efficiency of the assays, because of the reduction of PBMC required
and time to run the assays. Other benefits of the application of antigen include the
potential to measure responses from all epitopes in DEN proteins to see the
response of DEN specific IFN-γ cells, and prior understanding of antigenic peptides
is not necessary.

45
Here we report the application of ELISPOT and IC-FC to quantify DEN
specific IFN- γ producing T lymphocytes in pigtail macaques. Application of these
techniques would enhance the study of pigtail macaque as animal model for DEN
studies. Further study is needed to understand whether these two assays may
complement one another or be considered as alternative.
ACKNOWLEDGMENT
The authors thank Dr. Herman Kosasih for valuable advice, Deni Pepi
Butarbutar and Sri Hadiwijaya for technical assistance. This work was supported by
the Naval Medical Research Center work unit 61102A.S13.S.S1415. Authors (SW,
DP, VAS, ILW, GTB, THB, KRP, CGB, MW, PJB) as employees of the U.S.
Government or military service members, conducted the work as part of their
official duties. Title 17 U.S.C. §105 provides that ‘Copyright protection under this
title is not available for any work of the United States Government.’ Title 17
U.S.C. §101 defines a US Government work as a work prepared by a military
service member or employee of the US Government as part of that person’s official
duties.
REFERENCES
Bashyam HS, Green S, Rothman AL. 2006. Dengue virus-reactive CD8-T cells
display quantitative and qualitative differences in their response to variant
epitopes of heterologous viral serotypes. J Immunol 176:2817-2824
Beckett CG, Kosasih H, Faisal I, Nurhayati, Tan R, Widjaja S, Listiyaningsih E,
Ma'roef C, Wuryadi S, Bangs MJ, Samsi TK, Yuwono D, Hayes CG, Porter KR.
2005. Early detection of dengue infections using cluster sampling around index
cases. Am J Trop Med Hyg. 72(6):777-82
Bente DA, Rico-Hesse R. 2006. Models of dengue virus infection. Drug Discov
Today Dis 3(1):97-103.
Brennan G, Kozyrev Y, Kodam T, Hu S-L. 2007. Novel TRIM 5 isoforms
expressed by Macaca nemestrina. J Virol. 81:12210-12217.
Favre D, Lederer S, Kanwar B, Ma Z-M, Proll S, Kasakow Z, Mold J, Swainson L,
Barbour JD, Baskin CR, Palermo R, Pandrea I, Miller CJ, Katze MG, McCune
JM. 2009. Critical loss of the balance between Th17 and T regulatory cell
populations in pathogenic SIV infection. PLOS Pathog 5:1-17.

46
Gauduin M-C, Kaur A, Ahmad S, Yilma T, Lifson JD, Johnson RP. 2004.
Optimization of intracellular cytokine staining for the quantitation of antigen-
specific CD4+ T cell responses in rhesus macaques. J Immunol Methods 288:61-
79.
Koraka P, Benton S, Amerongen G, Stittelaar KJ, Osterhaus ADME. 2007a.
Characterization of humoral and cellular immune responses in cynomolgus
macaques upon primary and subsequent heterologous infections with dengue
virus. Microbes Infect. 9:940-6.
Koraka P, Benton S, Amerongen G, Stittelaar KJ, Osterhaus ADME. 2007b.
Efficacy of a live attenuated tetravalent candidate dengue vaccine in naive and
previously infected cynomolgus macaques. Vaccine. 25:5409-16.
Letch A, Scheibenbogen C. 2003. Quantification and characterization of specific T-
cells by antigen-specific cytokine production using ELISPOT assay ot
intrasellular cytokine staining. J Methods. 31:143-9.
Mathew A, Rothman A. 2008. Understanding the contribution of cellular immunity
to dengue diseases pathogenesis. Immunol Rev 255: 300-13
Mangada MM, Ennis FA, Rothman AL. 2004. Quantitation of dengue virus specific
CD4+ T cells by intracellular cytokine staining. J Immunol Methods. 284(1-
2):89-97.
Mori M, Kurane I, Janus J, Ennis FA. 1997. Cytokine production by dengue virus
antigen-responsive human T lymphocytes in vitro examined using a double
immunocytochemical technique. J Leukoc Biol. 61:338-45.
National Research Council. 1996. The guide for the care and use of laboratory
animals. Washington DC: National Academic Press.
Otten RA, Brown BG, Simon M, Lupo LD, Parekh BS, Lairmore MD, Schable CA,
Schochetman G, Rayfield MA. 1994. Differential replication and pathogenic
effects of HIV-1 and HIV-2 in Macaca nemestrina. AIDS 8:297-306.
Pahar B, Li J, Rourke T, Miller CJ, McCehesney MH. 2003. Detection of antigen-
specific interferon γ expression by ELISPOT and cytokine flow cytometry assays
in rhesus macaques. J Immunol Methods. 283:103-15.
Porter KR, Beckett CG, Kosasih H, Tan RI, Alisjahbana B, Rudiman PI, Widjaja S,
Listiyaningsih E, Ma'Roef CN, McArdle JL, Parwati I, Sudjana P, Jusuf H,
Yuwono D, Wuryadi S. 2005. Epidemiology of dengue and dengue hemorrhagic

47
fever in a cohort of adults living in Bandung, West Java, Indonesia. Am J Trop
Med Hyg 72(1):60-6
Raviprakash K, Defang G, Burgess T, Porter K. 2009. Advances in dengue vaccine
development. Hum Vaccin 5(8):520-8
Thomas SJ, Hombach J, Barrett A. 2009. Scientific consultation on cell mediated
immunity (CMI) in dengue and dengue vaccine development. Vaccine
27(3):355-68.
Widjaja S, Purwaganda D, Hadiwijaya SH, Burgess TH, Blair PJ, Novriani H, Brice
GT. 2007. Characterization of antigen-specific T-cell responses following natural
exposure: Evaluation of different lysate preparations and cytokine assays.
Abstract. Third Asian Regional Dengue Research Network Meeting. Taipei,
Taiwan.
Widjaja S, Winoto I, Sturgiss J, Maroef CN, Listiyaningsih E, Tan R, Pamungkas J,
Iskandrati D, Blair PJ, Sajuthi D, Porter KR. 2010. Macaca nemestrina and
dengue virus infectivity: a potential model for evaluating dengue vaccine
candidates. J Microbiol Ind. 4:58-63.
Wen J, Duan Z, Jiang L. 2010. Identification of a dengue virus-specific HLA-
A*0201-restricted CD8+ T cell epitope. J Med Virol. 82:642-8.
World Health Organization. 2005. Dengue, Dengue hemorrhagic fever and dengue
shock syndrome in the context of the Integrated Management of Childhood
Illness.
http://www.who.int/child_adolescent_health/documents/fch_cah_05_13/en/index
.html [21 May 2010]

GENERAL DISCUSSION
Of the eighteen species of NHP have been studied as models for dengue
infections, several species are endangered, therefore, they are not available for
biomedical research, while others do not support DEN replication, leaving rhesus
and cynomolgus as the common NHP used for vaccine evaluation (Bente and Rico-
Hesse 2006, Raviprakash et al. 2009). Taking notes that DEN vaccine and antiviral
development are still a long-term process and DHF model is still very limited, the
availability of another species as a model strengthens DEN research in animal
model. Concerning host genetic factors, the use of different macaque species or
different origins, may be considered as an important issue to mimic the variability
found in human populations. Here, we explored another species of NHP, pigtail
macaque for its susceptibility to DEN infection and its immune responses.
Pigtail macaques supported the replication of all four DEN serotypes in vivo
by showing detectable viremia after DEN virus injection. Since there is no
standardized methods for DEN virus titration and detection, comparisons between
different macaque species regarding their ability to support viremia following live
dengue virus infection may be inappropriate. However, the same viral detection
methods were used to study viremia in human and similar results were found. The
length of viremia in pigtail and human were 4.8±1.4 days (3 to 8 days) vs 5±2 days
(1 to 8 days) and 6.2±1.7 days (4 to 8 days) vs 7±3 days (1 to 13 days) and by
C6/36 isolation and RT PCR, respectively (US NAMRU-2 unpublished data).
Freire et al. 2007 reported that several strains of DEN viruses failed to
produce viremia in rhesus. Furthermore, a remarkable finding in the development of
DHF animal model was obtained by injecting intravenously high titer of DEN-2
virus in naive Indian rhesus macaques, and resulting in hemorrhagic manifestations
(Onlamoon et al. 2010). The current study utilized only a single wild type DEN
viral isolate and a single dosage for each serotype. Further study in pigtail is
necessary to clarify whether the species of NHP, route of injection, virus titer,
serotype or strain of DEN virus that affects the pathogenesis of DHF.
Also similar with other NHPs, pigtail macaques did not show any clinical
sign. It is worth to note that the animals were observed under anaethesia, a condition
that may mask some clinical signs such as pulse, respiration and body temperature.
(b)

49
As leucopenia, thrombocytopenia and increasing hematocrit are used as supportive
evidence in DF/DHF diagnosis, I would suggest to apply these parameters as the
alternate of clinical signs in the future study of pigtail macaques as DEN animal
model.
In this study, primary and homologous secondary infections did not show any
clinical symptoms. However, in predominantly heterologous secondary infections in
human, the ratio of symptomatic and asymptomatic DEN infections were 1:5 (US
NAMRU-2 unpublished data). As such, it is interesting to know whether
heterologous secondary DEN infections in pigtail macaques would show any
clinical manifestations.
It is noted that predominant IgG subclass response in pigtail is IgG1. While
IgG1 and IgG3, which have strong binding to FcR on the surface of the
macrophages, are predominant subclass IgG in human (Koraka et al. 2001).
Futhermore, significant elevated IgG1 at early infection could be a risk marker for
the manifestations of hemorrhage and shock, while elevated IgG4 could be a risk
marker for shock (Koraka et al. 2001). Whether these findings can be applied in the
development of DHF or DSS pigtail model is worth to investigate.
The ELISPOT and IC-FC can be applied to measure DEN specific IFN-γ
producing T lymphocytes in pigtail macaques. The total PBMC tested by the
ELISPOT and IC-FC simultaneously were limited, therefore, T lymphocyte
responses to primary and secondary DEN infections could not be observed
individually. Furthermore, instead of DEN specific IFN- γ producing CD3+CD4+
T cells, DEN specific CD3+CD4- T cells could also be measured. Whether
CD3+CD4- cells that are CD3CD8 T cells can also be measured by the application
of DEN antigen should be further studied, since simultaneously measurement of
CD3+CD4+ T cells (T helper1) and Tc will enhance the capability to investigate T
lymphocyte functions and to study pigtail macaque as DEN animal model.
Nevertheless, as IFN-γ is not the only cytokine produced by T lymphocytes and
each cytokine has its function, the investigation for each of these cytokines will
give a better prediction of T cell roles. The measurements of DEN specific T cells
should be developed to quantify other functional T cells. The limitation of PBMC
used in this study also limited the observation of T cell serotype cross-reactivity.
Therefore, the total blood volume collected should be calculated by the needs to run

50
simultaneous assays for sample from each individual animal, to study several
cytokines and to observe T cell cross-reactivity. Even it should be taken less than
1% the total of body weigth, the body weight of pigtail would support the
requirements of blood for these purposes.

CONCLUDING REMARKS
Pigtail macaques have showns evidence to serve as a novel NHP model for
DEN vaccine and antiviral evaluations, since their susceptibility to all four
serotypes of DEN virus results in three to eight days consistent viremia in one or
two days post-infections. This finding is similar with viremia in human. Their
antibody responses are also similar with human responses. The results also show
that primary infections induce sufficient immunity to protect the animals from
secondary homologous infection. The route of injection, virus titer, strain or
serotype of DEN virus and heterologous secondary infection should be studied to
understand the possibility of pigtail macaques as DHF model.
ELISPOT and IC-FC can be used to quantify DEN specific IFN-γ producing
T lymphocytes in pigtail macaques after in vitro stimulation of PBMC by DEN ag.
The results show that primary infections increase at least twice number of DEN
specific IFN-γ producing cells and secondary infections augment the number of
cells. Further study is needed to clarify whether CD8 T cells (Tc) can be measured
by DEN antigen application, and to understand whether ELISPOT and IC-FC may
complement one another or be considered as alternative. The development of
ELISPOT and IC-FC to detect other cytokine producing T lymphocytes should also
be considered. The measurements of DEN specific IFN-γ producing T lymphocyte
will enhace the understanding of T lymphocyte functions in the study of pigtail
macaque as DEN animal model. An integrated investigation of DEN specific
antibody and T cells in pigtails may reveal a better understanding of adaptive
immunity functions in the protection and pathogenesis of DHF.
.

REFERENCES
Ajariyakhajorn C, Mammen MP, Endy TP, Gettayacamin M, Nisalak A,
Nimmannitya S, D. H. Libraty DH. Randomized, placebo-controlled trial of
nonpegylated and pegylated forms of recombinant human alpha interferon 2a
for suppression of dengue virus viremia in rhesus monkeys. Antimicrobial
Agents and Chemotherapy. 49(11): 4508–4514 Vol. 49, No. 11
Baribaud F, Pohlmann S, Sparwasser T, Kimata MT, Choi YK, Haggarty BS,
Ahmad N, Macfarlan T, Edwards TG, Leslie GJ, Arnason J, Reinhart TA,
Kimata JT, Littman DR, Hoxie JA, Doms RW. 2001. Functional and antigenic
characterization of human, rhesus macaque, pigtailed macaque and murine DC-
SIGN. J Virol 75:10281-9.
Beckett CG, Kosasih H, Faisal I, Nurhayati, Tan R, Widjaja S, Listiyaningsih E,
Ma'roef C, Wuryadi S, Bangs MJ, Samsi TK, Yuwono D, Hayes CG, Porter
KR. 2005. Early detection of dengue infections using cluster sampling around
index cases. Am J Trop Med Hyg. 72(6):777-82
Bente DA, Rico-Hesse R. 2006. Models of dengue virus infection. Drug Discov
Today 3:97-103.
Brennan G, Kozyrev Y, Kodama T, Hu S. 2007. Novel TRIM5 isoforms expressed
by Macaca nemestrina. J Virol 22:12210-7.
Calisher H, Gould EA. 2003. Taxonomy of the virus family Flaviviridae. In:
Chamber TJ, Monath TP, eds. Advance in Virus Research. Volume 59. San
Diego: Academic Pr. p 1-17.
Chaturvedi UC, Shivastava R, Nagar R. 2005. Dengue vaccines: Problem &
prospects, Indian J Med Res. 121:639-652.
Clyde K, Kyle JL, Harris E. 2006. Recent advances in deciphering viral and host
determinants of dengue virus replication and pathogenesis. J Virol 23
(80):11418–31.
Dit Jen P2MPL. 2008. Laporan tahunan demam berdarah dengue di Indonesia.
Fink J, Gu F, Vasudevan SG. 2006. Role of T cells, cytokines and antibody in
dengue fever and dengue haemorrhagic fever. Rev Med Virol 16: 263–275.

53
Halstead SB. 2003. Neutralization and antibody-dependent enhancement of dengue
viruses. In: Chamber TJ, Monath TP, eds. Advance in Virus Research.
Volume 60. San Diego: Academic Pr. p 421-67.
Halstead SB, Rojanasuphot S, Sangkawibha N. 1983. Original antigenic sin in
dengue. Am J Trop Med Hyg 32:154-6.
Hickling J. 1998. Measuring T-lymphocyte functions. Expert Rev Mol Med.
http://www-ermm.cbcu.cam.ac.uk/ [27 January 2005]
Irie K, Mohan PM, Sasagouri Y, Putnak R, Padmanabhan R. 1989. Sequence
analysis of cloned dengue virus type 2 genome (New Guiena-C strain). Gene
75:197-211.
Janeway CA, Travers P, Walport M, Shlomchik.2001. Immunobiology. New York:
Garland Publishing.
Koraka P, Suharti C, Setiati E, MAiruhu TA, Van Gorp, Hack CE, Juffrie M,
Sutaryo J, Van Der Meer GM, Groen J, Oesterhaus ADME. 2001. Kinetic of
dengue virus-specific serum immunoglobulin classes and subclasses correlate
with clinical outcome of infection. J Clin Microbiol 39: 4332-8.
Lindenbach BD, Rice CM. 2001. Flaviviridae: The viruses ans their replication. In:
Fields BN, Howley PM, eds. Fields’ Virology. 4th
edition. Philadelphia:
Lippincott & Raven. p. 991-1125.
Lindenbach BD, Rice CM. 2003. Molecular biology of Flavivirus. In: Chamber TJ,
Monath TP, eds. Advance in Virus Research. Volume 59. San Diego:
Academic Pr. p 177-223.
Littaua R, Kurane I, Ennis FA. 1990. Human IgG Fc receptor II mediates antibody-
dependent enhancement of dengue virus infection. J Immunol 144:3183–3186.
Mathew A, Rothman AL. 2008. Understanding the contribution of cellular
immunity to dengue disease pathogenesis. Immunol Rev 225:300-313.
Malinoski FJ, Hasty SE, Ussery MA, Dalrymple JM. 1990. Prophylactic ribavirin
treatment of dengue type 1 infection in rhesus monkeys. Antiviral Research.
13: 139-150.
Noble CG, Chen Y, Dong H, Gu F, Lim SP, Schul W, Wang Q, Shi P. 2010.
Strategies for development of dengue virus inhibitors. Antiviral Research 85:
450–462

54
Onlamoon N, Noisakran S, Hsiao H, Duncan A, Villinger F, Ansari AA, Perng GC.
2010. Dengue virus-induced hemorrhage in nonhuman primate model. Blood
115(9):1823-34.
Raviprakash K, Defang G, Burgess T, Porter K. 2009. Advances in dengue vaccine
development. Hum Vaccin 5(8):520-8
Rothman AL. 2004. Dengue: defining protective versus pathologic immunity. J Clin
Inves 113(7): 946-951.
World Health Organization. 2005. Dengue, Dengue hemorrhagic fever and dengue
shock syndrome in the context of the Integrated Management of Childhood
Illness.
http://www.who.int/child_adolescent_health/documents/fch_cah_05_13/en/ind
ex.html [21 May 2010]

APPENDIX 1
1 List of reagents used for dengue virus isolation and mosquito inoculation
Reagents Manufacturer/ Catalog number
Minimum Essential medium-1018 Sigma/ M1018
Fetal Bovine Serum Gibco/ 26140-079
Penicillin-Streptomycin Gibco/ 15140-148
L-glutamine Sigma Aldrich/G5763
Non essential aminoacid Sigma/ M7145
Hepes Gibco/15630-080
MEM vitamin solution Gibco/11120-052
Monoclonal antibody to DEN-1 Chemicon/MAB8401
Monoclonal antibody to DEN-2 Chemicon/MAB8402
Monoclonal antibody to DEN-3 Chemicon/MAB8403
Monoclonal antibody to DEN-4 Chemicon/MAB8404
Flavivirus hyper-immune ascitic fluid In-house
Alphavirus hyperimmune ascitic fluid in-house
Anti-mouse IgG FITC Kirkegaard & Perry Laboratory Inc./ 021806
Aceton Merck/ K38024114
Trypan Blue Solution (0.4%) Sigma Aldrich / T-154
NaCl Sigma Aldrich / S 3014
KH2PO4 Sigma Aldrich / P 5655
Na2HPO4 Sigma Aldrich / S – 9763
Evans Blue Sigma Aldrich / E2129
Glycerol Sigma Aldrich / 5516
2 List of reagents used for dengue RT PCR
Reagents Manufacturer/ Catalog number
Access Reverse Transcription system Promega/A12601
GeneAmp PCR Core Reagents Applied Biosystems/4312765
10x TBE Promega/V4251
Agarose Promega/V3841
Ethidium bromide Sigma Aldrich/E-7637
100 bp DNA ladder Roche/11721933001
Gel loading solution Promega/G1881

Primers and Sequences
Primer Sequence
Genome
position Size (in bp)
D1 5'-TCAATATGCTGAAACGCGCGAGAAACCG-3' 134-161
D2 5'-TTGCACCAACAGTCAATGTCTTCAGGTTC-3' 616-644 511 (D1 and D2)
TS1 5'-CGTCTCAGTGATCCGGGGG-3' 568-586 482 (D1 and TS1)
TS2 5'-CGCCACAAGGGCCATGAACAG-3' 232-252 119 (D1 and TS2)
TS3 5'-TAACATCATCATGAGACAGAGC-3' 400-421 290 (D1 and TS3)
TS4 5'-CTCTGTTGTCTTAAACAAGAGA-3' 506-527 392 (D1 and TS4)
3 List of reagents used for IgM, IgG, avidity IgG and subclases IgG ELISA
Reagents Manufacturer/ Catalog number
Dengue fever ELISA IgM Focus Technology/EL1500M
Dengue fever ELISA IgG Focus Technology/EL1500G
Phosphate buffered saline 0.01M, pH 7.4 Sigma Aldrich / 1000-3
Tween-20 Sigma Aldrich / P-1379
Skim milk powder Carnation
ABTS substrate system Kirkegaard & Perry Laboratory Inc./ 506200
Mouse anti-human IgG (H&L) HRP Accurate/JMH035098
Urea Sigma Aldrich/U5378
Sheep anti human IgG1 peroxidase Binding Site/AP006
Sheep anti human IgG2 peroxidase Binding Site/AP007
Sheep anti human IgG3 peroxidase Binding Site/AP008
Sheep anti human IgG4 peroxidase Binding Site/AP009
4 List of reagents used for plaque reduction neutralization test
Reagents Manufacturer/ Catalog number
Minimum Essential medium Sigma Aldrich/ M-1018
Fetal Bovine Serum Gibco/ 26140-079
Penicillin-Streptomycin Gibco/ 15140-148
Carboxymethyl cellulose (CMC) Sigma Aldrich/C-4888
Minimum Essential Medium without
phenol Red
SigmaAldrich/M-3024
Sodium Bicarbonate Gibco/11810-025
Naphtol- blue black, dye content 80% Sigma Aldrich/19524-3
Sodium Acetate, anhydrous Sigma Aldrich/S-5636
EDTA Sigma Aldrich/E-5134

Tricine Sigma Aldrich/T-5816
Trypsin Gibco/27250-042
Acetic acid glacial Sigma Aldrich/A-0808

APPENDIX 2
1 RT PCR Cycle Condition:
48°C 45 min
94°C 2 min
94°C 30 sec
60°C 1 min 40 cycles
68°C 2 min
68°C 7 min
4°C ∞
2 Nested PCR Cycle condition:
94°C 2 min
94°C 1 min
55°C 1 min 20 cycles
72°C 2 min
72°C 10 min
4°C ∞

APPENDIX 3
1 List of reagents used for ELISPOT and intracellular cytokine staining
Reagents Manufacturer/ Catalogue number
RPMI 1640 Gibco/72400120
Fetal Bovine Serum Gibco/ 26140079
Penicillin-Streptomycin Gibco/ 15140-148
L-glutamine Sigma Aldrich/G5763
Benzonase Merck/71205-3
PMA Sigma Aldrich/P1585
Ionomycin Sigma Aldrich/I0634
Monkey ELISPOT IFN gamma Mabtech/3420M-2H
Phosphate buffered saline 0.01M, pH 7.4 Sigma Aldrich / 1000-3
Tween-20 Sigma Aldrich / P-1379
Dulbecco's PBS Gibco/21600069
Golgi Plug BD Biosciences/555029
Monoclonal antibody anti CD4 PerCP BD Biosciences/550631
Monoclonal antibody anti CD3 APC Cy7 BD Biosciences/557757
Monoclonal antibody anti IFN-γ BD Biosciences/554702
Cytofix/Cytoperm BD Biosciences/554714
Perm/wash 10x BD Biosciences/554723