ventromedial hypothalamic lesions produced by gold thioglucose do not impair induction of npy mrna...
TRANSCRIPT

BRAIN RESEARCH
ELSEVIER Brain Research 707 (1996) 266-271
Research report
Ventromedial hypothalamic lesions produced by gold thioglucose do not impair induction of NPY mRNA in the arcuate nucleus by fasting
Hugo T. Bergen ~'*, Charles V. Mobbs b
Dept. of Anatomy, Unil~ersity of Manitoba, 730 William At,e., Winnipeg, Man., Canada R3E OW3 Fishberg Center in Neurobiology, Mt. Sinai School of Medicine, One Gustaee Let,y Place, New York, NY 10029, USA
Accepted 28 September 1995
Abstract
There is increasing evidence that neuropeptide Y (NPY) plays an important role in the regulation of food intake. Neuropeptide Y mRNA in the arcuate nucleus increases after fasting and it has been proposed that this increase in NPY activity occurs as a result of the decreased circulating levels of both insulin and glucose associated with a fast. Glucose-responsive neurons in the ventromedial nucleus (VMN) of the hypothalamus alter their activity in response to changes in circulating glucose levels and these neurons have been proposed to be involved in the regulation of feeding behavior and metabolism. However, it is not known if these glucose-responsive neurons are involved in the response of NPY mRNA in the arcuate nucleus to fasting. To address this relationship, mice were injected with either saline or gold thioglucose (GTG), which appears to act on glucose-responsive neurons, and killed 6 weeks later after a 72 h fast or under ad lib fed conditions. In situ hybridization histochemistry for NPY mRNA was performed on hypothalamic sections containing the arcuate nucleus. The number of labelled cells was counted and the density of autoradiographic silver grains overlying the cells was also quantified (i.e. pixels per cell). Fasting resulted in increased levels of total NPY mRNA (number of labelled cells multiplied by the pixels per cell) in the arcuate nucleus of both control and GTG-treated mice. In addition, the relative fasting-induced increase (i.e. the fasted to fed ratio) in number of cells detected, number of pixels per cell, and total NPY mRNA was similar in both the control and GTG-treated mice. These data suggest that GTG-sensitive VMN neurons play little role in the induction of NPY mRNA by fasting in the arcuate nucleus.
Keywords: Neuropeptide Y; mRNA; Arcuate nucleus; Gold thioglucose; Ventromedial nucleus; Food deprivation
1. Introduct ion
Although both the ventromedial nucleus (VMN) and the arcuate nucleus of the hypothalamus are implicated in the regulation of body weight and feeding behavior, how neurons in these nuclei might interact to regulate body weight is not clear. Neurons that are responsive to glucose are found in or near the VMN [15,23-25]. These glucose- responsive neurons, which are also sensitive to insulin, appear to be particularly relevant to feeding behavior [15] and, presumably, glucose homeostasis. In mice, a single intraperitoneal (ip) injection of gold-thioglucose (GTG) causes long-term impairments in glucose homeostasis simi- lar to those produced by a VMN lesion ([2,3,12]; Bergen and Mobbs, submitted). At low doses, GTG is thought to act (at least initially) primarily on glucoreceptor cells in the VMN since (1) related gold compounds that lack
* Corresponding author. Fax: (1) (204) 772-1)622.
0006-8993/96/$15.00 © 1996 Elsevier Science B.V. All rights reserved SSDI 0 0 0 6 - 8 9 9 3 ( 9 5 ) 0 1 270-2
glucose do not induce VMN lesions [21], (2) inhibitors of glucose transport (e.g. phlorizin) prevent GTG-induced lesions in the VMN [8,13], and (3) GTG does not induce VMN lesions in the absence of insulin [11,12]. The pres- ence of glucoreceptors in the VMN is also supported by the recent finding that destruction of cell bodies in the VMN greatly attenuates the sympathetic response to hypo- glyemia [4]. Furthermore, this counterregulatory response to hypoglycemia is mimicked by the delivery of 2-de- oxyglucose (2-DG; which effectively produces the same intracellular changes as hypoglycemia) into the VMN [5]. These studies suggest that neurons in the VMN play a role in the response to hypoglycemia. These results may be of additional significance since hypoglycemia also appears to be a factor in the initiation of food intake. It has been shown that spontaneous feeding is preceded by a decline in blood glucose levels and that an experimentally induced drop in blood glucose levels can trigger the onset of a meal [9,18,29]. In addition, it has been shown that intracer- broventricular infusion glucose inhibits, and 2-DG stimu-

H.T. Bergen, C.V. Mobbs / Brain Research 707 (1996) 266-271 267
lates, food irttake in rats [16,30]. Interestingly, the inhibi- tion of feeding by glucose and the stimulation of feeding by 2-DG is impaired in Zucker rats, which, like VMN-le- sioned rats, are obese [30]. Similarly, we have preliminary evidence that under some circumstances, the induction of feeding behavior by 2-DG is impaired in mice treated with GTG (unpublished data). Taken together, these data sug- gest that neurons in the VMN may mediate not only effects of glucose on food intake, but may also play a role in mediating some effects of fasting.
There is increasing evidence that neurons in the arcuate nucleus which synthesize NPY and project to the paraven- tricular nucleus also play an important role in the regula- tion of food intake. For example, NPY peptide secretion in the paraventricular nucleus increases in association with increased appetite [14]. Similarly, NPY mRNA in the arcuate nucleus increases in fasting animals [10,19]. The increase of NPY during fasting is probably due both to the fall in insulin and the fall in glucose which occurs in fasted animals. Fasted animals injected with insulin exhibit an attenuated elevation of hypothalamic NPY mRNA [19,28]. Since administration of insulin only partially suppressed the increase in NPY, and there was a significant inverse correlation between plasma glucose and NPY, it was sug- gested that plasma glucose also inhibits hypothalamic NPY, and that the decline in plasma glucose levels during fasting may play a role in the fasting-induced increase in hypotha- lamic NPY levels [19]. Similarly, insulin deficiency due to streptozotocin treatment causes an increase in hypothala- mic NPY which can be reversed by insulin [22]. However, reduction of plasma glucose by fasting in streptozotocin- treated rats also causes an increase in hypothalamic NPY with no change in insulin levels, leading the authors to suggest that with regard to the effects of insulin to inhibit hypothalamic NPY mRNA, "insulin acts by facilitating glucose entry to sensitive (glucostat) neurons" [22]. A role for glucose metabolism in the regulation of hypothalamic NPY is also supported by studies demonstrating that ad- ministration of 2-DG, which blocks glucose utilization, increases NPY in the arcuate nucleus without altering insulin levels [1]. Taken together, these data suggest the hypothesis that glucose-responsive neurons in the VMN may mediate some effects of fasting on NPY neurons. A previous study has reported that an ip injection of GTG caused an elevation in NPY mRNA in the arcuate nucleus of ad lib fed mice [32], but the regulation of NPY by fasting was not examined. The present study, which ad- dressed this question, indicated that the regulation of NPY mRNA in the arcuate nucleus is not impaired by destruc- tion of GTG-sensitive VMN neurons.
2. Materials and methods
Male mice of the CBA strain were singly housed in a temperature- (23°C) and light-controlled room (lights on
from 07.00 h to 19.00 h). Rodent chow and tap water were available ad libitum except where otherwise noted and the mice weighed between 25 and 30 g at the beginning of the experiment. Gold thioglucose (aurothioglucose, Sigma) was dissolved in 0.1 M sodium citrate (250 mg/ml) immedi- ately prior to i.p. injection. The dosage used (0.5 m g / g body weight) was comparable to those previously reported to produce hypothalamic lesions and increase body weight [171.
Two weeks after they arrived the mice were weighed and then injected i.p. with either GTG or vehicle between 09.00 h and 10.00 h. The mice were weighed at weekly intervals and killed 7 weeks after GTG injection. In both control and GTG-treated groups, the mice were divided into 2 groups and either fasted for 72 h, or fed ad libitum, until the morning of the day of sacrifice. Mice were killed by CO 2 asphyxiation, decapitated, and their brains were rapidly removed and frozen with powdered dry ice. The brains were kept frozen at -70°C until sectioned on a cryostat.
In a follow up experiment male mice of the CBA strain were injected as above with either GTG or vehicle (Day 0). This experiment was performed to determine whether NPY mRNA levels are elevated in GTG-treated mice at a time point when they are hyperphagic relative to control mice. The mice were housed in suspended wire cages in which rodent chow and water were available ad libitum. Beginning with day 0, food intake was measured on a daily basis by weighing the food hopper and collecting the chow that had fallen through the cage floor. The mice were killed, as described above, during the afternoon of Day 12. The brains were frozen with dry ice and kept frozen until sectioned on a cryostat.
In situ hybridization histochemistry for NPY mRNA was performed on hypothalamic sections according to a well established protocol. The brains were cut (10 /zm) in a cryostat and the hypothalamic coronal sections were thaw-mounted onto subbed slides. The sections were fixed in 4% paraformaldehyde and dehydrated in alcohols and stored frozen ( -20°C) until in situ hybridization was performed. Every 20th section through the hypothalamus was stained with Cresyl violet and coverslipped with per- mount after being fixed with paraformaldehyde. These sections were later used for the matching of sections from the different mice. The probe was a single-stranded DNA probe complementary to NPY mRNA. The radiolabelled probe was synthesized using asymmetric polymerase chain reaction and tritiated nucleotides [7] (plasmid kindly sup- plied by S. Sabol). Matched sections of the arcuate nucleus were brought to room temperature and prehybridization buffer was applied to the sections. After 3 h the prehy- bridization buffer was removed and hybridization buffer containing 100,000 cpm/20 ml of the tritiated DNA probe was applied to the section. It had been previously demon- strated that this was sufficient probe to saturate the mes- sage in the sections. Alternate sections were hybridized
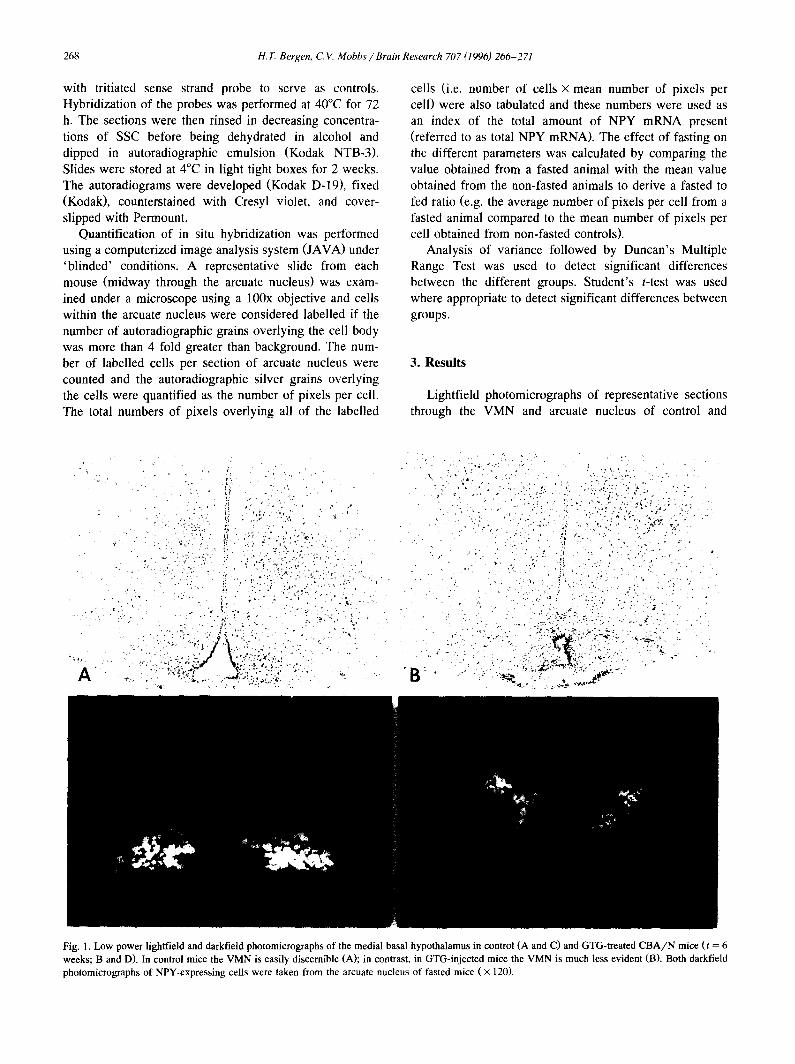
268 H. 1". Bergen, C.V. Mobbs / Brain Research 707 (1996)266-271
with tritiated sense strand probe to serve as controls. Hybridization of the probes was performed at 40°C for 72 h. The sections were then rinsed in decreasing concentra- tions of SSC before being dehydrated in alcohol and dipped in autoradiographic emulsion (Kodak NTB-3). Slides were stored at 4°C in light tight boxes for 2 weeks. The autoradiograms were developed (Kodak D-19), fixed (Kodak), counterstained with Cresyl violet, and cover- slipped with Permount.
Quantification of in situ hybridization was performed using a computerized image analysis system (JAVA) under 'blinded' conditions. A representative slide from each mouse (midway through the arcuate nucleus) was exam- ined under a microscope using a 100x objective and cells within the arcuate nucleus were considered labelled if the number of autoradiographic grains overlying the cell body was more than 4 fold greater than background. The num- ber of labelled cells per section of arcuate nucleus were counted and the autoradiographic silver grains overlying the cells were quantified as the number of pixels per cell. The total numbers of pixels overlying all of the labelled
cells (i.e. number of cells × mean number of pixels per cell) were also tabulated and these numbers were used as an index of the total amount of NPY mRNA present (referred to as total NPY mRNA). The effect of fasting on the different parameters was calculated by comparing the value obtained from a fasted animal with the mean value obtained from the non-fasted animals to derive a fasted to fed ratio (e.g. the average number of pixels per cell from a fasted animal compared to the mean number of pixels per cell obtained from non-fasted controls).
Analysis of variance followed by Duncan's Multiple Range Test was used to detect significant differences between the different groups. Student's t-test was used where appropriate to detect significant differences between groups.
3. Results
Lightfield photomicrographs of representative sections through the VMN and arcuate nucleus of control and
A
i
: . ;,. -., ,.. . .'-.,. ,-
- , . , , . . ~ r
T ~
D
Fig. 1. Low power lightfield and darkfield photomicrographs of the medial basal hypothalamus in control (A and C) and GTG-treated C B A / N mice (t = 6 weeks; B and D). In control mice the VMN is easily discernible (A); in contrast, in GTG-injected mice the VMN is much less evident (B). Both darkfield photomicrographs of NPY-expressing cells were taken from the arcuate nucleus of fasted mice ( × 120).

H.T. Bergen, C. V. Mobbs / Brain Research 707 (1996) 266-271 269
g- o x
o..
z
ioo
8O
60
40 a
0 Control ~IG
Fed
a
T • :~!ii~i}!:i!::ii~:-:i:i!;~ii:;i: b
~":"=~*=="" ; U
Fasted Fig. 2. Effect of GTG on total NPY mRNA levels in the arcuate nucleus of control and GTG-treated mice under fed and fasted conditions. The number of labelled cells in the section was multiplied by the mean number of pixels per cell (in that section) and is directly proportional to the total number of autoradiographic grains overlying all of the labelled cells in a section. Bars with the same letters are significantly different from each other (P < 0.05).
El-
1000 I
750 t
Control GTG
Fed
@
°1/i ',ontrol GTG
Fasted Fig. 4. Effect of GTG on the relative levels of NPY mRNA per labelled cell in the arcuate nucleus, under fasted and fed conditions. The number of pixels per cell is representative of the number of autoradiographic silver grains overlying the cell, and the amount of NPY mRNA in the cell. Bars with the same letters are significantly different from each other (P < 0.05).
GTG-treated mice are shown in Fig. 1 (A and B, respec- tively). Darkfield photomicrographs of representative auto- radiograms from control and GTG-treated mice (both fasted) are also shown in Fig. 1 (C and D, respectively) and these sections are at the same level as the sections shown in the lightfield photomicrographs. As an index of the total amount of NPY mRNA present in a section, the number of labelled cells in the section was multiplied by the mean number of pixels per cell (in that section). This is directly proportional to the total number of autoradio- graphic grains overlying all of the labelled cells in a section. Analysis of variance showed a significant effect of both GTG and fasting on total NPY mRNA (P < 0.05). In addition, fasting resulted in a significant increase in NPY mRNA in both control and GTG-treated mice ( P < 0.05) (Fig. 2).
80 a , b
o 70 ¢Y
o 50
'-' 40
" 30
a
Control GTG Fed
lit-- Control GTG
Fasted Fig. 3. Effect of GTG on the number of cells expressing NPY mRNA in the arcuate nucleus, under fasted and fed conditions. The number repre- sents mean number of labelled cells counted on one side of the brain (one section per mouse; n = 4 and 5 in both of the fasted and fed groups, respectively). Bars with the same letters are significantly different from each other (P < 0.05).
Analysis of variance detected a significant effect of fasting (P < 0.05) and GTG treatment (P < 0.01) on the number of NPY mRNA-containing cells present in the arcuate nucleus. Specifically, fasting increased while GTG treatment decreased the number of labelled cells detected. In addition there were significantly more labelled cells detected in the fasted control mice (n = 4) than in both the fed controls (n = 5) and the fasted GTG-treated mice (n = 4) ( P < 0.05) (Fig. 3). Although cell numbers tended to be higher in the GTG-fasted mice relative to the GTG-fed mice (n = 5), this did not reach the level of statistical significance (P = 0.089).
Analysis of variance also detected a significant effect of fasting on the number of pixels per cell (P < 0.05) and although GTG treatment tended to result in decreased pixels per cell (i.e. number of autoradiographic grains per cell) this did not achieve statistical significance ( P = 0.052). The number of pixels per cell was also greater in fasted controls as compared to fed controls ( P < 0.05) (Fig. 4). The numbers of pixels per cell between fed and fasted GTG-treated mice were not significantly different, however the degree of induction by fasting in the GTG- treated mice was similar to that observed in the control mice (Table 1).
The effect of fasting on the different parameters mea- sured, as determined by the fasted to fed ratio, is shown in
Table 1 The effect of a 72 h fast, as compared to ad lib fed controls, on the different parameters used to quantitate NPY activity in the arcuate nucleus. The numbers represent the relative increase observed in the fasted animals as compared to the ad lib fed animals
Total NPY mRNA Numbers of cells Pixels/Cell
Control 2.45 + 0.57 1.51 + 0.24 1.58 + 0.27 GTG 2.35+0.61 1.52+0.29 1.50+0.15

270 H.T. Bergen, C. V. Mobbs / Brain Research 707 (1996) 266-271
Table 1. The fasted to fed ratios of the different parameters were similar between control and GTG-treated mice. For example, in both GTG-treated mice and controls, fasting resulted in an approximately 2.4 fold increase in the amount of total NPY mRNA. Therefore although GTG decreased the number of NPY mRNA-containing cells detected and the relative of NPY mRNA present in the arcuate nucleus compared to controls (see above), it did not alter the degree to which a fast increases the amount of NPY mRNA in the arcuate nucleus.
Since the first experiment failed to demonstrate an elevation of NPY mRNA after GTG injection, as had been previously reported [32], a follow-up experiment was un- dertaken to assess if an induction of NPY mRNA might be exhibited at an earlier time-point following GTG treat- ment. Mice were killed 12 days after either GTG or vehicle injection. The GTG-treated mice weighed signifi- cantly more than the controls at the time of being killed (27.6 ± 0.8 g vs. 24.2 + 0.4 g, respectively; P < 0.05). During the 48 h period prior to being killed the GTG mice ate significantly more food than the controls (10.9 + 0.7 g vs. 8.4___ 0.2 g, respectively; P < 0.05). The number of labelled cells detected in the arcuate nucleus was similar in the GTG-injected and control mice (35.0 + 8.0 and 34.3 + 8.4 cells, respectively). Similar to what was reported above, the number of pixels per cell tended to be lower in the GTG-injected mice than in controls (517 + 83 vs. 968 + 178, respectively) but this did not reach the level of statistical significance. Total NPY mRNA levels in GTG- injected mice (n = 3) were not significantly different than controls (n = 4) (1.7 + 0.9 × 10 4 pixels and 3.2 + 1.3 × 10 4 pixels, respectively).
4. Discussion
In this study, GTG induced VMN lesions but did not impair the induction of NPY mRNA by fasting. These data suggest that glucose-sensitive, or at least GTG-sensitive VMN neurons do not mediate the effects of fasting on NPY mRNA in the arcuate nucleus. It has also been reported that GTG can lesion cells in the nucleus of the tractus solitarius and the dorsal motor nucleus of the vagus [26]. The presence of lesions in these sites would suggest that these neurons also play little role in the induction of NPY by fasting. The degree of induction of NPY mRNA by fasting (i.e. the fasted to fed ratio of the various parameters) was almost identical in GTG-treated animals and controls. In addition, the magnitude of the increase in arcuate NPY mRNA brought about by fasting in both GTG-treated and control mice (i.e. 2.4 fold), is similar to that reported in other in situ hybridization studies in which rats were fasted 48 to 96 h (1.5 to 3 fold increases) [6,20,27,28,31]. Therefore, it does not appear that the factors that play an important role in the stimulation of
NPY cells during a fast require the integrity of GTG-sensi- tive neurons in the VMN (or possibly elsewhere).
We also failed to observe an increase in NPY mRNA levels after injection with GTG, which is in contrast to a previous report [32]. The reasons for this discrepancy are not clear. One possibility that can be readily discounted is that GTG in our experiment was ineffective in inducing a lesion. Histological analysis revealed that a lesion in the VMN was observed in GTG-treated mice. In addition, GTG-treated mice weighed more than controls, and during the onset of obesity food intake was elevated in GTG- treated mice relative to controls. These findings support the contention that GTG was effective in inducing a VMN lesion. A second possibility is that the methods used in the present study might not be sensitive enough to detect the changes reported in the previous study. However, the effects of fasting demonstrated in the present study were approximately equal in magnitude to the effects of GTG reported in the previous study. Furthermore, it is striking that in both studies reported here, NPY mRNA was actu- ally lower after GTG injection. Considering the confidence interval obtained in the study (although with a relatively limited number of mice), the probability of observing large numbers of GTG-injected mice with much higher levels of NPY mRNA would appear to be small. The use of differ- ent mouse strains, differences in sampling within the arcu- ate nucleus between the studies, and differences in the type of probe used could account for the difference between our results and those reported previously [32]. In any case, whereas the metabolic perturbations that follow GTG-in- duced lesions in the VMN might be associated with an increase in NPY mRNA in the arcuate nucleus under certain circumstances [32], the present study strongly indi- cates that these metabolic impairments do not require a persistent elevation in NPY mRNA. Nevertheless, the pre- sent study does not address the possibility that altered NPY peptide release might mediate some effects of a GTG lesion, without a change in NPY mRNA levels. Conceiv- ably, metabolic effects of the GTG lesion could occur even in the absence of NPY in the arcuate nucleus, such as occurs after neonatal treatment of mice with monosodium glutamate. We are currently assessing this possibility. Taken together, these results suggest that NPY neurons in the arcuate nucleus and glucose-responsive neurons in the VMN may regulate metabolism through largely indepen- dent mechanisms.
Acknowledgements
These studies were supported by the American Diabetes Association (C.V.M.), the American Federation for Aging Research (C.V.M.), the Manitoba Health Research Council (H.T.B.), and the Glenn Foundation for Medical Research, of which CVM was a Fellow.

H.T. Bergen, C.V. Mobbs / Brain Research 707 (1996) 266-271 271
References
[1] Akabayashi, A., Zaia, C.T.B., Siva, I., Chae, H..I. and Leibowitz, S.F., Neuropeptide Y in the arcuate nucleus is modulated by alter- ations in glucose utilization, Brain Res., 621 (1993) 343-348.
[2] Blair, S.C., Caterson, I.D. and Cooney, G.J., Insulin response to an intravenous glucose load during development of obesity in gold thioglucose-injected mice, Diabetes, 42 (1993) 1153-1158.
[3] Blair, S.C., Cooney, G.J., Denyer, G.S., Williams, P.F. and Cater- son, I.D., Differences in lipogenesis in tissues of control and gold- thioglucose obese mice after an isocaloric meal, Biochim. Biophys. Acta, 1085 (1991) 385-388.
[4] Borg, W.P., During, M.J., Sherwin, R.S., Borg, M.A., Brines, M.L. and Shulman, G.I., Ventromedial hypothalamic lesions in rats sup- press counterregulatory responses to hypoglycemia, J. Clin. Invest., 93 (1994) 1677-1682.
[5] Borg, W.P., Sherwin, R.S., During, M.J., Borg, M.A. and Shulman, G.I., Local ventromedial hypothalamus glucopenia triggers counter- regulatory hormone release, Diabetes, 44 (1995) 180-184.
[6] Brady, L.S., Smith, M.A., Gold, P.W. and Herkenham, M., Altered expression of hypothalamic neuropeptide mRNAs in food-restricted and food-deprived rats, Neuroendocrinology, 52 (1990) 441-447.
[7] Brooks, P.J., Kaplitt, M.G., Kleopoulos, S.P., Funabashi, T., Mobbs, C.V. and Pfaff, D.W., Detection of messenger RNA and low-abun- dance heteronuclear RNA with single-stranded DNA probes pro- duced by amplified primer extension labeling, J. Histochem. Cy- tochem., 41 (1993) 1761-1766.
[8] Brown, D.F. and Viles, J.M., Systemic phlorizin prevents gold thioglucose necrosis in the ventromedial hypothalamus, Brain Res. Bull., 8 (1982) 347-351.
[9] Campfield, L.A., Brandon, P. and Smith, F., On-line continuous measurement of blood glucose and meal pattern in free-feeding rats: the role of glucose in meal initiation, Brain Res. Bull,, 14 (1985) 605-616.
[10] Chua, J.S.C., Brown, A.W., Kim, J., Hennessey, K.L., Leibel, R.L. and Hirsch, J., Food deprivation and hypothalamic neuropeptide gene expression: effects of strain background and the diabetes mutation, Mot. Brain Res., 11 (1991) 291-299.
[11] Debons, A.F., Krimsky, I., Likuski, H.J., From, A. and Cloutier, R.J., Gold thioglucose damage to the satiety center: inhibition in diabetes, Am. J. Physiol., 214 (1968) 652-658.
[12] Debons, A.F., Krimsky, I., Maayan, M.L., Fani, K. and Jimenez, F.A., Gold thioglucose obesity syndrome, Fed. Proc., 36 (1977) 143-147.
[13] Debons, A.F., Krimsky, I., From, A. and Pattinian, H., Phlorizin inhibition of hypothalamic necrosis induced by gold thioglucose, Am. J. Physiol., 226 (1974) 574-578.
[14] Kalra, S.P., Dube, M.G., Sahu, A., Phelps, C.P. and Kalra, P.S., Neuropeptide Y secretion increases in the paraventricular nucleus in association with increased appetite for food, Proc. Natl. Acad. Sci., 88 (1991) 10931-10935.
[15] Kow, L.-M. and Pfaff, D.W., Actions of feeding relevant agents on hypothalamic glucose responsive neurons in vitro, Brain Res. Bull,, 15 (1985) 509-513.
[16] Kurata, K., Fujimoto, K., Sakata, T., Etou, H. and Fukagawa, K.,
D-Glucose suppression of eating after intra-third ventricle infusion in rat, Physiol. Behav., 37 (1986) 615-620.
[17] Liebelt, R.A., Sekiba, K., Liebelt, A.G. and Perry, J.H., Genetic susceptibility to goldthioglucose-induced obesity in mice, Proc. Soc. Exp. Biol. Med., 104 (1960) 689-692.
[18] Louis-Sylvestre, J. and LeMagnen, J., A fall in blood glucose level precedes meal onset in free-feeding rats, Neurosci. Biobehav. Rev., 4 (Suppl. 1) (1980) 13-15.
[19] Malabu, U.H., McCarthy, H.D., McKibbin, P.E. and Williams, G., Peripheral insulin administration attenuates the increase in neuropep- tide Y concentrations in the hypothalamic arcuate nucleus of fasted rats, Peptides, 13 (1992) 1097-1102.
[20] Marks, J.L., Li, M., Schwartz, M., Porte, J.D. and Baskin, D.G., Effect of fasting on regional levels of neuropeptide Y mRNA and insulin receptors in the rat hypothalamus, Mol. Cell, Neurosci., 3 (1992) 199-205.
[21] Mayer, J. and Marshall, N.B., Specificity of goldthioglucose for ventromedial hypothalamic lesions and hyperphagia, Nature, 178 (1956) 1399-1400.
[22] McKibbin, P.E., McCarthy, H.D., Shaw, P. and Williams, G., Insulin deficiency is a specific stimulus to neuropeptide Y: a com- parison of the effects of insulin replacement and food restriction in streptozotocin-diabetic rats, Peptides, 13 (1992) 721-727.
[23] Ono, T., Sasaki, K. and Shibata R., Feeding- and chemical-related activity of ventromedial hypothalamic neurones in freely behaving rats, J. Physiol., 394 (1987) 221-237.
[24] Oomura, Y., Glucose as a regulator of neuronal activity, Adv. Metab. Disord., 10 (1983) 31-65.
[25] Oomura, Y., Ono, T., Ooyama, H. and Wayner, M.J., Glucose and osmosensitive neurons of the rat hypothalamus, Nature, 222 (1969) 282-284.
[26] Powley, T.L. and Prechtl, J.C., Gold thioglucose selectively dam- ages dorsal vagal nuclei, Brain Res., 367 (1986) 192-200.
[27] Schwartz, M.W., Sipols, A.J., Grubin, C.E. and Baskin, D.G., Differential effect of fasting on hypothalamic expression of genes encoding neuropeptide Y, galanin, and glutamic acid decarboxylase, Brain Res. Bull,, 31 (1993) 361-367.
[28] Schwartz, M.W., Sipols, A.J., Marks, J.L., Sanacora, G., White, J.D., Schuerink, A., Kahn, S.E., Baskin, D.G., Woods, S.C., Figlewicz, D.P. and Porte, J.D., Inhibition of hypothalamic neu- ropeptide Y expression by insulin, Endocrinology, 130 (1992) 3608-3616.
[29] Smith, F.J. and Campfield, L.A., Meal initiation occurs after experi- mental induction of transient declines in blood glucose, Am. J. Physiol., 265 (1993) R1423-R1429.
[30] Tsujii, S. and Bray, G.A., Effects of glucose, 2-deoxyglucose, phlorizin, and insulin on food intake of lean and fatty rats, Am. J. Physiol., 258 (1990) E476-E481.
[31] White, J.D. and Kershaw, M., Increased hypothalamic neuropeptide Y expression following food deprivation, Mol. Cell, Neurosci., 1 (1989) 41-48.
[32] Young, J.K., McKenzie, J.C., Brady, L.S. and Herkenham, M., Hypothalamic lesions increase neuropeptide Y mRNA in the arcuate nucleus of mice, Neurosci. Lett., 165 (1994) 13-17.