variabilità genetica di mhc di classe ii locus drb in...
TRANSCRIPT

Variabilità genetica di MHC di classe II locus DRB in popolazioni
selvatiche di camoscio alpino (Rupicapra r. rupicapra) e
camoscio pirenaico (Rupicapra p. pyrenaica)
Tesi di Dottorato di Ricerca in Scienze di Sanità Pubblica e Microbiologia
Curriculum Microbiologia e Parassitologia
Dr.ssa Sofia Ingrosso


Tra i petroglifici presenti sulle pareti rocciose
del Parco archeologico presso Vila Nova de
Fez Côa (Pirenei della Catalogna), risalenti a
20 30 mila anni fa, nel Paleolitico Superiore, si
può riconoscere la figura di un capride, forse
un camoscio. Nel profilo ottenuto mediante
una difficile picchiettatura irregolare anziché
con un'incisione lineare, si distinguono il
corpo, le zampe, la testa e le corna appenna
accennate.


Indagare l’imprevedibilità del certo, ridurre ai minimi termini. Il comune denominatore
si chiama conoscenza e la ricerca inizia …
Iago.




Dottorato di Ricerca in Scienze di Sanità Pubblica e Microbiologia
Curriculum Microbiologia e Parassitologia
XXI ciclo AA 2008/2009
Variabilità genetica di MHC di classe II locus DRB in popolazioni selvatiche
di camoscio alpino (Rupicapra r. rupicapra) e camoscio pirenaico
(Rupicapra p. pyrenaica)
Dottorando: SOFIA INGROSSO
Docente Guida Coordinatore
Prof. Stefano D’Amelio Prof. Gianfranco Tarsitani


INDICE
1. INTRODUZIONE 5
1.1 Introduzione generale 5 1.1.1 I markers neutrali 6 1.1.2 I markers dei processi adattativi 7 1.1.3 Studi sulla variabilità genetica delle MHC nei vertebrati 8
1.2 Il Complesso Maggiore di Istocompatibilità 10 1.2.1 Struttura e funzione delle proteine delle MHC 11 1.2.2 L’organizzazione genetica delle MHC 16 1.2.3 Il polimorfismo delle MHC 17 1.2.4 I meccanismi di selezione 19
1.3 Caratteristiche generali del camoscio 21 1.3.1 Origine ed evoluzione del camoscio 24 1.3.2 Distribuzione geografica 26 1.3.3 Biologia del camoscio 29 1.3.4 Una malattia parassitaria: la rogna sarcoptica 32 1.3.5 Breve introduzione alla genetica della conservazione 37 1.3.6 L’importanza della genetica per la conservazione delle popolazioni di camoscio 39
1.4 Obiettivi della ricerca 40
2. MATERIALI E METODI 43
2.1 Campioni analizzati 43
2.2 L’area di studio 49 2.2.1 La Provincia di Belluno 49 2.2.2 La Provincia di Lecco 52 2.2.3 La Spagna pirenaica 52
2.3 Il campionamento 52
2.4 Caratterizzazione molecolare 54 2.4.1 Estrazione ed isolamento del DNA 55
2.4.2 Amplificazione dell’esone 2 delle MHC di classe II 56
2.5 Analisi dei dati 60 2.5.1 Codifica delle sequenze 60 2.5.2 Ricostruzione degli aplotipi con metodi statistici 61

3. RISULTATI 65
3.1 Polimorfismo dell’esone 2 delle MHC di classe locus DRB 65
3.2 Diversità genetica intra-popolazioni 83
3.3 Diversità genetica tra-popolazioni 91
3.4 Alberi di Neighbor-Joining 95
4. DISCUSSIONE DEI DATI E CONSIDERAZIONI 97
4.1 Variabilità genetica delle popolazioni di camoscio 98
4.2 Variabilità nucleotidica delle popolazioni di camoscio 102
5. CONCLUSIONI 105
RIFERIMENTI BIBLIOGRAFICI 107
ALLEGATI 125

5
INTRODUZIONE
1.1 Introduzione generale
Negli ultimi decenni, i geni del complesso maggiore di istocompatibilità (Major
Histocompatibility Complex, MHC) hanno attirato l’interesse dei ricercatori
principalmente spinti dal proposito di comprendere i processi che sostengono il
polimorfismo genetico (Klein 1986; Apanius et al. 1977; Bernatchez & Landry 2003).
Dopo la scoperta dello straordinario numero degli alleli delle MHC nelle popolazioni
naturali, è stato supposto che la selezione naturale è responsabile del loro
mantenimento (Clarke & Kirby 1966). La relazione tra i genotipi delle MHC e
l’insorgenza o il decorso delle infezioni è stata ampiamente comprovata (Apanius et
al. 1966; Bernatchez & Landry 2003). Alcuni Autori hanno dimostrato una
significativa associazione tra aplotipi o alleli delle MHC e le malattie (Briles et al.
1977; Hill et al. 1991; Decamposlima et al. 1993; Godot et al. 2000; Langefors et al.
2001; Wegner et al. 2003 b; Harf & Sommer 2005), altri Autori hanno dimostrato il
vantaggio dell’eterozigosi delle MHC (Penn et al. 2002; McClelland et al. 2003),
oppure la superiorità competitiva degli individui con un elevato numero di
sequenze varianti delle MHC (Wegner et al. 2003 a; Bonneaud et al. 2004; Wegner
et al. 2004).
L'obiettivo della presente ricerca è di valutare la variabilità genetica intra ed
inter specifica in popolazioni selvatiche di camoscio alpino e pirenaico e di valutare
se la pressione selettiva esercitata da infezioni parassitarie può modulare la
variabilità genetica degli ospiti e se tale variabilità può essere correlata con la
diversa capacità dell’ospite di sviluppare una resistenza al parassita e con le
percentuali di sopravvivenza agli eventi epidemici.

6
1.1.1 I markers neutrali
Studi genetici condotti su animali selvatici sono stati eseguiti negli ultimi venti
anni, per quantificare il differenziamento genetico ed i livelli di diversità genetica in
individui e popolazioni potenzialmente isolate, utilizzando dei markers neutrali,
come sequenze di DNA mitocondriale, microsatelliti o polimorfismi di singoli
nucleotidi (SNPs) (Avise 2000; Sunnucks 2003; Lowe et al. 2004). Mentre questi loci
sono molto importanti per analizzare gli aspetti demografici ed evolutivi come la
ricostruzione filogenetica e la storia delle popolazioni, i livelli e le modalità di flusso
genico tra le popolazioni, i livelli di ibridazione tra specie e sottospecie, il grado di
parentela tra gli individui (Blouin et al. 1996; Balloux & Lugon-Moulin 2002;
Brumfield et al. 2003; Morin et al. 2004), la variabilità dei loci neutrali tende però a
sottostimare o ignorare i processi adattativi importanti e non ci da informazioni
dirette in merito ai processi selettivi coinvolti nella relazione tra individui ed il loro
ambiente ovvero sulla capacità di adattamento ai cambiamenti futuri (Meyers &
Bull 2002, Van Tienderen et al. 2002). Questi aspetti, comunque, sono rilevanti in
studi di ecologia evolutiva e di biologia della conservazione (Crandall et al. 2000;
Stockwell et al. 2003). Studi recenti, inoltre, condotti in diversi taxa e condizioni
hanno rilevato che l’evoluzione spesso si realizza in una scala temporale piuttosto
breve, ad esempio in decadi (Stockwell et al. 2003). In alcuni casi, il tempo
trascorso tra la separazione di popolazioni è troppo breve per lasciare un “segno”
evidenziabile in loci neutrali così le differenze tra le popolazioni possono essere
rilevate solo nei geni sottoposti a selezione (Cohen 2002), come ad esempio i geni
altamente variabili del complesso maggiore di istocompatibilità. Contrariamente ai
markers neutrali, la variabilità delle MHC può darci informazioni riguardo i processi
evolutivi ed adattativi. Il confronto tra i markers neutrali, che sono loci non tradotti
in proteine e ritenuti quindi debolmente sottoposti a pressioni selettive, e loci
codificanti e chiaramente soggetti alla selezione naturale come i geni delle MHC,
sono molto importanti per distinguere i ruoli relativi che hanno avuto i processi

7
demografici e quelli selettivi nel modellare la variabilità genetica intra e tra
popolazioni, e per capire direttamente i livelli di variabilità genetica adattativa
(Hedrick et al. 2001 a, b; Sommer 2005; Mona et al. 2008). Lo studio della
variabilità genetica adattativa delle MHC è importante sia per rispondere alle
questioni di tipo ecologico relative all’individuo stesso ed all’interazione tra
organismi ed il loro ambiente (fitness: sopravvivenza, capacità riproduttiva, scelta
ed attrazione del maschio per la riproduzione, fecondità, resistenza alle malattie,
etc.), ma ha anche un valore negli studi di conservazione e gestione della
variabilità genetica di una specie, e quindi della specie stessa (Amos 2000, Slate
et al. 2000, Acevedo-Whitehouse et al. 2003).
1.1.2 I markers dei processi adattativi
Molte popolazioni naturali di vertebrati sono minacciate non solo dalla
drammatica diminuzione dell’areale totale del proprio habitat ma anche dalla
crescente frammentazione e degradazione dello stesso, che ha causato una
riduzione della dimensione delle popolazioni attraverso la generazione di barriere
che hanno ostacolato il flusso genico tra gli individui (Wahlberg et al. 1996; Meffe
& Carral 1997; Peacock & Smith 1997). Le piccole popolazioni soffrono della
riduzione della diversità genetica dovuta alla deriva genetica ed agli effetti
dell’inbreeding (Harrison & Astings 1996; Dudash & Fenster 2000; Keller & Waller
2002). Gli effetti negativi, quali l’aumento della velocità della perdita allelica, la
fissazione di alleli svantaggiosi e la minore disponibilità di individui eterozigoti,
sono stati osservati in studi teorici ed empirici (Primack 1993; Saccheri et al. 1998).
La perdita di variabilità genetica può essere associata alla riduzione a breve termine
dei componenti della fitness come la sopravvivenza, la capacità riproduttiva, la
velocità di accrescimento ed alla ridotta abilità di adattarsi ai cambiamenti a lungo
termine dell’ambiente (Primack 1993; Lande 1988; Brewer et al. 1990; Lacy 1997;
Frankham & Ralls 1998; Biilsma et al. 2000). Un crescente numero di studi indicano
che la diversità genetica dell’ospite gioca una ruolo rilevante nella resistenza delle

8
popolazioni contro patogeni ed epidemie in generale (Keller & Waller 2002;
Coltman et al. 1999; Meagher 1999; Cassinello et al. 2001; Little 2002; Altizer et al.
2003; Bernatchez & Landry 2003; Spielman et al. 2004). Le diverse funzioni e
caratteristiche che coinvolgono le MHC relativamente ai processi biologici ed
ecologici degli organismi, rendono i geni delle MHC quali candidati ottimali per
studiare i meccanismi ed il significato dell’adattamento molecolare nei vertebrati.
1.1.3 Studi sulla variabilità genetica delle MHC nei vertebrati
Il complesso maggiore di istocompatibilità comprende una regione di geni
altamente polimorfici i cui prodotti sono espressi da una gran varietà di cellule, che
giocano un ruolo centrale nel meccanismo di risposta immunitaria ad antigeni
proteici. I prodotti di questi geni, infatti, forniscono il sistema per rendere
riconoscibili peptidi antigenici ai linfociti T. Nelle popolazioni esistono diverse
forme alleliche codominanti nell’MHC ognuno dei quali può avere una diversa
capacità di legare e presentare determinati antigeni proteici. Geni dell’MHC di
classe II sono stati studiati in molti vertebrati tra i quali alcune specie ittiche, quali il
salmone, la carpa e lo spinarello (Bakker & Zbinden 2001; Reusch et al. 2001, 2004;
Ottovà et al. 2005). Tali studi hanno evidenziato una elevata variabilità non solo
intra-specifica ma anche trans-specifica con l’esistenza di linee evolutive comuni
condivise tra specie diverse. La variabilità genetica delle MHC, principalmente
concentrata in una regione del gene denominata DRB (Domine Region of Binding)
che codifica per la regione PBR, (Peptide-Binding Region) è correlata alla capacità
di rispondere e di riconoscere una elevata varietà di peptidi antigenici diversi. I
parassiti possono rappresentare un fattore potenziale per modulare la diversità
dell’MHC. Questo può avvenire sia per un vantaggio dell’eterozigote, in quanto
capace di riconoscere uno spettro più ampio di antigeni derivanti da patogeni, sia
per un vantaggio degli alleli rari, in quanto potenzialmente in grado di rispondere a
nuove forme di parassiti (Hughes & Nei 1992; Jeffery & Bangham 2000). D’altro
canto va sottolineato che questo processo di aumento della variabilità può non

9
essere necessariamente un beneficio per le specie e può essere limitato dalla
necessità di eliminare i linfociti T che reagiscono contro combinazioni tra MHC e
peptidi di origine endogena (Wegner et al. 2003 b).
Numerosi studi sul polimorfismo dell’MHC sono stati condotti su mammiferi
ungulati e in particolare su bovini e ovini (Lewin et al. 1999), sulla capra (Takada et
al. 1998), sul cervo (Swarbrick et al. 1995), sull’alce (Mikko & Anderson 1995), sul
capriolo, la renna e il daino (Mikko et al. 1999). Tali studi hanno messo in evidenza
una significativa differenza tra specie e specie nei parametri di variabilità genetica.
In alcuni casi elevati valori di polimorfismo dell’MHC sono stati registrati anche
successivamente a significativi “colli di bottiglia” come quello causato dalla peste
bovina nel bufalo africano alla fine del XIX secolo (Wenink et al. 1998).
Studi sull’impatto della presenza di patogeni e in particolare degli agenti di
parassitosi sulla variabilità del complesso maggiore di istocompatibilità hanno
dimostrato che elevati livelli di polimorfismo possono essere correlati con la
capacità di difendersi dagli agenti patogeni (Hedrick 2002) e che popolazioni che
mostrano bassi livelli di polimorfismo genetico possono essere più suscettibili alle
malattie parassitari.
Per quanto riguarda il camoscio alpino (Rupicapra rupicapra rupicapra), studi
sulla variabilità genetica dell’MHC, principalmente concentrata nella regione del
gene denominata PBR (Peptide-Binding Region) hanno dimostrato alti livelli di
polimorfismo nel gene DRB (Schaschal et al. 2004, 2005, 2006; Alvarez-Busto et al.
2007; Mona et al. 2008). Tali alti livelli di polimorfismo erano inaspettati poiché
l’habitat alpino del camoscio dovrebbe fornire scarsità d’infezioni parassitarie, per
le condizioni climatiche avverse, per i periodi più brevi a disposizione degli stadi
infettivi dei patogeni, e per una generalizzata diminuzione della biodiversità. La
storia demografica di questa specie, caratterizzata da elevata frammentazione degli
habitat frequentati e le forti riduzioni della popolazione dovuta alle infestazioni di

10
rogna sarcoptica (Rossi et al. 1995) avrebbero dovuto, inoltre, influenzare
negativamente i livelli di variabilità genetica delle popolazioni di camoscio.
1.2 Il Complesso Maggiore di Istocompatibilità
Il complesso maggiore di istocompatibilità (MHC) è stato identificato
inizialmente attraverso il suo ruolo nel rigetto dei trapianti. La capacità di
distinguere il sé dal non-sé è una caratteristica protettiva di quasi tutti gli organismi
multicellulari; questa capacità risponde ad una esigenza di preservazione
dell’individualità e, in generale, serve ad assicurare che i meccanismi di difesa siano
diretti verso microrganismi infettanti ed altre molecole estranee senza causare
danni ai tessuti dell’ospite.
Le strutture fondamentali dell’MHC sono proteine di superficie altamente
polimorfiche. Due sono le classi di molecole identificate nel sistema dei vertebrati,
codificate da un tratto di DNA di 2000-4000 Kilobasi. Si tratta di un gruppo di geni
strettamente connessi che costituiscono il più importante componente genetico
del sistema immunitario dei mammiferi (Klein 1986).
Le molecole di classe I, i classici “antigine da trapianto”, sono presenti sulla
superficie di tutte le cellule nucleate (Piertney & Oliver 2006) e sono coinvolte nel
riconoscimento delle cellule bersaglio da parte delle cellule T citotossiche. Le
cellule bersaglio possono essere sia cellule autologhe infettate da virus sia cellule
tumorali: le cellule T citotossiche riconoscono, sulla superficie di queste cellule, le
molecole di classe I autologhe in associazione con antigeni virali o agenti tumorali
(il sé modificato); nel caso di cellule di un trapianto, gli antigeni MHC estranei
espressi su queste cellule sono riconosciuti dalle cellule T citotossiche in quanto
non-sé (Klein 1986).

11
Un secondo gruppo di geni codificano per le molecole di classe II delle MHC. Le
molecole di classe II sono espresse principalmente sulle cellule che “presentano”
l’antigene (APC) e sui linfociti B. Le molecole di classe II hanno un ruolo
fondamentale nella “presentazione” dell’antigene ai linfociti T helper (Th) da parte
delle APC. Le cellule Th , infatti, riconoscono l’antigene solo se esposto sulla
superficie delle APC in associazione con le molecole di classe II delle MHC (Klein &
Horejsi 1997; Dengjel et al. 2005).
1.2.1 Struttura e funzione delle proteine delle MHC
Le molecole di classe I consistono di un eterodimero formato da una catena
pesante (43 kDa) legata, non covalentemente, ad una catena più piccola (11 kDa),
chiamata β2 microglobulina, monomorfica, codificata al di fuori delle MHC (fig. 1.1).
La catena pesante è organizzata in tre domini globulari esposti all’esterno ed è
ancorata alla membrana da un tratto idrofobico, terminando con un corto
segmento idrofilico C-terminale all’interno del citoplasma.

12
In base alla loro sequenza e
struttura cristallina queste molecole
si dividono in quattro domini: un
dominio N-terminale legante
peptidi, uno extra-cellulare simile a
quello dell’emoglobina, un dominio
trans-membrana e il dominio
citoplasmatico. Due di questi
domini (α1 e α2) formano una tasca
che può ospitare un peptide di 9−11
ΑΑ, che una volta legato viene
presentato alle cellule T; gli altri due
domini (α3 e β1) stabilizzano la
molecola e vengono detti “dominio
simil Ig”.
Figura 1.1 Strutture delle molecole MHC di classe I e II. Sono rappresentati i domini, la parte transmembranaria e le tasche di legame per il peptide antigene. Immagine tratta da: Abbas Lichtman, Fondamenti di Immunologia, Piccin.
Le molecole di classe II sono anch’esse glicoproteine transmembranarie, e
consistono di una catena alfa e di una catena beta di peso molecolare 34 e 28 kDa
rispettivamente. Sono costituite ciascuna da due domini globulari ed ancorate alla
membrana in modo analogo alle molecole di classe I. Anche per le molecole di
classe II ritroviamo i quattro domini: un dominio legante peptidi, formato dalle
catene α1 e β1, un dominio simil Ig formato dalle catene α2 e β2, un dominio
transmembrana e un dominio citoplasmatico.

13
Sia le molecole di classe I che quelle di classe II fungono da bersaglio per i
linfociti T, che regolano la risposta immunitaria. Gli antigeni espressi in associazione
all'MHC di classe I vengono riconosciuti dai linfociti T citotossici, anche detti linfociti
T CD8+ che vengono attivati agendo da effettori per la risposta immunitaria cellulo-
mediata, ossia risposte immunitarie specifiche, provocando la lisi delle cellule che
esprimono tali proteine. Queste ultime possono essere cellule tumorali o cellule in
cui stanno avvenendo processi di replicazione di virus. L'MHC di seconda classe sarà
riconosciuto invece dai recettori dei linfociti T helper, altrimenti detti linfociti T
CD4+, che rispondo alla stimolazione producendo citochine.
Le cellule T, in dettaglio, riconoscono gli antigeni nel contesto delle molecole
delle MHC, non riconoscono di fatto gli antigeni nella loro forma nativa, ma
piuttosto dei loro frammenti peptidici generati dalle APC. Le APC infatti, dopo aver
captato l’antigeni, lo internalizzano e lo sottopongono a proteolisi. Solo dopo aver
così modificato l’antigene, le APC ne espongono i frammenti in membrana,
associati alle molecole MHC. Ma come avviene l’interazione fra le molecole MHC
ed i frammenti dell’antigene derivati dalla proteolisi? La risposta a questa domanda
viene da due osservazioni sperimentali. Confrontando le sequenze aminoacidiche
di differenti forme alleliche delle molecole di classe I e classe II, si è osservato che
le variazioni alleliche non sono distribuite a caso e che i residui polimorfici sono
raggruppati in regioni particolari della molecola. Recentemente, inoltre, la
definizione della struttura tridimensionale per le molecola delle MHC di classe I e di
classe II, ha messo in evidenza il fatto che i residui aminoacidici variabili di queste
proteine sono disposti in una profonda tasca della molecola, mentre i residui
invarianti restano all’esterno. E’ ovvio che la posizione dei residui variabili
all’interno della tasca è strategica, perché determina la capacità delle molecole
delle MHC di formare un sito accettore per i frammenti peptidici originati dalla
degradazione dell’antigene (Fig. 1.2).

14
Figura 1.3 Le molecole di classe II, insieme al peptide a esse associato, vengono riconosciute dal TCR. La parte invariante della molecola MHC è legata invece dalla molecola CD4.
Figura 1.4 Le molecole di classe I, insieme al peptide ad esse associato, vengono riconosciute dal TCR. La parte invariante della molecola MHC è legata invece dalla molecola CD8.
Il riconoscimento dell’antigene, quindi, da parte dei linfociti T avviene
attraverso un recettore che riconosce il complesso dell’antigene alloggiato in un
sito formato dalla parte polimorfica delle molecole MHC. Questi siti polimorfici si
Fig. 1.2 La tasca di legame per l’antigene è formata solo dalla catena α nella classe I e dalle catene α e β nella classe II. La classe I può alloggiare peptidi di 8-11 aa, mentre la classe II può alloggiare peptidi di 10-30 aa. Il legame è possibile se pochi requisiti strutturali vengono soddisfatti. Immagine tratta da: Abbas Lichtman, Fondamenti di Immunologia, Piccin.

15
trovano nella parte N-terminale delle catene alfa e beta delle molecole di classe II
(Fig. 1.3) e nei domini α1 e α2 delle molecole d classe I (Fig. 1.4).
Le molecole CD4 e CD8 hanno struttura invariabile ed interagiscono con parti
invarianti delle molecole MHC, stabilizzando l’interazione TCR con il complesso Ag-
MHC.
Nella figura seguente vengono sintetizzate le modalità di azione delle molecole
di classe I e di classe II delle MHC.
Figura 1.5 Rappresentazione schematica dei meccanismi di azione delle molecole di classe I e classe II delle MHC. Vie di ingresso degli antigeni nella cellula e vie di processamento.
In merito ai geni codificanti per le molecole di classe II, molte ricerche in
mammiferi si sono concentrate sul secondo esone codificante le porzioni funzionali
dei siti di legame dell’antigene (Peptide Binding Region, PBR)(Ohta 1998). I patterns
osservati per i loci DRB codificanti per la regione PBR, comunque, sono in generale
dei buoni indicatori della variabilità genetica delle MHC costituendo uno dei loci
polimorfici più conosciuti nei vertebrati (Hedrick 1994; Huges & Huges 1995). La

16
variabilità delle molecole delle MHC è correlata con la diversità dei recettori dei
linfociti T che determinano la resistenza a malattie o parassitosi di un organismo ed
in conclusione potrebbe influenzare a lungo termine la probabilità di sopravvivenza
di una popolazione (Paterson et al. 1998; Hedrick et al. 2001 a, b; Langefors et al.
2001).
1.2.2 L’organizzazione genetica delle MHC
L’elevato polimorfismo ritrovato nelle MHC di classe II si concentra
principalmente nell’esone 2, che codifica per il β1-dominio del sito di legame
dell’antigene, PBR. In alcune specie di ruminanti ungulati come bovini (Bos taurus)
ed ovini (Ovis aries) sono stati caratterizzati i seguenti geni delle MHC di classe II:
DRA, DRB, DQA e DQB. Nei bovini la regione DR comprende 3 geni DRB ed un gene
DRA (Lewin et al. 1999). Tra i tre loci DRB, il DRB3 viene espresso maggiormente
rispetto agli altri loci e mostra un maggiore polimorfismo. Più di 100 DRB3 alleli
sono ad oggi conosciuti nei bovini (Bola Committee, Web site:
http://www.projects.roslin.ac.uk/bola/DRBtab.html).
Negli ovini l’organizzazione delle MHC di classe II è similare a quella dei bovini,
con la differenza che il locus maggiormente polimorfico è il DRB1 (Dutia et al. 1994;
Schwaiger et al. 1996). Altri studi confermano l’elevato polimorfismo del locus DRB
in Capra aegagrus e Capra hircus (Schwaiger et al. 1993; Takada et al. 1998), Ovis
canadensis (Gutierrez-Espeleta et al. 2001), nel cervo Odocoileus virginianus (Van
der Bussche et al. 1999) e Cervus elaphus (Swarbrick et al. 1995). D’altra parte,
Mikko & Andersson (1995) e Ellengren et al. (1996) rilevarono un basso livello di
variabilità genetica in corrispondenza dei medesimi loci, in Alces alces del Nord
America e del Nord Europa. Inoltre, una limitata diversità nel DRB locus è stata
anche riscontrata in Capreulus capreulus e Rangifer tarandus della Norvegia e della
Svezia, ed in Ovibos moschatus e Cervus doma in cui questo locus è monomorfico
(Mikko et al. 1999). Nel bufalo africano (Syncerus caffer), infine, è stato rilevato una

17
elevata variabilità genetica del locus DRB3 a dispetto degli abbattimenti causati da
una epidemia di peste scoppiata alla fine del XIX secolo (Wenink et al. 1998). La
stessa Ovis canadensis ha mostrato un aumento dei livelli di polimorfismo in
corrispondenza di un declino della popolazione provocato principalmente da una
epidemia trasmessa dai bovini (Gutierrez-Espeleta et al. 2001).
Per quanto riguarda il camoscio alpino (Rupicapra rupicapra rupicapra), studi
sulla variabilità genetica delle MHC, principalmente concentrata nella regione del
gene PBR (Peptide-Binding Region) hanno dimostrato alti livelli di polimorfismo nel
gene DRB (Schaschl et al. 2004, 2006). Tali alti livelli di polimorfismo erano
inaspettati poiché l’habitat alpino del camoscio dovrebbe fornire scarsità
d’infezioni parassitarie, per le condizioni climatiche avverse, per i periodi più brevi a
disposizione degli stadi infettivi dei patogeni, e per una generalizzata diminuzione
della biodiversità. La storia demografica di questa specie, caratterizzata da elevata
frammentazione degli habitat frequentati e le forti riduzioni della popolazione
dovuta alle infezioni di rogna sarcoptica (Rossi et al. 1995) avrebbero dovuto,
inoltre, influenzare negativamente la variabilità genetica delle popolazioni di
camoscio.
1.2.3 Il polimorfismo delle MHC
Uno degli aspetti peculiari del complesso MHC è il suo incredibile polimorfismo
(Robinson et al. 2003), mostrando in confronto con altri loci una poco comune alta
variabilità nel numero di alleli ed elevata variabilità nucleotidica (Parham & Ohta
1996). Considerando la funzione dei prodotti genici delle MHC, è facile immaginare
una buona ragione per spiegare il vantaggio del polimorfismo di questo locus. Gli
antigeni sono visti nel contesto dei prodotti genici delle MHC, ed antigeni diversi
interagiscono in modo efficace con alcune, ma non tutte, le proteine delle MHC.
L’esteso polimorfismo dei geni MHC garantisce una più alta probabilità che un
individuo possa esprimere almeno una proteina MHC in grado di interagire bene

18
con un dato antigene, rendendo il soggetto responsivo a quella sostanza. A livello di
popolazione, il polimorfismo aumenta fortemente la probabilità che almeno alcuni
individui della popolazione sopravvivano in caso di improvvisa “entrata in scena” di
un nuovo antigene, per esempio un virus o un parassita mortale.
L’elevata variabilità, è stato visto concentrarsi in una zona ben definita delle
MHC che corrisponderebbe al sito di legame dell’antigene che mostra alti livelli di
varianti non solo per quanto concerne il numero degli alleli codificanti coinvolti ma
anche per l’estensione delle sequenze varianti degli alleli stessi (Huges & Yeager
1998). Secondo l’ipotesi nulla, si è dimostrato che la velocità delle sostituzioni
sinonime dei nucleotidi (ds) è più alta (ds>dn) della velocità delle sostituzioni non-
sinonime (dn) perché le sostituzione non sinonime variano la composizione degli
aminoacidi delle proteine provocando cambiamenti deleteri (Hughes & Nei 1988,
1989); molti studi, comunque, dimostrano che il sito di legame dell’antigene
peptidico (PBR) mostra più sostituzioni non sinonime che sostituzioni sinonime
(dn>ds) (Brown et al. 1988, 1993). Questo non può essere spiegato da una più
elevata velocità di mutazione in questa specifica regione (Huges & Yeager 1998;
Brown et al. 1988, 1993). Un ruolo fondamentale nell’arrangiamento della
variabilità nucleotidica a livello dei geni delle MHC è svolto dall’azione della
selezione bilanciante che favorisce positivamente la diversificazione dei loci
(Hughes & Nei 1988, 1989; Bergstrom & Gyllensten 1995; Bernatchez & Landry
2003). La selezione bilanciante non agisce solo nel mantenere un elevato numero di
alleli nelle popolazioni ma opera anche nella persistenza della diversità allelica in
periodi estremamente lunghi relativamente alla variazione genetica neutrale
(Richman 2000), definita come “evoluzione trans-specifica del polimorfismo” (Klein
1986). La conseguente alterazione del PBR assicura il “processamento” di diversi
tipi di antigeni (Brown et al. 1988, 1993; Janeway & Travers 2002).
Il mantenimento del polimorfismo intra popolazione dipende dal risultato
dell’intensità di selezione, dalla velocità di mutazione e dalla grandezza della

19
popolazione stessa (Huges & Yagger 1998). In determinate circostanze la forza della
selezione operante sugli alleli delle MHC può risultare insufficiente a mantenere la
variazione quando la popolazione è troppo piccola numericamente o troppo
frammentata (Sommer 2005). La ricombinazione intragenica sembra essere uno
dei meccanismi più importanti deputati a sostenere la variabilità genomica delle
MHC (Gyllensten et al. 1991; Andersson & Mikko 1995; Bergstrom et al. 1998;
Richman et al. 2003; Schaschl 2006). L’abilità di una popolazione naturale di
mantenere la variabilità genetica in geni funzionali dipende dalle pressioni selettive
coinvolte. La selezione bilanciante contrasta gli effetti della deriva genetica e
ritarda la fissazione di alleli
1.2.4 I meccanismi di selezione
I due principali meccanismi della selezione bilanciante (ipotesi del vantaggio
dell’eterozigote e selezione frequenza-dipendente) sono stati considerati i maggiori
responsabili degli alti livelli di polimorfismo delle MHC nell’uomo e nei vertebrati
(Hedrick 1994, 1999; Jeffery & Bergham 2000; Richman 2000; Miller et al. 2001;
Penn 2002; Bernatchez & Landry 2003).
La selezione naturale, in generale, opera attraverso due meccanismi favorendo
gli eterozigoti o gli alleli rari; gli individui eterozigoti sarebbero favoriti perché sono
in grado di far fronte a un numero più grande di antigeni. Alternativamente se gli
alleli sono vantaggiosi per il portatore quando sono rari, in quanto i patogeni sono
selezionati per sfuggire al maggior numero di alleli MHC, la variabilità genetica è
mantenuta in un tipico ciclo di coevoluzione ospite-parassita.
In particolare, nel valutare il potenziale evolutivo dei meccanismi del vantaggio
dell’eterozigote bisogna fare una differenziazione tra dominanza (vantaggio
dell’eterozigote in senso stretto) e sovradominanza (superiorità dell’eterozigote). Il
termine dominanza si riferisce al fatto che alla eterozigosi è associata la resistenza

20
a malattie per esempio, ma di fatto possono mascherare alleli suscettibili recessivi.
La sovradominanza, invece, è il tipico meccanismo che promuove la variabilità delle
MHC (Penn 2002; Penn et al. 2002). In questo caso, infatti, gli eterozigoti mostrano
una maggiore fitness rispetto ai parenti omozigoti, poiché posseggono una
maggiore varietà di MHC capaci di rispondere ad un ampio range di antigeni
(Hughes & Nei 1989; Takahata & Nei 1990).
Il secondo meccanismo, selezione frequenza-dipendente, si realizza quando un
allele o genotipo è favorevole o meno se portato con una certa frequenza
(Takahata & Nei 1990; Clarke & Kirby 1966). Le dinamiche ospite-parassita possono
essere considerate come un ciclo coevolutivo. I patogeni si adattano ad infettare i
più comuni genotipi ospiti, lasciando i genotipi ospiti rari non infettati (Lively &
Dybdahl 2000). Se gli alleli sono favoriti quando sono rari, ma contro selezionati
quando sono comuni, ne risulta un polimorfismo bilanciato. Gli alleli delle MHC che
forniscono la resistenza ad un maggior numero di parassiti conferiscono un
vantaggio dell’ospite, si trasmettono alla popolazione e divengono comuni. Questo
meccanismo aumenta la selezione dei parassiti capaci di evadere il riconoscimento
da parte di questi alleli comuni. Come la patogenicità dei parassiti varia, la fitness
relativa dei genotipi comuni dell’ospite diminuisce e crea un vantaggio selettivo in
favore degli alleli rari.
In più, meccanismi riproduttivi come la scelta del compagno disassortativa e le
interazioni materno-fetali potrebbero essere considerati come meccanismi
alternativi o complementari del mantenimento della diversità delle MHC (Potts &
Wakeland 1993; Edwards & Potts 1996; Penn & Potts 1999; Penn 2002; Penn &
Fisher 2004). La preferenza del compagno con maggiore variabilità delle MHC
favorirebbe: i. l’aumento della persistenza degli eterozigoti nella prole (Zuk 1990);
ii. promuovere figli con vari target di alleli delle MHC come protezione contro
patogeni che si adattano rapidamente ai genotipi parentali (Clarke & Kirby 1966;
Takahata & Nei 1990; Penn & Potts 1999); iii. sfavorire l’inbreeding o

21
l’incompatibilità genetica (Brown & Eklund 1994; Acevedo-Whitehouse et al. 2003);
iv. assicurare una diversità ottimale nella prole per rispondere alla resistenza del
parassita (Hedrick 2004).
1.3 Caratteristiche generali del camoscio
Il camoscio è un mammifero tipico degli ambienti montani che appartiene al
superordine degli Ungulati (il termine ungulato significa “provvisto di zoccolo”); la
sua classificazione completa è riportata in Tabella 1:
Camoscio alpino
Classificazione Caratteristiche generali
Phylum Cordati Parte terminale delle dita ricoperte da robuste unghie.
Subphylum Vertebrati Numero di dita pari.
Classe Mammalia Stomaco con quattro camere.
Superordine Ungulata Corna cave, non caduche, crescita annuale, presenza in entrambi i sessi.
Ordine Artiodactyla
Sottordine Ruminantia
Famiglia Bovidae
Sottofamiglia Caprinae
Tribù Rupicaprini Genere Rupicapra
Specie Rupicapra rupicapra rupicapra, Linnaeus, 1758
Tabella 1: Inquadramento tassonomico del camoscio delle Alpi.
A livello specifico, la tassonomia del camoscio ha subito recentemente alcune
revisioni, e ancora adesso non è del tutto chiara e condivisa dai diversi autori. Fino
alla metà degli anni ottanta si pensava che il genere possedesse un’unica specie
sulla base dei dati morfologici e dei reperti fossili (Lydekker 1913; Couturier 1938;
Dolan 1963). Questa specie era R. rupicapra, che era suddivisa in dieci sottospecie
sulla base delle aree geografiche di provenienza.

22
Negli anni seguenti furono condotte delle analisi più approfondite dei caratteri
morfologici (Lovari & Scala 1980; Lovari 1987; Masini & Lovari 1988), etologici
(Lovari 1985) e genetici (Nascetti et al. 1985) che portarono all’identificazione di
due specie: R. rupicapra distribuito in tutta l’Europa centro-orientale e suddiviso in
7 sottospecie (cartusiana, rupicapra, tatrica, carpatica, balcanica, asiatica e
caucasica) e R. pyrenaica localizzato solamente sulla catena dei Pirenei e in centro
Italia, e comprendente 3 sottospecie (parva, pyrenaica e ornata) (Fig. 1.7 a e b).
Queste distinzioni sono state recentemente confermate da studi basati su
marcatori genetici molecolari (microsatelliti, d-loop, ecc.). Negli ultimi anni si è
sviluppata l’idea, basandosi anche su dati genetici (Pérez et al. 2002), che l’alta
differenziazione morfologica riscontrata in R. pyrenaica ornata (il camoscio
d’Abruzzo) possa essere ricondotta ad uno status di nuova specie: il camoscio
Appenninico. Questa differenziazione potrebbe essere dovuta al forte isolamento
geografico che caratterizza questa popolazione del centro Italia da lungo tempo.
Agli stessi risultati arriva anche un lavoro recente che analizza delle popolazioni di
camoscio provenienti da diverse regioni europee, tra cui alcuni campioni di R.
pyrenaica ornata. Questo studio arriva alla conclusione che ci sarebbero tre linee
evolutive differenti nel genere Rupicapra: quella del camoscio nord-orientale,
quella sud-occidentale e una linea indipendente per R. pyrenaica ornata
(Crestanello 2002-2004).
Il camoscio alpino e quello dei Pirenei possiedono alcune caratteristiche
distintive:
il camoscio dei Pirenei ha dimensioni inferiori;
il mantello invernale ha una diversa colorazione. La macchia golare
chiara nella specie R. pyrenaica è nettamente più estesa, e arriva
fino al petto dell’animale; due bande scure la separano dalle parti
biancastre o isabelline poste ai lati del collo che scendono fino alle

23
spalle. Queste chiazze non sono presenti nel manto del camoscio
settentrionale così come quelle localizzate nella coscia.
la fontanella etmoidale, cioè quella parte di cranio compresa tra
l’osso frontale e l’osso mascellare, appare più lunga e
completamente suturata nella specie pyrenaica, mentre è più corta
e ancora aperta nella specie rupicapra. Inoltre la distanza tra le
corna alla base e le dimensioni medie del cranio sono più grandi in
rupicapra, mentre l’angolo tra le corna e il cranio é maggiore in
pyrenaica (le corna in questa specie sono inclinate all’indietro).
Le due specie hanno sviluppato nel corso dell’evoluzione moduli
comportamentali diversi, soprattutto nella sfera riproduttiva.
Se la suddivisione in due e probabilmente tre specie viene ormai accettata da
molti, c’è sicuramente più disaccordo sulla suddivisione nelle varie sottospecie,
visto anche che i caratteri morfologici non sono in grado da soli di differenziarle in
modo sicuro.
Figura 1.7 a Rupicapra rupicapra rupicapra (foto scaricata dal sito www.stelviopark.it/.../Bolzano/bz_04.htm

24
Figura 1.7 b Rupicapra pyrenaica pyrenaica (foto di David Shackleton)
1.3.1 Origine ed evoluzione del camoscio
Per comprendere le differenze nella distribuzione attuale delle diverse specie di
camoscio, è necessario vedere quali sono le ipotesi sulla storia evolutiva della tribù
dei Rupicaprini. Uno dei possibili progenitori dei Rupicaprini si pensa possa essere
Pachigazzella grangeri, un genere comparso nel Pliocene in Cina, circa 5-7 milioni
di anni fa (Masini & Lovari 1988).
Dall’Asia centrale nel tardo Miocene (10 milioni di anni fa) i Rupicaprini si
sarebbero diffusi verso sud, est e nord-ovest sino ad occupare il loro attuale areale
distributivo. In particolare si ritiene che gli avi del camoscio si siano diffusi verso
occidente, a partire da una regione ad ovest dell’Himalaya, lungo le catene
montuose del sistema asiatico durante il Pleistocene medio (da 120.000 a 700.000
anni fa), approfittando delle glaciazioni per estendere l’areale distributivo verso
ovest (Fig.1.8).

25
In Europa, l’esistenza del camoscio durante la glaciazione del Riss (250-150 mila
anni fa) è suggerita da un unico reperto fossile, rinvenuto sui Pirenei francesi
(Caune de l’Arago) e attribuito alla forma pyrenaica.
A partire dalla glaciazione del Würm in avanti (tra 80.000 e i 12.000 anni fa) i
fossili di questo rupicaprino diventano comuni e confermano la distinzione già
esistente tra le specie rupicapra e pyrenaica. La specie pyrenaica si sarebbe evoluta
in Europa occidentale direttamente dai Rupicaprini immigranti nel medio
Pleistocene, durante la glaciazione del Riss, mentre rupicapra si sarebbe evoluta in
Europa orientale o in Asia Minore durante un periodo di isolamento interglaciale
(Lovari & Scala 1980).
Figura 1.8: Probabile radiazione dei Rupicaprini (da Lovari & Scala 1980).
Durante la glaciazione del Würm, i nuovi camosci di tipo rupicapra avrebbero
raggiunto, con una seconda ondata colonizzatrice, l’Europa centro-meridionale.
L’areale distributivo quindi sarebbe stato molto più ampio e continuo dell’attuale e
avrebbe compreso gran parte dell’Europa centrale e centro-meridionale, con la
specie pyrenaica già geograficamente diffusa nella penisola Iberica e negli

26
Appennini, e la specie rupicapra diffusa dal Caucaso all’arco alpino, fino alle Alpi
Apuane e, forse, agli Appennini settentrionali.
Alla fine della glaciazione del Würm (circa 12.000 anni fa), con l’innalzamento
della temperatura e i conseguenti cambiamenti ambientali, i camosci di tipo alpino
furono costretti a ritirarsi sui rilievi montuosi, l’areale della specie si frammentò ed
iniziarono così a differenziarsi le diverse sottospecie.
Questa è solo una delle teorie esistenti sulla differenziazione delle varie
sottospecie di Rupicapra. Una seconda ipotesi più semplice prevede che ci sia stato
un unico evento di colonizzazione durante la glaciazione del Riss (da 300.000 a
120.000 anni fa), che portò il camoscio ad invadere tutte le pianure dell’Europa
meridionale, le Alpi e i Pirenei. In seguito, durante periodi più caldi, ci sarebbe stato
il ritiro verso le cime delle principali catene montuose, dove si sarebbero suddivise
specie e sottospecie (Lovari & Scala 1980).
1.3.2 Distribuzione geografica
Il genere Rupicapra è attualmente presente nelle aree montane dell’Europa
occidentale, centrale e dell’Asia Minore, con una distribuzione disomogenea in
relazione ai principali rilievi montuosi, risultato della diffusione naturale e, in parte,
di immissioni effettuate dall’uomo (Figura 1.9). Il camoscio nord-orientale (R.
rupicapra) è naturalmente presente in tutte le regioni montane dell’Europa centro-
occidentale fino ai monti del Caucaso e alla Turchia (massiccio della Chartreuse,
Alpi, Alti e Bassi Tatra, Carpazi rumeni e penisola Balcanica, Bosnia, Montenegro,
Macedonia, Albania, Grecia e Bulgaria), mentre il camoscio dei Pirenei (R.
pyrenaica) è presente in modo discontinuo in alcune aree montane dell’Europa
sud-occidentale (Pirenei, Monti Cantabrici e Appennino meridionale).

27
Figura 1.9 Distribuzione europea del camoscio (in blu R. pyrenaica, in rosso R. rupicapra).
Elenchiamo brevemente la distribuzione delle sottospecie:
R. pyrenaica parva (Cabrera, 1911): presente in Spagna sui monti
cantatrici.
R. pyrenaica pyrenaica (Bonaparte, 1845): presente in Spagna e in
Francia sui Pirenei.
R. pyrenaica ornata (Neumann, 1899): presente in Italia in Abruzzo.
Composta attualmente da 350-400 individui, ora si trovano solo nel
Parco Nazionale d’Abruzzo, ma fino alla fine dell’Ottocento era
presente su tutto l’Appennino centrale.
R. rupicapra cartusiana (Couturier, 1938): presente in Francia nel
massiccio della Chartreuse. E’ a rischio d’estinzione a causa
dell’elevata diminuzione numerica.
R. rupicapra tatrica (Blahout, 1971): presente in Cecoslovacchia e
Polonia sui monti Tatra nei Carpazi occidentali.

28
R. rupicapra balcanica (Bolkay, 1925): presente nei monti della
Jugoslavia centro-meridionale, dell’Albania, della Grecia sulla
catena dei Balcani.
R. rupicapra carpatica (Couturier, 1938): presente nei Carpazi
orientali.
R. rupicapra caucasica (Lydekker, 1910): presente nel Caucaso.
R. rupicapra asiatica (Lydekker, 1908): presente sul Tauro,
Antitauro, Catena dei Monti Pontici.
R. rupicapra rupicapra (Linneo, 1758): presente nelle Alpi francesi,
italiane, svizzere, austriache, jugoslave settentrionali, bavaresi, del
Liechtenstein, nel Giura e introdotto nei Vosgi, nelle regioni
montuose dell’Elba e nella Foresta Nera. Questa sottospecie è la
più numerosa, con l’Italia al terzo posto preceduta da Austria e
Svizzera. In Italia questo camoscio è presente su tutto l’arco alpino
con concentrazioni maggiori nelle aree protette (Parco Nazionale
del Gran Paradiso, Parco Regionale Valdieri-Entracque, Parco
Nazionale dello Stelvio) e in Trentino Alto Adige (Mustoni et al.,
2003).
In Italia il camoscio alpino è uniformemente diffuso, anche se con densità
differenti, in tutte le regioni alpine, dal Friuli - Venezia Giulia (la distribuzione
appare continua nelle province di Pordenone e Udine e puntiforme in quella di
Trieste), Veneto, Trentino–Alto Adige, Lombardia, Piemonte, Valle d’Aosta, sino alla
Liguria (presente in Provincia di Imperia e sporadiche comparse in Provincia di
Savona). Le consistenze e le densità appaiono però estremamente eterogenee a
seconda della zona geografica considerata. L’estensione dell’areale distributivo
ammonta comunque a 41.130 Km2. Il camoscio era molto diffuso su tutto l’arco
alpino fino alla seconda metà del 1700; ebbe poi un notevole calo e, solo dopo la
Seconda Guerra Mondiale, si espanse nuovamente per raggiungere l’attuale

29
consistenza, ma si pensa che questo fenomeno di generale espansione non sia
ancora terminato. Al progressivo miglioramento dello status di questi animali ha
contribuito in maniera determinante la creazione di parchi e di altri istituti di
protezione che hanno favorito un più rapido incremento e stabilizzazione dei nuclei
presenti. Una raccolta e sintesi di informazioni sullo status del camoscio alpino é
stata condotta dal ”Istituto Nazionale per la Fauna Selvatica” che ha preso in
considerazione anche le popolazioni presenti all’interno delle aree protette e nelle
zone precluse all’attività venatoria.
Il numero di esemplari attualmente presente sull’arco alpino è pari a circa
137.000, con un incremento dell’11% rispetto a quanto rilevato nel 2000, dovuto
principalmente dalla crescita delle popolazioni nella parte occidentale dell’arco
alpino. La specie si distribuisce con densità (riferite all'estensione delle sole aree
ritenute idonee alla presenza del Camoscio per ciascuna provincia) comprese fra un
minimo di 1,3 capi/km2 in provincia di Brescia ed un massimo di oltre 10 capi/ km2
in provincia di Lecco.
Nei Parchi e nelle Aziende faunistico venatorie vivono circa 20.000 individui,
pari al 16% dell’intera popolazione italiana. Le consistenze più elevate si registrano
nelle province autonome di Trento e Bolzano e in Piemonte, nei cui territori risulta
al momento presente il 62% dei camosci alpini italiani. I contingenti più numerosi
sono presenti nelle province di Trento e Bolzano (23.000 ciascuna), Belluno, Cuneo,
Torino e Aosta (dai 9.000 ai 12.000) e Sondrio (8.500) (Mustoni et al. 2003).
1.3.3 Biologia del camoscio
Il camoscio è un animale che può arrivare al massimo ad un’età di 20 – 25 anni;
raramente però si raggiungono queste età, infatti le femmine raggiungono
generalmente i 15 – 18 anni e i maschi i 9 – 10 anni. Questo si deve alle difficili
condizioni di vita e, per quanto riguarda i maschi, ad un forte affaticamento nel

30
periodo riproduttivo che li rende più vulnerabili alle insidie invernali. Un altro
fattore che impedisce il raggiungimento da parte del camoscio di età elevate è
l’attività venatoria.
La struttura corporea del camoscio è compatta, con zampe lunghe e forti;
nonostante queste caratteristiche si presenta come un animale agile. Rispetto agli
altri ungulati, è un considerato di media taglia; in età adulta raggiunge pesi di 30–
45 Kg per il maschio e 25–35 Kg per la femmina.
Il mantello si presenta di colore variabile con la stagione, viene cambiato due
volte l’anno con due mute che avvengono in maniera graduale e lenta. Il manto
estivo ha una durata di circa tre mesi e mezzo. In autunno, si verifica la prima muta
che modifica, in previsione del freddo invernale, il colore e la lunghezza del pelo. Il
mantello invernale ha una durata di 8 mesi e mezzo. Con l’arrivo della primavera si
verifica la seconda muta che comporta la perdita dei lunghi e dritti peli scuri e dei
sottili peli di lana, che hanno contribuito all’isolamento dal freddo durante
l’inverno.
Le corna del camoscio sono presenti in entrambe i sessi e si presentano
uncinate e di color ebano. Le corna sono permanenti, a crescita continua;
raggiungono in genere i 22 –26 cm di altezza nei maschi e i 14 – 18 cm nelle
femmine.
Gli arti del camoscio hanno una struttura scheletrica ideale per lo scatto e per
la corsa.
La comunicazione sociale tra i camosci avviene mediante segnali di tipo visivo;
questo animale è dotato di una buona vista anche se non particolarmente acuta, la
visione binoculare (quella tridimensionale) sembra essere ristretta a un campo di
circa 45°. Il riconoscimento tra partner secondo le classi sociali, avviene grazie alla

31
valutazione visiva di alcune caratteristiche riguardanti il colore e la struttura del
corpo e della testa. Anche l’olfatto gioca un ruolo importante nelle interazioni
sociali e nella difesa dai predatori. L’udito è discreto e i padiglioni auricolari
estremamente mobili favoriscono la percezione dei suoni. Le femmine e i piccoli
emettono una specie di belato, mentre i maschi durante la stagione riproduttiva
emettono un suono rauco gutturale.
La vita di relazione del camoscio è influenzata in gran misura dal grado di
maturazione sociale dell’individuo. La struttura sociale di base è di tipo matriarcale
ed è fondata sul legame madre–piccolo, che si scioglie nel secondo anno di vita del
capretto.
Gli animali vengono distinti in capretti (fino al compimento di un anno), giovani
(maschi di 1-2 anni, femmine di un anno), subadulti (maschi di 3-4 anni, femmine di
2-3 anni), adulti (maschi di 5-9 anni, femmine di 4-11 anni) e anziani (maschi di 10 o
più anni e femmine con 12 o più anni). La maggior parte dei camosci vive in gruppi
di dimensioni e composizione variabili a seconda del periodo dell’anno e della zona
geografica.
Il periodo degli amori inizia a fine ottobre e si conclude a metà dicembre, con
un picco nell’attività riproduttiva tra il 20 e 30 novembre. Alcuni maschi, detti
territoriali, difendono piccole aree (diametro di 200-500 metri), da cui scacciano
tutti gli altri maschi adulti e subadulti e in cui tentano di mantenere un gruppo di
femmine mediante comportamenti di corteggiamento ritualizzati. I combattimenti
avvengono principalmente fra i giovani maschi vaganti verso la fine del periodo
degli amori, quando i maschi territoriali e dominanti si ritirano. Al termine della
stagione riproduttiva, che giunge circa a metà dicembre, gli animali si spostano
verso le zone di svernamento dove rimarranno fino alla primavera.

32
In figura 1.10 vediamo il ciclo vitale del camoscio. La gestazione dura circa 6
mesi (25-27 settimane). Il parto avviene in zone tranquille, localizzate su versanti
ripidi e scoscesi, quasi sempre al di sotto del limite degli alberi, dove minimi
risultano il disturbo ed il possibile impatto dei predatori. Tra il 10 maggio e il 10
giugno le femmine partoriscono 1, al massimo 2 (anche se raramente) piccoli.
L’allattamento si protrae con regolarità per 2-3 mesi (sino a 6), ma già a 20-30
giorni il capretto comincia a brucare. Il capretto è in grado di seguire la madre
immediatamente dopo il parto.
Figura 1.10 Rappresentazione schematica del ciclo biologico annuale del camoscio
1.3.4 Una malattia parassitaria: la rogna sarcoptica
La rogna o meglio “Le rogne” sono malattie parassitarie sostenute da acari che
colpiscono la maggior parte degli esseri viventi, compresi i rettili e gli uccelli, anche
se, per l’aspetto clinico, di “rogna vera e propria” si può parlare solo quando sono
coinvolti gli acari del genere Sarcoptes, che colpiscono essenzialmente i mammiferi,
compreso l’uomo.

33
Questo genere, infatti, visibile solamente al microscopio ottico, a differenza di
altri che vivono esclusivamente sulla superficie cutanea dell’ospite nutrendosi di
annessi epidermici (penne o pelo) senza creare evidenti lesioni alla pelle, penetra
nello spessore dell’epidermide infilandosi nei canalicoli delle ghiandole sebacee.
Il passaggio degli acari da un animale all’altro della stessa specie avviene per
contatto diretto, che si realizza attraverso le cure parentali, gli accoppiamenti e le
relazioni sociali o più raramente per via indiretta, attraverso la frequentazione di
luoghi comuni contaminati, quali saline e giacigli dove l’acaro può sopravvivere
pochi giorni, in funzione delle condizioni ambientali che trova. Questa seconda
modalità spiega inoltre, la trasmissione tra alcune specie diverse, com’è avvenuto
inizialmente per la rogna del camoscio che ha colpito pesantemente anche lo
stambecco e la capra selvatica. Va comunque precisato che gli acari della rogna
sono essenzialmente specie-specifici, in quanto l’acaro che colpisce il camoscio,
Sarcoptes scabiei var. rupicaprae non può colpire il cinghiale e la volpe, che
vengono parassitati rispettivamente da Sarcoptes scabiei var. suis e Sarcoptes
scabiei var. canis.
Questo dato, supportato da lavori di ricerca scientifica, esclude in maniera
pressochè certa, la possibilità che la volpe possa fungere da serbatoio o
trasmettere la malattia agli ungulati.
I sintomi nel camoscio (lo stambecco presenta un decorso diverso) si
manifestano entro poco tempo dal contagio, da qualche giorno a tre settimane; al
prurito sempre più intenso, gli animali rispondono con evidente nervosismo e
grattamento incoercibile, strofinandosi su qualsiasi superficie ruvida a disposizione.
Successivamente, compaiono l’alopecia (perdita di peli, non sempre visibile) e
lesioni crostose screpolate e maleodoranti (odore di grasso rancido) a partire dal
muso e dal collo in estensione a tutto il corpo.

34
In questa seconda fase, si manifestano lesioni dermatologiche secondarie a
infezioni microbiche, un calo delle difese immunitarie e un conseguente cambio del
comportamento, che porta l’animale ad isolarsi, ad abbassarsi di quota e a perdere
l’appetito, fino a giungere a morte entro 2-4 mesi dal primo contagio.
a b c
Figura 1.11 Camoscio colpito da rogna sarcoptica (a e b foto di Moma Marzio, c di Luca Rossi)
Non tutti però muoiono, alcuni individui (circa il 10%), dotati di resistenza
genetica all’infezione, riescono a superare la malattia andando incontro a
guarigione spontanea, anche se la risposta immunitaria, seppur presente, non
genera nell’animale sufficiente protezione. Nel caso in cui una popolazione vergine
e numericamente consistente di camosci venga parassitata da questo acaro,
(decorso acuto) la mortalità potrà raggiungere valori molto alti (75 – 90%), come
avvenne in alcune zone del tarvisiano negli anni ’90, e si arresterà solamente
quando la densità della popolazione si aggirerà intorno a 1,5 capi/100 ha (soglia

35
dell’estinzione dell’ospite), esaurendosi in genere in 5-7 anni, spontaneamente. La
malattia, una volta divenuta endemica, acquisisce carattere ciclico e tende a
ripresentarsi ogni 7 -12 anni con virulenza via via sempre più attenuata e tassi di
mortalità sempre più ridotti, che si attestano intorno al 10 - 15%.
La specie Sarcoptes scabiei è divisa in varietà indistinguibili morfologicamente
ma che hanno però un alto grado di specificità verso l’ospite (Berrilli et al. 2002).
Non esiste una vera e propria tassonomia tra i vari tipi di acaro che infettano diversi
ospiti, ma vengono identificate diverse varianti per la specie che infettano in
maniera predominante. Ad esempio, l’acaro che infetta il camoscio viene detto S.
scaibei var. rupicaprae (Pence & Ueckermann 2002).
L’acaro Sarcoptes scabiei (Fig. 1.12) possiede quattro paia di zampe, e riesce ad
invadere gli strati più superficiali dell’epidermide, formando gallerie che si
estendono a volte fino allo strato granuloso e che appaiono in rilievo e grigi per le
feci del parassita che vi si accumulano. Le femmine a temperature superiori ai 20-
25 °C sono particolarmente mobili e sono quindi capaci di penetrare
nell’epidermide in appena 20 minuti, per una profondità di 2,5-3 cm.
L’accoppiamento avviene in superficie; il maschio muore subito dopo senza
penetrare nel derma, le femmine fecondate restano in superficie fino a quando le
uova sono quasi mature. Ciascuna femmina depone dalle due alle quattro uova al
giorno. Dopo circa sette giorni fuoriescono le larve (larve d’acaro); queste ultime
subiscono tre mute prima di trasformarsi in elementi adulti, capaci di accoppiarsi
ed iniziare così di nuovo la deposizione delle uova. Il ciclo vitale è di 6 settimane
(De Carneri 1997). Gli acari sopravvivono per poco tempo al di fuori dell’ospite
naturale; la trasmissione avviene per contatto diretto da animale infettato ad
animale sano, oppure per contatto indiretto in quanto i parassiti possono vivere
per un certo periodo di tempo fuori dai loro ospiti, magari nei luoghi di riposo
(Pence & Ueckermann 2002).

36
Figura 1.12 Femmina di Sarcoptes scabiei e sezione di derma di animale colpito da rogna sarcoptica
La rogna sarcoptica nei camosci delle Alpi venne descritta in letteratura, per la
prima volta, in Austria e nelle Alpi Bavaresi all’inizio del XIX secolo. La malattia si
diffuse più tardi nelle regioni più a sud dell’Austria e dal 1949 raggiunse le Alpi
orientali italiane (Alpi Carniche in Provincia di Udine). Da qui si sviluppò poi, sia
verso est che verso ovest lungo tutte l’arco alpino.
Il primo evento epizootico successivo a quello avvenuto nelle Alpi Carniche si
ebbe nel Sud Tirolo (Provincia di Bolzano) nel 1976 (Gagliardi et al. 1985); la
malattia colpì molto duramente le popolazioni di camoscio e stambecco presenti a
nord del fiume Rienza. Dopo quattro anni, nel 1980, il secondo evento di diffusione
della malattia partì dalla Slovenia e colpì le Alpi Giulie, in Provincia di Udine (Rodolfi
1990; Rossi et al. 1995).
L’epidemia di rogna che ha interessato e sta tuttora colpendo la Provincia di
Belluno è iniziata nel 1995, facendo registrare i primi casi in Val Cengia e Val
Marzòn, a sud delle Tre Cime di Lavaredo (Dolomiti di Sesto). Due anni dopo la

37
malattia si allargò anche verso nord colpendo la Provincia di Bolzano. In seguito,
tutte le Dolomiti bellunesi, dove erano presenti grandi popolazioni di camoscio,
vennero colpite, e nel 2001 la malattia raggiunse anche la Provincia di Trento (Fig.
1.13). Finora, in 10 anni di malattia, sono stati registrati più di 1800 ce (Progetto
Rogna Sarcoptica, 2001-2005). L’evoluzione della malattia dal 1995 al 2005, anno
per anno, è riportata nell’Allegato 1.
Figura 1.13 In colore scuro l’area interessata dall’epidemia di rogna sarcoptica tra il 1976 e il 1996; in colore chiaro l’area colpita fino al 2005
1.3.5 Breve introduzione alla genetica della conservazione
La genetica della conservazione è una disciplina delineatasi all’inizio degli anni
’80 (Schonewald-Cox et al. 1983; Falk & Holsinger 1991; Ellstrand & Elam 1993) e
consiste nell’applicazione delle tecniche genetiche molecolari e della genetica delle
popolazioni per aiutare la gestione e la conservazione della specie come entità
dinamiche capaci di sopravvivere e far fronte ai cambiamenti ambientali.
Comprende principalmente la gestione genetica di piccole popolazioni, la
risoluzione di incertezze tassonomiche, la definizione di un’unità all’interno delle

38
specie e la comprensione della biologia e dell’evoluzione delle specie (Frankham et
al. 2004).
Scendendo più nel particolare questa disciplina si occupa di:
Ridurre il rischio di estinzione minimizzando l’inbreeding
(accoppiamento di consanguinei) che porterebbe alla diminuzione
della fecondità e della sopravvivenza (inbreeding depression);
Studiare la perdita di diversità genetica e quindi dell’abilità ad
evolvere in risposta ai cambiamenti ambientali;
Studiare la frammentazione delle popolazioni e la riduzione del
flusso genico, processi che generalmente aumentano il rischio di
estinzione;
Considerare l’adattamento genetico alla cattività e i suoi effetti
avversi sul successo delle reintroduzioni;
Utilizzare le analisi genetiche molecolari in studi forensi soprattutto
per identificare il bracconaggio ed il commercio di specie protette.
Nell’ultimo secolo, i cambiamenti ambientali associati alle trasformazioni sociali
ed economiche hanno causato distruzione, degradazione e frammentazione di una
gran parte degli habitat naturali. Come conseguenza, i tassi di estinzione sono
destinati ad aumentare considerevolmente. La durata media della vita di una
specie animale nei record fossili, dall’origine all’estinzione, è di circa 1-10 milioni di
anni; per uccelli e mammiferi il tasso di estinzione documentato nel secolo passato
corrisponde alla durata della vita di una specie di 10.000 anni e se il trend continua
diventerà 200-400 anni (Frankham et al. 2004). I piani di conservazione operano a
molti livelli, partendo dagli interi ecosistemi e comunità fino al livello di singoli
individui; ad ognuno di questi livelli le tecniche di genetica molecolare possono
essere utilizzate per valutare meglio i processi e per sviluppare nuove strategie.

39
Approcci di genetica molecolare sono molto utili, ad esempio per identificare
eventuali divisioni biogeografiche che esistono all’interno delle specie, e
proteggere separatamente i diversi gruppi. Inoltre, a livello di individuo le tecniche
di genetica molecolare possono essere usate per dedurre la discendenza in
popolazioni selvatiche e in cattività se vengono campionati tutti i potenziali genitori
e la loro prole. Anche in popolazioni per le quali sono disponibili poche
informazioni, si possono fare inferenze da dati genetici e molecolari riguardo alla
struttura di accoppiamento.
La quantità totale di DNA richiesto per le analisi molecolari può essere minima
e può provenire da peli, siero, pelle mutata, penne, feci, campioni di museo o
archeologici di pelle o osso; possono essere quindi usati metodi non invasivi, che
evitano di disturbare gli animali, essenziale precauzione quando si studiano specie
a rischio (Frankham et al. 2004).
1.3.6 L’importanza della genetica per la conservazione delle popolazioni di
camoscio
Sebbene il camoscio non sia una specie minacciata o in pericolo di estinzione è
comunque elencato nell’appendice n. 3 (1979) della Convenzione per la
Conservazione della Fauna Selvatica e degli Ambienti Naturali Europei, che implica
di seguire particolari misure di protezione per gestire le popolazioni. Il
miglioramento della politica di conservazione delle popolazioni di camoscio viene
riposta sull’identificazione di aree sensibili, e sulla definizione di eventuali strategie
di reintroduzione o traslocazioni di individui. Le popolazioni di camoscio sono in
gran parte soggette a prelievo venatorio e questo le rende una specie a cui
l’opinione pubblica è particolarmente sensibile. Quando inoltre, si presenta una
grave patologia come la rogna sarcoptica, entra in gioco il pericolo di estinzioni
locali e quindi nasce un interrogativo sulla corretta gestione venatoria da attuare.

40
Quindi per rispondere a questi programmi di conservazione è importante usare
anche approcci di genetica molecolare e lavorare a due livelli: la stima della
variabilità genetica delle popolazioni analizzate e relazioni genetiche tra queste
popolazioni, come già accennato nel paragrafo 1.3.1.
In questa tesi, per rispondere alle domande della genetica di conservazione del
camoscio, abbiamo affrontato lo studio di marcatori dei processi adattativi che
vede il complesso maggiore di istocompatibilità quale indicatore ottimale delle
risposte alle pressioni selettive patogeno mediate.
1.4 Obiettivi della ricerca
Questa tesi fa parte di un progetto di ricerca iniziato dal Parco Nazionale delle
Dolomiti Bellunesi che ha lo scopo di valutare la struttura e la variabilità genetica
delle popolazioni di camoscio alpino all’interno di questa area protetta. La presente
ricerca, si pone come obiettivo primario la comprensione delle dinamiche che
agiscono nel mantenimento di una elevata variabilità genetica, utilizzando markes
adattativi, quali il locus MHC, codificante per proteine coinvolte nella risposta
immunitaria. Inoltre, ci si pone lo scopo di cercare di comprendere, attraverso lo
studio dei meccanismi a favore della variabilità genetica, l’evoluzione delle MHC a
livello inter ed intra specifico del genere Rupicapra limitatamente alle specie di
camoscio alpino e pirenaico.
Obiettivi specifici sono pertanto quelli di (1) analizzare la variabilità genetica
nell’MHC di classe II in popolazioni di camoscio caratterizzati da diversa pressione
selettiva da parte di parassiti con diverso effetto patogeno quali gli acari agenti
eziologici della rogna sarcoptica (Sarcoptes scabiei); (2) utilizzare i dati ottenuti
come marcatori del differenziamento genetico tra le diverse popolazioni di

41
camoscio da località alpine diverse; (3) osservare la variabilità genetica tra
popolazioni di camoscio alpino e pirenaico soggette a parassitosi;
I campioni sono stati raccolti in modo da rappresentare meta-popolazioni 1)
provenienti da una zona dove non è nota la presenza di Sarcoptes; 2) affetti da
rogna sarcoptica, con effetto patogeno basso sulla popolazione di camosci e basse
percentuali di mortalità (meta-popolazioni resistenti); 3) pesantemente colpiti da
rogna sarcoptica, con significativo effetto patogeno sulla popolazione di camosci ed
elevate percentuali di mortalità (meta-popolazioni sensibili); 4) altamente sensibili,
ma attualmente in fase di ripresa numerica post-epidemia; in modo tale da poter
confrontare se effettivamente a livello genetico mutazioni abbiano indotto
resistenza a parassitosi del portatore aumentandone la fitness.
I risultati ottenuti sulle popolazioni di camoscio alpino prelevati dalla Provincia
di Belluno sono stati confrontati con i dati rilevati da popolazioni di camoscio alpino
provenienti dalla Provincia di Lecco e con popolazioni di camoscio pirenaico
provenienti dalla regione nord-orientale dei Pirenei spagnoli in modo da analizzare
l’evoluzione delle MHC a livello inter ed intra specifico.

42

43
MATERIALI E METODI
2.1 Campioni analizzati
I campioni delle Alpi Orientali sono stati raccolti grazie alla cooperazione
dei cacciatori e degli Agenti del Corpo Forestale dello Stato, sotto la supervisione
delle amministrazioni locali, come l’Ufficio Caccia e Pesca della Provincia di Belluno
ed il Parco Nazionale delle Dolomiti Bellunesi. La raccolta dei campioni è stata
gestita da una collaborazione tra l’Università degli Studi di Ferrara e l’Università
degli Studi di Torino.
I camosci provenienti dalla Provincia di Lecco sono stati reperiti grazie alla
collaborazione con il Prof. Paolo Lanfranchi dell’Università di Milano sempre in
accordo con i cacciatori e le autorità locali di gestione delle popolazioni di
camoscio.
Per quel che riguarda i campioni di camoscio pirenaico gli individui sono stati
prelevati dalle riserve di caccia nazionali e sono stati fatti pervenire dalla Spagna
grazie ad una collaborazione con il Prof. Jorge Ramon Lopez Olvera dell’Università
Autonoma di Barcellona.
Il numero complessivo di campioni analizzati è di 182.
I campioni provenienti dalla Provincia di Belluno sono stati prelevati durante
l’attività venatoria nelle riserve alpine di caccia. Interessante per questo lavoro è la
disponibilità di due gruppi di campioni che derivano dalla stessa riserva alpina di
caccia. Uno comprende campioni di camoscio prelevati nel 1997/98 e che
corrispondono ad animali trovati morti a causa dell’epidemia di rogna sarcoptica.
L’altro invece è costituito da animali cacciati durante la stagione venatoria del

44
2004/05, e quindi sono animali che hanno resistito all’epidemia oppure sono i figli
dei sopravvissuti.
I campioni provenienti dalla Provincia di Lecco sono stati prelevati durante la
stagione venatoria del 2004. Gli individui sono stati campionati prima e dopo un
evento epidemico di polmonite che ha pesantemente colpito le popolazioni di
camoscio di questa Provincia.
I camosci pirenaici, infine, sono stati prelevati tra il 1991 ed il 2009 durante
diverse stagioni venatorie. Gli individui sono risultati sani o affetti da diverse
patologie o da pestivirus.
La localizzazione geografica ed il numero di campioni delle varie popolazioni è
riportata nelle figure e nelle tabella seguenti. (Fig. 2.1a,b,c; Tab. 2a,b,c)

45
Figura 2.1a Localizzazione geografica dei siti di campionamento di Rupicapra r. rupicapra della Provincia di Belluno

46
Figura 2.1b Localizzazione geografica dei siti di campionamento di Rupicapra r. rupicapra della Provincia di Lecco

47
Figura 2.1c Localizzazione geografica dei siti di campionamento di Rupicapra p. pyrenaica della regione Dei Pirenei spagnoli
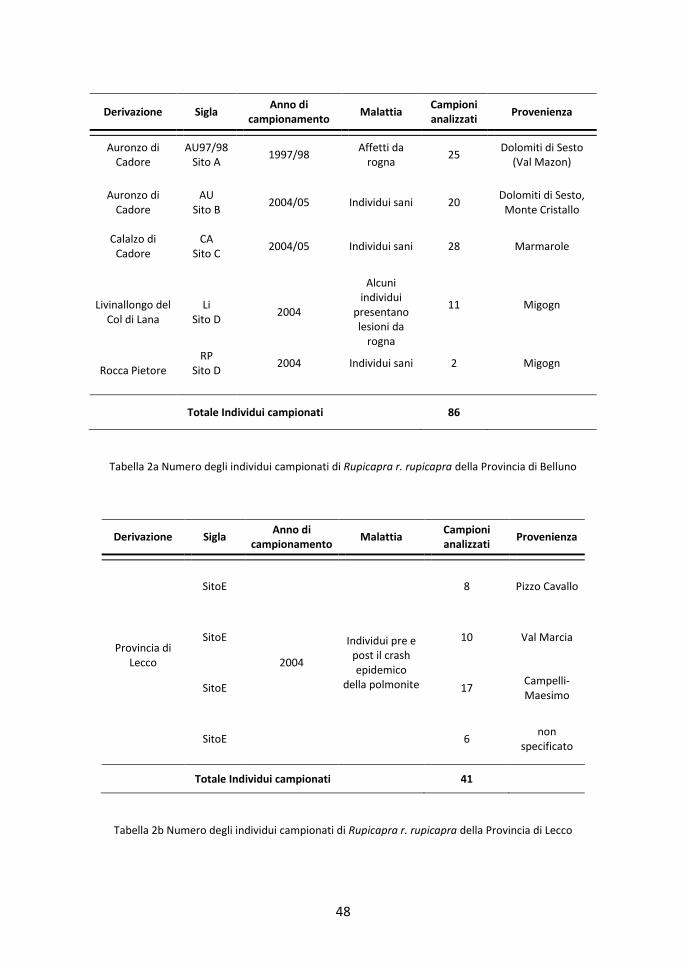
48
Derivazione Sigla Anno di
campionamento Malattia
Campioni analizzati
Provenienza
Auronzo di Cadore
AU97/98 Sito A
1997/98 Affetti da
rogna 25
Dolomiti di Sesto (Val Mazon)
Auronzo di
Cadore
AU
Sito B
2004/05
Individui sani
20
Dolomiti di Sesto,
Monte Cristallo
Calalzo di Cadore
CA
Sito C
2004/05
Individui sani
28
Marmarole
Livinallongo del
Col di Lana
Li
Sito D
2004
Alcuni
individui presentano lesioni da
rogna
11 Migogn
Rocca Pietore
RP Sito D
2004 Individui sani 2 Migogn
Totale Individui campionati 86
Tabella 2a Numero degli individui campionati di Rupicapra r. rupicapra della Provincia di Belluno
Derivazione Sigla Anno di
campionamento Malattia
Campioni analizzati
Provenienza
Provincia di Lecco
SitoE
2004
Individui pre e post il crash epidemico
della polmonite
8 Pizzo Cavallo
SitoE 10 Val Marcia
SitoE 17 Campelli-Maesimo
SitoE 6 non
specificato
Totale Individui campionati 41
Tabella 2b Numero degli individui campionati di Rupicapra r. rupicapra della Provincia di Lecco

49
Derivazione Sigla Anno di
campionamento Malattia
Campioni analizzati
Provenienza
Pirenei Spagnoli Sito 1 1991/2007
Pestivirus ed
altre patologie
17 Cadì, Cadì-Be
Pirenei Spagnoli Sito 2 2001/08
Pestivirus ed
altre patologie
21 APA, PN, Pallars,
Aran
Pirenei Spagnoli Sito 3 1996-2007/08
Individui sani,
affetti da pestivirus e/o da altre patologie
13 Freser
Pirenei Spagnoli Sito 4 2005 Affetti da Pestivirus
4 Cerdanya
Totale Individui campionati 55
Tabella 2c Numero degli individui campionati di camoscio pirenaico della Regione Pirenaica Spagnola nord-orientale
Per maggiori informazioni sui campioni, vedere l’allegato 2.
2.2 L’area di studio
2.2.1 La Provincia di Belluno
La Provincia di Belluno fa parte del Veneto, confina con la Provincia Autonoma
di Bolzano e con l’Austria a nord, le province di Udine e Pordenone ad est, la
Provincia di Treviso a sud, la Provincia di Vicenza e quella autonoma di Trento a
ovest. Con tutte queste entità, la Provincia condivide alcune delle popolazioni di
camoscio.

50
L’idrografia della Provincia è dominata dal fiume Piave, che forma l’omonima
valle, e da tutti i suoi affluenti. Questi creano delle vallate che sono dei fattori di
divisione per le popolazioni di camoscio che trovano difficile attraversarli. L’intera
Provincia è suddivisa in 68 riserve alpine di caccia, corrispondenti
fondamentalmente alla suddivisione politica del territorio in comuni.
Per quanto riguarda il camoscio, la distribuzione nel territorio varia molto,
poiché è una specie con particolari esigenze ecologiche e richiede particolari
caratteristiche ambientali rispetto agli altri ungulati. Il camoscio, infatti, è
distribuito nella parte alta della Provincia, dove l’ambiente è definito esalpico ed
endalpico, riferiti a temperature inferiori ai 13-14°C e a vegetazione di latifoglie e
conifere (Provincia di Belluno e Università di Padova, 2003). Qui, le popolazioni di
camoscio (Fig. 2.2 le varie unità di popolazione) sono demograficamente in declino
a causa dell’epidemia di rogna sarcoptica che le sta colpendo, mentre nel settore
meridionale della Provincia le popolazioni stanno aumentando come quantità di
individui. Le densità dell’ultimo censimento parlano di una media per riserva di 1-
30 capi/100 ha (Provincia di Belluno e Università di Padova, 2003). I campioni da
me analizzati derivano da quattro riserve alpine di caccia del nord della Provincia,
che sono:
Auronzo di Cadore: è la riserva più grande, confinante con la riserva di Calalzo
di Cadore, comprende le unità di popolazione della parte meridionale del gruppo
delle Dolomiti di Sesto, la parte orientale del gruppo del Monte Cristallo, e una
parte del Gruppo delle Marmarole. Da questa riserva sono stati forniti 21 campioni
di camoscio raccolti nella stagione venatoria 2004/05 (sigla AU), mentre sono stati
analizzati anche 26 campioni provenienti da questa riserva ma raccolti nel 1997/98
(sigla AU97/98), anni in cui l’epidemia di rogna sarcoptica ha colpito più duramente
questa zona (Tab. 2.a).

51
Calalzo di Cadore: comprende l’unità di popolazione del Gruppo delle
Marmarole, che condivide con la riserva di Auronzo di Cadore. Di questa riserva
sono stati analizzati 28 campioni di camoscio, 16 prelevati nel 2004 e 12 nel 2005 a
seguito dell’attività venatoria dei cacciatori attivi in questa zona (sigla CA).
Livinallongo del Col di Lana: è la riserva alpina confinante con Rocca Pietore, e
comprende le unità di popolazione del gruppo Fanes-Tofane, del gruppo del Sella e
quello del Viel del Pan-Migogn. Erano disponibili 10 campioni derivanti dall’attività
venatoria 2004 di questa riserva (sigla LI).
Rocca Pietore: è la riserva a confine con Livinallongo del Col di Lana, con cui
condivide le unità di popolazione del gruppo Viel del Pan-Migogn, oltre a
comprendere anche il Gruppo della Marmolada-Monzoni. Di questa riserva è stato
possibile analizzare solo 2 campioni (sigla RP).
Figura 2.2 Distribuzione delle unità di popolazione di camoscio tra le province di Belluno, Bolzano e Trento

52
2.2.2 La Provincia di Lecco
La Provincia di Lecco è situata nella Regione Lombardia e si estende attorno al
ramo orientale del lago di Como. Confina a nord con la Provincia di Sondrio, a nord-
ovest con la Provincia di Como, ad est con la Provincia di Bergamo ed a sud con la
Provincia di Monza-Brianza.
La Provincia di Lecco è quasi interamente collinare e montuosa, come
abbarbicata sulle ripide sponde del lago di Como.
L’aumento delle aree boscate ed arbustive, la riduzione delle aree pascolate ed
aperte di alta quota ha ridotto l’estensione dell’areale disponibile per il camoscio
nella Provincia, soprattutto per le zone a media vocazionalità. Di fatto, però, ad
oggi, la Provincia di Lecco sembra essere, rispetto a quanto rilevato nel 2000 dai
censimenti, la zona in cui la popolazione di camoscio ha la maggior densità
relativamente al suo areale di distribuzione dell’arco alpino (10 capi/Km2).
2.2.3 La Spagna pirenaica
I campioni pervenuti per il presente studio sono stati prelevati da aree incluse
nelle riserve di caccia del National Hunting Riserve dei Pirenei della Catalogna. Le
popolazioni presenti in queste aree sono state pesantemente colpite da epidemie
da pestivirus e da altri tipi di malattie che hanno causato pesanti crolli demografici
delle popolazioni colpite.
2.3 Il campionamento
Per quanto riguarda i camosci derivanti dalle riserve alpine di caccia, i campioni
sono stati raccolti dai cacciatori e dagli Agenti del Corpo Forestale dello Stato,

53
durante la normale attività venatoria ovvero trovati morti a seguito di infestazioni
da rogna sarcoptica. Nel momento in cui un cacciatore ha prelevato un animale, ha
tagliato un frammento di muscolo sottopelle, in genere dalla coscia, di circa 1 cm3 e
lo ha posto in una provetta eppendorf e lo ha conservato poi in congelatore a –20°.
Successivamente, al termine della stagione venatoria, secondo gli accordi presi con
l’Università di Torino e di Ferrara, i frammenti giunti negli Atenei sono stati divisi in
provette più piccole, messi in etanolo assoluto ed assicurato il mantenimento dei
campioni a lungo termine grazie ad un continuo ricambio di etanolo ogni 6 mesi. I
campioni da analizzare mi sono stati fatti pervenire per posta grazie alla
collaborazione con il Prof. Luca Rossi dell’Università di Torino. Una parte dei
campioni, identificati con la sigla AU97/98 sono stati prelevati personalmente dal
Prof. Luca Rossi da animali malati di rogna sarcoptica e trovati morti in Val Marzòn,
nella riserva alpina di caccia di Auronzo di Cadore. Qui si ebbe il primo focolaio
della malattia negli anni 1995/96.
I campioni provenienti dalla Provincia di Lecco, invece, sono stati prelevati
grazie ad una collaborazione con gli enti forestali locali e cacciatori da individui sani
o trovati morti a seguito di una epidemia di polmonite. Anche in questo caso i
campioni mi sono stati fatti pervenire grazie al coordinamento con il Prof. Luca
Rossi ed alla collaborazione con il Prof. Paolo Lanfranchi dell’Università di Milano.
I frammenti di pelle di camoscio pirenaico, sono giunti al Laboratorio di
Parassitologia del Dipartimento di Scienze di Sanità Pubblica della Sapienza
Università di Roma, a seguito di un accordo con il Prof. Jorge Ramon Lopez Olvera
dell’Università Autonoma di Barcellona. Si tratta sempre di frammenti cutanei di
individui cacciati e trovati sani o affetti da pestivirus o da altre parassitosi.

54
2.4 Caratterizzazione molecolare
Come già accennato, nello studio della variabilità genetica e nella genetica
della conservazione, si utilizza un approccio di tipo molecolare. Questo è reso
possibile da una tecnica ampiamente utilizzata in tutte le analisi di laboratorio, la
reazione polimerasica a catena (PCR, Mullis et al. 1986), che permette di copiare ed
amplificare specifiche sequenze di DNA scelto di una lunghezza massima di 1
kilobase. Per usare questo metodo è necessario conoscere la sequenza
fiancheggiante il tratto di DNA scelto che si sceglie di amplificare in quanto le due
sequenze (una all’estremità 5’ del filamento senso e una all’estremità 3’ del
filamento anti-senso) servono da stampo per costruire i primers, che sono degli
oligonucleotidi a singola catena su cui l’enzima deputato alla replicazione del DNA,
la polimerasi, si attacca per copiare il tratto di DNA scelto. La reazione consiste in
cicli ripetuti di denaturazione (DNA melting), appaiamento (DNA annealing) ed
estensione (DNA synthesis), caratterizzati da una specifica temperatura e durata.
Per far questo l’enzima DNA polimerasi ha bisogno di una certa quantità di
ossiribonucleotidi trifosfato (dNTP) e di un buffer di reazione specifico contenente
magnesio.
Reazioni di amplificazione
Per le singole reazioni di PCR sono state utilizzate provette da 200 µl; 4 µl di
DNA genomico di ogni esemplare, sono stati amplificati, ciascuno in 50 µl di volume
finale di reazione, contenente oltre il templato, i due oligonucleotidi che servono
da primers, e definiscono i punti d’inizio della sintesi di DNA, la DNA polimerasi,
Cloruro di Magnesio, un buffer di amplificazione ed infine i quattro nucleotidi
precursori, con le seguenti quantità:
30,5 µl di acqua distillata sterile.

55
5 µl di GeneAmp 10x PCR Buffer II (100mM Tris- HCl, pH 8.3, 500mM
KCl; Applied Biosystems)
5 µl di MgCl2, 25mM (Applied Biosystems)
4 µl della soluzione contenente NTP’s (dCTP, dGTP, dATP, dTTP)
ciascuno alla concentrazione di 2mM. Commercialmente (Promega)
ogni deossinucleoside trifosfato è disponibile in una soluzione
100mM con 10mM di Tris-HCl, pH7.5. Quindi, una volta diluiti, sono
stati suddivisi in aliquote da 500 µl in Eppendorf sterili e conservati a
–20°C;
0.3 µl di primer 1 (forward), 0.5 µM (MWG-Biotech)
0.3 µl di primer 2 (reverse), 0.5 µM (MWG-Biotech)
0.4 µl di AmpliTaq Gold polimerasi, 5U/ µl (Applied Biosystems)
Tutti i reagenti sono conservati a -20°C
2.4.1 Estrazione ed isolamento del DNA
L’estrazione del DNA è stata effettuata su campioni di diaframma e/o di cute
conservati in alcool al 70%. Dai campioni sopra citati sono state prelevate porzioni
della grandezza di circa un 1 cm3, le quali sono state sminuzzate con bisturi e
forbicine ed inserite in provette eppendorf sterili da 1.5 ml. Successivamente i
campioni sono stati posti in azoto liquido per alcuni secondi, per facilitare la rottura
delle membrane cellulari, ed omogeneizzati mediante pestelli sterili.
Wizard Genomic Dna-purification kit (PROMEGA)
Per ogni campione in esame è stato preparato un mix composto da 120 µl di
EDTA 0.5 M (pH 8.0) e 500 ml di Nuclei Lysis Solution in una provetta mantenuta in
ghiaccio per 5 minuti. Di questo mix sono stati prelevati 600 µl e aggiunti ad ogni
campione insieme a 17.5 µl di Proteinasi K 20 mg/ml. I campioni sono stati poi

56
incubati per tre ore a 55°C in un bagnetto termico. Dopo l'incubazione, 3 µl di
RNase Solution sono stati aggiunti al lisato nucleare, i campioni invertiti più volte,
sono stati posti per 30 minuti in una camera termica a 37°C. Dopo 5 minuti, utili per
riportare i campioni a temperature ambiente, sono stati aggiunti 200 µl di Protein
Precipitation Solution per consentire la precipitazione delle proteine e vortexando
vigorosamente ad alta velocità per 20 secondi. Le provette sono state mantenute in
ghiaccio per 5 minuti e sottoposte a centrifugazione per 4 minuti a 14000 giri. Le
proteine precipitate formano un pellet biancastro sul fondo delle provette. Il
surnatante contenente il DNA è stato accuratamente rimosso e trasferito in una
nuova eppendorf da 1.5 ml contenente 600 µl di isopropanolo a temperatura
ambiente. Invertendo delicatamente più volte le eppendorf compare una massa
visibile di DNA. Dopo una seconda centrifugazione a 14000 giri per 1 minuto, il DNA
decantato è riconoscibile come un piccolo pellet bianco. Eliminato il surnatante, al
pellet sono stati aggiunti 600 ml di etanolo al 70% e le provette agitate per
sciacquare il DNA. Dopo questa operazione è stato allontanato l'etanolo e l’eccesso
di alcool è stato fatto evaporare tenendo i tappi delle eppendorf aperti a
temperatura ambiente per circa 45 minuti. Infine il DNA è stato risospeso in 100 µl
di DNA Rehydration Solution ed incubato a 65°C per 1 ora (nel caso in cui si è voluti
procedere subito all’amplificazione del DNA), oppure reidratato incubando la
soluzione per una notte a temperatura ambiente o a +4°C. Il DNA è stato poi
conservato a -20°C.
2.4.2 Amplificazione dell’esone 2 delle MHC di classe II
La regione di nostro interesse è nel genoma nucleare, e in particolare si tratta
della regione dell’esone 2 del gene DRB che deve fungere da marcatore genetico
per studiare il livello di variabilità inter ed intra specifica in relazione a pressioni di
tipo parassitologico. L’amplificazione dell’esone 2 del gene DRB è stata effettuata
utilizzando due primers caratterizzati nei bovini e che hanno già dimostrato la loro
efficacia nel camoscio (Schaschal et al. 2004). Allo scopo, inoltre, di aumentare la

57
specificità dell’amplificazione, una PCR semi-nested con condizioni altamente
stringenti è stata messa a punto utilizzando un primer nested.
Condizioni di amplificazione
La sequenza di DNA dell’esone 2 del gene DRB è stata amplificata utilizzando la
macchina da PCR Perkin Elmer – Gene Amp PCR System 2400, con le seguenti
condizioni:
prima amplificazione - 10 minuti a 95°C (denaturazione iniziale, separazione
dei due filamenti del DNA stampo) seguiti da 10 cicli di 1 minuto a 95°C
(denaturazione completa del DNA), 1 minuto a 60°C (annealing, appaiamento dei
primers alle sequenze bersaglio sul filamento stampo), 1 minuto e 30 sec. a 72°C
(estensione dei primers da parte dalla polimerasi, quindi polimerizzazione), seguiti
da 6 min a 72°C (allungamento finale); primers utilizzati:
HL030 for- 5’-ATCCTCTCTCTGCAGCACATTTCC-3’ ,
HL031 rev- 5’-TTTAAATTCGCGCTCACCTCGCCGCT-3’;
seconda amplificazione - 10 minuti a 95°C, seguiti da 25 cicli di 1 minuto a
95°C, 30 sec. a 65°C, 1 minuto a 72°C, seguiti da 10 min a 72°C; primers utilizzati:
HL030 for- 5’-ATCCTCTCTCTGCAGCACATTTCC-3’ ,
HL032 rev- 5’-TCGCCGCTGCACAGTGAAACTCTC-3’.
Elettroforesi su gel
Terminata la seconda reazione di amplificazione (Fig.2.3), i prodotti di
amplificazione di ciascun campione sono stati sottoposti a corsa elettroforetica su
gel d'agarosio. Il gel è stato ottenuto sciogliendo agarosio all'1% in tampone TBE

58
0.5X. A questa soluzione è stato aggiunto bromuro di etidio, 2 µl alla
concentrazione di 0,5 µg/ml. La miscela è stata versata in una vaschetta per
elettroforesi al cui apice è stato posto un pettinino con lo scopo di generare nel gel
dei pozzetti (in numero di 15). Si è atteso circa 1 ora per consentire la gelificazione
della soluzione. Rimosso il pettinino sono stati caricati nei pozzetti i prodotti di
amplificazione.
Figura 2.3 Risultato della seconda amplificazione visualizzato su gel di agarosio.
Il bromuro di etidio è il colorante più usato per la colorazione del DNA poiché si
intercala tra le coppie di basi nucleotidiche e quando il gel viene illuminato con luce
ultravioletta il bromuro fa in modo che le bande di DNA siano visibili, rendendole
fluorescenti.
Il DNA migra sul gel nel momento in cui viene applicata una differenza di
potenziale agli elettrodi dell' apparato di elettroforesi perché questa
macromolecola è carica negativamente e quindi migra verso il catodo con una
velocità inversamente proporzionale alle sue dimensioni; in questo modo si
separano i diversi frammenti. Dopo 20 min di corsa a 110V con macchina ESP 200 –
Pharmacia Biotech Electrophoresis Power Supplay, questi frammenti amplificati
500 pb

59
sono stati visualizzati esponendo il gel sotto la luce ultravioletta. Per l’osservazione
e la catalogazione delle immagini ottenute dall’esposizione alla luce ultravioletta è
stato usato il programma MULTI-ANALIST (versione 1.1-BIORAD).
Purificazione dei prodotti di PCR.
I prodotti di PCR sono stati preparati per il sequenziamento mediante
purificazione su colonna usando il kit Nucleo-spin extract 2 in 1 della MACHEREY-
NAGEL. Il buffer NT2 è stato aggiunto a 45 µl del prodotto di PCR e trasferiti nella
colonna di purificazione contenuta in un tubo da 2 ml. Mediante centrifugazione
per 1 minuto a 14000 giri il DNA viene trattenuto dalla colonna; è stato gettato via
il precipitato e, aggiunti 600 µl di buffer NT3 è stata effettuata una nuova
centrifuga di un minuto. Gettato via nuovamente il precipitato e aggiunti altri 200
µl di NT3 il DNA è stato sottoposto ad una nuova centrifuga per 2 minuti. A questo
punto per eliminare un possibile residuo di etanolo (contenuto nell' NT3), che
potrebbe inibire le seguenti reazioni, le provette sono state poste aperte, per 4
minuti, in bagnetto a 70°C in modo da fare evaporare l'etanolo. Dopo questo
tempo le colonne sono state trasferite in eppendorf da 1.5 ml , aggiunti 30
microlitri di elution buffer NE e dopo avere lasciato a temperatura ambiente per un
minuto, una nuova centrifuga di un minuto ha permesso la precipitazione del DNA
purificato.
Sequenziamento
I frammenti di PCR della regione in esame, purificati col metodo sopra
descritto, sono stati sequenziati dall'MWG-BIOTECH che mi hanno inoltrato in
formato elettronico i dati grezzi letti dal sequenziatore, che sono costituiti da 4
serie di picchi di fluorescenza parzialmente risolti, uno per ciascuna delle 4 basi.
Ciascun picco corrisponde al segnale emesso da un frammento di DNA che termina
con un ddNTP marcato con il fluorocromo. Un algoritmo di processamento del
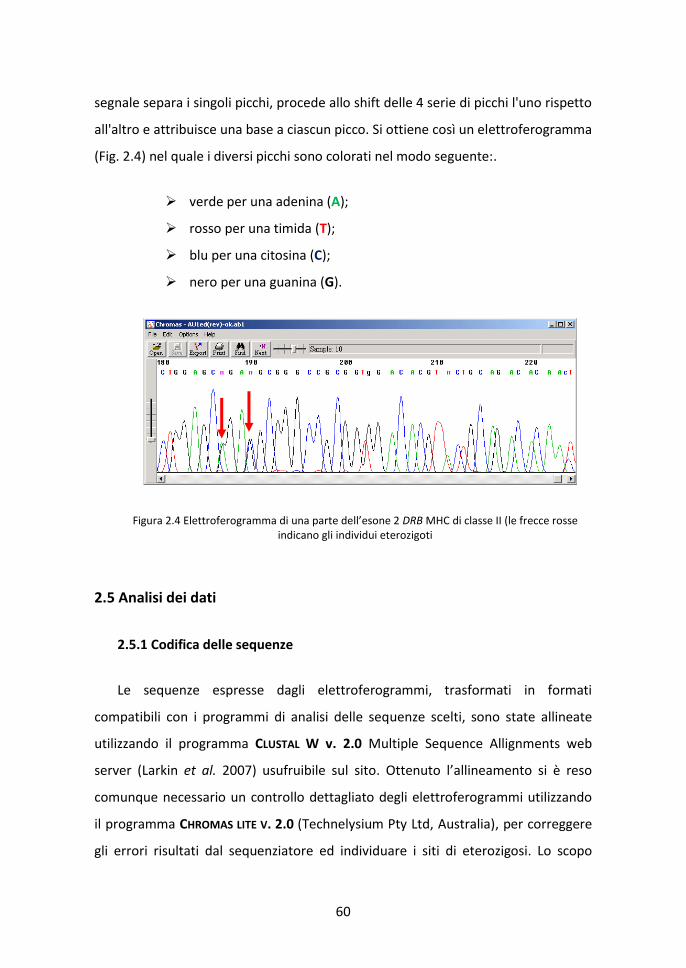
60
segnale separa i singoli picchi, procede allo shift delle 4 serie di picchi l'uno rispetto
all'altro e attribuisce una base a ciascun picco. Si ottiene così un elettroferogramma
(Fig. 2.4) nel quale i diversi picchi sono colorati nel modo seguente:.
verde per una adenina (A);
rosso per una timida (T);
blu per una citosina (C);
nero per una guanina (G).
Figura 2.4 Elettroferogramma di una parte dell’esone 2 DRB MHC di classe II (le frecce rosse indicano gli individui eterozigoti
2.5 Analisi dei dati
2.5.1 Codifica delle sequenze
Le sequenze espresse dagli elettroferogrammi, trasformati in formati
compatibili con i programmi di analisi delle sequenze scelti, sono state allineate
utilizzando il programma CLUSTAL W v. 2.0 Multiple Sequence Allignments web
server (Larkin et al. 2007) usufruibile sul sito. Ottenuto l’allineamento si è reso
comunque necessario un controllo dettagliato degli elettroferogrammi utilizzando
il programma CHROMAS LITE V. 2.0 (Technelysium Pty Ltd, Australia), per correggere
gli errori risultati dal sequenziatore ed individuare i siti di eterozigosi. Lo scopo

61
dell'allineamento è di posizionare i nucleotidi che si ritengono omologhi nella
stessa colonna così da confrontare immediatamente le diverse sequenze ed
evidenziare i siti variabili (Fig. 2.4). Le sequenze in cui si è verificata la presenza di
picchi doppi, sono quelle in cui la sequenza dei due alleli non è conosciuta
("unphased genotype data”) e vengono determinate grazie all’aiuto delle
successive analisi.
2.5.2 Ricostruzione degli aplotipi con metodi statistici
Per determinare le sequenze degli aplotipi di ogni individuo eterozigote è stato
utilizzato il programma HAP (Haplotype Reconstruction using Phylogeny, Halperin
& Eskin 2004) che ipotizza un modello probabilistico per la ricostruzione degli
aplotipi. Il programma genera aplotipi doppi (diplotipi) per ogni filamento di DNA,
per cui si otterrà un numero doppio di sequenze rispetto agli individui analizzati.
Allineamento degli aplotipi ricostruiti e confronti con quelli in letteratura
Gli aplotipi ricostruiti con il software HAP (Halperin & Eskin 2004) sono stati
analizzati mediante DnaSP v. 5 (Librado & Rozas 2009) in modo da evidenziare
eventuali corrispondenze tra gli aplotipi trovati nelle popolazioni di camoscio alpino
e pirenaico, ma anche per evidenziare un eventuale variabilità genetica inter e tra
popolazioni di camoscio. DnaSP v. 5 quantifica il numero dei siti variabili e delle
mutazioni in corrispondenza del frammento di DNA analizzato, la diversità
nucleotidica ed aplotipica, la quantità in GC, la frequenza delle sostituzioni
sinonime e non sinonime.
Gli aplotipi una volta ricostruiti sono stati allineati con le sequenze presenti in
letteratura e pubblicate da Schaschal nel 2004, 2005 (Ruru-DRB*01 - 40 per quanto
riguarda il camoscio alpino e Rupy-DRB*01 – 13 per la specie Rupicapra pyrenaica).
Diversità genetica entro popolazioni

62
I classici parametri genetici di popolazione per la diversità genetica vengono
stimati per ogni popolazione usando il software ARLEQUIN v. 3.1 (Excoffier et al
2005). Questi parametri sono:
le frequenze alleliche
l’eterozigosità osservata (HO)
l’eterozigosità attesa (HE).
Diversità genetica tra popolazioni
1. Analisi basate sui dati genotipici degli individui. La differenziazione genetica
tra popolazioni è stata testata usando il “test esatto di differenziazione dei
campioni basato sulla frequenza allelica” (Raymond & Rousset 1995), integrato da
ARLEQUIN v. 3.1. È stato anche possibile calcolare l’indice di fissazione FST (Wright
1951) che quantifica la diversità delle popolazioni. Usando questo software FST è
stimato con un’analisi della varianza (Weir & Cockerham 1984).
FST indica la frazione della variabilità totale dovuta alle differenze tra le
popolazioni. Il valore di tale indice varia tra 0 ed 1: più tale valore si avvicina ad 1
più le popolazioni sono diverse tra loro.
FST è definito anche come la devianza standardizzata delle frequenze alleliche
delle popolazioni in esame dalla frequenza allelica media, che dovrebbe
rispecchiare la frequenza allelica della popolazione ancestrale da cui queste hanno
origine. FST misura quindi la correlazione tra coppie di geni omologhi all’interno
delle sottopopolazioni, in relazione a coppie di geni presi a caso dalla popolazione
totale.
2. Analisi basate sui dati aplotipici degli individui. L’analisi della varianza
molecolare (AMOVA) per definire la struttura genetica del campione è stata
condotta utilizzando il software ARLEQUIN v. 3.1. Per applicare questo tipo di

63
analisi è necessario strutturare gli individui da analizzare in popolazioni e gruppi di
ordine gerarchico superiore. Nel nostro caso sono stati applicati due criteri di
divisione: il primo criterio è stato quello geografico, ovvero si sono divisi i campioni
in due gruppi principali (camosci italiani appartenenti alla specie R. r. rupicapra e
camosci spagnoli appartenenti alla specie R. p. pyrenaica) e all’interno di ogni
gruppo è stata fatta un’ulteriore divisione in popolazioni a seconda della località di
provenienza; il secondo criterio di divisione è stato quello di considerare gruppi di
camosci sottoposti a pressione selettiva da patogeno. Gli individui delle popolazioni
di camosci spagnoli sono stati suddivisi in ndividui sani ed affetti da pestivirus e/o
da altre patologie, mentre gli individui delle popolazioni italiane sono stati suddivisi
in sani, morti per rogna sarcoptica, resistenti alla rogna perché individui
sopravvissuti ad un evento epidemico oppure perché appartenenti a popolazioni di
figli di individui precedentemente affetti, inoltre una ulteriore suddivisione è stata
fatta per le popolazione di camoscio alpino analizzati prima e dopo il crash
epidemico della polmonite.
Questo tipo di approccio gerarchico per l’analisi della varianza molecolare
consente di dividere la varianza totale in componenti di covarianza, dovute alle
differenze intra-individuali, intra-popolazione e inter-gruppi. Da un confronto tra
queste quantità è possibile stimare due componenti della varianza, che
rappresentano l’effetto della variabilità interna (varintra) e della suddivisione in
popolazioni (vartra). La percentuale di varianza interna alle popolazioni e tra le
popolazioni è quindi calcolata come:
% di varianza entro popolazioni = varintra / vartot
% di varianza tra popolazioni = vartra / vartot

64
Questo tipo di analisi essenzialmente è simile agli altri tipi di approcci basati
sull’analisi della varianza della frequenza genica, ma tiene in conto il numero delle
mutazioni fra gli aplotipi (Excoffier et al 2005).
La significatività degli indici di fissazione è valutata usando l’approccio di
permutazione non parametrica (1000 repliche).
Alberi Neighbor-Joining
Uno dei metodi classici per analizzare le relazioni tra popolazioni, espressa in
forma di matrici di distanze, è rappresentato dai cladogrammi o alberi. Si tratta di
un metodo descrittivo che permette di visualizzare i rapporti di somiglianza
esistenti e che rappresenta anche i successivi eventi di divergenza tra gruppi.
Utilizzando l’algoritmo di Neighbor-Joining (Saitou & Nei 1987) costruisce un
albero per gli alleli MHC (o per gli aplotipi) basato sulle sequenze nucleotidiche,
mediante l’utilizzo del programma MEGA v. 4.0 (Tamura et al. 2007). La robustezza
dell’albero NJ ottenuto è stata testata mediante una tecnica statistica di
ricampionamento casuale con rimpiazzo nota con il termine di bootstrap 1000
replicati. Gli aplotipi presenti in letteratura sono stati inclusi in questa analisi.

65
RISULTATI
La caratterizzazione genetica dell’esone 2 del gene DRB del complesso
maggiore di istocompatibilità (MHC) di classe II è stata eseguita su 182 campioni; le
sequenze ottenute sono state 86 per i campioni di camoscio alpino provenienti
dalla Provincia di Belluno ed esposti alla rogna sarcoptica, 41 provenienti dalla
Provincia di Lecco ed esposti alla polmonite e 55 campioni di camoscio pirenaico
provenienti dal versante spagnolo dei Pirenei orientali. I prodotti di amplificazione,
ottenuti utilizzando i primers HL030 ed HL031 ed il nested primer HL032, hanno
fornito un frammento costituito da circa 300 paia di basi, in cui è inclusa la regione
dell’esone 2 del DRB dell’MHC di classe II.
3.1 Polimorfismo dell’esone 2 delle MHC di classe locus DRB
Gli ampliconi, una volta sequenziati, sono stati allineati mediante CLUSTAL W,
implementato in MEGA v. 4.0. L’allineamento è stato effettuato separatamente per
le specie di camoscio analizzate. L’allineamento ottenuto (Allegato 3a e 3b),
modificato manualmente per migliorare l’omologia delle regioni meno conservate,
è costituito da sequenze complessivamente di 206 nucleotidi di lunghezza.
L’allineamento delle sequenze di R. r. rupicapra ha evidenziato 31 siti polimorfici
(vedi Fig. 3.1 a e b per l’allineamento dei soli siti variabili), mentre in R. pyrenaica è
stato possibile evidenziare 24 siti polimorfici. Negli allineamenti sono state
comprese anche le sequenze di camoscio, alpino e pirenaico, disponibili in
GenBank. Osservando le singole sequenze e i singoli elettroferogrammi è stato
possibile identificare, in corrispondenza dei siti polimorfici, dei picchi indicativi di
eterozigosi. Abbiamo, perciò, utilizzato i seguenti codici IUPAC per evidenziare i siti
eterozigoti: R (A/G), Y (C/T), M (A/C), S (C/G) e W (A/T).

66
111111 1111111111 1
11155578 8899012334 4445557889 9
7845628906 7907469013 5681463581 7
Ruru-DRB*01 CAGCGTGACT GTGGAACGAC ACGGCCAGGG C
Ruru-DRB*02 ......T... .......... ......G... .
Ruru-DRB*03 AC........ .......... GA..T..... .
Ruru-DRB*04 AG........ .....T.... GA..T...A. .
Ruru-DRB*05 .G......T. .....TT... GA..T...A. .
Ruru-DRB*06 .......... .....T.... GA..T...A. .
Ruru-DRB*07 .......... C....T.... ........A. .
Ruru-DRB*08 .......... .......... ........A. .
Ruru-DRB*09 .G...C..TA CC.A...... GA..T..A.. .
Ruru-DRB*10 .G......TA CC.A...... GA..T...A. .
Ruru-DRB*11 AG.....GTA CCCAG....G GAC....AC. .
Ruru-DRB*12 AG......TA CC.A.....G GAA....ACA .
Ruru-DRB*13 AG......TA CC.A.....G GA.....AC. .
Ruru-DRB*14 AG..A...TA CG.A.....G GA.C...AC. .
Ruru-DRB*15 ........TA CC.A...... GA..T..AC. .
Ruru-DRB*16 A.......T. ...A...... GA..T..A.. .
Ruru-DRB*17 .......... C..A...... G......A.. A
Ruru-DRB*18 AG........ .......... .......AC. .
Ruru-DRB*19 ACAA...... .......... .......AC. .
Ruru-DRB*20 .....C..TA CC.A...... GA..T..A.. .
Ruru-DRB*21 ........TA CC.A...... .......AC. .
Ruru-DRB*22 ........T. C..A...... G......A.. A
Ruru-DRB*23 .......... C......... G....G.... .
Ruru-DRB*24 .......... C......... G....G.... .
Ruru-DRB*25 .......... .......... ....T..... .
Ruru-DRB*26 .......... .......... .......AC. .
Ruru-DRB*27 .......... .........G GA........ .
Ruru-DRB*28 .G......TA CC.A...... GA..T..AC. .
Ruru-DRB*29 .G........ .......... .......AC. .
Ruru-DRB*30 .G........ .....T.... GA..T...A. .
Ruru-DRB*31 .G........ .....T.... G...T...A. .
Ruru-DRB*32 A.......TA CC.A.....G GA.....AC. .
Ruru-DRB*33 A.AA...... .......... .......AC. .
Ruru-DRB*34 ACAA...... .......... .......A.. .
Ruru-DRB*35 AG...C..TA CC.A...... GA..T..A.. .
Ruru-DRB*36 AG...C..TA CC.A...... GA..T..... .
Ruru-DRB*37 AG...C..TA CC.A...... ....T..A.. .
Ruru-DRB*38 AG......TA CC.A.....G GA.....AC. .
Ruru-DRB*39 AG........ .......... .......... .
Ruru-DRB*40 AG........ .....T.... GA......A. .
1SUSCET .......... .......... .......... .
0)SUSCET MMRM...... .......... .......RS. .
0+SUSCETT MSRM.Y..YW SY.R...... RM..Y..AS. .
122SUSCET .......... .......... .......... .
123SUSCET .......... .......... R......... .
128SUSCET .......... .......... .......... .
129SUSCET MR........ .....T.... GM..Y...R. .
130SUSCET .......... .........S RM........ .
132SUSCET M.RM...... .......... .......RS. .
135SUSCET MR........ .......... .......RS. .
190897SUS MR........ .....T.... RM..Y...R. .
2aaf98SUS .......... .......... .......... .
2aam97SUS .......... .......... .......... .
3aaf97SUS .......... .........S RM........ .
3aaf98SUS .......... .........S RM........ .
3aam97SUS .......... S..A...... G......A.. A
3aam98SUS MGRM...... .....W.... RM..Y..RY. .
4aaf98SUS M.RA...... .......... .......RS. .
5aaf98SUS MMRM...... .......... .......AS. .
5aam97SUS .......... .......... .......... .

67
6aaSUSCET MR........ .......... .......AS. .
7aaf98SUS .R...Y..YW SY.R...... RM..Y..R.. .
9aam97SUS .......... .......... .......... .
9mSUSCETT ASRM...... .......... .......AC. .
AU1SOPRAV MMRM...... .......... .......RS. .
AU2SOPRAV MMRM...... .......... .......RS. .
AU3SOPRAV .......... .......... .......... .
CA1RESIST MSRM.Y..YW SY.R...... RM..Y..AS. .
CA10RESIS .R...Y..YW SY.R...... RM..Y..R.. .
CA11RESIS .......... .......... .......... .
CA12RESIS MMRM...... .......... .......AS. .
CA13RESIS .R...Y..YW SY.R...... RM..Y..R.. .
CA14RESIS .R...Y..YW SY.R...... RM..Y..R.. .
CA15RESIS .......... .......... .......... .
CA16RESIS MR........ .....W.... RM..Y...R. .
CA19RESIS MMRM...... .......... .......RS. .
CA2RESIST MG...Y..YW SY.R.W.... GA..T..RR. .
CA20RESIS .......... .......... .......... .
CA21RESIS .......... .......... .......... .
CA22RESIS .R...Y..YW SY.R...... RM..Y..R.. .
CA23RESIS .R...Y..YW SY.R...... RM..Y..R.. .
CA24RESIS .R...Y..YW SY.R...... RM..Y..R.. .
CA25RESIS MR......YW SY.R.....S RM.....AS. .
CA26RESIS .......... S..R...... R......R.. M
CA27RESIS MSRM.Y..YW SY.R...... RM..Y..AS. .
CA28RESIS MR........ .....W.... RM..Y...R. .
CA29RESIS ASRM...... .....W.... RM..Y..RM. .
CA3RESIST .R...Y..YW SY.R...... RM..Y..R.. .
CA30RESIS MR........ .....W.... RM..Y...R. .
CA4RESIST .R...Y..YW SY.R...... RM..Y..R.. .
CA5RESIST MR........ .....W.... RM..Y...R. .
CA6RESIST .R......YW SY.R...... RM..Y..RS. .
CA8RESIST .R......YW SY.R...... RM..Y..RS. .
CA7RESIST ACAA...... .......... .......AC. .
CA9RESIST MG...Y..YW SY.R.W.... GA..T..RR. .
Li1RESIST MMRM...... .......... .......RS. .
Li13RESIS MR......YW SY.R.....S RM.....RS. .
Li10RESIS .......... .......... .......... .
Li11RESIS .R...Y..YW SY.R...... RM..Y..R.. .
Li14RESIS .......... .......... .......... .
Li2RESIST MG...Y..YW SY.R...... RM..Y..AS. .
Li5RESIST .......... .......... .......... .
Li6RESIST .R...Y..YW RY.R...... RM..Y..R.. .
Li7RESIST ASRM...... .....W.... RM..Y..RM. .
Li8RESIST .......... S..R...... R......R.. M
Li9RESIST .......... .......... .......... .
NSSUSCETT .......... .......... .......... .
RP1RESIST MG...Y..TA CC.A.....S GA..Y..AS. .
RP4RESIST .......... .......... .......... .
Au22RESIS MR........ .......... .......RS. .
Au23RESIS MR.Y...... .......... .......RS. .
Au24RESIS .......... .......... .......... .
Au25RESIS .M...Y..YW SY.R...... RM..Y..R.. .
Au26RESIS .......... .......... .......... .
Au27RESIS .R...Y..YW SY.R...... RM..Y..R.. .
Au28RESIS M.RM...... .......... .......RS. .
Au29RESIS .......... .......... .......... .
Au30RESIS .......... .......... .......... .
Au31RESIS .......... .......... .......... .
Au32RESIS MR........ .....W.... RM..Y...R. .
Au33RESIS AG........ .....T.... GA..T...A. .
Au34RESIS .......... S..R...... R......R.. M
Au35RESIS .......... S..R...... R......R.. M
Au36RESIS .......... .......... .......... .
68
Au37RESIS .R...Y..TA CC.A...... RM..Y..AS. .
Au38RESIS .......... S..R...... R......R.. M
4359cavPOST MSRA.?..YW SY.R...... RM..Y..AS. .
4360cavaPRE MR......YW SW.R.....S RS.....RS. .
4373cavPOST .......... .......... .......... .
4402cavaPRE AG...Y..TA CC.A.....S GA..Y..AS. .
4403cavaPRE .......... .......... .......... .
4405cavaPRE ........YW SY.R...... .......RS. .
4409cavPOST .......... .......... .......... .
4410cavaPRE MS......YW SY.R.....S RM.....RS. .
4486marPOST .......... .......... .......... .
4487marPOST MR........ .......... .......RS. .
4488marCPRE .......... .......... .......... .
4489marPOST MR........ .....W.... RM..Y...R. .
4493marPOST .......... .......... .......... .
4503marPOST .R...Y..YW SY.R...... RM..Y..R.. .
4504marPOST .......... .......... .......... .
4505marPOST MR........ .......... .......RS. .
4506marcPRE .......... .......... .......... .
4507marCPRE .......... .......... .......... .
4542campPOS .......... .......... .......... .
4544campPOS .......... .......... .......... .
4547campPOS .......... .......... .......... .
4548campPOS .......... .......... .......... .
4553campPOS MR......YW SY.R.....S RM.....RS. .
4554campPOS .......... .......... .......... .
4555campPOS .......... .......... .......... .
4556campPOS AG...Y..TA CC.A.....S GA..Y..AS. .
4557campPOS .R...Y..YW SY.R...... RM..Y..A.. .
4562campPRE .......... .......... .......... .
4564campPOS .......... .......... .......... .
4564age3camPO .......... .......... .......... .
4571campPOS .......... .......AG. .......... .
4572mcamPRE MR...Y..YW SY.R...... RM..Y..S.. .
4572fcamPOS MR...Y..YW SY.R...... RM..Y..R.. .
4573campPRE .......... .......... .......... .
4575campPRE ........YW SY.R...... .......RS. .
PN62POSTOUT MMRM...... .......... .......RS. .
PN81POSTOUT AG...C..TA CC.A...... GA..T..A.. .
PN123OUTBRE .......... .......... .......... .
PN323OUTBRE .......... .......... .......... .
PN331OUTBRE .......... .......... .......... .
PP1355OUTBR MR......YW SY.R.....S RM.....RS. .
Figura 3.1 a Siti variabili presenti nell’allineamento dei campioni italiani a confronto con le sequenze depositate in GenBank di R. rupicapra (Schaschal et al 2004). Si evidenziano le sequenze omozigoti.

69
11111111 1111
11567888 8914445588 8999
7845250678 9763564656 8017
Rupy-DRB1*01A CAGCTACTCG TAACACCCAG GCGA
Rupy-DRB1*02A ........G. .G......G. ...C
Rupy-DRB1*03A AG..C.TA.. C......... ....
Rupy-DRB1*04A AG....TA.. C..GGA.... C..C
Rupy-DRB1*05A ACAA...... .G.GGA.... C...
Rupy-DRB1*06A .....TTA.C ....GA.... C..C
Rupy-DRB1*07A ACAA...... .G..GAT.G. C...
Rupy-DRB1*08A AG..C.TA.. C...GAT... ...C
Rupy-DRB1*09A AG........ .GT.G..GG. ....
Rupy-DRB1*10A ........G. .G......GA ...C
Rupy-DRB1*11A AG..C.TA.. C......... ...C
Rupy-DRB1*12A .......... .G..G..GG. ...C
Rupy-DRB1*13A ......TA.. C...GAT... C..C
16.06.HL032 .G........ .......... ...C
15.06 .G....T.G. ........R. ...C
18.06 .G....T.G. ........R. ...C
RP02.08 ........G. .G......G. ...C
RP2.96 ........G. ...G....R. S..C
RP03.04 .G....T.G. ........R. ...C
RP3.08 ........G. .G......G. ...C
RP3.99 AG..C.TA.. C......... ...C
RP4.03 ........G. .G......G. ...C
RP05.05 ........G. .G......G. ...C
RP5.99 ........G. .G......G. ...C
RP06.05 ........G. .G......G. ...C
RP06.07 AG..C.TA.. C......... ...C
38.07 MR..Y.YWS. YR......R. ...C
12.07 ........G. .G......G. ...C
RP33.07 AG..C.TA.. C......... ...C
RP34.07 AG..C.TA.. C......... ...C
RP35.07 MR....YWS. YR.SRM..R. S..C
RP36.07 MR....YWS. YR.SRM..R. SGCC
RP37.07 ........G. .G......G. ...C
RP38.06 ........G. .G......G. ...C
RP48.06 MR....YWS. YR.SRM..R. ...C
RP7.01 ........G. .G......G. ...C
RP10.02 AG..C.TA.. C......... ...C
RP10.03 MR..Y.YWS. YR......R. ...C
RP12.08 ........G. .G......G. ...C
RP13.02 ........G. .G......G. ...C
RP13.03 MR..Y.YWS. YR......G. ...C
RP13.07 MR..Y.YWS. YR......R. ...C
RP14.02 ........G. .G......G. ...C
RP15.02 MR..Y.YWS. YR......R. ...C
RP4.HL032 AG..Y.TA.. C..SRM.... S..C
RP23.HL032 AG..Y.TA.. C..SRM.... S..C
RPC20.HL032 ........G. .G......G. ...C
RP708.HL032 ........G. .G......G. ...C
RP808.HL032 MR....YWS. YR.SRM..R. S..C
RP09.03 MR..Y.YWS. YR......R. ...C
RPC8 MR....YWS. YR.SRM..R. S..C
RP19.06 ----Y.YWS. YR......R. ...C
RP22.07 ----....G. .G......G. ...C
35.05 ........G. .G......G. ...C
26.05 ........G. .G......G. ...C
1.04 ........G. .G......G. ...C
39.06 AG..C.TA.. C......... ...C
12.05 AG..C.TA.. C......... ...C
15.02 MR..Y.YWS. YR......R. ...C
30.05 AG..Y.TA.. C..SRM.... S..C

70
01.09 ........G. .G......G. ...C
19.06 MR..Y.YWS. YR......R. ...C
05.06 MR..Y.YWS. YR......R. ...C
12.06 ........G. .G......G. ...C
49.05 ........G. .G......G. ...C
04.04 MR..Y.YWS. YR......R. ...C
05.04 ........G. .G......G. ...C
06.05 MR..Y.YWS. YR......R. ...C
Figura 3.1 b Siti variabili emersi dal confronto delle sequenze dei campioni spagnoli e di quelle depositate in GenBank per R. pyrenaica. Evidenziate in grigio, le 31 sequenze in omozigosi.
Tra le 127 sequenze analizzate di camoscio alpino, le sequenze omozigoti
risultano essere 46 (sequenze evidenziate nella Fig.3.1a).
Dei 31 siti variabili, il sito in posizione 7 risulta polimorfico in 42 sequenze, otto
sequenze, inoltre, hanno una adenina in tale sito invece della citosina. In posizione
8, risultano polimorfiche 55 sequenze, di cui: 9 mostrano eterozigosi tipo M (A/C)
ed 8 mostrano eterozigosi tipo S (C/G), 38 sequenze invece mostrano eterozigosi
tipo R (A/G). Le posizioni 14 e 15 sono polimorfiche, entrambe i siti sono
contemporaneamente polimorfici (RM) in 17 sequenze. Nel sito 14 abbiamo
eterozigosi adenina-guanina e nel sito 15 abbiamo eterozigosi adenina-citosina. In
posizione 52, 26 sequenze rivelano eterozigosi timina-citosina, una sola sequenza
ha in questa posizione una citosina invece di una timina. In posizione 70, 32
sequenze rilevano eterozigosi timina-citosina, in questo sito variabile, 5 sequenze
hanno la timina, tutte le altre presentano la citosina. 33 sequenze rivelano
eterozigosi contemporaneamente nelle posizioni 86, 87, 89, e 97 rispettivamente
per i nucleotidi adenina-timina, guanina-citosina, citosina-timina, guanina-adenina;
contemporaneamente nelle posizione 87 e 97 altre 5 sequenze mostrano
eterozigosi tipo S (C/G) e R (A/G). La posizione 116 rivela eterozigosi timina-adenina
in 11 campioni, solo 3 sequenze hanno in questa posizione una timina, tutte le altre
hanno l’adenina. Per le posizioni 130 e 131 solo una sequenza presenta in
entrambe i siti una variazione, una adenina al posto di una guanina ed una guanina
al posto di una adenina, rispettivamente. I siti 143, 145, 146 e 154 risultano

71
variabili. In particolare 8 sequenze mostrano contemporaneamente eterozigosi tipo
S (C/G), R (A/G), ed M (A/C) in 143, 145 e 146; nei siti 145, 146 e 154, 35 sequenze
presentano contemporaneamente eterozigosi tipo R (A/G), M (A/C) ed Y (C/T). 6
campioni hanno eterozigosi in posizione 143. I siti 185 e 188 sono polimorfici. In
entrambe i siti 185 e 188, 23 sequenze sono eterozigoti tipo R (A/G) per la
posizione 185 ed S (C/G) per la posizione 188; il sito 188 risulta molto variabile
rivelando per 36 volte eterozigosi tipo S (C/G), 9 volte mostra eterozigosi tipo R
(A/G), una volta mostra eterozigosi tipo M (A/C) ed una volta tipo Y (C/T). In
posizione 197, 5 sequenze mostrano eterozigosi citosina-adenina, una sola
sequenza mostra una adenina tutte le altre hanno la citosina.
I siti 16, 58, 59, 90, 104 e 129 risultano variabili solo nelle sequenze scaricate da
GenBank ed allineate con i camosci alpini di questa ricerca. Perciò i siti
effettivamente variabili del nostro campione sono 25 (12,1% su 206 nucleotidi).
Le 55 sequenze di camoscio pirenaico, mostrano in totale 24 siti variabili. Le
sequenze omologhe sono in numero di 31 (56,4% ).
Nello specifico, nelle posizioni 7 ed 8, 16 sequenze mostrano eterozigosi per
entrambe i siti di tipo M (A/C) ed R (A/G) rispettivamente. Inoltre, nella posizione 7,
10 sequenze hanno una adenina al posto di una citosina e nella posizione 8, 14
sequenze hanno una guanina al posto di una adenina. Nel sito 52, 7 sequenze
hanno una citosina al posto di una timina e 15 sequenze mostrano eterozigosi di
tipo Y (C/T). I siti 70, 86, 87, 89 e 97 sono contemporaneamente in eterozigosi di
tipo Y (C/T), W (A/T), S (C/G), Y (C/T) ed R (A/G) in 17 sequenze di cui 16 hanno
anche le eterozigosi nelle posizioni 7 ed 8. Inoltre, nelle posizioni 70, 86 e 89, 10
sequenze hanno contemporaneamente una timina al posto di una citosina, una
adenina al posto di una citosina e una citosina la posto di una timina; nella
posizione 70 altre 3 sequenze hanno una timina in sostituzione della citosina; per i
siti 87, 97 e 185, 23 sequenze hanno contemporaneamente e rispettivamente una

72
guanina al posto di una citosina, una guanina al posto di una adenina ed una
guanina invece di una adenina. Altre 3 sequenze hanno in posizione 70 la
variazione guanina al posto di citosina. I siti variabili 143, 145 e 146 sono
eterozigoti contemporaneamente in 8 sequenze. Si hanno rispettivamente
eterozigosi di tipo S (C/G), R (A/G) ed M (A/C). Il sito 185 e variabile ed in 20
sequenze c’è eterozigosi di tipo R (A/G). 8 sequenze sono eterozigoti di tipo S (C/G)
nella posizione 188. Le posizioni 190 e 191 sono variabili in un solo individuo,
mentre per la posizione 197 i camosci analizzati hanno tutti una citosina invece di
una adenina rispetto alle sequenze di camoscio pirenaico, scaricate ed allineate con
i nostri campioni.
I siti 14, 15, 65, 88, 116, 154, 156 e 186 sono variabili solo nelle sequenze
scaricate da GenBank, per cui i siti variabili effettivi del nostro campione è in
numero di 16 (7.8% su 206 nucleotidi).
I siti variabili in comune tra tutti i campioni analizzati sono 19.
Questo polimorfismo corrisponde, in accordo con la ricostruzione aplotipica di
HAP e con l’analisi aplotipica eseguita con i software DnaSP v. 5 e ARLEQUIN v. 3.1,
a 58 aplotipi (Tab. 3). Di questi, due aplotipi sono condivisi tra le popolazioni di
camosci italiani e spagnoli: Hap_1, che corrisponde agli aplotipi depositati Ruru-
DRB*01, Rupy-DRB*02 e Hap_20 che corrisponde a Rupy-DRB*04, Ruru-DRB*13,
Ruru-DRB*38: Questi aplotipi risultano essere trans-specifici, ovvero presenti in
individui di entrambe le specie. Hap_1 è anche l’aplotipo con la frequenza relativa
più alta ed è quindi l’aplotipo più comune.
Gli aplotipi privati degli individui delle popolazioni di camoscio alpino sono in
numero di 42 mentre 12 sono esclusivi delle popolazioni di camoscio pirenaico.
Nove aplotipi sono specifici per le popolazioni del sito di campionamento indicato
con il codice SitoA (Auronzo 97/98; Hap_5,6,8,11,12,29,30,31 e 34); sei aplotipi

73
(Hap_45-50) sono stati riscontrati nelle popolazioni di Auronzo del 2004/05 (SitoB);
quattro aplotipi (Hap_35-36, Hap_39-40) sono riconducibili alle popolazioni
campionate a Calalzo di Cadore (SitoC); un aplotipo (Hap_41) è specifico per il SitoD
(Livinallongo, Rocca Pietore); otto aplotipi (Hap_51-58) sono specifici per i camosci
alpini campionati nella Provincia di Lecco (SitoE).
In merito ai camosci pirenaici 4 aplotipi (Hap_23, 24-26 e 28) sono specifici
della popolazione del Sito 1 (Cadì e Cadì-Be); 5 aplotipi (Hap_17-18,21-22 e 27)
sono specifici delle popolazioni campionate nel Sito 3 (Freser).
Il valore di diversità aplotipica (Hd) è pari allo 0.684 (varianza 0.00078 e
deviazione standard 0.028); il valore di diversità nucleotidica (Pi) è pari a 0,02475; il
numero medio di differenze nucleotidiche (k) è pari a 5.099.
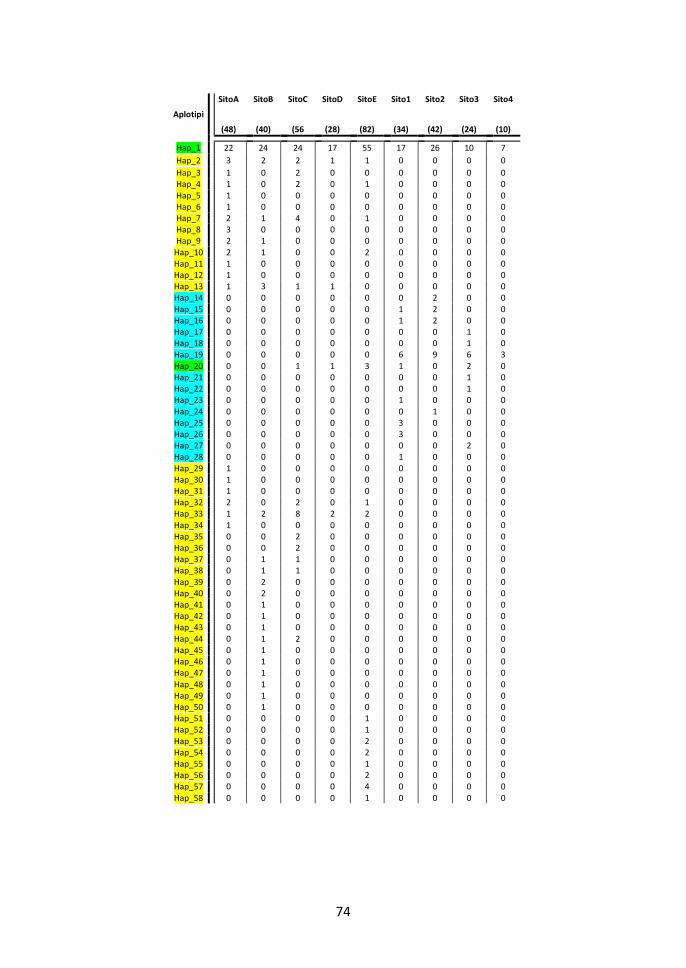
74
Aplotipi
SitoA
(48)
SitoB
(40)
SitoC
(56
SitoD
(28)
SitoE
(82)
Sito1
(34)
Sito2
(42)
Sito3
(24)
Sito4
(10)
Hap_1 22 24 24 17 55 17 26 10 7
Hap_2 3 2 2 1 1 0 0 0 0
Hap_3 1 0 2 0 0 0 0 0 0 Hap_4 1 0 2 0 1 0 0 0 0 Hap_5 1 0 0 0 0 0 0 0 0 Hap_6 1 0 0 0 0 0 0 0 0 Hap_7 2 1 4 0 1 0 0 0 0 Hap_8 3 0 0 0 0 0 0 0 0 Hap_9 2 1 0 0 0 0 0 0 0
Hap_10 2 1 0 0 2 0 0 0 0 Hap_11 1 0 0 0 0 0 0 0 0 Hap_12 1 0 0 0 0 0 0 0 0 Hap_13 1 3 1 1 0 0 0 0 0 Hap_14 0 0 0 0 0 0 2 0 0 Hap_15 0 0 0 0 0 1 2 0 0 Hap_16 0 0 0 0 0 1 2 0 0 Hap_17 0 0 0 0 0 0 0 1 0 Hap_18 0 0 0 0 0 0 0 1 0 Hap_19 0 0 0 0 0 6 9 6 3 Hap_20 0 0 1 1 3 1 0 2 0 Hap_21 0 0 0 0 0 0 0 1 0 Hap_22 0 0 0 0 0 0 0 1 0 Hap_23 0 0 0 0 0 1 0 0 0 Hap_24 0 0 0 0 0 0 1 0 0 Hap_25 0 0 0 0 0 3 0 0 0 Hap_26 0 0 0 0 0 3 0 0 0 Hap_27 0 0 0 0 0 0 0 2 0 Hap_28 0 0 0 0 0 1 0 0 0 Hap_29 1 0 0 0 0 0 0 0 0 Hap_30 1 0 0 0 0 0 0 0 0 Hap_31 1 0 0 0 0 0 0 0 0 Hap_32 2 0 2 0 1 0 0 0 0 Hap_33 1 2 8 2 2 0 0 0 0 Hap_34 1 0 0 0 0 0 0 0 0 Hap_35 0 0 2 0 0 0 0 0 0 Hap_36 0 0 2 0 0 0 0 0 0 Hap_37 0 1 1 0 0 0 0 0 0 Hap_38 0 1 1 0 0 0 0 0 0 Hap_39 0 2 0 0 0 0 0 0 0 Hap_40 0 2 0 0 0 0 0 0 0 Hap_41 0 1 0 0 0 0 0 0 0 Hap_42 0 1 0 0 0 0 0 0 0 Hap_43 0 1 0 0 0 0 0 0 0 Hap_44 0 1 2 0 0 0 0 0 0 Hap_45 0 1 0 0 0 0 0 0 0 Hap_46 0 1 0 0 0 0 0 0 0 Hap_47 0 1 0 0 0 0 0 0 0 Hap_48 0 1 0 0 0 0 0 0 0 Hap_49 0 1 0 0 0 0 0 0 0 Hap_50 0 1 0 0 0 0 0 0 0 Hap_51 0 0 0 0 1 0 0 0 0 Hap_52 0 0 0 0 1 0 0 0 0 Hap_53 0 0 0 0 2 0 0 0 0 Hap_54 0 0 0 0 2 0 0 0 0 Hap_55 0 0 0 0 1 0 0 0 0 Hap_56 0 0 0 0 2 0 0 0 0 Hap_57 0 0 0 0 4 0 0 0 0 Hap_58 0 0 0 0 1 0 0 0 0

75
Tabella 3 Lista delle frequenze degli aplotipi identificati nel presente lavoro di studio. I siti indicati rappresentano le divisioni delle popolazioni degli individui analizzati in base alla provenienza geografica. Gli aplotipi evidenziati in verde sono condivisi da entrambe le specie (R. rupicapra e R. pyrenaica), mentre quelli evidenziati in giallo e in azzurro sono esclusivi rispettivamente per R. rupicapra e per R. pyrenaica. SitoA: AU97/98; SitoB: AU; SitoC: CA; SitoD: LI e RP; SitoE: Provincia di Lecco; Sito1: Cadì e Cadì-Be; Sito2: Aran, Apa, PN, Pallars; Sito3: Freser; Sito4: Cardanya.

76
Lo studio del polimorfismo, oltre che associato ai siti di campionamento, è stato
anche effettuato per evidenziare eventuali corrispondenze tra gli aplotipi e lo
status degli individui, in relazione ad una pressione selettiva da patogeno. Gli
individui sono stati, perciò, raggruppati in popolazioni come specificato nella
Tabella 4.

77
Aplotipi
morti per
rogna
(50)
rogna soprav.
(42)
rogna resist
(82)
polm. preout
(34)
polm. post
(48)
spagna pestvirus
(26)
spagna altre
malattie
(40)
spagna sani
(44)
Hap_1 24 24 39 20 35 15 21 24
Hap_2 3 2 3 1 0 0 0 0
Hap_3 1 0 2 0 0 0 0 0 Hap_4 1 1 2 0 1 0 0 0 Hap_5 1 0 0 0 0 0 0 0 Hap_6 1 0 0 0 0 0 0 0 Hap_7 2 1 0 0 1 0 0 0 Hap_8 3 0 4 0 0 0 0 0 Hap_9 2 1 0 0 0 0 0 0
Hap_10 2 1 0 0 2 0 0 0 Hap_11 1 0 0 0 0 0 0 0 Hap_12 1 0 0 0 0 0 0 0 Hap_13 1 3 2 0 0 0 0 0 Hap_14 0 0 0 0 0 0 2 0 Hap_15 0 0 0 0 0 0 3 0 Hap_16 0 0 0 0 0 0 3 0 Hap_17 0 0 0 0 0 0 1 0 Hap_18 0 0 0 0 0 0 1 0 Hap_19 0 0 0 0 0 10 6 8 Hap_20 0 0 2 1 2 0 0 3 Hap_21 0 0 0 0 0 0 0 1 Hap_22 0 0 0 0 0 0 0 1 Hap_23 0 0 0 0 0 1 0 0 Hap_24 0 0 0 0 0 0 0 1 Hap_25 0 0 0 0 0 0 1 2 Hap_26 0 0 0 0 0 0 1 2 Hap_27 0 0 0 0 0 0 0 2 Hap_28 0 0 0 0 0 0 1 0 Hap_29 1 0 0 0 0 0 0 0 Hap_30 1 0 0 0 0 0 0 0 Hap_31 1 0 0 0 0 0 0 0 Hap_32 2 2 2 1 0 0 0 0 Hap_33 1 0 10 1 1 0 0 0 Hap_34 1 0 0 0 0 0 0 0 Hap_35 0 0 2 0 0 0 0 0 Hap_36 0 0 2 0 0 0 0 0 Hap_37 0 0 2 0 0 0 0 0 Hap_38 0 0 2 0 0 0 0 0 Hap_39 0 0 2 0 0 0 0 0 Hap_40 0 0 2 1 1 0 0 0 Hap_41 0 0 1 0 0 0 0 0 Hap_42 0 0 1 0 0 0 0 0 Hap_43 0 0 1 0 0 0 0 0 Hap_44 0 0 1 0 0 0 0 0 Hap_45 0 1 0 0 0 0 0 0 Hap_46 0 1 0 0 0 0 0 0 Hap_47 0 1 0 0 0 0 0 0 Hap_48 0 1 0 0 0 0 0 0 Hap_49 0 1 0 0 0 0 0 0 Hap_50 0 1 0 0 0 0 0 0 Hap_51 0 1 0 0 1 0 0 0 Hap_52 0 0 0 1 0 0 0 0 Hap_53 0 0 0 1 1 0 0 0 Hap_54 0 0 0 2 0 0 0 0 Hap_55 0 0 0 1 0 0 0 0 Hap_56 0 0 0 0 2 0 0 0 Hap_57 0 0 0 4 0 0 0 0 Hap_58 0 0 0 0 1 0 0 0

78
Tabella 4 Lista delle frequenze degli aplotipi riscontrati nei gruppi di individui di camoscio alpino identificati come: morti per rogna, sopravvissuti e resistenti all’epidemia (quindi sani), individui prelevati prima e dopo il crash epidemico causato da una epidemia di polmonite nella Provincia di Lecco. I gruppi di individui di camoscio pirenaico identificano i seguenti status: affetti da pesti virus, malati di altre parassitosi, individui sani. Gli aplotipi evidenziati in verde sono condivisi da entrambe le specie (R. r. rupicapra e R. p. pyrenaica), mentre quelli evidenziati in giallo e in azzurro sono esclusivi rispettivamente per R. r. rupicapra e per R. p. pyrenaica.

79
In allegato 5 si fornisce una lista delle corrispondenze fra i 58 aplotipi emersi
dall’analisi della regione esonica del locus DRB del presente studio e la
nomenclatura in uso per la definizione degli aplotipi della medesima regione.
Partendo dai 58 aplotipi, mediante l’uso del programma MEGA v. 4.0, abbiamo
costruito le sequenze degli amminoacidi che corrispondono ai nostri alleli. In totale
(Figure 3.2) da 206 nucleotidi risultano 68 codoni che codificano per 68 aminoacidi.
Dallo studio dell’allineamento risultano 18 siti amminoacidici variabili (26,47% di
siti polimorfici). Nella Figura di seguito vengono, inoltre, evidenziati i codoni
coinvolti nella regione di legame dell’antigene delle MHC di classe II, la regione
PBR, nell’uomo (Brown et al. 1993; Schaschal et al. 2004).

80
+ + + + + + ++ + + ++ + +
* * * * * *** * * * ** *
Hap_1 EYHKSECHFF NGTERVRFLD RYFHNGEELV RFDSDWGEYR AVAELGRPTA EHWNSQKEIL [60]
Hap_2 ..S.K..... .......... .......... .......... .......... .......... [60]
Hap_3 ..P....... .......... .......... .......... .......... .......... [60]
Hap_4 ..S.K..... .......L.. ...Y....YA ..N....... ........D. .Y........ [60]
Hap_5 .......... .......... .......... .......... ........A. .......... [60]
Hap_6 .......... .......... .......... ........F. ........A. .......... [60]
Hap_7 ..S....... .......... .......... ........F. ........D. .Y........ [60]
Hap_8 .......... .......... .......... .......... .......RD. .......... [60]
Hap_9 ..N.K..... .......... .......... .......... .......... .......... [60]
Hap_10 ..S....... .......... .......... .......... .......... .......... [60]
Hap_11 .......... .......... .......... ........F. .......... .......... [60]
Hap_12 .......... .......... .......... ..N....... ........A. .......... [60]
Hap_13 .......... .......... ........F. ..N....... ........A. .......... [60]
Hap_14 ..R....... .......... ........F. ..N....... .......... .......... [60]
Hap_15 ..R....... .......... ...Y...... ..N....... .......... .......... [60]
Hap_16 ..R....... .......... ...Y...... ..N....... .......... .......... [60]
Hap_17 .......... .......... .......... ..N....... .......R.. .......... [60]
Hap_18 .......... .......... .......... ..N....... .......R.. .......... [60]
Hap_19 ..S....... .......L.. ...Y....YA ..N....... .......... .......... [60]
Hap_20 ..S....... .......... ...Y....YA ..N....... .......RD. .......... [60]
Hap_21 .......... .......... .......... .......... .......... .......... [60]
Hap_22 ..S....... .......... ...Y....YA ..N....... .......RD. .......... [60]
Hap_23 ..S....... .......... ...Y....YA ..N....... .......RD. .......... [60]
Hap_24 ..S....... .......L.. ...Y....YA ..N....... .......... .......... [60]
Hap_25 ..S....... .......... ...Y....YA ..N....... .......... .......... [60]
Hap_26 ..S....... .......L.. ...Y....YA ..N....... .......RD. .......... [60]
Hap_27 .......... .......... .......V.. .......... .......... .......... [60]
Hap_28 .......... .......L.. ...Y....YA ..N....... .......... .......... [60]
Hap_29 ..R....... .......... .......... .......... .......... .......... [60]
Hap_30 ..S.K..... .......... .......... ........F. ........D. .Y........ [60]
Hap_31 ....R..... .......... .......... .......... .......... .......... [60]
Hap_32 .......... .......... .......... .......... .......... .......... [60]
Hap_33 ..R....... .......L.. ...Y....YA ..N....... ........D. .Y........ [60]
Hap_34 ..T....... .......... .......... .......... .......... .......... [60]
Hap_35 ..R....... .......... .......... .......... ........D. .Y........ [60]
Hap_36 ..S....... .......L.. ...Y....YA ..N.....F. ........D. .Y........ [60]
Hap_37 ..T....... .......... .......... .......... .......... .......... [60]
Hap_38 ..S.K..... .......... .......... ........F. ........D. .Y........ [60]
Hap_39 ..R....... .......... ...Y....YA ..N....... ........D. .Y........ [60]
Hap_40 ..T.K..... .......... .......... .......... .......... .......... [60]
Hap_41 ..R....... .......... .......... .......... .......... .......... [60]
Hap_42 ..S....... .......L.. ...Y....YA ..N....... ........D. .Y........ [60]
Hap_43 ..R....... .......... ...Y....YA ..N....... ........D. .......... [60]
Hap_44 ..S....... .......L.. ...Y....YA ..N....... .......RD. .Y........ [60]
Hap_45 ..S....... .......... .......... .......... .......... .......... [60]
Hap_46 ....R..... .......... .......... .......... .......... .......... [60]
Hap_47 ..S....... .......... .......... ........F. ........D. .Y........ [60]
Hap_48 ..S....... .......... .......... ........F. ........D. .Y........ [60]
Hap_49 .......... .......... ...Y....YA ..N....... .......... .......... [60]
Hap_50 ..R....... .......L.. ...Y....YA ..N....... ........D. .Y........ [60]
Hap_51 ..P.R..... .......... .......... .......... .......... .......... [60]
Hap_52 ..S....... .......... ...Y....YE ..N....... .......RD. .......... [60]
Hap_53 ..S....... .......... ...Y....YA ..N....... ........D. .......... [60]
Hap_54 .......... .......... ...Y....YA ..N....... .......... .......... [60]
Hap_55 ..P....... .......... .......... .......... .......... .......... [60]
Hap_56 .......... .......... .......... .......... ...R...... .......... [60]
Hap_57 ..S....... .......L.. ...Y....YA ..N....... ........D. .Y........ [60]
Hap_58 A......... .......... .......... .......... .......... .......... [60]
++ +
*** *
Hap_1 ERRRAAVD [68]
Hap_2 .QT..... [68]
Hap_3 .Q...... [68]
Hap_4 .QT..... [68]
Hap_5 ........ [68]
Hap_6 ........ [68]

81
Hap_7 ..K..... [68]
Hap_8 ........ [68]
Hap_9 .QT..... [68]
Hap_10 .QT..... [68]
Hap_11 ........ [68]
Hap_12 .Q...E.. [68]
Hap_13 .Q...E.. [68]
Hap_14 .Q...... [68]
Hap_15 ........ [68]
Hap_16 .Q...... [68]
Hap_17 ........ [68]
Hap_18 .QT..... [68]
Hap_19 .Q...... [68]
Hap_20 .QT..... [68]
Hap_21 ...A.... [68]
Hap_22 .QTA.... [68]
Hap_23 .Q...... [68]
Hap_24 ........ [68]
Hap_25 .Q...... [68]
Hap_26 .QT..... [68]
Hap_27 ........ [68]
Hap_28 .Q...... [68]
Hap_29 ..M..... [68]
Hap_30 .QT..... [68]
Hap_31 ........ [68]
Hap_32 .Q...... [68]
Hap_33 .Q...... [68]
Hap_34 .QT..... [68]
Hap_35 ........ [68]
Hap_36 .QK..... [68]
Hap_37 ........ [68]
Hap_38 .QK..... [68]
Hap_39 .QT..... [68]
Hap_40 .QT..... [68]
Hap_41 .Q...... [68]
Hap_42 .QT..... [68]
Hap_43 .Q...... [68]
Hap_44 .QT..... [68]
Hap_45 ........ [68]
Hap_46 .QT..... [68]
Hap_47 ........ [68]
Hap_48 ..T..... [68]
Hap_49 .Q...... [68]
Hap_50 .QT..... [68]
Hap_51 .Q...... [68]
Hap_52 .QT..... [68]
Hap_53 .Q...... [68]
Hap_54 .QT..... [68]
Hap_55 ........ [68]
Hap_56 ........ [68]
Hap_57 .Q...... [68]
Hap_58 ........ [68]
La regione sequenziata interamente codificante è formata da 68 aminoacidi.
Per i camosci alpini, su 206 nucleotidi, il numero di sostituzioni sinonime è 22 (ds –
tasso medio di sostituzioni sinonime = 0.0053) mentre il numero di sostituzioni non
sinonime è 9 (dn – tasso medio di sostituzioni non sinonime = 0.0532). Il rapporto
dn/ds perciò è pari a 10,04. Per i camosci pirenaici il numero di sostituzioni sinonime
è 20 (ds = 0.0019) mentre il numero di sostituzioni non sinonime è 4 (dn = 0.0532). Il
rapporto dn/ds perciò è pari a 28.
Figura 3.2 Allineamento delle sequenze aminoacidiche derivate dagli aplotipi trovati. Si evidenziano con * i siti polimorfici e con + i siti coinvolti nella codificazione della regione PBR delle molecole delle MHC di
classe II nell’uomo secondo Brown et al. 1993.

82
Considerando l’allineamento nucleotidico dei 58 aplotipi ricostruiti nel
seguente studio, sono state identificate 24 posizioni variabili. Sul totale, 8 variazioni
cadono in prima posizione, 14 in seconda posizione codonica e solo 2 in terza
posizione. L’allineamento nucleotidico genera un insieme di 68 codoni, di cui 18
variabili.
Analizzando invece le variazioni a livello aminoacidico, il cambiamento da acido
glutammico ad alanina in posizione 1 comporta un cambiamento di carica, in
quanto il primo è un aminoacido idrofilico contenente il gruppo acido COO- mentre
il secondo è un aminoacido alifatico idrofobico; in posizione 3 si possono ritrovare 6
aminoacidi diversi (istidina, serina, prolina, asparagina, arginina, treonina) di questi,
5 presentano caratteristiche idrofiliche e solo uno, la prolina (presente nell’aplotipo
3, 51 e 55), presenta invece caratteristiche idrofobiche. In posizione 5 sono
presenti la serina (contenente il gruppo OH-), la lisina e l’arginina (contenenti il
gruppo NH3+), tutti con caratteristiche idrofiliche. In posizione 18 ritroviamo due
aminoacidi: la fenilalanina e la leucina, il primo aromatico e il secondo alifatico, ma
entrambi con caratteristiche idrofobiche. In posizione 24 sono presenti 2
aminoacidi, istidina e tirosina, il primo idrofilico il secondo idrofobico. In posizione
28 sono presenti l’acido glutammico e la valina, e come nel caso della posizione 1 e
della posizione 24 ritroviamo che la sostituzione aminoacidica comporta un
cambiamento totale delle caratteristiche di legame dell’aminoacido (il primo acido
e idrofilico, il secondo alifatico e idrofobico). Nella posizione successiva, 29,
ritroviamo 3 aminoacidi variabili: la leucina, la fenilalanina e la tirosina; i primi due,
sebbene abbiano strutture chimiche diverse mantengono entrambi caratteristiche
di idrofobicità, mentre l’ultimo è idrofilico. In posizione 30 si ritrovano di nuovo 3
aminoacidi variabili: valina, alanina e acido glutammico. In posizione 33 sono
presenti l’acido aspartico e l’asparagina, il primo spiccatamente acido e idrofilico, il
secondo moderatamente idrofilico e senza una carica netta. In posizione 39 sono
presenti la fenilalanina e la tirosina; in posizione 44 sono presenti l’acido

83
glutammico e l’arginina ed in posizione 48 ritroviamo la prolina e l’arginina: in tutti i
casi ritroviamo aminoacidi con caratteristiche opposte. In posizione 49 sono
presenti tirosina, acido aspartico e alanina tutti e tre con caratteristiche diverse fra
loro; in posizione 52 sono presenti istidina e tirosina, con caratteristiche diverse; in
posizione 62 ritroviamo arginina e glutammina, la prima idrofilia e carica
positivamente, la seconda senza una carica netta. In posizione 63 ritroviamo 4
aminoacidi diversi: arginina, lisina, treonina e metionina; in posizione 64 sono
presenti arginina e alanina e in ultima posizione variabile ritroviamo alanina e acido
glutammico.
Come si evince da questi risultati, i cambiamenti aminoacidici dovuti alla
grande variazione presente in prima e seconda posizione codonica
dell’allineamento nucleotidico, comportano di solito cambiamenti significativi nella
struttura chimica e nelle caratteristiche di idrofobicità e idrofilicità degli aminoacidi
in questione. Essendo una regione interamente codificante, inclusa la regione che
lega il peptide (PBR), tali cambiamenti rappresentano una diversa capacità di
discriminare epitopi di antigeni di agenti patogeni molto diversi fra loro.
3.2 Diversità genetica intra-popolazioni
Per le popolazioni campionate nei diversi siti (SitoA, B, C, D, E, 1, 2, 3, 4), le
frequenze relative alleliche e gli indici di diversità genetica (Nei 1987, nA, H0, He) e
gli indici di diversità molecolare secondo (Ewens 1972; Zouros 1979; Tajima 1983;
Nei 1987; Tajima 1993) sono elencati nella tabella 5.
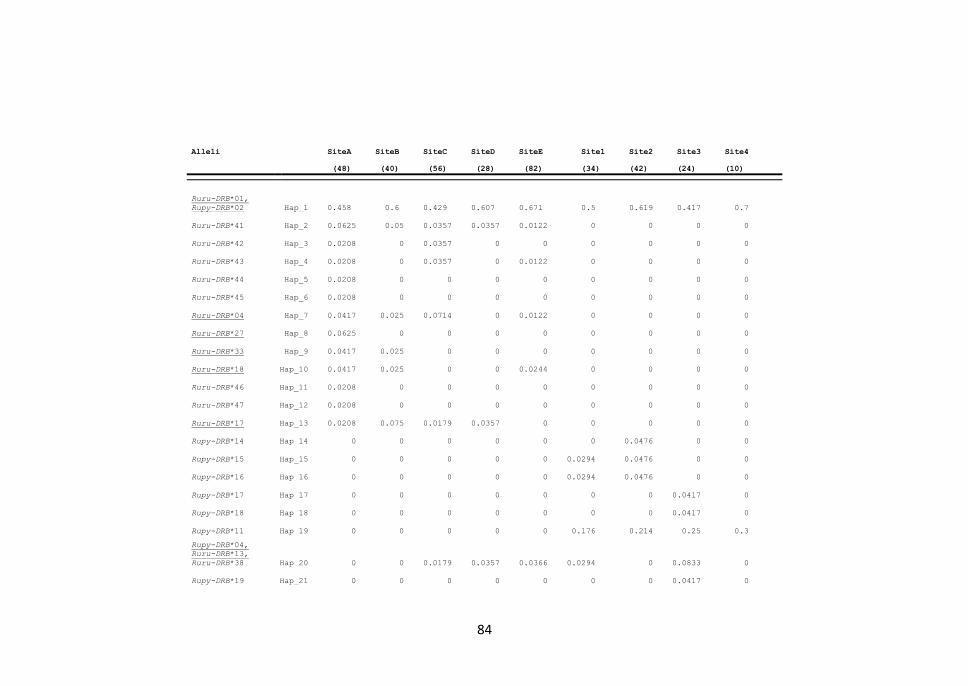
84
Alleli SiteA SiteB SiteC SiteD SiteE Site1 Site2 Site3 Site4
(48) (40) (56) (28) (82) (34) (42) (24) (10)
Ruru-DRB*01,
Rupy-DRB*02 Hap_1 0.458 0.6 0.429 0.607 0.671 0.5 0.619 0.417 0.7
Ruru-DRB*41 Hap_2 0.0625 0.05 0.0357 0.0357 0.0122 0 0 0 0
Ruru-DRB*42 Hap_3 0.0208 0 0.0357 0 0 0 0 0 0
Ruru-DRB*43 Hap_4 0.0208 0 0.0357 0 0.0122 0 0 0 0
Ruru-DRB*44 Hap_5 0.0208 0 0 0 0 0 0 0 0
Ruru-DRB*45 Hap_6 0.0208 0 0 0 0 0 0 0 0
Ruru-DRB*04 Hap_7 0.0417 0.025 0.0714 0 0.0122 0 0 0 0
Ruru-DRB*27 Hap_8 0.0625 0 0 0 0 0 0 0 0
Ruru-DRB*33 Hap_9 0.0417 0.025 0 0 0 0 0 0 0
Ruru-DRB*18 Hap_10 0.0417 0.025 0 0 0.0244 0 0 0 0
Ruru-DRB*46 Hap_11 0.0208 0 0 0 0 0 0 0 0
Ruru-DRB*47 Hap_12 0.0208 0 0 0 0 0 0 0 0
Ruru-DRB*17 Hap_13 0.0208 0.075 0.0179 0.0357 0 0 0 0 0
Rupy-DRB*14 Hap_14 0 0 0 0 0 0 0.0476 0 0
Rupy-DRB*15 Hap_15 0 0 0 0 0 0.0294 0.0476 0 0
Rupy-DRB*16 Hap_16 0 0 0 0 0 0.0294 0.0476 0 0
Rupy-DRB*17 Hap_17 0 0 0 0 0 0 0 0.0417 0
Rupy-DRB*18 Hap_18 0 0 0 0 0 0 0 0.0417 0
Rupy-DRB*11 Hap_19 0 0 0 0 0 0.176 0.214 0.25 0.3
Rupy-DRB*04,
Ruru-DRB*13,
Ruru-DRB*38 Hap_20 0 0 0.0179 0.0357 0.0366 0.0294 0 0.0833 0
Rupy-DRB*19 Hap_21 0 0 0 0 0 0 0 0.0417 0

85
Rupy-DRB*20 Hap_22 0 0 0 0 0 0 0 0.0417 0
Rupy-DRB*21 Hap_23 0 0 0 0 0 0.0294 0 0 0
Rupy-DRB*22 Hap_24 0 0 0 0 0 0 0.0238 0 0
Rupy-DRB*23 Hap_25 0 0 0 0 0 0.0882 0 0 0
Rupy-DRB*24 Hap_26 0 0 0 0 0 0.0882 0 0 0
Rupy-DRB*25 Hap_27 0 0 0 0 0 0 0 0.0833 0
Rupy-DRB*26 Hap_28 0 0 0 0 0 0.0294 0 0 0
Ruru-DRB*48 Hap_29 0.0208 0 0 0 0 0 0 0 0
Ruru-DRB*49 Hap_30 0.0208 0 0 0 0 0 0 0 0
Ruru-DRB*50 Hap_31 0.0208 0 0 0 0 0 0 0 0
Ruru-DRB*51 Hap_32 0.0417 0 0.0357 0 0.0122 0 0 0 0
Ruru-DRB*09 Hap_33 0.0208 0.05 0.143 0.0714 0.0244 0 0 0 0
Ruru-DRB*52 Hap_34 0.0208 0 0 0 0 0 0 0 0
Ruru-DRB*53 Hap_35 0 0 0.0357 0 0 0 0 0 0
Ruru-DRB*54 Hap_36 0 0 0.0357 0 0 0 0 0 0
Ruru-DRB*55 Hap_37 0 0 0.0179 0.0357 0 0 0 0 0
Ruru-DRB*56 Hap_38 0 0 0.0179 0.0357 0 0 0 0 0
Ruru-DRB*28 Hap_39 0 0 0.0357 0 0 0 0 0 0
Ruru-DRB*57 Hap_40 0 0 0.0357 0 0 0 0 0 0
Ruru-DRB*58 Hap_41 0 0 0 0.0357 0 0 0 0 0
Ruru-DRB*59 Hap_42 0 0 0 0.0357 0 0 0 0 0
Ruru-DRB*60 Hap_43 0 0 0 0.0357 0 0 0 0 0
Ruru-DRB*61 Hap_44 0 0 0 0.0357 0.0244 0 0 0 0
Ruru-DRB*39 Hap_45 0 0.025 0 0 0 0 0 0 0
Ruru-DRB*62 Hap_46 0 0.025 0 0 0 0 0 0 0
Ruru-DRB*63 Hap_47 0 0.025 0 0 0 0 0 0 0
Ruru-DRB*64 Hap_48 0 0.025 0 0 0 0 0 0 0

86
Ruru-DRB*65 Hap_49 0 0.025 0 0 0 0 0 0 0
Ruru-DRB*66 Hap_50 0 0.025 0 0 0 0 0 0 0
Ruru-DRB*67 Hap_51 0 0 0 0 0.0122 0 0 0 0
Ruru-DRB*68 Hap_52 0 0 0 0 0.0122 0 0 0 0
Ruru-DRB*69 Hap_53 0 0 0 0 0.0244 0 0 0 0
Ruru-DRB*21 Hap_54 0 0 0 0 0.0244 0 0 0 0
Ruru-DRB*70 Hap_55 0 0 0 0 0.0122 0 0 0 0
Ruru-DRB*71 Hap_56 0 0 0 0 0.0244 0 0 0 0
Ruru-DRB*35,
Rupy-DRB*08 Hap_57 0 0 0 0 0.0488 0 0 0 0
Ruru-DRB*72 Hap_58 0 0 0 0 0.0122 0 0 0 0
N 24 20 28 13 41 17 21 13 4
nA 19 13 15 11 17 9 6 8 2
H0 media 0,786 0,639 0,793 0,637 0,548 0,720 0,577 0,775 0,466
ds (±0,016) (±0,016) (±0,049) (±0,105) (±0,067) (±0,071) (±0,105) (±0,066) (±0,131) He media 0,227 0,250 0,361 0,299 0,266 0,422 0,418 0,370 0,466
ds 0,134 0,092 0,150 0,130 0,133 0,119 0,045 0,138 0,000
transiz. 10 10 10 10 12 7 6 7 6
trasvers 10 8 10 10 11 6 3 9 3
sostituz. 20 18 20 10 20 13 9 16 9
Tabella 5 Frequenze relative degli aplotipi dei campioni esaminati divisi per sito di campionamento. Gli alleli che corrispondono a sequenze presenti in letteratura (Schaschal et al. 2004) sono sottolineate. N=numero di campioni esaminati per sito di campionamento; nA= numero di alleli per sito; H0= eterozigosità media osservata; He= eterozigosità media attesa; ds= deviazione standard; divesità genetica; numero di transizioni, trasversioni e sostituzioni osservato.

87
Possiamo notare che l’allele Ruru-DRB01 è presente in tutti i campioni con una
frequenza molto alta (circa 50%). Tutti i restanti alleli sono presenti con una
frequenza ridotta, con un range che va da 1,22% a 30%.
La variabilità genetica ha un valore piuttosto elevato, con una eterozigosità
media attesa He che va da un valore di 0,227 per i camosci alpini di Auronzo morti
per rogna sarcoptica a 0,466 per il Sito4, per i camosci pirenaici.
Considerando le popolazioni raggrupppate secondo lo status possiamo evidenziare
altre caratteristiche di variabilità (Tab. 6). L’eterozigosi osservata, in questo caso
presenta valori H0 da 0,469 a 0,766.

88
Haplotype: rogna_morti rogna_soprav. rogna_res. sp_pest+ sp_altremal. sp_sani pol_preout polm_postout
(50) (42) (82) (26) (40) (44) (34) (48)
Hap_1 0.48 0.571 0.476 0.577 0.525 0.545 0.588 0.729
Hap_2 0.06 0.0476 0.0366 0 0 0 0.0294 0
Hap_3 0.02 0 0.0244 0 0 0 0 0
Hap_4 0.02 0.0238 0.0244 0 0 0 0 0.0208
Hap_5 0.02 0 0 0 0 0 0 0
Hap_6 0.02 0 0 0 0 0 0 0
Hap_7 0.04 0.0238 0.0488 0 0 0 0 0.0208
Hap_8 0.06 0 0 0 0 0 0 0
Hap_9 0.04 0.0238 0 0 0 0 0 0
Hap_10 0.04 0.0238 0 0 0 0 0 0.0417
Hap_11 0.02 0 0 0 0 0 0 0
Hap_12 0.02 0 0 0 0 0 0 0
Hap_13 0.02 0.0714 0.0244 0 0 0 0 0
Hap_14 0 0 0 0 0.05 0 0 0
Hap_15 0 0 0 0 0.075 0 0 0
Hap_16 0 0 0 0 0.075 0 0 0
Hap_17 0 0 0 0 0.025 0 0 0
Hap_18 0 0 0 0 0.025 0 0 0
Hap_19 0 0 0 0.385 0.15 0.182 0 0
Hap_20 0 0 0.0244 0 0 0.0682 0.0294 0.0417
Hap_21 0 0 0 0 0 0.0227 0 0
Hap_22 0 0 0 0 0 0.0227 0 0
Hap_23 0 0 0 0.0385 0 0 0 0
Hap_24 0 0 0 0 0 0.0227 0 0

89
Hap_25 0 0 0 0 0.025 0.0455 0 0
Hap_26 0 0 0 0 0.025 0.0455 0 0
Hap_27 0 0 0 0 0 0.0455 0 0
Hap_28 0 0 0 0 0.025 0 0 0
Hap_29 0.02 0 0 0 0 0 0 0
Hap_30 0.02 0 0 0 0 0 0 0
Hap_31 0.02 0 0 0 0 0 0 0
Hap_32 0.04 0 0.0244 0 0 0 0.0294 0
Hap_33 0.02 0.0476 0.122 0 0 0 0.0294 0.0208
Hap_34 0.02 0 0 0 0 0 0 0
Hap_35 0 0 0.0244 0 0 0 0 0
Hap_36 0 0 0.0244 0 0 0 0 0
Hap_37 0 0 0.0244 0 0 0 0 0
Hap_38 0 0 0.0244 0 0 0 0 0
Hap_39 0 0 0.0244 0 0 0 0 0
Hap_40 0 0 0.0244 0 0 0 0 0
Hap_41 0 0 0.0122 0 0 0 0 0
Hap_42 0 0 0.0122 0 0 0 0 0
Hap_43 0 0 0.0122 0 0 0 0 0
Hap_44 0 0 0.0122 0 0 0 0.0294 0.0208
Hap_45 0 0.0238 0 0 0 0 0 0
Hap_46 0 0.0238 0 0 0 0 0 0
Hap_47 0 0.0238 0 0 0 0 0 0
Hap_48 0 0.0238 0 0 0 0 0 0
Hap_49 0 0.0238 0 0 0 0 0 0
Hap_50 0 0.0238 0 0 0 0 0 0
Hap_51 0 0.0238 0 0 0 0 0 0.0208
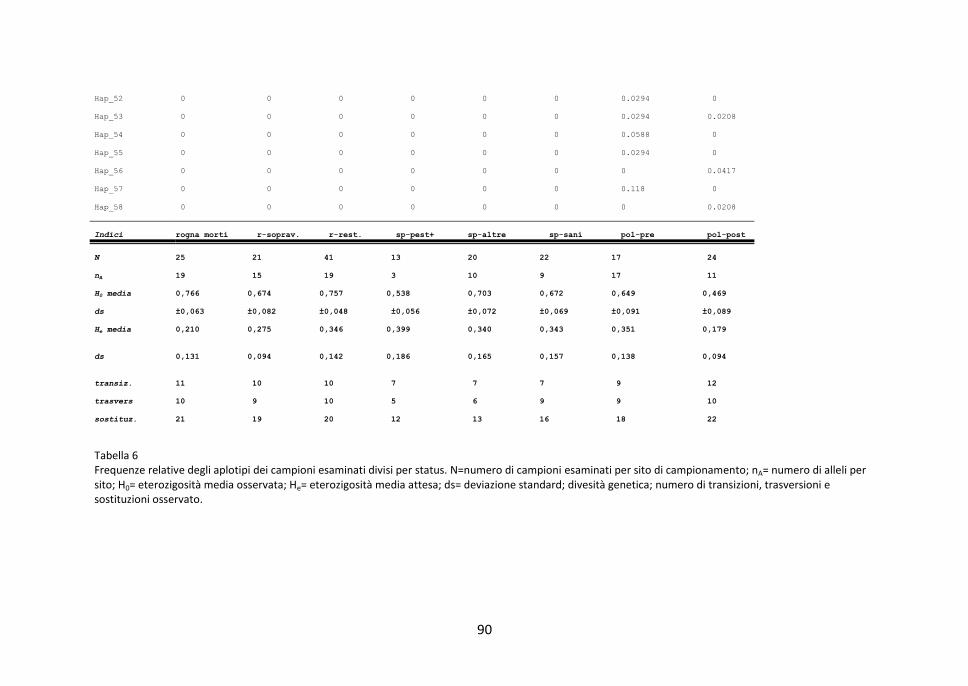
90
Hap_52 0 0 0 0 0 0 0.0294 0
Hap_53 0 0 0 0 0 0 0.0294 0.0208
Hap_54 0 0 0 0 0 0 0.0588 0
Hap_55 0 0 0 0 0 0 0.0294 0
Hap_56 0 0 0 0 0 0 0 0.0417
Hap_57 0 0 0 0 0 0 0.118 0
Hap_58 0 0 0 0 0 0 0 0.0208
Indici rogna morti r-soprav. r-rest. sp-pest+ sp-altre sp-sani pol-pre pol-post
N 25 21 41 13 20 22 17 24
nA 19 15 19 3 10 9 17 11
H0 media 0,766 0,674 0,757 0,538 0,703 0,672 0,649 0,469
ds ±0,063 ±0,082 ±0,048 ±0,056 ±0,072 ±0,069 ±0,091 ±0,089
He media 0,210 0,275 0,346 0,399 0,340 0,343 0,351 0,179
ds 0,131 0,094 0,142 0,186 0,165 0,157 0,138 0,094
transiz. 11 10 10 7 7 7 9 12
trasvers 10 9 10 5 6 9 9 10
sostituz. 21 19 20 12 13 16 18 22
Tabella 6 Frequenze relative degli aplotipi dei campioni esaminati divisi per status. N=numero di campioni esaminati per sito di campionamento; nA= numero di alleli per sito; H0= eterozigosità media osservata; He= eterozigosità media attesa; ds= deviazione standard; divesità genetica; numero di transizioni, trasversioni e sostituzioni osservato.

91
3.3 Diversità genetica tra-popolazioni
L’analisi della varianza molecolare (AMOVA), è stata condotta utilizzando il
software ARLEQUIN 3.1. Per applicare questo tipo di analisi è necessario
strutturare gli individui da analizzare in popolazioni e gruppi di ordine gerarchico
superiore. Nel nostro caso sono stati applicati due criteri di divisione: il primo
criterio è stato quello geografico (Tab. 7), ovvero si sono divisi i campioni in due
gruppi principali (camosci italiani appartenenti alla specie R. r. rupicapra e camosci
spagnoli appartenenti alla specie R. p. pyrenaica) e all’interno di ogni gruppo è
stata fatta un’ulteriore divisione in popolazioni a seconda della località di
provenienza; il secondo criterio di divisione è stato quello di considerare gruppi di
camosci sottoposti a pressione selettiva da patogeno (Tab. 8). Alcuni individui delle
popolazioni di camosci spagnoli sono stati affetti da Pestivirus e da altre patologie,
mentre gli individui delle popolazioni italiane sono stati affetti da rogna sarcoptica
e da polmonite.
Questo tipo di approccio gerarchico per l’analisi della varianza molecolare
consente di dividere la varianza totale in componenti di covarianza, dovute alle
differenze intra-individuali, intra-popolazione e inter-gruppi.
Questo tipo di analisi essenzialmente è simile agli altri tipi di approcci basati
sull’analisi della varianza delle frequenze genotipiche, ma tiene in conto il numero
delle mutazioni fra gli aplotipi (Excoffier et al. 2005).
La significatività degli indici di fissazione è valutata usando l’approccio di
permutazione non parametrica (1000 repliche).
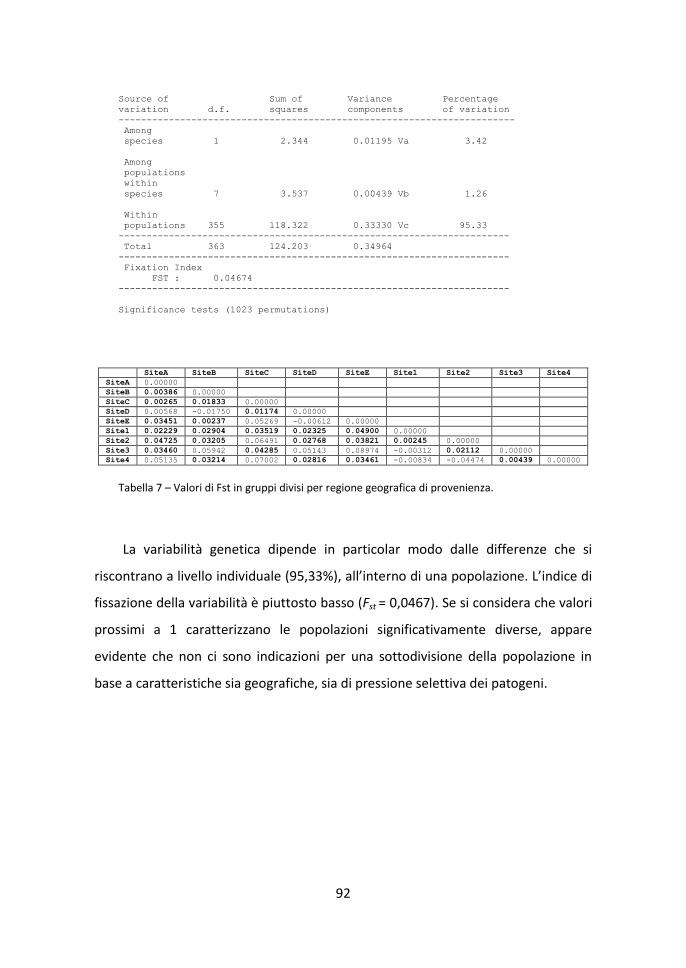
92
Source of Sum of Variance Percentage
variation d.f. squares components of variation
-----------------------------------------------------------------------
Among
species 1 2.344 0.01195 Va 3.42
Among
populations
within
species 7 3.537 0.00439 Vb 1.26
Within
populations 355 118.322 0.33330 Vc 95.33
----------------------------------------------------------------------
Total 363 124.203 0.34964
----------------------------------------------------------------------
Fixation Index
FST : 0.04674
----------------------------------------------------------------------
Significance tests (1023 permutations)
SiteA SiteB SiteC SiteD SiteE Site1 Site2 Site3 Site4
SiteA 0.00000
SiteB 0.00386 0.00000
SiteC 0.00265 0.01833 0.00000
SiteD 0.00568 -0.01750 0.01174 0.00000
SiteE 0.03451 0.00237 0.05269 -0.00612 0.00000
Site1 0.02229 0.02904 0.03519 0.02325 0.04900 0.00000
Site2 0.04725 0.03205 0.06491 0.02768 0.03821 0.00245 0.00000
Site3 0.03460 0.05942 0.04285 0.05143 0.08974 -0.00312 0.02112 0.00000
Site4 0.05135 0.03214 0.07002 0.02816 0.03461 -0.00834 -0.04474 0.00439 0.00000
Tabella 7 – Valori di Fst in gruppi divisi per regione geografica di provenienza.
La variabilità genetica dipende in particolar modo dalle differenze che si
riscontrano a livello individuale (95,33%), all’interno di una popolazione. L’indice di
fissazione della variabilità è piuttosto basso (Fst = 0,0467). Se si considera che valori
prossimi a 1 caratterizzano le popolazioni significativamente diverse, appare
evidente che non ci sono indicazioni per una sottodivisione della popolazione in
base a caratteristiche sia geografiche, sia di pressione selettiva dei patogeni.

93
----------------------------------------------------------------------
Source of Sum of Variance Percentage
variation d.f. squares components of variation
----------------------------------------------------------------------
Healthy/
sick 2 3.544 0.01170 Va 3.36
Among
populations
within
above groups 5 2.065 0.00176 Vb 0.50
Within
populations 358 119.893 0.33490 Vc 96.14
----------------------------------------------------------------------
Total 365 125.503 0.34835
----------------------------------------------------------------------
Fixation Indices
FST : 0.03862
----------------------------------------------------------------------
Significance tests (1023 permutations)
------------------
Rogna
morti
Rogna
Spravv.
Rogna
Resist.
Spagna
Pestivirus
Spagna
malattie
Spagna
sani
Polmonite
Preout.
Polm.
Postout.
Rogna
morti
0.00000
Rogna
Spravv
-0.0043 0.00000
Rogna
Resist.
0.00105 0.00169 0.00000
Spagna
Pest.
0.09031 0.09228 0.09587 0.00000
Spagna
malattie
0.01769 0.01703 0.02636 0.02711 0.00000
Spagna
sani
0.02514 0.02229 0.03148 0.01413 -0.00484 0.00000
Polmonite
Preout.
0.00810 -0.00037 0.01327 0.09955 0.02215 0.02430 0.00000
Polmointe
Postout.
0.04364 0.01469 0.04824 0.13307 0.05185 0.04796 0.01707 0.00000
Tabella 8 Valori di Fst in gruppi sottoposti a pressioni selettive da patogeno.
La percentuale di variabilità riscontrata nei gruppi costituiti a seconda dello
status degli individui rispetto alla pressione da patogeno dipende soprattutto dalla
diversità presente tra gli individui stessi (96,14%), l’indice di fissazione della
variabilità è pari a Fst = 0,0386.
Di seguito si schematizza il valore della diversità genetica tra le diverse
polazioni campionate raggruppate per sito di campionamento e per status (Tab. 9 e
Fig. 3.3a e b).

94
Gene diversity SiteA SiteB SiteC SiteD SiteE Site1 Site2 Site3 Site4
0.7863 0.6397 0.7935 0.6376 0.5483 0.7201 0.5772 0.7754 0.4667
Gene diversity
Rogna morti
Rogna Spravv.
Rogna Resist.
Spagna Pestivirus
Spagna altre malattie
Spagna sani
Polmonite Preout.
Polmointe Postout.
0.7665 0.6736 0.7573 0.5385 0.7026 0.6723 0.6488 0.4699
Tabella 9 Risultati della variabilità genetica in gruppi suddivisi per siti di campionamento e per status
a
b
Fig. 3.3 Istogrammi rappresentanti la variabilità genetica in gruppi suddivisi per sito di campionamento (a) e sottoposti a pressioni selettive da patogeno (b).
0 0,2 0,4 0,6 0,8
SiteA
SiteB
SiteC
SiteD
SiteE
Site1
Site2
Site3
Site4
Serie1
0 0,2 0,4 0,6 0,8
Rogna morti
Rogna soprav
Rogna resist
Polmonite pre
Polmonite post
Pestivirus
Altre patolog
Sani
Serie1

95
3.4 Alberi fenetici secondo il metodo Neighbor-Joining
Per evidenziare le relazioni tra i 58 aplotipi evidenziati nelle due specie di
camoscio è stato realizzato un albero NJ (Saitou & Nei 1987) basato sulle sequenze
nucleotidiche dell’esone 2 locus DRB delle MHC di classe II (Fig. 3.4). Le distanze
evolutive sono state calcolate con il metodo Kimura 2-parameters (Kimura 1980).
Le analisi sono state condotte utilizzando il programma MEGA v. 4.0. Va ricordato
che l’albero ottenuto si riferisce agli aplotipi evidenziati e non agli individui
analizzati. Le sequenze dei diversi campioni sono distribuite in modo eterogeneo
tra i principali gruppi.
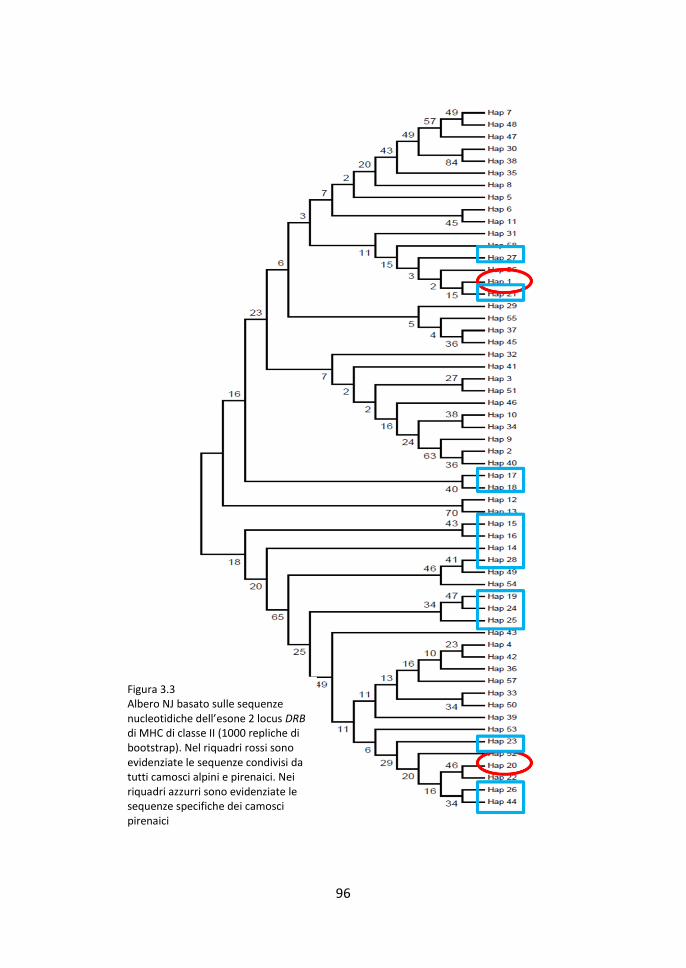
96
Figura 3.3 Albero NJ basato sulle sequenze nucleotidiche dell’esone 2 locus DRB di MHC di classe II (1000 repliche di bootstrap). Nel riquadri rossi sono evidenziate le sequenze condivisi da tutti camosci alpini e pirenaici. Nei riquadri azzurri sono evidenziate le sequenze specifiche dei camosci pirenaici

97
DISCUSSIONE DEI DATI E CONSIDERAZIONI
Il camoscio è una specie di grande importanza in Italia e nei Paesi in cui si
espande il suo areale, sia per il fatto che è molto abbondante, sia perché è soggetto
a prelievo venatorio. La presenza di parchi naturali e oasi di rifugio, di riproduzione
e sosta, in cui gli animali non possono essere cacciati, permettono di mantenere
queste specie in diverse aree a buoni livelli di densità, anche se tale densità è
piuttosto variabile da zona a zona.
Questo studio è stato effettuato nella Provincia di Belluno, zona dolomitica in
cui il camoscio è una delle specie più diffuse e rappresentative, nella Provincia di
Lecco in cui vi sono numerose aree protette e zone adibite alla caccia, e nel
versante spagnolo dei Pirenei. In tutte queste zone le popolazioni di camoscio siano
esse di camoscio alpino o piraneaico sono continuamente minacciate da parassitosi
ed epidemie di vario genere che causano ciclicamente pesanti perdite di individui,
ed anche se il camoscio non è una specie in pericolo di estinzione certamente la
drastica diminuzione della sua densità in determinate aree, potrebbe implicare
problematiche relativamente alla politica di conservazione ed all’interesse
venatorio a cui è soggetta questa specie.
Le popolazioni di camoscio alpino sono state soggette ciclicamente (ogni 15
anni circa) a drastiche riduzioni della densità di popolazione a causa in particolar
modo della rogna sarcoptica (Guberti & Zamboni 2000), infestazione che ha
pesantemente colpito le popolazioni della Provincia di Belluno.
La comprensione dei processi di generazione e mantenimento della variabilità
genetica con l’individuazione di markers che forniscono ai portatori una resistenza

98
a specifici patogeni, potrebbe risultare fondamentale nella gestione delle risorse e
nel controllo sanitario delle popolazioni di camoscio.
Questo studio perciò si colloca in una serie di ricerche che sono cominciate, in
Italia, a seguito del primo evento di rogna sarcoptica nel Bellunese che nel 1995
provocò un collasso demografico di camosci alpini pari all’80% degli individui delle
popolazioni colpite, e che hanno il fine di comprendere il livello e la distribuzione
della variabilità genetica adattativa nei camosci.
Nel corso dello studio, si sono aggiunti alla ricerca campioni provenienti da altre
regioni e sottoposti ad altri agenti patogeni, offrendo a possibilità di studiare in
modo più approfondito la variabilità a livello intra popolazioni e inter specifico.
Gli scopi di questa tesi di dottorato sono volti a ottenere informazioni sulla
variabilità genetica e sulla struttura delle popolazioni di camoscio alpino della
Provinica di Belluno e della Provincia di Lecco, e delle popolazioni di camoscio
pirenaico del versante spagnolo dei Pirenei utilizzando un gene codificante per
proteine del complesso maggiore di istocompatibilità coinvolto nella risposta
immunitaria e quindi sicuramente sottoposto a processi selettivi adattativi.
4.1 Variabilità genetica delle popolazioni di camoscio
La variabilità dei geni del sistema MHC è un indicatore per la resistenza a
parassiti e patogeni che a turno possono influenzare la probabilità di sopravvivenza
a lungo termine delle popolazioni. Il polimorfismo del locus MHC si pensa sia
mantenuto dalla selezione bilanciante (Hedrick & Thompson 1987), che può essere
il risultato di una superiorità degli eterozigoti oppure di un vantaggio per gli alleli
rari (Hughes & Yeage, 1998). La prima ipotesi prevede una selezione a favore degli
eterozigoti rispetto agli individui omozigoti, per il fatto che i primi potrebbero

99
essere più resistenti a diversi patogeni grazie alla possibilità di disporre di differenti
arrangiamenti di MHC adatti a presentare un maggior numero di antigeni ai
linfociti. La seconda ipotesi prevede che gli alleli rari siano selezionati
positivamente (per il fatto che i patogeni sono adattati a sfuggire alle forme più
frequenti di molecole MHC), e che quindi si instauri un classico ciclo ospite-
parassita di selezione frequenza dipendente.
I risultati della presente ricerca hanno evidenziato come in generale le
sequenze analizzate hanno un elevato valore di variabilità a livello aplotipico, ma
certamente interessante risulta essere una differenza tra la variabilità attesa e
quella osservata nei camosci di Auronzo morti per rogna. Le analisi degli aplotipi
mostrano infatti un valore di eterozigosità osservata piuttosto elevato H0 = 0,7863,
con un cospicuo numero di alleli (19).
La selezione non sembrerebbe favorire gli eterozigoti ma in questo caso
sembrerebbero selezionati positivamente più gli individui portatori di un allele raro.
Inoltre, si potrebbe pensare che per un camoscio possedere l’allele più frequente
(Ruru-DRB*01), forse già selezionato in precedenti episodi epidemici e ancora
efficiente contro il parassita, potrebbe aver conferito una maggiore resistenza. Ma
analizzando il genotipo si è riscontrato che questo aplotipo è frequente in
omozigosi tra gli individui morti di rogna. Al contrario, è il genotipo Ruru-DRB*01-
Ruru-DRB (non 01), ovvero il genotipo eterozigote per l’allele Ruru-DRB*01 in
combinazione con un qualsiasi altro allele, che nel campione di animali ammalati
risulta in numero inferiore (Tab. 10).

100
Codice Genotipo
1(rr1) Ruru-DRB*01
0(rr2) Ruru-DRB*01/ Ruru-DRB*41
0+(rr3) Ruru-DRB*42/ Ruru-DRB*43
122/97(rr4) Ruru-DRB*01
123/97(rr5) Ruru-DRB*01/ Ruru-DRB*44
128/98(rr6) Ruru-DRB*01
129/98(rr7) Ruru-DRB*04/ Ruru-DRB*45
130/98(rr8) Ruru-DRB*01/ Ruru-DRB*27
132/98(rr9) Ruru-DRB*01/ Ruru-DRB*33
135/98(rr10) Ruru-DRB*01/ Ruru-DRB*18
19/08/1997(rr11) Ruru-DRB*04/ Ruru-DRB*46
2aaf(rr12) Ruru-DRB*01
2aam(rr13) Ruru-DRB*01
3aaf97(rr14) Ruru-DRB*01/ Ruru-DRB*27
3aam98(rr15) Ruru-DRB*01/ Ruru-DRB*27
3aam97(rr16) Ruru-DRB*17/ Ruru-DRB*47
3aam98(rr17) Ruru-DRB*48/ Ruru-DRB*49
4aaf97(rr18) Ruru-DRB*50/ Ruru-DRB*33
5aaf98(rr19) Ruru-DRB*41/ Ruru-DRB*51
6aam98(rr20) Ruru-DRB*01
6aam97(rr21) Ruru-DRB*18/ Ruru-DRB*51
7aaf98(rr22) Ruru-DRB*01/ Ruru-DRB*09
9aam97(rr23) Ruru-DRB*01
9m(rr24) Ruru-DRB*41/ Ruru-DRB*52
NS(rr67) Ruru-DRB*01
Tabella 10 Individui omozigoti prelevati da Auronzo 1997/98 morti per rogna sarcoptica e relatipo genotipo
Sebbene ulteriori analisi siano necessarie, anche attraverso l’utilizzo di
campioni più numerosi, questo risultato è molto interessante, in quanto
sembrerebbe indicare che gli animali meglio resistenti all’epidemia del 1997/98
sono stati quelli eterozigoti che possedevano però un allele Ruru-DRB*01. Quindi,
si tratterebbe di un processo di selezione a favore di uno specifico set di
eterozigoti, che sarebbero in grado di garantire una maggiore possibilità di
riconoscimento dell’antigene, ma anche, grazie alla presenza di uno specifico
aplotipo (Ruru-DRB*01 nel nostro caso), di resistere ad una malattia prevalente. La
complessità della relazione tra variabilità del sistema MHC e diversità dei ceppi
patogeni è confermata da diversi studi (Paterson et al., 1998; Langefors et al.,
2001; Bernatchez & Landry 2003), e quindi sarebbe molto interessante

101
approfondire meglio l’ipotesi avanzata in questa tesi sulla situazione del camoscio
alpino.
Altro punto da discutere è il risultato della diversità genetica riscontrata nel
campione della Provincia di Lecco relativa agli individui prelevati successivamente
all’epidemia di polmonite che colpi questa regione nel 2004. Gli individui
sopravvissuti hanno un valore di H0 = 0,469, inferiore alla diversità genetica
riscontrata nelle popolazioni campionate prima dell’epidemia di polmonite della
stessa regione (H0 = 0,6488) e delle popolazioni sopravvissute (H0 = 0,674) e
resistenti (H0 = 0,757) alla rogna sarcoptica della Provincia di Belluno. Un’ipotesi
plausibile potrebbe essere l’effetto del collo di bottiglia post epidemico. L’epidemia
avrebbe causato la perdita di variabilità genetica e quindi di eterozigosi, In questo
campione, infatti, si riscontra il più alto numero di omozigosi (13) rispetto al
campione prelevato prima del crash epidemico (9) e agli altri campioni della
Provincia di Belluno (tra 1 e 8). Questo dato concorda con i risultati di altri Autori
(Hedrick et al. 2000; Mikko & Andersson 1995; Mikko et al. 1999; Schmulder et al.
2003; Schaschal et al. 2005).
A livello inter specifico si è notata una maggiore variabilità genetica per il
camoscio alpino rispetto al camoscio pirenaico. Questo risultato è in accordo con
altri Autori (Alvarez-Busto et al. 2007) ma è probabilmente anche connesso alla
dimensione del campione esaminato (127 individui italiani contro 55 individui
spagnoli). La ridotta variabilità genetica delle MHC è stata riscontrata anche in studi
su altri ungulati a seguito di colli di bottigli (Ellegren et al. 1993; Mikko & Andersson
1995, Mikko et al. 1999).
Gli aplotipi Ruru-DRB*01- Rupy-DRB*02, infine, oltre a risultare i più comuni tra
gli individui campionati, vengono condivisi dalle due specie esaminate sulla base
dei 206 nucleotidi esaminati nel presente studio. Si tratta, probabilmente, di un

102
gene ancestrale ereditato dalle due specie a seguito di selezione conservativa come
suggerito da Doxiadis et al 2006.
4.2 Variabilità nucleotidica delle popolazioni di camoscio
La selezione positiva dell’esone 2 delle MHC è determinante per entrambe le
specie esaminate. La variabilità dei riarrangiamenti amminoacidici corrispondenti al
sito di legame della regione PBR dell’MHC che permette il riconoscimento di
antigeni provenienti da patogeni, è fondamentale per il persistere di una
popolazione. Maggiore è la variabilità di questa regione maggiore è la possibilità di
rispondere a patogeni diversi.
Tra i 58 aplotipi trovati, tutti codificanti, sono state identificate 24 posizioni
variabili. Sul totale, 8 variazioni cadono in prima posizione codonica, 14 in seconda
posizione codonica e solo 2 in terza posizione codonica. Questa disposizione dei siti
variabili nelle prime due posizioni delle triplette codoniche è piuttosto inusuale
nelle sequenze codificanti. Nelle regioni genomiche che codificano per proteine, la
variabilità genetica si accumula in genere nella terza posizione del codone, poiché
le variazioni nella prima e seconda posizione della tripletta viene in genere
selezionata negativamente, in quanto in grado di alterare la funzionalità della
proteina codificata. Al contrario nella regione del gene DRB, la variazione
nucleotidica che altera la sequenza aminoacidica codificata viene selezionata
positivamente (Yang & Bielawski 2000; Hughes & Nei 1988, 1989; Bergstrom &
Gyllensten 1995; Bernatchez & Landry 2003), poiché essa corrisponde a una
maggiore possibilità di riconoscere antigeni diversi e per questo motivo contribuire
a una maggiore efficienza nel riconoscimento di patogeni diversi e/o di varianti
dello stesso organismo patogeno.

103
L’analisi condotta mediante AMOVA (Analysis of MOlecular VAriance) permette
di collegare la variazione genica con caratteristiche biologiche scelte volta per
volta. Nel corso del presente studio le popolazioni sono state suddivise in due
gruppi, quelle sensibili agli eventi epidemici, sia da rogna sarcoptica sia da
polmonite dalla Provincia di Lecco, e quelle in qualche modo considerate resistenti,
includendo anche quella popolazione di camoscio sopravvissuta all’epidemia di
polmonite. I risultati di tale analisi, non hanno evidenziato un differenziamento
significativo nell’AMOVA. Tuttavia, i valori di varianza molecolare osservati nel
confronto sani/malati (3,36 della percentuale di varianza) è maggiore della varianza
tra popolazioni nell’ambito dei due gruppi sopra citati (0,5). Questo dato, sebbene
preliminare, non permette di rifiutare l’ipotesi che i dati in nostro possesso
possano evidenziare una varianza molecolare significativa tra le popolazioni
resistenti e quelle suscettibili.
Inoltre, i dati indicherebbero un sostanziale e significativo differenziamento
genetico della popolazione resistente di Calalzo.

104

105
CONCLUSIONI
Lo studio condotto ha permesso di analizzare in profondità la struttura genetica
di diverse popolazioni di camoscio provenienti dal Parco delle Dolomiti Bellunesi,
anche a confronto con popolazioni provenienti dalla Provincia di Lecco, prima e
dopo un evento epidemico di polmonite. In più ha contribuito alla maggiore
conoscenza della variabilità genetica delle MHC della specie Rupicapra p.
pyrenaica.
Sebbene l’analisi effettuata non abbia permesso di individuare con certezza
aplotipi e/o domini caratteristici delle popolazioni resistenti, e che pertanto
potessero essere utilizzati come markers per la resistenza agli organismi patogeni, i
risultati ottenuti hanno condotto a una definizione della struttura genetica delle
popolazioni di camoscio caratterizzate da un diverso rapporto con le infestazioni
dovute all’agente eziologico della rogna sarcoptica, Sarcoptes scabiei. Di particolare
utilità è stato il confronto con le popolazioni di camoscio dalla Provincia di Lecco, in
quanto campionate in due momenti diversi, prima e dopo un’epidemia di
polmonite e con le popolazioni di camoscio pirenaico affette da pestivirus e da altre
patologie.
Numerosi autori hanno evidenziato che lo studio della variabilità genetica nelle
popolazioni ospiti fornisce spesso indicazioni contrastanti, dipendenti dal modello
ospite-parassita, dalla possibilità di co-infezioni simultanee e dalla variabilità
genetica della popolazione parassita. Alcuni patogeni di origine virale e batterica
possono indurre, grazie alla loro variabilità, un aumento dell’eterozigosi, in
considerazione del fatto che gli individui eterozigoti dimostrerebbero un vantaggio

106
selettivo nella risposta al patogeno. Questo vantaggio dell’eterozigote non è stato
però confermato in sistemi ospite/parassita dove il patogeno era un organismo
pluricellulare, complesso.
Le infestazioni da rogna sarcoptica, provocando un’elevata mortalità,
potrebbero vanificare l’effetto del vantaggio conferito agli individui e alle
popolazioni più variabili, poiché il collo di bottiglia determinato dalla mortalità degli
individui porta necessariamente a una riduzione sensibile della variabilità genetica.
Si ritiene fondamentale un monitoraggio continuato nel tempo, che possa
evidenziare cambiamenti nella struttura genetica delle popolazioni e il selezionarsi
di individui capaci di resistere alle infezioni parassitarie, a confronto con la
dinamica delle popolazioni, con l’andamento demografico e con altri fattori (clima,
altitudine, effetti dell’attività dell’uomo) che possano influenzarne le
caratteristiche.

107
Riferimenti bibliografici
Acevedo-Whitehouse K, Gulland F, Greig D, Amos W (2003) Disease susceptibility in
California sea lions. Nature 422:35
Altizer S, Harvell D, Friedle E (2003) Rapid evolutionary dynamics and disease
threats to biodiversity. Trends Ecol Evol 18:589-596
Alvarez-Busto J, Garcı´a-Etxebarria K, Herrero J, Garin I, Jugon BM (2007) Diversity
and evolution of the Mhc-DRB1 gene in the two endemic Iberian subspecies of
Pyrenean chamois, Rupicapra pyrenaica. Heredity 99:406-413
Amos W, Wilmer J, Fullard K, et al. (2001) The influence of parental relatedness on
reproductive success. Proc R Soc Lond B 268:2021-2027
Andersson L & Mikko S (1995) Generation of MHC class II diversity by intra- and
intergenic recombination. Immunol Rev 143:5–12
Apanius V, Penn D, Slev PR, Ruff LR, Potts WK (1997) The nature of selection of the
major histocompatibility complex. Crit Rev Immunol 17:179–224
Avise JC (2000) Phylogeography. Cambridge MA, Harvard University Press.
Bakker TC, Zbinden M. (2001). Counting on immunity. Nature. 414 (6861): 262-263.
Balloux F & Lugon-Moulin N (2002) The estimation of population differentiation
with microsatellite markers. Mol Ecol 11:155-165
Bergström TF & Gyllensten U (1995) Evolution of Mhc class II polymorphism: the
rise and fall of class II gene function in primates. Immunol Rev 143:13-31

108
Bergström TF, Josefsson A, Erlich HA, Gyllensten U (1998) Recent origin of HLA-
DRB1 alleles and implications for human evolution. Nat Genet 18:237–242
Bernatchez L & Landry C (2003) MHC studies in nonmodel vertebrates: what have
we learned about natural selection in 15 years? J Evol Biol 16: 363-377
Berrilli F, D’Amelio S, Rossi L (2002) Ribosomal and mitochondrial DNA sequence
variation in Sarcoptes mites from different hosts and geographical regions.
Parasitol Res 88:772–777
Biilsma R, Bundgaard J, Boerema AC (2000) Does inbreeding affect the extinction
risk of small populations?: predictions from Drosophila. J Evol Biol 13:502-514
Blouin MS, Parsons M, Lacaille V, Lotz S (1996) Use of microsatellite loci to classify
individuals by relatedness. Mol Ecol 5:393-401
Bonneaud C, Mazuc J, Chastel O, Westerdahl H, Sorci G (2004) Terminal investment
induced by immune challenge and fitness traits associated with major
histocompatibility complex in the house sparrow. Evolution 58(12):2823-30
Brewer BA, Lacy RC, Foster ML, Alaks G (1990) Inbreeding depression in insular and
central populations of Peromyscus mice. J Hered 81:257-266
Briles WE, Stone HA, Cole RK (1977) Marek's disease: effects of B histocompatibility
alloalleles in resistant and susceptible chicken lines. Science 195(4274):193-5
Brown JH, Jardetzky TS, Saper MA, Samraoui B, Bjorkman PJ, Wiley DC (1988) A
hypothetical model of foreign antigen binding site of class II histocompatibility
molecules. Nature 332:845-850

109
Brown JH, Jardetzky TS, Gorga JC, Stern LJ, Urban RG, Strominger JL, Wiley DC
(1993) Three-dimensional structure of the human class II histocompatibility antigen
HLA-DR1. Nature 364:33-39
Brown JL & Eklund A (1994) Kin recognition and the major histocompatibility
complex: an integrative review. Am Nat 143:435-461
Brumfield RT, Beerli P, Nickerson DA, Edwards SV (2003) The utility of single
nucleotide polymorphisms in inferences of population history. Trends Ecol Evol
18:249-256
Cassinello J, Gomendio M, Roldan ERS (2001) Relationship between coefficient of
inbreeding and parasite burden in endangered gazelles. Conserv Biol 15:1171-1174.
Clarke B & Kirby DR (1966) Maintenance of histocompatibility polymorphisms.
Nature 211:999-1000
Cohen S (2002) Strong positive selection and habitat-specific amino acid
substitution patterns in MHC from an estuary fish under intense pollution stress.
Mol Biol Evol 19:1870-1880
Coltman DW, Pilkington JG, Smith JA, Pemberton JM (1999) Parasitemediated
selection against inbred Soay sheep in a free-living, island population. Evol
53:1259-1267
Coulson T, Albon S, Slate J, Pemberton J (1999) Microsatellite loci reveal sex-
dependent responses to inbreeding and outbreeding in red deer calves. Evol
53:1951-1960
Couturier MAJ (1938) Le chamois. B. Arthaud-Editeur, Grenoble. 855 pp.

110
Crandall KA, Bininda-Emonds ORP, Mace GM, Wayne RK (2000) Considering
evolutionary processes in evolutionary biology. Trends Ecol Evol 15:290-295
Crestanello B (2002/2004) Variabilità genetica, divergenza e traslocazione nel
camoscio (Rupicapra ssp.): implicazioni per la tassonomia e la gestione. Dottorato
di ricerca in biologia, Università degli studi di Ferrara. pp. 120
De Carneri I (1997) Parassitologia generale e umana. Ambrosiana ed.
Dengjel J, Schoor O, Fischer R, Reich M, Kraus M, et al. (2005) Autophagy promotes
MHC class II presentation of peptides from intracellular source proteins. Proc Nat
Acad Sci USA 102:7922-7927
Decamposlima PO, Gavioli R, Zhang QJ, Wallace LE, Dolcetti R, Rowe M, Rickinson
AB, Masucci MG (2003) HLA-A11 epitope loss isolates of Epstein-Barr-virus from a
highly A11+ population. Science 260:98-100
Dolan JM (1963) Beitrag zur systematischen Gliederung des Tribus Kupicupini
Simpson 1945. Zeitschrift fur zoologische Systematik und Evolutionsforschung. 1:
311-407
Doxiadis GG, Rouweler AJ, de Groot NG, Louwerse A, Otting N, Verschoor EJ et al.
(2006). Extensive sharing of MHC class II alleles between rhesus and cynomolgus
macaques.Immunogenetics 58: 259–268
Dudash MR & Fenster CB (2000) Inbreeding and outbreeding depression. In
Genetics, Demography and Viability of Fragmented Populations. Edited by: Clarke
G. Cambridge, Cambridge University Press.

111
Dutia BM, McConnell I, Ballingall KT, Keating P, Hopkins J (1994) Evidence of the
expression of two distinct MHC class II DRB like molecules in the sheep. Anim Genet
25:35–241
Edwards SV, Potts WK (1996) Polymorphism of genes in the major
histocompatibility complex (MHC): implications for conservation genetics. In
Molecular Genetic Approaches in Conservation Edited by: Smith TB and Wayne RK.
New York, Oxford University Press 214-237
Ellegren H, Hartman G, Johansson M, Andersson L (1993) Major histocompatibility
complex monomorphism and low levels of DNA fingerprinting variability in a
reintroduced and rapidly expanding population of beavers. Proc Natl Acad Sci USA
90: 8150–8153
Ellengren H, Mikko S, Wallin K, Andersson L (1996) Limited polymorphism at major
histo-compatibility complex (MHC) loci in the Swedish moose A. alces. Mol Ecol
5:3–9
Ellstrand NC & Elam DR (1993) Population genetic consequences of small
population size-implications for plant conservation. Ann Rev Ecol Syst 24: 217-242
Excoffier L, Laval G, Schneider S (2005) Arlequin (version 3.0): anintegrated
software package for population genetics data analysis. Evolutionary Bioinformatics
1:47–50
Falk DA & Holsinger KE eds (1991) Genetics and Conservation of rare plants. Oxford
University press, New York.
Frankham R & Ralls K (1998) Inbreeding leads to extinction. Nature 392:441-442

112
Frankham R, Ballou JD, Briscoe DA (2004) A Primer of Conservation Genetics.
Cambridge University Press. Cambridge UK
Gagliardi G, Pacetti AI, Cappelli C (1985) Parchi naturali: significato biologico di
malattia nelle popolazioni animali protette. In: Proc Inter Symp “La
cheratocongiuntivite infettiva del camoscio” Vercelli: pp. 159-162
Gyllensten UB, Sundvall M, Erlich HA (1991) Allelic diversity is generated by
intraexon sequence exchange at the DRB1 locus of primates. Proc Natl Acad Sci
USA 88: 3686–3690
Godot V, Harraga S, Beurton I, Tiberghien P, Sarciron E, Gottstein B, Vuitton DA
(2000) Resistance/susceptibility to Echinococcus multilocularis infection and
cytocine profile in humans. II. Influence of the HLA B8, DR3, DQ2 haplotype. Clin
Exp Immunol 121:491-498
Guberti V & Zamboni L (2000) Can the host restistance hypothesis explain the cyclic
patterns observed in Sarcoptes scabiei in Chamois (Rupicapra rupicapra)?
Parassitologia 42 (suppl.): 72
Gutierrez-Ezpeleta GA, Hedrick PW, Kalinowski ST, Garrigan D, Boyce WM (2001). Is
the decline of desert bighorn sheep from infectious disease the result of low MHC
variation? Heredity 86: 439–450
Halperin E & Eskin E (2004) Haplotype reconstruction from genotype data using
Imperfect Phylogeny. Bioinformatics 20:1842–1849
Harf R & Sommer S (2005) Association between MHC Class II DRB alleles and
parasite load in the hairy-footed gerbil, Gerbillurus paeba, in the Southern Kalahari.
Mol Ecol 14:85-91

113
Harrison S & Hastings A (1996) Genetic and evolutionary consequences of a
metapopulation structure. Trends Ecol Evol 11:180-183
Hedrick PW (1994) Evolutionary genetics at the major histocompatibility complex.
Am Nat 143:945-964
Hedrick PW (1999) Balancing selection and the MHC. Genetica 104:207-214
Hedrick PW (2002) Pathogen resistance and genetic variation at MHC loci. Evolution
Int J Org Evolution 56 (10): 1902-1908
Hedrick PW (2004) Comment on 'parasite selection for immunogenetic optimality".
Science 303:957
Hedrick PW & Thompson G (1987) Evidence for balancing selection at HLA. Genetics
104: 449-456
Hedrick PW, Parker KM, Gutierrez Espeleta GA, Rattink A, Lievers K (2000) Major
histocompatibility complex variation in the Arabian oryx. Evolution 54:2145–2151
Hedrick PW, Kim TJ, Parker KM (2001a) Parasite resistance and genetic variation in
the endangered Gila topminow. Anim Cons 4:103-109
Hedrick PW, Parker KM, Miller EL, Miller PS (2001b) Major histocompatiblity
complex variation in the endangered Przewalski's horse. Genetics 152:1701-1710
Hill AVS, Allsopp CEM, Kwiatkowski D, Anstey NM, Tumasi P, Rowe PA, Bennett S,
Brewster D, McMichael AJ, Greenwood BM (1991) Common West-African HLA
antigens are associated with protection from severe malaria. Nature 352:595-600.
Hughes A & Hughes M (1995) Natural selection on the peptide-binding regions of
major histocompatibility complex molecules. Immunogenetics 42:233-243

114
Hughes AL & Nei M (1988) Pattern of nucleotide substitution at major
histocompatibility complex class I loci reveals overdominant selection. Nature
335:167-170
Hughes AL & Nei M (1989) Nucleotide substitution at major histocompatibility
complex class II loci: Evidence for overdominant selection. Proc Natl Acad Sci USA
86:948-962
Hughes AL & Yeager M (1998) Natural selection at major histocompatibility
complex loci of vertebrates. Ann Rev Genet 32:415-434
Hughes AL & Nei M. (1992) Models of host-parasite interaction and MHC
polymorphism. Genetics 132 (3): 863-864.
Janeway CA & Travers P (2002) Immunologie. Heidelberg, Berlin, Oxford, Spektrum
Akademischer Verlag GmbH
Jeffery KJ, Bangham CR (2000) Do infectious diseases drive MHC diversity? Microb
Infect 2:1335-41
Keller L & Waller D (2002) Inbreeding effects in wild populations. Trends Ecol Evol
17:230-241
Kimura M (1980) A simple method for estimating evolutionary rate of base
substitutions through comparative studies of nucleotide sequences. Journal of
Molecular Evolution 16:111–120
Klein J (1986) Natural History of the Major Histocompatibility Complex. New York,
Wiley & Son
Klein J & Horejsi V (1997) Immunology. Oxford, Blackwell Science

115
Lacy RC (1997) Importance of genetic variation to the viability of mammalian
populations. J Mamm 78:320-335.
Lande R (1988) Genetics and demography in biological conservation. Science
241:1455-1460
Langefors A, Lohm J, Grahn M, Andersen O, von Schantz T (2001) Association
between major histocombatibility complex class IIB alleles and resistance to
Aeromonas salmonicida in Atlantic salmon. Proc R Soc Lond B 268:479-485
Larkin MA, Blackshields G, Brown NP, Chenna R, McGettigan PA, McWilliam H,
Valentin F, Wallace IM, Wilm A, Lopez R, Thompson JD, Gibson TJ, Higgins DG.
(2007) Clustal W and Clustal X version 2.0. Bioinformatics 23: 2947-2948
Lewin HA, Russell GC, Glass EJ. (1999) Comparative organization and function of the
major histocompatibility complex of domesticated cattle. Immunol Rev 167: 145-
158
Librado P & Rozas J (2009) DnaSP v5: A software for comprehensive analysis of DNA
polymorphism data. Bioinformatics 25: 1451-1452
Lydekker R (1913) Artiodactyla, family Bovidae, subfamilies Bovinae to Ovibovinae
(cattle, sheep, goats, chamois, serows, takin, musk-oxen, etc.). In: Catalogue of the
ungulate mammals in the British Museum (Natural History). Trustees of the British
Museum (Nat. Hist.), London. Vol. I. 249 pp.
Little TJ (2002) The evolutionary significance of parasitism: do parasite-driven
genetic dynamics occur ex silico? J Evol Biol 15:1-9
Lively CM & Dybdahl MF (2000) Parasite adaptation to locally common host
genotypes. Nature 405:679-681

116
López-Olvera JR, D’Amelio S, Ignasi M, Cavallero S, Fernández L (2009)
Characterization of MHC class II DRB1 allelic diversity in Pyrenean chamois
(Rupicapra pyrenaica pyrenaica) Santiago Lavín Atti V World Conference on
Mountain Ungulates. Granada
Lowe A, Harris S, Ashton P (2004) Ecological Genetics: Design, Analysis, and
Application. Oxford, UK, Blackwell Publishing Ltd.
Lovari S (1985) Behavioural repertoire of the Abruzzo Chamois (Rupicapra
pyrenaica ornata). Saugetierkundliche Mitteilungen 32:113-136
Lovari S (1987) Evolutionary aspects of the biology of chamois, Rupicapra spp.
(Bovidae, Caprinae). In: The biology and management of Capricornis and related
mountain antelopes. Soma H. (ed). Croom-Helm: London. pp 51-61
Lovari S & Scala C (1980) Revision of Rupicapra Genus. I. A statistical reevaluation
of Couturier’s data on the morphometry of six chamois subspecies. Boll Zool
47:113–124
Masini F & Lovari S (1988) Systematics, Phylogenetic Relationships and dispersal of
the Chamois (Rupicapra spp.). Quaternary Research 30:339-349
McClelland EE, Penn DJ, Potts WK (2003) Major histocompatibility complex
heterozygote superiority during coinfection. Infect Immun 71:2079-2086
Meagher S (1999) Genetic diversity and Capillaria hepatica (Nematoda) prevalence
in Michigan deer mouse populations. Evol 53:1318-1324
Meffe GK & Carroll CG (1997) contributors: Principles of Conservation Biology. 2nd
edition. Sunderland, Massachusetts, Sinauer Associates.

117
Meyers LA & Bull JJ (2002) Fighting change with change: adaptive variation in an
uncertain world. Trends Ecol Evol 17:551-557
Miller KM, Kaukinen KH, Beacham TD, Withler RE (2001) Geographic heterogeneity
in natural selection on an MHC locus in sockeye salmon. Genetica 111:237-257
Mikko S & Andersson L (1995) Low major histocompatibility comlex class II diversity
in European and North American moose. Proc Natl Acad Sci US 92: 4259–4263
Mikko S, Roed K, Schmutz S, Andersson L (1999) Monomorphism and
polymorphism at MHC DRB loci in domestic and wild ruminants. Immunol Rev 167:
169–178
Mona S, Crestanello B, Bankhead-Dronnet S, Pecchioli E, Ingrosso S, D’AmelioS,
Rossi L, Meneguz PG, Bertorelle G (2008) Disentangling the effects of
recombination, selection, and demography on the genetic variation at a major
histocompatibility complex class II gene in the alpine chamois. Mol Ecol 17:4053-
4067
Morin PA, Luikart G, Wayne RK, the SNP workshop group (2004) SNPs in ecology,
evolution and conservation. Trends Ecol Evol 19:208-216
Mullis K, Faloona F, Scharf S, Saiki R, Horn G, Erlich H (1986) Specific enzymatic
amplification of DNA in vitro: the polymerase chain reaction. Cold Spring Harb
Symp Quant Biol 51 Pt 1: 263-73
Mustoni A, Pedrotti L, Zanon E, Tosi G (2003) Ungulati delle Alpi: Biologia,
riconoscimento e gestione. Nitida Immagine Editrice. 559 pp.
Nascetti G, Lovari S, Lanfranchi P, Berducou C, Mattiucci S, Rossi L, Bullini L (1985)
Revision of Rupicapra genus III: electrophoretic studies demonstrating species

118
distinction of chamois populations of the Alps from those of the Apennines and
Pyrenees. In: Lovari S (ed) Biology and management of mountain ungulates.
Croom–Helm, London, pp 57–62
Nei M (1987) Molecular Evolutionary Genetics. Columbia University Press, New
York, NY, USA
Ohta T (1998) On the pattern of polymorphisms at major histocompatibility
complex loci. J Mol Evol 46:633-638
Ottová E, Simková A, Martin JF, de Bellocq JG, Gelnar M, Allienne JF, Morand S
(2005) Evolution and trans-species polymorphism of MHC class IIbeta genes in
cyprinid fish. Fish Shellfish Immunol 18(3):199-222
Parham P & Ohta T (1996) Population biology of antigen presentation by MHC class
I molecules. Science 272:67–74
Paterson S, Wilson K, Pemberton JM (1998) Major histocompatibility complex
variation associated with juvenile survival and parasite resistance in a large
unmanaged ungulate population (Ovis aries L.). Evol 95:3714-3719
Peacock MM & Smith AT (1997) The effects of habitat fragmentation on dispersal
patterns, mating behavior and genetic variation in a pica (Ochotona princeps)
metapopulation. Oecol 112:524-533
Pence DB & Ueckermann E (2002) Sarcoptic mange in wildlife. Rev. Sc Epiz 21:385-
398
Penn DJ (2002) The Scent of Genetic Compatibility: Sexual Selection and the Major
Histocompatibility Complex. Ethol 108:1-21

119
Penn DJ, Damjanovich K, Potts WK (2002) MHC heterozygosity confers a selective
advantage against multi-strain infections. Proc Natl Acad Sci 99:11260-11264
Penn DJ & Fisher S (2004) Sniffing out genetic compatibility. Biologist
Penn DJ & Potts WK (1999) The evolution of mating preferences and major
histocompatibility complex genes. Am Nat 153:145-163
Pérez T, Albornoz J, Dominguez A (2002) Phylogeography oh chamois (Rupicapra
spp.) inferred from microsatellites. Mol Phylogenet Evol 25:524-34
Piertney SB & Oliver MK (2006) The evolutionary ecology of the major
histocompatibility complex.. Heredity 96(1):7-21
Potts WK & Wakeland EK (1993) Evolution of MHC genetic diversity: a tale of incest,
pestilence and sexual preference. Trends Genet 9:408-412
Primack RB (1993) Essentials of Conservation Biology. Sunderland, Massachusetts,
Sinauer Associates.
Provincia di Belluno e Università degli Studi di Padova (2003) Piano faunistico
venatorio-provinciale. Aggiornamento 2003-2008. 96 pp
Provincia di Belluno e Università degli Studi di Torino (1999) Il camoscio. Gestione e
società. Atti dell’euroconvegno 26/27 febbraio 1999. 138 pp
Raymond M & Rousset F (1995) GENEPOP Version 1.2 : population genetics
software for xact tests and ecumenicism. Journal of Heredity 86: 248-249
Reusch TB, Haberli MA, Aeschlimann PB, Milinski M (2001) Female sticklebacks
count alleles in a strategy of sexual selection explaining MHC polymorphism.
Nature 414 (6861):300-2

120
Reusch TBH, Schaschl H, Wegner KM (2004) Recent duplication and inter-locus
gene conversion in major histocompatibility class II genes in a teleost, the three-
spined stickleback. Immunogenetics 56:427-437
Richman A (2000) Evolution of balanced genetic polymorphism. Mol Ecol 9:1953-
1963
Richman AD, Herrera LG, Nash D, Schierup HM (2003) Relative roles of mutation
and recombination in generating allelic polymorphism at an MHC class II locus in
Peromyscus maniculatus. Genet Res Camb 82:89–99
Robinson J, Waller MJ, Parham P, de Groot N, Bontrop R, Kennedy LJ et al. (2003)
IMGT/HLA and IMGT/MHC: sequence databases for the study of the major
histocompatibility complex. Nucleic Acids Res 31:311–314
Rodolfi M (1990) Foresta di Tarvisio. Epizoozia di rogna sarcoptica dei camosci.
Aggiornamento 1990. Unpublished Report, Ministero Agricoltura e Foreste, Ufficio
Amministrazione Tarvisio. 34 pp.
Rossi L, Meneguz PG, De Martin P, Rodolfi M (1995) The epizootiology of sarcoptic
mange in chamois Rupicapra rupicapra, from the Italian Eastern Alps.
Parassitologia 37: 233-240
Saccheri I, Kuussaari M, Kankare M, Vikman P, Fortelius W, Hanski I (1998)
Inbreeding and extinction in a butterfly metapopulation. Nature 392
Saitou N & Nei M (1987) The neighbor-joining method: a new method for
reconstructing phylogenetic trees. Molecular Biology and Evolution 4: 406-425

121
Schaschl H, Goodman SJ, Suchentrunk F (2004) Sequence analysis of the MHC class
II DRB alleles in Alpine chamois (Rupicapra r. rupicapra). Dev Comp Immunol 91:
125–135
Schaschl H, Suchentrunk F, Hammer S, Goodman SJ (2005) Recombination and the
origin of sequence diversity in the DRB MHC Class II locus in chamois (Rupicapra
sp.). Immunogenetics 57: 108–115
Schaschl H, Wandeler P, Suchentrunk F, Obexer-Ruff G and Goodman SJ (2006)
Selection and recombination drive the evolution of MHC class II DRB diversity in
ungulates. Heredity 97: 427–437
Schmulder MJM, Snoek LB, Booy G, Vosman B (2003) Complete loss of MHC genetic
diversity in the common hamster (Cricetus cricetus) population in The Netherlands:
consequences for conservation strategies. Conserv Genetics 4:441–451
Schonewald-Cox CM, Chambers SM, MacBryde B, Thomas L (1983) Genetics and
conservation. Benjamin Cummings, Menlo Park. California, USA.
Schwaiger FW, Weyers E, Epplen C, Brun J, Ruff G, Crawford A, Epplen JT (1993) The
paradox of MHC-DRB exon/intron evolution: alpha-helix and beta-sheet encoding
regions diverge while hypervariable intronic simple repeats coevolve with beta-
sheet codons. J Mol Evol 37:260–72
Schwaiger FW, Maddox J, Ballingall J, Buitkamp J, Crawford AM, Dutia BM, Epplen
JT, Ferguson ED, Groth D, Hopkins J, Rhind SM, Sargan D, Wetherall J, Wright H
(1996) The ovine major histocompatibility complex. In: Schook LB, Lamont SJ,
editors. The major histocompatibility complex of domestic species. Boca Raton:
CRC p. 121–76

122
Slate J, Kruuk L, Marshall T, Pemberton J, Clutton-Brook T (2000) Inbreeding
depression influences lifetime breeding success in a wild population of red deer
(Cervus elaphus). Proc R Soc Lond B 267:1657-1662
Sommer S (2005) The importance of immune gene variability (MHC) in evolutionary
ecology and conservation. Frontiers in Zoology 2:16
Spielman D, Brook BW, Briscoe DA, Frankham R (2004) Does inbreeding and loss of
genetic diversity decrease disease resistance? Conserv Genet 5:439-448
Stockwell CA, Hendry AP, Kinnison MT (2003) Contemporary evolution meets
conservation biology. Trends Ecol Evol 18:94-101
Sunnucks P (2000) Efficient genetic markers for population biology. Trends Ecol Evol
15:199-203
Swarbrick PA, Schwaiger FW, Epplen JT, Buchan GS, Griffin JF, Crawfird AM (1995)
Cloning and sequencing of expressed DRB genes of the red deer (Cervus elaphus)
Mhc. Immunogenetics 42: 1–9
Takada T, Kikkawa Y, Yonekawa H, Amano T. (1998). Analysis of goat MHC class II
DRA and DRB genes: identification of the expressed gene and new DRB alleles.
Immunogenetics 48:408–12
Takahata N & Nei M (1990) Allelic genealogy under overdominant and frequency-
dependent selection and polymorphism of major histocompatibility complex loci.
Genetics 124:967-978.
Tamura K, Dudley J, Nei M & Kumar S (2007) MEGA4: Molecular Evolutionary
Genetics Analysis (MEGA) software version 4.0. Molecular Biology and Evolution
24:1596-1599

123
Van Den Bussche RA, Hoofer SR, Lochmiller RL (1999) Characterisation of Mhc-DRB
allelic diversity in white-tailed deer (Odocoileus virginianus) provides insight into
Mhc-DRB allelic evolution within Cervidae. Immunogenetics 49:429–37
Van Tienderen PH, de Haan AA, van der Linden G, Vosman B (2002) Biodiversity
assessment using markers for ecologically important traits. Trends Ecol Evol 17:577-
582
Ufficio Caccia e Pesca, Ripartizione foreste, Provincia Autonoma di Bolzano, 2001-
2005. Progetto rogna sarcoptica. Rapporti intermedi.
Wahlberg N, Moilanen A, Hanski I (1996) Predicting the occurrence of endangered
species in fragmented landscapes. Science 273:1536-1538
Wegner KM, Reusch TBH, Kalbe M (2003a) Multiple infections drive major
histocompatibility complex polymorphism in the wild. J Evol Biol 16:224-232
Wegner KM, Kalbe M, Kurtz J, Reusch TB, Milinski M (2003b) Parasite selection for
immunogenetic optimality. Science 301(5638):1343
Wegner KM, Kalbe M, Schaschl H, Reusch TBH (2004) Parasites and individual major
histocompatiblity diversity – an optimal choice. Microb Infect 6:1110-1116
Weir BS & Cockerham CC (1984) Estimating F-statistics for the analysis of
population structure. Evolution 38:1358-1370
Wenink PW, Groen AF, Roelke-Parker ME, Prins HHT (1998) African buffalo
maintain high genetic diversity in the major histocompatibility complex in spite of
historically known population bottlenecks. Mol Ecol 7:1315–1322
Wright S (1951) The genetical structure of populations. Annals of Eugenics 15:323-
354

124
Wright S (1965) The interpretation of population structure by F-statistics with
special regard to systems of mating. Evolution 19: 395-420
Yang Z, Bielawski (2000) Statistical methods for detecting molecular adaptation.
Trends Ecol Evol 15: 496–503
Zuk M (1990) Reproductive strategies and disease susceptibility: an evolutionary
viewpoint. Parasitol Today 6:231-233
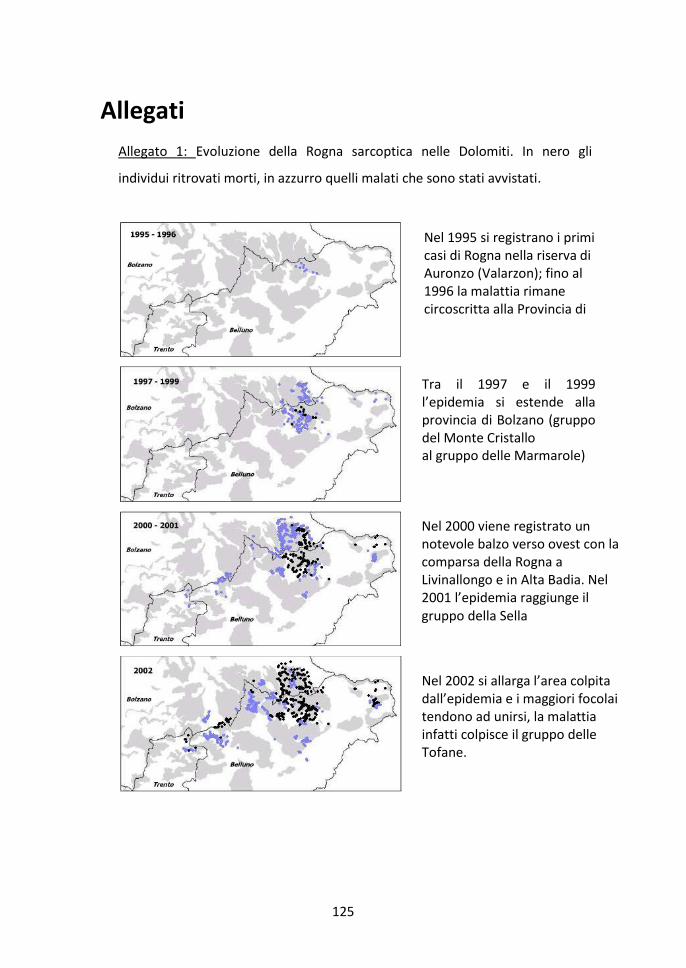
125
Allegati
Allegato 1: Evoluzione della Rogna sarcoptica nelle Dolomiti. In nero gli
individui ritrovati morti, in azzurro quelli malati che sono stati avvistati.
Nel 1995 si registrano i primi casi di Rogna nella riserva di Auronzo (Valarzon); fino al 1996 la malattia rimane circoscritta alla Provincia di Belluno.
Tra il 1997 e il 1999 l’epidemia si estende alla provincia di Bolzano (gruppo del Monte Cristallo al gruppo delle Marmarole)
Nel 2000 viene registrato un notevole balzo verso ovest con la comparsa della Rogna a Livinallongo e in Alta Badia. Nel 2001 l’epidemia raggiunge il gruppo della Sella
Nel 2002 si allarga l’area colpita dall’epidemia e i maggiori focolai tendono ad unirsi, la malattia infatti colpisce il gruppo delle Tofane.

126
Nel 2003 l’area maggiormente colpita si trova tra il gruppo delle Tofane e quello della Croda Rossa (colpendo il Parco Regole d’Ampezzo). Inoltre l’epidemia trapassa il fiume Piave.
Nel 2004 l’epidemia colpisce fortemente le province di Bolzano e Trento, mentre in provincia di Belluno sono interessate due nuove zone: il versante sud del gruppo delle Marmarole e il gruppo della Marmolada.

127
Allegato 2a: Caratteristiche specifiche dei campioni esaminati prelevati dalla
Provincia di Belluno
protocollo Codice
campione Provenienza Località sito data sesso età Status
1 rr1
Auronzo di Cadore 97/98
Dolomiti di Sesto
Val Marzon
06/01/1997
morti di rogna
O rr2 30/01/1997 M O
0+ rr3 03/03/1998 F 0+
122/97 rr4 1997
123/97 rr5 1997
128/98 rr6 1998
129/98 rr7 1998
130/98 rr8 1998
132/98 rr9 1998
135/98 rr10 1998
19/08/1997 rr11 19/08/1997 M ?
2aaf rr12 20/03/1998 F 2
2aam rr13 05/11/1997 M 2
3aaf97 rr14 08/01/1997 F 3
3aaf98 rr15 16/03/1998 F 3
3aam97 rr16 06/12/1997 M 3
3aam98 rr17 15/03/1998 M 3
4aaf98 rr18 22/03/1998 F 4
5aaf98 rr19 22/03/1998 F 5
5aam97 rr20 20/11/1997 M 5
6aam98 rr21 26/03/1998 M 5
7aaf98 rr22 12/03/1998 F 7
9aam97 rr23 18/12/1997 M 9
9m rr24 08/03/1998 F 9mesi
NS rr67 np
m NON SEQ. 12/11/1998 M
AU1 rr25
Auronzo di Cadore
Dolomiti di Sesto
Tre Cime 6/11/2004 0
sopravvissuti AU2 rr26 Maraia 12/12/2004 0
AU3 rr27 Gravasecca 18/11/2004 M
CA1 rr28
Calalzo di Cadore
Marmarole
26/9/2004 F 1 resistenti
CA2 rr37 27/11/2004 M 3
CA3 rr48 27/11/2004 F 0
CA4 rr50 4/12/2004 F 1
CA5 rr51 3/11/2004 M 2
CA6 rr52 30/9/2004 F 2
CA7 rr54 24/11/2004 M 0
CA8 rr53 Col Piccolo 16/10/2004 F 1
CA9 rr55 4/12/2004 F 1
CA10 rr29 21/11/2004 F

128
CA11 rr30 4/12/2004 M 2
CA12 rr31 8/12/2004 F 5
CA13 rr32 18/11/2004 F
CA14 rr33 5/12/2004 M 1
CA15 rr34 18/11/2004 F
CA16 rr35 Col Gran 9/10/2004 F 11
Li1 rr56
Livinallongo del Col di
Lana Migogn
Forfes 7/11/2004 M 2 resistenti
Li2 rr61 Ossario Pordoi 13/11/2004 M 2
Li6 rr63 Col Sentil Boè 19/9/2004 F 1
Li5 rr62 Gruppo di Sella 25/10/2004 M 11
Li7 rr64 Pale Monte Ornella 25/9/2004 F 9 affetto da rogna
Li8 rr65 Pizac 26/9/2004 M 1 resistenti
Li9 rr66 Pala Cuzinela 7/11/2004 M 3
Li10 rr58 Pale Monte Ornella 25/9/2004 F 1
Li11 rr59 Pale Pizac 14/10/2004 F 12
Li13 rr57 Bec De Le Forfes 17/10/2004 M 5
Li14 rr60 Selegacia Monte Foppa 4/11/2004 M 6
RP1 rr68 Rocca Pietore
Migogn Marmolada-Monzoni 23/10/2004 resistenti
RP4 rr69 6/11/2004
Au22 rr70
Auronzo di Cadore
Dolomiti di Sesto
Aiarnola 17-nov-05 F 1 post epidemico
Au23 rr71 Cengia 20-nov-05 F 2
Au24 rr72 Monte Cristallo
Pausa Marza 14-dic-05 M 0
Au25 rr73 Pausa Marza 30-nov-05 M 2
Au26 rr74
Dolomiti di Sesto
Arghena 05-nov-05 M 1
Au27 rr75 Carpi 06-nov-05 F 3
Au28 rr76 Monte Piana 19-nov-05 M 8 sopravvissuto
Au29 rr77 Pian Cavallo 12-nov-05 M 9
Au30 rr78 Ciadin Verde 16-nov-05 M 2 post epidemico
Au31 rr79 Monte
Cristallo Popena 20-nov-05
F 2
Au32 rr80
Dolomiti di Sesto
Carpi 24-nov-05 F 1
Au33 rr81 Carpi 19-nov-05 M 2
Au34 rr82 Cengia 10-dic-05 M 1
Au35 rr83 Tre Cime 07-dic-05 F 2
Au36 rr84 Cengia 09-nov-05 M 11 sopravvissuto
Au37 rr85 Monte Cristallo
Pausa Marza 23-nov-05 M 8
Au38 rr86 Pausa Marza 12-dic-05 F 2 post epidemico
Au39 NON SEQ. Dolomiti di
Sesto Cadini Misurina 31-ott-05 F 1
CA19 rr36
Calalzo di Cadore
Marmarole
Pis 10-dic-05 M 3 resistenti
CA20 rr38 Col Gran 25-set-05 F 1
CA21 rr39 Col Gran 07-dic-05 F 2
CA22 rr40 Scotter 19-nov-05 M 0
CA23 rr41 Col Gran 08-dic-05 M 5

129
CA24 rr42 Pociaval 15-set-05 F 1
CA25 rr43 Vanedel 24-set-05 F 1
CA26 rr44 Pedolas 10-dic-05 F 8
CA27 rr45 Pociaval 13-nov-05 M 3
CA28 rr46 Pociaval 12-dic-05 M 3
CA29 rr47 Pociaval 13-nov-05 M 2
CA30 rr49 Tiesse 17-nov-05 F 6
Allegato 2b: Caratteristiche specifiche dei campioni prelevati dalla Provincia di
lecco
protocollo codice
campione sito di
campionamento sesso età
Data di campionamento
Status
4359 rr87
Pizzo Cavallo
F 1 2004 post-outbreak
4360 rr88 F 7 2004 pre-outbreak
4373 rr89 F 1 2004 post-outbreak
4402 rr90 F 5 2004 pre-outbreak
4403 rr91 M 10 2004 pre-outbreak
4405 rr92 M 8 2004 pre-outbreak
4409 rr93 M 3 2004 post-outbreak
4410 rr94 F 6 2004 pre-outbreak
4486 rr95
Val Marcia
M 0 2004 post-outbreak
4487 rr96 F 2 2004 post-outbreak
4488 rr97 F 10 2004 pre-outbreak
4489 rr98 F 2 2004 post-outbreak
4493 rr99 M 1 2004 post-outbreak
4497 M 5 2004 pre-outbreak non seq
4503 rr100 F 1 2004 post-outbreak
4504 rr101 F 1 2004 post-outbreak
4505 rr102 M 1 2004 post-outbreak
4506 rr103 M 6 2004 pre-outbreak
4507 rr104 F 9 2004 pre-outbreak
4542 rr105
Campelli-Maesimo
M 3 2004 post-outbreak
4544 rr106 M 3 2004 post-outbreak
4547 rr107 F 3 2004 post-outbreak
4548 rr108 F 1 2004 post-outbreak
4553 rr109 M 3 2004 post-outbreak
4554 rr110 F 1 2004 post-outbreak

130
4555 rr111 M 1 2004 post-outbreak
4556 rr112 ignoto 0 2004 post-outbreak
4557 rr113 F 4 2004 pre-outbreak
4558 ignoto 1 2004 post-outbreak non seq
4559 M 5 2004 pre-outbreak non seq
4562 rr114 F 4 2004 pre-outbreak
4564 rr115 M 3 2004 post-outbreak
4564 rr116 M 1 2004 post-outbreak
4571 rr117 F 1 2004 post-outbreak
4572 rr118 F 4 2004 pre-outbreak
4572 rr119 M 4 2004 pre-outbreak non seq
4573 rr120 F 6 2004 pre-outbreak
4575 rr121 M 10 2004 pre-outbreak
PN62/2002 rr122
Provincia di Lecco
M adulto 2004 pre-outbreak
PN81/2002 rr123 M 5 2004 pre-outbreak
PN123/2002 rr124 M 1 2004 post-outbreak
PN322/2000 M 3 2004 pre-outbreak non seq
PN323/2000 rr125 F 3 2004 pre-outbreak
PN331/2000 rr126 F piccolo 2004 post-outbreak
PP1355/2000 rr127 piccolo 2004 post-outbreak
Allegato 2c: Caratteristiche specifiche dei campioni pervenuti dalla regione
spagnola dei Pirenei (S= individui sani; M= individui parassitati; P= individui
affetti da pestivirus)
PROTOCOLLO CODICE AREA SEX YEAR HEALTH STATUS
RP-04 rp45 CADÍ F 1991 S
RP-23 rp46 CADÍ F 1991 M
RP-02/96 rp18 FRESER F 1996 M
RP-03/99 rp21 CADÍ F 1999 S
RP-05/99 rp24 CADÍ F 1999 M
RP-C-8 rp51 CADÍ M 2000 S
RP-07/01 rp36 APA F 2001 P
RP-C-20 rp47 CADÍ F 2001 S
RP-10/02 rp37 APA F 2002 P
RP-13/02 rp40 CADÍ F 2002 M
131
RP-14/02 rp43 PN F 2002 M
RP-15/02 rp44 ARAN M 2002 M
RP-15/02 rp59 ARAN M 2002 M
RP-04/03 rp22 APA M 2003 M
RP-09/03 rp50 APA F 2003 M
RP-10/03 rp38 CADÍ F 2003 S
RP-13/03 rp41 ARAN M 2003 S
RP-01/04 rp56 ARAN M 2004 S
RP-03/04 rp19 ARAN M 2004 M
RP-04/04 rp66 ARAN M 2004 M
RP-05/04 rp67 ARAN F 2004 S
RP-05/05 rp23 CERDANYA M 2005 P
RP-06/05 rp25 CERDANYA M 2005 P
RP-06/05 rp68 CERDANYA M 2005 P
RP-12/05 rp58 CERDANYA M 2005 P
RP-26/05 rp55 ARAN F 2005 P
RP-30/05 rp60 CADÍ M 2005 S
RP-35/05 rp54 CADÍ F 2005 M
RP-49/05 rp65 ARAN M 2005 S
RP-05/06 rp63 APA F 2006 M
RP-12/06 rp64 ARAN M 2006 M
RP-15/06 rp15 APA M 2006 M
RP-16/06 rp14 APA F 2006 M
RP-18/06 rp16 CADÍ F 2006 M
RP-19/06 rp52 CADÍ-Be M 2006 M
RP-19/06 rp62 CADÍ-Be M 2006 M
RP-38/06 rp34 CADÍ-Be M 2006 P
RP-39/06 rp57 CADÍ M 2006 P
RP-48/06 rp35 CADÍ F 2006 P
RP-06/07 rp26 APA F 2007 P
RP-12/07 rp28 ARAN F 2007 M
RP-13/07 rp42 FRESER M 2007 P
RP-22/07 rp53 CADÍ M 2007 P
RP-33/07 rp29 FRESER F 2007 S
RP-34/07 rp30 FRESER M 2007 S
RP-35/07 rp31 FRESER M 2007 S
RP-36/07 rp32 FRESER F 2007 S
RP-37/07 rp33 FRESER M 2007 S

132
RP-38/07 rp27 FRESER F 2007 S
RP-01/08 rp61 FRESER F 2008 S
RP-02/08 rp17 FRESER M 2008 S
RP-03/08 rp20 FRESER F 2008 S
RP-07/08 rp48 FRESER F 2008 S
RP-08/08 rp49 FRESER F 2008 S
RP-12/08 rp39 PALLARS F 2008 P

133
Allegato 3a: Allineamento nucleotidico delle sequenze dei campioni italiani e di
quelle depositate in GenBank per R. r. rupicapra
Ruru-DRB*01 GAG TAT CAT AAG AGC GAG TGT CAT TTC TTC AAC GGG ACC GAG CGG GTG CGG TTC CTG GAC [ 60]
Ruru-DRB*02 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... T.. [ 60]
Ruru-DRB*03 ... ... AC. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Ruru-DRB*04 ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Ruru-DRB*05 ... ... .G. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Ruru-DRB*06 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Ruru-DRB*07 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Ruru-DRB*08 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Ruru-DRB*09 ... ... .G. ... ... ... ... ... ... ... ... ... ... ... ... ... ... C.. ... ... [ 60]
Ruru-DRB*10 ... ... .G. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Ruru-DRB*11 ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... .G. [ 60]
Ruru-DRB*12 ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Ruru-DRB*13 ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Ruru-DRB*14 ... ... AG. ... ... A.. ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Ruru-DRB*15 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Ruru-DRB*16 ... ... A.. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Ruru-DRB*17 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Ruru-DRB*18 ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Ruru-DRB*19 ... ... AC. ... .AA ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Ruru-DRB*20 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... C.. ... ... [ 60]
Ruru-DRB*21 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Ruru-DRB*22 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Ruru-DRB*23 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Ruru-DRB*24 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Ruru-DRB*25 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Ruru-DRB*26 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Ruru-DRB*27 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Ruru-DRB*28 ... ... .G. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Ruru-DRB*29 ... ... .G. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Ruru-DRB*30 ... ... .G. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Ruru-DRB*31 ... ... .G. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Ruru-DRB*32 ... ... A.. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Ruru-DRB*33 ... ... A.. ... .AA ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Ruru-DRB*34 ... ... AC. ... .AA ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Ruru-DRB*35 ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... C.. ... ... [ 60]
Ruru-DRB*36 ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... C.. ... ... [ 60]
Ruru-DRB*37 ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... C.. ... ... [ 60]
Ruru-DRB*38 ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Ruru-DRB*39 ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Ruru-DRB*40 ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
1SUSCET ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
0)SUSCET ... ... MM. ... .RM ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
0+SUSCETT ... ... MS. ... .RM ... ... ... ... ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
122SUSCET ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
123SUSCET ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
128SUSCET ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
129SUSCET ... ... MR. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
130SUSCET ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
132SUSCET ... ... M.. ... .RM ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
135SUSCET ... ... MR. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
190897SUS ... ... MR. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
2aaf98SUS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
2aam97SUS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
3aaf97SUS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
3aaf98SUS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
3aam97SUS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
3aam98SUS ... ... MG. ... .RM ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
4aaf98SUS ... ... M.. ... .RA ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
5aaf98SUS ... ... MM. ... .RM ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
5aam97SUS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
6aaSUSCET ... ... MR. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
7aaf98SUS ... ... .R. ... ... ... ... ... ... ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
9aam97SUS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
9mSUSCETT ... ... AS. ... .RM ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
AU1SOPRAV ... ... MM. ... .RM ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
AU2SOPRAV ... ... MM. ... .RM ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
AU3SOPRAV ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
CA1RESIST ... ... MS. ... .RM ... ... ... ... ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
CA10RESIS ... ... .R. ... ... ... ... ... ... ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
CA11RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
CA12RESIS ... ... MM. ... .RM ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
CA13RESIS ... ... .R. ... ... ... ... ... ... ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
CA14RESIS ... ... .R. ... ... ... ... ... ... ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
CA15RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
CA16RESIS ... ... MR. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
CA19RESIS ... ... MM. ... .RM ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
CA2RESIST ... ... MG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
CA20RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
CA21RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]

134
CA22RESIS ... ... .R. ... ... ... ... ... ... ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
CA23RESIS ... ... .R. ... ... ... ... ... ... ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
CA24RESIS ... ... .R. ... ... ... ... ... ... ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
CA25RESIS ... ... MR. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
CA26RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
CA27RESIS ... ... MS. ... .RM ... ... ... ... ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
CA28RESIS ... ... MR. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
CA29RESIS ... ... AS. ... .RM ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
CA3RESIST ... ... .R. ... ... ... ... ... ... ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
CA30RESIS ... ... MR. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
CA4RESIST ... ... .R. ... ... ... ... ... ... ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
CA5RESIST ... ... MR. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
CA6RESIST ... ... .R. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
CA8RESIST ... ... .R. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
CA7RESIST ... ... AC. ... .AA ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
CA9RESIST ... ... MG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
Li1RESIST ... ... MM. ... .RM ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Li13RESIS ... ... MR. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Li10RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Li11RESIS ... ... .R. ... ... ... ... ... ... ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
Li14RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Li2RESIST ... ... MG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
Li5RESIST ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Li6RESIST ... ... .R. ... ... ... ... ... ... ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
Li7RESIST ... ... AS. ... .RM ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Li8RESIST ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Li9RESIST ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
NSSUSCETT ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
RP1RESIST ... ... MG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
RP4RESIST ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Au22RESIS ... ... MR. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Au23RESIS ... ... MR. ... ..Y ... ... ... ..Y ... ... ... ... ... ... ... ... ... ... ... [ 60]
Au24RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Au25RESIS ... ... .M. ... ... ... ... ... ..Y ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
Au26RESIS ... ... ... ... ... ... ... ... ..Y ... ... ... ... ... ... ... ... ... ... ... [ 60]
Au27RESIS ... ... .R. ... ... ... ... ... ... ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
Au28RESIS ... ... M.. ... .RM ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Au29RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Au30RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Au31RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Au32RESIS ... ... MR. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Au33RESIS ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Au34RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Au35RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Au36RESIS ... ... ... ... ... ... ... ... ..Y ... ... ... ... ... ... ... ... ... ... ... [ 60]
Au37RESIS ... ... .R. ... ... ... ... ... ... ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
Au38RESIS ... ... ... ... ... ... ... ... ..Y ... ... ... ... ... ... ... ... ... ... ... [ 60]
4359cavPOST ... ... MS. ... .RA ... ... ... ... ... ... ... ... ... ... ... ... ?.. ... ... [ 60]
4360cavaPRE ... ... MR. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
4373cavPOST ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
4402cavaPRE ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
4403cavaPRE ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
4405cavaPRE ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
4409cavPOST ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
4410cavaPRE ... ... MS. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
4486marPOST ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
4487marPOST ... ... MR. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
4488marCPRE ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
4489marPOST ... ... MR. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
4493marPOST ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
4503marPOST ... ... .R. ... ... ... ... ... ... ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
4504marPOST ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
4505marPOST ... ... MR. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
4506marcPRE ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
4507marCPRE ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
4542campPOS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
4544campPOS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
4547campPOS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
4548campPOS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
4553campPOS ... ... MR. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
4554campPOS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
4555campPOS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
4556campPOS ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
4557campPOS ... ... .R. ... ... ... ... ... ... ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
4562campPRE ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
4564campPOS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
4564age3camPO ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
4571campPOS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
4572mcamPRE ... ... MR. ... ... ... ... ... ... ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
4572fcamPOS ... ... MR. ... ... ... ... ... ... ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
4573campPRE ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
4575campPRE ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
PN62POSTOUT ... ... MM. ... .RM ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
PN81POSTOUT ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... C.. ... ... [ 60]
PN123OUTBRE ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
PN323OUTBRE ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]

135
PN331OUTBRE .?. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
PP1355OUTBR ... ... MR. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Ruru-DRB*01 AGA TAC TTC CAT AAT GGA GAA GAG TTG GTG CGC TTC GAC AGC GAC TGG GGC GAG TAC CGG [120]
Ruru-DRB*02 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Ruru-DRB*03 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Ruru-DRB*04 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... .T. ... [120]
Ruru-DRB*05 ... ... ... T.. ... ... ... ... ... ... ... ... ... ... ... ... ... ... .T. ... [120]
Ruru-DRB*06 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... .T. ... [120]
Ruru-DRB*07 ... ... ... ... ... ... ... ... ..C ... ... ... ... ... ... ... ... ... .T. ... [120]
Ruru-DRB*08 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Ruru-DRB*09 ... ... ... T.. ... ... ... ... .AC .C. ... ... A.. ... ... ... ... ... ... ... [120]
Ruru-DRB*10 ... ... ... T.. ... ... ... ... .AC .C. ... ... A.. ... ... ... ... ... ... ... [120]
Ruru-DRB*11 ... ... ... T.. ... ... ... ... .AC .CC ... ... A.. ... .G. ... ... ... ... ... [120]
Ruru-DRB*12 ... ... ... T.. ... ... ... ... .AC .C. ... ... A.. ... ... ... ... ... ... ... [120]
Ruru-DRB*13 ... ... ... T.. ... ... ... ... .AC .C. ... ... A.. ... ... ... ... ... ... ... [120]
Ruru-DRB*14 ... ... ... T.. ... ... ... ... .AC .G. ... ... A.. ... ... ... ... ... ... ... [120]
Ruru-DRB*15 ... ... ... T.. ... ... ... ... .AC .C. ... ... A.. ... ... ... ... ... ... ... [120]
Ruru-DRB*16 ... ... ... T.. ... ... ... ... ... ... ... ... A.. ... ... ... ... ... ... ... [120]
Ruru-DRB*17 ... ... ... ... ... ... ... ... ..C ... ... ... A.. ... ... ... ... ... ... ... [120]
Ruru-DRB*18 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Ruru-DRB*19 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Ruru-DRB*20 ... ... ... T.. ... ... ... ... .AC .C. ... ... A.. ... ... ... ... ... ... ... [120]
Ruru-DRB*21 ... ... ... T.. ... ... ... ... .AC .C. ... ... A.. ... ... ... ... ... ... ... [120]
Ruru-DRB*22 ... ... ... T.. ... ... ... ... ..C ... ... ... A.. ... ... ... ... ... ... ... [120]
Ruru-DRB*23 ... ... ... ... ... ... ... ... ..C ... ... ... ... ... ... ... ... ... ... ... [120]
Ruru-DRB*24 ... ... ... ... ... ... ... ... ..C ... ... ... ... ... ... ... ... ... ... ... [120]
Ruru-DRB*25 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Ruru-DRB*26 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Ruru-DRB*27 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Ruru-DRB*28 ... ... ... T.. ... ... ... ... .AC .C. ... ... A.. ... ... ... ... ... ... ... [120]
Ruru-DRB*29 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Ruru-DRB*30 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... .T. ... [120]
Ruru-DRB*31 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... .T. ... [120]
Ruru-DRB*32 ... ... ... T.. ... ... ... ... .AC .C. ... ... A.. ... ... ... ... ... ... ... [120]
Ruru-DRB*33 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Ruru-DRB*34 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Ruru-DRB*35 ... ... ... T.. ... ... ... ... .AC .C. ... ... A.. ... ... ... ... ... ... ... [120]
Ruru-DRB*36 ... ... ... T.. ... ... ... ... .AC .C. ... ... A.. ... ... ... ... ... ... ... [120]
Ruru-DRB*37 ... ... ... T.. ... ... ... ... .AC .C. ... ... A.. ... ... ... ... ... ... ... [120]
Ruru-DRB*38 ... ... ... T.. ... ... ... ... .AC .C. ... ... A.. ... ... ... ... ... ... ... [120]
Ruru-DRB*39 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Ruru-DRB*40 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... .T. ... [120]
1SUSCET ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
0)SUSCET ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
0+SUSCETT ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
122SUSCET ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
123SUSCET ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
128SUSCET ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
129SUSCET ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... .T. ... [120]
130SUSCET ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
132SUSCET ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
135SUSCET ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
190897SUS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... .T. ... [120]
2aaf98SUS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
2aam97SUS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
3aaf97SUS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
3aaf98SUS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
3aam97SUS ... ... ... ... ... ... ... ... ..S ... ... ... A.. ... ... ... ... ... ... ... [120]
3aam98SUS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... .W. ... [120]
4aaf98SUS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
5aaf98SUS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
5aam97SUS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
6aaSUSCET ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
7aaf98SUS ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
9aam97SUS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
9mSUSCETT ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
AU1SOPRAV ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
AU2SOPRAV ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
AU3SOPRAV ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
CA1RESIST ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
CA10RESIS ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
CA11RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
CA12RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
CA13RESIS ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
CA14RESIS ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
CA15RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
CA16RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... .W. ... [120]
CA19RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
CA2RESIST ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... .W. ... [120]
CA20RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
CA21RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
CA22RESIS ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
CA23RESIS ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
CA24RESIS ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
CA25RESIS ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]

136
CA26RESIS ... ... ... ... ... ... ... ... ..S ... ... ... R.. ... ... ... ... ... ... ... [120]
CA27RESIS ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
CA28RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... .W. ... [120]
CA29RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... .W. ... [120]
CA3RESIST ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
CA30RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... .W. ... [120]
CA4RESIST ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
CA5RESIST ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... .W. ... [120]
CA6RESIST ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
CA8RESIST ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
CA7RESIST ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
CA9RESIST ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... .W. ... [120]
Li1RESIST ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Li13RESIS ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
Li10RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Li11RESIS ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
Li14RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Li2RESIST ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
Li5RESIST ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Li6RESIST ... ... ... Y.. ... ... ... ... .WR .Y. ... ... R.. ... ... ... ... ... ... ... [120]
Li7RESIST ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... .W. ... [120]
Li8RESIST ... ... ... ... ... ... ... ... ..S ... ... ... R.. ... ... ... ... ... ... ... [120]
Li9RESIST ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
NSSUSCETT ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
RP1RESIST ... ... ... T.. ... ... ... ... .AC .C. ... ... A.. ... ... ... ... ... ... ... [120]
RP4RESIST ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Au22RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Au23RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Au24RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Au25RESIS ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
Au26RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Au27RESIS ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
Au28RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Au29RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Au30RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Au31RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Au32RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... .W. ... [120]
Au33RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... .T. ... [120]
Au34RESIS ... ... ... ... ... ... ... ... ..S ... ... ... R.. ... ... ... ... ... ... ... [120]
Au35RESIS ... ... ... ... ... ... ... ... ..S ... ... ... R.. ... ... ... ... ... ... ... [120]
Au36RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Au37RESIS ... ... ... T.. ... ... ... ... .AC .C. ... ... A.. ... ... ... ... ... ... ... [120]
Au38RESIS ... ... ... ... ... ... ... ... ..S ... ... ... R.. ... ... ... ... ... ... ... [120]
4359cavPOST ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
4360cavaPRE ... ... ... Y.. ... ... ... ... .WS .W. ... ... R.. ... ... ... ... ... ... ... [120]
4373cavPOST ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
4402cavaPRE ... ... ... T.. ... ... ... ... .AC .C. ... ... A.. ... ... ... ... ... ... ... [120]
4403cavaPRE ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
4405cavaPRE ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
4409cavPOST ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
4410cavaPRE ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
4486marPOST ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
4487marPOST ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
4488marCPRE ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
4489marPOST ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... .W. ... [120]
4493marPOST ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
4503marPOST ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
4504marPOST ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
4505marPOST ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
4506marcPRE ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
4507marCPRE ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
4542campPOS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
4544campPOS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
4547campPOS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
4548campPOS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
4553campPOS ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
4554campPOS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
4555campPOS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
4556campPOS ... ... ... T.. ... ... ... ... .AC .C. ... ... A.. ... ... ... ... ... ... ... [120]
4557campPOS ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
4562campPRE ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
4564campPOS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
4564age3camPO ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
4571campPOS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
4572mcamPRE ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
4572fcamPOS ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
4573campPRE ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
4575campPRE ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
PN62POSTOUT ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
PN81POSTOUT ... ... ... T.. ... ... ... ... .AC .C. ... ... A.. ... ... ... ... ... ... ... [120]
PN123OUTBRE ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
PN323OUTBRE ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
PN331OUTBRE ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
PP1355OUTBR ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
Ruru-DRB*01 GCG GTG GCC GAG CTG GGG CGG CCG ACC GCC GAG CAC TGG AAC AGC CAG AAG GAG ATT CTG [180]

137
Ruru-DRB*02 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... .G. ... ... [180]
Ruru-DRB*03 ... ... ... ... ... ... ... ... GA. ... ... T.. ... ... ... ... ... ... ... ... [180]
Ruru-DRB*04 ... ... ... ... ... ... ... ... GA. ... ... T.. ... ... ... ... ... ... ... ... [180]
Ruru-DRB*05 ... ... ..T ... ... ... ... ... GA. ... ... T.. ... ... ... ... ... ... ... ... [180]
Ruru-DRB*06 ... ... ... ... ... ... ... ... GA. ... ... T.. ... ... ... ... ... ... ... ... [180]
Ruru-DRB*07 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Ruru-DRB*08 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Ruru-DRB*09 ... ... ... ... ... ... ... ... GA. ... ... T.. ... ... ... ... ... ... ... ... [180]
Ruru-DRB*10 ... ... ... ... ... ... ... ... GA. ... ... T.. ... ... ... ... ... ... ... ... [180]
Ruru-DRB*11 ... ... ... ... ... ... ... .G. GA. C.. ... ... ... ... ... ... ... ... ... ... [180]
Ruru-DRB*12 ... ... ... ... ... ... ... .G. GA. A.. ... ... ... ... ... ... ... ... ... ... [180]
Ruru-DRB*13 ... ... ... ... ... ... ... .G. GA. ... ... ... ... ... ... ... ... ... ... ... [180]
Ruru-DRB*14 ... ... ... ... ... ... ... .G. GA. ... C.. ... ... ... ... ... ... ... ... ... [180]
Ruru-DRB*15 ... ... ... ... ... ... ... ... GA. ... ... T.. ... ... ... ... ... ... ... ... [180]
Ruru-DRB*16 ... ... ... ... ... ... ... ... GA. ... ... T.. ... ... ... ... ... ... ... ... [180]
Ruru-DRB*17 ... ... ... ... ... ... ... ... G.. ... ... ... ... ... ... ... ... ... ... ... [180]
Ruru-DRB*18 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Ruru-DRB*19 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Ruru-DRB*20 ... ... ... ... ... ... ... ... GA. ... ... T.. ... ... ... ... ... ... ... ... [180]
Ruru-DRB*21 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Ruru-DRB*22 ... ... ... ... ... ... ... ... G.. ... ... ... ... ... ... ... ... ... ... ... [180]
Ruru-DRB*23 ... ... ... ... ... ... ... ... G.. ... ... ..G ... ... ... ... ... ... ... ... [180]
Ruru-DRB*24 ... ... ... ... ... ... ... ... G.. ... ... ..G ... ... ... ... ... ... ... ... [180]
Ruru-DRB*25 ... ... ... ... ... ... ... ... ... ... ... T.. ... ... ... ... ... ... ... ... [180]
Ruru-DRB*26 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Ruru-DRB*27 ... ... ... ... ... ... ... .G. GA. ... ... ... ... ... ... ... ... ... ... ... [180]
Ruru-DRB*28 ... ... ... ... ... ... ... ... GA. ... ... T.. ... ... ... ... ... ... ... ... [180]
Ruru-DRB*29 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Ruru-DRB*30 ... ... ... ... ... ... ... ... GA. ... ... T.. ... ... ... ... ... ... ... ... [180]
Ruru-DRB*31 ... ... ... ... ... ... ... ... G.. ... ... T.. ... ... ... ... ... ... ... ... [180]
Ruru-DRB*32 ... ... ... ... ... ... ... .G. GA. ... ... ... ... ... ... ... ... ... ... ... [180]
Ruru-DRB*33 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Ruru-DRB*34 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Ruru-DRB*35 ... ... ... ... ... ... ... ... GA. ... ... T.. ... ... ... ... ... ... ... ... [180]
Ruru-DRB*36 ... ... ... ... ... ... ... ... GA. ... ... T.. ... ... ... ... ... ... ... ... [180]
Ruru-DRB*37 ... ... ... ... ... ... ... ... ... ... ... T.. ... ... ... ... ... ... ... ... [180]
Ruru-DRB*38 ... ... ... ... ... ... ... .G. GA. ... ... ... ... ... ... ... ... ... ... ... [180]
Ruru-DRB*39 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Ruru-DRB*40 ... ... ... ... ... ... ... ... GA. ... ... ... ... ... ... ... ... ... ... ... [180]
1SUSCET ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
0)SUSCET ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
0+SUSCETT ... ... ... ... ... ... ... ... RM. ... ... Y.. ... ... ... ... ... ... ... ... [180]
122SUSCET ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
123SUSCET ... ... ... ... ... ... ... ... R.. ... ... ... ... ... ... ... ... ... ... ... [180]
128SUSCET ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
129SUSCET ... ... ... ... ... ... ... ... GM. ... ... Y.. ... ... ... ... ... ... ... ... [180]
130SUSCET ... ... ... ... ... ... ... .S. RM. ... ... ... ... ... ... ... ... ... ... ... [180]
132SUSCET ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
135SUSCET ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
190897SUS ... ... ... ... ... ... ... ... RM. ... ... Y.. ... ... ... ... ... ... ... ... [180]
2aaf98SUS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
2aam97SUS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
3aaf97SUS ... ... ... ... ... ... ... .S. RM. ... ... ... ... ... ... ... ... ... ... ... [180]
3aaf98SUS ... ... ... ... ... ... ... .S. RM. ... ... ... ... ... ... ... ... ... ... ... [180]
3aam97SUS ... ... ... ... ... ... ... ... G.. ... ... ... ... ... ... ... ... ... ... ... [180]
3aam98SUS ... ... ... ... ... ... ... ... RM. ... ... Y.. ... ... ... ... ... ... ... ... [180]
4aaf98SUS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
5aaf98SUS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
5aam97SUS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
6aaSUSCET ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
7aaf98SUS ... ... ... ... ... ... ... ... RM. ... ... Y.. ... ... ... ... ... ... ... ... [180]
9aam97SUS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
9mSUSCETT ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
AU1SOPRAV ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
AU2SOPRAV ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
AU3SOPRAV ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
CA1RESIST ... ... ... ... ... ... ... ... RM. ... ... Y.. ... ... ... ... ... ... ... ... [180]
CA10RESIS ... ... ... ... ... ... ... ... RM. ... ... Y.. ... ... ... ... ... ... ... ... [180]
CA11RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
CA12RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
CA13RESIS ... ... ... ... ... ... ... ... RM. ... ... Y.. ... ... ... ... ... ... ... ... [180]
CA14RESIS ... ... ... ... ... ... ... ... RM. ... ... Y.. ... ... ... ... ... ... ... ... [180]
CA15RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
CA16RESIS ... ... ... ... ... ... ... ... RM. ... ... Y.. ... ... ... ... ... ... ... ... [180]
CA19RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
CA2RESIST ... ... ... ... ... ... ... ... GA. ... ... T.. ... ... ... ... ... ... ... ... [180]
CA20RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
CA21RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
CA22RESIS ... ... ... ... ... ... ... ... RM. ... ... Y.. ... ... ... ... ... ... ... ... [180]
CA23RESIS ... ... ... ... ... ... ... ... RM. ... ... Y.. ... ... ... ... ... ... ... ... [180]
CA24RESIS ... ... ... ... ... ... ... ... RM. ... ... Y.. ... ... ... ... ... ... ... ... [180]
CA25RESIS ... ... ... ... ... ... ... .S. RM. ... ... ... ... ... ... ... ... ... ... ... [180]
CA26RESIS ... ... ... ... ... ... ... ... R.. ... ... ... ... ... ... ... ... ... ... ... [180]
CA27RESIS ... ... ... ... ... ... ... ... RM. ... ... Y.. ... ... ... ... ... ... ... ... [180]
CA28RESIS ... ... ... ... ... ... ... ... RM. ... ... Y.. ... ... ... ... ... ... ... ... [180]
CA29RESIS ... ... ... ... ... ... ... ... RM. ... ... Y.. ... ... ... ... ... ... ... ... [180]

138
CA3RESIST ... ... ... ... ... ... ... ... RM. ... ... Y.. ... ... ... ... ... ... ... ... [180]
CA30RESIS ... ... ... ... ... ... ... ... RM. ... ... Y.. ... ... ... ... ... ... ... ... [180]
CA4RESIST ... ... ... ... ... ... ... ... RM. ... ... Y.. ... ... ... ... ... ... ... ... [180]
CA5RESIST ... ... ... ... ... ... ... ... RM. ... ... Y.. ... ... ... ... ... ... ... ... [180]
CA6RESIST ... ... ... ... ... ... ... ... RM. ... ... Y.. ... ... ... ... ... ... ... ... [180]
CA8RESIST ... ... ... ... ... ... ... ... RM. ... ... Y.. ... ... ... ... ... ... ... ... [180]
CA7RESIST ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
CA9RESIST ... ... ... ... ... ... ... ... GA. ... ... T.. ... ... ... ... ... ... ... ... [180]
Li1RESIST ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Li13RESIS ... ... ... ... ... ... ... .S. RM. ... ... ... ... ... ... ... ... ... ... ... [180]
Li10RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Li11RESIS ... ... ... ... ... ... ... ... RM. ... ... Y.. ... ... ... ... ... ... ... ... [180]
Li14RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Li2RESIST ... ... ... ... ... ... ... ... RM. ... ... Y.. ... ... ... ... ... ... ... ... [180]
Li5RESIST ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Li6RESIST ... ... ... ... ... ... ... ... RM. ... ... Y.. ... ... ... ... ... ... ... ... [180]
Li7RESIST ... ... ... ... ... ... ... ... RM. ... ... Y.. ... ... ... ... ... ... ... ... [180]
Li8RESIST ... ... ... ... ... ... ... ... R.. ... ... ... ... ... ... ... ... ... ... ... [180]
Li9RESIST ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
NSSUSCETT ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
RP1RESIST ... ... ... ... ... ... ... .S. GA. ... ... Y.. ... ... ... ... ... ... ... ... [180]
RP4RESIST ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Au22RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Au23RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Au24RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Au25RESIS ... ... ... ... ... ... ... ... RM. ... ... Y.. ... ... ... ... ... ... ... ... [180]
Au26RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Au27RESIS ... ... ... ... ... ... ... ... RM. ... ... Y.. ... ... ... ... ... ... ... ... [180]
Au28RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Au29RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Au30RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Au31RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Au32RESIS ... ... ... ... ... ... ... ... RM. ... ... Y.. ... ... ... ... ... ... ... ... [180]
Au33RESIS ... ... ... ... ... ... ... ... GA. ... ... T.. ... ... ... ... ... ... ... ... [180]
Au34RESIS ... ... ... ... ... ... ... ... R.. ... ... ... ... ... ... ... ... ... ... ... [180]
Au35RESIS ... ... ... ... ... ... ... ... R.. ... ... ... ... ... ... ... ... ... ... ... [180]
Au36RESIS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Au37RESIS ... ... ... ... ... ... ... ... RM. ... ... Y.. ... ... ... ... ... ... ... ... [180]
Au38RESIS ... ... ... ... ... ... ... ... R.. ... ... ... ... ... ... ... ... ... ... ... [180]
4359cavPOST ... ... ... ... ... ... ... ... RM. ... ... Y.. ... ... ... ... ... ... ... ... [180]
4360cavaPRE ... ... ... ... ... ... ... .S. RS. ... ... ... ... ... ... ... ... ... ... ... [180]
4373cavPOST ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
4402cavaPRE ... ... ... ... ... ... ... .S. GA. ... ... Y.. ... ... ... ... ... ... ... ... [180]
4403cavaPRE ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
4405cavaPRE ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
4409cavPOST ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
4410cavaPRE ... ... ... ... ... ... ... .S. RM. ... ... ... ... ... ... ... ... ... ... ... [180]
4486marPOST ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
4487marPOST ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
4488marCPRE ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
4489marPOST ... ... ... ... ... ... ... ... RM. ... ... Y.. ... ... ... ... ... ... ... ... [180]
4493marPOST ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
4503marPOST ... ... ... ... ... ... ... ... RM. ... ... Y.. ... ... ... ... ... ... ... ... [180]
4504marPOST ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
4505marPOST ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
4506marcPRE ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
4507marCPRE ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
4542campPOS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
4544campPOS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
4547campPOS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
4548campPOS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
4553campPOS ... ... ... ... ... ... ... .S. RM. ... ... ... ... ... ... ... ... ... ... ... [180]
4554campPOS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
4555campPOS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
4556campPOS ... ... ... ... ... ... ... .S. GA. ... ... Y.. ... ... ... ... ... ... ... ... [180]
4557campPOS ... ... ... ... ... ... ... ... RM. ... ... Y.. ... ... ... ... ... ... ... ... [180]
4562campPRE ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
4564campPOS ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
4564age3camPO ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
4571campPOS ... ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
4572mcamPRE ... ... ... ... ... ... ... ... RM. ... ... Y.. ... ... ... ... ... ... ... ... [180]
4572fcamPOS ... ... ... ... ... ... ... ... RM. ... ... Y.. ... ... ... ... ... ... ... ... [180]
4573campPRE ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
4575campPRE ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
PN62POSTOUT ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
PN81POSTOUT ... ... ... ... ... ... ... ... GA. ... ... T.. ... ... ... ... ... ... ... ... [180]
PN123OUTBRE ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
PN323OUTBRE ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
PN331OUTBRE ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
PP1355OUTBR ... ... ... ... ... ... ... .S. RM. ... ... ... ... ... ... ... ... ... ... ... [180]
Ruru-DRB*01 GAG CGG AGG CGG GCC GCG GTG GAC A [205]
Ruru-DRB*02 ... ... ... ... ... ... ... ... . [205]
Ruru-DRB*03 ... ... ... ... ... ... ... ... . [205]
Ruru-DRB*04 ... ... .A. ... ... ... ... ... . [205]
Ruru-DRB*05 ... ... .A. ... ... ... ... ... . [205]

139
Ruru-DRB*06 ... ... .A. ... ... ... ... ... . [205]
Ruru-DRB*07 ... ... .A. ... ... ... ... ... . [205]
Ruru-DRB*08 ... ... .A. ... ... ... ... ... . [205]
Ruru-DRB*09 ... .A. ... ... ... ... ... ... . [205]
Ruru-DRB*10 ... ... .A. ... ... ... ... ... . [205]
Ruru-DRB*11 ... .A. .C. ... ... ... ... ... . [205]
Ruru-DRB*12 ... .A. .C. .A. ... ... ... ... . [205]
Ruru-DRB*13 ... .A. .C. ... ... ... ... ... . [205]
Ruru-DRB*14 ... .A. .C. ... ... ... ... ... . [205]
Ruru-DRB*15 ... .A. .C. ... ... ... ... ... . [205]
Ruru-DRB*16 ... .A. ... ... ... ... ... ... . [205]
Ruru-DRB*17 ... .A. ... ... ... .A. ... ... . [205]
Ruru-DRB*18 ... .A. .C. ... ... ... ... ... . [205]
Ruru-DRB*19 ... .A. .C. ... ... ... ... ... . [205]
Ruru-DRB*20 ... .A. ... ... ... ... ... ... . [205]
Ruru-DRB*21 ... .A. .C. ... ... ... ... ... . [205]
Ruru-DRB*22 ... .A. ... ... ... .A. ... ... . [205]
Ruru-DRB*23 ... ... ... ... ... ... ... ... . [205]
Ruru-DRB*24 ... ... ... ... ... ... ... ... . [205]
Ruru-DRB*25 ... ... ... ... ... ... ... ... . [205]
Ruru-DRB*26 ... .A. .C. ... ... ... ... ... . [205]
Ruru-DRB*27 ... ... ... ... ... ... ... ... . [205]
Ruru-DRB*28 ... .A. .C. ... ... ... ... ... . [205]
Ruru-DRB*29 ... .A. .C. ... ... ... ... ... . [205]
Ruru-DRB*30 ... ... .A. ... ... ... ... ... . [205]
Ruru-DRB*31 ... ... .A. ... ... ... ... ... . [205]
Ruru-DRB*32 ... .A. .C. ... ... ... ... ... . [205]
Ruru-DRB*33 ... .A. .C. ... ... ... ... ... . [205]
Ruru-DRB*34 ... .A. ... ... ... ... ... ... . [205]
Ruru-DRB*35 ... .A. ... ... ... ... ... ... . [205]
Ruru-DRB*36 ... ... ... ... ... ... ... ... . [205]
Ruru-DRB*37 ... .A. ... ... ... ... ... ... . [205]
Ruru-DRB*38 ... .A. .C. ... ... ... ... ... . [205]
Ruru-DRB*39 ... ... ... ... ... ... ... ... . [205]
Ruru-DRB*40 ... ... .A. ... ... ... ... ... . [205]
1SUSCET ... ... ... ... ... ... ... ... . [205]
0)SUSCET ... .R. .S. ... ... ... ... ... . [205]
0+SUSCETT ... .A. .S. ... ... ... ... ... . [205]
122SUSCET ... ... ... ... ... ... ... ... . [205]
123SUSCET ... ... ... ... ... ... ... ... . [205]
128SUSCET ... ... ... ... ... ... ... ... . [205]
129SUSCET ... ... .R. ... ... ... ... ... . [205]
130SUSCET ... ... ... ... ... ... ... ... . [205]
132SUSCET ... .R. .S. ... ... ... ... ... . [205]
135SUSCET ... .R. .S. ... ... ... ... ... . [205]
190897SUS ... ... .R. ... ... ... ... ... . [205]
2aaf98SUS ... ... ... ... ... ... ... ... . [205]
2aam97SUS ... ... ... ... ... ... ... ... . [205]
3aaf97SUS ... ... ... ... ... ... ... ... . [205]
3aaf98SUS ... ... ... ... ... ... ... ... . [205]
3aam97SUS ... .A. ... ... ... .A. ... ... . [205]
3aam98SUS ... .R. .Y. ... ... ... ... ... . [205]
4aaf98SUS ... .R. .S. ... ... ... ... ... . [205]
5aaf98SUS ... .A. .S. ... ... ... ... ... . [205]
5aam97SUS ... ... ... ... ... ... ... ... . [205]
6aaSUSCET ... .A. .S. ... ... ... ... ... . [205]
7aaf98SUS ... .R. ... ... ... ... ... ... . [205]
9aam97SUS ... ... ... ... ... ... ... ... . [205]
9mSUSCETT ... .A. .C. ... ... ... ... ... . [205]
AU1SOPRAV ... .R. .S. ... ... ... ... ... . [205]
AU2SOPRAV ... .R. .S. ... ... ... ... ... . [205]
AU3SOPRAV ... ... ... ... ... ... ... ... . [205]
CA1RESIST ... .A. .S. ... ... ... ... ... . [205]
CA10RESIS ... .R. ... ... ... ... ... ... . [205]
CA11RESIS ... ... ... ... ... ... ... ... . [205]
CA12RESIS ... .A. .S. ... ... ... ... ... . [205]
CA13RESIS ... .R. ... ... ... ... ... ... . [205]
CA14RESIS ... .R. ... ... ... ... ... ... . [205]
CA15RESIS ... ... ... ... ... ... ... ... . [205]
CA16RESIS ... ... .R. ... ... ... ... ... . [205]
CA19RESIS ... .R. .S. ... ... ... ... ... . [205]
CA2RESIST ... .R. .R. ... ... ... ... ... . [205]
CA20RESIS ... ... ... ... ... ... ... ... . [205]
CA21RESIS ... ... ... ... ... ... ... ... . [205]
CA22RESIS ... .R. ... ... ... ... ... ... . [205]
CA23RESIS ... .R. ... ... ... ... ... ... . [205]
CA24RESIS ... .R. ... ... ... ... ... ... . [205]
CA25RESIS ... .A. .S. ... ... ... ... ... . [205]
CA26RESIS ... .R. ... ... ... .M. ... ... . [205]
CA27RESIS ... .A. .S. ... ... ... ... ... . [205]
CA28RESIS ... ... .R. ... ... ... ... ... . [205]
CA29RESIS ... .R. .M. ... ... ... ... ... . [205]
CA3RESIST ... .R. ... ... ... ... ... ... . [205]
CA30RESIS ... ... .R. ... ... ... ... ... . [205]
CA4RESIST ... .R. ... ... ... ... ... ... . [205]
CA5RESIST ... ... .R. ... ... ... ... ... . [205]

140
CA6RESIST ... .R. .S. ... ... ... ... ... . [205]
CA8RESIST ... .R. .S. ... ... ... ... ... . [205]
CA7RESIST ... .A. .C. ... ... ... ... ... . [205]
CA9RESIST ... .R. .R. ... ... ... ... ... . [205]
Li1RESIST ... .R. .S. ... ... ... ... ... . [205]
Li13RESIS ... .R. .S. ... ... ... ... ... . [205]
Li10RESIS ... ... ... ... ... ... ... ... . [205]
Li11RESIS ... .R. ... ... ... ... ... ... . [205]
Li14RESIS ... ... ... ... ... ... ... ... . [205]
Li2RESIST ... .A. .S. ... ... ... ... ... . [205]
Li5RESIST ... ... ... ... ... ... ... ... . [205]
Li6RESIST ... .R. ... ... ... ... ... ... . [205]
Li7RESIST ... .R. .M. ... ... ... ... ... . [205]
Li8RESIST ... .R. ... ... ... .M. ... ... . [205]
Li9RESIST ... ... ... ... ... ... ... ... . [205]
NSSUSCETT ... ... ... ... ... ... ... ... . [205]
RP1RESIST ... .A. .S. ... ... ... ... ... . [205]
RP4RESIST ... ... ... ... ... ... ... ... . [205]
Au22RESIS ... .R. .S. ... ... ... ... ... . [205]
Au23RESIS ... .R. .S. ... ... ... ... ... . [205]
Au24RESIS ... ... ... ... ... ... ... ... . [205]
Au25RESIS ... .R. ... ... ... ... ... ... . [205]
Au26RESIS ... ... ... ... ... ... ... ... . [205]
Au27RESIS ... .R. ... ... ... ... ... ... . [205]
Au28RESIS ... .R. .S. ... ... ... ... ... . [205]
Au29RESIS ... ... ... ... ... ... ... ... . [205]
Au30RESIS ... ... ... ... ... ... ... ... . [205]
Au31RESIS ... ... ... ... ... ... ... ... . [205]
Au32RESIS ... ... .R. ... ... ... ... ... . [205]
Au33RESIS ... ... .A. ... ... ... ... ... . [205]
Au34RESIS ... .R. ... ... ... .M. ... ... . [205]
Au35RESIS ... .R. ... ... ... .M. ... ... . [205]
Au36RESIS ... ... ... ... ... ... ... ... . [205]
Au37RESIS ... .A. .S. ... ... ... ... ... . [205]
Au38RESIS ... .R. ... ... ... .M. ... ... . [205]
4359cavPOST ... .A. .S. ... ... ... ... ... . [205]
4360cavaPRE ... .R. .S. ... ... ... ... ... . [205]
4373cavPOST ... ... ... ... ... ... ... ... . [205]
4402cavaPRE ... .A. .S. ... ... ... ... ... . [205]
4403cavaPRE ... ... ... ... ... ... ... ... . [205]
4405cavaPRE ... .R. .S. ... ... ... ... ... . [205]
4409cavPOST ... ... ... ... ... ... ... ... . [205]
4410cavaPRE ... .R. .S. ... ... ... ... ... . [205]
4486marPOST ... ... ... ... ... ... ... ... . [205]
4487marPOST ... .R. .S. ... ... ... ... ... . [205]
4488marCPRE ... ... ... ... ... ... ... ... . [205]
4489marPOST ... ... .R. ... ... ... ... ... . [205]
4493marPOST ... ... ... ... ... ... ... ... . [205]
4503marPOST ... .R. ... ... ... ... ... ... . [205]
4504marPOST ... ... ... ... ... ... ... ... . [205]
4505marPOST ... .R. .S. ... ... ... ... ... . [205]
4506marcPRE ... ... ... ... ... ... ... ... . [205]
4507marCPRE ... ... ... ... ... ... ... ... . [205]
4542campPOS ... ... ... ... ... ... ... ... . [205]
4544campPOS ... ... ... ... ... ... ... ... . [205]
4547campPOS ... ... ... ... ... ... ... ... . [205]
4548campPOS ... ... ... ... ... ... ... ... . [205]
4553campPOS ... .R. .S. ... ... ... ... ... . [205]
4554campPOS ... ... ... ... ... ... ... ... . [205]
4555campPOS ... ... ... ... ... ... ... ... . [205]
4556campPOS ... .A. .S. ... ... ... ... ... . [205]
4557campPOS ... .A. ... ... ... ... ... ... . [205]
4562campPRE ... ... ... ... ... ... ... ... . [205]
4564campPOS ... ... ... ... ... ... ... ... . [205]
4564age3camPO ... ... ... ... ... ... ... ... . [205]
4571campPOS ... ... ... ... ... ... ... ... . [205]
4572mcamPRE ... .S. ... ... ... ... ... ... . [205]
4572fcamPOS ... .R. ... ... ... ... ... ... . [205]
4573campPRE ... ... ... ... ... ... ... ... . [205]
4575campPRE ... .R. .S. ... ... ... ... ... . [205]
PN62POSTOUT ... .R. .S. ... ... ... ... ... . [205]
PN81POSTOUT ... .A. ... ... ... ... ... ... . [205]
PN123OUTBRE ... ... ... ... ... ... ... ... . [205]
PN323OUTBRE ... ... ... ... ... ... ... ... . [205]
PN331OUTBRE ... ... ... ... ... ... ... ... . [205]
PP1355OUTBR ... .R. .S. ... ... ... ... ... . [205]
Allegato 3b: Allineamento nucleotidico delle sequenze dei campioni spagnoli e di quelle depositate in GenBank per R. p. pyrenaica

141
Rupy-DRB1*01A GAG TAT CAT AAG AGC GAG TGT CAT TTC TTC AAC GGG ACC GAG CGG GTG CGG TTC CTG GAC [ 60]
Rupy-DRB1*02A ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Rupy-DRB1*03A ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... C.. ... ... [ 60]
Rupy-DRB1*04A ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Rupy-DRB1*05A ... ... AC. ... .AA ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Rupy-DRB1*06A ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Rupy-DRB1*07A ... ... AC. ... .AA ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Rupy-DRB1*08A ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... C.. ... ... [ 60]
Rupy-DRB1*09A ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Rupy-DRB1*10A ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Rupy-DRB1*11A ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... C.. ... ... [ 60]
Rupy-DRB1*12A ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Rupy-DRB1*13A ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
16.06.HL032 ... ... .G. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
15.06 ... ... .G. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
18.06 ... ... .G. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
RP02.08 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
RP2.96 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
RP03.04 ... ... .G. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
RP3.08 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
RP3.99 ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... C.. ... ... [ 60]
RP4.03 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
RP05.05 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
RP5.99 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
RP06.05 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
RP06.07 ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... C.. ... ... [ 60]
38.07 ... ... MR. ... ... ... ... ... ... ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
12.07 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
RP33.07 ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... C.. ... ... [ 60]
RP34.07 ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... C.. ... ... [ 60]
RP35.07 ... ... MR. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
RP36.07 ... ... MR. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
RP37.07 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
RP38.06 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
RP48.06 ... ... MR. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
RP7.01 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
RP10.02 ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... C.. ... ... [ 60]
RP10.03 ... ... MR. ... ... ... ... ... ... ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
RP12.08 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
RP13.02 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
RP13.03 ... ... MR. ... ... ... ... ... ... ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
RP13.07 ... ... MR. ... ... ... ... ... ... ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
RP14.02 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
RP15.02 ... ... MR. ... ... ... ... ... ... ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
RP4.HL032 ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
RP23.HL032 ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
RPC20.HL032 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
RP708.HL032 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
RP808.HL032 ... ... MR. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
RP09.03 ... ... MR. ... ... ... ... ... ... ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
RPC8 ... ... MR. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
RP19.06 --- --- --- --- --- ... ... ... ... ... .-. ... ... ... ... ... ... Y.. ... ... [ 60]
RP22.07 --- --- --- --- --- ... ... ... ... ... .-. ... ... ... ... ... ... ... ... ... [ 60]
35.05 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
26.05 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
1.04 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
39.06 ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... C.. ... ... [ 60]
12.05 ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... C.. ... ... [ 60]
15.02 ... ... MR. ... ... ... ... ... ... ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
30.05 ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
01.09 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
19.06 ... ... MR. ... ... ... ... ... ... ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
05.06 ... ... MR. ... ... ... ... ... ... ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
12.06 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
49.05 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
04.04 ... ... MR. ... ... ... ... ... ... ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
05.04 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
06.05 ... ... MR. ... ... ... ... ... ... ... ... ... ... ... ... ... ... Y.. ... ... [ 60]
Rupy-DRB1*01A AGA TAC TTC CAT AAT GGA GAA GAG TTC GTG CGC TTC AAC AGC GAC TGG GGC GAG TAC CGG [120]
Rupy-DRB1*02A ... ... ... ... ... ... ... ... ..G ... ... ... G.. ... ... ... ... ... ... ... [120]
Rupy-DRB1*03A ... ... ... T.. ... ... ... ... .A. .C. ... ... ... ... ... ... ... ... ... ... [120]
Rupy-DRB1*04A ... ... ... T.. ... ... ... ... .A. .C. ... ... ... ... ... ... ... ... ... ... [120]
Rupy-DRB1*05A ... ... ... ... ... ... ... ... ... ... ... ... G.. ... ... ... ... ... ... ... [120]
Rupy-DRB1*06A ... .T. ... T.. ... ... ... ... .A. C.. ... ... ... ... ... ... ... ... ... ... [120]
Rupy-DRB1*07A ... ... ... ... ... ... ... ... ... ... ... ... G.. ... ... ... ... ... ... ... [120]
Rupy-DRB1*08A ... ... ... T.. ... ... ... ... .A. .C. ... ... ... ... ... ... ... ... ... ... [120]
Rupy-DRB1*09A ... ... ... ... ... ... ... ... ... ... ... ... G.. ... ... ... ... ... .T. ... [120]
Rupy-DRB1*10A ... ... ... ... ... ... ... ... ..G ... ... ... G.. ... ... ... ... ... ... ... [120]
Rupy-DRB1*11A ... ... ... T.. ... ... ... ... .A. .C. ... ... ... ... ... ... ... ... ... ... [120]
Rupy-DRB1*12A ... ... ... ... ... ... ... ... ... ... ... ... G.. ... ... ... ... ... ... ... [120]
Rupy-DRB1*13A ... ... ... T.. ... ... ... ... .A. .C. ... ... ... ... ... ... ... ... ... ... [120]
16.06.HL032 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
15.06 ... ... ... T.. ... ... ... ... ..G ... ... ... ... ... ... ... ... ... ... ... [120]
18.06 ... ... ... T.. ... ... ... ... ..G ... ... ... ... ... ... ... ... ... ... ... [120]
RP02.08 ... ... ... ... ... ... ... ... ..G ... ... ... G.. ... ... ... ... ... ... ... [120]

142
RP2.96 ... ... ... ... ... ... ... ... ..G ... ... ... ... ... ... ... ... ... ... ... [120]
RP03.04 ... ... ... T.. ... ... ... ... ..G ... ... ... ... ... ... ... ... ... ... ... [120]
RP3.08 ... ... ... ... ... ... ... ... ..G ... ... ... G.. ... ... ... ... ... ... ... [120]
RP3.99 ... ... ... T.. ... ... ... ... .A. .C. ... ... ... ... ... ... ... ... ... ... [120]
RP4.03 ... ... ... ... ... ... ... ... ..G ... ... ... G.. ... ... ... ... ... ... ... [120]
RP05.05 ... ... ... ... ... ... ... ... ..G ... ... ... G.. ... ... ... ... ... ... ... [120]
RP5.99 ... ... ... ... ... ... ... ... ..G ... ... ... G.. ... ... ... ... ... ... ... [120]
RP06.05 ... ... ... ... ... ... ... ... ..G ... ... ... G.. ... ... ... ... ... ... ... [120]
RP06.07 ... ... ... T.. ... ... ... ... .A. .C. ... ... ... ... ... ... ... ... ... ... [120]
38.07 ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
12.07 ... ... ... ... ... ... ... ... ..G ... ... ... G.. ... ... ... ... ... ... ... [120]
RP33.07 ... ... ... T.. ... ... ... ... .A. .C. ... ... ... ... ... ... ... ... ... ... [120]
RP34.07 ... ... ... T.. ... ... ... ... .A. .C. ... ... ... ... ... ... ... ... ... ... [120]
RP35.07 ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
RP36.07 ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
RP37.07 ... ... ... ... ... ... ... ... ..G ... ... ... G.. ... ... ... ... ... ... ... [120]
RP38.06 ... ... ... ... ... ... ... ... ..G ... ... ... G.. ... ... ... ... ... ... ... [120]
RP48.06 ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
RP7.01 ... ... ... ... ... ... ... ... ..G ... ... ... G.. ... ... ... ... ... ... ... [120]
RP10.02 ... ... ... T.. ... ... ... ... .A. .C. ... ... ... ... ... ... ... ... ... ... [120]
RP10.03 ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
RP12.08 ... ... ... ... ... ... ... ... ..G ... ... ... G.. ... ... ... ... ... ... ... [120]
RP13.02 ... ... ... ... ... ... ... ... ..G ... ... ... G.. ... ... ... ... ... ... ... [120]
RP13.03 ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
RP13.07 ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
RP14.02 ... ... ... ... ... ... ... ... ..G ... ... ... G.. ... ... ... ... ... ... ... [120]
RP15.02 ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
RP4.HL032 ... ... ... T.. ... ... ... ... .A. .C. ... ... ... ... ... ... ... ... ... ... [120]
RP23.HL032 ... ... ... T.. ... ... ... ... .A. .C. ... ... ... ... ... ... ... ... ... ... [120]
RPC20.HL032 ... ... ... ... ... ... ... ... ..G ... ... ... G.. ... ... ... ... ... ... ... [120]
RP708.HL032 ... ... ... ... ... ... ... .W. ..G ... ... ... G.. ... ... ... ... ... ... ... [120]
RP808.HL032 ... ... ... Y.. ... ... ... .W. .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
RP09.03 ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
RPC8 ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
RP19.06 ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
RP22.07 ... ... ... ... ... ... ... ... ..G ... ... ... G.. ... ... ... ... ... ... ... [120]
35.05 ... ... ... ... ... ... ... ... ..G ... ... ... G.. ... ... ... ... ... ... ... [120]
26.05 ... ... ... ... ... ... ... ... ..G ... ... ... G.. ... ... ... ... ... ... ... [120]
1.04 ... ... ... ... ... ... ... ... ..G ... ... ... G.. ... ... ... ... ... ... ... [120]
39.06 ... ... ... T.. ... ... ... ... .A. .C. ... ... ... ... ... ... ... ... ... ... [120]
12.05 ... ... ... T.. ... ... ... ... .A. .C. ... ... ... ... ... ... ... ... ... ... [120]
15.02 ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
30.05 ... ... ... T.. ... ... ... ... .A. .C. ... ... ... ... ... ... ... ... ... ... [120]
01.09 ... ... ... ... ... ... ... ... ..G ... ... ... G.. ... ... ... ... ... ... ... [120]
19.06 ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
05.06 ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
12.06 ... ... ... ... ... ... ... ... ..G ... ... ... G.. ... ... ... ... ... ... ... [120]
49.05 ... ... ... ... ... ... ... ... ..G ... ... ... G.. ... ... ... ... ... ... ... [120]
04.04 ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
05.04 ... ... ... ... ... ... ... ... ..G ... ... ... G.. ... ... ... ... ... ... ... [120]
06.05 ... ... ... Y.. ... ... ... ... .WS .Y. ... ... R.. ... ... ... ... ... ... ... [120]
Rupy-DRB1*01A GCG GTG GCC GAG CTG GGG CGG CCG ACC GCC GAG CAC TGG AAC AGC CAG AAG GAG ATT CTG [180]
Rupy-DRB1*02A ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Rupy-DRB1*03A ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Rupy-DRB1*04A ... ... ... ... ... ... ... .G. GA. ... ... ... ... ... ... ... ... ... ... ... [180]
Rupy-DRB1*05A ... ... ... ... ... ... ... .G. GA. ... ... ... ... ... ... ... ... ... ... ... [180]
Rupy-DRB1*06A ... ... ... ... ... ... ... ... GA. ... ... ... ... ... ... ... ... ... ... ... [180]
Rupy-DRB1*07A ... ... ... ... ... ... ... ... GA. ... ... T.. ... ... ... ... ... ... ... ... [180]
Rupy-DRB1*08A ... ... ... ... ... ... ... ... GA. ... ... T.. ... ... ... ... ... ... ... ... [180]
Rupy-DRB1*09A ... ... ... ... ... ... ... ... G.. ... ... ..G ... ... ... ... ... ... ... ... [180]
Rupy-DRB1*10A ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Rupy-DRB1*11A ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Rupy-DRB1*12A ... ... ... ... ... ... ... ... G.. ... ... ..G ... ... ... ... ... ... ... ... [180]
Rupy-DRB1*13A ... ... ... ... ... ... ... ... GA. ... ... T.. ... ... ... ... ... ... ... ... [180]
16.06.HL032 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
15.06 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
18.06 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
RP02.08 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
RP2.96 ... ... ... ... ... ... ... .G. ... ... ... ... ... ... ... ... ... ... ... ... [180]
RP03.04 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
RP3.08 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
RP3.99 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
RP4.03 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
RP05.05 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
RP5.99 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
RP06.05 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
RP06.07 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
38.07 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
12.07 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
RP33.07 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
RP34.07 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
RP35.07 ... ... ... ... ... ... ... .S. RM. ... ... ... ... ... ... ... ... ... ... ... [180]
RP36.07 ... ... ... ... ... ... ... .S. RM. ... ... ... ... ... ... ... ... ... ... ... [180]
RP37.07 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
RP38.06 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]

143
RP48.06 ... ... ... ... ... ... ... .S. RM. ... ... ... ... ... ... ... ... ... ... ... [180]
RP7.01 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
RP10.02 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
RP10.03 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
RP12.08 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
RP13.02 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
RP13.03 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
RP13.07 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
RP14.02 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
RP15.02 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
RP4.HL032 ... ... ... ... ... ... ... .S. RM. ... ... ... ... ... ... ... ... ... ... ... [180]
RP23.HL032 ... ... ... ... ... ... ... .S. RM. ... ... ... ... ... ... ... ... ... ... ... [180]
RPC20.HL032 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
RP708.HL032 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
RP808.HL032 ... ... ... ... ... ... ... .S. RM. ... ... ... ... ... ... ... ... ... ... ... [180]
RP09.03 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
RPC8 ... ... ... ... ... ... ... .S. RM. ... ... ... ... ... ... ... ... ... ... ... [180]
RP19.06 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
RP22.07 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
35.05 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
26.05 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
1.04 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
39.06 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
12.05 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
15.02 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
30.05 ... ... ... ... ... ... ... .S. RM. ... ... ... ... ... ... ... ... ... ... ... [180]
01.09 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
19.06 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
05.06 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
12.06 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
49.05 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
04.04 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
05.04 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
06.05 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Rupy-DRB1*01A GAG CAG AGG CGG GCC GAG GTG GAC AC [206]
Rupy-DRB1*02A ... .G. ... ... ... .C. ... ... .. [206]
Rupy-DRB1*03A ... ... ... ... ... ... ... ... .. [206]
Rupy-DRB1*04A ... ... .C. ... ... .C. ... ... .. [206]
Rupy-DRB1*05A ... ... .C. ... ... ... ... ... .. [206]
Rupy-DRB1*06A ... ... .C. ... ... .C. ... ... .. [206]
Rupy-DRB1*07A ... .G. .C. ... ... ... ... ... .. [206]
Rupy-DRB1*08A ... ... ... ... ... .C. ... ... .. [206]
Rupy-DRB1*09A ... .G. ... ... ... ... ... ... .. [206]
Rupy-DRB1*10A ... .GA ... ... ... .C. ... ... .. [206]
Rupy-DRB1*11A ... ... ... ... ... .C. ... ... .. [206]
Rupy-DRB1*12A ... .G. ... ... ... .C. ... ... .. [206]
Rupy-DRB1*13A ... ... .C. ... ... .C. ... ... .. [206]
16.06.HL032 ... ... ... ... ... .C. ... ... .. [206]
15.06 ... .R. ... ... ... .C. ... ... .. [206]
18.06 ... .R. ... ... ... .C. ... ... .. [206]
RP02.08 ... .G. ... ... ... .C. ... ... .. [206]
RP2.96 ... .R. .S. ... ... .C. ... ... .. [206]
RP03.04 ... .R. ... ... ... .C. ... ... .. [206]
RP3.08 ... .G. ... ... ... .C. ... ... .. [206]
RP3.99 ... ... ... ... ... .C. ... ... .. [206]
RP4.03 ... .G. ... ... ... .C. ... ... .. [206]
RP05.05 ... .G. ... ... ... .C. ... ... .. [206]
RP5.99 ... .G. ... ... ... .C. ... ... .. [206]
RP06.05 ... .G. ... ... ... .C. ... ... .. [206]
RP06.07 ... ... ... ... ... .C. ... ... .. [206]
38.07 ... .R. ... ... ... .C. ... ... .. [206]
12.07 ... .G. ... ... ... .C. ... ... .. [206]
RP33.07 ... ... ... ... ... .C. ... ... .. [206]
RP34.07 ... ... ... ... ... .C. ... ... .. [206]
RP35.07 ... .R. .S. ... ... .C. ... ... .. [206]
RP36.07 ... .R. .S. GC. ... .C. ... ... .. [206]
RP37.07 ... .G. ... ... ... .C. ... ... .. [206]
RP38.06 ... .G. ... ... ... .C. ... ... .. [206]
RP48.06 ... .R. ... ... ... .C. ... ... .. [206]
RP7.01 ... .G. ... ... ... .C. ... ... .. [206]
RP10.02 ... ... ... ... ... .C. ... ... .. [206]
RP10.03 ... .R. ... ... ... .C. ... ... .. [206]
RP12.08 ... .G. ... ... ... .C. ... ... .. [206]
RP13.02 ... .G. ... ... ... .C. ... ... .. [206]
RP13.03 ... .G. ... ... ... .C. ... ... .. [206]
RP13.07 ... .R. ... ... ... .C. ... ... .. [206]
RP14.02 ... .G. ... ... ... .C. ... ... .. [206]
RP15.02 ... .R. ... ... ... .C. ... ... .. [206]
RP4.HL032 ... ... .S. ... ... .C. ... ... .. [206]
RP23.HL032 ... ... .S. ... ... .C. ... ... .. [206]
RPC20.HL032 ... .G. ... ... ... .C. ... ... .. [206]
RP708.HL032 ... .G. ... ... ... .C. ... ... .. [206]
RP808.HL032 ... .R. .S. ... ... .C. ... ... .. [206]
RP09.03 ... .R. ... ... ... .C. ... ... .. [206]
RPC8 ... .R. .S. ... ... .C. ... ... .. [206]

144
RP19.06 ... .R. ... ... ... .C. ... ... .. [206]
RP22.07 ... .G. ... ... ... .C. ... ... .. [206]
35.05 ... .G. ... ... ... .C. ... ... .. [206]
26.05 ... .G. ... ... ... .C. ... ... .. [206]
1.04 ... .G. ... ... ... .C. ... ... .. [206]
39.06 ... ... ... ... ... .C. ... ... .. [206]
12.05 ... ... ... ... ... .C. ... ... .. [206]
15.02 ... .R. ... ... ... .C. ... ... .. [206]
30.05 ... ... .S. ... ... .C. ... ... .. [206]
01.09 ... .G. ... ... ... .C. ... ... .. [206]
19.06 ... .R. ... ... ... .C. ... ... .. [206]
05.06 ... .R. ... ... ... .C. ... ... .. [206]
12.06 ... .G. ... ... ... .C. ... ... .. [206]
49.05 ... .G. ... ... ... .C. ... ... .. [206]
04.04 ... .R. ... ... ... .C. ... ... .. [206]
05.04 ... .G. ... ... ... .C. ... ... .. [206]
06.05 ... .R. ... ... ... .C. ... ... .. [206]

145
Allegato 4: Allineamento nucleotidico dei 58 aplotipi ricostruiti per R. r. rupicapra e R. p. pyrenaica. Il risultato ottenuto dalla ricostruzione degli aplotipi (diplotipi) mediante HAP (A e B indicano i 2 aplotipi ottenuti per ogni individuo analizzato) e dall’analisi degli aplotipi mediante DnaSP5. La ricostruzione aplotipica è stata condotta mediante confronto con gli aplotipi depositati in GenBank in riferimento alle due specie R. r. rupicapra e R. p. pyrenaica (Rispettivamente RuruDRB*01-RuruDRB*40 e RupyDRB*01-RupyDRB*13). In base agli aplotipi doppi ottenuti per ogni individuo è stato creato il file in DnaSP che ha permesso l’analisi degli aplotipi stessi (assegnando gli individui agli aplotipi ottenuti) e successivamente è stato creato il file input per l’analisi di popolazione con Arlequin.
(Hap_n= nome dell’aplotipo assegnato da DnaSP; il numero dopo i due punti indica quanti individui hanno quell’aplotipo; nelle parentesi ci sono gli individui in riferimento all’aplotipo) Quando è indicato un aplotipo depositato in parentesi dopo l’aplotipo ricostruito significa che c’è identità fra i due aplotipi.
Hap_1 GAG TAT CAT AAG AGC GAG TGT CAT TTC TTC AAC GGG ACC GAG CGG GTG CGG TTC CTG GAC [ 60]
Hap_2 ... ... AG. ... .AA ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_3 ... ... .C. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_4 ... ... AG. ... .AA ... ... ... ... ... ... ... ... ... ... ... ... C.. ... ... [ 60]
Hap_5 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_6 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_7 ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_8 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_9 ... ... A.. ... .AA ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_10 ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_11 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_12 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_13 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_14 ... ... .G. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_15 ... ... .G. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_16 ... ... .G. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_17 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_18 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_19 ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... C.. ... ... [ 60]
Hap_20 ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_21 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_22 ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_23 ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_24 ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... C.. ... ... [ 60]
Hap_25 ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_26 ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... C.. ... ... [ 60]
Hap_27 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_28 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... C.. ... ... [ 60]
Hap_29 ... ... .G. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_30 ... ... AG. ... .AA ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_31 ... ... ... ... ..A ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_32 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_33 ... ... .G. ... ... ... ... ... ... ... ... ... ... ... ... ... ... C.. ... ... [ 60]
Hap_34 ... ... AC. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_35 ... ... .G. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_36 ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... C.. ... ... [ 60]
Hap_37 ... ... AC. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_38 ... ... AG. ... .AA ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_39 ... ... .G. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_40 ... ... AC. ... .AA ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_41 ... ... .G. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_42 ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... C.. ... ... [ 60]
Hap_43 ... ... .G. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_44 ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... C.. ... ... [ 60]
Hap_45 ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_46 ... ... ... ... ..A ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_47 ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_48 ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_49 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_50 ... ... .G. ... ... ... ... ... ... ... ... ... ... ... ... ... ... C.. ... ... [ 60]
Hap_51 ... ... .C. ... ..A ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_52 ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_53 ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_54 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]

146
Hap_55 ... ... .C. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_56 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_57 ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... C.. ... ... [ 60]
Hap_58 .C. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [ 60]
Hap_1 AGA TAC TTC CAT AAT GGA GAA GAG TTG GTG CGC TTC GAC AGC GAC TGG GGC GAG TAC CGG [120]
Hap_2 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Hap_3 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Hap_4 ... ... ... T.. ... ... ... ... .AC .C. ... ... A.. ... ... ... ... ... ... ... [120]
Hap_5 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Hap_6 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... .T. ... [120]
Hap_7 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... .T. ... [120]
Hap_8 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Hap_9 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Hap_10 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Hap_11 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... .T. ... [120]
Hap_12 ... ... ... ... ... ... ... ... ... ... ... ... A.. ... ... ... ... ... ... ... [120]
Hap_13 ... ... ... ... ... ... ... ... ..C ... ... ... A.. ... ... ... ... ... ... ... [120]
Hap_14 ... ... ... ... ... ... ... ... ..C ... ... ... A.. ... ... ... ... ... ... ... [120]
Hap_15 ... ... ... T.. ... ... ... ... ... ... ... ... A.. ... ... ... ... ... ... ... [120]
Hap_16 ... ... ... T.. ... ... ... ... ... ... ... ... A.. ... ... ... ... ... ... ... [120]
Hap_17 ... ... ... ... ... ... ... ... ... ... ... ... A.. ... ... ... ... ... ... ... [120]
Hap_18 ... ... ... ... ... ... ... ... ... ... ... ... A.. ... ... ... ... ... ... ... [120]
Hap_19 ... ... ... T.. ... ... ... ... .AC .C. ... ... A.. ... ... ... ... ... ... ... [120]
Hap_20 ... ... ... T.. ... ... ... ... .AC .C. ... ... A.. ... ... ... ... ... ... ... [120]
Hap_21 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Hap_22 ... ... ... T.. ... ... ... ... .AC .C. ... ... A.. ... ... ... ... ... ... ... [120]
Hap_23 ... ... ... T.. ... ... ... ... .AC .C. ... ... A.. ... ... ... ... ... ... ... [120]
Hap_24 ... ... ... T.. ... ... ... ... .AC .C. ... ... A.. ... ... ... ... ... ... ... [120]
Hap_25 ... ... ... T.. ... ... ... ... .AC .C. ... ... A.. ... ... ... ... ... ... ... [120]
Hap_26 ... ... ... T.. ... ... ... ... .AC .C. ... ... A.. ... ... ... ... ... ... ... [120]
Hap_27 ... ... ... ... ... ... ... .T. ... ... ... ... ... ... ... ... ... ... ... ... [120]
Hap_28 ... ... ... T.. ... ... ... ... .AC .C. ... ... A.. ... ... ... ... ... ... ... [120]
Hap_29 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Hap_30 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... .T. ... [120]
Hap_31 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Hap_32 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Hap_33 ... ... ... T.. ... ... ... ... .AC .C. ... ... A.. ... ... ... ... ... ... ... [120]
Hap_34 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Hap_35 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Hap_36 ... ... ... T.. ... ... ... ... .AC .C. ... ... A.. ... ... ... ... ... .T. ... [120]
Hap_37 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Hap_38 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... .T. ... [120]
Hap_39 ... ... ... T.. ... ... ... ... .AC .C. ... ... A.. ... ... ... ... ... ... ... [120]
Hap_40 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Hap_41 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Hap_42 ... ... ... T.. ... ... ... ... .AC .C. ... ... A.. ... ... ... ... ... ... ... [120]
Hap_43 ... ... ... T.. ... ... ... ... .AC .C. ... ... A.. ... ... ... ... ... ... ... [120]
Hap_44 ... ... ... T.. ... ... ... ... .AC .C. ... ... A.. ... ... ... ... ... ... ... [120]
Hap_45 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Hap_46 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Hap_47 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... .T. ... [120]
Hap_48 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... .T. ... [120]
Hap_49 ... ... ... T.. ... ... ... ... .AC .C. ... ... A.. ... ... ... ... ... ... ... [120]
Hap_50 ... ... ... T.. ... ... ... ... .AC .C. ... ... A.. ... ... ... ... ... ... ... [120]
Hap_51 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Hap_52 ... ... ... T.. ... ... ... ... .AC .A. ... ... A.. ... ... ... ... ... ... ... [120]
Hap_53 ... ... ... T.. ... ... ... ... .AC .C. ... ... A.. ... ... ... ... ... ... ... [120]
Hap_54 ... ... ... T.. ... ... ... ... .AC .C. ... ... A.. ... ... ... ... ... ... ... [120]
Hap_55 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Hap_56 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Hap_57 ... ... ... T.. ... ... ... ... .AC .C. ... ... A.. ... ... ... ... ... ... ... [120]
Hap_58 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [120]
Hap_1 GCG GTG GCC GAG CTG GGG CGG CCG ACC GCC GAG CAC TGG AAC AGC CAG AAG GAG ATT CTG [180]
Hap_2 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_3 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_4 ... ... ... ... ... ... ... ... GA. ... ... T.. ... ... ... ... ... ... ... ... [180]
Hap_5 ... ... ... ... ... ... ... ... G.. ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_6 ... ... ... ... ... ... ... ... G.. ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_7 ... ... ... ... ... ... ... ... GA. ... ... T.. ... ... ... ... ... ... ... ... [180]
Hap_8 ... ... ... ... ... ... ... .G. GA. ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_9 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_10 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_11 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_12 ... ... ... ... ... ... ... ... G.. ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_13 ... ... ... ... ... ... ... ... G.. ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_14 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_15 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_16 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_17 ... ... ... ... ... ... ... .G. ... ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_18 ... ... ... ... ... ... ... .G. ... ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_19 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_20 ... ... ... ... ... ... ... .G. GA. ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_21 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_22 ... ... ... ... ... ... ... .G. GA. ... ... ... ... ... ... ... ... ... ... ... [180]

147
Hap_23 ... ... ... ... ... ... ... .G. GA. ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_24 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_25 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_26 ... ... ... ... ... ... ... .G. GA. ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_27 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_28 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_29 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_30 ... ... ... ... ... ... ... ... GA. ... ... T.. ... ... ... ... ... ... ... ... [180]
Hap_31 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_32 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_33 ... ... ... ... ... ... ... ... GA. ... ... T.. ... ... ... ... ... ... ... ... [180]
Hap_34 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_35 ... ... ... ... ... ... ... ... GA. ... ... T.. ... ... ... ... ... ... ... ... [180]
Hap_36 ... ... ... ... ... ... ... ... GA. ... ... T.. ... ... ... ... ... ... ... ... [180]
Hap_37 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_38 ... ... ... ... ... ... ... ... GA. ... ... T.. ... ... ... ... ... ... ... ... [180]
Hap_39 ... ... ... ... ... ... ... ... GA. ... ... T.. ... ... ... ... ... ... ... ... [180]
Hap_40 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_41 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_42 ... ... ... ... ... ... ... ... GA. ... ... T.. ... ... ... ... ... ... ... ... [180]
Hap_43 ... ... ... ... ... ... ... ... GA. ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_44 ... ... ... ... ... ... ... .G. GA. ... ... T.. ... ... ... ... ... ... ... ... [180]
Hap_45 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_46 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_47 ... ... ... ... ... ... ... ... GA. ... ... T.. ... ... ... ... ... ... ... ... [180]
Hap_48 ... ... ... ... ... ... ... ... GA. ... ... T.. ... ... ... ... ... ... ... ... [180]
Hap_49 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_50 ... ... ... ... ... ... ... ... GA. ... ... T.. ... ... ... ... ... ... ... ... [180]
Hap_51 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_52 ... ... ... ... ... ... ... .G. GA. ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_53 ... ... ... ... ... ... ... ... GA. ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_54 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_55 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_56 ... ... ... AG. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_57 ... ... ... ... ... ... ... ... GA. ... ... T.. ... ... ... ... ... ... ... ... [180]
Hap_58 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... [180]
Hap_1 GAG CGG AGG CGG GCC GCG GTG GAC AC [206]
Hap_2 ... .A. .C. ... ... ... ... ... .. [206]
Hap_3 ... .A. ... ... ... ... ... ... .. [206]
Hap_4 ... .A. .C. ... ... ... ... ... .. [206]
Hap_5 ... ... ... ... ... ... ... ... .. [206]
Hap_6 ... ... ... ... ... ... ... ... .. [206]
Hap_7 ... ... .A. ... ... ... ... ... .. [206]
Hap_8 ... ... ... ... ... ... ... ... .. [206]
Hap_9 ... .A. .C. ... ... ... ... ... .. [206]
Hap_10 ... .A. .C. ... ... ... ... ... .. [206]
Hap_11 ... ... ... ... ... ... ... ... .. [206]
Hap_12 ... .A. ... ... ... .A. ... ... .. [206]
Hap_13 ... .A. ... ... ... .A. ... ... .. [206]
Hap_14 ... .A. ... ... ... ... ... ... .. [206]
Hap_15 ... ... ... ... ... ... ... ... .. [206]
Hap_16 ... .A. ... ... ... ... ... ... .. [206]
Hap_17 ... ... ... ... ... ... ... ... .. [206]
Hap_18 ... .A. .C. ... ... ... ... ... .. [206]
Hap_19 ... .A. ... ... ... ... ... ... .. [206]
Hap_20 ... .A. .C. ... ... ... ... ... .. [206]
Hap_21 ... ... ... GC. ... ... ... ... .. [206]
Hap_22 ... .A. .C. GC. ... ... ... ... .. [206]
Hap_23 ... .A. ... ... ... ... ... ... .. [206]
Hap_24 ... ... ... ... ... ... ... ... .. [206]
Hap_25 ... .A. ... ... ... ... ... ... .. [206]
Hap_26 ... .A. .C. ... ... ... ... ... .. [206]
Hap_27 ... ... ... ... ... ... ... ... .. [206]
Hap_28 ... .A. ... ... ... ... ... ... .. [206]
Hap_29 ... ... .T. ... ... ... ... ... .. [206]
Hap_30 ... .A. .C. ... ... ... ... ... .. [206]
Hap_31 ... ... ... ... ... ... ... ... .. [206]
Hap_32 ... .A. ... ... ... ... ... ... .. [206]
Hap_33 ... .A. ... ... ... ... ... ... .. [206]
Hap_34 ... .A. .C. ... ... ... ... ... .. [206]
Hap_35 ... ... ... ... ... ... ... ... .. [206]
Hap_36 ... .A. .A. ... ... ... ... ... .. [206]
Hap_37 ... ... ... ... ... ... ... ... .. [206]
Hap_38 ... .A. .A. ... ... ... ... ... .. [206]
Hap_39 ... .A. .C. ... ... ... ... ... .. [206]
Hap_40 ... .A. .C. ... ... ... ... ... .. [206]
Hap_41 ... .A. ... ... ... ... ... ... .. [206]
Hap_42 ... .A. .C. ... ... ... ... ... .. [206]
Hap_43 ... .A. ... ... ... ... ... ... .. [206]
Hap_44 ... .A. .C. ... ... ... ... ... .. [206]
Hap_45 ... ... ... ... ... ... ... ... .. [206]
Hap_46 ... .A. .C. ... ... ... ... ... .. [206]
Hap_47 ... ... ... ... ... ... ... ... .. [206]
Hap_48 ... ... .C. ... ... ... ... ... .. [206]
Hap_49 ... .A. ... ... ... ... ... ... .. [206]

148
Hap_50 ... .A. .C. ... ... ... ... ... .. [206]
Hap_51 ... .A. ... ... ... ... ... ... .. [206]
Hap_52 ... .A. .C. ... ... ... ... ... .. [206]
Hap_53 ... .A. ... ... ... ... ... ... .. [206]
Hap_54 ... .A. .C. ... ... ... ... ... .. [206]
Hap_55 ... ... ... ... ... ... ... ... .. [206]
Hap_56 ... ... ... ... ... ... ... ... .. [206]
Hap_57 ... .A. ... ... ... ... ... ... .. [206]
Hap_58 ... ... ... ... ... ... ... ... .. [206]

149
Allegato 5: Lista delle corrispondenze fra i 58 aplotipi emersi dall’analisi della regione esonica del locus DRB nel presente studio e la nomenclatura in uso per la definizione degli aplotipi della medesima regione. Gli aplotipi che vanno da Ruru-DRB*01 a Ruru-DRB*40 e da Rupy-DRB*01 a Rupy-DRB*13 sono depositati in GenBank, gli aplotipi da Rupy-DRB*14 a Rupy-DRB*26 sono stati identificati nello studio di Lopez-Olvera J.R. et al (2009). (Riferimento del poster: Characterization of MHC class II DRB1 allelic diversity in Pyrenean chamois (Rupicapra pyrenaica pyrenaica) Jorge Ramón López-Olvera, Stefano D’Amelio, Ignasi Marco, Serena Cavallero, Laura Fernández, Santiago Lavín). Tali aplotipi sono stati utilizzati come confronto per la ricostruzione degli aplotipi degli individui appartenenti alle popolazioni di camosci italiani e come confronto per le successive analisi di popolazione. Hap_1: Ruru-DRB*01, Rupy-DRB*02 Hap_2: Ruru-DRB*41 Hap_3: Ruru-DRB*42 Hap_4: Ruru-DRB*43 Hap_5: Ruru-DRB*44 Hap_6: Ruru-DRB*45 Hap_7: Ruru-DRB*04 Hap_8: Ruru-DRB*27 Hap_9: Ruru-DRB*33 Hap_10: Ruru-DRB*18 Hap_11: Ruru-DRB*46 Hap_12: Ruru-DRB*47 Hap_13: Ruru-DRB*17 Hap_14: Rupy-DRB*14 Hap_15: Rupy-DRB*15 Hap_16: Rupy-DRB*16 Hap_17: Rupy-DRB*17 Hap_18: Rupy-DRB*18 Hap_19: Rupy-DRB*11 Hap_20: Rupy-DRB*04, Ruru-DRB*13, Ruru-DRB*38 Hap_21: Rupy-DRB*19 Hap_22: Rupy-DRB*20 Hap_23: Rupy-DRB*21 Hap_24: Rupy-DRB*22 Hap_25: Rupy-DRB*23 Hap_26: Rupy-DRB*24 Hap_27: Rupy-DRB*25 Hap_28: Rupy-DRB*26 Hap_29: Ruru-DRB*48 Hap_30: Ruru-DRB*49

150
Hap_31: Ruru-DRB*50 Hap_32: Ruru-DRB*51 Hap_33: Ruru-DRB*09 Hap_34: Ruru-DRB*52 Hap_35: Ruru-DRB*53 Hap_36: Ruru-DRB*54 Hap_37: Ruru-DRB*55 Hap_38: Ruru-DRB*56 Hap_39: Ruru-DRB*28 Hap_40: Ruru-DRB*57 Hap_41: Ruru-DRB*58 Hap_42: Ruru-DRB*59 Hap_43: Ruru-DRB*60 Hap_44: Ruru-DRB*61 Hap_45: Ruru-DRB*39 Hap_46: Ruru-DRB*62 Hap_47: Ruru-DRB*63 Hap_48: Ruru-DRB*64 Hap_49: Ruru-DRB*65 Hap_50: Ruru-DRB*66 Hap_51: Ruru-DRB*67 Hap_52: Ruru-DRB*68 Hap_53: Ruru-DRB*69 Hap_54: Ruru-DRB*21 Hap_55: Ruru-DRB*70 Hap_56: Ruru-DRB*71 Hap_57: Ruru-DRB*35, Rupy-DRB*08 Hap_58: Ruru-DRB*72