uva photolysis using the protein-bound sensitizers present in human lens
TRANSCRIPT

Photochemistry and Photobiology, Vol. 60, No. 1, pp. 53-60, 1994 Printed in the United States. All rights reserved
0031-8655/93 $05.00+0.00 0 1994 American Society for Photobiology
UVA PHOTOLYSIS USING THE PROTEIN-BOUND SENSITIZERS PRESENT IN HUMAN LENS
B. J. ORTWERTH* and P. R. OLESEN Mason Institute of Ophthalmology, University of Missouri, Columbia, MO 652 12, USA
(Received 24 November 1993; accepted 1 March 1994)
Abstract -This research was undertaken to demonstrate that the protein-bound chromophores in aged human lens can act as sensitizers for protein damage by UVA light. The water-insoluble (WI) proteins from pooled human and bovine lenses were solubilized by sonication in water and illuminated with UV light similar in output to that transmitted by the cornea. Analysis of the irradiated proteins showed a linear decrease in sulfiydryl groups with a 30% loss after 2 h. No loss was seen when native a-crystallin was irradiated under the same conditions. A 25% loss of histidine residues was also observed with the human lens WI fraction, and sodium dodecyl sulfate polyacrylamide gels indicated considerable protein cross-linking. Similar photodamage was seen with a WI fraction from old bovine lenses. While the data show the presence of UVA sensitizers, some histidine destruction and protein cross-linking were also obtained with a-crystallin and with lysozyme, which argue that part of the histidine loss in the human WISS was likely due to tryptophan acting as a sensitizer.
A preparation of human WI proteins was irradiated with a total of 200 J/cm2 of absorbed light at 10 nm intervals from 290 to 400 nm. Photodamage of cysteine SH groups (35%) and methionine (28Y0) was maximum at 330 nm and diminished linearly at longer wavelengths. The major loss oftryptophan (80%) occurred at 290 nm, but destruction was observed throughout the UVA range. Tyrosine was 35% destroyed at 290 nm but decreased sharply to only 50/0 at 330 nm. A constant loss of histidine (20%) was seen at all wavelengths from 290 to 360 nm, with some loss (7- 8%) even at 400 nm. These action spectra show that the human lens WI fraction contains a collection of protein- bound UVA sensitizers that can cause protein photodamage similar to that seen in cataractous lenses.
INTRODUCTION
Many epidemiological studies have implicated sunlight in the formation of age-onset cataracts in the human popula- tion,I4 and this is thought to be due to the UV portion of the spectrum. While a correlation with cataract has been made with UVB light (280-320 nm),5,6 much of this light is known to be filtered out by the Ultraviolet-A light (320-380 nm), however, is transmitted by the cornea and is almost completely absorbed by the lens. An action spectrum for cataract formation in rabbits9 showed that light from 295 to 3 15 nm was most effective. Also, studies camed out with UVB light, employing the tryptophan in lens protein as a sensitizer, have demonstrated protein damage in vitro.l&l3 The relevance of this work is diminished, however, because the little UVB light that reaches the lens would be absorbed in the first millimeter or so of lens tissue. Consistent with this is the fact that direct irradiation with UVB light in vitro causes the formation of anterior subcapsular cataract^.^^,'^ The role of tryptophan as a UVB sensitizer in vivo is also questionable because amino acid analysis of lens proteins show no loss of tryptophan in either aging l e n s e ~ ' ~ . ' ~ or cat- aracts. l8.l9
Significant lens protein damage can be demonstrated with UVA light provided an exogenous sensitizer is Sen- sitizers that produce singlet oxygen cause a photodestruction of amino acids, which resembles that seen in ~ a t a r a c t . ~ ~ , ~ ~ Twenty years ago Pirie14 and vanHeyningenZ5 demonstrated
*To whom correspondence should be addressed. tAbbreviutions: PAGE, polyacrylamide gel electrophoresis; SDS, so-
dium dodecyl sulfate; WI, water-insoluble fraction; WISS, the wa- ter-insoluble fraction solubilized by sonication.
that lens proteins exposed to sunlight for several weeks dis- played a loss of tryptophan, histidine and methionine in the absence of added sensitizer. This was presumed to be due to UVA light because of the absorption of UVB radiation by the test tube glass.
Chromophores, which absorb light above 320 nm, are present in older lenses, and increase markedly in brunescent lenses.26 Localization studies have shown these to be present mainly in the water-insoluble (WI)? fraction in the central nucleus of the lens27-29 and that they are almost exclusively bound to protein in the WI fraction upon homogen i~a t ion .~~ Zigler and G o o ~ e y ~ l . ~ ~ irradiated a mixture of lens-soluble protein with a WI fraction from brunescent cataracts and demonstrated UVA-induced protein cross-linking. Protein cross-linking, which is a feature that always accompanies senile cataract formation, has been proposed to result from singlet oxygenJ3 generated in the lens by UV light.
The work presented here not only documents that protein damage can be produced by UVA light using sensitizers bound to the WI proteins from human lens, but determines for the first time the photolysis spectrum for each amino acid de- stroyed. The spectrum of amino acids destroyed is consistent with a singlet oxygen mediator.
MATERIALS AND METHODS
Preparation of the lens WI proteins. Normal human lenses were obtained from the Lions Eye Tissue Bank of Missouri and stored frozen in Hank's salts. Twenty human lenses were decapsulated, pooled and homogenized in 10 mL of deionized water with a Dounce homogenizer. After centrifugation at 30 000 g, the pelleted WI frac- tion was resuspended in the original volume of water, stirred and repelleted. This wash procedure to remove soluble proteins was re- peated twice more, after which the WI proteins were suspended in 10 mL water and sonicated in ice for 5 min at a power rating of 4
53
54 B. J. ORTWERTH and P. R. OLESEN
and a 40% duty cycle as described previou~ly.~' The solubilized protein was recovered after centrifugation at 30 000 g, and the pellet was resuspended and sonicated again. The second supernatant, when combined with the first, represented 95% ofthe total lens WI fraction and was designated the WI sonicate supernatant (WISS). This protein solution was diluted with water to 4.0 mg/mL, sterile filtered into sterile plastic tubes and stored in the cold in aluminum foil-wrapped test tubes.
The bovine WISS was prepared as above, except that the WI fraction was prepared using lens nuclei from older lenses only which were homogenized with 5.0 mL of deionized waterllens. a-Crystallin was isolated from the water-soluble fraction by Sephadex G-200 gel filtration chromatography, followed by Agarose A-5m chromatog- raphy to remove any high molecular aggregates.
Illuniination conditions. Protein solutions (3.5 mL) containing 1 .O-2.0 mg/mL protein as determined by amino acid analysis were placed in a 4.0 mL glass stoppered cuvette (1 .O cm light path). The cuvette was placed in the beam from a 200 W super pressure mercury lamp (Oriel Corp, Stratford, CT) equipped with a 20 cm water filter to absorb infrared light and a 320 nm cutoff filter to remove UVB light. An illumination of 54 mW/cm2 for light between 320 and 380 nm was measured at the cuvette position with a YSI-Kettering Model 65 radiometer and confirmed with a uranyl oxalate actinometer. The solution was continuously stirred during a 2 h irradiation and sam- ples were removed at 30, 60, 90 and 120 min. Each aliquot was placed in a sterile Nunc tube and immediately stored in liquid ni- trogen. Equivalent samples were placed in Nunc tubes initially and incubated at room temperature in a dark chamber as control.
Illuminations at various wavelengths were camed out at 2.0 mg/ mL protein in a stirred, stoppered cuvette mounted directly in front of the YSI-Kettering detector. Light from a high pressure 1000 W mercury/xenon lamp (Oriel Corp) was directed through a Bausch and Lomb 500 mm monochrometer with the slits set at a total 10 nni bandwidth. The light absorbed was directly measured at each wavelength, and illumination was continued until a radiance of 200 J/cm2 was attained. The irradiated sample and the dark control (3.0 mL each) were removed and placed directly into liquid nitrogen.
Chemical analyses. Sulfhydryl groups were analyzed by the meth- od of Sedlack and Lind~ay.'~ Aliquots of each protein sample (0.1 mL) were added to 0.85 mL of 6.0 M urea in 0.1 MTns buffer, pH 8.0. After 15 min to denature the proteins, 50 mLofO.O1 Mdithiodi- nitrobenzoic acid in absolute methanol was added, and after 25 min the absorbance at 4 12 nm was read and compared to a glutathione standard curve also in 6.0 M urea. Sulfhydryl groups were deter- mined in duplicate for the 2 h UVA irradiations and in quadruplicate for the action spectrum.
The tryptophan content of the proteins was determined exactly as described by Spies and Chambers.j6 Values were determined in tnp- licate on 100 mg aliquots and compared to an N-acetyl-tryptophan- arnide standard curve from 0 to 40 ng per assay.
Histidine, tyrosine and methionine values were taken from a stan- dard amino acid analysis carried out as described previ~usly.~~ Ali- quots of 100 pg were analyzed in triplicate.
The values determined at each wavelength were compared to the average of the 12 dark control analyses. The loss of amino acid was expressed per 100 pg protein as determined by amino acid analysis of the original sample. Sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) was carried out as described by Laem mli .38
RESULTS
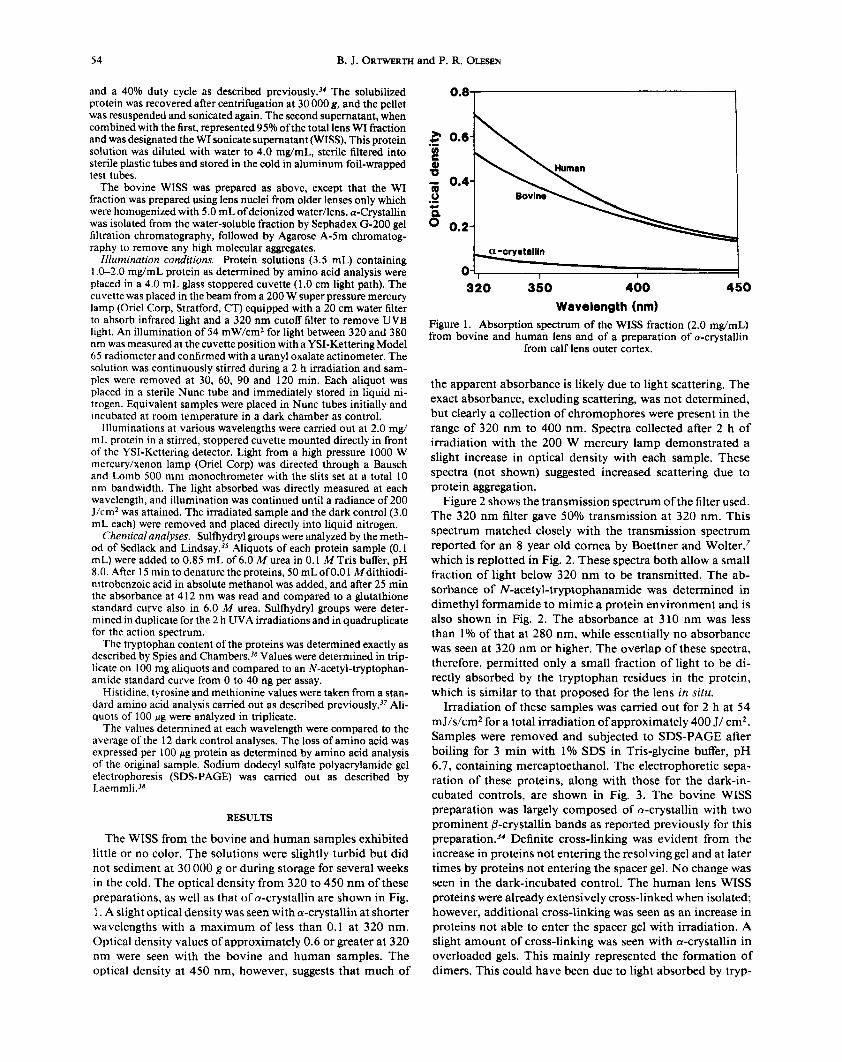
The WISS from the bovine and human samples exhibited little or no color. The solutions were slightly turbid but did not sediment at 30 000 g or during storage for several weeks in the cold. The optical density from 320 to 450 nm of these preparations, as well as that of a-crystallin are shown in Fig. 1. A slight optical density was seen with a-crystallin at shorter wavelengths with a maximum of less than 0.1 at 320 nm. Optical density values of approximately 0.6 or greater at 320 nm were seen with the bovine and human samples. The optical density at 450 nm, however, suggests that much of
I
320 350 400 450 Wavelength (nm)
Figure 1. Absorption spectrum of the WISS fraction (2.0 mg/mL) from bovine and human lens and of a preparation of a-crystallin
from calf lens outer cortex.
the apparent absorbance is likely due to light scattering. The exact absorbance, excluding scattering, was not determined, but clearly a collection of chromophores were present in the range of 320 nm to 400 nm. Spectra collected after 2 h of irradiation with the 200 W mercury lamp demonstrated a slight increase in optical density with each sample. These spectra (not shown) suggested increased scattering due to protein aggregation.
Figure 2 shows the transmission spectrum of the filter used. The 320 nm filter gave 50% transmission at 320 nm. This spectrum matched closely with the transmission spectrum reported for an 8 year old cornea by Boettner and Wolter,' which is replotted in Fig. 2. These spectra both allow a small fraction of light below 320 nm to be transmitted. The ab- sorbance of N-acetyl-tryptophanamide was determined in dimethyl formamide to mimic a protein environment and is also shown in Fig. 2. The absorbance at 310 nm was less than 1% of that at 280 nm, while essentially no absorbance was seen at 320 nm or higher. The overlap of these spectra, therefore, permitted only a small fraction of light to be di- rectly absorbed by the tryptophan residues in the protein, which is similar to that proposed for the lens in situ.
Irradiation of these samples was camed out for 2 h at 54 mJ/s/cm2 for a total irradiation of approximately 400 J/ cm2. Samples were removed and subjected to SDS-PAGE after boiling for 3 min with 1% SDS in Tris-glycine buffer, pH 6.7, containing mercaptoethanol. The electrophoretic sepa- ration of these proteins, along with those for the dark-in- cubated controls, are shown in Fig. 3. The bovine WISS preparation was largely composed of a-crystallin with two prominent P-crystallin bands as reported previously for this preparati0n.j' Definite cross-linking was evident from the increase in proteins not entering the resolving gel and at later times by proteins not entering the spacer gel. No change was seen in the dark-incubated control. The human lens WISS proteins were already extensively cross-linked when isolated; however, additional cross-linking was seen as an increase in proteins not able to enter the spacer gel with irradiation. A slight amount of cross-linking was seen with a-crystallin in overloaded gels. This mainly represented the formation of dimers. This could have been due to light absorbed by tryp-

UVA photolysis spectrum of human lens proteins 5 5
roo 1.0-
0 g 1.2-
?! 4 0.8-
0.4-
290 300 310 320 330
Wavelength (nm)
Figure 2. A comparison of the transmission spectrum of the 320 nm filter used in the broad-spectrum illumination and the published transmission spectrum of the human cornea taken from Boettner and Walter.' The absorption spectrum of the N-acetyl-tryptophan- amide in dimethylformamide is also shown to indicate the spectral
overlap.
0 280
tophan or a modification-derived sensitizer already present in a-crystallin. A control irradiation of chromatographically purified lysozyme, however, produced the same cross-linking as a-crystallin, but no cross-linking was seen with RNase A (data not shown). These data suggest a small portion of the damage was due to tryptophan acting as sensitizer.
A loss of sulfhydryl groups was similarly measured during the 2 h irradiation. Figure 4 shows an essentially linear loss in sulfhydryl groups with time of irradiation with an overall loss of 25-30% with each WISS preparation compared to the dark control. Similar assays with native a-crystallin gave no sulfhydryl destruction, showing that tryptophan is not a sen- sitizer for sulfhydryl destruction. The products of this oxi-
7 0
x F
9
94 67
43
30
20
14
030 60 90120 D 0 306090 1200 0 30 60QO120 D
Bovine WISS Human WlSS a-crystallin Figure 3. The effect of increasing time of illumination on the cross- linking of lens proteins. The first lane of each set represents the initial protein sample before irradiation, and the last lane, marked D, rep- resents the incubated dark control. The four experimental lanes show
the effect of irradiation for the time shown in minutes.
C a-Crymtallln
0 30 60 ab 120 0 30 So 80 120 0 30 So 00 120
Figure 4. The photodestruction of sulfhydryl groups in the three protein fractions by the broad spectrum light shown in Fig. 2. The 100% values were 70 nmol/mg for the human WISS, 125 nmol/mg for the bovine WISS and 46 nmol/mg for the a-crystallin sample. Aliquots from the irradiated samples are shown as open circles (I)
and the dark control as filled circles (D).
Tlme of irradlatlon (mid
dation are not certain. Amino acid analysis consistently showed an increase in cysteic acid compared to the dark controls, but the increase was variable and did not directly correspond to the loss of sulihydryl groups. The cysteine loss did not represent sulfhydryl groups that had become inac- cessible due to the formation of larger protein aggregates during the illumination as the proteins were denatured in 6.0 A4 urea prior to the SH determination. Also the incubation of illuminated samples with 6.0 A4 urea for 60 min prior to the SH determination showed no increase in SH values with time. A sample of 2 h irradiated protein was left for an additional 30 min after the lamp was turned off. This sample showed no further sulfhydryl loss, suggesting that oxidizing reagents do not accumulate during the irradiation period.
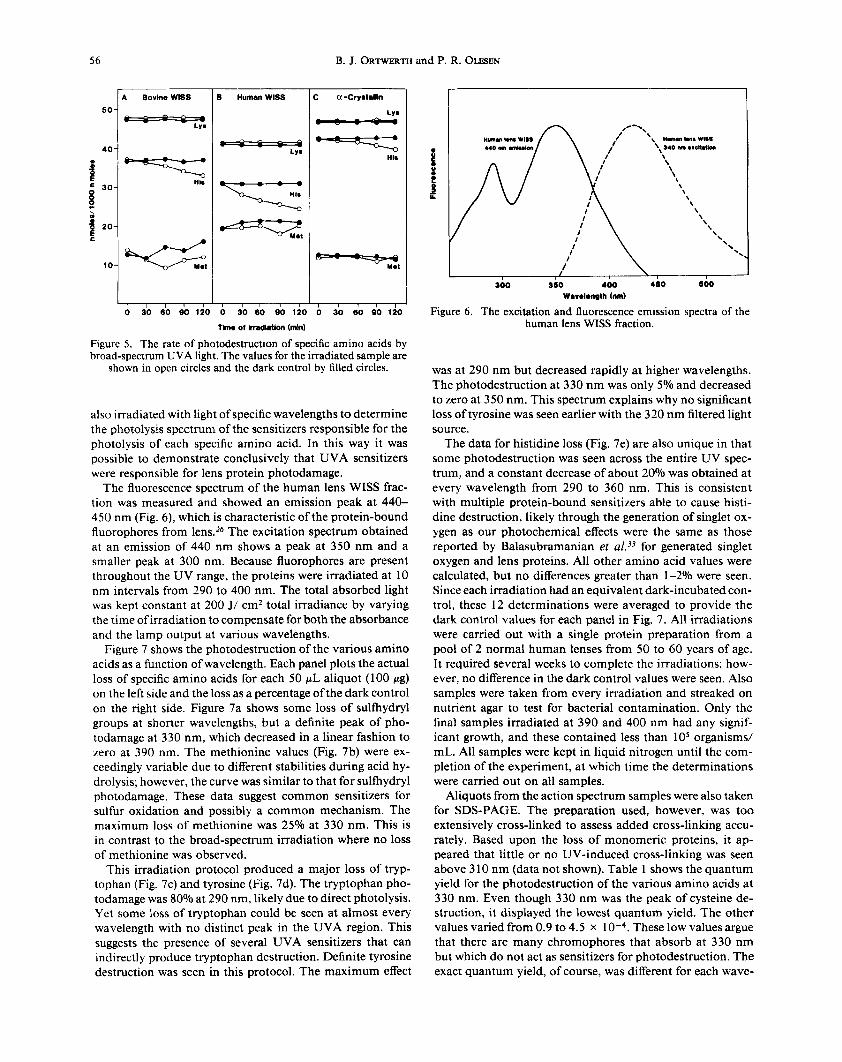
Aliquots were also subjected to amino acid analysis. No significant change in any of the amino acids was observed except cysteine and histidine. Figure 5 shows the values de- termined for histidine along with lysine as a control amino acid and methionine. No loss of lysine was seen, but histidine decreased linearly with every protein preparation. The great- est decrease was seen for the human preparation with lesser decreases for the bovine and the a-crystallin preparations.
The illumination of a 2 mg/mL solution of lysozyme (0.9 mM in tryptophan content) caused a loss of only 2 nmol of histidine/ 1000 nmol of protein, whereas the illumination of RNase A, which contains no tryptophan, had no effect on histidine content (data not shown). Therefore, while tryp- tophan appears to be a sensitizer for part of the histidine loss in our system, it was responsible for no more than 35% of the loss seen with a 2 mg/mL solution of the human WISS proteins (0.4 mM in tryptophan content).
No significant loss of tyrosine was seen, and, while ex- pected, no significant loss of methionine could be docu- mented. The loss of histidine did not apparently result in the presence of a new peak in the amino acid profile, nor was an increase in glutamic acid routinely observed. The mechanism for the photodamage is not known, but the correlation be- tween cross-linking and histidine loss supports a singlet ox- ygen m e d i a t ~ r . ~ ~ , ~ ~ , ' ~
Optimum wavelength for photolysis The previous experiments were camed out with broad-
spectrum irradiation. A preparation of human lens WISS was
56 B. J. ORTWERTH and P. R. OLESEN
I A BovheWISS
4 0 4
f -==G i 30 i
I Human WISS C a-Cryatallin
LYS - -=-z
HIS
0 30 60 eo I20 0 30 60 &I 120 b 30 do do d o Tima of hadlathon (mhl
Figure 5. The rate of photodestruction of specific amino acids by broad-spectrum UVA light. The values for the irradiated sample are
shown in open circles and the dark control by filled circles.
also irradiated with light of specific wavelengths to determine the photolysis spectrum of the sensitizers responsible for the photolysis of each specific amino acid. In this way it was possible to demonstrate conclusively that UVA sensitizers were responsible for lens protein photodamage.
The fluorescence spectrum of the human lens WISS frac- tion was measured and showed an emission peak at 440- 450 nm (Fig. 6), which is characteristic of the protein-bound fluorophores from lens.26 The excitation spectrum obtained at an emission of 440 nm shows a peak at 350 nm and a smaller peak at 300 nm. Because fluorophores are present throughout the UV range, the proteins were irradiated at 10 nm intervals from 290 to 400 nm. The total absorbed light was kept constant at 200 J/ cmz total irradiance by varying the time of irradiation to compensate for both the absorbance and the lamp output at various wavelengths.
Figure 7 shows the photodestruction of the various amino acids as a function of wavelength. Each panel plots the actual loss of specific amino acids for each 50 pL aliquot (100 pg) on the left side and the loss as a percentage of the dark control on the right side. Figure 7a shows some loss of sulfhydryl groups at shorter wavelengths, but a definite peak of pho- todamage at 330 nm, which decreased in a linear fashion to zero at 390 nm. The methionine values (Fig. 7b) were ex- ceedingly variable due to different stabilities during acid hy- drolysis; however, the curve was similar to that for sulfhydryl photodamage. These data suggest common sensitizers for sulfur oxidation and possibly a common mechanism. The maximum loss of methionine was 25% at 330 nm. This is in contrast to the broad-spectrum irradiation where no loss of methionine was observed.
This irradiation protocol produced a major loss of tryp- tophan (Fig. 7c) and tyrosine (Fig. 7d). The tryptophan pho- todamage was 80% at 290 nm, likely due to direct photolysis. Yet some loss of tryptophan could be seen at almost every wavelength with no distinct peak in the UVA region. This suggests the presence of several UVA sensitizers that can indirectly produce tryptophan destruction. Definite tyrosine destruction was seen in this protocol. The maximum effect
,
\ \ \ \
\\ \
'\\
300 360 400 460 600 Warolonpth (nml
Figure 6 . The excitation and fluorescence emission spectra of the human lens WISS fraction.
was at 290 nm but decreased rapidly at higher wavelengths. The photodestruction at 330 nm was only 5% and decreased to zero at 350 nm. This spectrum explains why no significant loss of tyrosine was seen earlier with the 320 nm filtered light source.
The data for histidine loss (Fig. 7e) are also unique in that some photodestruction was seen across the entire UV spec- trum, and a constant decrease of about 20% was obtained at every wavelength from 290 to 360 nm. This is consistent with multiple protein-bound sensitizers able to cause histi- dine destruction, likely through the generation of singlet ox- ygen as our photochemical effects were the same as those reported by Balasubramanian et ~ 2 1 . ~ ~ for generated singlet oxygen and lens proteins. All other amino acid values were calculated, but no differences greater than 1-2% were seen. Since each irradiation had an equivalent dark-incubated con- trol, these 12 determinations were averaged to provide the dark control values for each panel in Fig. 7. All irradiations were carried out with a single protein preparation from a pool of 2 normal human lenses from 50 to 60 years of age. It required several weeks to complete the irradiations; how- ever, no difference in the dark control values were seen. Also samples were taken from every irradiation and streaked on nutrient agar to test for bacterial contamination. Only the final samples irradiated at 390 and 400 nm had any signif- icant growth, and these contained less than lo5 organisms/ mL. All samples were kept in liquid nitrogen until the com- pletion of the experiment, at which time the determinations were carried out on all samples.
Aliquots from the action spectrum samples were also taken for SDS-PAGE. The preparation used, however, was too extensively cross-linked to assess added cross-linking accu- rately. Based upon the loss of monomeric proteins, it ap- peared that little or no UV-induced cross-linking was seen above 3 10 nm (data not shown). Table 1 shows the quantum yield for the photodestruction of the various amino acids at 330 nm. Even though 330 nm was the peak of cysteine de- struction, it displayed the lowest quantum yield. The other values varied from 0.9 to 4.5 x These low values argue that there are many chromophores that absorb at 330 nm but which do not act as sensitizers for photodestruction. The exact quantum yield, of course, was different for each wave-

UVA photolysis spectrum of human lens proteins
10-
8-
6-
4-
2-
0-
57
10-
8-
6-
4-
2-
0-
a)
I I I
300 340 380 Wavelength (nm)
I 1 I I
300 340 380 Wavelength (nm)
el I
'0 cp
50 2 z s P g
25 3 0
CL T
o_ 0
W 6
100 8 z 0, a
50 0
CL T
o_ 0
I I I 1 I
300 340 380 Wavelength (nm)
30 g CL
I s
t o
I b,
1 I I I
300 340 380 Wavelength (nm)
I I I 1
300 340 380 Wavelength (nm)
W
cp 50 2
2 s P
25 0
8 = 0
0
50 W
5 2 z s
25 n X 0 0 2 0
0
Figure 7. Photolysis spectra for the photodamage of specific amino acids. Aliquots of a single human lens WISS preparation were irradiated with light of different wavelengths to a total Iight absorption of 200 J/cm2. a, Sulfhydryl groups; b, methionine; c, tryptophan; d, tyrosine and e, histidine. All values are presented as nmol lost/100 pg protein and as percent loss compared to the average of 12 dark control values.
Error bars represent the standard deviation of either three or four determinations.

58 B. J. ORTWERTH and P. R. OLESEN
length. An overall photolysis at 330 nm was approximately 350 nmol/mg protein or 4-5% of the total amino acids pres- ent in the protein.
DISCUSSION
Due to the central role of the lens in focusing light on the retina, the lens proteins are constantly subjected to solar radiation. While the cornea removes most of the UVB light,7 all of the light below 400 nm is absorbed by the l e n ~ . ~ ' , ~ ~ Since there is little or no turnover of lens proteins in the central part of the lens, protein modifications accumulate and it is these protein-bound chromophores that absorb most of the UVA light.
Numerous studies have measured protein damage from UV light. Almost all of these studies have employed UVB light, measuring the photodamage generated with tryptophan as sen~i t izer~ ' , '~J~ or with UVA light using exogenously add- ed sensitizers such as riboflavin, methylene blue or rose ben- ga1.21.23J3.43 Direct UVA photodamage by sunlight, however, was carried out with lens proteins using only endogenous sensitizers by vanHeyninganZ5 and Zigler and Goos- ey.jl
We have investigated the effects of UV light at high in- tensity and short duration and have attempted to mimic the in vivo situation by using a filter that approximates the human cornea. The irradiated solution contains only the washed, WI fraction suspended as large protein aggregates in water.34 This system showed the photodestruction of cysteine, tryptophan and histidine. The cross-linking appeared to correlate with the loss of histidine as reported by Balasubramanian et ~ 1 . ~ ~ This was most clearly seen with a-crystallin where there was no cysteine loss, but a definite loss of histidine and protein dimer formation was observed. These data argue for different sensitizers causing cysteine and histidine photodamage. In every case the destruction was linear, indicating that sensi- tizers were not destroyed or formed during the irradiation. This is in contrast to the irradiation of N-acetyl-tryptophan- amide, which showed a sharply accelerated tryptophan pho- todestruction during the last hour of irradiation due to the generation of tryptophan photoproducts. This could be easily seen by the formation of yellow color. Little or no increase in yellow color, however, was observed upon irradiation of the lens protein solutions (data not shown). The 320 nm filter transmitted light that overlapped the longer wavelength tail of the tryptophan spectrum. This overlap may have been responsible for part of the photodestruction of tryptophan (data not shown) and histidine and for the protein cross- linking but not for the oxidation of sulfhydryl groups. The UVB effects on whole lens would only be seen near the an- terior surface of the lens due to the poor penetration of UVB light into the protein-rich lens.
None of the original studies on UVA photodamage in- cluded a careful determination of an amino acid photolysis at different wavelengths. One report, however, determined cataract formation at three different wavelengths using laser irradiation of calf lenses in vitro.I5 This showed highest opa- cification and protein cross-linking at 308 nm, a slight effect at 337 nm and no difference from dark control lenses at 350 nm. We have for the first time determined the optimum wavelength for the photodestruction of individual amino
Table 1. Quantum yield for photodestruction at 330 nm
Amino acid Quantum yield
( X 10-4)
Cysteine Methionine Tryptophan Tyrosine Histidine
0.7 2.4 9.0 1.8 2.0
acids in lens protein. Some damage was seen at almost every wavelength, suggesting the presence of multiple sensitizers. A comparison of the various photolysis spectra to the exci- tation spectrum for the 440 nm fluorescence shows no re- lationship. Apparently the major light energy-absorbing mol- ecules in our preparation primarily produce fluorescence when illuminated with UVA light. This is consistent with the low quantum yields, which show that only a fraction of the light absorbed results in amino acid photodestruction.
The major sensitizer for cysteine and methionine oxidation displayed a maximum at 330 nm. The mechanism is un- known, but the chemistry would suggest an involvement of singlet oxygen or hydrogen peroxide." It is also significant that both cysteine and methionine are oxidized in brunescent ~ataract.~', '~ This cataract also features the accumulation of protein-bound browning pigments that absorb light in this area of the spectrum.
The photolysis spectrum for the destruction of tryptophan extends past the absorption spectrum of tryptophan, indi- cating additional sensitizers for this amino acid. This spec- trum matched most closely the overall absorption spectrum of the isolated lens protein preparation. The spectrum for tyrosine loss, however, did resemble the absorption spectrum of tryptophan. This may represent a direct transfer of elec- trons from tryptophan to tyrosines within the protein or to some extent the direct absorption of light by tyrosine. The tyrosine action spectrum also explains why no loss of tyrosine was observed in direct irradiations with light above 320 nm. This is also consistent with cataract data, which do not report a loss of t y r ~ s i n e . ~ ~ , ' ~ Measurements for the loss of trypto- phan in cataract are conflicting. The original work of PineZ4 and vanHeyningad5 reported large losses of tryptophan in cataract lenses, but others have not seen similar losses. Re- cent measurements with a Raman probe argue that there is, if anything, an increase in tryptophan in the core of aged human lenses and cataract^.'^ Therefore, the 2O-2S0/o loss of the tryptophan we observed at higher wavelengths i n vitro are possibly not seen in cataract lenses due to protective molecules present in the lens.
Histidine photodestruction was constant from 290 to 360 nm. These data argue for multiple protein-bound UVA sen- sitizers, which are able to damage histidine via a singlet ox- ygen mediator39 and are consistent with the loss of histidine content in human cataract^.^^,^',^^
Taken as a whole, it is clear that UVA sensitizers are present in the lens, and that the major amino acid photo- destruction seen in cataract lenses is due to UVA as opposed to UVB light. This UVA effect would explain how UV light could cause a nuclear cataract, as most of the UVB light

UVA photolysis spectrum of human lens proteins 59
would be absorbed i n the first millimeter of the lens and cause an anterior subcapsular cataract as reported by Li and BorkmanI5 using a 308 n m laser in intact lenses. Ultraviolet-A light, on the other hand, would readily penetrate the normal lens fiber cells and be absorbed selectively by the protein- bound sensitizers that accumulate in the lens nucleus with age. These data also explain the correlation between sunlight exposure and the formation of brunescent cataracts.
It must also be emphasized that the illumination used for the action spectrum (200 J/cm2 of total light absorbed) rep- resents the amount of light at 330 nm that would be accu- mulated by a 1 month exposure to sunlight. The illumination (1 .O mJ/s/cm2) was only about an order of magnitude greater than sunlight (0.1 mJls/cm2) on a clear day in Cleveland a t the same ~ a v e l e n g t h . ~ ~ It must be pointed out, however, that these experiments were carried out in the absence of protec- tive molecules such as glutathione and ascorbic acid, which are present at millimolar levels even in the lens nucleus.50 Similarly, the enzymes responsible for metabolizing super- oxide anion and hydrogen peroxide were absent.
While it is difficult to assess the protective effect of these molecules, we would like t o point out that the data gathered here did not use sensitizers in solution but rather sensitizers located within large protein aggregates. The solubilized ag- gregates used here are greater than lo7 Da,j4 and aggregates greater than 1 O8 have been reported i n intact l e n ~ e s . ~ ’ J ~ Light and oxygen can readily penetrate these aggregates and, there- fore, the free radical chemistry would be carried out in the hydrophobic protein interior. This would allow the oxygen free radicals to have a greater lifetime and cause more dam- age.53 Also external antioxidants may be less effective against oxygen free radicals generated in the interior. This would suggest that UVA light is no threat until these huge protein aggregates form in abundance, which occurs after 40 years of age.54 The photodamage in our system of solubilized pro- tein aggregates from human lens, therefore, offers a rationale to explain how protein damage could accumulate in the cen- ter of the lens, how damage can occur in the presence of overwhelming protective mechanisms, and argue that UVB would be of diminished significance in senile nuclear cataract. The kinetics of photodestruction and the photolysis spectrum of the UVA sensitizers in these aggregates are presented here for the first time.
Acknowledgements-The authors express their gratitude to Dr. Ca- millo Ghiron and Dr. Robert Kuntz for their invaluable help and advice. Without their efforts this work could not have been com- pleted. The assistance of Ms. Sherry DiMaggio in preparing this manuscript is also gratefully acknowledged. This work was supported in part by NIH grant EY02035 and in part by Research to Prevent Blindness Inc.
1.
2.
3.
4.
5.
REFERENCES
Zigman, S., M. Datiles and E. Torczynski (1 979) Sunlight and human cataract. Invest. Ophthalmol. Vis. Sci. 18, 462467. Zigman, S. (1983) The role of sunlight in human cataract formation. Surv. Ophthalmol. 27, 31 7-326. Leske, M. C., L. T. Chylack Jr. and S.-Y. Wu (1992) The lens opacities case-control study. Arch. Ophthalmol. 109, 244-25 1. Young, R. W. (1992) Sunlight and age-related eye disease. J. Natl. Med. Assoc. 84, 353-358. Taylor, H. R., S. K. West, F. S . Rosenthal, B. Munoz, H. S. Newland, H. Abbey and E. A. Emmett (1988) Effect of ultra-
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
22.
23.
24.
25.
26.
27.
28.
violet radiation on cataract formation. N. Engl. J. Med. 319,
Schein, 0. D., S. West, B. Munoz, S. Vitale, M. Maguire, H. R. Taylor and N. R. Bressler ( I 994) Cortical lenticular opacifi- cation: distribution and location in a longitudinal study. invest. Ophthalmol. Vis. Sci. 35, 363-366, Boettner, E. A. and J. R. Wolter (1962) Transmission of the ocular media. Invest. Ophthalmol. 1, 776-183. Schmidt, J., S. Brettner, A. Wegener and 0. Hockwin (1990) Ultraviolet-induced changes in corneal transmission properties and influence on Scheimpflug photography. Ophthalmic Res.
Pitts, D. G., A. P. Cullen and P. D. Hacker (1977) Ocular effects of ultraviolet irradiation from 295 to 365 nrn. Invest. Ophthal. Vis. Sci. 16, 932-939. Weiter, J. J. and S. Subramanian (1 978) Free radicals produced by a biphotonic process. Invest. Ophthalmol. Vis. Sci. 17, 869- 873. Andley, U. P., P. Sutherland, J. N. Liang and B. Chakrabarti (1 984) Changes in tertiary structure of calf-lens a-crystallin by near-UV irradiation: role of hydrogen peroxide. Photochem. Photobiol. 40, 343-349. Li, D.-Y., R. F. Borkrnan, R.-H. Wang and J. Dillon (1990) Mechanisms of photochemically produced turbidity in lens pro- tein solutions. Exp. Eye Res. 51, 663-669. Hoyt, J. L. and R. F. Borkman (1992) Analysis of photooxi- dized amino acids in tryptic peptides of calf lens y-I1 crystallin. Photochem. Photobiol. 56, 257-262. MacKeen, D., S. Fine and B. S. Fine (1973) Production of cataracts in rabbits with ultraviolet laser. Ophthalmol. Res. 5,
Li, D.-Y. and R. F. Borkman (1990) Photodamage to calf lenses in vitro by excimer laser radiation at 308, 337 and 350 nm. Invest. Ophthalmol. Vis. Sci. 31, 2180-2184. Bessems, G. J. H. and H. J. Hoenders (1987) Distribution of aromatic and fluorescent compounds within single human lens- es. Exp. Eye Res. 44, 8 17-824. Smeets, M. H., G. F. J. M. Vrensen, K. Otto, G. J. Puppels and J. Greve (1993) Local variations in protein structure in the human eye lens: a Raman spectroscopic study. Biochim. Bio- phys. Acta 1164, 236-242. Dilley, K. J. and A. Pine (1974) Changes to the proteins of the human lens nucleus in cataract. Exp. Eye Res. 19, 59-72. Zigler, J. S., Jr., J. B. Sidbury, B. S. Yamanaski and M. Wol- barssht (1976) Studies on brunescent cataracts. I. Analysis of free and protein-bound amino acids. Ophthal. Res. 8,379-387. Goosey, J. D., J. S. Zigler Jr. and J. H. Kinoshita ( 1 980) Cross- linking of lens crystallins in a photodynamic system: a process mediated by singlet oxygen. Science 208, 1278-1280. Mandal, K., S. K. Bose and B. Chakrabarti (1 986) Sensitizer- induced conformational changes in lens crystallin I. Photody- namic action of methylene blue and N-formylkynurenine on bovine a-crystallin. Photochem. Photobiol. 43, 5 1 5-523. Roberts, J. E., J. S. Kinley, A. R. Young, G. Jenkins, S. J. Atherton and J. Dillon (199 I) in vivo and photophysical stud- ies on photo-oxidative damage to lens proteins and their pro- tection by radioprotectors. Photochem. Photobiol. 53, 33-38. McDermott, M., R. Chiesa, J. E. Roberts and J. Dillon (1 99 1) Photo-oxidation of specific residues in a-crystallin polypeptides. Biochemistry 30, 8 6 5 3-8 660. Pine, A. (1972) Photo-oxidation of proteins and comparison of photo-oxidized proteins with those of the cataractous lens. Isr. J. Med. Sci. 8, 1567-1573. vanHeyningen, R. (1973) Photo-oxidation of lens proteins by sunlight in the presence of fluorescent derivatives of kynurenine, isolated from human lens. Exp. Eye Rex 17, 137-147. Satoh, K., M. Bando and A. Nakajima (1973) Fluorescence in human lens. Exp. Eye Res. 16, 167-1 72. Augusteyn, R. C. (1975) Distribution of fluorescence in the human cataractous lens. Ophthalmic Res. 7, 2 17-224. Yu, N.-T., M. N. Bando and J. F. R. Kuck Jr. (1 983) Metabolic production of a blue-green fluorophore in lenses of dark-adapted mice and its increase with age. Invest. Ophthalmol. Vis. Sci. 24,
1429-1 433.
22, 365-370.
317-324.
1157-1161.

60 B. J. ORTWERTH and P. R. OLESEN
29. Yu, N.-T., M.-Z. Cai, D. J.-Y. Ho, and J. F. R. Kuck Jr. (1988) Automated laser-scanning microbeam fluorescence/Raman im- age analysis of human lens with multichannel detection: evi- dence for metabolic production of a green fluorophore. Proc. Natl. Acad. Sci. USA 85, 103-106.
30. Pine, A. (1968) Color and solubility ofthe proteins of human cataracts. Invest. Ophthalmol. 7, 634-650.
3 I . Zigler, J. S., Jr. and J. D. Goosey (1 98 1) Photosensitized ox- idation in the ocular lens: evidence for photosensitizers endog- cnous to the human lens. Photochem. Photobiol. 33, 869-874.
32. Zigler, J. S., Jr. and J. D. Goosey (1984) Singlet oxygen as a possible factor in human senile nuclear cataract development. Curr. Eye Res. 3, 59-65.
33. Balasubramanian, D., X. Du and J. S. Zigler Jr. (1990) The reactions of singlet oxygen with proteins, with special reference to crystallins. Photochem. Photobiol. 52, 76 1-768.
34. Ortwerth, B. J. and P. R. Olesen (1989) Studies on the nature of the water-insoluble fraction from aged bovine lens. Exp. Eye Res. 48, 605-619.
35. Sedlack, J. and R. H. Lindsay (1968) Estimation of total, pro- tein-bound and nonprotein sulfhydryl groups in tissue with Ell- man’s reagent. Anal. Biochem. 25, 192-205.
36. Spies, J. R. and D. C. Chambers (1948) Chemical determi- nation of tryptophan. Anal. Chem. 20, 30-39.
37. Ortwerth, B. J.. K. Krishna Sharma and P. R. Olesen (1992) The effect of urea on the aggregate state and elastase inhibitor activity of the water-insoluble fraction from bovine and human lens. Exp. Eye Res. 54, 573-581.
38. Laemmli, U. K. (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 277,680- 685.
39. Matheson, I. B. C. and J. Lee (1979) Chemical reaction rates of amino acids with singlet oxygen. Photochem. Photobiol. 29,
40. Verwey, H., T. M. A. R. Dubbleman and J. van Steveninck ( I 98 I ) Photodynamic protein crosslinking. Biochim. Biophys. Acta 647, 87-94.
41. Pokorny, J., V. C. Smith and M. Lutze (1987) Aging of the human lens. Appl. Optics 26, 1437-1440.
42. Barker, F. M., G. M. Brainard and P. Doyhaw-Barker (1991) Transmittance of the human lens as a function of age. Invest. Ophthalmol. Vis. Sci. Suppl. 32, 1083.
879-88 1.
43. Jernigan, H. M., Jr., H. N. Fukui, J . D. Goosey and J. H. Ki- noshita (1981) Photodynamic effects of rose bengal or ribo- flavin on carrier-mediated transport systems in rat lens. Exp. Eye Res. 32, 461-466.
44. Siezen, R. J., C. M. Coffin, E. D. Kaplan, D. Dwyer and J. A. Thomsom (1989) Oxidative modifications to crystallins in- duced in calf lenses in vitro by hydrogen peroxide. Exp. Eye Res. 48, 225-235.
45. Truscott, R. J. W. and R. C. Augusteyn (1977) Oxidative changes in human lens proteins during senile nuclear cataract formation. Biochirn. Biophys. Acta 492, 43-52.
46. Gamer, M. H. and A. Spector (1980) Sulfur oxidation in se- lected human cortical cataracts and nuclear cataracts. Exp. Eye Res. 31, 361-369.
47. Clark, R., S. Zigman and S. Lerman (1969) Studies on the structural proteins of lens. Exp. Eye Res. 8, 172-182.
48. Ortwerth, B. J. and P. R. Olesen (1992) Studies on the solu- bilization of the water-insoluble fraction from human lens and cataract. Exp. Eye Res. 55, 777-783.
49. Koller, L. R. (1965) Ultraviolet Radiation. John Wiley and Sons, New York.
50. Pau, H., P. Graf and H. Sies (1990) Glutathione levels in human lens: regional distribution in different forms of cataract. Exp. Eye Res. 50, 17-20.
5 1. Caldwell, K. D., B. J. Compton, J. C. Giddings and R. J. Olson (1 984) Sedimentation field-flow fractionation: a method for studying particulates in cataractous lens. Invest. Ophthalmol. Vis. Sci. 25, 153-159.
52. Merkel, P. B. and D. R. Kearns (1972) Radiationless decay of singlet molecular oxygen in solution. An experimental and the- oretical study of electronic-to-vibrational energy transfer. J. Am. Chem. SOC. 94, 7244-7253.
53. Pyskir, M., B. Grzegorzewski and A. Gutsze (1991) Compo- nents of light scattered by eye lens. Lens and Eye Toxicol. Res.
54. Suarez, G., A. L. Oronsky and M. H. L. J. Koch (1993) Age- dependent structural changes in intact human lenses detected by synchroton radiation X-ray scattering. J. Biol. Chem. 268,
8, 109-120.
17716-1772 1.