utilization of labelled mineral and organic phosphorus ... · utilization of labelled mineral and...
TRANSCRIPT

Aust. J. Soil Res., 1993, 31, 271-83
Soil Fertility and Plant Nutrition
Utilization of Labelled Mineral and Organic Phosphorus Sources by Grasses Common to Semi-arid Mulga Shrublands
R. D. ArmstrongA and K. R. HelyarB
A Queensland Department of Primary Industries, P.M.B. 6, Emerald, Qld 4720.
Wagga Agricultural Research Institute, P.M.B., Wagga Wagga, N.S.W. 2650.
Abstract
We examined whether the differing growth responses to varying concentrations of soil P by grasses common to the mulga shrublands of south-west Queensland were due to differences in their ability to utilize sparingly soluble sources of P in two glasshouse trials.
The uptake of three sparingly soluble sources of mineral P (amorphous FeP04, crystalline FeP04 (strengite), and amorphous AlP04) was compared with that of a soluble P source, KH2P04, over 34 days. All P sources were uniformly labelled with 3 2 ~ . For the four grasses studied (Cenchrus ciliaris, Aristida armata, Digitaria ammophila and Thyridolepis mitchelliana), the addition of strengite produced the lowest yields and plant P content, followed by FeP04, and then Alp04 and KH2P04. Recovery of the labelled P by the grasses was in the order: strengite < FeP04 < Alp04 < KH2P04. C. ciliaris recovered more of the added strengite, but less FeP04 and AlP04, than the other grasses. There were no differences in specific activity between the grasses for KH2P04. Where soil treatments were the same, there was no evidence that these species differed in their ability to utilize sparingly soluble sources of mineral P.
The reutilization of P from 3 2 ~ labelled plant residues by two grasses was examined in a second experiment. The experiment consisted of a factorial combination of two species (D. ammophila and A. armata ) and a control (non-planted pots), two phosphorus systems (low and high P) , and four harvests (15, 23, 31, and 39 days). The net release of P from the residues was significantly increased in the presence of plants but was not affected by the species present. D. ammophila and A. armata obtained similar proportions of their P content from the added residues (6.6%).
Keywords: mineral phosphates, organic phosphate, plant residues, mineralization, 3 2 ~ ,
Cenchrus ciliaris, Aristida armata, Digitaria ammophila, Thyridolepis mitchelliana.
Introduction
Phosphorus (P) is the principal soil nutrient limiting plant growth in the mulga shrublands of south-western Queensland (Dawson and Ahern 1973). Previous studies have demonstrated that grasses native to mulga shrublands such as Aristida armata, Digitaria ammophila and Thyridolepis mitchelliana and exotic grasses, such as buffel grass (Cenchrus ciliaris L.), vary significantly in both their relative yields without added P and in their responsiveness to P fertilizers (Silcock et al. 1976; Armstrong and Helyar 1993). The differing responses of these grasses to varying concentrations of soil P has been attributed to eco-physiological adaptations such as low relative growth rates (Christie and Moorby 1975; Armstrong and

R. D. Armstrong and K. R. Helyar
Helyar 1993) and infection with vesicular arbuscular mycorrhiza (Armstrong e t al. 1992). However, none of these studies have considered whether the differential ability of these grasses to grow in these soils is related to an ability to access sparingly soluble sources of soil P.
In the acid, strongly weathered soils that characterize mulga shrublands, only a small proportion of the total soil P would be readily soluble in the short term (Grierson e t al. 1991). The majority of soil P comprises both (1) sparingly soluble sources of mineral P and (2) organic P compounds and P derived from recycling of plant residues. Mineral P in these acid soils is likely to be associated with cations such as aluminium (Al) and iron (Fe) (Chang and Jackson 1958). The amount of these various compounds present depends on several factors including soil pH, weathering history, and parent material (Enwezor 1977).
Despite vast amounts of literature examining differences between species in ability to utilize readily soluble P, comparatively few studies have examined whether plants vary in their ability to utilize sparingly soluble sources of P. Most of these studies have examined differences between cereals/grasses and legumes rather than between species within a family. Probert (1972), for example, found that L-values of ryegrass increased with time at a greater rate than for clover. This increase in L-value was attributed to mobilization of sparingly soluble P which, in turn, was related to differences in P uptake between the species. VA-mycorrhiza infection (Bolan et. al. 1987) and alteration of rhizosphere pH by the plant have also been shown to affect the uptake of sparingly soluble sources of mineral P (Aguilar and Van Diest 1981). No study, to our knowledge, has examined whether species differ in their ability to access P derived from plant residues.
In this paper, we examined whether previous observations of varying growth responses to different soil P concentrations by four grasses common to mulga shrublands (Armstrong and Helyar 1993) were related to the ability of plants to utilize different sources of soil P. We studied the ability of the grasses to utilize P from synthetic 32P labelled compounds, comprising both amorphous and crystalline A1 and Fe-P, as well as that of a soluble P source, KH2P04. There is some uncertainty as to what form sparingly soluble P occurs in the soil (Sanyal and De Datta 1991) and there is a widespread perception that crystalline P compounds such as strengite are unlikely to occur naturally in the soil (Bache 1963; Norrish and Rosser 1983). However, we argue that the synthetic compounds used in this study represent extremes of a continuum of P solubility ranging from amorphous A1 and Fe-P of high solubility to highly insoluble crystalline forms such as strengite. Within this continuum, continuous interchange is likely between different (unidentified) phases in accordance with solubility (Lindsay 1979; Hetrick and Schwab 1992) and kinetic (Sposito 1985) principles. Consequently, the synthetic compounds used in this study were used to test the ability of plants to utilize sparingly soluble sources of (unidentified) soil P. Because of the importance of recycling of P from plant residues to the maintenance of the P cycle in natural systems (Harrison 1982), we also investigated whether two of the grasses used in the first study differed in their ability to reutilize P derived from labelled organic matter.

Utilization of P Sources by Grasses
Materials and Methods Labelling of Mineral Phosphates
Colloidal (subsequently referred to as FeP04) and crystalline iron phosphate (strengite) was prepared by the method of Cate et al. (1959)'using carbonyl iron powder dissolved in orthophosphoric acid. After colloidal ferric phosphate was precipitated, it was washed repeatedly with distilled water until the pH of the bathing solution was greater than pH 3.6 to remove free phosphoric acid. This precipitate was digested in boiling water to induce crystallization. Amorphous aluminium phosphate (AlP04) was prepared by the method of Deming and Cate (1963). Aluminium wire was dissolved in phosphoric acid and the precipitate repeatedly washed with distilled water until the filtrate was greater than pH 3.5 in order to remove any free phosphoric acid.
Both the iron and aluminium compounds were oven dried at 50°C for several days and finely ground with a mortar and pestle before analysis with X-ray diffraction to identify the compounds.
After drying over anhydrous CaC12, the compounds were uniformly labelled with 3 2 ~ by subjecting them to neutron irradiation (5 x 1013 neutrons cm-2 s-l) in the HIFAR reactor at the Australian Nuclear Science and Technology Organization Laboratories, Lucas Heights, Sydney. Temperatures during irradiation did not exceed 50°C and microscopic examination of the compounds before and after irradiation showed no apparent signs of crystal decomposition induced by irradiation (Armstrong et al. 1993).
Glasshouse %a1
This glasshouse experiment consisted of a completely randomized design with three replicates and factorial combinations of the following treatments: four P sources (amorphous FeP04, crystalline FeP04 (strengite), amorphous A1P04 and KH2P04) and four species (Cenchms ciliaris cv. U.S.A., Digitaria ammophila, Aristida armata and Thyridolepis mitchelliana). C. ciliaris is an introduced species whereas the other grasses are indigenous to the mulga shrublands of Queensland.
Topsoil (600 g) collected from a sandy red earth (Gn2.12; Northcote 1971) profile in a mulga (Acacia aneura) shrubland was placed in 100 mm diameter pots lined with plastic bags. General chemical characteristics of this soil are bicarbonate extractable P (Olsen et al. 1954), 2.9 pg g-l; pH (lo-' M CaClz), 4.1; cation exchange capacity, 1.58 cmol(+) kg-'; organic carbon, 9.1 g kgF1; and P buffering capacity (Ozanne and Shaw 1967), 45 pg P sorbed g-l/pg m ~ - l at 0.3 pg P rn~- ' . A basal nutrient application of K (114 mg KCl), S (137 mg NazS04) and N (17.25 mg NH4N03) was incorporated into the soil of each pot. Further N (17.25 mg NH4N03) was added as a solution to the surface of pots on days 17, 22 and 29. In the case of C. ciliaris, CaC03 was added at the rate of 0.64 g per pot to overcome the previously observed severe aluminium toxicity suffered by this species on this soil. The addition of CaC03 raised soil pH from 4.1 to 5.8, measured in lo-' M CaC12. CaC03 was not added to the native grasses as it can cause growth depressions in these species (Armstrong 1988). An addition of soluble P (2.5 mg P per pot as NaHzP04.2HzO) was made to all pots. This low rate of P was added so as to ensure thorough root exploitation of the soil within the time period that 3 2 ~ could be measured. This rate of addition was still deficient for growth of all species (Armstrong 1988).
The labelled phosphorus compounds were added to each pot as a stirred suspension. Potassium dihydrogen orthophosphate was mixed with commercially supplied (ANSTO) carrier-free ~ 3 ~ ~ ~ 0 4 to give the same specific activity as the irradiated compounds. The radioactive compounds were added at a rate designed to add 3.05 mg P and approximately 0.53 MBq per pot. Subsequent analysis showed the actual specific activity of the various compounds differed by f 20% (due to the nature of the irradiation). Consequently, it was necessary to correct for initial differences in specific activity between compounds when calculating 3 2 ~ uptake.
After all compounds were added to the pots and allowed to dry, the soil in each pot was thoroughly mixed, and then wetted to field capacity (0.125 g water g-l soil). Pre-germinated seedlings of the grasses were then transplanted at the 1- to 2-leaf stage to give four plants per pot. The grasses were grown in a glasshouse with a 30/27OC daylnight cycle under natural lighting. Shoot and roots of the grasses were harvested 34 days after transplanting.

R. D. Armstrong and K. R. Helyar
Chemical Analysis
Following harvest, the plants were oven dried at 60°C for 36 h, weighed, ground (<1 mm) and digested in nitric and perchloric acids. Inorganic 3 1 ~ in the digests was determined using the ascorbic acid/molybdenum blue method (Murphy and Riley 1962). The amount of 3 2 ~ in the digest was determined by measuring Cerenkov radiation in a liquid scintillation counter, and corrected for decay.
Preparation of Labelled Organic Matter
Cenchrus ciliaris cv. U.S.A. seedlings were grown in the mulga sandy red earth topsoil in 125 mm diameter pots (six seedlings per pot) lined with plastic bags. Each pot contained 1.2 kg of air dry soil and was amended with basal nutrients (KC1, 218 mg; NazS04, 278 mg; NH4N03, 69 mg). Two contrasting tissue P levels were achieved by adding NaH2P04.2HzO 'at a rate of 30 and 60 mg P per pot, designated as the low and high phosphorus systems, respectively. These comparatively high rates of applied P were used because (i) it was necessary to achieve rapid growth rates due to the short half life of 3 2 ~ and (ii) the tissue P concentrations approached levels more realistic of field conditions where there was more soil available per plant than in the pots. At 26 days post transplanting, tops of seedlings were removed at 3 cm and carrier free 3 2 ~ (370 MBq) was applied over the soil surface. Immediately after the 3 2 ~ was added, a further 15 mg of P (as NaHzP04.2H20) was added to the high P pots in order to replace P exported in the harvest. The grass was grown for a further 15 days before the shoots were harvested. After harvesting, the plant material was oven dried for 24 h at 40°C then ground (<1 mm). The plant residues contained 1.09 mg P g-l and 119 MBq g-l P in the low P system, whereas that from the high P system contained 1.71 mg P g-l and 50 MBq g-l P (corrected to the time of sowing the following trial).
Glasshouse Dial
This experiment consisted of a completely randomized factorial design of: plants present or absent (subsequently referred to as planted and control pots respectively); two species (Arzstida armata and Digitaria ammophila); two soil/residue phosphorus systems comprising a low P system (2.5 mg P per pot added to soil as NaH~P04.H20 together with the low P residues) and a high P system (7.5 mg P per pot added to soil plus high P residues); and four harvests (15, 23, 31 and 39 days). The control pots were replicated twice and other treatments replicated three times.
Soil similar to that used to prepare the labelled material was used in the pot study. Labelled plant material was thoroughly mixed with the soil at a rate of 0.6 g per 600 g of soil. Basal nutrients of K (144 mg KCl), S (139 mg Na2S04) and N (34.5 mg NH4N03) were added to each pot and, together with the labelled residues, were incorporated into the soil. Immediately after the residues were added, all pots were watered to field capacity (0.12 g water g-l soil) and 5 day old seedlings (1- to 2-leaf stage) were transplanted into the pots (four plants per pot). Pots were watered daily to weight and grown in an air conditioned glasshouse maintained at 30&3OC day and 27f 3OC night temperature.
Chemical Analysis After harvesting, plants were oven dried at 80°C and soil at 40°C for 24 h. Plant samples
were weighed, ground and digested in nitric/perchloric acid. Soil inorganic and organic P fractions were determined using the ignition method of Saunders and Williams (1958). The 3 1 ~ and 3 2 ~ concentrations in both the soil and plant material were then analysed as in the first experiment (Labelling of Mineral Phosphates). The net phosphorus release from the labelled residues (NPR) was calculated as
NPR (%) = ( 3 2 ~ in soil inorganic fraction +32 P in shoots and roots)
x 100/(total 3 2 ~ initially added in residues)
Where appropiate, data were transformed for analysis by ANOVA and re-transformed for presentation.

Utilization of P Sources by Grasses
Table 1. Effect of P source on the total plant weight (shoot and root) and total plant P uptake of C. ciliaris, A. armata, D. ammophila and T. mitchelliana
-- - -
Species P source Mean FeP04 Strengite Alp04 KH2P04
Total dry weight (g/pot)
C. ciliaris 2.36 1.94 3.16 3.07 2.63 A. annata 1.62 1.28 2.25 1.76 1.73 D. ammop hila 2-77 2.55 2.86 3.23 2.85 T. mitchelliana 1.28 1.12 1.47 1.60 1.37
Mean 2.01 1.72 2.44 2.41 1.s.d. ( P > 0.05) P source (0.289) Species (0.289) PxS ( n . ~ . ) ~
Total (shoot~root) P uptake (mg P/pot)
C. ciliam's 1.48 1.29 1.97 1.88 1.66 A. armata 1.93 1.49 2.42 2.16 2.00 D. ammophila 2.02 1.44 2.32 2.84 2.16 T. mitchelliana 1.70 1.34 1.87 1.88 1.70
Mean 1.78 1.39 2.14 2.19 1.s.d. ( P > 0.05) P source (0.220) Species (0,220) PxS (n.s.)
A n.s., ANOVA not significant (P > 0.05).
Results
Experiment 1. Utilization of Mineral P
There was no significant interaction (P > 0.05) between plant species and P source for either whole plant dry matter or plant P uptake (Table 1). The addition of strengite produced the lowest yields and P uptake in all the grasses, followed by FeP04, whereas adding Alp04 and KH2P04 produced the highest yields and P uptake. C. ciliaris and T. mitchelliana had significantly lower P uptake than A. armata and D. ammophila.
The percentage of each labelled P source recovered by the grasses depended on the species (Table 2). Lowest recoveries always occurred for strengite and, to a lesser extent, FeP04. Whereas C. ciliaris recovered more of the added strengite than the other species, it recovered less FeP04 and AlP04. D. ammophila recovered more KH2P04 than A. a m a t a which, in turn, recovered more KHzP04 than C. ciliam's and T. mitchelliana.
The proportion of P in the plant derived from the labelled P sources was calculated as
where SA, is the specific activity of P in the plant (shoot plus root) and SA, is the specific activity of P in the P source. The percentage of plant P derived from the labelled P sources ranged from a low of 0 - 4 for strengite to nearly 32% for Alp04 (Table 2). The percentage of plant P derived from the labelled P sources followed similar trends to the percentage recovery of FeP04 and strengite. However, AlP04 contributed a greater proportion of plant P than KH2P04 for the native grasses, whereas the opposite tended to occur for the percentage of P source recovered.

R. D. Armstrong and K. R. Helyar
Table 2. The percentage of the labelled P sources recovered and the proportion of P in the plant derived from the labelled P sources by C. ciliaris, A. armata, D. ammophila and
T. mitchelliana Means followed by the same letter are not significantly different ( P > 0.05). Data were transformed for analysis and retransformed for presentation. Non-transformed 1.s.d. ( P > 0.05):
(1) percentage recovery of P source, 3.00; (2) proportion of plant P, 2.92
P source FeP04 Strengite Alp04 KHzP04
C. ciliaris A, armata D. ammophila T. mitchelliana
C. ciliaris A. armata D. ammophila T. mitchelliana
Recovery of P source (%) 3 . 6 8 ~ 1.00b 5.95d 0.12a 7.19de 0.13a 5.98d 0.14a
Proportion of plant P (%) 1 2 . 0 1 ~ 3.91b 14.93d 0.42a 17.42d 0.46a 17.33d 0.54a
Table 3. The specific activity (kBq mg plant P-l) and comparative availability (in parentheses) of the labelled mineral P sources compared to KHzP04 (=loo)
Means followed by the same letter are not significantly different (P > 0.05). Specific activity data were log transformed and comparative availability data were square root transformed for analysis before retransforming for presentation. Values were corrected for differences in initial specific activity between sources and corrected for decay so that the day the P sources
were added to pots equals Time0 - -
Species P source FeP04 Strengite Alp04 KHzP04
C. ciliaris 21.ld 6 . 7 ~ 36.7f 60.3ghij (35 .4~) (1l . lb) (61 .0e) (1oog)
A. armata 26.2e 0.7a 54.7ghi 69.4j (38. OC) (1. Oa) (78.9f) (10%)
D. ammophila 30.6ef 0.7a 53.1gh 66.5ij (46. ld) (1.2a) (80. Of) (10%)
T. mitchelliana 30.5ef 0.9b 51.2g 63.4hij (48. ld) (1.5a) (80. lf) (10%)
The specific activity of the P taken up from the KH232P04 did not significantly (P > 0.05) differ between the grasses (Table 3). However, the comparative availability of the mineral P sources differed between the grasses. Strengite had 11.1% of the availability of KH2P04 (=loo) for C. ciliaris, whereas it was less than 1.5% availability for the other grasses. FeP04 was less available to C. ciliaris and A. armata (35 to 38%) than to D. ammophila and T. mitchelliana (46-48%). AlP04 had up to 80% of the availability of KH2P04 and was more available than the Fe compounds. Similar to FeP04, Alp04 was less available to C. ciliaris than the native grasses.
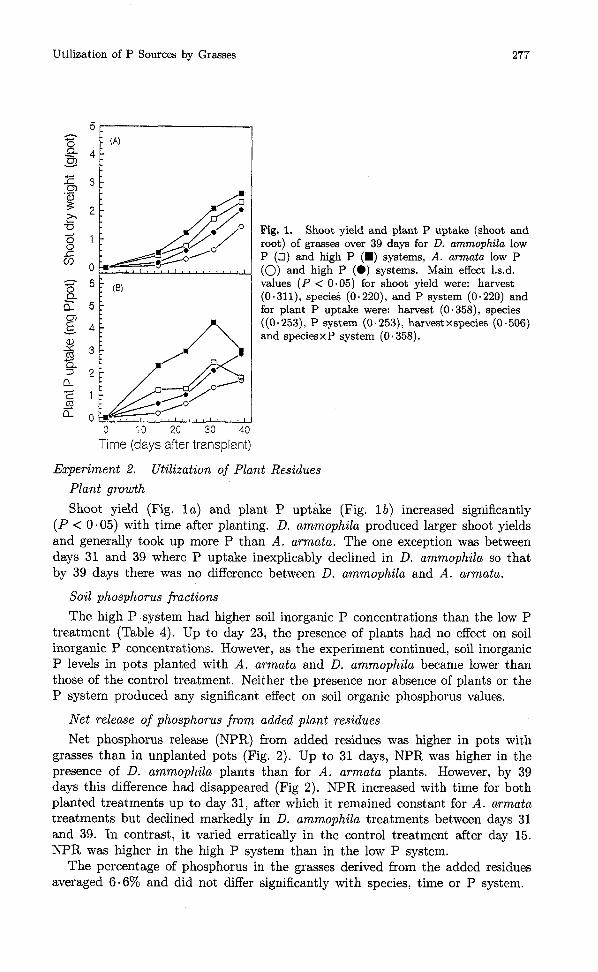
Utilization of P Sources by Grasses
5~
Fig. 1. Shoot yield and plant P uptake (shoot and root) of grasses over 39 days for D. ammophila low P (0) and high P (W) systems, A. armata low P (0) and high P (0) systems. Main effect 1.s.d. values (P < 0.05) for shoot yield were: harvest (0.311), species (0.220), and P system (0.220) and for plant P uptake were: harvest (0.358), species ((0 - 253), P system (0.253), harvest xspecies (0.506) and speciesx P system (0.358).
Time (days after transplant)
Experiment 2. Utilization of Plant Residues
Plant growth
Shoot yield (Fig. l a ) and plant P uptake (Fig. l b ) increased significantly (P < 0.05) with time after planting. D. ammophila produced larger shoot yields and generally took up more P than A. armata. The one exception was between days 31 and 39 where P uptake inexplicably declined in D. ammophila so that by 39 days there was no difference between D. ammophila and A. armata.
Soil phosphorus fractions
The high P system had higher soil inorganic P concentrations than the low P treatment (Table 4). Up to day 23, the presence of plants had no effect on soil inorganic P concentrations. However, as the experiment continued, soil inorganic P levels in pots planted with A. armata and D. ammophila became lower than those of the control treatment. Neither the presence nor absence of plants or the P system produced any significant effect on soil organic phosphorus values.
Net release of phosphorus from added plant residues
Net phosphorus release (NPR) from added residues was higher in pots with grasses than in unplanted pots (Fig. 2). Up to 31 days, NPR was higher in the presence of D. ammophila plants than for A. armata plants. However, by 39 days this difference had disappeared (Fig 2). NPR increased with time for both planted treatments up to day 31, after which it remained constant for A. armata treatments but declined markedly in D. ammophila treatments between days 31 and 39. In contrast, it varied erratically in the control treatment after day 15. NPR was higher in the high P system than in the low P system.
The percentage of phosphorus in the grasses derived from the added residues averaged 6.6% and did not differ significantly with species, time or P system.

R. D. Armstrong and K. R. Helyar
Discussion
These experiments demonstrated that under the experimental conditions used, grasses common to mulga shrublands can derive a major proportion of their plant P from sparingly soluble sources of P. However, there was little unequivocal evidence that the varying abilities of these grasses to grow in this soil without
Table 4a. Soil inorganic and organic phosphorus levels in relation to phosphorus level and the presence/absence of plants
Phosphorus Days since planting level 15 23 31 39
Inorg. Org. Inorg. Org. Inorg. Org. Inorg. Org. ( C L ~ p/g soil)
A. a m a t a
LOW 9 . 7 26.2 6.2 24.2 7.4 22.6 7 . 9 23.2 High 13.4 24.2 10.8 22.3 10.7 23.0 9 .5 23.5
D. ammophila
LOW 9.8 24.9 6 . 1 23.5 7.0 23.3 6.9 23.1 High 12.7 25.7 9.4 22.3 9 .1 24.9 10.4 24.1
No. plants
LOW 9 . 1 23.7 7 . 3 22.3 9 .5 25.3 8 . 3 24.1 High 12.8 25.0 12.6 22.8 14.4 24.5 13.8 21.6
Table 4b. Statistical analysis of phosphorus levels
ANOVA Inorganic 1.s.d. Organic 1.s.d.
P (5%) P (5%)
Harvest ** 0.2 * 1 . 3
Plants ** 0.6 n.s. -
P level ** 0.5 n.s. - Harvest x plants ** 1.2 n.s. - Harvest x P level n . ~ . ~ - n.s. - Plants x P level ** 0.9 n.s. - Harvest x Plants x P level n.s. - n.s.
A n.s., ANOVA was not significant (P > 0.05).
Fig. 2. The net release of phosphorus from added residues for D. ammophila low P (0) and high P (M) systems, A. a m a t a low P (0) and high P (e) systems, and unplanted (control) for low P (A) and high P (A) systems. Main effect 1.s.d. values (P < 0.05) were harvest (2.70), plants (2 -33), P system (1.91), and harvest xplants (4.67).
Harvest (days)

Utilization of P Sources by Grasses
added P, or to respond to added soluble P (Armstrong and Helyar 1993), could be explained in terms of differential utilization of sparingly soluble sources of mineral P or P derived from plant residues.
Mineral Phosphate
The use of labelled compounds provided unequivocal evidence that the grasses derived P from the labelled mineral P compounds added to the soil. The present results support those from another study using a sequential fractionation procedure which found that these grasses utilized soil P that was in equilibrium with chemical extractants that are claimed to recover sparingly soluble forms of P (Armstrong and Helyar 1992). These extractants (0.5 M NaHC03, 0.1 M
NaOH, and 1 .0 M HC1) are claimed to extract iron and aluminium phosphates, resistant soil organic phosphates and inorganic Ca phosphates such as apatite (Anderson 1964, Smith 1966, Holford and Crocker 1988). However, the allocation of different soil P pools to different extracts in P fractionation schemes is arbitrary and has been repeatedly criticized (eg. Russell 1973). In the current experiments, radioactive labelled P compounds were used to show that labelled P in the grasses was originally derived from the labelled sources of mineral P.
The availability of the various P sources to the grasses compared to KH2P04 (100%) was 35 to 49% for FeP04, 1-11% for strengite, and 61-80% for Alp04 (Table 4). Although it is difficult to determine the exact availability of the mineral P compounds because of the varying effect of isotopic exchange on P compounds of different solubility through time (McLaughlin and Alston 1986), these findings agree qualitatively with studies using both non-labelled and labelled compounds for several tropical crop species (Taylor e t al. 1960; Lindsay and DeMent 1961; Juo and Ellis 1968; Armstrong e t al. 1993). The comparative availability of the P sources to the grasses is controlled by their solubility, which is a function of the surface area of the mineral (Huffman e t al. 1960), and the ability of plants to reduce the solution P concentration at the mineral surface so that ion activity product is less than the Ks, of the mineral. Surface areas of 20-27.5, 10.5, and 0 86-1.98 m2 g-I have been measured for FeP04, AlP04, and strengite, respectively using compounds prepared by similar methodologies to those used in this study (Huffman e t al. 1960; Juo and Ellis 1968). The higher availability of the AlP04 compared to the amorphous FeP04 occurred despite its smaller surface area, possibly reflecting the tendency of iron phosphate to crystallize more rapidly in soil (Juo and Ellis 1968). Alternatively, it may indicate that the solubility of A1 and Fe in the soil favoured higher solubility of Alp04 than FeP04.
There were no significant differences in the specific activity of plant P when K H ~ ~ ~ P O ~ was the source, indicating equal accessibility to all the grasses. This finding is not unexpected as most studies have found no difference between species in their ability to utilize isotopically exchangeable P, once results are corrected for P uptake (Nye and Foster 1958; Probert 1972). Therefore, it appears that differences between these grasses in their responsiveness to soluble P fertilizers (Armstrong and Helyar 1993) are probably due to differences in ability to locate P sources rather than by enhancing its mobilization.

R. D. Armstrong and K. R. Helyar
Similarly, there was little difference in the proportion of plant P derived from the mineral P compounds between the grasses despite significant differences in P uptake. However, there was one exception. C. ciliaris, which was grown on limed soil, derived more of its P content from strengite and less from AlP04 compared with the native grasses. This change in availability of strengite is consistent with expectations from the effect of pH on the solubility of P in the FeP04-Fe(OH)3 system (Lindsay 1979) and higher P solubility resulting from the decreasing solubility of Fe as pH increases (Chang and Jackson 1958). Liming acid soils has been observed to increase neutral salt extractable P from unfertilized soils and from fertilizer residues following several months of reaction (Holford 1989). Other studies have shown that increasing soil pH in the range 4.0-5.5 increases the availability of iron and aluminium phosphates (Huffman et al. 1960; Lindsay and DeMent 1961; Lindsay 1979). In addition to increased solubility as the soil pH increases, greater precipitation of ferric oxides may be favoured with a congruent release of phosphorus (Chang and Jackson 1957). In contrast, the decrease in availability of AlP04 and the small decrease in availability of FeP04 with liming (C. ciliaris) is opposite to expectations of P solubility changes with pH in the A1P04-A1(OH)3 and FeP04-Fe(OH)3 systems. In these cases the effect of liming on A1 and Fe phosphate recrystallization rates, or on adsorption/desorption reactions, may have dominated the effect of pH on P availability. Differences in soil pH of a similar magnitude to that induced by liming the C. ciliaris pots occur between different microsites in mulga shrublands. C. ciliaris usually only establishes in microsites beneath Eucalyptus populnea trees (Ebersohn and Lucas 1965) where the soil pH is similar to that used in C. ciliaris pots and is significantly higher than that of surrounding areas.
Availability of P from Organic Residues
The presence of the grasses produced a significant increase in the release of P from the residues. However, the proportion of plant P derived from the added residues did not differ between A. armata and D. ammophila, and nor did the net release of P, despite marked differences in dry matter production and P uptake between these grasses in the earlier harvests. These results indicate that both grasses possessed a (unidentified) mechanism that either directly or indirectly enhanced the release of the P in the residues. Soil analysis detected decreases in the inorganic P fraction as a result ofoPlant growth, confirming results from other experiments using these grasses (Armstrong and Helyar 1992). The failure of the grasses to deplete soil organic P may reflect the relatively small amount of P absorbed by the grasses relative to the size of this pool in the soil. Alternatively, it may indicate that organic P cycling processes in the soil effectively buffered changes in this fraction.
The two grasses obtained less than 7% of their P from the added residues during the 39 days of the experiment, indicating that this source of P contributes only a small proportion of plant P in the short term. Furthermore, the amount of P released from the residues did not increase between days 15 and 39 suggesting that most of the P release occurred during the first 15 days. This is not surprising as a high proportion of P in rapidly dried residues can occur as inorganic P and is readily leached from the residues (Bromfield and Jones 1972). This effect may be less pronounced in the field as inorganic P contained in residues can

Utilization of P Sources by Grasses
be immobilized (Bromfield and Jones 1972) and is therefore less likely to be leached when subjected to major rainfall events in the following growth season. Differences between P released from the residues and that utilized by the plant indicates that a significant proportion of this P was already lost to other sources, such as microbial biomass (McLaughlin and Alston 1986) or adsorption onto soil particles.
Conclusions
Phosphorus derived from both sparingly soluble sources of mineral P and plant residues can potentially contribute a significant proportion of the P absorbed by these pasture grasses. However, there was little difference between species in their ability to absorb P from the different sources (both mineral and organic) once species differences confounded with soil pH were disregarded (Experiment 1). Therefore, differences between these grasses in their growth responses to varying soil P concentrations can not be explained in terms of differing abilities to mobilize sparingly soluble sources of mineral P or P derived from plant residues.
Acknowledgments
We would like to thank Dr P Sorby (Australian Nuclear Science and Technology Organisation, Lucas Heights) for irradiating the mineral P sources and Drs M. Probert and M. McLaughlin (CSIRO) for comments on the manuscript and A. Milnes and G. Riley for X-ray diffraction measurements.
References Aguilar, S., and Van Diest, A. (1981). Rock-phosphate mobilisation induced by the alkaline
uptake pattern of legumes utilising symbiotically fixed nitrogen. Plant Soil 61, 27-42. Anderson, G. (1964). Investigations on the analysis of inositol hexaphosphate in soils. 8th Int.
Congr. Soil Sci., Bucharest, Romania. Part IV. pp. 563-71. Armstrong, R. D. (1988). The phosphorus nutrition of mulga pasture grasses with reference
to invasion of Aristida armata. Ph.D. Thesis, School of Australian Environmental Studies, Griffith University, Nathan, Queensland.
Armstrong, R. D., and Helyar, K. R. (1992). Changes in soil phosphate fractions in the rhizosphere of semi-arid grasses. Aust. J. Soil Res. 30, 13143.
Armstrong, R. D., and Helyar, K. R. (1993). The relative importance of soil phosphorus to the growth of wiregrass (Aristida ama ta ) compared with other grasses common to the mulga shrublands of Queensland. Aust. J. Exp. Agric. 33, (in press).
Armstrong, R. D., Helyar, K. R., and Prangnell, R. (1993). Direct assessment of mineral phosphorus availability to tropical crops using 3 2 ~ labelled compounds. Plant Soil, (in press).
Bache, B. W. (1963). Aluminium and iron phosphate studies relating to soils. I. Solution and hydrolysis of variscite and strengite. J. Soil Sci. 14, 113-23.
Bolan, N. S., Robson, A. D., and Barrow, N. J. (1987). Effects of vesicular arbuscular mycorrhiza on the availability of iron phosphates to plants. Plant Soil 99, 401-10.
Bromfield, S. M., and Jones, 0. L. (1972). The initial leaching of hayed off pasture plants in relation to the recycling of phosphorus. Aust. J. Agric. Res. 23, 811-24.
Cate, W. E., Huffman, E. O., and Deming, M. E. (1959). Preparation of crystalline ferric phosphates. Soil Sci. 88, 130-2.
Chang, S. C., and Jackson, M. L. (1957). Fractionation of soil phosphorus. Soil Sci. 83, 133-44. Chang, S. C., and Jackson, M. L. (1958). Soil phosphate fractions of some representative soils.
J. Soil Sci. 9, 109-19.

R. D. Armstrong and K. R. Helyar
Christie, E. K., and Moorby, J. (1975). Physiological responses of semi-arid grasses. I. The influence of phosphorus supply on growth and phosphorus absorption. Aust. J. Agric. Res. 26, 423-36.
Dawson, N. M., and Ahern, C. R. (1973). Soils and landscapes of mulga lands with special reference to south-west Queensland. %pica1 Grassl. 7, 23-34.
Deming, M. E., and Cate, W. E. (1963). Preparation of variscite. Soil Sci. 95, 206-7. Ebersohn, J. P., and Lucas, P. (1965). Trees and soil nutrients in south-western Queensland.
Qld J. Agric. Anim. Sci. 22, 431-5. Enwezor, W. 0. (1977). Soil testing for phosphorus in some Nigerian soils. 30 Forms of
phosphorus in soils of south-eastern Nigeria and their relationship to plant available phosphorus. Soil Sci. 124, 27-33.
Grierson, I., Bull, B., and Graham, R. (1991). Soil management and fertiliser strategies. In 'Dryland Farming: A Systems Approach'. (Eds V. Squires and P. Tow.) pp. 134-45. (Sydney University Press/Oxford University Press: Melbourne, Australia.)
Harrison, A. F. (1982). Labile organic phosphorus mineralisation in relationship to soil properties. Soil Biol. Biochem. 14, 343-51.
Hetrick, J. A., and Schwab, A. P. (1992). Changes in aluminium and phosphorus solubilities in response to long-term fertilization. Soil Sci. Soc. Am. J. 56, 755-61.
Holford, I. C. R. (1985). Effects of lime on yields and phosphate uptake by clover in relation to changes in soil phosphate and related characteristics. Aust. J. Soil Res. 23, 75-83.
Holford, I. C. R. (1989). Effects of lime on the residual effectiveness of phosphate in acid soils. Aust. J. Soil Res. 27, 425-32.
Holford, I. C. R., and Crocker, G. J. (1988). Efficacy of various soil phosphate tests for predicting phosphate responsiveness and requirements of clover pastures on acidic Tableland soils. Aust. J. Soil Res. 26, 479-88.
Huffman, E. O., Cate, W. E., and Deming, M. E. (1960). Rates and mechanisms of dissolution of some ferric phosphates. Soil Sci. 90, 8-15.
Juo, A. S. R., and Ellis, B. G. (1968). Chemical and physical properties of iron and aluminium phosphates and their relation to phosphorus availability. Soil Scz. Soc. Am. Proc. 32, 216-21.
Lindsay, W. L. (1979). 'Chemical Equilibria in Soils'. (John Wiley: New York.) Lindsay, W. L., and DeMent, J. D. (1961). Effectiveness of some iron phosphates as sources
of phosphorus for plants. Plant Soil 14, 118-26. McLaughlin, M. J . , and Alston, A. M. (1986). The relative contribution of plant residues and
fertiliser to the phosphorus nutrition of wheat in a pasture/cereal system. Aust. J. Soil Res. 24, 517-26.
Murphy, J., and Riley, J. P. (1962). A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 27, 31-6.
Norrish, K., and Rosser, H. (1983). Mineral phosphate. In 'Soils: An Australian Viewpoint'. pp. 336-61. (CSIRO Aust./Academic Press: Melbourne.)
Northcote, K. H. (1971). 'A Factual Key for the Recognition of Australian Soils'. (CSIRO Aust./Rellim Tech. Publ.: Adelaide.)
Nye, P. H., and Foster, W. N. M. (1958). A study of the mechanism of soil-phosphate uptake in relation to plant species. Plant Soil 9, 338-52.
Olsen, S. R., Cole, C. V., Watanabe, F. S., and Dean, L. A. (1954). Estimation of available phosphorus in soils by extraction with sodium bicarbonate. Circular No. 939. (United States Department of Agriculture: Washington.)
Ozanne, P. G., and Shaw, T. C. (1967). Phosphate sorption by soils as a measure of the phosphate requirement for pasture growth. Aust. J. Agric. Res. 18, 601-12.
Probert, M. E. (1972). The dependence of isotopically exchangeable phosphate (L-value) on phosphate uptake. Plant Soil 36, 141-48.
Russell, E. W. (1973). 'Soil Conditions and Plant Growth'. 10th edn. (Longmans: London.) Sanyal, S. K., and De Datta, S. K. (1991). Chemistry of phosphorus transformations in soil.
Adv. Soil Sci. 16, 1-120. Saunders, W. M. H., and Williams, E. G. (1955). Observations on the determination of total
organic phosphorus in soils. J. Soil Sci. 6, 245-67.

Utilization of P Sources by Grasses
Silcock, R. G., Noble, A., and Whalley, R. D. B. (1976). Importance of phosphorus and nitrogen in the nutrition of grass seedlings growing in mulga soil. Aust. J. Agric. Res. 27, 583-92.
Smith, A. N. (1966). The role of inorganic phosphate in supplying phosphorus to plants. Agrochimica 11, 79-89.
Sposito, G. (1985). Chemical models of weathering in soils. In 'The Chemistry of Weathering'. (Ed. J. I. Drever.) pp. 1-18. (Reidel: Boston.)
Taylor, A. W., Gurney, E. L., and Lindsay, W. L. (1960). An evaluation of some iron and aluminium phosphates as sources of phosphate for plants. Soil Sci. 90, 22-31.
Manuscript received 11 May 1992, accepted 9 February 1993