university of veterinary medicine hannover mosquito-borne
TRANSCRIPT

University of Veterinary Medicine Hannover
Mosquito-borne Flaviviruses: Vector and Avian Host
Susceptibility for West Nile Virus and Usutu Virus in Germany
Inaugural-Dissertation
in fulfillment of the academic degree Doctor medicinae veterinariae
(Dr. med. vet)
submitted by
Cora Marielle Holicki
Stade
Hannover 2020

Academic supervision: Prof. Dr. Martin H. Groschup Friedrich-Loeffler-Institut Federal Research Institute for Animal Health Institute of Novel and Emerging Infectious Diseases Greifswald-Insel Riems
1. Referee: Prof. Dr. Martin H. Groschup
Friedrich-Loeffler-Institut Federal Research Institute for Animal Health Institute of Novel and Emerging Infectious Diseases Greifswald-Insel Riems
2. Referee: Prof. Dr. Stefanie Becker
University of Veterinary Medicine Hannover Institute for Parasitology Research Center for Emerging Infections and Zoonoses Hannover
Day of the oral examination: 10th of November 2020 Sponsorship: This work was supported by the German Federal
Ministry of Food and Agriculture (BMEL) through the Federal Office for Agriculture and Food (BLE) (project: CuliFo).

To my beloved family


Parts of this work were already published in the following journals: West Nile virus lineage 2 vector competence of indigenous Culex and Aedes mosquitoes
from Germany at temperate climate conditions
Cora M. Holicki, Ute Ziegler, Cristian Răileanu, Helge Kampen, Doreen Werner, Jana Schulz, Cornelia Silaghi, Martin H. Groschup, and Ana Vasić Viruses (2020), Volume 12, Issue 5, 561
Pathogenicity of West Nile virus lineage 1 to German poultry
Cora M. Holicki, Friederike Michel, Ana Vasić, Christine Fast, Martin Eiden, Cristian Răileanu, Helge Kampen, Doreen Werner, Martin H. Groschup, and Ute Ziegler Vaccines (2020), Volume 8, Issue 3, 507 Manuscripts extracted from the doctorate project: German Culex pipiens biotype molestus and Culex torrentium are vector-competent
for Usutu virus
Cora M. Holicki, Dorothee E. Scheuch, Ute Ziegler, Julia Lettow, Helge Kampen, Doreen Werner, and Martin H. Groschup (This manuscript has been submitted and peer-reviewed) Intrathoracic injection of German Culex mosquitoes with West Nile virus to secure virus
infection and transmission Cora M. Holicki, Ute Ziegler, Cristian Răileanu, Helge Kampen, Doreen Werner, Cornelia Silaghi, Martin H. Groschup, and Ana Vasić (To be submitted) Further publications: Comparison of vector competence of Aedes vexans Green River and Culex pipiens
biotype pipiens for West Nile virus lineages 1 and 2
Elisabeth Wöhnke, Ana Vasić, Cristian Răileanu, Cora Marielle Holicki, Birke Andrea Tews, and Cornelia Silaghi Zoonoses Public Health (2020), Volume 67, Issue 4, Pages 416-424 West Nile virus mosquito vectors (Diptera: Culicidae) in Germany
Helge Kampen, Cora M. Holicki, Ute Ziegler, Martin H. Groschup, Birke Andrea Tews, and Doreen Werner Viruses (2020), Volume 12, Issue 5, 493

Experimental data were also published at national and international conferences:
Vector capacity of indigenous mosquitoes for Rift Valley fever virus and Usutu virus
and their transmission to vertebrate hosts
Cora M. Holicki, Dorothee E. Scheuch, Ute Ziegler, Julia Lettow, Helge Kampen, and Martin H. Groschup 6. FLI- Junior Scientist Symposium, Braunschweig, Germany, 20./21./22.09.2017, Poster presentation
Transmission of West Nile virus from indigenous mosquitoes to domestic poultry
Cora M. Holicki, Friederike Michel, Ute Ziegler, Anna Heitmann, Stephanie Jansen, Helge Kampen, Doreen Walther, and Martin H. Groschup 7. FLI- Junior Scientist Symposium, Greifswald-Insel Riems, Germany, 24./25./26.09.2018, Poster presentation
Flaviviruses in Germany: Vector competence of northern and southern European Culex
pipiens biotype molestus for Usutu and West Nile virus
Cora M. Holicki, Ana Vasić, Dorothee E. Scheuch, Ute Ziegler, Julia Lettow, Doreen Werner, Dušan Petrić, Helge Kampen, Cornelia Silaghi, and Martin H. Groschup 13th EPIZONE Annual Meeting "Breaking Walls", Berlin, Germany, 26./28.08.2019, Poster presentation
West West Nile virus in Germany: Vector competence of native Culex pipiens and Aedes
albopictus Cora M. Holicki, Ute Ziegler, Helge Kampen, Doreen Werner, Cornelia Silaghi, Martin H. Groschup, and Ana Vasić Zoonoses 2019 - International Symposium on Zoonoses Research, Berlin, Germany, 16./17./18.09.2019, Oral presentation

Table of Contents i
TABLE OF CONTENTS
LIST OF ABBREVIATIONS ............................................................................. v
LIST OF TABLES ............................................................................................ vii
LIST OF FIGURES ........................................................................................... ix
1 INTRODUCTION ....................................................................................... 1
2 LITERATURE REVIEW ........................................................................... 5
2.1 History and Epidemiology ................................................................ 5
2.1.1 West Nile Virus .................................................................... 5
2.1.2 Usutu Virus ........................................................................... 6
2.2 Taxonomy and Virus Structure ......................................................... 7
2.3 Replication ........................................................................................ 8
2.4 Transmission ................................................................................... 10
2.4.1 Enzootic Cycle .................................................................... 10
2.4.2 Epidemic/Epizootic Cycle .................................................. 12
2.4.3 Infection Pattern in a Competent Mosquito Vector ............ 13
2.4.4 Vectorial Capacity and Vector Competence ...................... 15
2.4.5 Infection Pattern in a Susceptible Avian Host .................... 16
2.5 Clinical Presentation and Pathology ............................................... 17
2.5.1 West Nile Virus .................................................................. 17
2.5.2 Usutu Virus ......................................................................... 19
2.6 Detection ......................................................................................... 20
2.7 Prevention and Treatment ............................................................... 20
3 MATERIALS AND METHODS ............................................................. 23
3.1 Mosquito Infection .......................................................................... 23
3.2 Vector Competence Indices ............................................................ 23
3.3 Animal Trials .................................................................................. 24
3.4 RNA Extraction and RT-qPCR ....................................................... 25

ii Table of Contents
4 MANUSCRIPT I ....................................................................................... 27
German Culex pipiens Biotype molestus and Culex torrentium are Vector Competent for Usutu Virus
4.1 Abstract ........................................................................................... 28
4.2 Introduction ..................................................................................... 28
4.3 Materials and Methods .................................................................... 30
4.3.1 Mosquito Collection, Rearing, and Identification .............. 30
4.3.2 Virus Strain and Cultivation .............................................. 31
4.3.3 Oral Infection with USUV .................................................. 31
4.3.4 Forced Salivation Assay ..................................................... 32
4.3.5 Pathogen Screening in Mosquito Bodies and Legs plus Wings .................................................................................. 32
4.3.6 Interpretation of Results ..................................................... 33
4.3.7 Data Analysis ...................................................................... 33
4.3.8 Ethics Statement ................................................................. 34
4.4 Results ............................................................................................. 34
4.4.1 Feeding and Survival Rates ................................................ 34
4.4.2 Infection, Dissemination, and Transmission Rates ............ 34
4.5 Discussion ....................................................................................... 35
4.6 Conclusion ...................................................................................... 39
4.7 Tables .............................................................................................. 40
4.8 Figures ............................................................................................. 42
4.9 Acknowledgments ........................................................................... 44
5 MANUSCRIPT II ...................................................................................... 45
West Nile Virus Lineage 2 Vector Competence of Indigenous Culex and Aedes Mosquitoes from Germany at Temperate Climate Conditions
5.1 Abstract ........................................................................................... 45
6 MANUSCRIPT III .................................................................................... 47

Table of Contents iii
Intrathoracic Injection of German Culex Mosquitoes with West Nile Virus to Secure Virus Infection and Transmission
6.1 Abstract ........................................................................................... 48
6.2 Introduction ..................................................................................... 48
6.3 Materials and Methods .................................................................... 49
6.3.1 Mosquito Origin and Rearing ............................................. 49
6.3.2 Mosquito Infection .............................................................. 49
6.3.3 Mosquito Processing ........................................................... 50
6.3.4 Vector Competence Indices and Data Analysis ................. 50
6.4 Results ............................................................................................. 51
6.5 Discussion ....................................................................................... 51
6.6 Tables .............................................................................................. 52
6.7 Figures ............................................................................................. 53
6.8 Acknowledgments ........................................................................... 54
7 MANUSCRIPT IV .................................................................................... 55
Pathogenicity of West Nile Virus Lineage 1 to German Poultry
7.1 Abstract ........................................................................................... 55
8 GENERAL DISCUSSION........................................................................ 57
9 SUMMARY ................................................................................................ 69
10 ZUSAMMENFASSUNG .......................................................................... 73
AUTHORS’ CONTRIBUTION ....................................................................... 77
ACKNOWLEDGMENTS ................................................................................ 79
BIBLIOGRAPHY ............................................................................................. 81
SUPPLEMENTS ............................................................................................. 109
A. Manuscript I (Chapter 4) ....................................................................... 111
B. Manuscript III (Chapter 6) .................................................................... 115


List of Abbreviations v
LIST OF ABBREVIATIONS
arbovirus Arthropod-borne virus BNITM Bernhard-Nocht-Institut für Tropenmedizin BSL Biosafety level C protein Capsid protein CCLV Collection of Cell Lines in Veterinary Medicine CDC Centers for Disease Control and Prevention CHIKV Chikungunya virus CI Confidence interval CNS Central nervous system CO2 Carbon dioxide DC-SIGN Dendritic cell-specific intercellular adhesion molecule-3-grabbing
non-integrin DENV Dengue virus dpi Day(s) post infection DNA Deoxyribonucleic acid DR Dissemination rate DWD Deutscher Wetter Dienst e.g. Exempli gratia (“for example”) ELISA Enzyme linked immunosorbent assay E protein Envelope protein ECDC European Centre for Disease Prevention and Control ER Endoplasmic reticulum et al. Et alia (“and others”) etc. Et cetera (“and other similar things”) FCS Fetal calf serum FLI Friedrich-Loeffler-Institut GLM Generalized binomial regression model h Hours Hz Hertz i.e. Id est (“that is”) IHC Immunohistochemistry IMD Immune deficiency IR Infection rate JAK Janus Kinase JEV Japanese encephalitis virus LSM Least-squares mean M protein Membrane protein MEM Minimal essential medium

vi List of Abbreviations
mL Milliliter NA Not applicable nm Nanometer NS Nonstructural PBS Phosphate buffered saline PCR Polymerase chain reaction PFU Plaque forming units PrM protein Premembrane protein PRNT Plaque reduction neutralization test RNA Ribonucleic acid RNAi RNA interference rpm Revolutions per minute RT-qPCR Quantitative real-time reverse transcription polymerase chain reaction s.l. Sensu lato (“in a broad sense”) spp. Species pluralis (+)ssRNA Positive-sense, single-stranded RNA (-)ssRNA Negative-sense, single-stranded RNA STAT Signal Transducer and Activator Transcription Sup. Supplemental TBEV Tick-borne encephalitis virus TCID50 Tissue culture infective dose 50 TE Transmission efficiency TR Transmission rate UK The United Kingdom µL Microliter µm Micrometer USA The United States of America USDA United States Department of Agriculture USUV Usutu virus vs. Versus VNT Virus neutralization test WNND West Nile neuroinvasive disease WNV West Nile virus YFV Yellow fever virus ZIKV Zika virus

List of Tables vii
LIST OF TABLES
Table 3.1 Definition of vector competence indices. ........................................... 24
Table 4.1 Infection, dissemination, and transmission rates of mosquitoes
infected with the German USUV Africa 2 strain. ............................................... 40
Table 6.1 Infection, dissemination, and transmission rates of Culex species after
intrathoracic injection with West Nile virus (WNV). ......................................... 52
Sup. Table 4.1 Overview on explanatory variables included in the final
generalized binomial regression models (GLM) for the investigated rates at 25
°C. ...................................................................................................................... 111
Sup. Table 4.2 p-values of the fixed effects in the least-square means analysis
when comparing the rates between all species at 25 °C in the final generalized
binomial regression models (GLM) specified in Sup. Table 4.1. ..................... 112
Sup. Table 4.3 p-values of the interaction term in the least-square means
analysis for the survival, infection, and transmission rates at 25 °C. ............... 113
Sup. Table 6.1 p-values of the fixed effects in the least-square means analysis
when comparing the rates between all species at 25 °C in the final generalized
binomial regression models (GLM). ................................................................. 115


List of Figures ix
LIST OF FIGURES
Figure 2.1 Taxonomy of West Nile virus and Usutu virus .................................. 8
Figure 2.2 West Nile virus (WNV) genome organization (a) and WNV
replication (b). ....................................................................................................... 9
Figure 2.3 Transmission cycle of the flavivirus West Nile virus. ...................... 10
Figure 2.4 Arbovirus migratory route in vector and host................................... 14
Figure 2.5 Factors influencing vectorial capacity. ............................................. 15
Figure 4.1 Graphical abstract displaying work flow of vector competence
experiments. ........................................................................................................ 42
Figure 4.2 Comparison of the feeding and survival rates (from 0 to 14/16 dpi) of
the four tested mosquito populations. ................................................................. 43
Figure 6.1 Viral loads in (A) mosquito bodies and (B) legs plus wings of Culex
pipiens biotype pipiens (Cx. p. p.) and Cx. torrentium (Cx. tor.) populations
from Mecklenburg-Western Pomerania (M) and/or Brandenburg (B)
intrathoracically injected with WNV 10/11 or 22 days post infection (dpi),
respectively. ......................................................................................................... 53


1 Introduction 1
1 INTRODUCTION
West Nile virus (WNV) and Usutu virus (USUV) are closely related mosquito-borne flaviviruses that pose an ongoing threat to animal (avian and equine) and human health. Both viruses (co-)exist in a sylvatic/enzootic cycle between susceptible, predominantly free-ranging bird species and ornithophilic mosquito species (e.g., Culex species). There exist, however, also mosquito species that are considered opportunistic feeders, feeding indiscriminately on both birds and mammals (e.g., Cx. quinquefasciatus or Cx. pipiens hybrids of the pipiens and molestus biotypes). This behavioral characteristic allows them to transmit viruses to so called dead-end hosts, such as horses and humans. These incidental hosts can develop clinical symptoms but will not produce viremia levels high enough to infect naïve mosquitoes and are, therefore, inadequate in perpetuating the transmission cycles of WNV and USUV. Disease manifestation in susceptible hosts is species-dependent and can range from asymptomatic to febrile, with nonspecific symptoms, to life-threatening and fatal with meningitis and encephalitis.
To date, WNV is a global health concern, with virus detections in every continent aside from Antarctica. The virus was first isolated in 1937 from a febrile patient in the West Nile province of Uganda. In 1953, it was isolated from several avian species in Egypt, followed by two major epornitics in Israel (1997-2000). In 1999, WNV (lineage 1) crossed the Atlantic Ocean and reached North America, resulting in an unprecedented outbreak in New York City. The virus was associated with high fatality rates in crows (Corvus spp.) and numerous other avian species (up to 342 in total) as well as horses and humans. In Europe, primarily around the Mediterranean basin, WNV has been circulating since the 1950s and was at first only associated with sporadic cases or size- and time-limited human and equine outbreaks. Only in the early 1990s, with a reemergence of WNV, did incidence and disease severity shift. This was initiated by two significant human outbreaks in Romania (1996) and Russia (1999), followed by an unanticipated outbreak in France (2000). In 2004, a new genetic lineage of WNV, WNV lineage 2, was isolated from a dead Hungarian goshawk. This lineage rapidly spread throughout central and southern Europe, resulting in seasonal outbreaks inter alia in Greece, Romania, Italy, Serbia, and Spain. Since 2018, there have been numerous detections of WNV also in Germany.
Usutu virus also has its origin in Africa, with its first isolation near the Usutu River in Swaziland, South Africa. After multiple detections in mosquitoes in several other African countries, USUV was associated in 2001 with the mass mortality of avian species in Vienna, Austria. Eurasian blackbirds (Turdus merula) and great grey owls (Strix nebulosa) were primarily affected. A retrospective study of dead blackbirds in Italy, however, confirmed that USUV must have already been present in Europe since 1996. In the following years, the geographic range of USUV expanded to several European countries (Hungary, Switzerland, Spain, and Italy), with the first record in Germany in mosquitoes in 2010. Outbreaks in birds,

2 1 Introduction
for example, in 2011, 2016, and 2018 had large scale impacts on the avian wildlife in Germany, with high mortality rates of Eurasian blackbirds.
As both flaviviruses only recently emerged in Germany, research indicating which indigenous mosquito species are involved in the perpetuation of these viruses is scarce. This is especially true for USUV, for which vector competence experiments with German mosquito populations have not yet been performed. The first study described in this thesis, therefore, focused on the susceptibility of indigenous Cx. pipiens biotype molestus and Cx. torrentium for USUV lineage Africa 2. This strain was isolated in 2015 from two succumbed great grey owls from the zoological garden in Berlin and was linked to a distinct virus introduction from Africa to Europe. The Cx. pipiens biotype molestus colony was established in 2012 from egg rafts collected in northern Germany (Lower Saxony). Culex torrentium egg rafts were collected near Berlin and Bonn and the emerged mosquitoes (i.e., F0 generation) were directly used for the experiments. The results of the two German Culex populations were compared to those of an established Cx. pipiens biotype molestus colony from the Republic of Serbia and an Aedes aegypti colony from Malaysia. Adult female mosquitoes were allowed to feed on a virus-spiked blood meal (high or low dose) and engorged females were incubated for two or three weeks at 25 °C. After 14/16 or 21 days a saliva assay was performed to assess whether the saliva of individual mosquitoes contained viable virus particles (i.e., the mosquito species is vector-competent and can transmit USUV under laboratory settings). To investigate the infection of the midgut and the dissemination to secondary tissues the mosquito bodies (thorax and abdomen) and mosquito legs plus wings were also examined via nucleic acid amplification (i.e., polymerase chain reaction (PCR)).
In the second study, the vector competence of German Culex mosquitoes was examined for the newly registered German WNV lineage 2 strain. The established Cx. pipiens biotype molestus colony from Lower Saxony was used again, as well as two Cx. pipiens biotype pipiens populations collected in Brandenburg in 2018 (established colony) and in 2019 (F0 generation). An established colony of the invasive mosquito species Ae. albopictus, collected from Thuringia, Germany was also tested. Similar to the study with USUV, these results were again compared to those of the southern European Cx. pipiens biotype molestus from the Republic of Serbia. In view of the excessively warm and dry summers of 2018 and 2019, the temperature dependency of WNV vector competence was also analyzed.
After the identification of vector-competent mosquitoes for WNV, virus transmission between infectious mosquitoes and susceptible avian hosts was reproduced. Furthermore, the last studies also focused on uncovering the role domestic poultry plays in the epidemiology of WNV in Germany. First, juvenile three-week-old chickens, ducks, and geese were infected subcutaneously to assess the virulence and pathogenicity of an Italian WNV lineage 1 strain to domestic poultry. An Italian strain was used as at that time WNV had not yet been isolated in Germany and its introduction into the country was postulated to occur from the South. The bird’s wellbeing was monitored daily according to a score sheet and blood and swab samples

1 Introduction 3
(oropharyngeal and cloacal) were taken following a set time schedule. After three weeks the experiments were terminated and tissue samples were collected during necropsy. The most susceptible poultry species from the first studies (i.e., geese), was also exposed to infectious Culex mosquitoes in a further experiment. For this, Cx. pipiens biotype pipiens from Brandenburg (F0 generation) were intrathoracically injected with WNV ten days prior to being allowed to feed on the geese. For this the intrathoracic injection technique was implemented beforehand. Rather than artificial feeding, intrathoracic injections with WNV can ensure higher transmission efficiencies after a given incubation period. After exposure to the geese, the mosquitoes were subjected to a saliva assay. Saliva samples were inoculated onto mammalian cells to check for viable virus particles while mosquito heads, bodies (thorax and abdomen), and extremities (legs plus wings) were tested for WNV-specific ribonucleic acid (RNA). The geese were, as in the prior experiments, monitored and sampled for three weeks post exposure.


2 Literature Review 5
2 LITERATURE REVIEW
2.1 History and Epidemiology
2.1.1 West Nile Virus
With the exception of Antarctica, West Nile virus (WNV) is nowadays present in every continent in the world, making it the most widely distributed arthropod-borne virus (arbovirus) (Chancey et al. 2015). The virus was initially isolated from a febrile patient in Uganda in 1937 (Smithburn et al. 1940), followed by sporadic cases as well as outbreaks in Africa, Eurasia, Australia, and the middle East (Hayes 2001). For example, in Egypt (Schmidt and Elmansoury 1963, Taylor et al. 1956) and Israel (Bernkopf et al. 1953, Spigland et al. 1958) WNV was detected in mosquitoes, birds, and humans. In Europe, the first major outbreak of WNV was recorded in France (1962-1965) in wild and domestic horses, with only occasional detections in humans, and mosquitoes (Joubert et al. 1970, Murgue et al. 2001b, Pantheir et al. 1966). Up until 1996, epidemics with human infections, like the one in France, were rarely accompanied by severe neurological manifestations (Hayes 2001). However, with the transition into the 21st century, WNV virulence began to shift. The next major outbreak in Romania (1996-2000) was associated with severe neuroinvasive diseases in humans (Ceianu et al. 2001, Dinu et al. 2015, Han et al. 1999, Tsai et al. 1998). Concurrently, Russia (1999) also experienced an outbreak with 84 human cases of meningoencephalitis of which 40 were fatal (Platonov et al. 2001). In 1999, WNV reached the Western Hemisphere after a single point introduction into New York City (Ebel et al. 2001, Lanciotti et al. 1999). The virus was phylogenetically closely related to a strain isolated in Israel in the previous year (Lanciotti et al. 1999). Reports of succumbed exotic and domestic birds in the New York City area presaged infections in horses and humans (Steele et al. 2000). As WNV encountered a naïve environment in the Western Hemisphere, it quickly dispersed from the East to the West Coast of the United States of America (USA) and to Canada, Mexico, Central America, the Caribbean, and South America (Bosch et al. 2007, Elizondo-Quiroga et al. 2005, Komar and Clark 2006, Morales-Betoulle et al. 2006, Morales et al. 2006). In the USA, 24,657 neuroinvasive disease cases, with 2,330 deaths, have been reported by the Centers for Disease Control and Prevention (CDC) since the introduction of WNV up until 2018 (CDC 2019). These historic WNV outbreaks were all attributed to WNV lineage 1 (Chancey et al. 2015).
The appearance of WNV lineage 2 in Hungary (Bakonyi et al. 2006) and Russia (Platonov et al. 2011) in 2004 further increased disease severity and incidence. Unlike the Russian isolate, lineage 2 from Hungary rapidly expanded to central and southern Europe. Phylogeographic models propose that one group (Clade A) spread through Austria northwards to the Czech Republic or southwards to Italy, while Clade B spread south- and eastwards through the Balkans (Camp and Nowotny 2020). Numerous epidemics were recorded in the past two decades, such as in Austria in 2008 (Bakonyi et al. 2013), Greece in 2010 (Danis et al. 2011a), Romania in 2010 (Sirbu et al. 2011), Spain in 2010 (García-Bocanegra et al. 2011), Italy in 2011 (Bagnarelli

6 2.1 History and Epidemiology
et al. 2011, Magurano et al. 2012), and Serbia in 2012 (Popović et al. 2013). In 2018, a substantial outbreak hit central and southern Europe with the European Centre for Disease Prevention and Control (ECDC) reporting a 7.2-fold increase in the number of confirmed human infections (n = 2,083) (ECDC 2018). In that year, WNV was also registered in countries with a more temperate climate, such as in Germany, where it was isolated from several resident wild and aviary birds (Ziegler et al. 2019). In the following year, autochthonous WNV cases were documented also in equines and humans, and mosquitoes (ECDC 2019, Kampen et al. 2020, Ziegler et al. 2020). To date, Europe is facing seasonal and regional WNV lineage 2 outbreaks with virus activity from mid-June to mid-November, correlating with mosquito abundance (Sambri et al. 2013).
To date WNV has been most commonly classified into seven different genetic lineages (reviewed in Rizzoli et al. 2015) of which lineage 1 and lineage 2 are most frequent. Lineage 1 includes three subclades: clade 1a, 1b, and 1c. Subclade 1a is the Mediterranean and former eastern European subtype while 1b and 1c are only found in Australia (Kunjin virus strains) and India, respectively. Lineage 2 is associated with Africa and since 2004 also central Europe and it derived from the two independent introductions into Hungary and Russia. The remaining lineages are considered less relevant as they have not been connected to diseases in humans nor animals.
2.1.2 Usutu Virus
Usutu virus (USUV) was first isolated in 1959 from a Cx. neavei mosquito caught near the Usutu River in Swaziland, South Africa (Williams et al. 1964). Thereafter, it was detected in numerous African countries: Central African Republic, Senegal, Côte d´Ivoire, Nigeria, Uganda, Burkina Faso, Tunisia, and Morocco (Ben Hassine et al. 2017, Durand et al. 2016, Mossel et al. 2017, Nikolay et al. 2011). USUV dispersed from the African continent and appeared in 2001 in Europe in the city of Vienna, Austria (isolated from bird carcasses) (Weissenböck et al. 2002) and in 2014-2015 in Israel (isolated from mosquitoes) (Mannasse et al. 2017). In Vienna, USUV caused mass mortality events of Eurasian blackbirds and great grey owls (Weissenböck et al. 2002). Retrospective analysis of preserved paraffinated tissue blocks from succumbed Eurasian blackbirds in the Tuscany region of Italy, however, indicate that USUV was already present in Europe since 1996 (Weissenböck et al. 2013). Phylogenetic analyses suggest three unique USUV introductions from Africa into Europe with the help of migratory birds as long-distance dispersal vehicles: twice (1950s and 1990s) into Spain along an eastern Atlantic migratory route and once (1980s) into central Europe along a Black Sea/Mediterranean migratory route (Clé et al. 2019, Engel et al. 2016, Weissenböck et al. 2013). At present, surveillance studies, nevertheless, provide evidence for ongoing introduction events into Europe from Africa. For example, only recently were African lineages detected in Germany (Ziegler et al. 2016) and France (Eiden et al. 2018). Until 2015, USUV infections had been detected in 12 European countries (Austria, Belgium, Croatia, France, Germany, Greece, Hungary, Italy, the Czech Republic, Serbia, Spain, and Switzerland (Ashraf et al. 2015, Eiden

2 Literature Review 7
et al. 2018, Kemenesi et al. 2018, Lecollinet et al. 2016, Scheuch et al. 2018, Vilibic-Cavlek et al. 2020)) and were associated with periodic small epornitics. In the summer of 2016 and 2018, more substantial USUV epidemics occurred affecting primarily the avifauna but also public health (Aberle et al. 2018, Cadar et al. 2017b, Michel et al. 2018, Michel et al. 2019).
In Germany, the earliest detection of USUV was in 2010, in a pool of Cx. pipiens mosquitoes from southwestern Germany (Jöst et al. 2011). Its introduction lead to mass mortalities in Eurasian blackbirds (2011) (Becker et al. 2012, Cadar et al. 2017a) and to at least two human infections in the following two years around the Upper Rhine Valley (Allering et al. 2012, Cadar et al. 2017b). Studies estimated that as a consequence the local blackbird population declined by more than 50% (Lühken et al. 2017). Nonetheless, confined to individual detections as in Bonn (2014) and Berlin (2015), USUV infections in birds steadily decreased from 2013 onwards (Michel et al. 2018). In 2016, USUV reemerged in multiple northern European countries like Belgium and the Netherlands but also Germany, where a large outbreak in North Rhine-Westphalia and around Leipzig, Saxony and Halle, Saxony-Anhalt occurred (Cadar et al. 2017a, Michel et al. 2018, Sieg et al. 2017). In the following year, USUV expanded further to the north of Germany reaching Hannover, Bremen, and Hamburg. In 2018, another detrimental outbreak hit Germany, involving for the first time all national federal states and the death of several hundred Eurasian blackbirds (Michel et al. 2019).
Phylogenetic studies clustered USUV into eight distinct lineages based on their geographic origin of isolation: Africa 1, 2, and 3 and Europe 1, 2, 3, 4, and 5. The African 2 strain SAAR-1776 is, thereby, only distantly related to the European strains (Gaibani and Rossini 2017). In Germany, USUV Africa 2, Africa 3, Europe 3, and Europe 5 are currently circulating and Europe 2 was detected in 2019 for the first time (Cadar et al. 2017a, Michel et al. 2019, Sieg et al. 2017).
2.2 Taxonomy and Virus Structure
The Flavivirus genus is one of four genera belonging to the family Flaviviridae and order Amarillovirales. The genus comprises over 70 arboviruses (a selection portrayed in Figure 2.1), which are classified into tick-borne and mosquito-borne virus groups. The mosquito-borne cluster (also known as the encephalitic clade) includes the largest of the ten serologic/genetic complexes, the Japanese encephalitis serocomplex. This serocomplex further consists of ten members, including WNV and USUV. These members are grouped together on the basis of their cross-neutralizing properties in polyclonal sera (International Committee on Taxonomy of Viruses (ICTV) 2020, Kuno et al. 1998, Poidinger et al. 1996).
The Flavivivirus virion is a small (40-60 nm in diameter), enveloped, icosahedral particle containing a single-stranded, positive-sense RNA genome of approximately 11 kilobase pairs (Brinton 2013, Rice et al. 1985, Simmonds et al. 2017). The translated single polyprotein is divided into three structural (capsid (C), envelope (E), and premembrane (PrM)) and seven

8 2.3 Replication
nonstructural proteins (NS1, NS2A, NS2B, NS3, NS4A, NS4B, and NS5) on the basis of host and viral proteases (Figure 2.2 a) (Barrows et al. 2018, Rice et al. 1985, Simmonds et al. 2017). While the structural proteins are necessary for virus entry, fusion, and encapsidation, the nonstructural proteins serve a broad range of functions as, for example, viral replication, virion assembly, and inhibiting the host’s innate immune response (Colpitts et al. 2012).
2.3 Replication
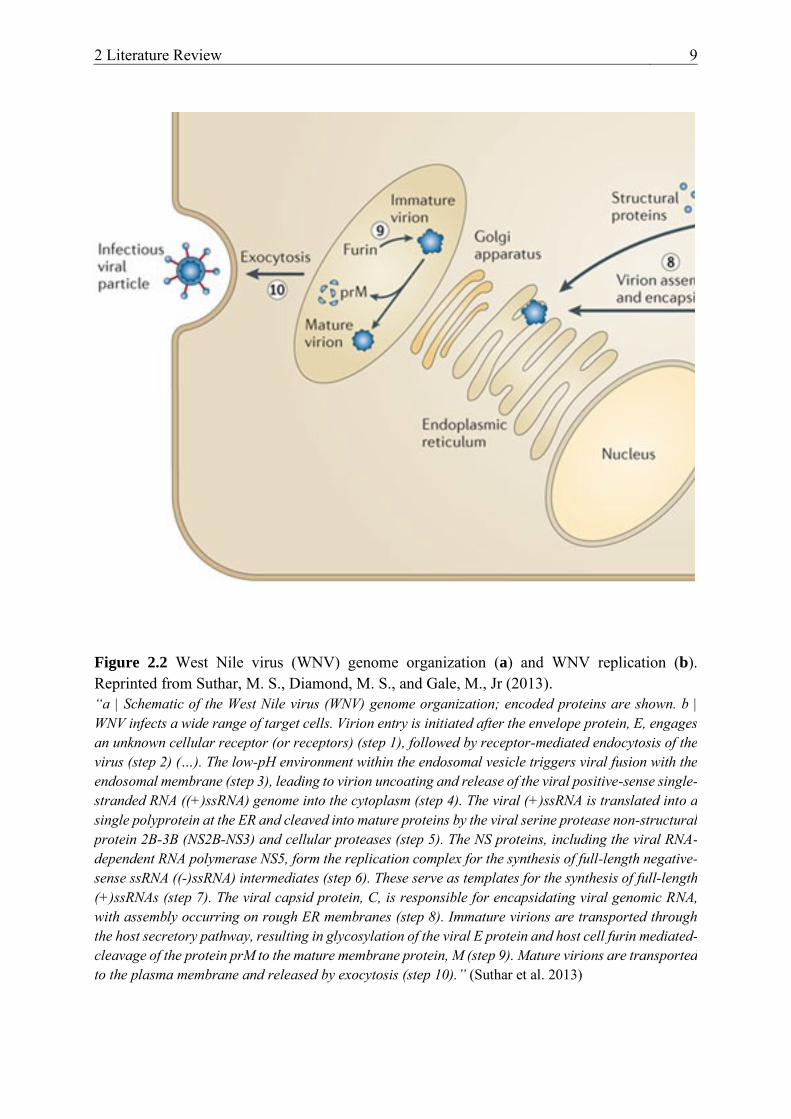
The replication of flaviviruses such as WNV (Figure 2.2 b) (nicely summarized by Colpitts et al. 2012) takes place in the cytoplasm and begins with virus entry via receptor-mediated endocytosis of clathrin coated pits (Chu and Ng 2004). For this, the envelope protein E binds with an undefined cellular receptor. Studies have proposed that lectins (e.g., dendritic cell-specific intercellular adhesion molecule-3-grabbing non-integrin (DC-SIGN)), mannose receptors, and several glycosaminoglycans play a role here (Davis et al. 2006, Lee et al. 2004, Tassaneetrithep et al. 2003). An acidic environment (pH 6.3-6.4) in the virus-containing endosome results in conformational changes of the envelope protein and fusion of the viral lipid membrane with the endosomal membrane. This is followed by virion uncoating and release of a viral, positive-sense, single-stranded RNA ((+)ssRNA) genome into the cytoplasm (Modis et al. 2004). The genome is translated into a single polyprotein at the endoplasmic reticulum and cleaved into ten mature viral proteins by viral and cellular proteins. These structural and non-structural proteins assist in further viral replication and virion assembly. At the same time, the viral RNA-dependent RNA polymerase copies complementary, negative polarity strands ((-)ssRNA) from the positive polarity genomic RNA template ((+)ssRNA). These in turn serve as templates for the synthesis of new positive-sense viral RNA. Next the structural proteins assemble onto membranes of the endoplasmic reticulum (ER) and the viral capsid structural protein promotes viral genomic RNA encapsidation. The virus buds into the cytoplasm through the Golgi network. Lastly, through maturation (furin mediated-cleavage of the protein prM to the mature membrane protein M) of the exocytic vesicle, mature virus is released from the cell surface (Brinton 2013, Lindenbach et al. 2007).
Figure 2.1 Taxonomy of West Nile virus and Usutu virus.
Amarillovirales
Flaviviridae
Flavivirus
Dengue virus
Japanese encephalitis
virus
Murray Valley
encephalitis virus
Saint Louis encephalitis
virus
Tick-borne encephaliltis
virusUsutu virus
West Nile virus
Yellow fever virus
Zika virus
Hepacivirus Pegivirus PestivirusGenus
Family
Species
Order
2 Literature Review 9
Figure 2.2 West Nile virus (WNV) genome organization (a) and WNV replication (b). Reprinted from Suthar, M. S., Diamond, M. S., and Gale, M., Jr (2013). “a | Schematic of the West Nile virus (WNV) genome organization; encoded proteins are shown. b | WNV infects a wide range of target cells. Virion entry is initiated after the envelope protein, E, engages an unknown cellular receptor (or receptors) (step 1), followed by receptor-mediated endocytosis of the virus (step 2) (…). The low-pH environment within the endosomal vesicle triggers viral fusion with the endosomal membrane (step 3), leading to virion uncoating and release of the viral positive-sense single-stranded RNA ((+)ssRNA) genome into the cytoplasm (step 4). The viral (+)ssRNA is translated into a single polyprotein at the ER and cleaved into mature proteins by the viral serine protease non-structural protein 2B-3B (NS2B-NS3) and cellular proteases (step 5). The NS proteins, including the viral RNA-dependent RNA polymerase NS5, form the replication complex for the synthesis of full-length negative-sense ssRNA ((-)ssRNA) intermediates (step 6). These serve as templates for the synthesis of full-length (+)ssRNAs (step 7). The viral capsid protein, C, is responsible for encapsidating viral genomic RNA, with assembly occurring on rough ER membranes (step 8). Immature virions are transported through the host secretory pathway, resulting in glycosylation of the viral E protein and host cell furin mediated-cleavage of the protein prM to the mature membrane protein, M (step 9). Mature virions are transported to the plasma membrane and released by exocytosis (step 10).” (Suthar et al. 2013)

10 2.4 Transmission
2.4 Transmission
2.4.1 Enzootic Cycle
In nature, the flaviviruses WNV and USUV circulate between predominantly ornithophilic mosquito species as vectors and a wide variety of bird species of the local avifauna as amplifying/reservoir hosts (Figure 2.3) (Brault 2009). The two viruses share many different vector and host species. For example, more than 34 avian species of 11 different orders are susceptible to both WNV and USUV. Most affected by both viruses are species belonging to the Family Corvidae of the order Passeriformes, such as the in Europe indigenous Eurasian jay (Garrulus gladarius), common starling (Sturnus vulgaris), and Eurasian magpie (Pica pica) (Nikolay 2015, Zannoli and Sambri 2019). However, WNV alone is chiefly associated with infections in crows, especially the American crow (Corvus brachyrhynchos) and the fish crow (Corvus ossifragus) in North America (Komar et al. 2003) and USUV is connected to large die-offs in Eurasian blackbird populations in Europe (Lühken et al. 2017, Michel et al. 2019). Both species also regularly infect birds of prey, including raptors (Falconiformes) and owls (Strigiformes) (Gancz et al. 2004, Höfle et al. 2008, Jiménez-Clavero et al. 2008, Michel et al. 2019, Nemeth et al. 2006a, Steele et al. 2000). Migratory birds, such as the white stork (Ciconia ciconia) (Malkinson et al. 2002), can act as long-distance dispersal vehicles (Owen et al. 2006).
In the case of WNV, non-vector-borne transmissions between birds have been confirmed in multiple bird species, such as in-contact transmission (oral-fecal, allopreening, or feather picking) in the common goose (Anser anser domesticus) (Banet-Noach et al. 2003), chicken (Gallus gallus domesticus) (Langevin et al. 2001), ring-billed gull (Larus delawarensis), blue jay (Cyanociatta cristata), black-billed magpie (Pica hudsonia), and the American crow
Figure 2.3 Transmission cycle of the flavivirus West Nile virus.

2 Literature Review 11
(Komar et al. 2003). The common grackle (Quiscalus quiscula), house sparrow (Passer domesticus), and the American crow were infected through the ingestion of WNV in aqueous solutions (Komar et al. 2003). Five bird species were experimentally infected through the ingestion of WNV-infected mice: the great horned owl (Bubo virginianus) (Komar et al. 2003, Nemeth et al. 2006a), Eastern screech owl (Megascops asio) (Nemeth et al. 2006b), black-billed magpie, the American crow (Komar et al. 2003, Marra et al. 2004), and the American kestrel (Falco sparverius) (Nemeth et al. 2006a). American crows became infected after ingesting infected house sparrow carcasses and house finches (Haemorhous mexicanus) after ingesting infectious mosquitoes (Komar et al. 2003). It is, therefore, a justifiable hypothesis that carcasses of highly susceptible bird species such as Corvidae constitute a source of contamination for birds of prey (Reisen 2013). The conceivable direct transmission of WNV places nestlings, one-year-hatch-birds, social species, and flocking social birds especially at risk (Marra et al. 2004). A similar scenario for USUV could not be confirmed yet. In two studies with chickens (Chvala et al. 2005) and geese (Chvala et al. 2006) direct transmission could not be demonstrated.
In the case of WNV and USUV the viruses can be transmitted by multiple mosquito species, with the genera Culex being the most essential. For example, more than 60 species have been described as potential vectors for WNV in the USA (Hayes et al. 2005). Culex pipiens pipiens was important for the avian amplification cycle in the northeast, Cx. pipiens quinquefasciatus was essential in the southeast, and Cx. tarsalis in the west of the USA (Andreadis 2012). In Europe, numerous species, including Cx. pipiens sensu lato (s.l.), Cx. torrentium, Cx. modestus, Ae. albopictus, Ae. detritus, and Ae. japonicus were experimentally proven to be susceptible to WNV (summarized by Vogels et al. 2017c). European vector competence studies concluded that Cx. pipiens is the main WNV vector as the species is not only highly abundant in Europe during the summer but is also associated with very high transmission rates (Engler et al. 2013, Vogels et al. 2017c). The species is especially interesting due to its two biotypes (pipiens and molestus). These possess different feeding preferences (ornithophilic vs. mammalophilic) (Osório et al. 2014), making them essential not only for the natural transmission cycle but also for spillover events into the human population (Rudolf et al. 2013, Zannoli and Sambri 2019). Both biotypes can co-exist, but also readily hybridize with the described synergistic effect on virus transmission (Ciota and Kramer 2013). Vertical passage (transoverial transmission) of WNV from infected females to hatched larvae has been confirmed in overwintering cohorts (Nelms et al. 2013).
USUV has also been isolated from a diverse panel of mosquito species, both in Africa (e.g., Senegal, Kenya, and Uganda) and in Europe (e.g., Italy, Austria, and Germany). Equally to WNV, USUV is most commonly found in species of the Culex genus, such as Cx. modestus, Cx. neavei, Cx. perexiguus, Cx. perfuscus, Cx. pipiens, Cx. quinquefasciatus, and Cx. univittatus (summarized by Clé et al. 2019). The in Europe invasive mosquito species Ae. albopictus (Calzolari et al. 2010, Mancini et al. 2017, Tamba et al. 2011) and Ae. japonicus (Camp et al. 2019) have also been associated with USUV. The importance of Culex species

12 2.4 Transmission
(Cx. pipiens, Cx. neavei and Cx. quinquefasciatus) has been established in vector competence experiments (Cook et al. 2018, Fros et al. 2015, Nikolay et al. 2012). Interestingly, the transmission efficiency of Dutch Cx. pipiens appeared to be higher for USUV than for WNV after incubation at elevated temperature regimes (28 °C) (Fros et al. 2015).
2.4.2 Epidemic/Epizootic Cycle
When a mosquito species feeds indiscriminately on both avian and mammalian species it can function as a bridge vector and transmit WNV and USUV from birds to vertebrates such as equids and humans (Kramer et al. 2008). These vertebrates are considered incidental or dead-end hosts as they cannot perpetuate the transmission cycle i.e., their viremia does not suffice to infect feeding mosquitoes (Chancey et al. 2015). However, epizootic and/or epidemic outbreaks can occur with the mass repetition of such spillover events (Fenton and Pedersen 2005, Lloyd-Smith et al. 2009). For WNV, unlike USUV, human-to-human infections have been described through blood transfusion, organ transplantation, breast-feeding, intrauterine exposure, and laboratory-acquired infections (Hayes and O'Leary 2004, Iwamoto et al. 2003, Kusne and Smilack 2005, Lindsey et al. 2010, Rios et al. 2006).
WNV infections have also been described in at least 100 additional free-ranging and captive mammalian species as well as amphibians and reptiles (Ariel 2011, Klenk and Komar 2003, Root and Bosco-Lauth 2019). Exposed mammals comprise predominantly artiodactyls (cloven-hooved mammals), carnivores and mesocarnivores, rodents, and non-human primates (good summary by Root and Bosco-Lauth 2019 and by van der Meulen et al. 2005). Species included various deer species, wild boars (Sus scrofa), sheep (Ovis domesticus), Rocky Mountain goats (Oreamnos americanus), black howlers (Aloutta caraya), mountain gorillas (Gorilla beringei beringei), rhesus macaques (Macaca mulatta), common marmosets (Callithrix jacchus), Virginia opossums (Didelphis virginiana), raccoons (Procyon lotor), striped skunks (Mephitis mephitis), captive and wild bear species (Ursus spp.), red foxes (Vulpes vulpes), stone martens (Martes foina), gray wolves (Canis lupus), domestic cats (Felis domesticus), and dogs (Canis familiaris), various squirrel species (Sciurus spp.), groundhogs (Marmota monax), commensal rats and mice, bats (e.g., Eptesicus fuscus), and elephants (Elephas maximus) (Root and Bosco-Lauth 2019, van der Meulen et al. 2005). Amphibians, such as the marsh frog (Pelophylax ridibundus) or bullfrog (Rana catesbeiana) and reptiles such as the garter snake (Thamnophis sirtalis), red-eared slider (Trachemys scripta), and green iguana (Iguana iguana) are even suggested to be WNV amplifiers (Klenk and Komar 2003). A WNV outbreak with a high mortality rate was described in an American alligator (Alligator mississippiensis) farm (Klenk et al. 2004, Miller et al. 2003).
Information about USUV circulation in vertebrate species besides humans and equines is scarce (Saiz and Blázquez 2017). USUV-specific RNA has been isolated from the brain of bats (Pipistrellus pipistrellus) in southwest Germany (Cadar et al. 2014) and from rodent and shrew species from Senegal (Diagne et al. 2019). USUV-specific neutralizing antibodies have been

2 Literature Review 13
detected in dogs (Durand et al. 2016, Montagnaro et al. 2019), wild boars (Bournez et al. 2019, Escribano-Romero et al. 2015), and wild ruminants (García-Bocanegra et al. 2016). Experimental studies suggest that newborn/weaning mice are sensitive to USUV (Weissenböck et al. 2004). Yet, unlike WNV (Morrey et al. 2008), USUV only causes neurologic lesions (neuronal apoptosis and demyelination) in less than one-week-old mice and does not induce a fatal illness (Weissenböck et al. 2004). Horses represent dead-end hosts for USUV and USUV-specific antibodies have been recorded in multiple European and African countries (Croatia (Barbic et al. 2013), Italy (Savini et al. 2011), Morocco (Durand et al. 2016), Serbia (Lupulovic et al. 2011) Spain (Vanhomwegen et al. 2017), Poland (Bażanów et al. 2018), and Tunisia (Ben Hassine et al. 2014)).
2.4.3 Infection Pattern in a Competent Mosquito Vector
A mosquito acquires the flaviviruses WNV or USUV by feeding on a viremic host. The life-cycle of flaviviruses in a mosquito has been well described (Franz et al. 2015, Vogels et al. 2017c). With the ingestion of the blood meal the virus travels through the foregut and cardia and enters the midgut (Okuda et al. 2002). There it enters the midgut epithelial monolayer (primarily the posterior portion of the midgut (Scholle et al. 2004)) via cellular membrane-associated receptors (Cheng et al. 2010) and initiates replication. The virus must subsequently disseminate to the hemocoel, the blood circuit of arthropods, in order to reach the salivary glands. For this the virus can follow several alternate routes. It can pass through the basal lamina (extracellular matrix) by budding from epithelial midgut cells (with or without virus replication in these cells) (Girard et al. 2005), via the passive passage through a transiently “leaky” midgut (Houk et al. 1979), or via the additional infection of the midgut trachea (Romoser et al. 2004). By means of the hemolymph the virus then disseminates to secondary tissues, including the fat body, hemocytes, nerve tissues, and muscles, until it finally reaches the paired salivary glands (Girard et al. 2004). Once more the virus must overcome the basal lamina before infecting the salivary gland epithelial monolayer (da Cunha Sais et al. 2003). Finally, the virus is released into the salivary gland lumen either via budding or apoptosis, thereby, increasing present viral titers in the saliva (Clem 2016, Hardy et al. 1983). The mosquito is now capable of transmitting the virus to another vertebrate host during feeding (infection pattern in vectors and hosts summarized by Franz et al. 2015, Vogels et al. 2017c and illustrated in Figure 2.4).
Hence, to fulfill the life cycle and ensure that a virus-exposed mosquito can become infectious, a virus must overcome three barriers within the mosquito body: the peritrophic membrane, the midgut barrier, and the salivary gland barrier (Vogels et al. 2017c). The peritrophic membrane is a chitinous sac comprised of proteins, glycoproteins, and chitin microfibrils. It forms within a few hours after uptake of a blood meal and separates the blood meal from the midgut epithelium (Lehane 1997). In some mosquito species its thickness can act as a mechanical barrier for certain viruses, yet it does not appear to pose a strong barrier for flaviviruses according to the literature (Kato et al. 2008, Whitfield et al. 1973). The midgut and salivary

14 2.4 Transmission
gland barriers are each split into an infection and an escape barrier. The midgut/salivary gland infection barriers are based on receptor recognition to enable successful virus endocytosis into the midgut or salivary gland epithelial cells. The midgut escape barrier comprises the basal lamina, with its size exclusion limit below the size of certain virions, and the salivary gland escape barrier refers to virus-triggered cell apoptosis. The latter can be required for virus release but can also have detrimental effects not only on the virus but also on its vector (Franz et al. 2015, Kenney and Brault 2014, Vogels et al. 2017c). These barriers can function mechanically, but the mosquitoes have also evolved several antiviral responses combating viral replication, such as RNA regulatory pathways (RNA interference (RNAi)), antiviral signaling cascades (Janus Kinase (JAK)/Signal Transducer and Activator of Transcription (STAT), Toll, immune deficiency (IMD)), and cellular processes of autophagy and apoptosis (Prasad et al. 2013). The effectiveness of the mechanical barriers and the antiviral immune response against certain flaviviruses is mosquito species dependent (Vogels et al. 2017c). In addition, the mosquito gut microbiome, consisting of a variety of bacteria and fungi, can interact with a virus (Jupatanakul et al. 2014). Also intracellular bacteria such as Wolbachia spp. (Johnson 2015) and co-infections with insect-specific viruses (Salas-Benito and De Nova-Ocampo 2015) or other arboviruses can influence the ability of a vector to replicate and transmit a virus.
Figure 2.4 Arbovirus migratory route in vector and host.
2 Literature Review 15
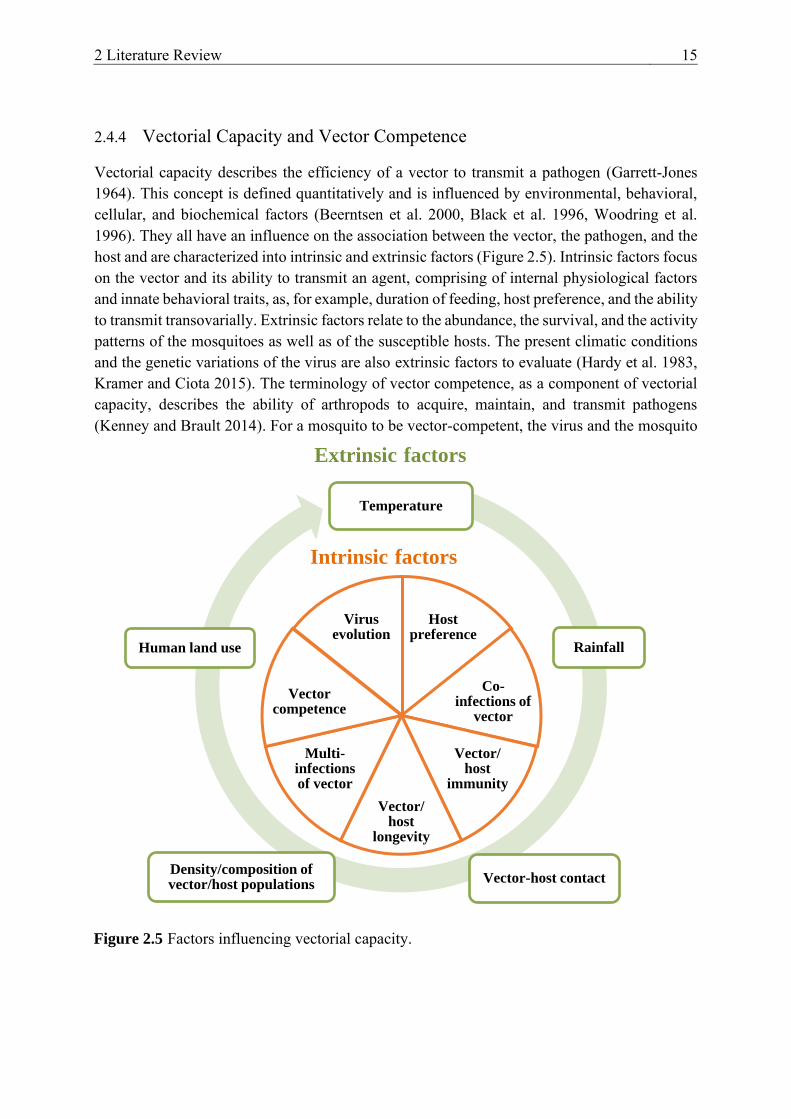
2.4.4 Vectorial Capacity and Vector Competence
Vectorial capacity describes the efficiency of a vector to transmit a pathogen (Garrett-Jones 1964). This concept is defined quantitatively and is influenced by environmental, behavioral, cellular, and biochemical factors (Beerntsen et al. 2000, Black et al. 1996, Woodring et al. 1996). They all have an influence on the association between the vector, the pathogen, and the host and are characterized into intrinsic and extrinsic factors (Figure 2.5). Intrinsic factors focus on the vector and its ability to transmit an agent, comprising of internal physiological factors and innate behavioral traits, as, for example, duration of feeding, host preference, and the ability to transmit transovarially. Extrinsic factors relate to the abundance, the survival, and the activity patterns of the mosquitoes as well as of the susceptible hosts. The present climatic conditions and the genetic variations of the virus are also extrinsic factors to evaluate (Hardy et al. 1983, Kramer and Ciota 2015). The terminology of vector competence, as a component of vectorial capacity, describes the ability of arthropods to acquire, maintain, and transmit pathogens (Kenney and Brault 2014). For a mosquito to be vector-competent, the virus and the mosquito
Figure 2.5 Factors influencing vectorial capacity.
Temperature
Rainfall
Vector-host contactDensity/composition of vector/host populations
Human land use
Extrinsic factors
Host preference
Co-infections of
vector
Vector/ host
immunity
Vector/ host
longevity
Multi-infections of vector
Vector competence
Virus evolution
Intrinsic factors

16 2.4 Transmission
species must be “compatible”, where the mosquito-related and virus-dependent barriers do not present a significant hindrance (Beerntsen et al. 2000, Vogels et al. 2017c).
2.4.5 Infection Pattern in a Susceptible Avian Host
In a susceptible host the pathogenesis of flaviviruses proceeds in three key phases: 1. local infection followed by primary viremia, 2. spread of the virus with replication in peripheral organs, and 3. neuroinvasion and neurovirulence (Figure 2.4) (King et al. 2007). The mechanism for WNV pathogenesis has been analyzed in detail and is very similar to that of USUV. After mosquito blood-feeding, the virus replicates in the keratinocytes, skin residential dendritic cells, and Langerhans cells of the epidermis. Via activated Langerhans cells it is then transported to the draining lymph nodes and from there it disseminates to visceral organs, such as the kidneys, spleen, liver, and heart (Gamino and Höfle 2013, Samuel and Diamond 2006). The exact mechanism how WNV crosses the blood-brain-barrier and enters the central nervous system (CNS) is not yet known, hematogenous and transneural pathways have been proposed (summarized by Bai et al. 2019, King et al. 2007). In mammals a tumor necrosis factor-alpha (TNF-α) and an Interleukin-1beta (IL-1β) mediated change in endothelial cell permeability may facilitate virus entry (Wang et al. 2004) aside from the infection or passive transport through endothelium or epithelial cells of the choroid plexus (McMinn 1997). The transport via infected immune cells (Garcia-Tapia et al. 2006) or directly via axonal retrograde transport from the olfactory or motor neurons is also possible (Monath et al. 1983, Samuel et al. 2007).
Pathogenesis of WNV and USUV in birds is virus and host specific. Numerous infection experiments give insight into the pathogenesis of WNV in free-ranging and captive avian species (summarized by Gamino and Höfle 2013). Experimentally infected birds usually develop viremia within 24 or 48 h (the latter being the case in less susceptible species such as chickens). Peak viremia is reached 2-3 days post infection (dpi) in Passeriformes (crows and jays) (Weingartl et al. 2004) and 4-6 dpi in Falconiformes (raptors) (Ziegler et al. 2013) and Strigiformes (owls) (Nemeth et al. 2006b). Viremia is the highest in birds that eventually succumb to the infection (Langevin et al. 2005). Virus load in the blood gradually declines with the last detection 6-7 dpi (Passeriformes, owls, and geese) or 10 dpi (raptors) (Banet-Noach et al. 2003, Lapointe et al. 2009, Nemeth et al. 2006b, Swayne et al. 2000, Ziegler et al. 2013). Disease development is associated with WNV replication in a wide variety of cell types and invasion of major organs such as the liver, spleen, kidney, heart and CNS (Gamino and Höfle 2013, Steele et al. 2000). For example, the virus can already be detected 1 dpi in the spleen of infected crows, reaching the highest viral titers 4 dpi. In Passeriformes (e.g., house finch and scrub-jay) WNV can persist up to six months in certain organs, like the spleen, kidney, eye, brain, or skin (Komar et al. 2003, Wheeler et al. 2012). The host immune response consists of an innate immune response involving 2´-5´-oligoadenylate synthase pathway, interferons, inflammatory cytokines, complement factors, and natural killer cells and the adaptive immune response with B- and T-cell activation and the production of neutralizing antibodies against the viral E glycoprotein and NS proteins (summarized by Bai et al. 2019, King et al. 2007). In most

2 Literature Review 17
species, seroconversion can be initially detected 4-6 dpi (Gamino and Höfle 2013). In certain species neutralizing antibodies can persist up to one year and maternal antibodies can provide protection to offspring (Gibbs et al. 2005, Hahn et al. 2006, Nemeth et al. 2009b, Wilcox et al. 2007).
It is probable that the course of an USUV infection in highly susceptible bird species (e.g., blackbirds and magpies) is comparable to that of WNV. Experimentally infected domestic canaries (Serinus canaria) confirmed that USUV also triggers early onset of viremia followed by the rapid invasion of vital organs, including brain, heart, liver, spleen, skin, and kidneys (Benzarti et al. 2020). A similar systemic infection was observed in naturally infected birds (Benzarti et al. 2019, Chvala et al. 2004). As was suggested for WNV (Gamino and Höfle 2013), the wide tissue tropism of USUV can be a result of the virus targeting immune cells, such as macrophages and dendritic cells (Chvala et al. 2004). In the canaries, USUV-specific neutralizing antibodies were found two weeks after infection in combination with a humeral immune response (Benzarti et al. 2020). Even though in-contact transmission has not been verified experimentally for USUV, oral-fecal virus transmission or via feather picking is conceivable. Viral shedding (oropharyngeal or cloacal) was described in experimentally infected domestic chickens (Chvala et al. 2005), geese (Chvala et al. 2006), and canaries (Benzarti et al. 2020) and in naturally infected song thrushes (Turdus philomelos) (Höfle et al. 2013). Furthermore, USUV-specific RNA was isolated from the immature feathers of infected canaries (Benzarti et al. 2020). It has been speculated for USUV as well as for WNV that ectoparasites can increase virus susceptibility in birds or possibly even serve as additional vectors. For example, an infestation with louse flies (Icosta americana) was often observed in conjunction with virus detections in captive owls (Gancz et al. 2004, Meister et al. 2008, Ziegler et al. 2016).
2.5 Clinical Presentation and Pathology
2.5.1 West Nile Virus
After infection with WNV most birds remain asymptomatic due to a rapid innate and adaptive immune response with virus elimination. Yet some birds may develop clinical signs, generally appearing 5 dpi, ranging from unspecific signs such as lethargy, anorexia, dehydration, and ruffled feathers to neurological signs including ataxia, abnormal head posture or head movements, tremors, disorientation, paresis, paralysis, and blindness (summarized by Gamino and Höfle 2013). Long-lived birds such as raptors or owls can even undergo relapses of neurological signs up to four years after an infection (Nemeth et al. 2006a, Nemeth et al. 2009a). As a result of the widespread cellular and tissue tropism of WNV there are no pathognomonic macroscopic lesions (Gamino and Höfle 2013). In birds that survive longer after an infection, chronic lesions can be detected, such as emaciation, dehydration, multiorgan hemorrhages, petechiae, and congestion (Lopes et al. 2007, Nemeth et al. 2006a, Steele et al. 2000, Wünschmann et al. 2005). Microscopic lesions can be a direct effect of the virus or due to the

18 2.5 Clinical Presentation and Pathology
host immune response (virus induced autophagy, apoptosis, inflammation, and necrosis). Pathological changes include necrotizing hepatitis, splenitis, myocardial degeneration, myocarditis, necrosis of striated muscles, non-suppurative encephalitis, and neuronal necrosis (summarized by Gamino and Höfle 2013).
For the most part, equines seroconvert without a clinical manifestation. However, approximately 8% of the infected horses develop severe neurological signs: weakness of the limbs, recumbency, muscle fasciculation, and ataxia. Less frequent signs include fever, cranial nerve deficits, hyperesthesia, teeth grinding, muscular tremor, photophobia, and blindness (summarized by Angenvoort et al. 2013). Behavioral shifts into a non-responsive, somnolent, disorientated, hyperexciteable, or aggressive state have also been documented (Abutarbush et al. 2004, García-Bocanegra et al. 2016, Long 2006, Ostlund et al. 2001, Porter et al. 2003). With the appearance of clinical signs the case fatality rate increases from 22% to 44% (García-Bocanegra et al. 2016, Murgue et al. 2001a, Ostlund et al. 2001, Schuler et al. 2004). Clinical signs can sustain in mild cases for two to seven days but recovery can take up to several months in severe cases (Trock et al. 2001, Venter and Swanepoel 2010). Horses can suffer under disease sequelae with weight loss, lethargy, ataxia, or cranial nerve deficits (Salazar et al. 2004). Unlike in birds, WNV exhibits a pronounced CNS tropism in horses. A mild to severe polioencephalomyelitis, primarily in the lower brain stem and ventral horns of the thoracolumbar spinal cord, is the most common cause for microscopic lesions (Cantile et al. 2001).
Similar to equines, the majority (> 80%) of human WNV infections are asymptomatic (Drebot and Artsob 2005, Petersen et al. 2003). Symptomatic infections, with an incubation period of approximately 2-15 days (Mostashari et al. 2001, Petersen and Marfin 2002), primarily represent themselves as mild, self-limiting, febrile illnesses known as West Nile fever (WNF) (Drebot and Artsob 2005, Petersen et al. 2003). Symptoms include headache, myalgia, nausea, vomiting, chills, and a roseolar or macropapular rash (Brilla et al. 2004, Hayes et al. 2005, Madden 2003, Petersen and Marfin 2002, Petersen et al. 2002, Tyler 2004). Of the symptomatic infections 5% develop a life-threatening neurological infection, known as West Nile neuroinvasive disease (WNND), with meningitis, encephalitis, or a poliomyelitis-like disease (acute flaccid paralysis) (Campbell et al. 2002). Pathophysiological findings can be traced back to virus-associated neuronal proliferation, cytotoxic immune responses, perivascular inflammation, and microglial nodule formation (Deubel et al. 2001, Sampson et al. 2000, Shieh et al. 2000). Primarily affected is the brain stem, yet the thalamus, cerebellum, and cerebral cortex can also be involved (Sampson et al. 2000). Patients can develop symptoms such as headache, photophobia, back pain, confusion, persistent fever, asymmetric weakness, absent reflexes, and tremors (Rossi et al. 2010). Long-term complications, such as fatigue, weakness, myalgia, arthralgia, headaches, and neurological complications, are common (Sejvar 2007). Of patients with WNND the case fatality rate is commonly under 1%, can, however, vary between

2 Literature Review 19
outbreaks (e.g., 4% in Romania (Tsai et al. 1998) and up to 17% in Greece (Danis et al. 2011b, Pervanidou et al. 2014)).
2.5.2 Usutu Virus
Regularly reported neurological signs in birds infected with USUV consist of prostration, disorientation, ataxia, and weight loss. Common macroscopic lesions are hepatomegaly and splenomegaly. Death is frequently attributed to multi-systemic organ failure with severe inflammation and necrosis, predominantly in the heart, liver, kidneys, spleen, and brains. In addition, glia nodules and neuronophagia can be observed in the brain (Bakonyi et al. 2007, Chvala et al. 2004, Clé et al. 2019). By contrast, horses are not highly susceptible to USUV with infections being asymptomatic. Nevertheless, they can be a good sentinel species due to high seroprevalence in certain countries, e.g., in Italy (89.2% in 2008 (Savini et al. 2011)) and in Poland (28.0% in 2012-2013 (Bażanów et al. 2018)). In other European countries USUV-specific neutralizing antibodies have also been observed, for example in Serbia (Lupulovic et al. 2011), Croatia (Barbic et al. 2013), and Spain (Vanhomwegen et al. 2017).
Human infections due to USUV appear to be scarce, and as with WNV, are commonly asymptomatic. Individual cases have been reported in Africa and Europe. In Africa the virus has only been isolated twice: in 1981 from a patient in the Central African Republic and in 2004 from a child in Burkina Faso. Both exhibited only mild clinical symptoms including fever, jaundice, and rashes (Nikolay et al. 2011). In 2009, the first human USUV infections were reported also in Europe, involving two immunocompromised patients in Italy diagnosed with meningoencephalitis (Cavrini et al. 2009, Pecorari et al. 2009). Retrospective studies in Italy enabled the confirmation of further USUV infections in patients with meningoencephalitis (Cavrini et al. 2011, Grottola et al. 2017). Outside of Italy, neuroinvasive USUV infections have been described in Croatia in 2013 and 2018 (Santini et al. 2015, Vilibic-Cavlek et al. 2019). The most frequent symptoms in all patients included headache, fever, nuchal rigidity, hand tremor, and hyperflexia (Saiz and Blázquez 2017). Individual cases of acute infections have also been described in Austria (Aberle et al. 2018, Bakonyi et al. 2017) and Germany (Cadar et al. 2017b) in asymptomatic blood donors and in France in a patient with the atypical presentation of “a frigore” facial paralysis (Simonin et al. 2018). Seroconversion was verified in all patients. Furthermore, screening of blood donor serum in Germany, Italy, and Serbia showed a low seroprevalence of USUV-specific antibodies of 0.02% to 1.1% (Allering et al. 2012, Cvjetković et al. 2016, Gaibani et al. 2012, Grottola et al. 2017, Pierro et al. 2013). These studies illustrate the non-negligible exposure risk to USUV that the human population is currently facing. It is suggested that, even though severe infections are rare, USUV circulates more actively than WNV in Europe and that the probability of human infections increases with an upsurge in USUV circulation in avian reservoir or vector species (Clé et al. 2019).

20 2.6 Detection
2.6 Detection
Acute flavivirus infections with WNV or USUV can be diagnosed by virus isolation in cell culture (mosquito or mammalian cell lines), nucleic acid amplification via a quantitative (real-time) reverse transcription PCR (RT-qPCR), or the identification of virus-specific immunoglobulin M (IgM) antibodies via multiple serological tests for horses and humans. To detect antibodies, an enzyme-linked immunosorbent assay (ELISA), immunofluorescence (IF), hemagglutination inhibition assay (HIA), or virus neutralization test (VNT) or plaque reduction neutralization test (PRNT) can be performed, preferably using serum or cerebral spinal fluid (Clé et al. 2019). For WNV, unlike USUV, commercial ELISAs are available (Malan et al. 2004). Virus neutralization tests (VNT/PRNT), however, are the gold standard for confirming WNV/USUV-specific antibodies as only these tests can determine pathogen-specific antibodies and rule out cross-reactive antibodies induced, for example, by Japanese encephalitis virus (JEV), tick-borne encephalitis virus (TBEV), yellow fever virus (YFV), dengue virus (DENV), or Zika virus (ZIKV) (Pauli et al. 2014, Rathore and St. John 2020).
2.7 Prevention and Treatment
No therapeutic treatments are available for WNV infections in humans and animals. In humans, treatment options are mainly supportive care measures and intensive rehabilitation (Kramer et al. 2007). Immune prophylaxis is also not possible as there are no vaccines approved by the Food and Drug Administration of the United States of America. Even though promising candidates have been evaluated (De Filette et al. 2012), the development process was stopped as a universal WNV vaccine was considered unlikely to be cost-effective due to market uncertainty (i.e., relatively low virus incidence) (Kaiser 2012). However, four vaccines, licensed by the United States Department of Agriculture (USDA), are available for equines: two consist of inactivated whole WNV (WN-InnovatorTM; Zoetis, Parsippany, NJ, USA (Hankins 2010) and VeteraTM WNV; Boehringer Ingelheim Vetmedica, Duluth, GA, USA), one is a recombinant live canary pox virus co-expressing WNV prM and E proteins (RecombitekTM Equine WNV; Merial, Athens, GA, USA (El Garch et al. 2008)), and one is an inactivated flavivirus chimeric vaccine (EquiNileTM; Merck Animal Health, Omaha, NE, USA) (Angenvoort et al. 2013, Kaiser and Barrett 2019, Ng et al. 2003). As the antibody response of animals commonly targets the E-protein, all vaccine approaches aim to best deliver this protein. These E-protein based WNV vaccines are protective against both WNV lineages, 1 and 2, (Ulbert and Magnusson 2014). They have been widely used throughout the USA and three have also recently been employed in Europe. In order to be protective, all four vaccines require two shots within a few weeks for the primary immunization and a yearly booster vaccination (Angenvoort et al. 2013). There are currently no licensed drugs or vaccines to fight USUV infections yet a cross-reactive vaccine for flaviviruses remains a possible option, especially for valuable birds (de Oya et al. 2019, Lobigs and Diamond 2012). For example, mice inoculated with a WNV vaccine (recombinant subviral particle) showed higher antibody levels upon

2 Literature Review 21
infection with USUV (Merino-Ramos et al. 2014). A further point of attack could be preventing mosquito bites (mosquito control programs, use of insect repellents and mosquito nets, elimination of mosquito breeding sites, and minimization of outdoor activities during mosquito peak-activity) (Saiz and Blázquez 2017).


3 Materials and Methods 23
3 MATERIALS AND METHODS
The following section summarizes the most frequently used diagnostic tests and materials. Detailed information to an individual procedure can be found in the corresponding manuscripts I-IV.
3.1 Mosquito Infection
Mosquito infection experiments (manuscripts I-IV) were performed with field-collected, as well as, laboratory-reared colonized mosquitoes. Focus was placed on in Germany indigenous mosquito species (e.g., Cx. pipiens and Cx. torrentium). Yet for comparison purposes, mosquito populations from other countries, such as Cx. pipiens biotype molestus from the Republic of Serbia or Ae. aegypti originally from Malaysia were also tested. Invasive species in Europe (e.g., Ae. albopictus) were also used as they are highly interesting, especially for the transmission of WNV. Field-collected mosquito populations were obtained by collecting egg-rafts, identifying the hatching larvae to species and biotype level, and gathering pupated adults into cages. For the infection of mosquitoes via a blood meal (manuscripts I and II) female mosquitoes were sorted into feeding chambers and allowed to feed on an infectious blood meal (i.e., spiked with WNV or USUV) either using the Hemotek PS5 Feeder (Hemotek Ltd, Lancashire, the United Kingdom (UK)) or by offering them two cotton stick ends soaked with the infectious blood meal. For the infection of mosquitoes via intrathoracic injections (manuscripts III-IV), female mosquitoes were injected with WNV into the thorax under carbon dioxide (CO2) anesthesia. After feeding or injection, engorged and/or surviving females were sorted into incubation chambers and kept for up to three weeks under fixed environmental conditions: temperature (18 °C, 25 °C, or 28 °C ± 1 °C), relative humidity (80%-85%), and 16 h light/ 8 h dark photocycle. At set time points saliva assays (for detailed information see manuscripts I-IV) were performed. The obtained saliva samples were then inoculated onto Vero cells to check if they contained viable virus particles. Mosquito bodies (thorax and abdomen), extremities (legs plus wings), and cell-culture supernatants of infectious saliva samples were examined in a specific RT-qPCR (Eiden et al. 2010, Jöst et al. 2011). To verify that all the mosquitoes used in manuscript IV to infect the domestic poultry could transmit virus in their saliva, a saliva assay was performed in addition to head-squashes that were analyzed via RT-qPCR.
3.2 Vector Competence Indices
Various indices are used and defined here (Table 3.1) to assess the vector competence of a mosquito species (manuscripts I-IV). The feeding rate refers to the number of engorged females out of the total number of females exposed to the infectious blood meal. The survival rate describes the number of females surviving a given period of time out of the total number of fed females. The infection rate is defined as the number of WNV-positive bodies in relation to the total number of mosquitoes examined. The dissemination rate is calculated as the number of

24 3.3 Animal Trials
specimens with WNV-positive extremities (legs plus wings) out of the total number of WNV-positive bodies. The transmission rate is the percentage of mosquitoes with infected bodies and legs plus wings that also had viable virus in their saliva. Transmission efficiency is the percentage of mosquitoes having viable virus in their saliva in relation to the total number of mosquitoes analyzed.
Table 3.1 Definition of vector competence indices.
Infection Rate
(IR) = mosquitoes with positive bodies/analyzed mosquitoes
Dissemination Rate
(DR) = mosquitoes with positive legs plus wings/mosquitoes with positive bodies
Transmission Rate
(TR) = mosquitoes with positive saliva/mosquitoes with positive legs plus wings
Transmission Efficiency
(TE) = mosquitoes with positive saliva/analyzed mosquitoes
3.3 Animal Trials
Domestic chickens, ducks, and geese were obtained from local commercial breeders for the animal infection studies (manuscript IV). All three species were injected subcutaneously into the knee fold with a WNV lineage 1 strain from Italy. Over a period of three weeks the health status of the birds was examined daily according to a pre-defined score sheet and the birds were sampled according to a fixed sampling schedule. Whole blood was collected either from the basilica vein (Vena cutenea ulnaris superficialis), the caudal tibial vein (V. metatarsalis plantaris superficialis), or after euthanasia from the jugular vein (V. jugularis). Serum samples were analyzed in a virus neutralization test (VNT) on Vero cells, based on the same virus strain used to infect the birds and with a commercial ID Screen® WN competition ELISA (IDVet, Grabels, France). RNA was extracted from the blood clots and was tested with a WNV-specific RT-qPCR (Eiden et al. 2010). Viral shedding was analyzed via RNA extraction and RT-qPCR of oropharyngeal and cloacal swabs.
During necropsy, a panel of various tissues was removed for the RT-qPCR and for the pathological examination (histopathology and immunohistochemistry (IHC)). Tissue samples were fixed in 4% neutral buffered formalin, embedded in paraffin, sectioned at 3 µm, stained with hematoxylin/eosin, and examined by a light microscope. An additional IHC was performed with samples that had been tested WNV-positive in the RT-qPCR. For this, a peroxidase-based polymer system (EnVisionTM; Dako Diagnostics, Hamburg, Germany) was used for the demonstration of WNV antigen and the in-house OM8, diluted in goat serum, as the primary polyclonal antibody. Staining/counterstaining was performed with diaminobenzidine-tetrahydrochloride and Mayer`s hematoxylin. For a more detailed description of the methods see manuscript IV.

3 Materials and Methods 25
3.4 RNA Extraction and RT-qPCR
For both the mosquito and animal infection experiments RNA was extracted either per hand with the RNeasy Mini Kit (Qiagen, Hilden, Germany) or with an extraction robot, BioSprint 96 (Qiagen) or KingFisher Flex Purification System (ThermoFischer, Darmstadt, Germany) using the NucleoMag VET Kit (Macherey-Nagel, Düren, Germany) and following manufacturer’s instructions. All RNA extracts were examined for WNV (manuscript II-IV) using a WNV-specific RT-qPCR assay targeting the 5' untranslated region for the simultaneous detection of lineage 1 and 2 strains (Eiden et al. 2010). For USUV (manuscript I), the USUV-specific RT-qPCR assay described by Jöst et al. 2011. A WNV and USUV calibration curve, established using synthetic RNA, was run in parallel for means of RNA-quantification.


4 Manuscript I 27
4 MANUSCRIPT I
German Culex pipiens Biotype molestus and Culex torrentium are Vector-
Competent for Usutu Virus
Cora M. Holicki 1, Dorothee E. Scheuch 1, Ute Ziegler 1, Julia Lettow 2,3, Helge Kampen 2,
Doreen Werner 4, and Martin H. Groschup 1,*
1 Institute of Novel and Emerging Infectious Diseases, Friedrich-Loeffler-Institut, Federal Research Institute for Animal Health, 17493 Greifswald-Insel Riems, Germany
2 Institute of Infectology, Friedrich-Loeffler-Institut, Federal Research Institute for Animal Health, 17493 Greifswald-Insel Riems, Germany
3 Present address: Department of Molecular Genetics and Infection Biology, University of Greifswald, Greifswald, Germany
4 Biodiversity of Aquatic and Semiaquatic Landscape Features, Leibniz-Centre for Agricultural Landscape Research, 15374 Muencheberg, Germany
Author to whom correspondence should be addressed: [email protected]
This manuscript has been submitted and peer-reviewed.

28 4.1 Abstract
4.1 Abstract
Background: Usutu virus (USUV) is a rapidly spreading zoonotic arbovirus (arthropod-borne virus) and a considerable threat to the global avifauna and in isolated cases to human health. It is maintained in an enzootic cycle involving ornithophilic mosquitoes as vectors and birds as reservoir hosts. Despite massive die-offs in wild bird populations and the detection of severe neurological symptoms in infected humans little is known about which mosquito species are involved in the propagation of USUV.
Methods: In the present study, the vector competence of a German (i.e., “Central European”) and a Serbian (i.e., “Southern European”) Culex pipiens biotype molestus laboratory colony was experimentally evaluated. For comparative purposes, Culex torrentium, a frequent species in Northern Europe, and Aedes aegypti, a primarily tropical species, were also tested. Adult female mosquitoes were exposed to bovine blood spiked with USUV Africa 2 and subsequently incubated at 25 °C. After two to three weeks saliva was collected from each individual mosquito to assess the ability of a mosquito species to transmit USUV.
Results: Culex pipiens biotype molestus mosquitoes originating from Germany and the Republic of Serbia and Cx. torrentium mosquitoes from Germany proved competent for USUV, as indicated by harboring viable virus in their saliva 21 days post infection. By contrast, Ae. aegypti mosquitoes were relatively refractory to an USUV infection, exhibiting low infection rates and lacking virus in their saliva.
Conclusions: Consistent with the high prevalences and abundances of Cx. pipiens biotype molestus and Cx. torrentium in Central and Northern Europe, these two species have most likely played a historic role in the spread, maintenance, and introduction of USUV into Germany. Identification of the key USUV vectors enables the establishment and implementation of rigorous entomological surveillance programs and the development of effective, evidence-based vector control interventions.
4.2 Introduction
An increase in international trade and travel, combined with climate change, has led to the recent introduction and spread of arthropod-borne pathogens into Europe (Medlock et al. 2012). The (re-)emergence of arboviruses threatens human and animal health and can cause high economic losses (Pfeffer and Dobler 2010). For example, West Nile virus (WNV) can cause encephalitis or meningoencephalitis, with a possible lethal outcome in infected humans (DeBiasi and Tyler 2006), and dengue and chikungunya viruses can lead to haemorrhagic and thereby sometimes fatal illnesses (Tomasello and Schlagenhauf 2013, Venturi et al. 2017).
Usutu virus (USUV; Flaviviridae, Flavivirus) is currently considered an emerging zoonotic arbovirus in Europe with an increase in registered cases and observed disease severity in both humans and animals. This flavivirus belongs to the Japanese encephalitis virus serocomplex

4 Manuscript I 29
with birds, mainly Eurasian blackbirds (Turdus merula), Eurasian magpies (Pica pica), house sparrows (Passer domesticus), and great grey owls (Strix nebulosa) as amplifying hosts (Clé et al. 2019, Roesch et al. 2019). Susceptible avian species show mild to severe clinical signs including encephalitis, myocardial degeneration, and necrosis of the liver and spleen (Chvala et al. 2004, Ziegler et al. 2016). Although USUV primarily causes a self-limiting, febrile illness with seroconversion in incidental hosts such as humans and horses, a neuroinvasive disease in humans, primarily in immunocompromised patients, should receive more attention. Recent studies even indicate that human USUV infections are not as sporadic as previously presumed (Clé et al. 2019, Grottola et al. 2017, Pecorari et al. 2009, Santini et al. 2015), with so far 49 reported acute infections over the years (1981-2018) in the Central African Republic, Burkina Faso, Italy, Croatia, Germany, France, and Austria (Clé et al. 2019).
USUV was first detected by McIntosh in 1959 in a Culex neavei mosquito (originally described as Cx. univittatus) near the Usutu River, Natal, South Africa (Williams et al. 1964). Phylogenetic analyses suggest a minimum of three unique USUV dispersions from Africa to Europe (in the 1950s, 1980s, and 1990s). Presumably, the virus arrived along the eastern Atlantic and Black Sea/Mediterranean routes (Clé et al. 2019) with migratory birds as long-distance dispersal vehicles (Engel et al. 2016). The first recorded outbreak of USUV in Europe, occurred in 2001 in Austria among Eurasian blackbirds and great grey owls (Chvala et al. 2007). Thereupon, USUV ribonucleic acid (RNA) was retrospectively isolated from the tissue samples of Eurasian blackbirds from 1996 in Italy (Weissenböck et al. 2013). Since then, USUV continued to colonize new ecological niches throughout Europe. In Germany, the first documented isolation of USUV was in 2010 from a pool of Cx. pipiens biotype pipiens mosquitoes collected in the city of Weinheim in the southwest of the country (Jöst et al. 2011). In the following two years, several epidemics in the Upper Rhine Valley resulted in substantial mortalities among the wild avifauna, primarily of the Eurasian blackbird (Ziegler et al. 2015). To date, USUV has spread to the north and east of Germany, covering all German federal states, with major epidemics in 2016 and 2018 (Michel et al. 2019).
USUV is currently classified into eight genetically distinct lineages (Cadar et al. 2017a) depending on their geographical origin of isolation (Engel et al. 2016), of which five circulate in Germany: “Africa 2”, “Africa 3”, “Europe 2”, “Europe 3”, and “Europe 5” (Cadar et al. 2017a, Michel et al. 2019, Ziegler et al. 2015, Ziegler et al. 2020). The Africa 2 clade, for example, originated from South Africa and was isolated in Germany in 2015 from two carcasses of juvenile great grey owls from the Zoological Garden Berlin (Ziegler et al. 2016). Till now, it is uncertain which vectors play an important role in the maintenance and spread of USUV in Germany.
USUV has been isolated from numerous mosquito species yet the primary vectors belong to the genus Culex (Clé et al. 2019). This genus contains species that already are known to be fundamental in the spread of multiple other arboviruses including Sindbis virus, St. Louis encephalitis virus, Rift Valley fever phlebovirus, and WNV (Turell 2012). So far Cx. neavei

30 4.3 Materials and Methods
from Senegal (Nikolay et al. 2012), Cx. pipiens and Cx. quinquefasciatus from North America (Cook et al. 2018), and Cx. pipiens from the Netherlands (Fros et al. 2015) proved to be highly competent for USUV under laboratory conditions. In Europe, Cx. pipiens is highly abundant (Hesson et al. 2014) and can be subdivided into two distinct biotypes: biotype pipiens (Linnaeus 1758) and biotype molestus (Forskål 1775), which vary greatly in their physiological and behavioural characteristics (Brugman et al. 2018). In Germany, both biotypes occur in sympatry or as hybrids, and their relevance for the spread of German USUV strains must be further clarified (Rudolf et al. 2013). Culex torrentium, another common species of temperate regions (e.g., Northern Europe), exhibits similar feeding-preferences as biotype pipiens, with both preferring to feed on birds, unlike the mammalophilic biotype molestus (Brugman et al. 2018). Primarily due to difficulties in the establishment of Cx. torrentium laboratory colonies, research with this species is very limited. Furthermore, there are only a few studies on the role of Aedes species such as Ae. aegypti or the invasive species Ae. japonicus and Ae. albopictus in the USUV transmission cycle (Abbo et al. 2020a, Cook et al. 2018, Puggioli et al. 2017). To date both an Italian and a North American Ae. albopictus population were found to be refractory to an USUV infection (Cook et al. 2018, Puggioli et al. 2017) while an Ae. japonicus population from the Netherlands could transmit USUV it its saliva (Abbo et al. 2020a).
In this research, vector competence studies were performed with German mosquito populations to better comprehend and predict the spread of enzootic USUV strains throughout Germany and of strains newly introduced into the country. Infection experiments were carried out with German Cx. pipiens biotype molestus (i.e., a mammalophilic species, possibly transmitting USUV to humans) and Cx. torrentium (i.e., an ornithophilic species, possibly essential for sustaining the enzootic transmission cycle between birds) to investigate their susceptibility for the USUV Africa 2 strain, “USUV-Berlin” (Ziegler et al. 2016). For comparison purposes, the vector competence of “Southern European”, Serbian Cx. pipiens biotype molestus and Malaysian Ae. aegypti colonies were also examined.
4.3 Materials and Methods
4.3.1 Mosquito Collection, Rearing, and Identification
For the infection experiments the following laboratory colonies were used: Cx. pipiens biotype molestus from the “Wendland”, Lower Saxony, Germany (established in 2012); Cx. pipiens biotype molestus from Novi Sad, Republic of Serbia (established in 2012); and Ae. aegypti originally from Malaysia (Bayer CropScience, Langenfeld, Germany). All the tested mosquito colonies were reared at 24 °C ± 1 °C with a relative humidity of approximately 60%-70% and a 16 h light/ 8 h dark photocycle. The adult mosquitoes were provided with a 5%-6% sugar solution ad libitum and for egg production with bovine or chicken (EDTA or heparin) whole blood, depending on the feeding preference of the species/biotype. Culex torrentium females were produced from egg rafts collected in the field from different populations near Berlin and Bonn, Germany, after species identification of two to three larvae hatching from each raft by

4 Manuscript I 31
means of a real-time polymerase chain reaction (PCR) (Rudolf et al. 2013). To ensure that all tested mosquito populations were free from flaviviruses prior to the experiments, individual non-engorged females per population were examined via an USUV-specific reverse transcription quantitative real-time PCR (RT-qPCR) (Jöst et al. 2011) and a SYBR® Green-based quantitative real-time pan-flavivirus assay (Viña-Rodríguez 2017).
4.3.2 Virus Strain and Cultivation
USUV Africa 2 strain was used for all infection experiments. The strain was isolated from the brain of a succumbed great grey owl in Berlin, Germany in 2015 (GenBank accession no. KU664608) (Ziegler et al. 2016). The virus was passaged twice on confluent monolayers of Vero cells (Collection of Cell Lines in Veterinary Medicine (CCLV), Friedrich-Loeffler-Institut (FLI), Greifswald-Insel Riems, Germany), and cell culture supernatant was harvested four days post infection (dpi). The virus stock was aliquoted and stored in cryovials at -70 °C until further use. For virus cultivation, Vero cells were maintained in minimum essential medium (MEM), supplemented with 2% fetal calf serum (FCS) and 1% antibiotics (penicillin and streptomycin; Merck, St. Louis, MO, The United States of America (USA)). Virus was quantified by means of an endpoint dilution assay, and the virus titer was calculated with the Spearman-Kaerber algorithm (Mayr et al. 1974). The used stock had a titer of approximately 108.1 50% tissue culture infective dose (TCID50) per mL.
4.3.3 Oral Infection with USUV
Adult mosquitoes (3-14 days old) were sorted into groups of ten individuals under 100% carbon dioxide (CO2) anesthesia one day prior to infection. After sugar-deprivation for 24-48 hours, Culex mosquitoes were fed artificially overnight. They were exposed to cotton swab ends soaked in an infectious blood meal. Aedes mosquitoes were allowed to feed for one hour on virus-spiked blood offered via a Hemotek PS5 feeder (Hemotek Ltd, Lancashire, The United Kingdom (UK)), using hog gut and parafilm to seal-off the reservoirs. The blood meal consisted of 10% virus stock, 60% bovine EDTA blood, and 30% sugar solution with a stock concentration of 5%-6%. To confirm that the used blood was free from WNV-specific antibodies, serum samples from the cows were examined with the ID Screen® WN competition enzyme-linked immunosorbent assay (ELISA) (IDVet, Grabels, France). After the preparation of the blood meal, a remainder of it was titrated on Vero cells to calculate the exact virus titer in TCID50/mL, which varied between the experiments. In two of the experiments, the virus titer of the blood meal had dropped for unknown reasons by about two-logs. The mosquitoes were, therefore, split into two groups, one was exposed to a high virus titer (107.4 TCID50/mL) and the other to a low virus titer (105.1 TCID50/mL).
Engorged mosquitoes were transferred under 100% CO2-sedation into chambers and kept in an incubator (climate test cabinet MKKL; Flohr Instruments, Nieuwegein, Netherlands and dehumidifier MG50; Munters, Bedfordshire, UK) under 25 °C ± 1 °C and a relative humidity

32 4.3 Materials and Methods
of 80%-85% and offered cotton pads soaked with 5%-6% sugar solution ad libitum. A minimum of one engorged female per species/population was frozen at -70 °C to confirm virus ingestion and as a baseline for virus replication in the mosquitoes.
4.3.4 Forced Salivation Assay
By means of forced salivation, saliva was collected after an incubation period of both 14 and 16 (14/16) or 21 days, respectively. After immobilization of the mosquitoes by removing their legs and wings under 100% CO2-anesthesia, their probosces were inserted into cut 10 µL filter tips filled with 10 µL of phosphate-buffered saline (PBS). Saliva was collected for half an hour, and the tip content was subsequently transferred into a 1.5 mL tube containing another 10 µL of PBS (Heitmann et al. 2018). Directly thereafter, the mosquito saliva samples were inoculated onto a 96-well-plate cell monolayer of Vero cells (i.e., 20 µL of each saliva solution was diluted in 150 µL of MEM + 2% FCS + 1% antibiotics). After seven days, the cells were examined under the light microscope, fixed with 7.5% neutral buffered formalin (Carl Roth, Karlsruhe, Germany), and stained with crystal violet (Carl Roth). If the saliva appeared to contain viable and replicable virus after the seven days (i.e., represented by a distinct cytopathogenic effect in the Vero cells), 140 µL of the cell culture supernatant was collected before fixing, and the RNA was extracted using the QIAamp Viral RNA Mini Kit (Qiagen, Hilden, Germany) according to manufacturer’s instructions. RNA extracts, eluted in 50 µL of the elution buffer, were tested with an USUV-specific RT-qPCR assay (5 µL at a time) (Ziegler et al. 2015) with a standard curve running in parallel (Pinho dos Reis 2018). All RT-qPCR assays were completed with the AgPath-ID One-Step RT-PCR Reagents (ThermoFischer Scientific, Darmstadt, Germany) and the CFX96TM Real-Time PCR Detection System (Bio-Rad Laboratories, Feldkirchen, Germany). To confirm correct species-association, mosquitoes which contained USUV in their saliva, were retroactively once again identified to species and biotype level. For the sake of completeness this was not only done with the field-derived mosquitoes (i.e., positive Cx. torrentium) but also with the mosquitoes originating from the laboratory colonies. For molecular identification a real-time PCR assay (Rudolf et al. 2013) was used for the Cx. pipiens complex and cytochrome c oxidase subunit 1 barcoding (Folmer et al. 1994, Hebert et al. 2003) for Ae. aegypti.
4.3.5 Pathogen Screening in Mosquito Bodies and Legs plus Wings
After saliva collection, the mosquito bodies (thorax and abdomen) and legs plus wings were stored separately in 2 mL screw cap tubes with two 3 mm steel beads, 560 µL of AVL viral lysis buffer, and carrier RNA (Qiagen) at -70 °C. For RNA extraction, mosquito bodies and legs plus wings were homogenized for 2 minutes at 30 Hz (TissueLyser II; Qiagen). Next, insoluble debris was pelleted by centrifugation for 1 minute at 13,000 rpm (5430R centrifuge; Eppendorf, Hamburg, Germany), and RNA was extracted from the supernatant with a BioSprint 96 (Qiagen) or KingFisher Flex Purification System (ThermoFischer) using the NucleoMag VET kit (Macherey-Nagel, Düren, Germany) and following the manufacturer’s instructions.

4 Manuscript I 33
RNA extracts were eluted in 100 µL elution buffer and stored at -70 °C. RNA solutions (5 µL at a time) were tested for viral RNA, using the same USUV-specific RT-qPCR assay as above (Jöst et al. 2011) with the standard curve for quantification (Pinho dos Reis 2018) and the CFX96TM Real-Time PCR Detection System (Bio-Rad Laboratories).
4.3.6 Interpretation of Results
Feeding rate refers to the number of engorged females compared to the overall number of females exposed to the infectious blood meal. Survival rate at a given time point is defined as the number of live mosquitoes out of the total number of engorged females subjected to the experiment (minus the day-0 samples). Mosquito bodies, legs plus wings, and saliva samples were considered USUV-positive if they contained USUV-specific RNA (cycle threshold below 36). A minimum of two technical replicates were completed for each positive sample in the RT-qPCR assay (Jöst et al. 2011). The infection rate describes the number of infected mosquitoes (i.e., USUV-positive bodies) out of the total number of mosquitoes analyzed. To verify that USUV infection and replication in the respective species were, however, not limited to the midgut and that viral dissemination to secondary tissues took place, legs plus wings were also examined. The dissemination rate is defined as the number of USUV-positive legs plus wings samples among the infected (i.e., USUV-positive bodies) mosquitoes. Nonetheless, a mosquito species was only considered competent for USUV when viral RNA was found in its saliva. Transmission rate is measured as the number of USUV-positive saliva samples out of the number of mosquitoes with a disseminated infection (i.e., USUV-positive bodies and legs plus wings). Transmission efficiency describes the total number of USUV-positive saliva samples out of the total number of mosquitoes analyzed.
4.3.7 Data Analysis
Statistical analyses and graphical displays were completed with the R version 3.6.0. (26 April 2019) (R Development Core Team 2019) and the additional package “lsmeans” (Lenth 2016). Generalized binomial regression models (GLM) were used to investigate the effect of mosquito species, the days post infection, and the virus titer in the blood meal including their interactions (explanatory variables) on the feeding, survival, infection, dissemination, and transmission rates and transmission efficiencies (response variables). Least-squares means (LSM) (Harvey 1960) were used for testing linear contrasts among predictions with Tukey’s adjustment for p-values (Tukey 1991). Results were considered statistically relevant when the p-values (summarized in Sup. Table 4.2 and 4.3) were less than 0.05. In the case of highly unbalanced mosquito numbers a Fisher’s exact test with Bonferroni correction was implemented instead.
Individual samples where the extremities were virus-positive but not the bodies were not included in the analyses. Two mosquitoes had USUV-positive bodies and saliva even though USUV-specific RNA quantities in their legs plus wings were close to the detection limit. However, before virus could be secreted with the saliva, the mosquitoes must have developed

34 4.4 Results
a disseminated infection. Therefore, the two samples were included when counting USUV-positive legs plus wings samples, and the low viral RNA quantities in the legs plus wings were attributed to errors in the extraction process.
4.3.8 Ethics Statement
Blood for mosquito feeding was collected from animals kept at the FLI, Greifswald-Insel Riems, Germany. The animals were held and sampled according to national and European legislation (Directive 2010/63/EU on the protection of animals used for scientific purposes), and the procedures were approved by the competent authority of the Federal State of Mecklenburg-Western Pomerania, Germany (reference number: 7221.3-2-041/17, approved 12 February 2018).
4.4 Results
4.4.1 Feeding and Survival Rates
Of the 284 German and the 101 Serbian Cx. pipiens biotype molestus, which were offered an infectious blood meal, 72.9% and 100%, respectively, were fully engorged after exposure (Figure 4.2). Similarly high was the feeding rate in the Ae. aegypti with 80.7% of the 197 exposed females. By contrast, of the F0 generation of the field-collected Cx. torrentium only 24.7% of the 89 females took a blood meal, significantly less than of the Ae. aegypti and both the Cx. pipiens biotype molestus laboratory colonies from Germany and Serbia (Fisher’s exact test: p < 0.001 for all). The few engorged Cx. torrentium females, however, showed high survival rates of 94.1% and 100% from 0 to 14/16 dpi and from 14/16 to 21 dpi (Figure 4.2; only including the survival rates from 0 to 14/16 dpi), respectively, while survival rates of the laboratory colonies after feeding from the infectious blood meal were lower. However, differences were only significant for groups of mosquitoes surviving from 0 to 14/16 dpi but not from 14/16 to 21 dpi (summarized in Sup. Table 4.3). For example, from 0 to 14/16 dpi the survival rate of the German Cx. pipiens biotype molestus was 45.6%, which was significantly lower than for Ae. aegypti with 67.1% (GLM and LSM: df = infinity, Z-ratio = 4.0, p = 1.4 x 10-3). Similarly, the survival rate of the Serbian Cx. pipiens biotype molestus was 27.3%, which was significantly lower than the survival rates of both Ae. aegypti (GLM and LSM: df = infinity, Z-ratio = 6.0, p < 0.001) and Cx. torrentium (GLM and LSM: df = infinity, Z-ratio = -3.6, p = 9.0 x 10-3).
4.4.2 Infection, Dissemination, and Transmission Rates
The infection, dissemination, and transmission rates of the four mosquito infection experiments conducted in this study are summarized in Table 4.1. The virus titers of the blood meals were distinguished as being either high with 107.4 TCID50/mL or low with 105.1 TCID50/mL. After uptake via the infectious blood meal, USUV was able to establish an infection in all of the tested species. However, only Cx. pipiens biotype molestus and Cx. torrentium proved competent for

4 Manuscript I 35
USUV. After oral exposure to a high virus titer (107.4 TCID50/mL) in the blood meals, viral RNA could be detected in the bodies, legs plus wings, and saliva samples of multiple mosquitoes from both the German and Serbian Cx. pipiens biotype molestus colonies. The saliva secreted by six of the German and six of the Serbian Cx. pipiens biotype molestus mosquitoes contained virus particles that successfully infected mammalian Vero cells with substantial virus replication 7 dpi. Significant differences, however, were not observed among the vector competence indices (infection, dissemination, and transmission rates), neither between the mosquito species nor between the collection dates (i.e., 14/16 and 21 dpi). This, however, may be a result of the small sample sizes per species and time point. By contrast, a low virus titer (105.1 TCID50/mL) in the blood meals yielded a very low infection rate of 5.6% (2/36) 14 dpi and 5.3% (1/19) 21 dpi in the German Cx. pipiens biotype molestus with no virus replication in secondary tissue. Nonetheless, the low virus titer still infected one out of a total of eight Cx. torrentium specimens 21 dpi, with subsequent virus dissemination and transmission. Extensive virus replication took place with RNA viral loads of 3.9 x 106 per µL of total RNA in the body and 4.7 x 104 per µL of total RNA in the legs plus wings. In comparison, the day-0 Cx. torrentium mosquitoes contained on average 3.5 x 104 per µL of total RNA. Furthermore, the virus titer in the blood meals not only affected the infection, dissemination, and transmission rates but also the viral loads in the individual mosquito samples. For example, after oral inoculation with a high titer, 6.3 x 105 virus copies per µL of total RNA were found on average in the bodies and 1.2 x 104 virus copies per µL of total RNA in the legs plus wings of the German Cx. pipiens biotype molestus 14/16 and 21 dpi. This is higher than the viral load found in the day-0 samples with 1.1 x 104 virus copies per µL of total RNA. A low virus titer resulted in a decline in the number of virus copies to 8.7 x 101 per µL of total RNA in the bodies and no virus copies in the legs plus wings of the German Cx. pipiens biotype molestus 14/16 and 21 dpi. Even when exposed to a high virus titer, Ae. aegypti had an infection rate of 0% 14/16 dpi and only 18.2% 21 dpi, with the detection of virus dissemination in only one mosquito. USUV could not be detected in the saliva of Ae. aegypti mosquitoes.
4.5 Discussion
The goal of this study was to identify German mosquito species that are vector-competent for USUV in order to establish evidence-based recommendations regarding the implementation of mosquito surveillance and control strategies. The study proved for the first time that Serbian and German Cx. pipiens biotype molestus and German Cx. torrentium mosquito populations are competent in replicating and transmitting the German USUV Africa 2 strain from Berlin. The vector competence of Culex mosquitoes for zoonotic viruses like USUV is, nevertheless, not a new discovery. Several infection experiments have been performed in the past with Culex species (Clé et al. 2019). This genus includes species which are not only abundant in Europe but can also function as bridge vectors, due to the high variability in their host-feeding patterns, transmitting zoonotic pathogens from birds to mammals (Hamer et al. 2008). A Cx. neavei population from Senegal, a species which is ubiquitous in Africa, was verified to be highly

36 4.5 Discussion
competent for the Africa 2 USUV strain SAAR-1776 (GenBank accession no. AY453412) and is therefore a likely endemic vector in the USUV transmission cycle in Africa. Fourteen days post exposure to a blood meal with a virus titer of 106.5 plaque forming units (PFU) per mL, its infection rate was 91%, its dissemination rate 40%, and its transmission rate 81% (Nikolay et al. 2012). These values are very similar to the rates found in this report for the German Cx. pipiens biotype molestus. Recently performed infection experiments with a Cx. pipiens population from the Netherlands (Fros et al. 2015) demonstrated for the first time the ability of European mosquitoes to transmit a Europe 2 USUV strain from Italy (Bologna/09; GenBank accession no. HM569263). Similarly to the African Cx. neavei and the German Cx. pipiens biotype molestus from this study, the infection rates were high (80%), with virus dissemination and accumulation rates in the saliva of 69% (Fros et al. 2015). However, contrary to the infection of the European mosquitoes with the Italian USUV strain, two Cx. pipiens populations from the UK (Hernández-Triana et al. 2018) showed a low susceptibility to the African SAAR-1776 strain. Infection rates ranged from 0% to 14.2%, and only one out of 48 Cx. pipiens was positive for USUV in its saliva 14 dpi (Hernández-Triana et al. 2018). The variation in susceptibility could be due to the genetic variability of the virus strains with in situ evolution and host-specific mutations in European but not African lineages (Engel et al. 2016). With respect thereto, it is especially interesting that Cx. pipiens and Cx. quinquefasciatus colonies from North America (Cook et al. 2018), where USUV has not been detected yet, proved highly susceptible to the very same Africa 2 strain (SAAR-1776). Culex pipiens showed an infection rate of 59%, and of these, 24% were positive in their saliva. Culex quinquefasciatus yielded similar results (Cook et al. 2018). It is, therefore, highly likely that varying blood meal virus titers (107.5 compared to 106.0 TCID50/mL) or disparities in the vector competence of geographically distinct mosquito populations of the same species also influenced the results.
So far, no vector competence studies have been performed with Cx. torrentium and USUV. This is, therefore, the first confirmation of USUV transmission by a Cx. torrentium specimen. Due to the limited availability of Cx. torrentium females for this study the vector competence of this species was only examined after feeding from a blood meal with a low virus titer. It would be highly interesting to also test the species’ vector competence after feeding from a high virus titer and compare it to that of other species. Only recently was a German Cx. torrentium population described to be highly competent for the closely related flavivirus WNV with transmission rates of up to 90% (Jansen et al. 2019). Culex torrentium even transmitted WNV to a greater degree than Cx. pipiens biotype molestus (Jansen et al. 2019).
In addition to the results from the vector competence experiments, the role of Culex mosquitoes in the transmission of USUV is reinforced through the findings of USUV in native mosquitoes in Africa and Europe. For example, USUV was detected in Cx. antennatus, Cx. modestus, Cx. neavei, Cx. perexiguus, Cx. perfuscus, Cx. pipiens, and Cx. quinquefasciatus (Clé et al. 2019). Since the first isolation of USUV in Germany in 2010, multiple detections in Cx. pipiens sensu lato (s.l.) have been described: in 2014 near Freiburg, Baden-Wuerttemberg (Europe 3)

4 Manuscript I 37
(Scheuch et al. 2018), in 2015 near Leipzig, Saxony-Anhalt (Africa 2) (Sieg et al. 2017), and in 2016 in Emsdetten, North Rhine-Westphalia (Africa 3) (Scheuch et al. 2018). To date, USUV-positive Cx. pipiens s.l. have also been collected in Austria, France, Italy, Serbia, Spain, and Switzerland (Clé et al. 2019). It must, however, be kept in mind that the mere isolation of USUV from field-collected, homogenized, whole mosquitoes cannot give a precise indication on the vector status of a specific mosquito species.
Of the invasive mosquito species in Europe, USUV nucleic acid was isolated from the Asian bush mosquito Ae. japonicus in Graz, Austria (Camp et al. 2019) and the Asian tiger mosquito Ae. albopictus in Emilia-Romagna region, Italy (Calzolari et al. 2010, Mancini et al. 2017, Tamba et al. 2011). Two vector competence studies have described Ae. albopictus as being refractory to an USUV infection, where neither an African (SAAR-1776) nor various Italian strains (GenBank accession no. KF055442, KF055441, and KF055440) resulted in virus accumulation in the mosquitoes’ saliva (Cook et al. 2018, Puggioli et al. 2017). In comparison, this study also investigated the USUV-vector competence of the yellow fever mosquito Ae. aegypti. This species is thermophilic and endemic in tropical and subtropical regions of the world. Multiple introductions into Europe have been described, such as into a German household (Kampen et al. 2016) and a Dutch airport (Ibañez-Justicia et al. 2017) by means of passive mosquito dispersal. Similarly to Ae. albopictus, the results of this study show that Ae. aegypti mosquitoes are relatively refractory to USUV. The species is, therefore, unlikely to be involved in the transmission of USUV in Europe despite virus replication and dissemination in individual specimens. The infection rates of both the German and Serbian Cx. pipiens biotype molestus colonies were higher than those of the tested Ae. aegypti. None of the tested Ae. aegypti saliva samples contained viable virus. Nonetheless, due to the ongoing expansion of invasive species such as Ae. albopictus (Kuhlisch et al. 2018, Walther et al. 2017) and Ae. japonicus (Kampen and Werner 2014, Zielke et al. 2016), the need to also investigate the role of these mosquito species in the transmission of USUV to incidental hosts (e.g., humans and equines) remains.
Multiple studies have demonstrated the virus-dose dependency of a mosquito species’ vector competence after an oral infection with USUV (Nikolay et al. 2012) and WNV (Vogels et al. 2017c). This correlates with the results in this report where a two-log drop in the virus titer resulted in the absence of USUV-specific RNA in secondary tissues and the saliva of German Cx. pipiens biotype molestus. The generalized binomial regression model calculated a significant increase in the infection rates after exposure to a high rather than a low virus titer in the blood meals (GLM and LSM: df = infinity, Z-ratio = -4.7, p < 0.001). Nonetheless, recurring USUV outbreaks amongst the German avifauna, such as in 2017 and 2018 (Michel et al. 2019), verify the probable presence of numerous wild bird species producing viral loads high enough to infect susceptible mosquito species. In the case of the closely related WNV, viremia levels in birds can reach 1012 TCID50/mL, yet most birds do not reach titers higher than 108 TCID50/mL (Vogels et al. 2017c).

38 4.5 Discussion
The virus titer is not the only variable to take into account, as there is also a strong temperature dependency of vector competence. For example, the Dutch Cx. pipiens infected with the Italian USUV strain had distinctly higher infection rates at 28 °C than at 18 °C (90% versus 11%) (Fros et al. 2015). This temperature dependency of vector competence is multifactorial and varies between mosquito species. The positive correlation between the vector competence for USUV and the ambient temperature is amongst others a result of the ectothermic nature of mosquitoes, where an increase in temperature directly results in an increase in the replication rate of the virus (Lambrechts et al. 2011). This, in turn, results in a reduction of the extrinsic incubation period (Lühken et al. 2017, Vogels et al. 2017a). Furthermore, a reduced effectiveness of the midgut barrier at higher temperatures, temperature-induced changes in the regulation of biotype-specific immune-responsive genes, and temperature-dependent activation of RNA-interference pathways can all influence vector competence (Vogels et al. 2017a). All experiments in this study were performed at 25 °C. This temperature resembles the average summer conditions around the Mediterranean Sea and southeastern European countries but is higher than the recorded long-term mean temperatures of German summers. Yet, extreme heat waves such as in 2003, 2018, and 2019 have clearly demonstrated that an increase from the long-term mean temperatures in Germany is becoming more frequent (DWD 2018b, 2019). 2018 was the second warmest summer in Germany ever recorded, possibly shortening the extrinsic incubation period and allowing a more rapid autochthonous virus transmission (Ziegler et al. 2019). Therefore, the occurrence of temperatures high enough to support the USUV vector-host transmission cycle in Germany is also conceivable and can help explain the increasing frequency of avian infections (Michel et al. 2018, Michel et al. 2019, Ziegler et al. 2015).
In this experiment, numerous other factors may have had an effect on the results. Aside from the Cx. torrentium specimens, all mosquitoes originated from established colonies with multiple generations in the laboratory. This may have had an impact on the genetic and phenotypic variations of the mosquito strains. The susceptibility of a mosquito for a virus cannot only be generation- but also time- and population-specific, with geographic variations in the vector competence (Richards et al. 2010, Vogels et al. 2017c). Furthermore, to obtain optimal feeding rates a sugar solution was applied to the blood meals. Even though a final sugar concentration of 1.5-1.8% in the blood meals is probably too low to influence a mosquito’s vector competence (Vaidyanathan et al. 2008), its influence on the mosquito’s immune response and its gut microbiota (Vaidyanathan et al. 2008, Muturi et al. 2019) and therefore also on its vector competence for USUV remains unclear. Not only can the gut microbiome or the presence of intracellular Wolbachia bacteria influence a mosquito’s vector competence but also coinfections with other viruses or pathogens. For example, the German and Serbian Cx. pipiens biotype molestus colonies tested competent for USUV in this study already proved their high transmission efficiency for a German WNV isolate (Holicki et al. 2020b). USUV and WNV coinfections are not only highly conceivable in mosquitoes but have also been described in 34 avian species and in horses in Europe (Roesch et al. 2019). Future studies should, therefore,

4 Manuscript I 39
focus on what effect viral interferences will have on the antibody production in susceptible hosts and the immune response of mosquitoes (Rizzoli et al. 2015).
4.6 Conclusion
Both the German (“Central European”) and Serbian (“Southern European”) Cx. pipiens biotype molestus laboratory colonies could transmit the USUV Africa 2 lineage at an extrinsic incubation temperature of 25 °C. A specimen obtained from field-collected egg-rafts of Cx. torrentium, a highly prevalent species in Northern Europe, was also capable of transmitting USUV. The tropical mosquito species Ae. aegypti, on the other hand, showed reduced susceptibility with no detection of virus in its saliva.

40 4.7 Tables
4.7 Tables
Table 4.1 Infection, dissemination, and transmission rates of mosquitoes infected with the German USUV Africa 2 strain.
Blood Meal
Virus Titer
(TCID50/mL)
Mosquito
Species Dpi
Infection
Rate
(%)
(95% CI)
Mean Viral
Load
Bodies
(Viral Copies
/µL of Total
RNA)
Dissemination
Rate
(%)
(95% CI)
Mean Viral Load
Legs plus Wings
(Viral Copies
/µL of Total RNA)
Transmission
Rate
(%)
(95% CI)
High Titer
107.4
Cx. pipiens biotype
molestus †
14 8/10
(80.0) (44.4-97.5)
6.9 x 105 3/8
(37.5) (8.5-75.5)
9.0 x 103 3/3
(100) (29.2-100)
21 4/6
(66.7) (22.3-95.7)
5.6 x 105 4/4
(100) (39.7-100)
1.5 x 104 3/4
(75.0) (19.4-99.4)
Cx. pipiens biotype
molestus ‡
16 13/16 (81.3)
(54.4-96.0) 1.9 x 106
13/13 (100)
(75.3-100) 7.8 x 104
2/13 (15.4)
(1.9-45.4)
21 8/10
(80.0) (44.4-97.5)
8.1 x 105 8/8
(100) (63.1-100)
7.8 x 104 4/8
(50.0) (15.7-84.3)
Ae. aegypti
14 0/53 (0.0)
(0.0-6.7) NA NA NA NA
21 4/22
(18.2) (5.2-40.3)
2.3 x 105 1/4
(25.0) (0.6-80.6)
5.5 x 103 0/1
(0.0) (0.0-97.5)
Low Titer
105.1
Cx. pipiens biotype
molestus †
14 2/36 (5.6)
(0.7-18.7) 1.2 x 102
0/2 (0.0)
(0.0-84.2) NA NA
21 1/19 (5.3)
(0.7-18.7) 5.4 x 101
0/1 (0.0)
(0.0-84.2) NA NA
Cx. torrentium §
14 1/8
(12.5) (0.3-52.7)
2.8 x 101 0/1
(0.0) (0.0-97.5)
NA NA
21 1/8
(12.5) (0.3-52.7)
3.9 x 106 1/1
(100) (2.5-100)
4.7 x 104 1/1
(100) (2.5-100)
Transmission rates include results from the saliva inoculation on Vero cells and from the RT-qPCRs of cell culture supernatants; All mosquitoes were incubated for 14/16 or 21 days; Absolute quantification of viral copies/µL of total RNA was performed via an RT-qPCR based calibration curve; CI, confidence interval; dpi, days post infection; NA, not applicable; †, Cx. pipiens biotype molestus laboratory colony from “Wendland”, Lower Saxony, Germany; ‡, Cx. pipiens biotype molestus laboratory colony from Novi Sad, the Republic of Serbia; §, Cx.

4 Manuscript I 41
torrentium field-collected colony near Berlin and Bonn, North Rhine-Westphalia, Germany; ¶, Ae. aegypti laboratory colony from Malaysia (Bayer CropScience, Langenfeld, Germany).

42 4.8 Figures
4.8 Figures
4.8 F
igu
res
Fig
ure 4
.1 G
raphical abstract displaying work flow
of vector competence experim
ents.

4 Manuscript I 43
Figure 4.2 Comparison of the feeding and survival rates (from 0 to 14/16 dpi) of the four tested mosquito populations. Data values above the bars indicate the number of fully engorged or survived females per species, respectively. Numbers in brackets specify the ratio of engorged and survived females to the total number of females exposed to a blood meal or subjected to the experiment (minus day-0 samples), respectively. Error bars represent 95% confidence intervals. * p < 0.05, ** p < 0.01, and *** p < 0.001 by generalized binomial regression models or Fisher’s exact test with Bonferroni correction. † Cx. pipiens biotype molestus laboratory colony from “Wendland”, Lower Saxony, Germany. ‡ Cx. pipiens biotype molestus laboratory colony from Novi Sad, Republic of Serbia. § Cx. torrentium field-collected colony near Berlin and Bonn, North Rhine-Westphalia, Germany. Ae. aegypti laboratory colony from Malaysia (Bayer CropScience, Langenfeld, Germany).

44 4.9 Acknowledgments
4.9 Acknowledgments
A special thank you goes to Jana Schulz (FLI) for her support with the statistical analyses of the data. We are grateful to René Schöttner and Cornelia Steffen (FLI) for excellent technical assistance in the laboratory. Furthermore, we want to thank René Klein, Marlene Hausner, and Juliane Horenk (FLI) for rearing the mosquitoes and their species identification. We would like to thank Dušan Petrić (Faculty of Agriculture, University of Novi Sad, Novi Sad, Republic of Serbia) for supplying the Serbian Cx. pipiens biotype molestus colony and Cornelia Silaghi (FLI) for her theoretical support. We are thankful to Vinícius Pinho dos Reis (FLI) for providing us with the synthetic USUV-RNA for the calibration curve and Bärbel Hammerschmidt (FLI) for supplying the cattle blood for the infectious blood meals. We would also like to thank Anna Heitmann and Stephanie Jansen (Bernhard-Nocht-Institut (BNITM), Hamburg, Germany) for their instructions and theoretical advice.
This work was financially supported by the German Federal Ministry of Food and Agriculture (BMEL) through the Federal Office for Agriculture and Food (BLE), grant numbers 2819104915, 2819104615, and 2818SE001.

5 Manuscript II 45
5 MANUSCRIPT II
West Nile Virus Lineage 2 Vector Competence of Indigenous Culex and
Aedes Mosquitoes from Germany at Temperate Climate Conditions
Cora M. Holicki 1, Ute Ziegler 1, Cristian Răileanu 2, Helge Kampen 2, Doreen Werner 3,
Jana Schulz 1, Cornelia Silaghi 2, Martin H. Groschup 1, and Ana Vasić 2,*
1 Institute of Novel and Emerging Infectious Diseases, Friedrich-Loeffler-Institut, Federal Research Institute for Animal Health, 17493 Greifswald-Insel Riems, Germany
2 Institute of Infectology, Friedrich-Loeffler-Institut, Federal Research Institute for Animal Health, 17493 Greifswald-Insel Riems, Germany
3 Biodiversity of Aquatic and Semiaquatic Landscape Features, Leibniz-Centre for Agricultural Landscape Research, 15374 Muencheberg, Germany
Author to whom correspondence should be addressed: [email protected]
5.1 Abstract
West Nile virus (WNV) is a widespread zoonotic arbovirus and a threat to public health in Germany since its first emergence in 2018. It has become of particular relevance in Germany in 2019 due to its rapid geographical spread and the detection of the first human clinical cases. The susceptibility of indigenous Culex pipiens (biotypes pipiens and molestus) for a German WNV lineage 2 strain was experimentally compared to that of Serbian Cx. pipiens biotype molestus and invasive German Aedes albopictus. All tested populations proved to be competent laboratory vectors of WNV. Culex pipiens biotype pipiens displayed the highest transmission efficiencies (40.0%-52.9%) at 25 °C. This biotype was also able to transmit WNV at 18 °C (transmission efficiencies of 4.4%-8.3%), proving that temperate climates in Central and Northern Europe may support WNV circulation. Furthermore, due to their feeding behaviors, Cx. pipiens biotype molestus and Ae. albopictus can act as bridge vectors, leading to human WNV infections.
Published in Viruses in 2020
Volume 12, Issue 5 https://doi.org/10.3390/v12050561


6 Manuscript III 47
6 MANUSCRIPT III
Intrathoracic Injection of German Culex Mosquitoes with West Nile Virus
to Secure Virus Infection and Transmission
Cora M. Holicki 1, Ute Ziegler 1, Cristian Răileanu 2, Helge Kampen 2, Doreen
Werner 3, Cornelia Silaghi 2, Martin H. Groschup 1, and Ana Vasić 2,*
1 Institute of Novel and Emerging Infectious Diseases, Friedrich-Loeffler-Institut, Federal Research Institute for Animal Health, 17493 Greifswald-Insel Riems, Germany
2 Institute of Infectology, Friedrich-Loeffler-Institut, Federal Research Institute for Animal Health, 17493 Greifswald-Insel Riems, Germany
3 Biodiversity of Aquatic and Semiaquatic Landscape Features, Leibniz-Centre for Agricultural Landscape Research, 15374 Muencheberg, Germany
Author to whom correspondence should be addressed: [email protected]
This manuscript (short communication) was prepared for submission.

48 6.1 Abstract
6.1 Abstract
West Nile virus (WNV) is a widely-dispersed zoonotic arbovirus. In 2018, the virus emerged in Germany with an unforeseeable effect on animal and human health. To better uncover the natural transmission cycle of WNV in central Europe, bird and mosquito transmission models are essential. For this purpose the vector competence of two ubiquitous Culex species (Cx. pipiens biotype pipiens and Cx. torrentium) infected intrathoracically with WNV were examined. 100% infection and dissemination rates were achieved after 10/11 and 22 days post infection (dpi), respectively in all tested specimens. The transmissions rates were around 50% in all populations after 10/11 dpi and reached 81.8% in the Cx. torrentium 22 dpi. It can, therefore, be concluded that the intrathoracic injection of Culex populations can guarantee high infection rates and high transmission rates, especially after longer incubation periods. These results are essential prerequisites for the establishment of animal infection models with a natural route of infection.
6.2 Introduction
West Nile virus (WNV) is a mosquito-borne flavivirus affecting both human and animal health. Flaviviruses are positive-sense, single-stranded RNA viruses (Rice et al. 1985), with WNV being a member of the Japanese encephalitis virus serocomplex (Calisher et al. 1989). In susceptible species WNV can induce asymptomatic infections as well as a wide range of clinical signs: from mild “flu-like” signs to severe neurological diseases (including meningitis, encephalitis, and acute flaccid paralysis) (Campbell et al. 2002). The sylvatic transmission cycle of WNV comprises different mosquito species as vectors and wild birds as amplifying hosts (Lanciotti et al. 1999). Mosquito species designated to be key vector species in Europe are part of the Culex pipiens complex (Vogels et al. 2016), of which, Cx. pipiens biotype pipiens and Cx. torrentium are ubiquitous in Germany (Rudolf et al. 2013). Even though these species are predominantly ornithophilic they can still act as bridge vectors transmitting the virus to susceptible humans and animals (Börstler et al. 2016, Rudolf et al. 2013). The intrathoracic injection of mosquitoes with virus enables bypassing of the midgut barrier and has produced stable virus infections in mosquitoes from various populations (Abbo et al. 2020b, Bergold et al. 1968, Terradas and McGraw 2019, Uraki et al. 2018). The aim of our study was, therefore, to demonstrate that the intrathoracic injection of WNV results in infectious mosquitoes independent of the species. The successful implementation of a standardized infection method guaranteeing WNV-positive mosquitoes that transmit virus in their saliva is a fundamental cornerstone for animal infection studies. In order to mimic natural WNV transmission in the field animal, species should be infected via mosquito bite. This enables an assessment of the influence of mosquito-vectors on the susceptibility and pathogenesis of WNV in vertebrate hosts.

6 Manuscript III 49
6.3 Materials and Methods
6.3.1 Mosquito Origin and Rearing
Culex pipiens biotype pipiens egg rafts were collected in 2018 in Niederhof, Mecklenburg-Western Pomerania, Germany (54.2562N, 13.1958E) (Wöhnke et al. 2020) and in 2019 from Brandenburg, Germany (“Schöneiche/Rehfelde” and “Groß Kreutz”). The Cx. torrentium egg rafts were also collected in the same region of Brandenburg (“Schöneiche/Rehfelde”) in 2018 and 2019. All egg rafts were transferred to the biosafety level 2 (BSL-2) insectary (Friedrich-Loeffler-Institut, (FLI), Greifswald-Insel Riems, Germany) where they were kept in plastic containers filled with tap water and 0.5 g of grounded fish food (TetraMin; Tetra, Melle, Germany). Hatched larvae (n = 1-3) from each egg raft were euthanized by submerging them in 70% ethanol and transferred to a 2 mL tube (Eppendorf, Hamburg, Germany) filled with 200 µL of modified minimum essential medium (MEM) (Collection of Cell Lines in Veterinary Medicine (CCLV), FLI) and a 3 mm steel bead (Qiagen, Heidelberg, Germany). Samples were subsequently grounded (Tissuelyser II; Qiagen) and deoxyribonucleic acid (DNA) was extracted following manufacturers’ instructions (QIAamp DNA Mini Kit; Qiagen). Species identification was performed by the sequencing of the mitochondrial cytochrome c oxidase subunit I gene (Folmer et al. 1994). Furthermore, a real-time polymerase chain reaction (PCR) was employed to distinguish between the two Cx. pipiens biotypes (Rudolf et al. 2013). Larvae from the same species and biotype were subsequently joined and reared together in a plastic basin. After pupation the pupae were transferred into mosquito breeding cages (BugDorm 30 x 30 x 30 cm; MegaView Science Co., Ltd., Taichung, Taiwan) and adults were reared at 24-26 °C, 65%-80% relative humidity, 16 h light/ 8 h dark photocycle, and with 5%-6% sugar solution ad libitum. A WNV-specific reverse transcription quantitative real-time PCR (RT-qPCR) (Eiden et al. 2010) was used to confirm that field-acquired mosquitoes were not infected with WNV (e.g., transovarially) prior to the experiments.
6.3.2 Mosquito Infection
Seven-to-14-day-old female Cx. pipiens biotype pipiens and Cx. torrentium mosquitoes were anesthetized with 100% carbon dioxide (CO2) and injected intrathoracically (in the thoracic spiracles) with approximately 500 TCID50 of an Italian WNV lineage 1 strain (TOS_09; GenBank accession no. HM991273/HM641225 (Rossini et al. 2011); passaged on Ae. albopictus C6/36 and mammalian Vero cells (Holicki et al. 2020a)) in a volume of 41.4 nL using a nanoliter injector (Nanoject II; Drummond Scientific Company, Broomall, PA, USA) and pulled 5 µL glass capillary needles (PUL-1000 Micropipette Puller; World Precision Instruments, Sarasota, FL, USA). All procedures were completed in a biosafety level 3 (BSL-3) insectary, inside of a glovebox (Terra Universal Inc, Fullerton, CA, USA), and under microscopic control (Leica Mikrosysteme Vertreib GmbH, Wetzlar, Germany). Injected mosquitoes were placed into modified Drosophila tubes (foam plug on one end and netting on

50 6.3 Materials and Methods
the other; Carl Roth, Karlsruhe, Germany). Injected mosquitoes were incubated for 10/11 or 22 days at 25 ± 1 °C, 80%-85% relative humidity, and fed ad libitum with a 5%-6% sugar solution.
6.3.3 Mosquito Processing
Ten days post injection (dpi) saliva was collected from the mosquitoes according to the protocol of Heitmann et al. 2018. Briefly summarized: under 100% CO2-anesthesia mosquito extremities (legs plus wings) were removed and the proboscises were inserted into cut 20 µL filter tips filled with 10 µL of phosphate-buffered saline (PBS). Mosquitoes were allowed to salivate for 45-60 min. Thereafter, the mosquito bodies and legs plus wings were added to separate 2 mL tubes with 560 µL AVL buffer (Qiagen) and two 3-mm steel beads (Qiagen). Samples were homogenized (30 Hz for 2 minutes; Tissuelyser II; Qiagen) and stored at -70 °C until further use.
Saliva samples were transferred into 1.5 mL tubes containing additional 10 µL of PBS and inoculated onto Vero B4 cell monolayers (CCLV) to enable the microscopic (Nikon Eclipse Ts2; Nikon, Tokyo, Japan) detection of a cytopathogenic effect a week later. To verify that an observed CPE was WNV-related 140 µL of the cell culture supernatant was added to 560 µL of AVL buffer (Qiagen) and stored at -70 °C until further use.
In preparation for RNA extraction all samples were centrifuged at room temperature for 1 minute at 13,000 rpm (5430R centrifuge; Eppendorf). Nucleic acid was extracted from 200 µL of the supernatant from mosquito bodies and legs plus wings using the NucleoMag VET kit (Macherey-Nagel, Düren, Germany) and the BioSprint 96 (Qiagen). RNA extracts were eluted in 100 µL of elution buffer and stored at -70 °C. However, RNA was extracted per hand (QIAamp Viral RNA Mini Kit; Qiagen) from the entire volume of saliva cell culture supernatant (700 µL). Amplification of RNA (5 µL) from mosquito bodies, legs plus wings, and saliva cell culture supernatants was performed with a WNV-specific rRT-PCR assay (Eiden et al. 2010) using the AgPath-ID One-Step RT-PCR Reagents (ThermoFischer Scientific, Darmstadt, Germany) and the CFX96TM Real-Time PCR Detection System (Bio-Rad Laboratories, Feldkirchen, Germany). For RNA quantification via RT-qPCR an absolute standard curve based on synthetic WNV RNA was run in parallel using ten-fold serial dilutions (Eiden et al. 2010).
6.3.4 Vector Competence Indices and Data Analysis
All statistical analyses and graphical displays were performed with R version 3.6.0 (26 April 2019) (R Development Core Team 2019). Generalized binominal regression models (GLM) were used to examine the effect of species, including their interactions (explanatory variables) on the infection, dissemination, and transmission rates (response variables). Least-squares means (Harvey 1960) were used for testing linear contrasts among predictions with Tukey adjustment for p-values (Tukey 1991). Results were deemed statistically relevant when the p-values were less than 0.05.

6 Manuscript III 51
6.4 Results
In total 140 Cx. pipiens biotype pipiens from Mecklenburg-Western Pomerania, 215 Cx. pipiens biotype pipiens, and 141 Cx. torrentium from Brandenburg were injected intrathoracically with the Italian WNV lineage 1 strain (results summarized in Table 6.1). Independent of the species and population all mosquitoes became infected (i.e., WNV-positive bodies) after an intrathoracic injection with WNV. Without exception disseminated infections (i.e., WNV-positive legs plus wings) were also observed in all specimens. However, only around half of the mosquitoes had infectious WNV in their saliva 10/11 dpi. By contrast, the ratio of mosquitoes expectorating WNV in their saliva increased for Cx. torrentium 22 dpi. Nonetheless, the statistical analysis revealed no significant differences between the tested populations regarding their vector competence indices (summarized in Sup. Table 6.1). The viral loads measured in the bodies were fairly similar between all species (Figure 6.1). The viral loads in the legs plus wings of Cx. pipiens biotype pipiens from Brandenburg were lower than those of Cx. pipiens biotype pipiens from Mecklenburg-Western Pomerania. Reduced viral loads were also observed for Cx. torrentium 22 dpi instead of 10/11 dpi.
6.5 Discussion
The intrathoracic injection of mosquitoes with a virus is a reliable and controlled method for producing infectious vectors and is, therefore, highly suitable for testing/establishing animal infection models following a natural route of infection. Not only does the process allow all mosquitoes to receive the same amount of virus but it also speeds up the infection process by bypassing the midgut barrier and the mosquito’s first innate immune defense mechanisms (Kuo et al. 2018). The intrathoracic injection of both Cx. pipiens biotype pipiens und Cx. torrentium with WNV in this study assured a disseminated infection in all specimens, reaching already 10/11 dpi high transmission rates. No significant differences in susceptibility (infection, dissemination, and transmission rates) were observed between the different populations and species tested. These results coincide with the 100% infection rates described for several mosquito populations (e.g., from North America, Italy and the Netherlands) after an intrathoracic injection with WNV (Fros et al. 2015, Vogels et al. 2017a). In this study, constant transmission rates of approximately 50% 10/11 dpi suggest the existence of a salivary gland barrier for WNV in all tested species. This is probably dependent on the incubation period as, for example for Cx. torrentium, transmission efficiencies were higher after 22 (81.8%) than 10/11 (52.5%) dpi. This study also provides proof for the existence of a midgut barrier for WNV in Culex species as the two injected Cx. pipiens biotype pipiens populations had higher transmission rates than a Cx. pipiens biotype pipiens population (from Schleswig-Holstein, Germany) infected via a virus-spiked blood meal (50.0% or 52.1% compared to 14.3% at 24 °C or 33.3% at 27 °C) (Jansen et al. 2019).
Interestingly, all heads of the Cx. pipiens biotype pipiens from Brandenburg (data shown in Holicki et al. 2020a) were WNV positive suggesting a discrepancy between mosquito heads
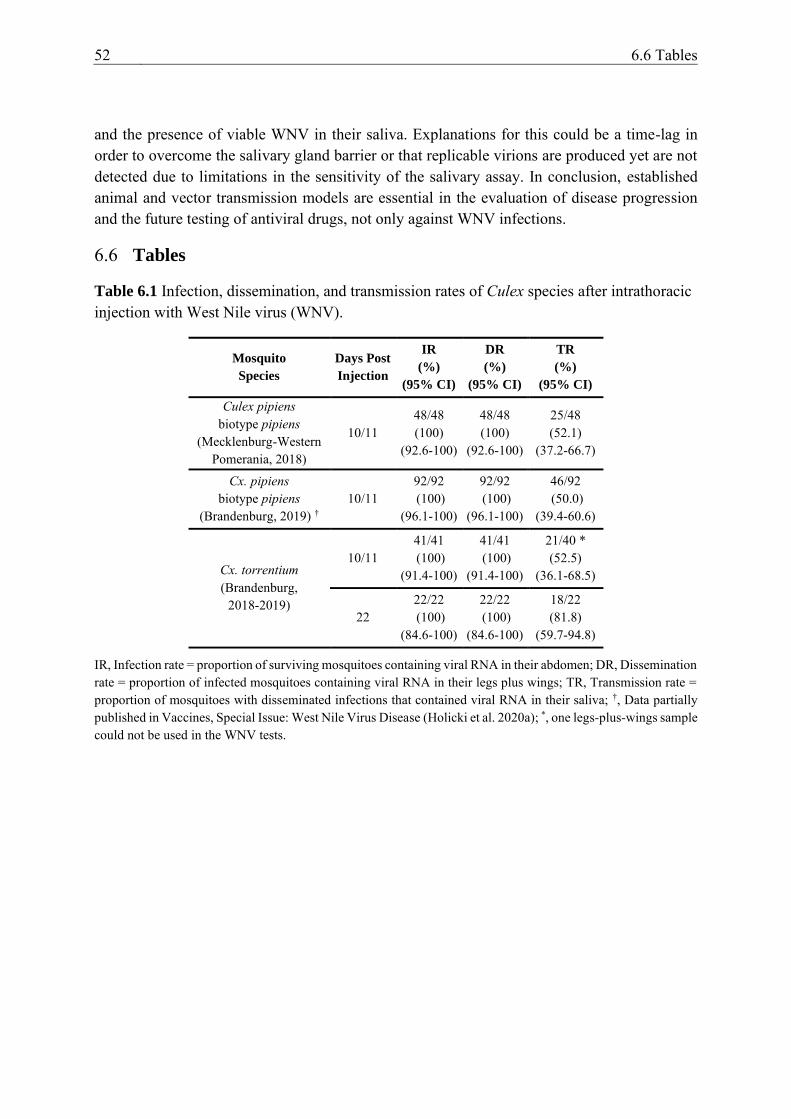
52 6.6 Tables
and the presence of viable WNV in their saliva. Explanations for this could be a time-lag in order to overcome the salivary gland barrier or that replicable virions are produced yet are not detected due to limitations in the sensitivity of the salivary assay. In conclusion, established animal and vector transmission models are essential in the evaluation of disease progression and the future testing of antiviral drugs, not only against WNV infections.
6.6 Tables
Table 6.1 Infection, dissemination, and transmission rates of Culex species after intrathoracic injection with West Nile virus (WNV).
Mosquito
Species
Days Post
Injection
IR
(%)
(95% CI)
DR
(%)
(95% CI)
TR
(%)
(95% CI)
Culex pipiens biotype pipiens
(Mecklenburg-Western Pomerania, 2018)
10/11 48/48 (100)
(92.6-100)
48/48 (100)
(92.6-100)
25/48 (52.1)
(37.2-66.7)
Cx. pipiens biotype pipiens
(Brandenburg, 2019) † 10/11
92/92 (100)
(96.1-100)
92/92 (100)
(96.1-100)
46/92 (50.0)
(39.4-60.6)
Cx. torrentium (Brandenburg,
2018-2019)
10/11 41/41 (100)
(91.4-100)
41/41 (100)
(91.4-100)
21/40 * (52.5)
(36.1-68.5)
22 22/22 (100)
(84.6-100)
22/22 (100)
(84.6-100)
18/22 (81.8)
(59.7-94.8)
IR, Infection rate = proportion of surviving mosquitoes containing viral RNA in their abdomen; DR, Dissemination rate = proportion of infected mosquitoes containing viral RNA in their legs plus wings; TR, Transmission rate = proportion of mosquitoes with disseminated infections that contained viral RNA in their saliva; †, Data partially published in Vaccines, Special Issue: West Nile Virus Disease (Holicki et al. 2020a); *, one legs-plus-wings sample could not be used in the WNV tests.
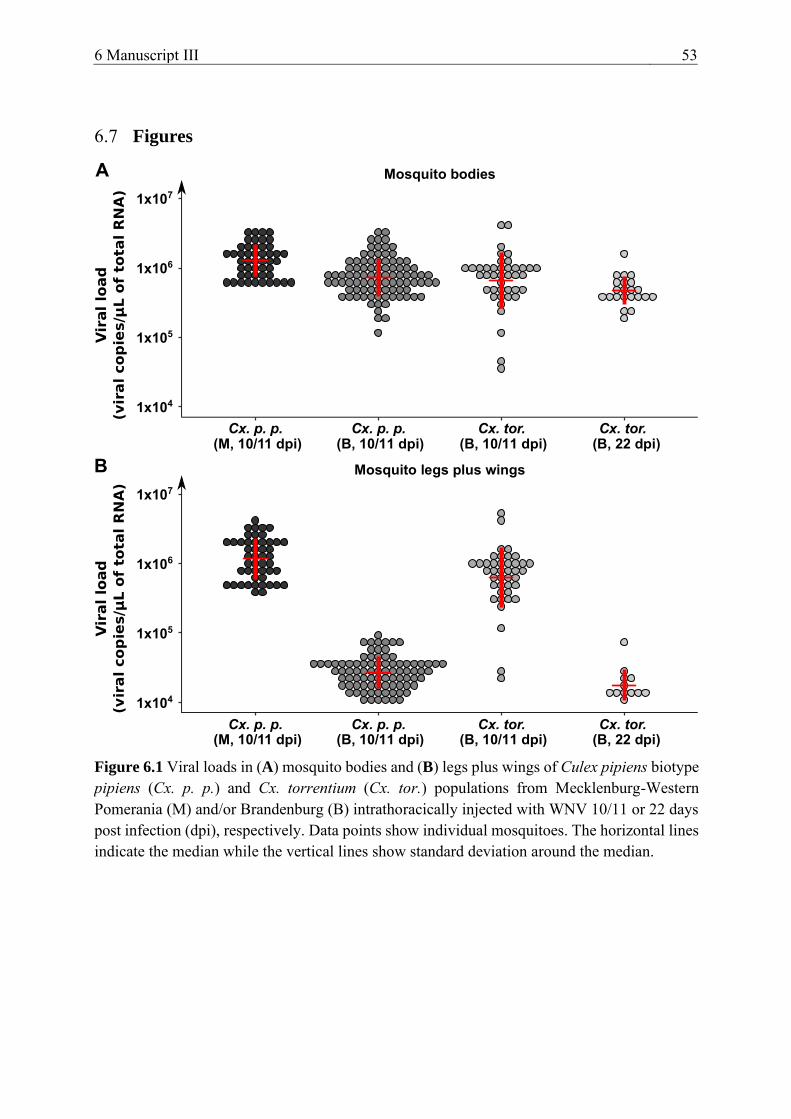
6 Manuscript III 53
6.7 Figures
Figure 6.1 Viral loads in (A) mosquito bodies and (B) legs plus wings of Culex pipiens biotype pipiens (Cx. p. p.) and Cx. torrentium (Cx. tor.) populations from Mecklenburg-Western Pomerania (M) and/or Brandenburg (B) intrathoracically injected with WNV 10/11 or 22 days post infection (dpi), respectively. Data points show individual mosquitoes. The horizontal lines indicate the median while the vertical lines show standard deviation around the median.

54 6.8 Acknowledgments
6.8 Acknowledgments
We thank Oliver Tauchmann, Cornelia Steffen, Friederike Michel, Stephan Löwe, and Dorothee E. Scheuch (FLI) for their excellent technical assistance in the laboratory. Furthermore, we want to thank René Klein, Marlene Hausner, and Maxi Uecker (FLI) for mosquito rearing and identification. A big thank you goes to Ariel Viña-Rodríguez (FLI) for supplying the synthetic WNV-RNA for the standard curve. We are thankful to Anna Heitmann and Stephanie Jansen (Bernhard-Nocht-Institut (BNITM), Hamburg, Germany) for their advice in designing the study. Lastly, we are grateful to Susanne Fischer (FLI) for assisting in the statistical analysis of the data.
This research was funded by the German Federal Ministry of Food and Agriculture (BMEL) through the Federal Office for Agriculture and Food (BLE), grant numbers 2819104915, 2819104615, and 2818SE001.

7 Manuscript IV 55
7 MANUSCRIPT IV
Pathogenicity of West Nile Virus Lineage 1 to German Poultry
Cora M. Holicki 1, Friederike Michel 1, Ana Vasić 2, Christine Fast 1, Martin Eiden 1,
Cristian Răileanu 2, Helge Kampen 2, Doreen Werner 3, Martin H. Groschup 1, and Ute
Ziegler 1,*
1 Institute of Novel and Emerging Infectious Diseases, Federal Research Institute for
Animal Health, Friedrich-Loeffler-Institut, 17493 Greifswald-Insel Riems, Germany 2 Institute of Infectology, Friedrich-Loeffler-Institut, Federal Research Institute for Animal
Health, 17493 Greifswald-Insel Riems, Germany 3 Biodiversity of Aquatic and Semiaquatic Landscape Features, Leibniz-Centre for
Agricultural Landscape Research, 15374 Muencheberg, Germany
Author to whom correspondence should be addressed: [email protected]
7.1 Abstract
West Nile virus (WNV) is a mosquito-borne virus that originates from Africa and at present causes neurological disease in birds, horses, and humans all around the globe. As West Nile fever is an important zoonosis, the role of free-ranging domestic poultry as a source of infection for humans should be evaluated. This study examined the pathogenicity of an Italian WNV lineage 1 strain for domestic poultry (chickens, ducks, and geese) held in Germany. All three species were subcutaneously injected with WNV, and the most susceptible species was also inoculated via mosquito bite. All species developed various degrees of viremia, viral shedding (oropharyngeal and cloacal), virus accumulation, and pathomorphological lesions. Geese were most susceptible, displaying the highest viremia levels. The tested waterfowl, geese, and especially ducks proved to be ideal sentinel species for WNV due to their high antibody levels and relatively low blood viral loads. None of the three poultry species can function as a reservoir/amplifying host for WNV, as their viremia levels most likely do not suffice to infect feeding mosquitoes. Due to the recent appearance of WNV in Germany, future pathogenicity studies should also include local virus strains.
Published in Vaccines in 2020
Volume 8, Issue 3 https://doi.org/10.3390/vaccines8030507


8 General Discussion 57
8 GENERAL DISCUSSION
Flaviviruses are a group of single-stranded RNA viruses, primarily transmitted by arthropods (e.g., mosquitoes and ticks), and causative agents of severe illnesses in humans and animals (Pierson and Diamond 2020). Since the last seven decades flaviviruses, such as DENV, WNV, and ZIKV, have become increasingly prevalent worldwide with detrimental epidemics on an annual basis (Pierson and Diamond 2020). These viruses have repeatedly proven their ability to rapidly spread throughout continents while causing considerable human morbidity (Pierson and Diamond 2018, Roehrig 2013). Flaviviruses also constitute important veterinary pathogens, infecting a broad spectrum of animals, including several domestic species. The (re-)emergence of flaviviruses around the globe, therefore, poses a constant threat to public as well as veterinary health (Mansfield et al. 2017, McLean et al. 2002, Pierson and Diamond 2020, Venter 2018) and the associated socio-economic impacts are possibly stagnating the development of countries struck by these viruses (Shepard et al. 2016, United Nations Development Programme (UNDP) 2017). Knowledge of the biology and ecology of flaviviruses as such is a cornerstone in understanding the epidemic potential of these viruses. Unique characteristics such as viral vector and host range and their geographical expansion in combination with fluctuating environmental settings and continuous globalization can substantially affect viral epidemiology (Tabachnick 2016, Young 2018).
The studies included in this thesis aimed to unravel the ecology of two closely related flaviviruses, WNV and Usutu virus (USUV), by investigating possible vector and hosts species. Both viruses have their origin in Africa (McIntosh 1985, Smithburn et al. 1940, Williams et al. 1964) and were dispersed around the globe via migratory birds (Engel et al. 2016, Owen et al. 2006). WNV was first isolated from a febrile patient in the West Nile district of Uganda in 1937 (Smithburn et al. 1940). Up until the early 1990s, the virus was only associated with sporadic outbreaks in Africa and Israel (Hayes 2001). Thereafter, new virus strains emerged in Russia (Platonov et al. 2011) and Southern and Eastern Europe (Bakonyi et al. 2006) with increasing disease severity in humans. In 1999, WNV reached the American continent, where it rapidly spread from the East to the West Coast of the USA (Ebel et al. 2001, Lanciotti et al. 1999). In Europe, the virus steadily expanded northwards, with the first detection in Germany in the summer of 2018 (Ziegler et al. 2019). Today, WNV is the most dispersed zoonotic arbovirus with detections in all continents, except Antarctica (Chancey et al. 2015). Usutu virus, on the other hand, was first isolated in 1959 in South Africa (McIntosh 1985, Williams et al. 1964) and spread throughout Africa (Ben Hassine et al. 2017, Durand et al. 2016, Mossel et al. 2017, Nikolay et al. 2011), Israel (Mannasse et al. 2017), and several European countries (Weissenböck et al. 2002). The first detection of USUV in Germany was in 2010 in mosquitoes (Jöst et al. 2011). Nowadays, the virus is receiving growing attention from the scientific community, primarily due to the speed at which the virus is expanding throughout Europe, the high toll it has on the avifauna (especially the Eurasian blackbird), and its increasing association to human infections (Roesch et al. 2019). For example, recent epidemics in 2016 and 2018

58 8 General Discussion
resulted in the expansion of USUV in northern and western Europe including multiple human deaths (Clé et al. 2019).
Both viruses belong to the Japanese encephalitis serocomplex and are associated with human diseases, e.g., (meningo-)encephalitis (Poidinger et al. 1996). They are maintained in a bird-mosquito-bird transmission cycle, where birds can act as amplifying/reservoir hosts and primarily Culex mosquitoes as vectors. Birds can develop mild to severe infections, ranging from “flu-like” clinical signs to a neuroinvasive disease with fatal outcome (Kramer et al. 2007). Wild avian species belonging to the orders Passeriformes, Falconiformes, and Strigiformes appear highly susceptible to both viruses and the overlap in susceptible bird species is considerable: up to 34 species from 11 different orders (Nikolay 2015). WNV infections of highly susceptible Corvidae such as crows or blue jays often lead to fatal diseases, while abundant passerine species such as common grackles are likely to act as virus reservoirs in certain areas, developing high viremia levels without necessarily succumbing to the infection (Komar et al. 2003). USUV, on the other hand, mainly affects blackbirds, magpies, and great grey owls, which can act as amplifying hosts (Clé et al. 2019). Both viruses can infect a variety of other mammalian species without causing any clinical signs. However, horses and humans are considered dead-end hosts as they do not produce viremia levels high enough to infect feeding mosquitoes and, thereby, cannot perpetuate the transmission cycles (Chancey et al. 2015). Nevertheless, they can develop a severe and fatal neuroinvasive disease (Bunning et al. 2002, Murgue et al. 2001b, Nash et al. 2001, Ostlund et al. 2001, Petersen and Marfin 2002).
WNV as well as USUV are predominantly transmitted by Culex mosquito species, with a few exceptions. Four European mosquito species have proven their vector competence for WNV: Cx. pipiens, Cx. modestus, Ae. albopictus, and Ae. detritus (Balenghien et al. 2007, Blagrove et al. 2016, Fortuna et al. 2015a, Fortuna et al. 2015b). USUV has been detected in Europe in several different mosquito species from five different genera (Aedes, Anopheles, Culex, Culiseta, and Ochlerotatus) (summarized by Clé et al. 2019, Nikolay 2015), of which the vector competence for USUV has been verified in the European Cx. pipiens species (Fros et al. 2015). This species can justly be considered the key vector for WNV and USUV in Europe (Fros et al. 2015). Vector competence is defined as the ability of a mosquito species to become infected after feeding from an infected host and to ultimately transmit the virus in its saliva to a new host (Kenney and Brault 2014). This ability strongly depends on the mosquito species and the viral lineage and strain. Vectorial capacity in turn is influenced by a mosquito’s vector competence, its abundance, its feeding behavior, its survival rate, and its environmental setting (Garrett-Jones 1964). Assessing the vector competence of specific mosquito species for a virus strain is the first step in identifying species that might act as important vectors in the field and, thereby, facilitate the implementation of evidence-based vector control measures (Kenney and Brault 2014).
Therefore, the first two manuscripts stated in this thesis (manuscript I and II) aimed to reveal indigenous mosquito species involved in the transmission cycle of WNV and USUV in

8 General Discussion 59
Germany. For this purpose, vector competence studies were performed with circulating German WNV and USUV virus strains and ubiquitous German mosquito populations (Cx. pipiens biotype pipiens, Cx. pipiens biotype molestus, and Cx. torrentium). Experiments were performed in a BSL-3 insectary where female mosquitoes (age: 3-14 days) were allowed to feed on a virus-spiked blood meal for a given timeframe. Thereafter, fully-engorged females were incubated at 18 °C, 25 °C, or 28 °C for two or three weeks until the saliva assays were conducted. Mosquito bodies, legs plus wings, and saliva samples were tested on their respective WNV or USUV content via a virus-specific RT-qPCR (Eiden et al. 2010, Jöst et al. 2011) or a cell culture system.
Regarding WNV (manuscript II), Cx. pipiens (both biotype pipiens and molestus) had the highest transmission efficiencies. Multiple mosquitoes from the two Cx. pipiens biotype pipiens populations collected in Brandenburg, Germany as well as from the one Cx. pipiens biotype molestus colony from northwestern Germany (Schleswig-Holstein) proved to be vector-competent for WNV lineage 2. However, of the two biotypes, mosquitoes of the biotype pipiens transmitted WNV in their saliva more frequently. Mosquitoes of the same Cx. pipiens biotype molestus colony could also transmit USUV (manuscript I), interestingly even to a higher percentage than for WNV (transmission efficiencies of 37.5%-50.0% compared to 6.7%-28.6%). For both viruses the vector competence indices were similar to those of mosquitoes from an additionally tested southern European (Serbian) Cx. pipiens biotype molestus colony. Furthermore, the infection studies discussed in this thesis also suggest for the first time the vector competence of German Cx. torrentium for USUV. An individual female of this species could transmit infectious USUV Africa 2 in its saliva three weeks post infection. Unfortunately, Cx. torrentium mosquitoes are very difficult to breed artificially. Therefore, experiments were always performed with females emerging from field-collected egg rafts, making it very difficult to obtain a large sample size of the exact same population within a given age span. Nonetheless, these results support the presumption that Cx. torrentium play a role in the transmission of USUV in Germany (Jansen et al. 2019). Culex torrentium (Martini 1925) and Cx. pipiens s.l. (Linnaeus 1758) are the most common mosquito species in Europe (Hesson et al. 2014) and can exist in sympatry in central Europe, including Germany (Lühken et al. 2015, Rudolf et al. 2013). In the past, both species were considered purely ornithophilic but only recent studies have described a certain variability in their host feeding preferences including also other mammals such as humans (Börstler et al. 2016). They might, therefore, also transmit WNV and USUV from infected birds to humans and should be considered possible bridge vectors, posing a threat to public health (Jansen et al. 2019). The same is true for Cx. pipiens hybrids of the biotypes pipiens and molestus (Chevillon et al. 1995, Ciota et al. 2013).
Not only indigenous mosquito species in Germany pose a threat to animal and human health. International trade, in combination with climatic and environmental changes, has increased the likelihood that invasive mosquito species, such as Ae. albopictus or Ae. japonicus, are introduced into and expand throughout Europe (Medlock et al. 2012). They are currently widely

60 8 General Discussion
established now in southern and central Europe (ECDC 2020). Ongoing studies with invasive mosquito species are analyzing their vector status for multiple arboviruses and, thereby, assessing whether they represent a considerable threat to the public health (Abbo et al. 2020a, Brustolin et al. 2018, Brustolin et al. 2016, Huber et al. 2014, Wagner et al. 2018). The Asian bush mosquito, Ae. japonicus, has proven its successful laboratory transmission for multiple flaviviruses including WNV, USUV, JEV, and ZIKV (Abbo et al. 2020a, Kampen and Werner 2014). This species is native to East Asia and was detected for the first time in Europe, in France, in 2000 (Schaffner et al. 2003). In Germany this species is now well established, with several populations in multiple federal states (Kampen and Werner 2014). The Asian tiger mosquito, Ae. albopictus, originated in Southeast Asia and is nowadays the most invasive mosquito species in the world. This was substantially facilitated by the international trade of tires, the import of agricultural products (such as “lucky bamboo”), and the transit of mosquitoes in public and private vehicles. This species is able to adapt to quite different climatic conditions found in temperate regions of the world as they can, for example, lay cold-resistant eggs (Benedict et al. 2007, Medlock et al. 2012). Aedes albopictus have been implicated as vectors for several zoonotic viruses such as the chikungunya virus (CHIKV), DENV, Eastern equine encephalitis virus, La crosse virus, and JEV (reviewed by Medlock et al. 2012). This work (manuscript II) demonstrated that an established German population of Ae. albopictus is vector-competent for a German WNV lineage 2 strain, even if to a significantly lower extent than the tested Culex species. Aedes albopictus are, not without reason, considered opportunistic feeders and serious biting nuisances. They can, therefore, become serious health threats in regions where they are well-established, usually close to human settlements (Benedict et al. 2007). Further infection studies with mosquito populations from the USA (Holick et al. 2002, Sardelis et al. 2002) or Europe (Fortuna et al. 2015b) came to similar conclusions.
Another potentially invasive mosquito species with a possible impact on European public health is the yellow fever mosquito, Ae. aegypti. To date it is found in tropical and subtropical regions of the Americas, Africa, and Asia, in addition to southeastern USA and northern Australia (Benedict et al. 2007). As most invasive mosquito species, it is referred to as an urban vector with adaptation to urban environments and a propensity to feed on multiple human blood sources, thereby, increasing disease transmission potential (Benedict et al. 2007). Major outbreaks of DENV (Rosen 1986, Wilder-Smith et al. 2010), CHIKV (Gould and Higgs 2009), and YFV (Chastel 1999) in numerous countries have been related to Ae. aegypti. This species, however, has only occasionally been reported in Europe, such as in France (Reiter 2010), Germany (Kampen et al. 2016), Madeira (Almeida et al. 2007), and the Netherlands (Scholte et al. 2010). It is unlikely that it will become established in central/northern European countries with temperate climate conditions as it, unlike Ae. albopictus, cannot overwinter as an adult (Christophers 1960). Nonetheless, due to climate change, the establishment of Ae. aegypti in Europe cannot be completely excluded. Therefore, in the frame of these experiments (manuscript I), a Malaysian Ae. aegypti colony was checked for its vector competence for USUV. Even though USUV-specific RNA could be isolated from their bodies after the

8 General Discussion 61
incubation period, none of the mosquitoes expectorated viable virus in their saliva. For this reason, Ae. aegypti, unlike Ae. japonicus, will probably not play a role in the dispersal of USUV in Germany in the future.
There are multiple conceivable reasons as to why the vector competence varies between mosquito species, biotypes, and/or populations. For example, interspecies and intraspecies genetics can influence the effectiveness of a mosquito’s midgut and salivary gland barriers as well as its antiviral immune response. Furthermore, aspects such as incubation time, incubation temperature, virus strain, and virus dose can also affect virus prevalence in the vector (Vogels et al. 2017c). As part of this work (manuscript II), the temperature dependency of vector competence was examined for WNV. For this the Cx. pipiens biotype pipiens population from Brandenburg was infected with WNV lineage 2 and incubated not only at 25 °C but also at 18 °C and 28 °C. Even though mosquito longevity appeared to be negatively affected by increasing temperatures, transmission efficiency was significantly higher at 25 °C and 28 °C than at 18 °C, especially after a longer incubation period. Within certain boundaries, temperature is directly positively correlated to virus replication rates and also to virus dissemination in ectothermic species (Ewing et al. 2016, Vogels et al. 2017c). Temperature can also have varying influences on a mosquito’s microbiome and innate immune system (Murdock et al. 2012). The results presented in this thesis correlate with those of other studies (summarized by Ciota and Keyel 2019). For example, the vector competence of Dutch and Italian Cx. pipiens increased when the temperature increased from 18 °C to 23 °C. However, the change was less pronounced from 23 °C to 28 °C (Vogels et al. 2017b), which is similar to the non-significant differences between the vector competences of the German Cx. pipiens at 25 °C and 28 °C. These results indicate a thermal limit of the temperature dependency. Remarkably though, individual Cx. pipiens mosquitoes were still capable of transmitting WNV after an incubation temperature of only 18 °C, being close to the average summer temperatures found in Germany (DWD 2018a). It is, therefore, justified to say that not only will climate change (i.e., increasing temperatures) shift arboviruses and their vectors polewards (Campbell-Lendrum et al. 2015) but that it is also likely that favorable conditions (i.e., susceptible vectors and hosts) are already present for the quick establishment of virus transmission cycles in new ecological niches in central and northern Europe.
The viral dose can also greatly affect the vector competence of mosquitoes. This was tested in this work (manuscript I) for indigenous Cx. pipiens biotype molestus and a German USUV isolate. Mosquitoes were fed with a blood meal spiked either with a high (107.4 TCID50/mL) or low (105.1 TCID50/mL) viral dose. Transmission rates of 100% and 75% were recorded 14 and 21 dpi, respectively after infection with a high viral dose compared to a complete cessation of virus dissemination and transmission via saliva after infection with a low virus dose. Similar findings were already described for Cx. quinquefasciatus (Anderson et al. 2010) and Cx. nigripalpus (Richards et al. 2011) infected with differing WNV doses. This dose dependency, however, seems to be connected to the genetic diversity of mosquitoes, as one Cx. torrentium

62 8 General Discussion
individual from the performed experiments could transmit virus in its saliva even after infection with the low titer. It is hypothesized that variations in virus dose affect the rate of midgut escape and salivary gland invasion. Increases in viral dose consequently increase the probability of mosquitoes becoming infected and developing a disseminated infection by facilitating primarily the escape from the midgut tissue barrier (Tesla et al. 2018). Mathematical models with ZIKV have shown that if mosquito populations feed on increasing viral doses (i.e., higher viremia levels) there will probably be more infectious bites causing more secondary ZIKV infections (Tesla et al. 2018). The same could be assumed for WNV and USUV circulation. Unlike the incubation temperature, the viral dose did not appear to influence mosquito longevity.
There are further parameters that can influence the vector competence for arboviruses: the incubation time, the viral strain, and intrinsic differences between mosquito species, biotypes, or distinct geographical populations (Vogels et al. 2017c). In this work (manuscript I and II) the differences in vector competence were strikingly high between Culex species and Aedes species for WNV and USUV. Furthermore, Cx. pipiens biotype pipiens had a greater vector competence for WNV than biotype molestus. However, no significant difference could be observed between different populations of the same species, as, for example, with the two Cx. pipiens biotype pipiens populations from Brandenburg for WNV.
The innate immune responses can play a leading role in the effectiveness of the midgut and salivary gland barriers for WNV (Vogels et al. 2017c) and it is probable that similar findings are true for USUV (Fros et al. 2015). Mosquito antiviral responses include RNAi, JAK/STAT, Toll, IMD, and lastly, apoptosis pathways (Prasad et al. 2013, Vogels et al. 2017c). The ingestion of an infectious blood meal can upregulate or activate the expression of antiviral genes (Vogels et al. 2017c). Their relevance is strongly dependent on the mosquito species and on the involved arbovirus. Studies with Cx. quinquefasciatus have confirmed that the activation of RNAi response leads to the destruction of WNV genomic RNA in the midgut (Brackney et al. 2009). Interestingly RNAi suppressors exist as countermeasures in the WNV genome, which can suppress the RNAi response for instance of Cx. pipiens and are, therefore, essential in overcoming the midgut barrier (Abbo et al. 2020a). Furthermore, the JAK/STAT pathway resembles the mammalian interferon response and can also suppress virus replication and cause cells to enter an antiviral state (Paradkar et al. 2012). The involvement of this pathway has been described for Ae. aegypti and Cx. quinquefasciatus for WNV (Bartholomay et al. 2010, Colpitts et al. 2011, Paradkar et al. 2012). However, the relevance of Toll and IMD pathway in the restriction of WNV in Culex mosquitoes is not fully understood (Bartholomay et al. 2010, Colpitts et al. 2011). Finally, WNV infection can cause apoptosis for self-preservation in the midgut and salivary gland epithelial cells of Culex species (Girard et al. 2010, Girard et al. 2007, Vaidyanathan and Scott 2006). Virus-induced apoptosis can, however, also facilitate virus transmission by releasing virus into the saliva (Vogels et al. 2017c).
Further factors influencing vector competence that were not examined as a part of this thesis include, for example the mosquito’s microbiome. This consists of bacterial and fungal

8 General Discussion 63
symbionts in the gut which impact the mosquito’s metabolism along with its pathogen interactions. This microbiome can negatively or positively affect infection and replication of flaviviruses (Jupatanakul et al. 2014). For example, endosymbionts, such as Wolbachia, can compete with arboviruses for nutrients and the cellular machinery required for replication (Jupatanakul et al. 2014). Unique bacteria species can even secrete anti-viral compounds (Běhal 2001). Data also propose the importance of bacteria in the activation of anti-pathogen immune responses and in immune priming (Dennison et al. 2014). Paradoxically though, Wolbachia has shown to enhance WNV infection in Cx. tarsalis through the downregulation of the Toll pathway-related genes (Dodson et al. 2014). The same, however, could not be verified for Cx. quinquefasciatus due to an insufficient density of Wolbachia (Micieli and Glaser 2014).
Mosquito infection studies can also be performed by infecting mosquitoes via the intrathoracic injection of a virus rather than through the ingestion of a virus-spiked blood meal. Even though this inoculation method does not mimic a natural infection it can aid in uncovering the relevance of mosquito escape barriers as this technique bypasses the midgut barrier. Furthermore, it is a helpful prerequisite for the implementation of mosquito-to-vertebrate infection models. Hence, in preparation for the poultry pathogenicity studies, two indigenous Cx. pipiens biotype pipiens laboratory populations and one indigenous Cx. torrentium field-derived population were injected intrathoracically with 500 TCID50 of an Italian WNV lineage 1 strain (manuscript III). Saliva assays were performed 10/11 or 22 dpi and vector competence indices were calculated. As might be expected, the infection and dissemination rates consistently reached 100% independent of the species and population while approximately every second mosquito transmitted WNV in its saliva 10/11 dpi. Only after a longer incubation period (i.e., 22 days) did the transmission rate of the Cx. torrentium increase up to 81.8%. In combination with the vector competence studies in manuscript II, these results give a clear indication for the greater relevance of the midgut rather than the salivary gland barrier on the vector competence of mosquitoes for flaviviruses, in this case WNV.
After identifying possible vector species for circulating WNV and USUV strains in Germany the last part of this work (manuscript IV) aimed to assess the relevance of domestic poultry as amplifying or reservoir hosts for flaviviruses, such as for WNV. The first step was to compare the susceptibility of indigenous juvenile chickens, ducks, and geese for WNV after subcutaneous injection. Birds from the species considered most susceptible to WNV, in this case the geese, were then also inoculated via the bite of infected mosquitoes. Eight 3.5-week-old chickens, ducks, and geese were inoculated under BSL-3 conditions with an Italian WNV lineage 1 strain; the same strain with which the intrathoracic injections had been performed (manuscript III). An Italian strain was used, as at that time WNV had not been isolated in Germany and it was presumed that WNV would enter from the South and not from the East (the Czech Republic) (Ziegler et al. 2019). Furthermore, the vector competence of German Cx. pipiens biotype pipiens for this strain had already been proven (Jansen et al. 2019). The infected birds and four control birds were kept for 3 weeks post infection and monitored for degree and

64 8 General Discussion
duration of clinical signs, viremia, viral shedding (oropharyngeal and cloacal), and seroconversion. Necropsy focused on macroscopic and microscopic pathological lesions as well as tissue tropism and IHC.
The majority of the birds did not develop any clinical signs, the only exceptions were two geese in their third week post infection. They displayed unspecific signs comprising lethargy, reduced appetite, body weight loss, impaired movement, skewed body posture, temporary muscle tremor, and ruffled/dirty plumage. Both geese had to be euthanized shortly before the planned termination of the experiment. Furthermore, one duck succumbed unexpectedly 2 dpi, without a clinically apparent disease. Throughout the experiments all three species manifested various degrees of viremia, viral shedding, seroconversion, tissue tropism, and (histo-)pathological lesions. In all birds viremia was detected from 2 dpi, which peaked already 2 dpi (chickens and geese) or 3 dpi (ducks). Other studies have described a similar time frame for peak viremia (i.e., 2-3 dpi) in passeriform species such as crows and jays (Weingartl et al. 2004) while in birds of prey it was slightly later (4-6 dpi) (Nemeth et al. 2006b, Ziegler et al. 2013). The viremia levels were the highest in the geese, followed by the ducks and lowest in the chickens, where the values were close to the detection limit. Compared to the other poultry species, the duration of viremia was also considerably longer in the geese with WNV-specific RNA being detected in the blood up to 10 dpi. For all three species viremia levels were, however, fairly low, especially when comparing them to highly susceptible species such as house sparrows (Langevin et al. 2005). Viral shedding was also observed in all three species from 2-5 dpi. Strikingly, the geese and the chickens shed the virus primarily through the oropharynx while in the case of the ducks the virus was detected more often in the cloacal swabs. Lastly, the systemic infection of birds activates the innate and adaptive immune response. The latter includes B- and T-cell activation with antibody production (Samuel and Diamond 2006, Suthar et al. 2013). In the pathogenicity studies all birds seroconverted with the highest antibody titers recorded in the ducks.
There are no pathognomonic macroscopic lesions for WNV as the virus poses a varied cellular and tissue tropism, differing greatly between species (Gamino and Höfle 2013). Species with a low susceptibility usually do not have any observable macroscopic lesions (Senne et al. 2000), as was the case in this study for the chickens. Birds that are more susceptible but usually survive an infection, such as birds of prey, can have more pronounced macroscopic lesions (emaciation, dehydration, multiorgan hemorrhages, spleno-/hepatomegaly, myocardial pallor, cerebral atrophy, etc.) (Lopes et al. 2007, Nemeth et al. 2006a, Steele et al. 2000, Wünschmann et al. 2005). In the studies referred to in this thesis, the majority of the ducks and a few of the geese developed myocardial pallor. All of the geese, most of the ducks and even individual chickens developed a weak/mild non-suppurative encephalitis. A common trait in all of these species was the detection of mild leucocytic infiltrations and degenerations within the perivascular cuffing. Such lesions in the brain are indicative of a viral entrance into the CNS via the bloodstream or through infected immune cells (Gamino and Höfle 2013, Weingartl et al. 2004). However, a distinct necrotizing component, as commonly described for WNV in other studies

8 General Discussion 65
(Byas and Ebel 2020, Feyer et al. 2020, Gamino and Höfle 2013, Senne et al. 2000, Ziegler et al. 2013), was not present in the majority of the birds. By contrast, the alterations seen in the spleen provided evidence for the aforementioned high variability of WNV infections. One chicken and two ducks showed a mild follicular hyperplasia of the spleen, even though this pattern is usually detected in geese. In addition to this, individual ducks and geese revealed necrotic lesions in the spleen, to some extent associated with vascular alterations. The heart is an organ often affected by WNV infections, yet in this study clear associations to a viral infection (i.e., necrotic lesions) could only be observed in a few of the ducks. All of the remaining birds revealed an unspecific mild non-suppurative interstitial myocarditis, the cause of which could not be clarified as it was also observed in one of the control chickens. Even though the IHC could not detect WNV antigen in the organ samples, the histopathological lesions in all of the birds were most probably associated to the viral infection. In two geese there were even indications for a persistent infection, as they showed clinical signs and necrosis in the spleen long after the cessation of viremia and viral shedding. Necrosis of the spleen is usually an indication for an acute infection (Gamino and Höfle 2013), as was also the case in the one duck that succumbed 2 dpi. Knowledge about the major target organs of WNV can not only improve our understanding of the immunocompetence of different species to WNV and the (neuro-)pathogenesis of the virus but also accelerate dead-bird surveillance programs.
Even though juvenile chickens, ducks, and geese were susceptible to the Italian WNV lineage 1 strain, their infection profiles were relatively weak compared to that of passeriform or corvid species. Due to their low and brief viremic phases they most likely do not constitute amplifying/reservoir hosts for the vector-borne sylvatic cycle of WNV in the field. A minimum titer of 106 PFU/mL has been declared a prerequisite to infect naïve mosquitoes and, thereby, perpetuate the transmission cycle (van der Meulen et al. 2005). Nonetheless, poultry species can still function as transmitters for WNV via direct contact between birds. Studies have already confirmed this for chickens (Langevin et al. 2001) and geese (Banet-Noach et al. 2003). There are multiple explanations for the in-contact transmission of WNV between birds: through the inhalation of infectious aerosols secreted with bodily fluids (i.e., through viral shedding), the ingestion of infected food and water, and the contact with viremic blood or infected organs (Pérez-Ramírez et al. 2014). The latter is most probable and can occur through cannibalism and feather picking (Banet-Noach et al. 2003, Langevin et al. 2001). Even though animals can become infected with WNV via infectious aerosols (Nir et al. 1965) the virus quantities shed via the oropharynx and cloaca are very low and WNV is also not very stable outside of the host (Langevin et al. 2001). In the here listed pathogenicity studies WNV-specific RNA was detected at the injection site of one duck 2 dpi, therefore, confirming that the skin constitutes a possible virus source. Interestingly, one goose showed delayed viremia and viral shedding which would fit into the picture of direct virus transmission. Yet, due to the also very likely intraspecies differences in susceptibility, further studies should be conducted collecting feather samples as well as additionally housing negative in-contact birds.

66 8 General Discussion
As chickens, ducks, and geese were uniformly susceptible to WNV and most likely do not play an essential role in the pathogen transmission cycle they are ideal candidates for sentinel bird-based surveillance of WNV. They rapidly seroconvert and in the majority of cases do not develop a clinical disease. In the past, alive and dead, captive and free-ranging birds have been used to monitor WNV transmission seasons and predict epizootic/epidemic outbreaks (Komar 2001). There are numerous benefits in using waterfowl and poultry flocks kept on backyard premises as sentinel species for avian diseases. Firstly, many influencing factors can be standardized, such as the age of the birds and their initial serological status, and they can easily be maintained in large flocks in multiple geographical areas, in close contact to urban settlements as well as with migrating birds. Sampling of poultry species is also less expensive than that of wild birds (Coman et al. 2014, European Commission 2003, Globig et al. 2009, Halliday et al. 2007, Komar 2001). For example, ducks and geese have been used for the active surveillance of AI (Coman et al. 2014, Globig et al. 2009) and chickens were usually the bird of choice for the monitoring of WNV. In the past, chickens have been used in numerous countries (e.g., Australia (Russell 1998), Greece (Chaintoutis et al. 2016), Romania (Cernescu et al. 2000), South Africa (McIntosh et al. 1967), and USA (Cherry et al. 2001, Komar et al. 2001, Marfin et al. 2001). However, their usage was not quite as successful as predicted with antibody detections only late in the transmission season and unfortunately often after onset of human disease (Komar 2001). From the pathogenicity studies with domestic poultry (manuscript IV) it can be concluded that, the usage of ducks as sentinels could be a good alternative. In the experiments they seroconverted faster and produced higher antibody titers with only weak viremia and minimal viral shedding. They are often kept in backyard, free-ranging flocks throughout the summer and as such are readily exposed to vector-competent mosquitoes (Komar 2001). A study analyzing the host-feeding patterns of mosquito species in Iran, even identified ducks as a preferred blood source (Shahhosseini et al. 2018) and domestic ducks held in the wetlands of Iran produced WNV-specific antibodies when WNV strains were circulating in the vicinity (Ziegler et al. 2010).
The last study within this work (manuscript IV) was conducted not only to establish a mosquito-to-bird model for WNV, but also to investigate mosquito saliva-induced modulation of WNV pathogenesis in birds. Eight juvenile geese were exposed to intrathoracically infected Cx. pipiens biotype pipiens mosquitoes. The intrathoracic injection technique for Culex species was established beforehand (manuscript III). Four geese infected via mosquito bite displayed similar infection profiles as those injected subcutaneously, with slightly higher viremia levels, delayed viral shedding, and seroconversion. The other four birds either were not bitten by the infectious mosquitoes or received an insufficient amount of WNV. The higher viremia levels indicate a mosquito induced enhancement of WNV pathogenesis, as was already discussed in other studies with mice and chickens (Schneider et al. 2006, Styer et al. 2006, Styer et al. 2011). Saliva contains pharmacologically active molecules that can modulate host hemostatic as well as immune responses (Fontaine et al. 2011, Ribeiro and Francischetti 2003, Titus et al. 2006). As the saliva composition differs between mosquito species (Ribeiro 2000), this study provides

8 General Discussion 67
the first indication of the effects of saliva from German Cx. pipiens biotype pipiens. Nonetheless, as viremia levels only differed marginally between the applied infection methods, other factors may have also influenced the results such as the viral load secreted by one or multiple mosquitoes and the probing/feeding time of the mosquitoes as well as the location of virus inoculation. Taking these factors into consideration it is very difficult to standardize mosquito-to-bird infection models. For example, in this study only one mosquito appeared blood-fed and it is, therefore, hypothesized that in three of the four geese the probing time was sufficient enough to infect the geese (i.e., engorgement of the mosquitoes is not necessary). The probing time is the time it takes from insertion of the mosquito’s mouthparts into the skin until engorgement begins (Das et al. 2010).
In summary, this work proved the vector competence of various German Cx. pipiens biotype pipiens and molestus populations for German USUV as well as WNV strains. The studies also demonstrated a temperature and viral dose dependency of mosquito-transmitted flaviviruses. To the greatest extent, the midgut escape barrier seems to play a role in the susceptibility of mosquitoes for WNV. Finally, domestic poultry was verified to be susceptible to a European WNV lineage 1 strain with geese developing the highest viremia levels and ducks the greatest antibody responses. A mosquito-to-bird infection model was also established with Cx. pipiens biotype pipiens mosquitoes from northern Germany and domestic geese. The study provided insight into the role of mosquito saliva in the pathogenesis of WNV. It could be concluded that chickens and ducks as well as geese probably do not play a vital role in the maintenance of WNV lineage 1 strain in the field as they constitute neither reservoir nor amplifying hosts but they, especially the ducks, are recommendable sentinel species.


9 Summary 69
9 SUMMARY
Mosquito-borne Flaviviruses: Vector and Avian Host
Susceptibility for West Nile Virus and Usutu Virus in Germany Cora Marielle Holicki
Flaviviruses are known to cause a variety of diseases in humans as well as animals, ranging from mild febrile illnesses to severe hemorrhagic or neuroinvasive manifestations. West Nile virus (WNV) and Usutu virus (USUV) are closely related flaviviruses circulating in an enzootic sylvatic cycle, consisting of mosquitoes (particularly Culex species) as vectors and birds as amplifying and/or reservoir hosts. Humans and equines, however, are considered dead-end hosts as they can develop severe clinical diseases but cannot in turn infect naïve feeding mosquitoes. Globalization, with increased international trade, in combination with climate change is facilitating the worldwide spread of vectors and their pathogens. To date WNV is the most dispersed zoonotic arbovirus, with detections in every continent aside Antarctica. Outbreaks are recorded yearly and the virus is becoming an increasing veterinary and public health concern. At the same time, USUV is also expanding its geographical range and colonizing new ecological niches.
Since the first isolation of USUV (2010) and WNV (2018) in Germany, both viruses have become endemic. They are overwintering either in infected mosquito females or in natural reservoir hosts with a yearly reemergence predominantly towards the end of the mosquito transmission seasons (from August to October). This has not only emphasized the necessity for evidence-based passive and active surveillance programs, but also for the completion of infection studies focusing on the pathogenesis of these viruses and the immunocompetence of their hosts. The experiments in the frame of this thesis, therefore, began to uncover vector species involved in the WNV and USUV transmission cycles in Germany and to tackle the question of whether domestic poultry could play a role as an amplifying and/or reservoir host for WNV.
In the first study (manuscript I) German Cx. pipiens biotype molestus and Cx. torrentium species proved to be vector-competent for an USUV Africa lineage 2 strain, isolated in Germany from a great grey owl (2015). Results were comparable to that of an established Serbian Cx. pipiens biotype molestus colony, while a Malaysian Ae. aegpyti colony could not transmit USUV in its saliva. To our knowledge, this is the first confirmation of Cx. torrentium transmitting USUV under laboratory settings. The transmission rate of the German Cx. pipiens biotype molestus colony was dependent on the viral titer in the blood meal. When the blood meal titer declined from 107.4 to 105.1 TCID50/mL the infection rate dropped and the mosquitoes

70 9 Summary
no longer developed a disseminated infection nor transmitted USUV in their saliva, neither after 14 nor 21 dpi.
With the first isolation of WNV in Germany in 2018, the question of possible vector species was no longer confined to USUV. The second study (manuscript II), therefore, comprised vector competence studies with indigenous mosquito species and the first recorded WNV lineage 2 strain isolated from a great grey owl in eastern Germany. Two populations of field-derived Cx. pipiens biotype pipiens from the northeast of Germany (Brandenburg) could readily transmit WNV in their saliva two and three weeks after feeding on an infectious blood meal. The same was true for a German Cx. pipiens biotype molestus colony from northern Germany (Lower Saxony). The transmission efficiency was, however, lower for biotype molestus than for biotype pipiens. The study also confirmed that the vector competence of Cx. pipiens biotype pipiens was dependent on the incubation temperature. The infection rates were fairly similar across all temperature gradients, yet the dissemination rates and transmission efficiencies were significantly greater at 25 °C and 28 °C compared to only 18 °C. Surprisingly, however, individual mosquitoes were still able to transmit WNV after incubation at 18 °C, an indication for the adaptation of the vector and the virus to temperate climates. An invasive Ae. albopictus population, established in central Germany (Thuringia), was also susceptible to WNV, but did not transmit infectious saliva to the same extent as the Culex species.
The third study (manuscript III) aimed to establish a method by which to successfully infect Culex species with WNV via intrathoracic injections. This procedure was not only an essential requirement for the mosquito-to-bird infection models, but also enabled an assessment of the functionality and importance of the midgut barrier for WNV in Culex species. The intrathoracic injection of Cx. pipiens biotype pipiens and Cx. torrentium with a European WNV lineage 1 strain guaranteed in both species a disseminated infection with approximately every second mosquito secreting infectious saliva 10/11 dpi. The results verified the significance of the midgut barrier, as it appears to limit initial virus replication in the midgut and subsequently also virus dissemination to secondary tissues.
Finally, the fourth study (manuscript IV) examined the interactions between susceptible vectors and hosts i.e., domestic poultry. It was designed to mimic the natural mosquito-to-bird transmission pathway of WNV and to compare subsequent disease pathogenesis to that of subcutaneously injected birds. In the first experiments, juvenile chickens, ducks, and geese were injected subcutaneously with a European WNV lineage 1 strain and monitored and sampled for three weeks post infection. The majority of the birds did not develop any clinical signs aside from one duck that succumbed 2 dpi (associated with technopathies) and two geese that possibly developed a persistent infection. They showed clinical manifestations only in the third week yet the pathological alterations (severe splenitis) were typical for an acute infection. Chickens were least susceptible, while ducks displayed the highest antibody levels and geese the highest viremia. Viral shedding was observed in all three species from 2 to 5 dpi, and was mainly via the oropharynx in chickens and geese and via the cloaca in ducks. Macroscopic

9 Summary 71
lesions were observed in the ducks and geese and included in particular pale and mottled appearance of the myocardium. All three species developed histological lesions, which are characteristic for a viral infection and were primarily detected in the hearts (myocarditis) and brains (encephalitis) of the ducks and the spleens (hyperplasia/splenitis) and brains (encephalitis) of the geese. As geese were most susceptible to WNV after its subcutaneous injection, this species was also used for the mosquito-to-bird infection model. For this purpose, field-derived Cx. pipiens biotype pipiens mosquitoes from northeast Germany (Brandenburg) were intrathoracically injected with the WNV lineage 1 strain and allowed to feed on juvenile geese 10 dpi. Four of the eight exposed geese developed viremia, shed virus, seroconverted, and exhibited pathological lesions. The geese infected via mosquito bite displayed higher viremia levels and more diverse pathological lesions than those injected subcutaneously, providing evidence for a mosquito saliva-induced modulation of WNV pathogenesis in geese.
In conclusion, these studies proved various Culex species to be vector-competent for USUV and/or WNV in Germany and verified that domestic poultry is unlikely to act as an amplifying or reservoir host for WNV lineage 1 in nature. Ducks and geese appear to be suitable sentinel species. They can monitor the spread of WNV and ideally detect WNV epidemics prior to the onset of human cases.


10 Zusammenfassung 73
10 ZUSAMMENFASSUNG
Stechmückenübertragene Flaviviren: Empfänglichkeit von
Vektoren und Wirten für West-Nil-Virus und Usutu-Virus in
Deutschland Cora Marielle Holicki
Flaviviren verursachen eine Vielzahl von Krankheiten in Menschen und Tieren. Die Bandbreite reicht von fieberhaften, grippeähnlichen Erkrankungen bis hin zu schweren hämorrhagischen und neuroinvasiven Formen. Das West-Nil-Virus (WNV) und das Usutu-Virus (USUV) sind engverwandte Flaviviren, die in einem enzootisch-sylvatischen Zyklus existieren, mit Stechmücken (insbesondere Culex-Arten) als Vektoren und Vögeln als Amplifikations-, beziehungsweise Reservoirwirten. Im Gegensatz dazu sind Menschen und Pferde Fehlwirte der Infektion, die selbst keine Virusquelle für Mücken darstellen, obwohl sie schwer erkranken können. Auf Grund der Globalisierung, insbesondere durch den verstärkten internationalen Handel und der Klimaerwärmung, ist es eine Leichtigkeit für Vektoren und Pathogene sich weltweit auszubreiten. WNV ist heute das am weitesten verbreitete zoonotische Arbovirus und zirkuliert auf allen Kontinenten außer der Antarktis. Jährlich wiederkehrende epidemische, beziehungsweise epizootische Ausbrüche stellen dabei eine Gefahr für die tierische und menschliche Gesundheit dar. Zeitgleich erobert auch das USUV stetig neue ökologische Nischen.
Beide Viren sind seit dem ersten Nachweis in Deutschland (USUV 2010; WNV 2018) auch hier in der Lage in infizierten Stechmückenweibchen sowie Reservoirwirten zu überwintern. Dadurch kommt es zu jährlich wiederkehrenden Ausbrüchen gegen Ende der Mückensaison (August bis Oktober). Dies unterstreicht die Notwendigkeit von evidenzbasierten, aktiven und passiven Überwachungsprogrammen sowie von gezielter Forschung zur Viruspathogenese in empfänglichen Wirten und deren Immunkompetenz. Diese Doktorarbeit befasst sich daher mit der Identifikation von vektorkompetenten Stechmücken für WNV und USUV in Deutschland und der Rolle des Wirtschaftsgeflügels im Übertragungszyklus von WNV.
In der ersten Studie (Manuskript I) konnten deutsche Cx. pipiens Biotyp molestus und Cx. torrentium Mücken als potentielle USUV-Vektoren bestätigt werden. Sie übertrugen erfolgreich mittels Speichel ein USUV-Afrika 2 Isolat, welches ursprünglich von einem Bartkauz (aus 2015) stammte. Diese Ergebnisse sind vergleichbar mit unserer etablierten serbischen Cx. pipiens Biotyp molestus Laborkolonie. Eine malaysische Ae. aegypti Laborkolonie erwies sich hingegen als vermindert empfänglich gegenüber dem Virus. Unseres Wissens nach ist dies der erste Nachweis, dass die Mückenart Cx. torrentium USUV unter Laborbedingungen übertragen kann. Zudem konnte eine hohe Korrelation zwischen der

74 10 Zusammenfassung
Transmissionsrate von Cx. pipiens Biotyp molestus und dem Virustiter in der Blutmahlzeit gezeigt werden. Eine Reduktion von 107.4 auf 105.1 Kulturinfektiöse Dosis 50 (KID50) pro mL führte, unabhängig von der Inkubationszeit, zu einem Absinken der Infektionsrate. Dabei ermöglichte der niedrigere Titer keine Virusdissemination in Sekundärgeweben wie den Speicheldrüsen.
Mit dem ersten Nachweis von WNV in Deutschland im Jahr 2018 stellte sich die Frage, welche Vektoren für die Ausbreitung und Etablierung von WNV eine Rolle spielen. Die zweite Studie (Manuskript II) beinhaltet daher Vektorkompetenzstudien mit einheimischen Stechmücken und einem WNV-Linie 2 Isolat, welches aus dem ersten in Deutschland registrierten WNV Vogel (Bartkauz) isoliert wurde. Zwei Cx. pipiens Biotyp pipiens Stechmückenpopulationen, gewonnen aus Eischiffchen aus dem Nordosten Deutschlands (Brandenburg), konnten nach Aufnahme einer infektiösen Blutmahlzeit mit ihrem Speichel WNV übertragen. Dies gilt auch für eine kürzlich etablierte Cx. pipiens Biotyp molestus Kolonie aus Norddeutschland (Niedersachsen). Die Transmissionseffizienz von Biotyp molestus war allerdings deutlich geringer als die vom Biotyp pipiens. Des Weiteren konnte die Temperaturabhängigkeit der Vektorkompetenz von Stechmücken bestätigt werden. Bei relativ konstanter Infektionsrate waren die Disseminationsraten und Transmissionseffizienzen bei 25 °C und 28 °C signifikant höher als im Vergleich zu 18 °C. Trotzdem konnten vereinzelte Stechmückenweibchen infektiösen Speichel auch nach einer zwei- bis dreiwöchigen Inkubation bei 18 °C absondern. Dies ist ein Indiz für die Adaptation vom Vektor und vom Virus an das hiesige Klima. Weiterhin erwies sich eine neuetablierte, invasive Ae. albopictus Population aus Mitteldeutschland (Thüringen) als weiterer möglicher Überträger von WNV, allerdings nicht in demselben Umfang wie die Culex-Arten.
Die dritte Studie (Manuskript III) diente der Etablierung von Infektionsmodellen basierend auf der intrathorakalen Injektion in einheimischen Culex-Arten mit WNV. Dies ist nicht nur ein wichtiger Baustein für die nachfolgende Studie, sondern ermöglichte auch die Beurteilung der Mitteldarmbarriere für die Virusausbreitung in den getesteten Mückenarten. Die intrathorakale Injektion gewährleistete eine disseminierte WNV-Infektion in Cx. pipiens Biotyp pipiens und Cx. torrentium Stechmückenweibchen, wobei ungefähr jede zweite Mücke nach 10 bis 11 Tagen infektiösen Speichel absonderte. Diese Ergebnisse bestätigen die Relevanz der Mitteldarmbarriere für die Minimierung der initialen Virusreplikation im Mitteldarm, sowie die Dissemination ins Sekundärgewebe.
Die letzte Studie (Manuskript IV) befasste sich mit der Etablierung eines Vektor-Wirts-Systems, um ein genaueres Verständnis der Interaktionen zwischen vektorkompetenten Stechmücken und Nutzgeflügelarten zu erarbeiten. Hierfür wurde der natürliche Infektionsweg des Virus über die Stechmücke in den Vogel nachgeahmt und anschließend die Pathogenese mit subkutan infiziertem Wirtschaftsgeflügel verglichen. Im ersten Teil wurden juvenile Hühner, Enten und Gänse mit einem Isolat der europäischen WNV-Linie 1 subkutan infiziert. Anschließend wurden die Vögel drei Wochen lang veterinärmedizinisch überwacht und

10 Zusammenfassung 75
beprobt. Fast alle Vögel zeigten dem Zeitraum über keinerlei klinische Symptomatik. Lediglich eine Ente erkrankte frühzeitig (Technopathie bedingt) und zwei Gänse zeigten unspezifische klinische Symptome zum Ende des Versuches hin, die zur Euthanasie führten. Die nachgewiesene hochgradige Splenitis, welche eher für eine akute virale Infektion spricht, lässt die Annahme zu, dass eine mögliche persistente WNV-Infektion vorliegen könnte, da zu diesem Zeitpunkt keine Virämie mehr nachweisbar war.
Vergleichsweise zeigten die Hühner die geringste Empfänglichkeit gegenüber WNV, während bei den Enten die stärkste Antikörperreaktion nachgewiesen wurde, sowie bei den Gänsen die stärkste Virämie. Bei allen drei Spezies erfolgte die Virusausscheidung vom zweiten bis zum fünften Tag nach der Infektion. Sie verlief bei den Hühnern und Gänsen vor allem oropharyngeal, bei den Enten vorranging kloakal. Abgesehen von den Hühnern konnten bei fast allen Vögeln makroskopische Aufhellungen an der Herzmuskulatur nachgewiesen werden. Bei allen drei Spezies deuteten die histologischen Läsionen auf eine virale Infektion hin. Bei den Enten war das Herz (Myokarditis) und das Gehirn (Enzephalitis) am stärksten betroffen und bei den Gänsen sowohl die Milz (Hyperplasie/Splenitis) als auch das Gehirn (Enzephalitis). Da sich die Gänse in diesem ersten Teilversuch am empfänglichsten erwiesen, wurde diese Spezies auch für das Vektor-Wirts-Modell verwendet. Zunächst wurden aus dem Nordosten Deutschlands stammende Cx. pipiens Biotyp pipiens Stechmücken intrathorakal mit WNV infiziert und nach einer Inkubationszeit von zehn Tagen an die Gänse gesetzt. Die Hälfte der exponierten Gänse zeigten eine Virämie, schieden Virus aus, serokonvertierten und entwickelten pathomorphologische Veränderungen. Interessanterweise entwickelten die Gänse nach einer Infektion durch Mückenstiche eine höhere Virämie und eine größere Bandbreite an pathologischen Läsionen als nach subkutaner Injektion mit WNV. Diese Ergebnisse bestätigen die Annahme, dass Mückenspeichel die Viruspathogenese positiv beeinflusst.
Die hier beschriebenen Studien zeigen, dass verschiedene Culex-Arten wichtige Vektoren in der Übertragung von USUV und WNV in Deutschland sind, sowie dass das Nutzgeflügel (Huhn, Ente, Gans) kein Amplifikations-/Reservoirwirt für WNV-Linie 1 darstellt. Trotzdem könnten Enten und Gänse in Freilandhaltung als Sentinel-Spezies genutzt werden, um sowohl die Ausbreitung von WNV in bestimmten Regionen zu überwachen, als auch Epidemien frühzeitig aufzuzeigen, möglichst noch bevor die ersten Humanfälle in neuen Gebieten auftreten.


Authors’ Contribution 77
AUTHORS’ CONTRIBUTION
Cora Marielle Holicki: Vector Competence Studies (Manuscripts I, II, and III) Establishment and optimization of vector competence studies with mosquitoes in the BSL-3 facility at the FLI for the first time. These included the infection of mosquitoes via intrathoracic injections as well as via a virus-spiked blood meal. Cultivation of the viruses beforehand to a high enough titer (>108 TCID50/mL) for the infection experiments. Preparation of the USUV and WNV standard for the quantification of viral RNA. Coordination and performance of numerous infection experiments with subsequent sole investigation of the collected samples (e.g., RNA extraction, RT-qPCR, and saliva assays under BSL-3 conditions). Data collection, data analysis, and data visualization with support from a statistician. Interpretation of the collected data and writing of the original drafts of the manuscripts. WNV Pathogenicity Studies in Chickens, Ducks, and Geese (Manuscript IV) Planning of the pathogenicity studies with help from published literature. Writing of the first draft of the application form for animal experiments according to German animal welfare legislation. Preparation and execution of the animal experiments under BSL-3 conditions (e.g., virus cultivation and titration, subcutaneous injection or mosquito bite inoculation of the birds, monitoring of their health status, blood and swab sample collection, euthanasia, and necropsy). Investigation of the generated samples (e.g., preparation of samples, RNA extraction, RT-qPCR). Assisting with the histopathological examinations (embedding of samples in paraffin, performing the immunohistochemical staining and subsequently evaluating slides). Data collection, data analysis, data visualization, data interpretation and writing of the original draft of the manuscript. Martin H. Groschup and Ute Ziegler supervised all the studies described in this thesis. Ute Ziegler, together with Friederike Michel, taught me BSL-3 experimentation skills and introduced me to classical virological methods as well as modern molecular diagnostic techniques such as RT-qPCR. In this context, Martin Eiden and Markus Keller gave me valuable technical advice especially concerning the implementation of a standard to quantify viral RNA loads via RT-qPCR. Dorothee E. Scheuch, Ute Ziegler, Helge Kampen, and later also Ana Vasić were key partners in setting up the protocols for the vector competence experiments in the BSL-3 insectary. Anna Heitmann and Stephanie Jansen (BNITM, Hamburg) also provided theoretical suggestions. Christine Fast introduced me to the IHC techniques and performed the histological examinations of the hematoxylin/eosin slides under the light microscope. Jana Schulz and Susanne Fischer assisted with the statistical analysis.


Acknowledgments 79
ACKNOWLEDGMENTS
Firstly, I would like to thank my academic supervisors Prof. Dr. Martin H. Groschup and Dr. Ute Ziegler for their consistent support and guidance throughout my dissertation. I am grateful to have been granted the opportunity to work at the Institute for Novel and Emerging Infectious Diseases of the Friedrich-Loeffler-Institut within the CuliFo project “Mosquitoes and mosquito-borne zoonoses in Germany”. After familiarization to laboratory techniques, I was allowed the freedom to implement and optimize protocols for the vector competence experiments with mosquitoes and BSL-2 and -3 viruses. I could put forward my own ideas and at the same time receive enough feedback and inspiration. I especially want to thank Dr. Ute Ziegler, for taking me under her wing and not only supporting me during the animal pathogenicity studies but also ensuring that I challenge my own work and that my projects progressed swiftly. She always found time for me when I needed technical advice or encouragement. Both Prof. Dr. Martin H. Groschup and Dr. Ute Ziegler played a considerable role in the revising of my thesis and its corresponding manuscripts.
My special gratitude goes to all those who spent long days together with me in the insectary. Dr. Dorothee E. Scheuch was not only my mentor in the first year but she also became a best friend of mine. Together we implemented the first vector competence experiments and from her I learned how to enjoy my work, never give up, and to learn from my mistakes. Dr. Ana Vasić supported me during my last year and due to her relentless perseverance we managed to successfully complete numerous vector competence studies. I admire her unbeatable optimism and her assurance in me. She was certain from the start that I would successfully complete my dissertation in time. I would not want to miss out on our friendship. I would also like to thank, Cristian Răileanu, Stephan Löwe, and Julia Lettow for temporarily assisting me with the mosquito vector competence studies.
Great thanks goes to Dr. Friederike Michel, who taught me all the necessary techniques required for the animal pathogenicity studies. She also supported me during the experiments in the stable as well as in the lab. I could always confine in her and she always offered me a helping hand, not only at the FLI.
I also want to thank Dr. Helge Kampen and Prof. Dr. Cornelia Silaghi for their entomological guidance and always providing me with enough mosquitoes for the studies. Thank you, Dr. Doreen Werner for always offering me field-collected egg rafts, I would never have imagined how grateful one would become to receive mosquito egg rafts.
It goes without saying how thankful I am to all my hard-working colleagues in the lab. Thank you, Cornelia Steffen, Katja Wittig, Gesine Kreplin, René Schöttner, and Oliver Tauchmann. I appreciate all the tender care René Klein and Marlene Hausner put into the rearing of the mosquitoes, ensuring I would have healthy and blood-thirsty mosquitoes for the studies. Without Dr. Susanne Fischer and Dr. Jana Schulz I would have never managed the statistical

80 Acknowledgments
evaluation of the data. The animal pathogenicity studies could not have been completed without the assistance of the animal care takers. I want to especially thank them for always being eager to jump into the full protective suits with me every day of the week.
I made a lot of great friends at the FLI and I want to expressively thank Dr. Veronika Dill. She was always there for me when I needed someone to share my thoughts with and she always had clever suggestions for work related as well as private obstacles. Thank you for being a loyal friend- together we could always laugh about our daily struggles and enjoy the small things in life.
A big thank you also goes to all my other friends and colleagues: Nicole Cichon, Dr. Benjamin Gutjahr, Julia Hartlaub, Rebecca König, Maren Penning, Dr. Melanie Rissmann, Dr. Ansgar Schulz, and Dr. Franziska Stoek.
Finally, I want to thank the most important people in my life, my family and my boyfriend. They accompanied me through the good but also the difficult times of my dissertation, especially in the final months. Thanks to their tolerance and patience, their relentless encouragement, and their unconditional faith in me I managed to also navigate the more emotionally challenging aspects of my dissertation.
I could not have made it through the last three years without all this support!

Bibliography 81
BIBLIOGRAPHY
Abbo, S. R.; Visser, T. M.; Wang, H.; Göertz, G. P.; Fros, J. J.; Abma-Henkens, M. H. C.;
Geertsema, C.; Vogels, C. B. F.; Koopmans, M. P. G.; Reusken, Cbem; Hall-Mendelin, S.; Hall, R. A.; van Oers, M. M.; Koenraadt, C. J. M.; Pijlman, G. P. The invasive Asian bush mosquito Aedes japonicus found in the Netherlands can experimentally transmit Zika virus and Usutu virus. PLoS Negl Trop Dis 2020a, 14 (4), e0008217, doi:10.1371/journal.pntd.0008217.
Abbo, S. R.; Vogels, C. B. F.; Visser, T. M.; Geertsema, C.; van Oers, M. M.; Koenraadt, C. J. M.; Pijlman, G. P. Forced Zika virus infection of Culex pipiens leads to limited virus accumulation in mosquito saliva. Viruses 2020b, 12 (6), 659, doi:10.3390/v12060659.
Aberle, S. W.; Kolodziejek, J.; Jungbauer, C.; Stiasny, K.; Aberle, J. H.; Zoufaly, A.; Hourfar, M. K.; Weidner, L.; Nowotny, N. Increase in human West Nile and Usutu virus infections, Austria, 2018. Euro Surveill 2018, 23 (43), pii=1800545, doi:10.2807/1560-7917.ES.2018.23.43.1800545.
Abutarbush, S. M.; O'Connor, B. P.; Clark, C.; Sampieri, F.; Naylor, J. M. Clinical West Nile virus infection in 2 horses in western Canada. Can Vet J 2004, 45 (4), 315-317.
Allering, L.; Jöst, H.; Emmerich, P.; Günther, S.; Lattwein, E.; Schmidt, M.; Seifried, E.; Sambri, V.; Hourfar, K.; Schmidt-Chanasit, J. Detection of Usutu virus infection in a healthy blood donor from south-west Germany, 2012. Euro Surveill 2012, 17 (50), pii=20341, doi:10.2807/esm.03.10.00094-fr.
Almeida, A. P.; Gonçalves, Y. M.; Novo, M. T.; Sousa, C. A.; Melim, M.; Gracio, A. J. Vector monitoring of Aedes aegypti in the autonomous region of Madeira, Portugal. Euro Surveill 2007, 12 (46), pii=3311, doi:10.2807/esw.12.46.03311-en.
Anderson, S. L.; Richards, S. L.; Tabachnick, W. J.; Smartt, C. T. Effects of West Nile virus dose and extrinsic incubation temperature on temporal progression of vector competence in Culex pipiens quinquefasciatus. J Am Mosq Control Assoc 2010, 26 (1), 103-107, doi:10.2987/09-5926.1.
Andreadis, T. G. The contribution of Culex pipiens complex mosquitoes to transmission and persistence of West Nile virus in North America. J Am Mosq Control Assoc 2012, 28 (4 Suppl), 137-151, doi:10.2987/8756-971X-28.4s.137.
Angenvoort, J.; Brault, A. C.; Bowen, R. A.; Groschup, M. H. West Nile viral infection of equids. Vet Microbiol 2013, 167 (1-2), 168-180, doi:10.1016/j.vetmic.2013.08.013.
Ariel, E. Viruses in reptiles. Vet Res 2011, 42, 100, doi:10.1186/1297-9716-42-100. Ashraf, U.; Ye, J.; Ruan, X.; Wan, S.; Zhu, B.; Cao, S. Usutu virus: An emerging flavivirus in
Europe. Viruses 2015, 7 (1), 219-238, doi:10.3390/v7010219. Bagnarelli, P.; Marinelli, K.; Trotta, D.; Monachetti, A.; Tavio, M.; Del Gobbo, R.;
Capobianchi, M.; Menzo, S.; Nicoletti, L.; Magurano, F.; Varaldo, P. Human case of autochthonous West Nile virus lineage 2 infection in Italy, September 2011. Euro Surveill 2011, 16 (43), pii=20002, doi:10.2807/ese.16.43.20002-en.
Bai, F.; Thompson, E. A.; Vig, P. J. S.; Leis, A. A. Current understanding of West Nile virus clinical manifestations, immune responses, neuroinvasion, and immunotherapeutic implications. Pathogens 2019, 8 (4), 193, doi:10.3390/pathogens8040193.

82 Bibliography
Bakonyi, T.; Erdélyi, K.; Ursu, K.; Ferenczi, E.; Csörgő, T.; Lussy, H.; Chvala, S.; Bukovsky, C.; Meister, T.; Weissenböck, H.; Nowotny, N. Emergence of Usutu virus in Hungary. J Clin Microbiol 2007, 45 (12), 3870-3874, doi:10.1128/JCM.01390-07.
Bakonyi, T.; Ferenczi, E.; Erdélyi, K.; Kutasi, O.; Csörgő, T.; Seidel, B.; Weissenböck, H.; Brugger, K.; Bán, E.; Nowotny, N. Explosive spread of a neuroinvasive lineage 2 West Nile virus in Central Europe, 2008/2009. Vet Microbiol 2013, 165 (1-2), 61-70, doi:10.1016/j.vetmic.2013.03.005.
Bakonyi, T.; Ivanics, E.; Erdélyi, K.; Ursu, K.; Ferenczi, E.; Weissenböck, H.; Nowotny, N. Lineage 1 and 2 strains of encephalitic West Nile virus, central Europe. Emerg Infect Dis 2006, 12 (4), 618-623, doi:10.3201/eid1204.051379.
Bakonyi, T.; Jungbauer, C.; Aberle, S. W.; Kolodziejek, J.; Dimmel, K.; Stiasny, K.; Allerberger, F.; Nowotny, N. Usutu virus infections among blood donors, Austria, July and August 2017 - Raising awareness for diagnostic challenges. Euro Surveill 2017, 22 (41), pii=17-00644, doi:10.2807/1560-7917.ES.2017.22.41.17-00644.
Balenghien, T.; Vazeille, M.; Reiter, P.; Schaffner, F.; Zeller, H.; Bicout, D. J. Evidence of laboratory vector competence of Culex modestus for West Nile virus. J Am Mosq Control Assoc 2007, 23 (2), 233-236,
doi:10.2987/8756-971X(2007)23[233:EOLVCO]2.0.CO;2. Banet-Noach, C.; Simanov, L.; Malkinson, M. Direct (non-vector) transmission of West Nile
virus in geese. Avian Pathol 2003, 32 (5), 489-494, doi:10.1080/0307945031000154080.
Barbic, L.; Vilibic-Cavlek, T.; Listes, E.; Stevanovic, V.; Gjenero-Margan, I.; Ljubin-Sternak, S.; Pem-Novosel, I.; Listes, I.; Mlinaric-Galinovic, G.; Di Gennaro, A.; Savini, G. Demonstration of Usutu virus antibodies in horses, Croatia. Vector Borne Zoonotic Dis 2013, 13 (10), 772-774, doi:10.1089/vbz.2012.1236.
Barrows, N. J.; Campos, R. K.; Liao, K. C.; Prasanth, K. R.; Soto-Acosta, R.; Yeh, S. C.; Schott-Lerner, G.; Pompon, J.; Sessions, O. M.; Bradrick, S. S.; Garcia-Blanco, M. A. Biochemistry and molecular biology of flaviviruses. Chem Rev 2018, 118 (8), 4448-4482, doi:10.1021/acs.chemrev.7b00719.
Bartholomay, L. C.; Waterhouse, R. M.; Mayhew, G. F.; Campbell, C. L.; Michel, K.; Zou, Z.; Ramirez, J. L.; Das, S.; Alvarez, K.; Arensburger, P.; Bryant, B.; Chapman, S. B.; Dong, Y.; Erickson, S. M.; Karunaratne, S. H.; Kokoza, V.; Kodira, C. D.; Pignatelli, P.; Shin, S. W.; Vanlandingham, D. L.; Atkinson, P. W.; Birren, B.; Christophides, G. K.; Clem, R. J.; Hemingway, J.; Higgs, S.; Megy, K.; Ranson, H.; Zdobnov, E. M.; Raikhel, A. S.; Christensen, B. M.; Dimopoulos, G.; Muskavitch, M. A. Pathogenomics of Culex quinquefasciatus and meta-analysis of infection responses to diverse pathogens. Science 2010, 330 (6000), 88-90, doi:10.1126/science.1193162.
Bażanów, B.; Jansen van Vuren, P.; Szymański, P.; Stygar, D.; Frącka, A.; Twardoń, J.; Kozdrowski, R.; Pawęska, J. T. A survey on West Nile and Usutu viruses in horses and birds in Poland. Viruses 2018, 10 (2), 87, doi:10.3390/v10020087.
Becker, N.; Jöst, H.; Ziegler, U.; Eiden, M.; Höper, D.; Emmerich, P.; Fichet-Calvet, E.; Ehichioya, D. U.; Czajka, C.; Gabriel, M.; Hoffmann, B.; Beer, M.; Tenner-Racz, K.; Racz, P.; Günther, S.; Wink, M.; Bosch, S.; Konrad, A.; Pfeffer, M.; Groschup, M. H.; Schmidt-Chanasit, J. Epizootic emergence of Usutu virus in wild and captive birds in Germany. PLoS One 2012, 7 (2), e32604, doi:10.1371/journal.pone.0032604.

Bibliography 83
Becker, N.; Petrić, D.; Zgomba, M.; Boase, C.; Madon, M.; Dahl, C.; Kaiser, A. Mosquitoes and their control, 2nd ed.; Springer-Verlag: Berlin/Heidelberg, Germany, 2010, doi: 10.1007/978-3-540-92874-4, ISBN 978-3-540-92874-4.
Beerntsen, B. T.; James, A. A.; Hristensen, B. M. Genetics of mosquito vector competence. Microbiol Mol Biol Rev 2000, 64 (1), 115–137, doi:10.1128/mmbr.64.1.115-137.2000.
Běhal, V. Nontraditional microbial bioactive metabolites. Folia Microbiol 2001, 46 (5), 363-370, doi:10.1007/BF02814422.
Ben Hassine, T.; Conte, A.; Calistri, P.; Candeloro, L.; Ippoliti, C.; De Massis, F.; Danzetta, M. L.; Bejaoui, M.; Hammami, S. Identification of suitable areas for West Nile virus circulation in Tunisia. Transbound Emerg Dis 2017, 64 (2), 449-458, doi:10.1111/tbed.12384.
Ben Hassine, T.; De Massis, F.; Calistri, P.; Savini, G.; BelHaj Mohamed, B.; Ranen, A.; Di Gennaro, A.; Sghaier, S.; Hammami, S. First detection of co-circulation of West Nile and Usutu viruses in equids in the south-west of Tunisia. Transbound Emerg Dis 2014, 61 (5), 385-389, doi:10.1111/tbed.12259.
Benedict, M. Q.; Levine, R. S.; Hawley, W. A.; Lounibos, L. P. Spread of the tiger: Global risk of invasion by the mosquito Aedes albopictus. Vector Borne Zoonotic Dis 2007, 7 (1), 76-85, doi:10.1089/vbz.2006.0562.
Benzarti, E.; Linden, A.; Desmecht, D.; Garigliany, M. Mosquito-borne epornitic flaviviruses: An update and review. J Gen Virol 2019, 100 (2), 119-132, doi:10.1099/jgv.0.001203.
Benzarti, E.; Rivas, J.; Sarlet, M.; Franssen, M.; Desmecht, D.; Schmidt-Chanasit, J.; Savini, G.; Lorusso, A.; Van Laere, A. S.; Garigliany, M. M. Experimental Usutu virus infection in domestic canaries Serinus canaria. Viruses 2020, 12 (2), 164, doi:10.3390/v12020164.
Bergold, G. H.; Suárez, O. M.; Munz, K. Multiplication in and transmission by Aedes aegypti of vesicular stomatitis virus. J Invertebr Pathol 1968, 11 (3), 406-428, doi:10.1016/0022-2011(68)90190-0.
Bernkopf, H.; Levine, S.; Nerson, R. Isolation of West Nile virus in Israel. J Infect Dis 1953, 93 (3), 207-218, doi:10.1093/infdis/93.3.207.
Black, W. C.; Moore, C. G.; Beaty, B. J.; Marquardt, W. C. Population biology as a tool for studying vector-borne diseases. In Biology of Disease Vectors, 2nd ed.; Marquardt, W. C. , Ed. Elsevier Academic Press: Burlington, MA, USA, 1996; pp. 393–416, ISBN 0-12-473276-3.
Blagrove, M. S.; Sherlock, K.; Chapman, G. E.; Impoinvil, D. E.; McCall, P. J.; Medlock, J. M.; Lycett, G.; Solomon, T.; Baylis, M. Evaluation of the vector competence of a native UK mosquito Ochlerotatus detritus (Aedes detritus) for dengue, chikungunya and West Nile viruses. Parasit Vectors 2016, 9, 452, doi:10.1186/s13071-016-1739-3.
Börstler, J.; Jöst, H.; Garms, R.; Krüger, A.; Tannich, E.; Becker, N.; Schmidt-Chanasit, J.; Lühken, R. Host-feeding patterns of mosquito species in Germany. Parasit Vectors 2016, 9 (1), 318, doi:10.1186/s13071-016-1597-z.
Bosch, I.; Herrera, F.; Navarro, J. C.; Lentino, M.; Dupuis, A.; Maffei, J.; Jones, M.; Fernández, E.; Pérez, N.; Pérez-Emán, J.; Guimarães, A. E.; Barrera, R.; Valero, N.; Ruiz, J.; Velásquez, G.; Martinez, J.; Comach, G.; Komar, N.; Spielman, A.; Kramer, L. West Nile virus, Venezuela. Emerg Infect Dis 2007, 13 (4), 651-653, doi:10.3201/eid1304.061383.
Bournez, L.; Umhang, G.; Faure, E.; Boucher, J. M.; Boué, F.; Jourdain, E.; Sarasa, M.; Llorente, F.; Jiménez-Clavero, M. A.; Moutailler, S.; Lacour, S. A.; Lecollinet, S.;

84 Bibliography
Beck, C. Exposure of wild ungulates to the Usutu and tick-borne encephalitis viruses in France in 2009-2014: Evidence of undetected flavivirus circulation a decade ago. Viruses 2019, 12 (1), 10, doi:10.3390/v12010010.
Brackney, D. E.; Beane, J. E.; Ebel, G. D. RNAi targeting of West Nile virus in mosquito midguts promotes virus diversification. PLoS Pathog 2009, 5 (7), e1000502, doi:10.1371/journal.ppat.1000502.
Brault, A. C. Changing patterns of West Nile virus transmission: Altered vector competence and host susceptibility. Vet Res 2009, 40 (2), 43, doi:10.1051/vetres/2009026.
Brilla, R.; Block, M.; Geremia, G.; Wichter, M. Clinical and neuroradiologic features of 39 consecutive cases of West Nile virus meningoencephalitis. J Neurol Sci 2004, 220 (1-2), 37-40, doi:10.1016/j.jns.2004.01.013.
Brinton, M. A. Replication cycle and molecular biology of the West Nile virus. Viruses 2013, 6 (1), 13-53, doi:10.3390/v6010013.
Brugman, V. A.; Hernández-Triana, L. M.; Medlock, J. M.; Fooks, A. R.; Carpenter, S.; Johnson, N. The role of Culex pipiens L. (Diptera: Culicidae) in virus transmission in Europe. Int J Environ Res Public Health 2018, 15 (2), 389, doi:10.3390/ijerph15020389.
Brustolin, M.; Santamaría, C.; Napp, S.; Verdún, M.; Rivas, R.; Pujol, N.; Talavera, S.; Busquets, N. Experimental study of the susceptibility of a European Aedes albopictus strain to dengue virus under a simulated Mediterranean temperature regime. Med Vet Entomol 2018, 32 (4), 393-398, doi:10.1111/mve.12325.
Brustolin, M.; Talavera, S.; Santamaría, C.; Rivas, R.; Pujol, N.; Aranda, C.; Marquès, E.; Valle, M.; Verdún, M.; Pagès, N.; Busquets, N. Culex pipiens and Stegomyia albopicta (= Aedes albopictus) populations as vectors for lineage 1 and 2 West Nile virus in Europe. Med Vet Entomol 2016, 30 (2), 166-173, doi:10.1111/mve.12164.
Bunning, M. L.; Bowen, R. A.; Cropp, C. B.; Sullivan, K. G.; Davis, B. S.; Komar, N.; Godsey, M. S.; Baker, D.; Hettler, D. L.; Holmes, D. A.; Biggerstaff, B. J.; Mitchell, C. J. Experimental infection of horses with West Nile virus. Emerg Infect Dis 2002, 8 (4), 380-386, doi:10.3201/eid0804.010239.
Byas, A. D.; Ebel, G. D. Comparative pathology of West Nile virus in humans and non-human animals. Pathogens 2020, 9 (1), 48, doi:10.3390/pathogens9010048.
Cadar, D.; Becker, N.; de Mendonca Campos, R.; Börstler, J.; Jöst, H.; Schmidt-Chanasit, J. Usutu virus in bats, Germany, 2013. Emerg Infect Dis 2014, 20 (10), 1771-1773, doi:10.3201/eid2010.140909.
Cadar, D.; Lühken, R.; van der Jeugd, H.; Garigliany, M.; Ziegler, U.; Keller, M.; Lahoreau, J.; Lachmann, L.; Becker, N.; Kik, M.; Oude Munnink, B. B.; Bosch, S.; Tannich, E.; Linden, A.; Schmidt, V.; Koopmans, M. P.; Rijks, J.; Desmecht, D.; Groschup, M. H.; Reusken, C.; Schmidt-Chanasit, J. Widespread activity of multiple lineages of Usutu virus, western Europe, 2016. Euro Surveill 2017a, 22 (4), pii=30452, doi:10.2807/1560-7917.ES.2017.22.4.30452.
Cadar, D.; Maier, P.; Müller, S.; Kress, J.; Chudy, M.; Bialonski, A.; Schlaphof, A.; Jansen, S.; Jöst, H.; Tannich, E.; Runkel, S.; Hitzler, W. E.; Hutschenreuter, G.; Wessiepe, M.; Schmidt-Chanasit, J. Blood donor screening for West Nile virus (WNV) revealed acute Usutu virus (USUV) infection, Germany, September 2016. Euro Surveill 2017b, 22 (14), pii=30501, doi:10.2807/1560-7917.ES.2017.22.14.30501.
Calisher, C. H.; Karabatsos, N.; Dalrymple, J. M.; Shope, R. E.; Porterfield, J. S.; Westaway, E. G.; Brandt, W. E. Antigenic relationships between flaviviruses as determined by

Bibliography 85
cross-neutralization tests with polyclonal antisera. J gen Virol 1989, 70 (1), 37-43, doi:10.1099/0022-1317-70-1-37.
Calzolari, M.; Bonilauri, P.; Bellini, R.; Albieri, A.; Defilippo, F.; Maioli, G.; Galletti, G.; Gelati, A.; Barbieri, I.; Tamba, M.; Lelli, D.; Carra, E.; Cordioli, P.; Angelini, P.; Dottori, M. Evidence of simultaneous circulation of West Nile and Usutu viruses in mosquitoes sampled in Emilia-Romagna region (Italy) in 2009. PLoS One 2010, 5 (12), e14324, doi:10.1371/journal.pone.0014324.
Camp, J. V.; Kolodziejek, J.; Nowotny, N. Targeted surveillance reveals native and invasive mosquito species infected with Usutu virus. Parasit Vectors 2019, 12 (1), 46, doi:10.1186/s13071-019-3316-z.
Camp, J. V.; Nowotny, N. The knowns and unknowns of West Nile virus in Europe: What did we learn from the 2018 outbreak? Expert Rev Anti Infect Ther 2020, 18 (2), 145-154, doi:10.1080/14787210.2020.1713751.
Campbell-Lendrum, D.; Manga, L.; Bagayoko, M.; Sommerfeld, J. Climate change and vector-borne diseases: What are the implications for public health research and policy? Phil Trans R Soc B 2015, 370 (1665), 20130552, doi:10.1098/rstb.2013.0552.
Campbell, G. L.; Marfin, A. A.; Lanciotti, R. S.; Gubler, D. J. West Nile virus. Lancet Infect Dis 2002, 2 (9), 519-529, doi:10.1016/s1473-3099(02)00368-7.
Cantile, C.; Del Piero, F.; Di Guardo, G.; Arispici, M. Pathologic and immunohistochemical findings in naturally occuring West Nile virus infection in horses. Vet Pathol 2001, 38 (4), 414-421, doi:10.1354/vp.38-4-414.
Cavrini, F.; Della Pepa, M. E.; Gaibani, P.; Pierro, A. M.; Rossini, G.; Landini, M. P.; Sambri, V. A rapid and specific real-time RT-PCR assay to identify Usutu virus in human plasma, serum, and cerebrospinal fluid. J Clin Virol 2011, 50 (3), 221-223, doi:10.1016/j.jcv.2010.11.008.
Cavrini, F.; Gaibani, P.; Longo, G.; Pierro, A. M.; Rossini, G.; Bonilauri, P.; Gerunda, G. E.; Di Benedetto, F.; Pasetto, A.; Girardis, M.; Dottori, M.; Landini, M. P.; Sambri, V. Usutu virus infection in a patient who underwent orthotropic liver transplantation, Italy, August-September 2009. Euro Surveill 2009, 14 (50), pii=19448, doi:10.2807/ese.14.50.19448-en.
CDC. Final cumulative maps & data for 1999–2018. 2019. Availabe online: https://www.cdc.gov/westnile/statsmaps/cumMapsData.html (accessed on 20 January 2020).
Ceianu, C. S.; Ungureanu, A.; Nicolescu, G.; Cernescu, C.; Nitescu, L.; Tardei, G.; Petrescu, A.; Pitigoi, D.; Martin, D.; Ciulacu-Purcarea, V.; Vladimirescu, A.; Savage, H. M. West Nile virus surveillance in Romania: 1997-2000. Viral Immunol 2001, 14 (3), 251-262, doi:10.1089/088282401753266765.
Cernescu, C.; Nedelcu, N. I.; Tardei, G.; Ruta, S.; Tsai, T. F. Continued transmission of West Nile virus to humans in southeastern Romania, 1997-1998. J Infect Dis 2000, 181 (2), 710-712, doi:10.1086/315225.
Chaintoutis, S. C.; Gewehr, S.; Mourelatos, S.; Dovas, C. I. Serological monitoring of backyard chickens in Central Macedonia-Greece can detect low transmission of West Nile virus in the absence of human neuroinvasive disease cases. Acta Trop 2016, 163, 26-31, doi:10.1016/j.actatropica.2016.07.018.
Chancey, C.; Grinev, A.; Volkova, E.; Rios, M. The global ecology and epidemiology of West Nile virus. Biomed Res Int 2015, 2015, 376230, doi:10.1155/2015/376230.

86 Bibliography
Chastel, C. [The "plague" of Barcelona. Yellow fever epidemic of 1821]. Bull Soc Pathol Exot 1999, 92 (5 Pt 2), 405-407.
Cheng, G.; Cox, J.; Wang, P.; Krishnan, M. N.; Dai, J.; Qian, F.; Anderson, J. F.; Fikrig, E. A C-type lectin collaborates with a CD45 phosphatase homolog to facilitate West Nile virus infection of mosquitoes. Cell 2010, 142 (5), 714-725, doi:10.1016/j.cell.2010.07.038.
Cherry, B.; Trock, S. C.; Glaser, A.; Kramer, L.; Ebel, G. D.; Glaser, C.; Miller, J. R. Sentinel chickens as a surveillance tool for West Nile virus in New York City, 2000. Ann N Y Acad Sci 2001, 951, 343-346, doi:10.1111/j.1749-6632.2001.tb02714.x.
Chevillon, C.; Eritja, R.; Pasteur, N.; Raymond, M. Commensalism, adaptation and gene flow: Mosquitos of the Culex pipiens complex in different habitats. Genet Res 1995, 66 (2), 147-157, doi:10.1017/S0016672300034492.
Christophers, S.R. Aedes aegypti (L.) the yellow fever mosquito: Its life history, bionomics and structure; Cambridge University Press: London, UK, 1960, ISBN 9780521046381.
Chu, J. J. H.; Ng, M. L. Infectious entry of West Nile virus occurs through a clathrin-mediated endocytic pathway. J Virol 2004, 78 (19), 10543-10555, doi:10.1128/Jvi.78.19.10543-10555.2004.
Chvala, S.; Bakonyi, T.; Bukovsky, C.; Meister, T.; Brugger, K.; Rubel, F.; Nowotny, N.; Weissenböck, H. Monitoring of Usutu virus activity and spread by using dead bird surveillance in Austria, 2003-2005. Vet Microbiol 2007, 122 (3-4), 237-245, doi:10.1016/j.vetmic.2007.01.029.
Chvala, S.; Bakonyi, T.; Hackl, R.; Hess, M.; Nowotny, N.; Weissenböck, H. Limited pathogenicity of Usutu virus for the domestic chicken (Gallus domesticus). Avian Pathol 2005, 34 (5), 392-395, doi:10.1080/03079450500268500.
Chvala, S.; Bakonyi, T.; Hackl, R.; Hess, M.; Nowotny, N.; Weissenböck, H. Limited pathogenicity of Usutu virus for the domestic goose (Anser anser f. domestica) following experimental inoculation. J Vet Med B Infect Dis Vet Public Health 2006, 53 (4), 171-175, doi:10.1111/j.1439-0450.2006.00942.x.
Chvala, S.; Kolodziejek, J.; Nowotny, N.; Weissenböck, H. Pathology and viral distribution in fatal Usutu virus infections of birds from the 2001 and 2002 outbreaks in Austria. J Comp Pathol 2004, 131 (2-3), 176-185, doi:10.1016/j.jcpa.2004.03.004.
Ciota, A. T.; Chin, P. A.; Kramer, L. D. The effect of hybridization of Culex pipiens complex mosquitoes on transmission of West Nile virus. Parasit Vectors 2013, 6 (1), 305, doi:10.1186/1756-3305-6-305.
Ciota, A. T.; Keyel, A. C. The role of temperature in transmission of zoonotic arboviruses. Viruses 2019, 11 (11), 1013, doi:10.3390/v11111013.
Ciota, A. T.; Kramer, L. D. Vector-virus interactions and transmission dynamics of West Nile virus. Viruses 2013, 5 (12), 3021-3047, doi:10.3390/v5123021.
Clé, M.; Beck, C.; Salinas, S.; Lecollinet, S.; Gutierrez, S.; Van de Perre, P.; Baldet, T.; Foulongne, V.; Simonin, Y. Usutu virus: A new threat? Epidemiol Infect 2019, 147, e232, doi:10.1017/S0950268819001213.
Clem, R. J. Arboviruses and apoptosis: The role of cell death in determining vector competence. J Gen Virol 2016, 97 (5), 1033-1036, doi:10.1099/jgv.0.000429.
Colpitts, T. M.; Conway, M. J.; Montgomery, R. R.; Fikrig, E. West Nile virus: Biology, transmission, and human infection. Clin Microbiol Rev 2012, 25 (4), 635-648, doi:10.1128/CMR.00045-12.

Bibliography 87
Colpitts, T. M.; Cox, J.; Vanlandingham, D. L.; Feitosa, F. M.; Cheng, G.; Kurscheid, S.; Wang, P.; Krishnan, M. N.; Higgs, S.; Fikrig, E. Alterations in the Aedes aegypti transcriptome during infection with West Nile, dengue and yellow fever viruses. PLoS Pathog 2011, 7 (9), e1002189, doi:10.1371/journal.ppat.1002189.
Coman, A.; Maftei, D. N.; Chereches, R. M.; Zavrotchi, E.; Bria, P.; Dragnea, C.; McKenzie, P. P.; Valentine, M. A.; Gray, G. C. Avian influenza surveillance in the Danube Delta using sentinel geese and ducks. Influenza Res Treat 2014, 2014, 965749, doi:10.1155/2014/965749.
Cook, C. L.; Huang, Y. S.; Lyons, A. C.; Alto, B. W.; Unlu, I.; Higgs, S.; Vanlandingham, D. L. North American Culex pipiens and Culex quinquefasciatus are competent vectors for Usutu virus. PLoS Negl Trop Dis 2018, 12 (8), e0006732, doi:10.1371/journal.pntd.0006732.
Cvjetković, I. H.; Petrović, T.; Petrić, D.; Cvjetković, D.; Kovačević, G.; Radovanov, J.; Galović, A. J.; Patić, A.; Nikolić, N.; Mikić, S. S.; Milošević, V. Seroprevalence of mosquito-born and tick-born microorganisms in human population of South Backa District. Arch Vet Med 2016 9(1), 23-30, doi:10.46784/e-avm.v9i1.94.
da Cunha Sais, T.; de Moraes, R. M.; Ribolla, P. E.; de Bianchi, A. G.; Marinotti, O.; Bijovsky, A. T. Morphological aspects of Culex quinquefasciatus salivary glands. Arthropod Struct Dev 2003, 32 (2-3), 219-226, doi:10.1016/S1467-8039(03)00035-5.
Danis, K.; Papa, A.; Papanikolaou, E.; Dougas, G.; Terzaki, I.; Baka, A.; Vrioni, G.; Kapsimali, V.; Tsakris, A.; Kansouzidou, A.; Tsiodras, S.; Vakalis, N.; Bonovas, S.; Kremastinou, J. Ongoing outbreak of West Nile virus infection in humans, Greece, July to August 2011. Euro Surveill 2011a, 16 (34), pii=19951, doi:10.2807/ese.16.34.19951-en.
Danis, K.; Papa, A.; Theocharopoulos, G.; Dougas, G.; Athanasiou, M.; Detsis, M.; Baka, A.; Lytras, T.; Mellou, K.; Bonovas, S.; Panagiotopoulos, T. Outbreak of West Nile virus infection in Greece, 2010. Emerg Infect Dis 2011b, 17 (10), 1868-1872, doi:10.3201/eid1710.110525.
Das, S.; Radtke, A.; Choi, Y. J.; Mendes, A. M.; Valenzuela, J. G.; Dimopoulos, G. Transcriptomic and functional analysis of the Anopheles gambiae salivary gland in relation to blood feeding. BMC Genomics 2010, 11, 566, doi:10.1186/1471-2164-11-566.
Davis, C. W.; Nguyen, H. Y.; Hanna, S. L.; Sánchez, M. D.; Doms, R. W.; Pierson, T. C. West Nile virus discriminates between DC-SIGN and DC-SIGNR for cellular attachment and infection. J Virol 2006, 80 (3), 1290-1301, doi:10.1128/Jvi.80.3.1290-1301.2006.
De Filette, M.; Ulbert, S.; Diamond, M.; Sanders, N. N. Recent progress in West Nile virus diagnosis and vaccination. Vet Res 2012, 43, 16, doi:10.1186/1297-9716-43-16.
de Oya, N. J.; Escribano-Romero, E.; Blázquez, A. B.; Martín-Acebes, M. A.; Saiz, J. C. Current progress of avian vaccines against West Nile virus. Vaccines 2019, 7 (4), 126, doi:10.3390/vaccines7040126.
DeBiasi, R. L.; Tyler, K. L. West Nile virus meningoencephalitis. Nat Clin Pract Neurol 2006, 2 (5), 264-275, doi:10.1038/ncpneuro0176.
Dennison, N. J.; Jupatanakul, N.; Dimopoulos, G. The mosquito microbiota influences vector competence for human pathogens. Curr Opin Insect Sci 2014, 3, 6-13, doi:10.1016/j.cois.2014.07.004.

88 Bibliography
Deubel, V.; Fiette, L.; Gounon, P.; Drouet, M. T.; Khun, H.; Huerre, M.; Banet, C.; Malkinson, M.; Desprès, P. Variations in biological features of West Nile viruses. Ann N Y Acad Sci 2001, 951, 195-206, doi:10.1111/j.1749-6632.2001.tb02697.x.
Diagne, M. M.; Ndione, M. H. D.; Di Paola, N.; Fall, G.; Bedekelabou, A. P.; Sembène, P. M.; Faye, O.; Zanotto, P. M. A.; Sall, A. A. Usutu virus isolated from rodents in Senegal. Viruses 2019, 11 (2), 181, doi:10.3390/v11020181.
Dinu, S.; Cotar, A. I.; Pănculescu-Gătej, I. R.; Fălcuţă, E.; Prioteasa, F. L.; Sîrbu, A.; Oprişan, G.; Bădescu, D.; Reiter, P.; Ceianu, C. S. West Nile virus circulation in south-eastern Romania, 2011 to 2013. Euro Surveill 2015, 20 (20), pii=21130, doi:10.2807/1560-7917.es2015.20.20.21130.
Dodson, B. L.; Hughes, G. L.; Paul, O.; Matacchiero, A. C.; Kramer, L. D.; Rasgon, J. L. Wolbachia enhances West Nile virus (WNV) infection in the mosquito Culex tarsalis. PLoS Negl Trop Dis 2014, 8 (7), e2965, doi:10.1371/journal.pntd.0002965.
Drebot, M. A.; Artsob, H. West Nile virus - Update for family physicians. Can Fam Physician 2005, 51, 1094-1099.
Durand, B.; Haskouri, H.; Lowenski, S.; Vachiery, N.; Beck, C.; Lecollinet, S. Seroprevalence of West Nile and Usutu viruses in military working horses and dogs, Morocco, 2012: Dog as an alternative WNV sentinel species? Epidemiol Infect 2016, 144 (9), 1857-1864, doi:10.1017/S095026881600011X.
DWD. Annual Report 2018. In Yearbook 2018 of the Deutscher Wetterdienst; DWD: Offenbach am Main, Germany, 2018a; p 16.
DWD. Deutschlandwetter im Sommer 2018; DWD: Offenbach am Main, Germany, 2018b. DWD. The weather in Germany in summer 2019; DWD: Offenbach am Main, Germany,
2019. Ebel, G. D.; Dupuis, A. P., 2nd; Ngo, K.; Nicholas, D.; Kauffman, E.; Jones, S. A.; Young, D.;
Maffei, J.; Shi, P. Y.; Bernard, K.; Kramer, L. D. Partial genetic characterization of West Nile virus strains, New York State, 2000. Emerg Infect Dis 2001, 7 (4), 650-653, doi:10.3201/eid0704.010408.
ECDC. Epidemiological update: West Nile virus transmission season in Europe, 2018. 2018. Availabe online: https://www.ecdc.europa.eu/en/news-events/epidemiological-update-west-nile-virus-transmission-season-europe-2018 (accessed on 8 June 2020).
ECDC. Epidemiological update: West Nile virus transmission season in Europe, 2019. 2019. Availabe online: https://www.ecdc.europa.eu/en/news-events/epidemiological-update-west-nile-virus-transmission-season-europe-2019 (accessed on 8 June 2020).
ECDC. Aedes albopictus - current known distribution: May 2020. 2020. Availabe online: https://www.ecdc.europa.eu/en/publications-data/aedes-albopictus-current-known-distribution-may-2020 (accessed on 20 January 2020).
Eiden, M.; Gil, P.; Ziegler, U.; Rakotoarivony, I.; Marie, A.; Frances, B.; L'Ambert, G.; Simonin, Y.; Foulongne, V.; Groschup, M. H.; Gutierrez, S.; Eloit, M. Emergence of two Usutu virus lineages in Culex pipiens mosquitoes in the Camargue, France, 2015. Infect Genet Evol 2018, 61, 151-154, doi:10.1016/j.meegid.2018.03.020.
Eiden, M.; Viña-Rodríguez, A.; Hoffmann, B.; Ziegler, U.; Groschup, M. H. Two new real-time quantitative reverse transcription polymerase chain reaction assays with unique target sites for the specific and sensitive detection of lineages 1 and 2 West Nile virus strains. J Vet Diagn Invest 2010, 22 (5), 748-753, doi:10.1177/104063871002200515.
El Garch, H.; Minke, J. M.; Rehder, J.; Richard, S.; Edlund Toulemonde, C.; Dinic, S.; Andreoni, C.; Audonnet, J. C.; Nordgren, R.; Juillard, V. A West Nile virus (WNV)

Bibliography 89
recombinant canarypox virus vaccine elicits WNV-specific neutralizing antibodies and cell-mediated immune responses in the horse. Vet Immunol Immunopathol 2008, 123 (3-4), 230-239, doi:10.1016/j.vetimm.2008.02.002.
Elizondo-Quiroga, D.; Davis, C. T.; Fernandez-Salas, I.; Escobar-Lopez, R.; Velasco Olmos, D.; Soto Gastalum, L. C.; Aviles Acosta, M.; Elizondo-Quiroga, A.; Gonzalez-Rojas, J. I.; Contreras Cordero, J. F.; Guzman, H.; Travassos da Rosa, A.; Blitvich, B. J.; Barrett, A. D.; Beaty, B. J.; Tesh, R. B. West Nile virus isolation in human and mosquitoes, Mexico. Emerg Infect Dis 2005, 11 (9), 1449-1452, doi:10.3201/eid1109.050121.
Engel, D.; Jöst, H.; Wink, M.; Börstler, J.; Bosch, S.; Garigliany, M. M.; Jöst, A.; Czajka, C.; Lühken, R.; Ziegler, U.; Groschup, M. H.; Pfeffer, M.; Becker, N.; Cadar, D.; Schmidt-Chanasit, J. Reconstruction of the evolutionary history and dispersal of Usutu virus, a neglected emerging arbovirus in Europe and Africa. MBio 2016, 7 (1), e01938-01915, doi:10.1128/mBio.01938-15.
Engler, O.; Savini, G.; Papa, A.; Figuerola, J.; Groschup, M. H.; Kampen, H.; Medlock, J.; Vaux, A.; Wilson, A. J.; Werner, D.; Jöst, H.; Goffredo, M.; Capelli, G.; Federici, V.; Tonolla, M.; Patocchi, N.; Flacio, E.; Portmann, J.; Rossi-Pedruzzi, A.; Mourelatos, S.; Ruiz, S.; Vázquez, A.; Calzolari, M.; Bonilauri, P.; Dottori, M.; Schaffner, F.; Mathis, A.; Johnson, N. European surveillance for West Nile virus in mosquito populations. Int J Environ Res Public Health 2013, 10 (10), 4869-4895, doi:10.3390/ijerph10104869.
Escribano-Romero, E.; Lupulović, D.; Merino-Ramos, T.; Blázquez, A. B.; Lazić, G.; Lazić, S.; Saiz, J. C.; Petrović, T. West Nile virus serosurveillance in pigs, wild boars, and roe deer in Serbia. Vet Microbiol 2015, 176 (3-4), 365-369, doi:10.1016/j.vetmic.2015.02.005.
European Commission. Opion of the scientific committee on veterinary measures relating to public health on West Nile virus (WNV). 2003. Availabe online: https://ec.europa.eu/food/sites/food/files/safety/docs/sci-com_scv_out67_en.pdf (accessed on 8 June 2020).
Ewing, D. A.; Cobbold, C. A.; Purse, B. V.; Nunn, M. A.; White, S. M. Modelling the effect of temperature on the seasonal population dynamics of temperate mosquitoes. J Theor Biol 2016, 400, 65-79, doi:10.1016/j.jtbi.2016.04.008.
Fenton, A.; Pedersen, A. B. Community epidemiology framework for classifying disease threats. Emerg Infect Dis 2005, 11 (12), 1815-1821, doi:10.3201/eid1112.050306.
Feyer, S.; Bartenschlager, F.; Bertram, C. A.; Ziegler, U.; Fast, C.; Klopfleisch, R.; Müller, K. Clinical, pathological and virological aspects of fatal West Nile virus infections in ten free-ranging goshawks (Accipiter gentilis) in Germany. Transbound Emerg Dis 2020, 00, 1-13, doi:10.1111/tbed.13759.
Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol Mar Biol Biotechnol 1994, 3 (5), 294-299.
Fontaine, A.; Diouf, I.; Bakkali, N.; Missé, D.; Pagès, F.; Fusai, T.; Rogier, C.; Almeras, L. Implication of haematophagous arthropod salivary proteins in host-vector interactions. Parasit Vectors 2011, 4, 187, doi:10.1186/1756-3305-4-187.
Fortuna, C.; Remoli, M. E.; Di Luca, M.; Severini, F.; Toma, L.; Benedetti, E.; Bucci, P.; Montarsi, F.; Minelli, G.; Boccolini, D.; Romi, R.; Ciufolini, M. G. Experimental studies on comparison of the vector competence of four Italian Culex pipiens

90 Bibliography
populations for West Nile virus. Parasit Vectors 2015a, 8, 463, doi:10.1186/s13071-015-1067-z.
Fortuna, C.; Remoli, M. E.; Severini, F.; Di Luca, M.; Toma, L.; Fois, F.; Bucci, P.; Boccolini, D.; Romi, R.; Ciufolini, M. G. Evaluation of vector competence for West Nile virus in Italian Stegomyia albopicta (=Aedes albopictus) mosquitoes. Med Vet Entomol 2015b, 29 (4), 430-433, doi:10.1111/mve.12133.
Franz, A. W.; Kantor, A. M.; Passarelli, A. L.; Clem, R. J. Tissue barriers to arbovirus infection in mosquitoes. Viruses 2015, 7 (7), 3741-3767, doi:10.3390/v7072795.
Fros, J. J.; Miesen, P.; Vogels, C. B.; Gaibani, P.; Sambri, V.; Martina, B. E.; Koenraadt, C. J.; van Rij, R. P.; Vlak, J. M.; Takken, W.; Pijlman, G. P. Comparative Usutu and West Nile virus transmission potential by local Culex pipiens mosquitoes in north-western Europe. One Health 2015, 1, 31-36, doi:10.1016/j.onehlt.2015.08.002.
Gaibani, P.; Pierro, A.; Alicino, R.; Rossini, G.; Cavrini, F.; Landini, M. P.; Sambri, V. Detection of Usutu-virus-specific IgG in blood donors from northern Italy. Vector Borne Zoonotic Dis 2012, 12 (5), 431-433, doi:10.1089/vbz.2011.0813.
Gaibani, P.; Rossini, G. An overview of Usutu virus. Microbes Infect 2017, 19 (7-8), 382-387, doi:10.1016/j.micinf.2017.05.003.
Gamino, V.; Höfle, U. Pathology and tissue tropism of natural West Nile virus infection in birds: A review. Vet Res 2013, 44, 39, doi:10.1186/1297-9716-44-39.
Gancz, A. Y.; Barker, I. K.; Lindsay, R.; Dibernardo, A.; McKeever, K.; Hunter, B. West Nile virus outbreak in North American owls, Ontario, 2002. Emerg Infect Dis 2004, 10 (12), 2135-2142, doi:10.3201/eid1012.040167.
García-Bocanegra, I.; Jaén-Téllez, J. A.; Napp, S.; Arenas-Montes, A.; Fernández-Morente, M.; Fernández-Molera, V.; Arenas, A. West Nile fever outbreak in horses and humans, Spain, 2010. Emerg Infect Dis 2011, 17 (12), 2397-2399, doi:10.3201/eid1712.110651.
García-Bocanegra, I.; Paniagua, J.; Gutiérrez-Guzman, A. V.; Lecollinet, S.; Boadella, M.; Arenas-Montes, A.; Cano-Terriza, D.; Lowenski, S.; Gortázar, C.; Höfle, U. Spatio-temporal trends and risk factors affecting West Nile virus and related flavivirus exposure in Spanish wild ruminants. BMC Vet Res 2016, 12 (1), 249, doi:10.1186/s12917-016-0876-4.
Garcia-Tapia, D.; Loiacono, C. M.; Kleiboeker, S. B. Replication of West Nile virus in equine peripheral blood mononuclear cells. Vet Immunol Immunopathol 2006, 110 (3-4), 229-244, doi:10.1016/j.vetimm.2005.10.003.
Garrett-Jones, C. Prognosis for interruption of malaria transmission through assessment of the mosquito's vectorial capacity. Nature 1964, 204, 1173-1175, doi:10.1038/2041173a0.
Gibbs, S. E.; Hoffman, D. M.; Stark, L. M.; Marlenee, N. L.; Blitvich, B. J.; Beaty, B. J.; Stallknecht, D. E. Persistence of antibodies to West Nile virus in naturally infected rock pigeons (Columba livia). Clin Diagn Lab Immunol 2005, 12 (5), 665-667, doi:10.1128/CDLI.12.5.665-667.2005.
Girard, Y. A.; Klingler, K. A.; Higgs, S. West Nile virus dissemination and tissue tropisms in orally infected Culex pipiens quinquefasciatus. Vector Borne Zoonotic Dis 2004, 4 (2), 109-122, doi:10.1089/1530366041210729.
Girard, Y. A.; Mayhew, G. F.; Fuchs, J. F.; Li, H.; Schneider, B. S.; McGee, C. E.; Rocheleau, T. A.; Helmy, H.; Christensen, B. M.; Higgs, S.; Bartholomay, L. C. Transcriptome changes in Culex quinquefasciatus (Diptera: Culicidae) salivary glands

Bibliography 91
during West Nile virus infection. J Med Entomol 2010, 47 (3), 421-435, doi:10.1603/me09249.
Girard, Y. A.; Popov, V.; Wen, J.; Han, V.; Higgs, S. Ultrastructural study of West Nile virus pathogenesis in Culex pipiens quinquefasciatus (Diptera: Culicidae). J Med Entomol 2005, 42 (3), 429-444, doi:10.1093/jmedent/42.3.429.
Girard, Y. A.; Schneider, B. S.; McGee, C. E.; Wen, J.; Han, V. C.; Popov, V.; Mason, P. W.; Higgs, S. Salivary gland morphology and virus transmission during long-term cytopathologic West Nile virus infection in Culex mosquitoes. Am J Trop Med Hyg 2007, 76 (1), 118-128.
Globig, A.; Baumer, A.; Revilla-Fernández, S.; Beer, M.; Wodak, E.; Fink, M.; Greber, N.; Harder, T. C.; Wilking, H.; Brunhart, I.; Matthes, D.; Kraatz, U.; Strunk, P.; Fiedler, W.; Fereidouni, S. R.; Staubach, C.; Conraths, F. J.; Griot, C.; Mettenleiter, T. C.; Stärk, K. D. C. Ducks as sentinels for avian influenza in wild birds. Emerg Infect Dis 2009, 15 (10), 1633-1636, doi:10.3201/eid1510.090439.
Gould, E. A.; Higgs, S. Impact of climate change and other factors on emerging arbovirus diseases. Trans R Soc Trop Med Hyg 2009, 103 (2), 109-121, doi:10.1016/j.trstmh.2008.07.025.
Grottola, A.; Marcacci, M.; Tagliazucchi, S.; Gennari, W.; Di Gennaro, A.; Orsini, M.; Monaco, F.; Marchegiano, P.; Marini, V.; Meacci, M.; Rumpianesi, F.; Lorusso, A.; Pecorari, M.; Savini, G. Usutu virus infections in humans: A retrospective analysis in the municipality of Modena, Italy. Clin Microbiol Infect 2017, 23 (1), 33-37, doi:10.1016/j.cmi.2016.09.019.
Hahn, D. C.; Nemeth, N. M.; Edwards, E.; Bright, P. R.; Komar, N. Passive West Nile virus antibody transfer from maternal Eastern screech-owls (Megascops asio) to progeny. Avian Dis 2006, 50 (3), 454-455, doi:10.1637/7509-012606R1.1.
Halliday, J. E. B.; Meredith, A. L.; Knobel, D. L.; Shaw, D. J.; Bronsvoort, B. M. D. C.; Cleaveland, S. A framework for evaluating animals as sentinels for infectious disease surveillance. J. R. Soc. Interface 2007, 4 (16), 973-984, doi:10.1098/rsif.2007.0237.
Hamer, G. L.; Kitron, U. D.; Brawn, J. D.; Loss, S. R.; Ruiz, M. O.; Goldberg, T. L.; Walker, E. D. Culex pipiens (Diptera: Culicidae): A bridge vector of West Nile virus to humans. J Med Entomol 2008, 45 (1), 125-128, doi:10.1093/jmedent/45.1.125.
Han, L. L.; Popovici, F.; Alexander, J. P., Jr.; Laurentia, V.; Tengelsen, L. A.; Cernescu, C.; Gary, H. E., Jr.; Ion-Nedelcu, N.; Campbell, G. L.; Tsai, T. F. Risk factors for West Nile virus infection and meningoencephalitis, Romania, 1996. J Infect Dis 1999, 179 (1), 230-233, doi:10.1086/314566.
Hankins, K. Significance of West Nile virus equine strain variations. In Pfizer Animal Health Technical Bulletin; Pfizer Animal Health: New York, NY, USA, 2010; pp 1-9.
Hardy, J. L.; Houk, E. J.; Kramer, L. D.; Reeves, W. C. Intrinsic factors affecting vector competence of mosquitoes for arboviruses. Annu Rev Entomol 1983, 28, 229-262, doi:10.1146/annurev.en.28.010183.001305.
Harvey, W. Least-Squares Analysis of Data with Unequal Subclass Numbers; USDA National Agricultural Library: Washington, DC, USA, 1960.
Hayes, C. G. West Nile virus: Uganda, 1937, to New York City, 1999. Ann N Y Acad Sci 2001, 951, 25-37, doi:10.1111/j.1749-6632.2001.tb02682.x.
Hayes, E. B.; Komar, N.; Nasci, R. S.; Montgomery, S. P.; O'Leary, D. R.; Campbell, G. L. Epidemiology and transmission dynamics of West Nile virus disease. Emerg Infect Dis 2005, 11 (8), 1167-1173, doi:10.3201/eid1108.050289a.

92 Bibliography
Hayes, E. B.; O'Leary, D. R. West Nile virus infection: A pediatric perspective. Pediatrics 2004, 113 (5), 1375-1381, doi:10.1542/peds.113.5.1375.
Hebert, P. D.; Ratnasingham, S.; deWaard, J. R. Barcoding animal life: Cytochrome c oxidase subunit 1 divergences among closely related species. Proc R Soc Lond 2003, 270, S96-S99, doi:10.1098/rsbl.2003.0025.
Heitmann, A.; Jansen, S.; Lühken, R.; Leggewie, M.; Schmidt-Chanasit, J.; Tannich, E. Forced salivation as a method to analyze vector competence of mosquitoes. J Vis Exp 2018, (138), e57980, doi:10.3791/57980.
Hernández-Triana, L. M.; de Marco, M. F.; Mansfield, K. L.; Thorne, L.; Lumley, S.; Marston, D.; Fooks, A. A.; Johnson, N. Assessment of vector competence of UK mosquitoes for Usutu virus of African origin. Parasit Vectors 2018, 11 (1), 381, doi:10.1186/s13071-018-2959-5.
Hesson, J. C.; Rettich, F.; Merdić, E.; Vignjević, G.; Östman, O.; Schäfer, M.; Schaffner, F.; Foussadier, R.; Besnard, G.; Medlock, J.; Scholte, E. J.; Lundström, J. O. The arbovirus vector Culex torrentium is more prevalent than Culex pipiens in northern and central Europe. Med Vet Entomol 2014, 28 (2), 179-186, doi:10.1111/mve.12024.
Höfle, U.; Blanco, J. M.; Crespo, E.; Naranjo, V.; Jiménez-Clavero, M. A.; Sanchez, A.; de la Fuente, J.; Gortazar, C. West Nile virus in the endangered Spanish imperial eagle. Vet Microbiol 2008, 129 (1-2), 171-178, doi:10.1016/j.vetmic.2007.11.006.
Höfle, U.; Gamino, V.; de Mera, I. G.; Mangold, A. J.; Ortíz, J. A.; de la Fuente, J. Usutu virus in migratory song thrushes, Spain. Emerg Infect Dis 2013, 19 (7), 1173-1175, doi:10.3201/eid1907.130199.
Holick, J.; Kyle, A.; Ferraro, W.; Delaney, R. R.; Iwaseczko, M. Discovery of Aedes albopictus infected with West Nile virus in southeastern Pennsylvania. J Am Mosq Control Assoc 2002, 18 (2), 131.
Holicki, C. M.; Michel, F.; Vasić, A.; Fast, C.; Eiden, M.; Răileanu, C.; Kampen, H.; Werner, D.; Groschup, M. H.; Ziegler, U. Pathogenicity of West Nile virus lineage 1 to German poultry. Vaccines 2020a, 8 (3), 507, doi:10.3390/vaccines8030507.
Holicki, C. M.; Ziegler, U.; Răileanu, C.; Kampen, H.; Werner, D.; Schulz, J.; Silaghi, C.; Groschup, M. H.; Vasić, A. West Nile virus lineage 2 vector competence of indigenous Culex and Aedes mosquitoes from Germany at temperate climate conditions. Viruses 2020b, 12 (5), 561, doi:10.3390/v12050561.
Houk, E. J.; Obie, F.; Hardy, J. L. Peritrophic membrane formation and the midgut barrier to arboviral infection in the mosquito, Culex tarsalis Coquillett (Insecta, Diptera). Acta Trop 1979, 36 (1), 39-45.
Huber, K.; Jansen, S.; Leggewie, M.; Badusche, M.; Schmidt-Chanasit, J.; Becker, N.; Tannich, E.; Becker, S. C. Aedes japonicus japonicus (Diptera: Culicidae) from Germany have vector competence for Japan encephalitis virus but are refractory to infection with West Nile virus. Parasitol Res 2014, 113 (9), 3195-3199, doi:10.1007/s00436-014-3983-9.
Ibañez-Justicia, A.; Gloria-Soria, A.; den Hartog, W.; Dik, M.; Jacobs, F.; Stroo, A. The first detected airline introductions of yellow fever mosquitoes (Aedes aegypti) to Europe, at Schiphol International Airport, the Netherlands. Parasit Vectors 2017, 10 (1), 603, doi:10.1186/s13071-017-2555-0.
International Committee on Taxonomy of Viruses (ICTV). Taxonomy. 2020. Availabe online: https://talk.ictvonline.org/taxonomy/ (accessed on 09 September 2020).

Bibliography 93
Iwamoto, M.; Jernigan, D. B.; Guasch, A.; Trepka, M. J.; Blackmore, C. G.; Hellinger, W. C.; Pham, S. M.; Zaki, S.; Lanciotti, R. S.; Lance-Parker, S. E.; DiazGranados, C. A.; Winquist, A. G.; Perlino, C. A.; Wiersma, S.; Hillyer, K. L.; Goodman, J. L.; Marfin, A. A.; Chamberland, M. E.; Petersen, L. R.; West Nile Virus in Transplant Recipients Investigation Team. Transmission of West Nile virus from an organ donor to four transplant recipients. N Engl J Med 2003, 348 (22), 2196-2203, doi:10.1056/NEJMoa022987.
Jansen, S.; Heitmann, A.; Lühken, R.; Leggewie, M.; Helms, M.; Badusche, M.; Rossini, G.; Schmidt-Chanasit, J.; Tannich, E. Culex torrentium: A potent vector for the transmission of West Nile virus in Central Europe. Viruses 2019, 11 (6), 492, doi:10.3390/v11060492.
Jiménez-Clavero, M. A.; Sotelo, E.; Fernández-Pinero, J.; Llorente, F.; Blanco, J. M.; Rodriguez-Ramos, J.; Pérez-Ramírez, E.; Höfle, U. West Nile virus in golden eagles, Spain, 2007. Emerg Infect Dis 2008, 14 (9), 1489-1491, doi:10.3201/eid1409.080190.
Johnson, K. N. The impact of Wolbachia on virus infection in mosquitoes. Viruses 2015, 7 (11), 5705-5717, doi:10.3390/v7112903.
Jöst, H.; Bialonski, A.; Maus, D.; Sambri, V.; Eiden, M.; Groschup, M. H.; Günther, S.; Becker, N.; Schmidt-Chanasit, J. Isolation of Usutu virus in Germany. Am J Trop Med Hyg 2011, 85 (3), 551-553, doi:10.4269/ajtmh.2011.11-0248.
Joubert, L.; Oudar, J.; Hannoun, C.; Beytout, D.; Corniou, B.; Guillon, J. C.; Panthier, R. [Epidemiology of the West Nile virus: Study of a focus in Camargue. IV. Meningo-encephalomyelitis of the horse]. Ann Inst Pasteur (Paris) 1970, 118 (2), 239-247.
Jupatanakul, N.; Sim, S.; Dimopoulos, G. The insect microbiome modulates vector competence for arboviruses. Viruses 2014, 6 (11), 4294-4313, doi:10.3390/v6114294.
Kaiser, J. Public health. Outbreak pattern stymies vaccine work. Science 2012, 337 (6098), 1030, doi:10.1126/science.337.6098.1030.
Kaiser, J. A.; Barrett, A. D. T. Twenty years of progress toward West Nile virus vaccine development. Viruses 2019, 11 (9), 823, doi:10.3390/v11090823.
Kampen, H.; Holicki, C. M.; Ziegler, U.; Groschup, M. H.; Tews, B. A.; Werner, D. West Nile virus mosquito vectors (Diptera: Culicidae) in Germany. Viruses 2020, 12 (5), 493, doi:10.3390/v12050493.
Kampen, H.; Jansen, S.; Schmidt-Chanasit, J.; Walther, D. Indoor development of Aedes aegypti in Germany, 2016. Euro Surveill 2016, 21 (47), pii=30407, doi:10.2807/1560-7917.ES.2016.21.47.30407.
Kampen, H.; Werner, D. Out of the bush: The Asian bush mosquito Aedes japonicus japonicus (Theobald, 1901) (Diptera, Culicidae) becomes invasive. Parasit Vectors 2014, 7, 59, doi:10.1186/1756-3305-7-59.
Kato, N.; Mueller, C. R.; Fuchs, J. F.; McElroy, K.; Wessely, V.; Higgs, S.; Christensen, B. M. Evaluation of the function of a type I peritrophic matrix as a physical barrier for midgut epithelium invasion by mosquito-borne pathogens in Aedes aegypti. Vector Borne Zoonotic Dis 2008, 8 (5), 701-712, doi:10.1089/vbz.2007.0270.
Kemenesi, G.; Buzás, D.; Zana, B.; Kurucz, K.; Krtinic, B.; Kepner, A.; Földes, F.; Jakab, F. First genetic characterization of Usutu virus from Culex pipiens mosquitoes Serbia, 2014. Infect Genet Evol 2018, 63, 58-61, doi:10.1016/j.meegid.2018.05.012.
Kenney, J. L.; Brault, A. C. The role of environmental, virological and vector interactions in dictating biological transmission of arthropod-borne viruses by mosquitoes. Adv Virus Res 2014, 89, 39-83, doi:10.1016/B978-0-12-800172-1.00002-1.

94 Bibliography
King, N. J.; Getts, D. R.; Getts, M. T.; Rana, S.; Shrestha, B.; Kesson, A. M. Immunopathology of flavivirus infections. Immunol Cell Biol 2007, 85 (1), 33-42, doi:10.1038/sj.icb.7100012.
Klenk, K.; Komar, N. Poor replication of West Nile virus (New York 1999 strain) in three reptilian and one amphibian species. Am J Trop Med Hyg 2003, 69 (3), 260-262.
Klenk, K.; Snow, J.; Morgan, K.; Bowen, R.; Stephens, M.; Foster, F.; Gordy, P.; Beckett, S.; Komar, N.; Gubler, D.; Bunning, M. Alligators as West Nile virus amplifiers. Emerg Infect Dis 2004, 10 (12), 2150-2155, doi:10.3201/eid1012.040264.
Komar, N. West Nile virus surveillance using sentinel birds. Ann N Y Acad Sci 2001, 951, 58-73, doi:10.1111/j.1749-6632.2001.tb02685.x.
Komar, N.; Clark, G. G. West Nile virus activity in Latin America and the Caribbean. Rev Panam Salud Publica 2006, 19 (2), 112-117, doi:10.1590/s1020-49892006000200006.
Komar, N.; Langevin, S.; Hinten, S.; Nemeth, N.; Edwards, E.; Hettler, D.; Davis, B.; Bowen, R.; Bunning, M. Experimental infection of North American birds with the New York 1999 strain of West Nile virus. Emerg Infect Dis 2003, 9 (3), 311-322, doi:10.3201/eid0903.020628.
Komar, N.; Panella, N. A.; Burns, J. E.; Dusza, S. W.; Mascarenhas, T. M.; Talbot, T. O. Serologic evidence for West Nile virus infection in birds in the New York City vicinity during an outbreak in 1999. Emerg Infect Dis 2001, 7 (4), 621-625, doi:10.3201/eid0704.010403.
Kramer, L. D.; Ciota, A. T. Dissecting vectorial capacity for mosquito-borne viruses. Curr Opin Virol 2015, 15, 112-118, doi:10.1016/j.coviro.2015.10.003.
Kramer, L. D.; Li, J.; Shi, P. Y. West Nile virus. Lancet Neurol 2007, 6 (2), 171-181, doi:10.1016/S1474-4422(07)70030-3.
Kramer, L. D.; Styer, L. M.; Ebel, G. D. A global perspective on the epidemiology of West Nile virus. Annu Rev Entomol 2008, 53, 61-81, doi:10.1146/annurev.ento.53.103106.093258.
Kuhlisch, C.; Kampen, H.; Walther, D. The Asian tiger mosquito Aedes albopictus (Diptera: Culicidae) in Central Germany: Surveillance in its northernmost distribution area. Acta Trop 2018, 188, 78-85, doi:10.1016/j.actatropica.2018.08.019.
Kuno, G.; Chang, G. J.; Tsuchiya, K. R.; Karabatsos, N.; Cropp, C. B. Phylogeny of the genus Flavivirus. J Virol 1998, 72 (1), 73-83, doi:10.1128/JVI.72.1.73-83.1998.
Kuo, Y. P.; Tsai, K. N.; Luo, Y. C.; Chung, P. J.; Su, Y. W.; Teng, Y.; Wu, M. S.; Lin, Y. F.; Lai, C. Y.; Chuang, T. H.; Dai, S. S.; Tseng, F. C.; Hsieh, C. H.; Tsai, D. J.; Tsai, W. T.; Chen, C. H.; Yu, G. Y. Establishment of a mouse model for the complete mosquito-mediated transmission cycle of Zika virus. PLoS Negl Trop Dis 2018, 12 (4), e0006417, doi:10.1371/journal.pntd.0006417.
Kusne, S.; Smilack, J. Transmission of West Nile virus by organ transplantation. Liver Transpl 2005, 11 (2), 239-241, doi:10.1002/lt.20350.
Lambrechts, L.; Paaijmans, K. P.; Fansiri, T.; Carrington, L. B.; Kramer, L. D.; Thomas, M. B.; Scott, T. W. Impact of daily temperature fluctuations on dengue virus transmission by Aedes aegypti. Proc Natl Acad Sci USA 2011, 108 (18), 7460-7465, doi:10.1073/pnas.1101377108.
Lanciotti, R. S.; Roehrig, J. T.; Deubel, V.; Smith, J.; Parker, M.; Steele, K.; Crise, B.; Volpe, K. E.; Crabtree, M. B.; Scherret, J. H.; Hall, R. A.; MacKenzie, J. S.; Cropp, C. B.; Panigrahy, B.; Ostlund, E.; Schmitt, B.; Malkinson, M.; Banet, C.; Weissman, J.; Komar, N.; Savage, H. M.; Stone, W.; McNamara, T.; Gubler, D. J. Origin of the

Bibliography 95
West Nile virus responsible for an outbreak of encephalitis in the northeastern United States. Science 1999, 286 (5448), 2333-2337, doi:10.1126/science.286.5448.2333.
Langevin, S. A.; Brault, A. C.; Panella, N. A.; Bowen, R. A.; Komar, N. Variation in virulence of West Nile virus strains for house sparrows (Passer domesticus). Am J Trop Med Hyg 2005, 72 (1), 99-102.
Langevin, S. A.; Bunning, M.; Davis, B.; Komar, N. Experimental infection of chickens as candidate sentinels for West Nile virus. Emerg Infect Dis 2001, 7 (4), 726-729, doi:10.3201/eid0704.010422.
Lapointe, D. A.; Hofmeister, E. K.; Atkinson, C. T.; Porter, R. E.; Dusek, R. J. Experimental infection of Hawai'i 'Amakihi (Hemignathus virens) with West Nile virus and competence of a co-occurring vector, Culex quinquefasciatus: Potential impacts on endemic Hawaiian avifauna. J Wildl Dis 2009, 45 (2), 257-271, doi:10.7589/0090-3558-45.2.257.
Lecollinet, S.; Blanchard, Y.; Manson, C.; Lowenski, S.; Laloy, E.; Quenault, H.; Touzain, F.; Lucas, P.; Eraud, C.; Bahuon, C.; Zientara, S.; Beck, C.; Decors, A. Dual emergence of Usutu virus in common blackbirds, Eastern France, 2015. Emerg Infect Dis 2016, 22 (12), 2225, doi:10.3201/eid2212.161272.
Lee, E.; Hall, R. A.; Lobigs, M. Common E protein determinants for attenuation of glycosaminoglycan-binding variants of Japanese encephalitis and West Nile viruses. Journal of Virology 2004, 78 (15), 8271-8280, doi:10.1128/Jvi.78.15.8271-8280.2004.
Lehane, M. J. Peritrophic matrix structure and function. Annu Rev Entomol 1997, 42, 525-550, doi:10.1146/annurev.ento.42.1.525.
Lenth, R. V. Least-squares means: The R package lsmeans. J Stat Softw 2016, 69, 1-33, doi:10.18637/jss.v069.i01.
Lindenbach, B. D.; Thiel, H. J.; Rice, C. M. Flaviviridae: The viruses and their replication. In Fields Virology, 5th ed.; Knipe, D. M., Howley, P. M., Eds. Lippincott-Raven Publishers: Philadelphia, PA, USA, 2007; pp. 1101-1152, ISBN 9780781760607.
Lindsey, N. P.; Staples, J. E.; Lehman, J. A.; Fischer, M.; Centers for Disease Control and Prevention (CDC). Surveillance for human West Nile virus disease - United States, 1999-2008. MMWR Surveill Summ 2010, 59 (2), 1-17.
Lloyd-Smith, J. O.; George, D.; Pepin, K. M.; Pitzer, V. E.; Pulliam, J. R. C.; Dobson, A. P.; Hudson, P. J.; Grenfell, B. T. Epidemic dynamics at the human-animal interface. Science 2009, 326 (5958), 1362-1367, doi:10.1126/science.1177345.
Lobigs, M.; Diamond, M. S. Feasibility of cross-protective vaccination against flaviviruses of the Japanese encephalitis serocomplex. Expert Rev Vaccines 2012, 11 (2), 177-187, doi:10.1586/Erv.11.180.
Long, M.T. Flavivirus infections. In Equine Infectious Diseases, 2nd ed.; Sellon, D. C., Long, M. T., Eds. Saunders Elsevier: St. Louis, MO, USA, 2006; pp. 198–206, ISBN 978-1-4557-0891-8.
Lopes, H.; Redig, P.; Glaser, A.; Armien, A.; Wünschmann, A. Clinical findings, lesions, and viral antigen distribution in great gray owls (Strix nebulosa) and barred owls (Strix varia) with spontaneous West Nile virus infection. Avian Dis 2007, 51 (1), 140-145, doi:10.1637/0005-2086(2007)051[0140:CFLAVA]2.0.CO;2.
Lühken, R.; Jöst, H.; Cadar, D.; Thomas, S. M.; Bosch, S.; Tannich, E.; Becker, N.; Ziegler, U.; Lachmann, L.; Schmidt-Chanasit, J. Distribution of Usutu virus in Germany and its effect on breeding bird populations. Emerg Infect Dis 2017, 23 (12), 1994-2001, doi:10.3201/eid2312.171257.

96 Bibliography
Lühken, R.; Steinke, S.; Leggewie, M.; Tannich, E.; Krüger, A.; Becker, S.; Kiel, E. Physico-chemical characteristics of Culex pipiens sensu lato and Culex torrentium (Diptera: Culicidae) breeding sites in Germany. J Med Entomol 2015, 52 (5), 932-936, doi:10.1093/jme/tjv070.
Lupulovic, D.; Martín-Acebes, M. A.; Lazic, S.; Alonso-Padilla, J.; Blázquez, A. B.; Escribano-Romero, E.; Petrovic, T.; Saiz, J. C. First serological evidence of West Nile virus activity in horses in Serbia. Vector Borne Zoonotic Dis 2011, 11 (9), 1303-1305, doi:10.1089/vbz.2010.0249.
Madden, K. West Nile virus infection and its neurological manifestations. Clin Med Res 2003, 1 (2), 145-150, doi:10.3121/cmr.1.2.145.
Magurano, F.; Remoli, M. E.; Baggieri, M.; Fortuna, C.; Marchi, A.; Fiorentini, C.; Bucci, P.; Benedetti, E.; Ciufolini, M. G.; Rizzo, C.; Piga, S.; Salcuni, P.; Rezza, G.; Nicoletti, L. Circulation of West Nile virus lineage 1 and 2 during an outbreak in Italy. Clin Microbiol Infect 2012, 18 (12), E545-E547, doi:10.1111/1469-0691.12018.
Malan, A. K.; Martins, T. B.; Hill, H. R.; Litwin, C. M. Evaluations of commercial West Nile virus immunoglobulin G (IgG) and IgM enzyme immunoassays show the value of continuous validation. J Clin Microbiol 2004, 42 (2), 727-733, doi:10.1128/jcm.42.2.727-733.2004.
Malkinson, M.; Banet, C.; Weisman, Y.; Pokamunski, S.; King, R.; Drouet, M. T.; Deubel, V. Introduction of West Nile virus in the Middle East by migrating white storks. Emerg Infect Dis 2002, 8 (4), 392-397, doi:10.3201/eid0804.010217.
Mancini, G.; Montarsi, F.; Calzolari, M.; Capelli, G.; Dottori, M.; Ravagnan, S.; Lelli, D.; Chiari, M.; Santilli, A.; Quaglia, M.; Quaglia, M.; Federici, V.; Monaco, F.; Goffredo, M.; Savini, G. Mosquito species involved in the circulation of West Nile and Usutu viruses in Italy. Vet Ital 2017, 53 (2), 97-110, doi:10.12834/VetIt.114.933.4764.2.
Mannasse, B.; Mendelson, E.; Orshan, L.; Mor, O.; Shalom, U.; Yeger, T.; Lustig, Y. Usutu virus RNA in Mosquitoes, Israel, 2014-2015. Emerg Infect Dis 2017, 23 (10), 1699-1702, doi:10.3201/eid2310.171017.
Mansfield, K. L.; Hernández-Triana, L. M.; Banyard, A. C.; Fooks, A. R.; Johnson, N. Japanese encephalitis virus infection, diagnosis and control in domestic animals. Vet Microbiol 2017, 201, 85-92, doi:10.1016/j.vetmic.2017.01.014.
Marfin, A. A.; Petersen, L. R.; Eidson, M.; Miller, J.; Hadler, J.; Farello, C.; Werner, B.; Campbell, G. L.; Layton, M.; Smith, P.; Bresnitz, E.; Cartter, M.; Scaletta, J.; Obiri, G.; Bunning, M.; Craven, R. C.; Roehrig, J. T.; Julian, K. G.; Hinten, S. R.; Gubler, D. J.; ArboNET Cooperative Surveillance Group. Widespread West Nile virus activity, eastern United States, 2000. Emerg Infect Dis 2001, 7 (4), 730-735, doi:10.3201/eid0704.010423.
Marra, P. P.; Griffing, S.; Caffrey, C.; Kilpatrick, A. M.; McLean, R.; Brand, C.; Saito, E.; Dupuis, A. P.; Kramer, L.; Novak, R. West Nile virus and wildlife. Bioscience 2004, 54 (5), 393-402, doi:10.1641/0006-3568(2004)054[0393:Wnvaw]2.0.Co;2.
Mayr, A.; Bachmann, P. A.; Bibrack, B.; Wittmann, G. Quantitative Bestimmung der Virusinfektiosität (Virustitration). In Virologische Arbeitsmethoden, Band I (Zellkulturen - Bebrütete Hühnereier - Versuchstiere), 1st ed.; Mayr, A., Bachmann, P. A., Bibrack, B., Wittmann, G., Eds. Gustav Fischer Verlag: Jena, Germany, 1974; pp. 35-39, ISBN 9783437301759.

Bibliography 97
McIntosh, B. M.; Jupp, P. G.; Dickinson, D. B.; McGillivray, G. M.; Sweetnam, J. Ecological studies on Sindbis and West Nile viruses in South Africa. I. Viral activity as revealed by infection of mosquitoes and sentinel fowls. S Afr J Med Sci 1967, 32 (1), 1-14.
McIntosh, B. M. Usutu (SAAr 1776), nouvel arbovirus du groupe B. Int Cat Arboviruses 1985, 3, 1059-1060.
McLean, R. G.; Ubico, S. R.; Bourne, D.; Komar, N. West Nile virus in livestock and wildlife. In Japanese Encephalitis and West Nile Viruses. Current Topics in Microbiology and Immunology, Mackenzie, J. S., Barrett, A. D. T., Deubel, V., Eds. Springer-Verlag: Berlin/Heidelberg, Germany, 2002; Vol. 267, pp. 271-308, doi: 10.1007/978-3-642-59403-8_14.
McMinn, P. C. The molecular basis of virulence of the encephalitogenic flaviviruses. J Gen Virol 1997, 78 (Pt 11), 2711-2722, doi:10.1099/0022-1317-78-11-2711.
Medlock, J. M.; Hansford, K. M.; Schaffner, F.; Versteirt, V.; Hendrickx, G.; Zeller, H.; Van Bortel, W. A review of the invasive mosquitoes in Europe: Ecology, public health risks, and control options. Vector Borne Zoonotic Dis 2012, 12 (6), 435-447, doi:10.1089/vbz.2011.0814.
Meister, T.; Lussy, H.; Bakonyi, T.; Sikutová, S.; Rudolf, I.; Vogl, W.; Winkler, H.; Frey, H.; Hubálek, Z.; Nowotny, N.; Weissenböck, H. Serological evidence of continuing high Usutu virus (Flaviviridae) activity and establishment of herd immunity in wild birds in Austria. Vet Microbiol 2008, 127 (3-4), 237-248, doi:10.1016/j.vetmic.2007.08.023.
Merino-Ramos, T.; Blázquez, A. B.; Escribano-Romero, E.; Cañas-Arranz, R.; Sobrino, F.; Saiz, J. C.; Martín-Acebes, M. A. Protection of a single dose West Nile virus recombinant subviral particle vaccine against lineage 1 or 2 strains and analysis of the cross-reactivity with Usutu virus. Plos One 2014, 9 (9), e108056, doi:10.1371/journal.pone.0108056.
Michel, F.; Fischer, D.; Eiden, M.; Fast, C.; Reuschel, M.; Müller, K.; Rinder, M.; Urbaniak, S.; Brandes, F.; Schwehn, R.; Lühken, R.; Groschup, M. H.; Ziegler, U. West Nile virus and Usutu virus monitoring of wild birds in Germany. Int J Environ Res Public Health 2018, 15 (1), 171, doi:10.3390/ijerph15010171.
Michel, F.; Sieg, M.; Fischer, D.; Keller, M.; Eiden, M.; Reuschel, M.; Schmidt, V.; Schwehn, R.; Rinder, M.; Urbaniak, S.; Müller, K.; Schmoock, M.; Lühken, R.; Wysocki, P.; Fast, C.; Lierz, M.; Korbel, R.; Vahlenkamp, T. W.; Groschup, M. H.; Ziegler, U. Evidence for West Nile virus and Usutu virus infections in wild and resident birds in Germany, 2017 and 2018. Viruses 2019, 11 (7), 674, doi:10.3390/v11070674.
Micieli, M. V.; Glaser, R. L. Somatic Wolbachia (Rickettsiales: Rickettsiaceae) levels in Culex quinquefasciatus and Culex pipiens (Diptera: Culicidae) and resistance to West Nile virus infection. J Med Entomol 2014, 51 (1), 189-199, doi:10.1603/me13152.
Miller, D. L.; Mauel, M. J.; Baldwin, C.; Burtle, G.; Ingram, D.; Hines, M. E., 2nd; Frazier, K. S. West Nile virus in farmed alligators. Emerg Infect Dis 2003, 9 (7), 794-799, doi:10.3201/eid0907.030085.
Modis, Y.; Ogata, S.; Clements, D.; Harrison, S. C. Structure of the dengue virus envelope protein after membrane fusion. Nature 2004, 427 (6972), 313-319, doi:10.1038/nature02165.
Monath, T. P.; Cropp, C. B.; Harrison, A. K. Mode of entry of a neurotropic arbovirus into the central nervous system. Reinvestigation of an old controversy. Lab Invest 1983, 48 (4), 399-410.

98 Bibliography
Montagnaro, S.; Piantedosi, D.; Ciarcia, R.; Loponte, R.; Veneziano, V.; Fusco, G.; Amoroso, M. G.; Ferrara, G.; Damiano, S.; Iovane, G.; Pagnini, U. Serological evidence of mosquito-borne flaviviruses circulation in hunting dogs in Campania region, Italy. Vector Borne Zoonotic Dis 2019, 19 (2), 142-147, doi:10.1089/vbz.2018.2337.
Morales-Betoulle, M. E.; Morales, H.; Blitvich, B. J.; Powers, A. M.; Davis, E. A.; Klein, R.; Cordon-Rosales, C. West Nile virus in horses, Guatemala. Emerg Infect Dis 2006, 12 (6), 1038-1039, doi:10.3201/eid1206.051615.
Morales, M. A.; Barrandeguy, M.; Fabbri, C.; Garcia, J. B.; Vissani, A.; Trono, K.; Gutierrez, G.; Pigretti, S.; Menchaca, H.; Garrido, N.; Taylor, N.; Fernandez, F.; Levis, S.; Enría, D. West Nile virus isolation from equines in Argentina, 2006. Emerg Infect Dis 2006, 12 (10), 1559-1561, doi:10.3201/eid1210.060852.
Morrey, J. D.; Olsen, A. L.; Siddharthan, V.; Motter, N. E.; Wang, H.; Taro, B. S.; Chen, D.; Ruffner, D.; Hall, J. O. Increased blood-brain barrier permeability is not a primary determinant for lethality of West Nile virus infection in rodents. J Gen Virol 2008, 89 (Pt 2), 467-473, doi:10.1099/vir.0.83345-0.
Mossel, E. C.; Crabtree, M. B.; Mutebi, J. P.; Lutwama, J. J.; Borland, E. M.; Powers, A. M.; Miller, B. R. Arboviruses isolated from mosquitoes collected in Uganda, 2008-2012. J Med Entomol 2017, 54 (5), 1403-1409, doi:10.1093/jme/tjx120.
Mostashari, F.; Bunning, M. L.; Kitsutani, P. T.; Singer, D. A.; Nash, D.; Cooper, M. J.; Katz, N.; Liljebjelke, K. A.; Biggerstaff, B. J.; Fine, A. D.; Layton, M. C.; Mullin, S. M.; Johnson, A. J.; Martin, D. A.; Hayes, E. B.; Campbell, G. L. Epidemic West Nile encephalitis, New York, 1999: Results of a household-based seroepidemiological survey. Lancet 2001, 358 (9278), 261-264, doi:10.1016/S0140-6736(01)05480-0.
Murdock, C. C.; Paaijmans, K. P.; Cox-Foster, D.; Read, A. F.; Thomas, M. B. Rethinking vector immunology: The role of environmental temperature in shaping resistance. Nat Rev Microbiol 2012, 10 (12), 869-876, doi:10.1038/nrmicro2900.
Murgue, B.; Murri, S.; Triki, H.; Deubel, V.; Zeller, H. G. West Nile in the Mediterranean basin: 1950-2000. Ann N Y Acad Sci 2001a, 951, 117-126, doi:10.1111/j.1749-6632.2001.tb02690.x.
Murgue, B.; Murri, S.; Zientara, S.; Durand, B.; Durand, J. P.; Zeller, H. West Nile outbreak in horses in southern France, 2000: The return after 35 years. Emerg Infect Dis 2001b, 7 (4), 692-696, doi:10.3201/eid0704.010417.
Muturi, E. J.; Dunlap, C.; Ramirez, J. L.; Rooney, A. P.; Kim, C. H. Host blood-meal source has a strong impact on gut microbiota of Aedes aegypti. FEMS Microbiol Ecol 2019, 95 (1), doi:10.1093/femsec/fiy213.
Nash, D.; Mostashari, F.; Fine, A.; Miller, J.; O'Leary, D.; Murray, K.; Huang, A.; Rosenberg, A.; Greenberg, A.; Sherman, M.; Wong, S.; Layton, M.; West Nile Outbreak Response Working Group. The outbreak of West Nile virus infection in the New York City area in 1999. N Engl J Med 2001, 344 (24), 1807-1814, doi:10.1056/NEJM200106143442401.
Nelms, B. M.; Fechter-Leggett, E.; Carroll, B. D.; Macedo, P.; Kluh, S.; Reisen, W. K. Experimental and natural vertical transmission of West Nile virus by California Culex (Diptera: Culicidae) mosquitoes. J Med Entomol 2013, 50 (2), 371-378, doi:10.1603/ME12264.
Nemeth, N.; Gould, D.; Bowen, R.; Komar, N. Natural and experimental West Nile virus infection in five raptor species. J Wildl Dis 2006a, 42 (1), 1-13, doi:10.7589/0090-3558-42.1.1.

Bibliography 99
Nemeth, N. M.; Hahn, D. C.; Gould, D. H.; Bowen, R. A. Experimental West Nile virus infection in Eastern screech owls (Megascops asio). Avian Dis 2006b, 50 (2), 252-258, doi:10.1637/7466-110105R1.1.
Nemeth, N. M.; Kratz, G. E.; Bates, R.; Scherpelz, J. A.; Bowen, R. A.; Komar, N. Clinical evaluation and outcomes of naturally acquired West Nile virus infection in raptors. J Zoo Wildl Med 2009a, 40 (1), 51-63, doi:10.1638/2007-0109.1.
Nemeth, N. M.; Oesterle, P. T.; Bowen, R. A. Humoral immunity to West Nile virus is long-lasting and protective in the house sparrow (Passer domesticus). Am J Trop Med Hyg 2009b, 80 (5), 864-869, doi:10.4269/ajtmh.2009.80.864.
Ng, T.; Hathaway, D.; Jennings, N.; Champ, D.; Chiang, Y. W.; Chu, H. J. Equine vaccine for West Nile virus. Dev Biol (Basel) 2003, 114, 221-227.
Nikolay, B. A review of West Nile and Usutu virus co-circulation in Europe: How much do transmission cycles overlap? Trans R Soc Trop Med Hyg 2015, 109 (10), 609-618, doi:10.1093/trstmh/trv066.
Nikolay, B.; Diallo, M.; Boye, C. S.; Sall, A. A. Usutu virus in Africa. Vector Borne Zoonotic Dis 2011, 11 (11), 1417-1423, doi:10.1089/vbz.2011.0631.
Nikolay, B.; Diallo, M.; Faye, O.; Boye, C. S.; Sall, A. A. Vector competence of Culex neavei (Diptera: Culicidae) for Usutu virus. Am J Trop Med Hyg 2012, 86 (6), 993-996, doi:10.4269/ajtmh.2012.11-0509.
Nir, Y.; Beemer, A.; Goldwasser, R. A. West Nile virus infection in mice following exposure to a viral aerosol. Br J Exp Pathol 1965, 46 (4), 443-449.
Okuda, K.; de Souza Caroci, A.; Ribolla, P. E.; de Bianchi, A. G.; Bijovsky, A. T. Functional morphology of adult female Culex quinquefasciatus midgut during blood digestion. Tissue Cell 2002, 34 (3), 210-219, doi:10.1016/s0040-8166(02)00032-0.
Osório, H. C.; Zé-Zé, L.; Amaro, F.; Nunes, A.; Alves, M. J. Sympatric occurrence of Culex pipiens (Diptera, Culicidae) biotypes pipiens, molestus and their hybrids in Portugal, Western Europe: Feeding patterns and habitat determinants. Med Vet Entomol 2014, 28 (1), 103-109, doi:10.1111/mve.12020.
Ostlund, E. N.; Crom, R. L.; Pedersen, D. D.; Johnson, D. J.; Williams, W. O.; Schmitt, B. J. Equine West Nile encephalitis, United States. Emerg Infect Dis 2001, 7 (4), 665-669, doi:10.3201/eid0704.010412.
Owen, J.; Moore, F.; Panella, N.; Edwards, E.; Bru, R.; Hughes, M.; Komar, N. Migrating birds as dispersal vehicles for West Nile virus. Ecohealth 2006, 3 (2), 79-85, doi:10.1007/s10393-006-0025-9.
Pantheir, R.; Hannoun, C.; Oudar, J.; Beytout, D.; Corniou, B.; Joubert, L.; Guillon, J. C.; Mouchet, J. [Isolation of West Nile virus in a Camarge horse with encephalomyelitis]. C R Acad Hebd Seances Acad Sci D 1966, 262 (11), 1308-1310.
Paradkar, P. N.; Trinidad, L.; Voysey, R.; Duchemin, J. B.; Walker, P. J. Secreted Vago restricts West Nile virus infection in Culex mosquito cells by activating the Jak-STAT pathway. Proc Natl Acad Sci USA 2012, 109 (46), 18915-18920, doi:10.1073/pnas.1205231109.
Pauli, G.; Bauerfeind, U.; Blümel, J.; Burger, R.; Drosten, C.; Gröner, A.; Gürtler, L.; Heiden, M.; Hildebrandt, M.; Jansen, B.; Offergeld, R.; Seitz, R.; Schlenkrich, U.; Schottstedt, V.; Strobel, J.; Willkommen, H. Usutu virus. Transfus Med Hemother 2014, 41 (1), 73-82, doi:10.1159/000357106.
Pecorari, M.; Longo, G.; Gennari, W.; Grottola, A.; Sabbatini, A.; Tagliazucchi, S.; Savini, G.; Monaco, F.; Simone, M.; Lelli, R.; Rumpianesi, F. First human case of Usutu virus

100 Bibliography
neuroinvasive infection, Italy, August-September 2009. Euro Surveill 2009, 14 (50), pii=19446, doi:10.2807/ese.14.50.19446-en.
Pérez-Ramírez, E.; Llorente, F.; Jiménez-Clavero, M. A. Experimental infections of wild birds with West Nile virus. Viruses 2014, 6 (2), 752-781, doi:10.3390/v6020752.
Pervanidou, D.; Detsis, M.; Danis, K.; Mellou, K.; Papanikolaou, E.; Terzaki, I.; Baka, A.; Veneti, L.; Vakali, A.; Dougas, G.; Politis, C.; Stamoulis, K.; Tsiodras, S.; Georgakopoulou, T.; Papa, A.; Tsakris, A.; Kremastinou, J.; Hadjichristodoulou, C. West Nile virus outbreak in humans, Greece, 2012: Third consecutive year of local transmission. Euro Surveill 2014, 19 (13), pii=20758, doi:10.2807/1560-7917.es2014.19.13.20758.
Petersen, L. R.; Marfin, A. A. West Nile virus: A primer for the clinician. Ann Intern Med 2002, 137 (3), 173-179, doi:10.7326/0003-4819-137-3-200208060-00009.
Petersen, L. R.; Marfin, A. A.; Gubler, D. J. West Nile virus. JAMA 2003, 290 (4), 524-528, doi:10.1001/jama.290.4.524.
Petersen, L. R.; Roehrig, J. T.; Hughes, J. M. West Nile virus encephalitis. N Engl J Med 2002, 347 (16), 1225-1226, doi:10.1056/NEJMo020128.
Pfeffer, M.; Dobler, G. Emergence of zoonotic arboviruses by animal trade and migration. Parasit Vectors 2010, 3 (1), 35, doi:10.1186/1756-3305-3-35.
Pierro, A.; Gaibani, P.; Spadafora, C.; Ruggeri, D.; Randi, V.; Parenti, S.; Finarelli, A. C.; Rossini, G.; Landini, M. P.; Sambri, V. Detection of specific antibodies against West Nile and Usutu viruses in healthy blood donors in northern Italy, 2010-2011. Clin Microbiol Infect 2013, 19 (10), E451-453, doi:10.1111/1469-0691.12241.
Pierson, T. C.; Diamond, M. S. The emergence of Zika virus and its new clinical syndromes. Nature 2018, 560 (7720), 573-581, doi:10.1038/s41586-018-0446-y.
Pierson, T. C.; Diamond, M. S. The continued threat of emerging flaviviruses. Nat Microbiol 2020, 5 (6), 796-812, doi:10.1038/s41564-020-0714-0.
Pinho dos Reis, V. The role of integrins in flavivirus infection. Ernst-Moritz-Arndt-Universität Greifswald: Greifswald, Germany, 2018.
Platonov, A. E.; Karan, L. S.; Shopenskaia, T. A.; Fedorova, M. V.; Koliasnikova, N. M.; Rusakova, N. M.; Shishkina, L. V.; Arshba, T. E.; Zhuravlev, V. I.; Govorukhina, M. V.; Valentseva, A. A.; Shipulin, G. A. [Genotyping of West Nile fever virus strains circulating in southern Russia as an epidemiological investigation method: principles and results]. Zh Mikrobiol Epidemiol Immunobiol 2011, (2), 29-37.
Platonov, A. E.; Shipulin, G. A.; Shipulina, O. Y.; Tyutyunnik, E. N.; Frolochkina, T. I.; Lanciotti, R. S.; Yazyshina, S.; Platonova, O. V.; Obukhov, I. L.; Zhukov, A. N.; Vengerov, Y. Y.; Pokrovskii, V. I. Outbreak of West Nile virus infection, Volgograd Region, Russia, 1999. Emerg Infect Dis 2001, 7 (1), 128-132, doi:10.3201/eid0701.010118.
Poidinger, M.; Hall, R. A.; Mackenzie, J. S. Molecular characterization of the Japanese encephalitis serocomplex of the flavivirus genus. Virology 1996, 218 (2), 417-421, doi:10.1006/viro.1996.0213.
Popović, N.; Milošević, B.; Urošević, A.; Poluga, J.; Lavadinović, L.; Nedelijković, J.; Jevtović, D.; Dulović, O. Outbreak of West Nile virus infection among humans in Serbia, August to October 2012. Euro Surveill 2013, 18 (43), pii=20613, doi:10.2807/1560-7917.Es2013.18.43.20613.
Porter, M. B.; Long, M. T.; Getman, L. M.; Giguère, S.; MacKay, R. J.; Lester, G. D.; Alleman, A. R.; Wamsley, H. L.; Franklin, R. P.; Jacks, S.; Buergelt, C. D.; Detrisac,

Bibliography 101
C. J. West Nile virus encephalomyelitis in horses: 46 cases (2001). J Am Vet Med Assoc 2003, 222 (9), 1241-1247, doi:10.2460/javma.2003.222.1241.
Prasad, A. N.; Brackney, D. E.; Ebel, G. D. The role of innate immunity in conditioning mosquito susceptibility to West Nile virus. Viruses 2013, 5 (12), 3142-3170, doi:10.3390/v5123142.
Puggioli, A.; Bonilauri, P.; Calzolari, M.; Lelli, D.; Carrieri, M.; Urbanelli, S.; Pudar, D.; Bellini, R. Does Aedes albopictus (Diptera: Culicidae) play any role in Usutu virus transmission in Northern Italy? Experimental oral infection and field evidences. Acta Trop 2017, 172, 192-196, doi:10.1016/j.actatropica.2017.05.006.
R Development Core Team. R: A language and environment for statistical computing; Global Biodiversity Information Facility: Vienna, Austria, 2019.
Rathore, A. P. S.; St. John, A. L. Cross-reactive immunity among flaviviruses. Front Immunol 2020, 11 (334), doi:10.3389/fimmu.2020.00334.
Reisen, W. K. Ecology of West Nile virus in North America. Viruses 2013, 5 (9), 2079-2105, doi:10.3390/v5092079.
Reiter, P. Yellow fever and dengue: A threat to Europe? Euro Surveill 2010, 15 (10), pii=19509, doi:10.2807/ese.15.10.19509-en.
Ribeiro, J. M. Blood-feeding in mosquitoes: Probing time and salivary gland anti-haemostatic activities in representatives of three genera (Aedes, Anopheles, Culex). Med Vet Entomol 2000, 14 (2), 142-148, doi:10.1046/j.1365-2915.2000.00227.x.
Ribeiro, J. M.; Francischetti, I. M. Role of arthropod saliva in blood feeding: Sialome and post-sialome perspectives. Annu Rev Entomol 2003, 48, 73-88, doi:10.1146/annurev.ento.48.060402.102812.
Rice, C. M.; Lenches, E. M.; Eddy, S. R.; Shin, S. J.; Sheets, R. L.; Strauss, J. H. Nucleotide sequence of yellow fever virus: Implications for flavivirus gene expression and evolution. Science 1985, 229 (4715), 726-733, doi:10.1126/science.4023707.
Richards, S. L.; Anderson, S. L.; Lord, C. C.; Tabachnick, W. J. Impact of West Nile virus dose and incubation period on vector competence of Culex nigripalpus (Diptera: Culicidae). Vector Borne Zoonotic Dis 2011, 11 (11), 1487-1491, doi:10.1089/vbz.2010.0229.
Richards, S. L.; Lord, C. C.; Pesko, K. N.; Tabachnick, W. J. Environmental and biological factors influencing Culex pipiens quinquefasciatus (Diptera: Culicidae) vector competence for West Nile virus. Am J Trop Med Hyg 2010, 83 (1), 126-134, doi:10.4269/ajtmh.2010.09-0776.
Rios, M.; Zhang, M. J.; Grinev, A.; Srinivasan, K.; Daniel, S.; Wood, O.; Hewlett, I. K.; Dayton, A. I. Monocytes-macrophages are a potential target in human infection with West Nile virus through blood transfusion. Transfusion 2006, 46 (4), 659-667, doi:10.1111/j.1537-2995.2006.00769.x.
Rizzoli, A.; Jiménez-Clavero, M. A.; Barzon, L.; Cordioli, P.; Figuerola, J.; Koraka, P.; Martina, B.; Moreno, A.; Nowotny, N.; Pardigon, N.; Sanders, N.; Ulbert, S.; Tenorio, A. The challenge of West Nile virus in Europe: Knowledge gaps and research priorities. Euro Surveill 2015, 20 (20), pii=21135, doi:10.2807/1560-7917.es2015.20.20.21135.
Roehrig, J. T. West nile virus in the United States - a historical perspective. Viruses 2013, 5 (12), 3088-3108, doi:10.3390/v5123088.
Roesch, F.; Fajardo, A.; Moratorio, G.; Vignuzzi, M. Usutu virus: An arbovirus on the rise. Viruses 2019, 11 (7), 640, doi:10.3390/v11070640.

102 Bibliography
Romoser, W. S.; Wasieloski, L. P., Jr.; Pushko, P.; Kondig, J. P.; Lerdthusnee, K.; Neira, M.; Ludwig, G. V. Evidence for arbovirus dissemination conduits from the mosquito (Diptera: Culicidae) midgut. J Med Entomol 2004, 41 (3), 467-475, doi:10.1603/0022-2585-41.3.467.
Root, J. J.; Bosco-Lauth, A. M. West Nile virus associations in wild mammals: An update. Viruses 2019, 11 (5), 459, doi:10.3390/v11050459.
Rosen, L. Dengue in Greece in 1927 and 1928 and the pathogenesis of dengue hemorrhagic fever: New data and a different conclusion. Am J Trop Med Hyg 1986, 35 (3), 642-653, doi:10.4269/ajtmh.1986.35.642.
Rossi, S. L.; Ross, T. M.; Evans, J. D. West Nile virus. Clin Lab Med 2010, 30 (1), 47-65, doi:10.1016/j.cll.2009.10.006.
Rossini, G.; Carletti, F.; Bordi, L.; Cavrini, F.; Gaibani, P.; Landini, M. P.; Pierro, A.; Capobianchi, M. R.; Di Caro, A.; Sambri, V. Phylogenetic analysis of West Nile virus isolates, Italy, 2008-2009. Emerg Infect Dis 2011, 17 (5), 903-906, doi:10.3201/eid1705.101569.
Rudolf, M.; Czajka, C.; Börstler, J.; Melaun, C.; Jöst, H.; von Thien, H.; Badusche, M.; Becker, N.; Schmidt-Chanasit, J.; Krüger, A.; Tannich, E.; Becker, S. First nationwide surveillance of Culex pipiens complex and Culex torrentium mosquitoes demonstrated the presence of Culex pipiens biotype pipiens/molestus hybrids in Germany. PLoS One 2013, 8 (9), e71832, doi:10.1371/journal.pone.0071832.
Russell, R. C. Mosquito-borne arboviruses in Australia: The current scene and implications of climate change for human health. Int J Parasitol 1998, 28 (6), 955-969, doi:10.1016/s0020-7519(98)00053-8.
Saiz, J. C.; Blázquez, A. B. Usutu virus: Current knowledge and future perspectives. Virus Adapt Treat 2017, 9, 27-40, doi:10.2147/VAAT.S123619.
Salas-Benito, J. S.; De Nova-Ocampo, M. Viral interference and persistence in mosquito-borne flaviviruses. J Immunol Res 2015, 2015, 873404, doi:10.1155/2015/873404.
Salazar, P.; Traub-Dargatz, J. L.; Morley, P. S.; Wilmot, D. D.; Steffen, D. J.; Cunningham, W. E.; Salman, M. D. Outcome of equids with clinical signs of West Nile virus infection and factors associated with death. J Am Vet Med Assoc 2004, 225 (2), 267-274, doi:10.2460/javma.2004.225.267.
Sambri, V.; Capobianchi, M.; Charrel, R.; Fyodorova, M.; Gaibani, P.; Gould, E.; Niedrig, M.; Papa, A.; Pierro, A.; Rossini, G.; Varani, S.; Vocale, C.; Landini, M. P. West Nile virus in Europe: Emergence, epidemiology, diagnosis, treatment, and prevention. Clin Microbiol Infect 2013, 19 (8), 699-704, doi:10.1111/1469-0691.12211.
Sampson, B. A.; Ambrosi, C.; Charlot, A.; Reiber, K.; Veress, J. F.; Armbrustmacher, V. The pathology of human West Nile virus infection. Hum Pathol 2000, 31 (5), 527-531, doi:10.1053/hp.2000.8047.
Samuel, M. A.; Diamond, M. S. Pathogenesis of West Nile virus infection: A balance between virulence, innate and adaptive immunity, and viral evasion. J Virol 2006, 80 (19), 9349-9360, doi:10.1128/Jvi.01122-06.
Samuel, M. A.; Wang, H.; Siddharthan, V.; Morrey, J. D.; Diamond, M. S. Axonal transport mediates West Nile virus entry into the central nervous system and induces acute flaccid paralysis. Proc Natl Acad Sci USA 2007, 104 (43), 17140-17145, doi:10.1073/pnas.0705837104.
Santini, M.; Vilibic-Cavlek, T.; Barsic, B.; Barbic, L.; Savic, V.; Stevanovic, V.; Listes, E.; Di Gennaro, A.; Savini, G. First cases of human Usutu virus neuroinvasive infection

Bibliography 103
in Croatia, August-September 2013: Clinical and laboratory features. J Neurovirol 2015, 21 (1), 92-97, doi:10.1007/s13365-014-0300-4.
Sardelis, M. R.; Turell, M. J.; O'Guinn, M. L.; Andre, R. G.; Roberts, D. R. Vector competence of three North American strains of Aedes albopictus for West Nile virus. J Am Mosq Control Assoc 2002, 18 (4), 284-289.
Savini, G.; Monaco, F.; Terregino, C.; Di Gennaro, A.; Bano, L.; Pinoni, C.; De Nardi, R.; Bonilauri, P.; Pecorari, M.; Di Gialleonardo, L.; Bonfanti, L.; Polci, A.; Calistri, P.; Lelli, R. Usutu virus in Italy: An emergence or a silent infection? Vet Microbiol 2011, 151 (3-4), 264-274, doi:10.1016/j.vetmic.2011.03.036.
Schaffner, F.; Chouin, S.; Guilloteau, J. First record of Ochlerotatus (Finlaya) japonicus japonicus (Theobald, 1901) in metropolitan France. J Am Mosq Control Assoc 2003, 19 (1), 1-5.
Scheuch, D. E.; Schäfer, M.; Eiden, M.; Heym, E. C.; Ziegler, U.; Walther, D.; Schmidt-Chanasit, J.; Keller, M.; Groschup, M. H.; Kampen, H. Detection of Usutu, Sindbis, and Batai viruses in mosquitoes (Diptera: Culicidae) collected in Germany, 2011-2016. Viruses 2018, 10 (7), 389, doi:10.3390/v10070389.
Schmidt, J. R.; Elmansoury, H. K. Natural and experimental infection of Egyptian equines with West Nile virus. Ann Trop Med Parasitol 1963, 57, 415-427, doi:10.1080/00034983.1963.11686194.
Schneider, B. S.; Soong, L.; Girard, Y. A.; Campbell, G.; Mason, P.; Higgs, S. Potentiation of West Nile encephalitis by mosquito feeding. Viral Immunol 2006, 19 (1), 74-82, doi:10.1089/vim.2006.19.74.
Scholle, F.; Girard, Y. A.; Zhao, Q.; Higgs, S.; Mason, P. W. Trans-packaged West Nile virus-like particles: Infectious properties in vitro and in infected mosquito vectors. J Virol 2004, 78 (21), 11605-11614, doi:10.1128/JVI.78.21.11605-11614.2004.
Scholte, E. J.; Den Hartog, W.; Dik, M.; Schoelitsz, B.; Brooks, M.; Schaffner, F.; Foussadier, R.; Braks, M.; Beeuwkes, J. Introduction and control of three invasive mosquito species in the Netherlands, July-October 2010. Euro Surveill 2010, 15 (45), pii=19710, doi:10.2807/ese.15.45.19710-en.
Schuler, L. A.; Khaitsa, M. L.; Dyer, N. W.; Stoltenow, C. L. Evaluation of an outbreak of West Nile virus infection in horses: 569 cases (2002). J Am Vet Med Assoc 2004, 225 (7), 1084-1089, doi:10.2460/javma.2004.225.1084.
Sejvar, J. J. The long-term outcomes of human West Nile virus infection. Clin Infect Dis 2007, 44 (12), 1617-1624, doi:10.1086/518281.
Senne, D. A.; Pedersen, J. C.; Hutto, D. L.; Taylor, W. D.; Schmitt, B. J.; Panigrahy, B. Pathogenicity of West Nile virus in chickens. Avian Dis 2000, 44 (3), 642-649.
Shahhosseini, N.; Friedrich, J.; Moosa-Kazemi, S. H.; Sedaghat, M. M.; Kayedi, M. H.; Tannich, E.; Schmidt-Chanasit, J.; Lühken, R. Host-feeding patterns of Culex mosquitoes in Iran. Parasit Vectors 2018, 11 (1), 669, doi:10.1186/s13071-018-3237-2.
Shepard, D. S.; Undurraga, E. A.; Halasa, Y. A.; Stanaway, J. D. The global economic burden of dengue: A systematic analysis. Lancet Infect Dis 2016, 16 (8), 935-941, doi:10.1016/S1473-3099(16)00146-8.
Shieh, W. J.; Guarner, J.; Layton, M.; Fine, A.; Miller, J.; Nash, D.; Campbell, G. L.; Roehrig, J. T.; Gubler, D. J.; Zaki, S. R. The role of pathology in an investigation of an outbreak of West Nile encephalitis in New York, 1999. Emerg Infect Dis 2000, 6 (4), 370-372, doi:10.3201/eid0604.000407.

104 Bibliography
Sieg, M.; Schmidt, V.; Ziegler, U.; Keller, M.; Höper, D.; Heenemann, K.; Rückner, A.; Nieper, H.; Muluneh, A.; Groschup, M. H.; Vahlenkamp, T. W. Outbreak and cocirculation of three different Usutu virus strains in Eastern Germany. Vector Borne Zoonotic Dis 2017, 17 (9), 662-664, doi:10.1089/vbz.2016.2096.
Simmonds, P.; Becher, P.; Bukh, J.; Gould, E. A.; Meyers, G.; Monath, T.; Muerhoff, S.; Pletnev, A.; Rico-Hesse, R.; Smith, D. B.; Stapleton, J. T.; Consortium, ICTV Report. ICTV Virus Taxonomy Profile: Flaviviridae. J Gen Virol 2017, 98 (1), 2-3, doi:10.1099/jgv.0.000672.
Simonin, Y.; Sillam, O.; Carles, M. J.; Gutierrez, S.; Gil, P.; Constant, O.; Martin, M. F.; Girard, G.; Van de Perre, P.; Salinas, S.; Leparc-Goffart, I.; Foulongne, V. Human Usutu virus infection with atypical neurologic presentation, Montpellier, France, 2016. Emerg Infect Dis 2018, 24 (5), 875-878, doi:10.3201/eid2405.171122.
Sirbu, A.; Ceianu, C. S.; Panculescu-Gatej, R. I.; Vázquez, A.; Tenorio, A.; Rebreanu, R.; Niedrig, M.; Nicolescu, G.; Pistol, A. Outbreak of West Nile virus infection in humans, Romania, July to October 2010. Euro Surveill 2011, 16 (2), pii=19762, doi:10.2807/ese.16.02.19762-en.
Smithburn, K. C.; Hughes, T. P.; Burke, A. W.; Paul J. H. A neurotropic virus isolated from the blood of a native of Uganda. Am J Trop Med Hyg 1940, Volume s1-20 (4), 471-492.
Spigland, I.; Jasinska-Klingberg, W.; Hofshi, E.; Goldblum, N. [Clinical and laboratory observations in an outbreak of West Nile fever in Israel in 1957]. Harefuah 1958, 54 (11), 275-280; English & French abstracts 280-2811.
Steele, K. E.; Linn, M. J.; Schoepp, R. J.; Komar, N.; Geisbert, T. W.; Manduca, R. M.; Calle, P. P.; Raphael, B. L.; Clippinger, T. L.; Larsen, T.; Smith, J.; Lanciotti, R. S.; Panella, N. A.; McNamara, T. S. Pathology of fatal West Nile virus infections in native and exotic birds during the 1999 outbreak in New York City, New York. Vet Pathol 2000, 37 (3), 208-224, doi:10.1354/vp.37-3-208.
Styer, L. M.; Bernard, K. A.; Kramer, L. D. Enhanced early West Nile virus infection in young chickens infected by mosquito bite: Effect of viral dose. Am J Trop Med Hyg 2006, 75 (2), 337-345.
Styer, L. M.; Lim, P. Y.; Louie, K. L.; Albright, R. G.; Kramer, L. D.; Bernard, K. A. Mosquito saliva causes enhancement of West Nile virus infection in mice. J Virol 2011, 85 (4), 1517-1527, doi:10.1128/JVI.01112-10.
Suthar, M. S.; Diamond, M. S.; Gale, M., Jr. West Nile virus infection and immunity. Nat Rev Microbiol 2013, 11 (2), 115-128, doi:10.1038/nrmicro2950.
Swayne, D. E.; Beck, J. R.; Zaki, S. Pathogenicity of West Nile virus for turkeys. Avian Dis 2000, 44 (4), 932-937, doi:10.2307/1593067.
Tabachnick, W. J. Climate change and the arboviruses: Lessons from the evolution of the dengue and yellow fever viruses. Annu Rev Virol 2016, 3 (1), 125-145, doi:10.1146/annurev-virology-110615-035630.
Tamba, M.; Bonilauri, P.; Bellini, R.; Calzolari, M.; Albieri, A.; Sambri, V.; Dottori, M.; Angelini, P. Detection of Usutu virus within a West Nile virus surveillance program in Northern Italy. Vector Borne Zoonotic Dis 2011, 11 (5), 551-557, doi:10.1089/vbz.2010.0055.
Tassaneetrithep, B.; Burgess, T. H.; Granelli-Piperno, A.; Trumpfherer, C.; Finke, J.; Sun, W.; Eller, M. A.; Pattanapanyasat, K.; Sarasombath, S.; Birx, D. L.; Steinman, R. M.; Schlesinger, S.; Marovich, M. A. DC-SIGN (CD209) mediates dengue virus infection

Bibliography 105
of human dendritic cells. J Exp Med 2003, 197 (7), 823-829, doi:10.1084/jem.20021840.
Taylor, R. M.; Work, T. H.; Hurlbut, H. S.; Rizk, F. A study of the ecology of West Nile virus in Egypt. Am J Trop Med Hyg 1956, 5 (4), 579-620, doi:10.4269/ajtmh.1956.5.579.
Terradas, G.; McGraw, E. A. Using genetic variation in Aedes aegypti to identify candidate anti-dengue virus genes. BMC Infect Dis 2019, 19 (1), 580, doi:10.1186/s12879-019-4212-z.
Tesla, B.; Demakovsky, L. R.; Packiam, H. S.; Mordecai, E. A.; Rodríguez, A. D.; Bonds, M. H.; Brindley, M. A.; Murdock, C. C. Estimating the effects of variation in viremia on mosquito susceptibility, infectiousness, and R0 of Zika in Aedes aegypti. PLoS Negl Trop Dis 2018, 12 (8), e0006733, doi:10.1371/journal.pntd.0006733.
Titus, R. G.; Bishop, J. V.; Mejia, J. S. The immunomodulatory factors of arthropod saliva and the potential for these factors to serve as vaccine targets to prevent pathogen transmission. Parasite Immunol 2006, 28 (4), 131-141, doi:10.1111/j.1365-3024.2006.00807.x.
Tomasello, D.; Schlagenhauf, P. Chikungunya and dengue autochthonous cases in Europe, 2007-2012. Travel Med Infect Dis 2013, 11 (5), 274-284, doi:10.1016/j.tmaid.2013.07.006.
Trock, S. C.; Meade, B. J.; Glaser, A. L.; Ostlund, E. N.; Lanciotti, R. S.; Cropp, B. C.; Kulasekera, V.; Kramer, L. D.; Komar, N. West Nile virus outbreak among horses in New York State, 1999 and 2000. Emerg Infect Dis 2001, 7 (4), 745-747, doi:10.3201/eid0704.010427.
Tsai, T. F.; Popovici, F.; Cernescu, C.; Campbell, G. L.; Nedelcu, N. I. West Nile encephalitis epidemic in southeastern Romania. Lancet 1998, 352 (9130), 767-771, doi:10.1016/s0140-6736(98)03538-7.
Tukey, W. J. The philosophy of multiple comparisons. Stat Sci 1991, 6 (1), 100-116. Turell, M. J. Members of the Culex pipiens complex as vectors of viruses. J Am Mosq Control
Assoc 2012, 28 (4 Suppl), 123-126, doi:10.2987/8756-971X-28.4.123. Tyler, K. L. West Nile virus infection in the United States. Arch Neurol 2004, 61 (8), 1190-
1195, doi:10.1001/archneur.61.8.1190. Ulbert, S.; Magnusson, S. E. Technologies for the development of West Nile virus vaccines.
Future Microbiol 2014, 9 (10), 1221-1232, doi:10.2217/fmb.14.67. United Nations Development Programme (UNDP), in partnership with the International
Federation of Red Cross and Red Crescent Societies (IFRC). A socio-economic impact assessment of the Zika virus in Latin America and the Caribbean: With a focus on Brazil, Colombia and Suriname. 2017. Availabe online: http://www.undp.org/content/dam/undp/library/HIV-AIDS/UNDP-Zika-04-03-2017-English-WEB.pdf (accessed on 08 April 2020).
Uraki, R.; Hastings, A. K.; Gloria-Soria, A.; Powell, J. R.; Fikrig, E. Altered vector competence in an experimental mosquito-mouse transmission model of Zika infection. PLoS Negl Trop Dis 2018, 12 (3), e0006350, doi:10.1371/journal.pntd.0006350.
Vaidyanathan, R.; Fleisher, A. E.; Minnick, S. L.; Simmons, K. A.; Scott, T. W. Nutritional stress affects mosquito survival and vector competence for West Nile virus. Vector Borne Zoonotic Dis 2008, 8 (6), 727-732, doi:10.1089/vbz.2007.0189.
Vaidyanathan, R.; Scott, T. W. Apoptosis in mosquito midgut epithelia associated with West Nile virus infection. Apoptosis 2006, 11 (9), 1643-1651, doi:10.1007/s10495-006-8783-y.

106 Bibliography
van der Meulen, K. M.; Pensaert, M. B.; Nauwynck, H. J. West Nile virus in the vertebrate world. Arch Virol 2005, 150 (4), 637-657, doi:10.1007/s00705-004-0463-z.
Vanhomwegen, J.; Beck, C.; Desprès, P.; Figuerola, A.; García, R.; Lecollinet, S.; López-Roig, M.; Manuguerra, J. C.; Serra-Cobo, J. Circulation of zoonotic arboviruses in equine populations of Mallorca Island (Spain). Vector Borne Zoonotic Dis 2017, 17 (5), 340-346, doi:10.1089/vbz.2016.2042.
Venter, M. Assessing the zoonotic potential of arboviruses of African origin. Curr Opin Virol 2018, 28, 74-84, doi:10.1016/j.coviro.2017.11.004.
Venter, M.; Swanepoel, R. West Nile virus lineage 2 as a cause of zoonotic neurological disease in humans and horses in southern Africa. Vector Borne Zoonotic Dis 2010, 10 (7), 659-664, doi:10.1089/vbz.2009.0230.
Venturi, G.; Di Luca, M.; Fortuna, C.; Remoli, M. E.; Riccardo, F.; Severini, F.; Toma, L.; Del Manso, M.; Benedetti, E.; Caporali, M. G.; Amendola, A.; Fiorentini, C.; De Liberato, C.; Giammattei, R.; Romi, R.; Pezzotti, P.; Rezza, G.; Rizzo, C. Detection of a chikungunya outbreak in Central Italy, August to September 2017. Euro Surveill 2017, 22 (39), pii=17-00646, doi:10.2807/1560-7917.ES.2017.22.39.17-00646.
Vilibic-Cavlek, T.; Petrovic, T.; Savic, V.; Barbic, L.; Tabain, I.; Stevanovic, V.; Klobucar, A.; Mrzljak, A.; Ilic, M.; Bogdanic, M.; Benvin, I.; Santini, M.; Capak, K.; Monaco, F.; Listes, E.; Savini, G. Epidemiology of Usutu virus: The European scenario. Pathogens 2020, 9 (9), 699, doi:10.3390/pathogens9090699.
Vilibic-Cavlek, T.; Savic, V.; Sabadi, D.; Peric, L.; Barbic, L.; Klobucar, A.; Miklausic, B.; Tabain, I.; Santini, M.; Vucelja, M.; Dvorski, E.; Butigan, T.; Kolaric-Sviben, G.; Potocnik-Hunjadi, T.; Balenovic, M.; Bogdanic, M.; Andric, Z.; Stevanovic, V.; Capak, K.; Balicevic, M.; Listes, E.; Savini, G. Prevalence and molecular epidemiology of West Nile and Usutu virus infections in Croatia in the 'One health' context, 2018. Transbound Emerg Dis 2019, 66 (5), 1946-1957, doi:10.1111/tbed.13225.
Viña-Rodríguez, A.; Sachse, K.; Ziegler, U.; Chaintoutis, S.C.; Keller, M.; Groschup, M. H.; Eiden, M. A novel pan-flavivirus detection and identification assay based on RT-qPCR and microarray. Biomed Res Int 2017, 2017, e4248756, doi:10.1155/2017/4248756.
Vogels, C. B. F.; Göertz, G. P.; Pijlman, G. P.; Koenraadt, C. J. M. Vector competence of northern and southern European Culex pipiens pipiens mosquitoes for West Nile virus across a gradient of temperatures. Med Vet Entomol 2017a, 31 (4), 358-364, doi:10.1111/mve.12251.
Vogels, C. B. F.; Hartemink, N.; Koenraadt, C. J. M. Modelling West Nile virus transmission risk in Europe: effect of temperature and mosquito biotypes on the basic reproduction number. Sci Rep 2017b, 7 (1), 5022, doi:10.1038/s41598-017-05185-4.
Vogels, C. B.; Fros, J. J.; Göertz, G. P.; Pijlman, G. P.; Koenraadt, C. J. Vector competence of northern European Culex pipiens biotypes and hybrids for West Nile virus is differentially affected by temperature. Parasit Vectors 2016, 9 (1), 393, doi:10.1186/s13071-016-1677-0.
Vogels, C. B.; Göertz, G. P.; Pijlman, G. P.; Koenraadt, C. J. Vector competence of European mosquitoes for West Nile virus. Emerg Microbes Infect 2017c, 6 (11), e96, doi:10.1038/emi.2017.82.
Wagner, S.; Mathis, A.; Schönenberger, A. C.; Becker, S.; Schmidt-Chanasit, J.; Silaghi, C.; Veronesi, E. Vector competence of field populations of the mosquito species Aedes

Bibliography 107
japonicus japonicus and Culex pipiens from Switzerland for two West Nile virus strains. Med Vet Entomol 2018, 32 (1), 121-124, doi:10.1111/mve.12273.
Walther, D.; Scheuch, D. E.; Kampen, H. The invasive Asian tiger mosquito Aedes albopictus (Diptera: Culicidae) in Germany: Local reproduction and overwintering. Acta Trop 2017, 166, 186-192, doi:10.1016/j.actatropica.2016.11.024.
Wang, T.; Town, T.; Alexopoulou, L.; Anderson, J. F.; Fikrig, E.; Flavell, R. A. Toll-like receptor 3 mediates West Nile virus entry into the brain causing lethal encephalitis. Nat Med 2004, 10 (12), 1366-1373, doi:10.1038/nm1140.
Weingartl, H. M.; Neufeld, J. L.; Copps, J.; Marszal, P. Experimental West Nile virus infection in blue jays (Cyanocitta cristata) and crows (Corvus brachyrhynchos). Vet Pathol 2004, 41 (4), 362-370, doi:10.1354/vp.41-4-362.
Weissenböck, H.; Bakonyi, T.; Chvala, S.; Nowotny, N. Experimental Usutu virus infection of suckling mice causes neuronal and glial cell apoptosis and demyelination. Acta Neuropathol 2004, 108 (5), 453-460, doi:10.1007/s00401-004-0916-1.
Weissenböck, H.; Bakonyi, T.; Rossi, G.; Mani, P.; Nowotny, N. Usutu virus, Italy, 1996. Emerg Infect Dis 2013, 19 (2), 274-277, doi:10.3201/eid1902.121191.
Weissenböck, H.; Kolodziejek, J.; Url, A.; Lussy, H.; Rebel-Bauder, B.; Nowotny, N. Emergence of Usutu virus, an African mosquito-borne flavivirus of the Japanese encephalitis virus group, central Europe. Emerg Infect Dis 2002, 8 (7), 652-656, doi:10.3201/eid0807.020094.
Wheeler, S. S.; Langevin, S. A.; Brault, A. C.; Woods, L.; Carroll, B. D.; Reisen, W. K. Detection of persistent West Nile virus RNA in experimentally and naturally infected avian hosts. Am J Trop Med Hyg 2012, 87 (3), 559-564, doi:10.4269/ajtmh.2012.11-0654.
Whitfield, S. G.; Murphy, F. A.; Sudia, W. D. St. Louis encephalitis virus: An ultrastructural study of infection in a mosquito vector. Virology 1973, 56 (1), 70-87, doi:10.1016/0042-6822(73)90288-2.
Wilcox, B. R.; Yabsley, M. J.; Ellis, A. E.; Stallknecht, D. E.; Gibbs, S. E. J. West Nile virus antibody prevalence in American crows (Corvus brachyrhynchos) and fish crows (Corvus ossifragus) in Georgia, USA. Avian Diseases 2007, 51 (1), 125-128, doi:10.1637/0005-2086(2007)051[0125:Wnvapi]2.0.Co;2.
Wilder-Smith, A.; Ooi, E. E.; Vasudevan, S. G.; Gubler, D. J. Update on dengue: Epidemiology, virus evolution, antiviral drugs, and vaccine development. Curr Infect Dis Rep 2010, 12 (3), 157-164, doi:10.1007/s11908-010-0102-7.
Williams, M. C.; Simpson, D. I.; Haddow, A. J.; Knight, E. M. The isolation of West Nile virus from man and of Usutu virus from the bird-biting mosquito Mansonia aurites (Theobald) in the Entebbe area of Uganda. Ann Trop Med Parasitol 1964, 58, 367-374, doi:10.1080/00034983.1964.11686258.
Wöhnke, E.; Vasić, A.; Răileanu, C.; Holicki, C. M.; Tews, B. A.; Silaghi, C. Comparison of vector competence of Aedes vexans Green River and Culex pipiens biotype pipiens for West Nile virus lineages 1 and 2. Zoonoses Public Health 2020, 67, 416-424, doi:10.1111/zph.12700.
Woodring, J. L.; Higgs, S.; Beaty, B. J. Natural cycles of vector-borne pathogens. In The biology of disease vectors, University Press of Colorado: Niwot, CO, USA, 1996; pp. 51–72.
Wünschmann, A.; Shivers, J.; Bender, J.; Carroll, L.; Fuller, S.; Saggese, M.; van Wettere, A.; Redig, P. Pathologic and immunohistochemical findings in goshawks (Accipiter

108 Bibliography
gentilis) and great horned owls (Bubo virginianus) naturally infected with West Nile virus. Avian Dis 2005, 49 (2), 252-259, doi:10.1637/7297-103104R.
Young, P. R. Arboviruses: A family on the move. Adv Exp Med Biol 2018, 1062, 1-10, doi:10.1007/978-981-10-8727-1_1.
Zannoli, S.; Sambri, V. West Nile virus and Usutu virus co-circulation in Europe: Epidemiology and implications. Microorganisms 2019, 7 (7), 184, doi:10.3390/microorganisms7070184.
Ziegler, U.; Angenvoort, J.; Fischer, D.; Fast, C.; Eiden, M.; Viña-Rodriguéz, A.; Revilla-Fernández, S.; Nowotny, N.; de la Fuente, J. G.; Lierz, M.; Groschup, M. H. Pathogenesis of West Nile virus lineage 1 and 2 in experimentally infected large falcons. Vet Microbiol 2013, 161 (3-4), 263-273, doi:10.1016/j.vetmic.2012.07.041.
Ziegler, U.; Fast, C.; Eiden, M.; Bock, S.; Schulze, C.; Höper, D.; Ochs, A.; Schlieben, P.; Keller, M.; Zielke, D. E.; Lühken, R.; Cadar, D.; Walther, D.; Schmidt-Chanasit, J.; Groschup, M. H. Evidence for an independent third Usutu virus introduction into Germany. Vet Microbiol 2016, 192, 60-66, doi:10.1016/j.vetmic.2016.06.007.
Ziegler, U.; Jöst, H.; Müller, K.; Fischer, D.; Rinder, M.; Tietze, D. T.; Danner, K. J.; Becker, N.; Skuballa, J.; Hamann, H. P.; Bosch, S.; Fast, C.; Eiden, M.; Schmidt-Chanasit, J.; Groschup, M. H. Epidemic spread of Usutu virus in Southwest Germany in 2011 to 2013 and monitoring of wild birds for Usutu and West Nile viruses. Vector Borne Zoonotic Dis 2015, 15 (8), 481-488, doi:10.1089/vbz.2014.1746.
Ziegler, U.; Lühken, R.; Keller, M.; Cadar, D.; van der Grinten, E.; Michel, F.; Albrecht, K.; Eiden, M.; Rinder, M.; Lachmann, L.; Höper, D.; Viña-Rodriguéz, A.; Gaede, W.; Pohl, A.; Schmidt-Chanasit, J.; Groschup, M. H. West Nile virus epizootic in Germany, 2018. Antiviral Res 2019, 162, 39-43, doi:10.1016/j.antiviral.2018.12.005.
Ziegler, U.; Santos, P. D.; Groschup, M. H.; Hattendorf, C.; Eiden, M.; Höper, D.; Eisermann, P.; Keller, M.; Michel, F.; Klopfleisch, R.; Müller, K.; Werner, D.; Kampen, H.; Beer, M.; Frank, C.; Tews, B. A.; Wylezich, C.; Rinder, M.; Lachmann, L.; Grünewald, T.; Szentiks, C.; Sieg, M.; Schmidt-Chanasit, J.; Cadar, D.; Lühken, R. West Nile virus epidemic in Germany triggered by epizootic emergence, 2019. Viruses 2020, 12 (4), 448, doi:10.3390/v12040448.
Ziegler, U.; Seidowski, D.; Globig, A.; Fereidouni, S. R.; Ulrich, R. G.; Groschup, M. H. Sentinel birds in wild-bird resting sites as potential indicators for West Nile virus infections in Germany. Arch Virol 2010, 155 (6), 965-969, doi:10.1007/s00705-010-0618-z.
Zielke, D. E.; Walther, D.; Kampen, H. Newly discovered population of Aedes japonicus japonicus (Diptera: Culicidae) in Upper Bavaria, Germany, and Salzburg, Austria, is closely related to the Austrian/Slovenian bush mosquito population. Parasit Vectors 2016, 9, 163, doi:10.1186/s13071-016-1447-z.

SUPPLEMENTS

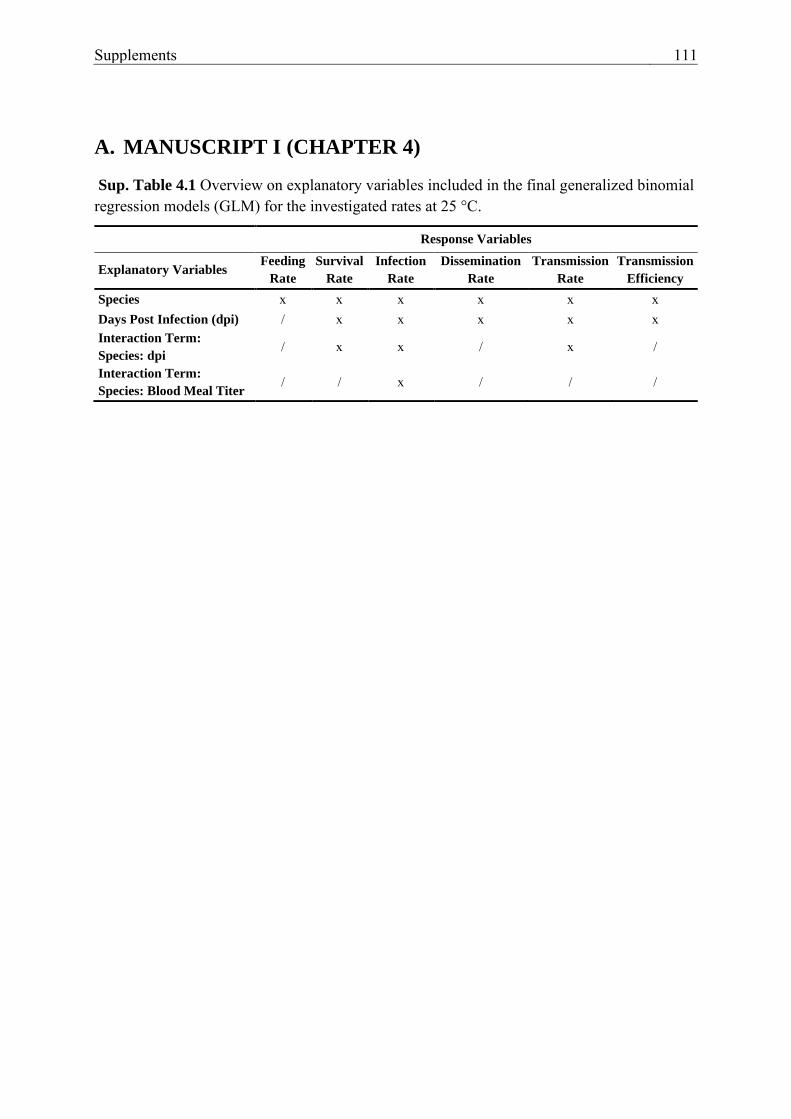
Supplements 111
A. MANUSCRIPT I (CHAPTER 4)
Sup. Table 4.1 Overview on explanatory variables included in the final generalized binomial regression models (GLM) for the investigated rates at 25 °C.
Response Variables
Explanatory Variables Feeding
Rate
Survival
Rate
Infection
Rate
Dissemination
Rate
Transmission
Rate
Transmission
Efficiency
Species x x x x x x Days Post Infection (dpi) / x x x x x Interaction Term:
Species: dpi / x x / x /
Interaction Term:
Species: Blood Meal Titer / / x / / /

112 A. Manuscript I (Chapter 4)
Su
p. T
ab
le 4
.2 p-values of the fixed effects in the least-square means analysis w
hen comparing the rates betw
een all species at 25 °C
in the final generalized binomial regression m
odels (GLM
) specified in Sup. Table 4.1. p-value adjustment w
as performed using the
Tukey method.
Fix
ed effects:
Feed
ing
Ra
te
Su
rviv
al
Ra
te +
Infectio
n
Ra
te +
Dissem
ina
tion
Ra
te
Tra
nsm
ission
Ra
te +
Tra
nsm
ission
Efficien
cy
C
on
trast
p-v
alu
e
(GL
M)
p-a
dju
sted
(Fish
er) ‡ p
-va
lue
p-v
alu
e p
-va
lue
p-v
alu
e p
-va
lue
Sp
ecies A
A_M
- CPM
_G
1.99 x 10-1
1.00 7.62 x 10
-1 1.00
3.26 x 10-1
NA
1.00
A
A_M
- CPM
_S 1.00
1.00 8.45 x 10
-1 1.00
1.00 N
A
1.00
A
A_M
- CT_G
4.29 x 10
-14 * 1.30 x 10
-5 * 1.00
1.00 6.86 x 10
-1 N
A
1.00
C
PM_G
- CPM
_S 1.00
3.86 x 10-1
6.12 x 10-1
9.64 x 10-1
1.00 1.00
7.21 x 10-1
C
PM_G
- CT_G
3.57 x 10
-13 * 4.58 x 10 -5 *
1.00 7.48 x 10
-1 9.97 x 10
-1 N
A
3.18 x 10-1
C
PM_S - C
T_G
1.00 8.65 x 10
-7 * 1.00
9.60 x 10-1
1.00 N
A
6.58 x 10-1
Da
ys P
ost
Infectio
n
14/16 - 21 N
A
NA
9.76 x 10
-1 9.91 x 10
-1 4.50 x 10
-2 * N
A
8.95 x 10-2
Blo
od
Mea
l
Viru
s Titer
(TC
ID50 /m
L)
10 5.1 - 7.4 N
A
NA
N
A
2.11 x 10-6 *
NA
N
A
NA
*, p < 0.05; +, Results for fixed effects m
ay be misleading due to involvem
ent in interactions (Sup. Table 4.3); ‡, Additional statistical
evaluation with Fisher’s exact test w
ith Bonferroni correction; N
A, not applicable; A
A_M
, Aedes aegypti from M
alaysia; CPM
_G, C
ulex pipiens biotype m
olestus from G
ermany; C
PM_S, C
x. pipiens biotype molestus from
the Republic of Serbia; C
T_G, C
x. torrentium from
G
ermany.
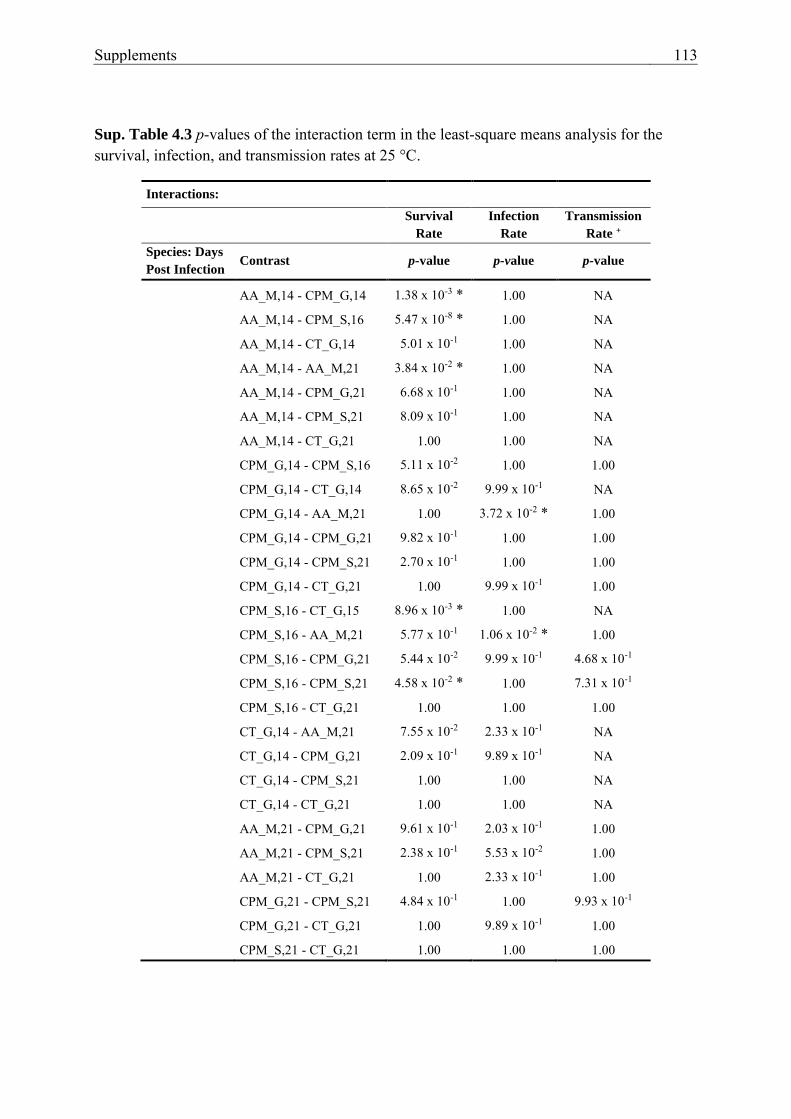
Supplements 113
Sup. Table 4.3 p-values of the interaction term in the least-square means analysis for the survival, infection, and transmission rates at 25 °C.
Interactions:
Survival
Rate
Infection
Rate
Transmission
Rate +
Species: Days
Post Infection Contrast p-value p-value p-value
AA_M,14 - CPM_G,14 1.38 x 10-3 * 1.00 NA AA_M,14 - CPM_S,16 5.47 x 10-8 * 1.00 NA AA_M,14 - CT_G,14 5.01 x 10-1 1.00 NA AA_M,14 - AA_M,21 3.84 x 10-2 * 1.00 NA AA_M,14 - CPM_G,21 6.68 x 10-1 1.00 NA AA_M,14 - CPM_S,21 8.09 x 10-1 1.00 NA AA_M,14 - CT_G,21 1.00 1.00 NA CPM_G,14 - CPM_S,16 5.11 x 10-2 1.00 1.00 CPM_G,14 - CT_G,14 8.65 x 10-2 9.99 x 10-1 NA CPM_G,14 - AA_M,21 1.00 3.72 x 10-2 * 1.00 CPM_G,14 - CPM_G,21 9.82 x 10-1 1.00 1.00 CPM_G,14 - CPM_S,21 2.70 x 10-1 1.00 1.00 CPM_G,14 - CT_G,21 1.00 9.99 x 10-1 1.00 CPM_S,16 - CT_G,15 8.96 x 10-3 * 1.00 NA CPM_S,16 - AA_M,21 5.77 x 10-1 1.06 x 10-2 * 1.00 CPM_S,16 - CPM_G,21 5.44 x 10-2 9.99 x 10-1 4.68 x 10-1 CPM_S,16 - CPM_S,21 4.58 x 10-2 * 1.00 7.31 x 10-1 CPM_S,16 - CT_G,21 1.00 1.00 1.00 CT_G,14 - AA_M,21 7.55 x 10-2 2.33 x 10-1 NA CT_G,14 - CPM_G,21 2.09 x 10-1 9.89 x 10-1 NA CT_G,14 - CPM_S,21 1.00 1.00 NA CT_G,14 - CT_G,21 1.00 1.00 NA AA_M,21 - CPM_G,21 9.61 x 10-1 2.03 x 10-1 1.00 AA_M,21 - CPM_S,21 2.38 x 10-1 5.53 x 10-2 1.00 AA_M,21 - CT_G,21 1.00 2.33 x 10-1 1.00 CPM_G,21 - CPM_S,21 4.84 x 10-1 1.00 9.93 x 10-1 CPM_G,21 - CT_G,21 1.00 9.89 x 10-1 1.00 CPM_S,21 - CT_G,21 1.00 1.00 1.00

114 A. Manuscript I (Chapter 4)
*, p < 0.05; NA, not applicable; AA_M, Aedes aegypti from Malaysia; CPM_G, Culex pipiens biotype molestus from Germany; CPM_S, Cx. pipiens biotype molestus from the Republic of Serbia; CT_G, Cx. torrentium from Germany.

Supplements 115
B. MANUSCRIPT III (CHAPTER 6)
Sup. Table 6.1 p-values of the fixed effects in the least-square means analysis when comparing the rates between all species at 25 °C in the final generalized binomial regression models (GLM). p-value adjustment was performed using the Tukey method.
Fixed Effects:
Infection
Rate
Dissemination
Rate
Transmission
Rate
Contrast p-value p-value p-value
Species CPP_B_10/11 - CPP_M_10/11 1.00 1.00 1.00 CPP_B_10/11 - CT_10/11 1.00 1.00 1.00 CPP_B_10/11 - CT_22 1.00 1.00 1.00 CPP_M_10/11 - CT_10/11 1.00 1.00 1.00 CPP_M_10/11 - CT_22 1.00 1.00 1.00 CT_10/11 - CT_22 1.00 1.00 1.00
CPP_B_10/11, Culex pipiens biotype pipiens from “Schöneiche/Rehfelde” and “Groß Kreutz”, Brandenburg, Germany 10/11 dpi; CPP_M_10/11, Cx. pipiens biotype pipiens from Niederhof, Mecklenburg Western-Pommerania, Germany 10/11 dpi; CT_10/11, Cx. torrentium from “Schöneiche/Rehfelde”, Brandenburg, Germany 10/11 dpi; CT_22, Cx. torrentium from “Schöneiche/Rehfelde”, Brandenburg, Germany 22 dpi.
