university of groningen rheologic changes of hypothermic
TRANSCRIPT

University of Groningen
Rheologic changes of hypothermic preserved red blood cellsHenkelman, Sandra
IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite fromit. Please check the document version below.
Document VersionPublisher's PDF, also known as Version of record
Publication date:2012
Link to publication in University of Groningen/UMCG research database
Citation for published version (APA):Henkelman, S. (2012). Rheologic changes of hypothermic preserved red blood cells. s.n.
CopyrightOther than for strictly personal use, it is not permitted to download or to forward/distribute the text or part of it without the consent of theauthor(s) and/or copyright holder(s), unless the work is under an open content license (like Creative Commons).
The publication may also be distributed here under the terms of Article 25fa of the Dutch Copyright Act, indicated by the “Taverne” license.More information can be found on the University of Groningen website: https://www.rug.nl/library/open-access/self-archiving-pure/taverne-amendment.
Take-down policyIf you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediatelyand investigate your claim.
Downloaded from the University of Groningen/UMCG research database (Pure): http://www.rug.nl/research/portal. For technical reasons thenumber of authors shown on this cover page is limited to 10 maximum.
Download date: 03-12-2021

Rheologic changes of hypothermic preserved red blood cells
Sandra Henkelman

Rheologic changes of hypothermic preserved red blood cells. By Sandra Henkelman
Department of BioMedical Engineering
University Medical Center Groningen, University of Groningen
Groningen, the Netherlands
ISBN (book) : 978-90-367-5399-9
ISBN (digital document) : 978-90-367-5400-2
Copyright © 2012 by S. Henkelman
All rights are reserved. No part of this publication may be reproduced or transmitted in any form or by any means, without permission of the author.
Cover design: In Zicht grafisch ontwerp
Lay-out: Ingrid Engelsman
Printed by: GVO drukkers & vormgevers B.V.

Rheologic changes of hypothermic preserved red blood cells
Proefschrift
ter verkrijging van het doctoraat in de Medische Wetenschappen
aan de Rijksuniversiteit Groningen op gezag van de
Rector Magnificus, dr. E. Sterken, in het openbaar te verdedigen op
woensdag 14 maart 2012 om 14.30 uur
door
Sandra Henkelman
geboren op 26 mei 1976
te Groningen

Promotores: Prof. dr. G. Rakhorst Prof. dr. ir. H.J. Busscher Copromotor: Dr. W. van Oeveren Beoordelingscommissie: Prof.dr. A.R. Absalom
Prof.dr. R.A.W. van Lier Prof.dr. G.J. Verkerke

Paranimfen: A.J. Munnike MD. O.C. Varnav Ir. G.E. Engels Ir. W. van der Houwen

The world is your Oyster
Voor Opa en Oma…

Contents Chapter 1. Introduction 1
Chapter 2. Use of hydroxyethyl starch for inducing red blood cell aggregation (Clin.Hemorheol.Micro. 2012)
19
Chapter 3. Red blood cell aggregation; an important phenomena in damage control resuscitation?
33
Chapter 4. Is red blood cell rheology preserved during routinely blood bank storage? (Transfusion 2010, 50: 941-948)
39
Chapter 5. Utilization of cryopreserved red blood cells in transfusion medicine.
57
Chapter 6. The effects of cryopreservation on red blood cell rheologic properties (Transfusion 2010, 50: 2393-2401)
77
Chapter 7. A new freezing technique allows storage of RBCs with 20% glycerol at - 80°C.
97
Chapter 8. General discussion 115
Summary 127 Samenvatting 133 Acknowledgements 139


List of Abbreviations
AABB American association of blood banks AI aggregation index ATP adenine triphosphate AS-3 additive solution 3 CPD citrate-phosphate-dextrose 2,3-DPG 2,3-diphosphoglycerate DEHP di(2-ethylhexyl)phthalate ECs endothelial cells EI elongation index EPM electrophoretic mobility FFP fresh frozen plasma FWB fresh whole blood Hb hemoglobin Hct hematocrit HES hydroxyethyl starch HGM high-glycerol method LGM low-glycerol method LORCA laser-assisted optical rotational cell analyzer MCV mean cell volume MCHC mean cell hemoglobin concentration MODS multiple organ dysfunction syndrome OD optical density PS phosphatidyl serine PVC polyvinylchloride PVP polyvinylpyrrolidone RBCs red blood cells SAGM saline-adenine-glucose-mannitol additive solution SIRS systemic inflammatory response syndrome SD standard deviation T ½ aggregation half-time T1DM type-1 diabetes mellitus TRALI transfusion related acute lung injury


Chapter 1
Introduction and aim of the thesis

Chapter 1
2
1.1 Introduction Blood transfusion experiments were practiced as early in the 17th century. However, it was the discovery of the ABO blood groups in the early 1900 that boosted transfusion medicine.1 Blood transfusion practice has considerably improved since then and nowadays refrigerated stored red blood cells (RBCs) are routinely infused to compensate for excessive blood loss or to correct for abnormal hemoglobin content. Yet, during refrigerated storage the RBCs undergo structural and biochemical alterations, collectively referred to as the storage lesion, which could compromise their function after infusion. Particularly, alterations in the RBC flow properties (i.e. rheologic properties) may impede the RBCs to properly function in the microcirculation. This chapter describes, from a rheologic perspective, the RBC characteristics and the cellular changes that are observed during refrigerated blood bank storage. In addition, cryopreservation of RBCs will be discussed as an alternative approach to counteract storage induced lesions and thus to extend the shelf life of RBCs.
Characteristics of RBCs RBCs are the most abundant cells in the blood. Adult humans have approximately 20-30 trillion circulating RBCs and on an average 2.6 million human RBCs are produced every second by the red bone marrow and released into the blood via marrow sinusoids.2 RBCs have unique structures and properties which enable them to distribute oxygen and collect carbon dioxide in the body. Mature RBCs are shaped as biconcave discs (Figure 1.1) that lack nuclei, mitochondria and other cell organelles. Yet, the RBCs are rich in hemoglobin (Hb), an iron-containing protein that is able to reversible bind oxygen and carbon dioxide.
Figure 1.1. Human red blood cells under physiological conditions.

Introduction
3
In the RBC, energy is provided via the anaerobic glycolysis pathway. In this pathway glucose is broken down in pyruvate with among others adenine-triphosphate (ATP) and 2,3-diphosphoglycerate (2,3-DPG) as intermediate metabolites. Subsequently, pyruvate is converted to lactate and transported out of the cell. ATP as an energy source is essential for the overall functioning of the RBC. Loss of ATP has been associated with shape changes, enhanced cation permeability, rigid cell membranes, altered surface expression of phospholipids, microvesiculation and decreased RBC viability.3-10 Furthermore, ATP released by the RBC is a potent vasodilatation signaling molecule that adapts the blood flow according to local needs.11,12 Recently, the ATP content was also correlated with the oxygenation capacity of the RBC.13,14 In the RBC binding of 2,3-DPG to the Hb induces a conformation state which will release oxygen from the Hb. Due to the high binding affinity of 2,3-DPG for deoxygenated Hb, oxygen is released in regions with low oxygen tension. Loss of 2,3-DPG will increase the oxygen affinity to the Hb which may hamper the oxygen delivery to the tissues.2 The RBC membrane is composed of a lipid bi-layer and an underlying viscoelastic cytoskeleton. This bi-layer consists of a layer of proteins which, due to the sialic acid groups, bears a net negative charge. The viscoelastic cytoskeleton enables the RBC to adapt their shape to maintain the flow behavior in the microvessels.
RBCs in the microcirculation The microcirculation consists of a network of arterioles, capillaries and venules with a diameter of less than 100 µm.15 In the capillaries exchange of oxygen and carbon dioxide between circulating RBCs and the surrounding tissues takes place. Most microcirculatory vessels are embedded within an organ. Adequate flow in the microcirculation is thus a requisite for normal organ perfusion and function. The RBC rheologic properties (i.e. the ability of RBC to aggregate, deform and adhere to endothelial cells) are important determinants of the flow behavior of blood and subsequently the oxygenation of the micro-vascular environment.16,17 The ability of RBCs to form linear aggregates (Rouleaux: Figure 1.2) or more complex three dimensional aggregates in regions with low shear rate is a phenomenon that has been studied for decades. RBC aggregation is depending on opposing shear forces. On the one hand RBC aggregation is counteracted by the repulsive force between negatively charged

Chapter 1
4
cells as well as the force generated by the blood flow. On the other hand, RBC aggregation is promoted by the presence of plasma proteins, most importantly fibrinogen, or by high molecular weight polymers.18,19 At present there are two theories proposed to explain the RBC aggregation mechanism: a bridging and a depletion theory.20 In the bridging theory, it is hypothesized that macromolecules adsorbed onto the RBC surface form bridges between adjacent RBCs, whereas in the depletion theory RBC aggregation is induced due to an osmotic gradient difference between the polymer concentration near the RBC surface versus the suspending medium. Although RBC aggregation markedly enhances the blood viscosity at low shear rate,21 the physiological role of this process is still elusive. Under normal physiological conditions, RBC aggregates are easily dispersed by the rise in blood flow rate. However, in certain pathologic states such as cardiovascular disorders, sepsis, pre-eclampsia, diabetes mellitus, chronic renal failure, rheumatoid arthritis, inflammation, hypertension, thalassemia and sickle cell disease,18,22-26 stronger and or larger aggregates are formed which are more resistant to dispersion by the blood flow. Essentially, enhanced RBC aggregation has the potential to impair the blood flow in the microcirculation and contribute to the occlusion of micro-vessels.26,27
Figure 1.2. RBC Rouleaux formation in the microcirculation.
ArteryVein Capillaries
Flow O2
CO2
O2

Introduction
5
The ability of RBCs to deform due to applied shear stress enables these cells to adapt their size to squeeze through narrow capillaries. It also allows RBCs to reduce the blood viscosity at high shear rate, so that blood remains fluid even at a high hematocrit. The RBC deformability is depending on the geometry of the cell, the viscosity of the cytoplasm as well as on the viscoelastic properties of the cytoskeleton.28 Alterations in these properties can make the RBC more rigid and impair or block the blood flow in the microcirculation. Ultimately, rigid RBCs will be sequestered by the spleen and destroyed. The RBC deformability is thus essential for adequate tissue perfusion and cell survival.28,29 Reduced RBC deformability has been observed in sepsis, diabetes mellitus, malaria, hypertension, thalassemia and sickle cell disease.23-25 The adherence of RBCs to the vascular endothelium is, under physiological conditions, negligible. However, structural changes in the RBC membrane may promote adherence to endothelial cells (ECs) and impair the microcirculatory blood flow.26,30Adherence of RBCs to ECs is mediated by expression of phosphatidylserine (PS) on the RBC surface.31,32 PS expression also triggers recognition by macrophages which clears the RBCs from the circulation.33-35 Enhanced RBC-EC interaction has been observed in malaria, diabetes mellitus, thalassemia, sickle cell disease.26 In summary, changes in the rheologic properties of RBCs may form a circulatory risk by hindering adequate tissue perfusion and contributing to ischemia or even infarction.26,36,37 It is therefore not surprisingly that flow disturbances in the microcirculation are closely associated with morbidity and mortality.38,39 These findings highlight the importance of RBC rheologic properties as functionality parameters in transfusion medicine.
Usage of the Laser-assisted Optical Rotational Cell Analyzer The laser-assisted optical rotational red cell analyzer (LORCA; R&R Mechatronics, Zwaag The Netherlands)40,41 is a useful device to determine the aggregation and deformation behavior of RBCs ex vivo.42,43 In general, a laser beam is directed through the RBC solution and RBC aggregation is determined by means of backscattered light. Initially, the RBC suspension is sheared to disperse pre-existing RBC aggregates. After several seconds the shear is halted and the light reflection from the suspension is recorded over a certain timeframe and subsequently analyzed by a computer program. As RBCs start to form side to side aggregates (Rouleaux) the intensity of the backscattered light decreases

Chapter 1
6
exponentially. The aggregation behavior of RBCs is described by the aggregation index (AI), which depends both on the kinetics and extent of aggregation. The RBC deformability can be determined at various fluid shear stresses by means of laser diffraction analysis. In general, a laser beam is directed through a sheared diluted RBC solution and the diffraction pattern produced by the deformed cells is analyzed by a computer program. Under the influence of shear stress, RBCs will gradual deform from a biconcave to ellipsoid shape. Based upon the geometry of the diffraction pattern the elongation index (EI) is calculated, in which an increased EI at any given shear stress indicates greater cell deformation.
RBC Senescence Human RBCs have an in vivo life span of approximately 120 days after which they are selectively removed from the circulation by macrophages.35,44 This mechanism is essential since it prevents Hb release from aged RBCs directly in the circulation. Aged RBCs are characterized by cell shrinkage, microvesiculation and PS exposure at the cell surface. Aged RBCs also have a decreased ability to deform and an enhanced tendency to aggregate.19,45-47 RBC injury due to energy depletion, osmotic shock or oxidative stress reduces the life span of the RBC.45 Refrigerated storage also induces cellular changes in the RBC that diminish the life span and possibly the functionality of RBCs.
1.2. Refrigerated storage of RBCs Worldwide RBCs are routinely infused in order to compensate for excessive blood loss as well as to improve the oxygen carrying capacity. Annually, developed countries transfuse approximately one RBC storage unit for every 20 inhabitants.48 Yet, recently RBC transfusions has been under intensive evaluation. This is mainly because infusion of prolonged refrigerated stored RBCs have been linked to adverse outcome in terms of postoperative infections, length of hospital stay, multiple organ dysfunction syndrome (MODS), transfusion related acute lung injury (TRALI), cancer progression and even mortality.49-56

Introduction
7
Requirements of refrigerated stored RBCs To date RBCs are routinely stored at 2-6°C for a maximum of 5 to 6 weeks, depending on the preservation solution used. Notably, in the Netherlands RBCs are refrigerated and stored in saline-adenine-glucose-mannitol (SAGM) solution for a maximum of 5 weeks. International guidelines require that the hemolysis in a refrigerated RBC storage unit must remain below allowable levels (i.e., 0.8% in Europe and 1% in The United States) and that at least 75% of the infused RBCs must still circulate 24 hours after infusion.57,58 Interestingly, these guidelines do not specifically reflect the RBC ability to function after infusion.
Quality of refrigerated stored RBCs Although storage at 4°C slows down the biochemical processes in the RBC, cellular metabolism is not completely suppressed at these temperatures. During refrigerated storage a variety of changes have been observed that could compromise the RBCs to function after infusion. These changes include decreased concentrations of 2,3-DPG, ATP and membrane sialic acid content, translocation of PS to the cell surface, oxidative injury to membrane lipids and proteins, shape change to spheroechinocytes, membrane blebbing and accumulation of potassium, free HB, cytokines, bioactive lipids and (pro-coagulant) microvesicles in the RBC storage unit.59-63 The RBCs rheologic properties also become impaired during refrigerated storage. RBCs demonstrate an increased tendency to aggregate, diminished deformability and augmented adhesiveness to ECs starting already from the second week of storage.64-68 These alterations could hamper the RBCs to properly function in the microcirculation. Especially, since the majority of the RBC units that are infused have a lifespan that exceeds 2 weeks of storage.69-71 Yet, studies which propose a detrimental association with the infusion of long-term refrigerated stored RBCs are often biased and not adequately corrected for known confounding factors.72-76 These factors include among others the number of RBC units transfused, mixture of RBCs of different durations, severity of illness and the utilization of leukoreduction.

Chapter 1
8
Leukofiltration of RBCs In most developed countries, leukoreduction of RBC units prior to storage is a mandatory practice. Initially leukoreduction was implemented to prevent HLA alloimmunization, however by reducing the leukocyte count from a mean of 109 to 106 cells per RBC unit, less detrimental substances such as cytokines, proteolytic enzymes and oxygen free radicals accumulate in the RBC storage unit.77-80 Notably, leukoreduction reduces the risk of HLA alloimmunization, febrile non-hemolytic transfusion reactions, immunomodulation and transmission of infectious agents.81-85 It has been reported that leukoreduction prior to RBC storage diminish the extent of storage induced RBC lesions.86,87 Hence, a lower degree of hemolysis, microvesiculation, potassium leakage and osmotic fragility was observed during refrigerated RBC storage as compared to their non-leukoreduced counterparts.80,88,89 In addition, leukoreduced refrigerated stored RBCs showed negligible PS exposure and adhesiveness to ECs with storage time.89-91 Interestingly, it was also observed that oxygen free radicals and enzymes derived from activated leukocytes can compromise the aggregation and deformation behavior of RBCs.86,92,93 Despite these findings, the aggregability and deformability (i.e. rheologic features) of leukoreduced refrigerated stored RBCs are yet to be determined. In general, leukoreduction has been widely regarded as a beneficial practice to transfusion medicine.81,94,95 However, whether leukoreduced refrigerated stored RBCs are functional in the microcirculation or might be deleterious to transfusion outcome is still unclear.15,96-99 As a result concerns regarding the infusion of prolonged stored RBCs are ongoing and a more restrictive transfusion policy is currently being favored.100,101 Alternatively, storage of RBC at ultra-low subzero temperatures halts the biochemical processes in the cell and could therefore represent a promising approach to circumvent storage induced lesions and subsequently to extend the shelf life of preserved RBCs.
1.3. Cryopreservation of RBCs Since the discovery in the 1950s that glycerol protects RBCs from freezing injury, cryopreserved RBCs have been under intensive investigation.48 Currently cryopreserved RBCs (commonly known as frozen RBCs) are primarily used for controlling an inventory in situations where the RBC availability is limited or unpredictable. Such as is the case for

Introduction
9
storage of RBC with rare blood groups or for usage in military settings.102,103 Occasionally cryopreservation is used to preserve autologous RBCs.
Freezing methods To date, there are two freezing methods approved for the preservation of RBCs.57,58 On the one hand, RBCs can be frozen rapidly using a low-glycerol method (LGM) with a final concentration of approximately 20% glycerol (wt/vol) at temperatures below -140°C. On the other hand, RBCs can be frozen slowly using a high-glycerol method (HGM), allowing storage of RBC units with a final concentration of approximately 40% (wt/vol) glycerol at temperatures between -65°C and -80°C. Cryopreserved RBCs can be preserved up to at least ten years if the correct storage temperature is guaranteed. The HGM cryopreserved RBCs tolerate wide fluctuations in temperature during freezing and are more stable during post-thaw storage.104,105 In addition, HGM cryopreserved RBCs did not require liquid nitrogen which eased storage and transportation conditions. Therefore, the HGM is currently the most applicable RBC freezing method in Europe and the United States.
Deglycerolization of thawed RBCs Usage of glycerol as a cryoprotectant requires an intensive deglycerolization washing procedure post-thaw in order to reduce the glycerol concentration within the cell. This is necessary, since incomplete deglycerolized RBCs will cause hemolytic transfusion reactions and renal failure after infusion.106,107 Washing also considerably reduces the amount of detrimental substances such as bioactive lipids, microparticles, cytokines, potassium and free Hb as well as leukocytes from the RBC unit.62,63,77,78,108-110 As a result, transfusion of cryopreserved RBCs are associated with less febrile transfusion reactions, alloimmunization as well as occurrences of TRALI and systemic inflammatory response syndrome (SIRS).48,111
Requirements of cryopreserved RBCs Preservation of RBCs at ultra-low subzero temperatures ceases the biological activity of the cells which enables them to be preserved for years.112,113 However, once thawed the shelf life of RBCs is limited. Deglycerolized RBCs are primarily stored in SAGM preservation

Chapter 1
10
solution for up to 48 hours or in AS-3 preservation solution for up to 14 days. Cryopreserved RBCs need to be deglycerolized to reduce the residual glycerol content to below 1%.104 Furthermore, international guidelines require that the hemolysis in the RBC units must remain below allowable levels (i.e. 0.8% in Europe and 1% in The United States) and that the RBC post-thaw recovery after deglycerolization (i.e. freeze-thaw-wash recovery) must exceed 80%. Also, at least 75% of cryopreserved RBCs must still circulate 24-hours after infusion.57,58 However, these guidelines also do not reflect the ability of RBCs to function after infusion. Hence, frozen storage subjects RBC to a range of chemical, thermal and mechanical forces, which might affect their oxygen delivering capacity after infusion.
Quality of thawed RBCs The quality of HGM cryopreserved RBCs is dependent on the pre-freeze and post-thaw storage time, the anticoagulant and additive solution used as well as on the biological variation among RBC units. The cooling process per se only minimally induces cellular damage when high glycerol concentrations were used.112,114 In general, RBCs intended for frozen storage are refrigerated stored prior to processing. The 2,3-DPG content of refrigerated stored RBCs rapidly declines due to the low pH of the storage solution. By limiting this pre-freezing storage time, higher 2,3-DPG values could be obtained post-thaw.114 Yet, the 2,3 DPG content continue to diminish during postthaw storage.115,116 Cryopreserved RBCs maintain high ATP content following post-thaw storage in SAGM and AS-3. However, during postthaw storage also the ATP content gradually declines. This decline is more prominent in AS-3 due to the lower pH of this storage solution.117 When the pre-freezing storage time was limited to three days no significant changes in PS exposure, CD-47 expression and membrane microvesiculation was observed with HGM cryopreserved RBCs that were post-thaw stored in SAGM solution.118 In the past the rheologic properties of cryopreserved RBCs have only briefly been addressed. It was demonstrated that the aggregability and deformability of LGM cryopreserved RBCs were diminished.119-121 Yet, the rheologic properties of HGM cryopreserved RBCs are still to be determined.

Introduction
11
Implementation of new freezing technology For routine clinical usage, cryopreserved RBCs must be safe, effective and reasonably priced. Cryopreserved RBCs are more expensive, inefficient and time consuming than refrigerated stored RBCs.48 However, ongoing technological advances could potentially expand the utilization of cryopreserved RBCs in transfusion medicine. A promising approach is the usage of a liquid Bio-freezer (Supachill, Lubbock, USA) with a high heat absorption capacity. This Bio-freezer consists of a reservoir of fluid in which samples are immersed. Consequently, the heat is rapidly dissipated from the sample and less freezing damage is observed. This Bio-freezer has been successfully used to freeze bovine sperm cells with a high post-thaw quality.122,123 The rate of heat release during freezing is also an important contributor to the post-thaw quality of RBCs.124 Traditional freezing of RBCs at –25°C in a mechanical freezer, normally requires high glycerol solutions to minimize ice formation.125 However, usage of high concentrations of cryoprotectants also exert detrimental effects on the cell.126 The rapid heat exchange of the liquid Bio-freezer, might allow RBCs to be frozen with lower glycerol concentrations at higher temperatures. The latter could reduce the osmotic stress and subsequently the freeze-thaw-wash related cell loss. This characteristic could make the Bio-freezer particular interesting for cryopreservation of RBCs.
1.4. Aim of this thesis The aim of this thesis is to gain a better understanding of the RBC quality from a rheologic perspective in transfusion medicine. Alternatively, the utilization of cryopreservation for long-term storage of RBCs will be explored.

Chapter 1
12
References
1. Giangrande PL. The history of blood transfusion. Br.J.Haematol. 2000, vol.110: 758-767.
2. Bray JJ, Cragg PA, Macknight ADC, Mills RG. Lecture notes on human physiology. fourth ed. Blackwell Science; 1999.
3. Betz T, Lenz M, Joanny JF, Sykes C. ATP-dependent mechanics of red blood cells. Proc.Natl.Acad.Sci. 2009, vol.106: 15320-15325.
4. Bucki R, Bachelot-Loza C, Zachowski A, Giraud F, Sulpice JC. Calcium induces phospholipid redistribution and microvesicle release in human erythrocyte membranes by independent pathways. Biochemistry. 1998, vol.37: 15383-15391.
5. Gov NS, Safran SA. Red blood cell membrane fluctuations and shape controlled by ATP-induced cytoskeletal defects. Biophys.J. 2005, vol.88: 1859-1874.
6. Gov NS. Active elastic network: cytoskeleton of the red blood cell. Phys.Rev.E.Stat.Nonlin.Soft.Matter Phys. 2007, vol.75: 011921.
7. Heaton WA. Evaluation of posttransfusion recovery and survival of transfused red cells. Transfus.Med.Rev. 1992, vol.6: 153-169.
8. Johnson RM. Membrane stress increases cation permeability in red cells. Biophys.J. 1994, vol.67:1876-1881.
9. Park Y, Best CA, Auth T, Gov NS, Safran SA, Popescu G, Suresh S, Feld MS. Metabolic remodeling of the human red blood cell membrane. Proc.Natl.Acad.Sci. 2010, vol.107: 1289-1294.
10. Verhoeven AJ, Hilarius PM, Dekkers DW, Lagerberg JW, de Korte D. Prolonged storage of red blood cells affects aminophospholipid translocase activity. Vox Sang. 2006, vol.91: 244-251.
11. Wan J, Ristenpart WD, Stone HA. Dynamics of shear-induced ATP release from red blood cells. Proc.Natl.Acad.Sci. 2008, vol.105: 16432-16437.
12. Andrews DA, Low PS. Role of red blood cells in thrombosis. Curr.Opin.Hematol. 1999, vol.6: 76-82.
13. Raat NJ, Verhoeven AJ, Mik EG, Gouwerok CW, Verhaar R, Goedhart PT, de Korte D, Ince C. The effect of storage time of human red cells on intestinal microcirculatory oxygenation in a rat isovolemic exchange model. Crit Care Med. 2005, vol.33: 39-45.
14. Raat NJ, Hilarius PM, Johannes T, de Korte D, Ince C, Verhoeven AJ. Rejuvenation of stored human red blood cells reverses the renal microvascular oxygenation deficit in an isovolemic transfusion model in rats. Transfusion. 2009, vol.49: 427-434.
15. Raat NJ, Ince C. Oxygenating the microcirculation: the perspective from blood transfusion and blood storage. Vox Sang. 2007, vol.93: 12-18.
16. Popel AS, Johnson PC. Microcirculation and Hemorheology. Annu.Rev.Fluid Mech. 2005 Jan 1;37:43-69.
17. Marossy A, Svorc P, Kron I, Gresova S. Hemorheology and circulation. Clin.Hemorheol.Microcirc. 2009, vol.42: 239-258.
18. Meiselman HJ. Red blood cell aggregation: 45 years being curious. Biorheology. 2009, vol.46: 1-19.
19. Rampling MW, Meiselman HJ, Neu B, Baskurt OK. Influence of cell-specific factors on red blood cell aggregation. Biorheology. 2004, vol.41: 91-112.

Introduction
13
20. Baumler H, Neu B, Donath E, Kiesewetter H. Basic phenomena of red blood cell Rouleaux formation. Biorheology. 1999, vol.36: 439-442.
21. Cabel M, Meiselman HJ, Popel AS, Johnson PC. Contribution of red blood cell aggregation to venous vascular resistance in skeletal muscle. Am.J.Physiol. 1997, vol.272: H1020-H1032.
22. Luquita A, Urli L, Svetaz MJ, Gennaro AM, Volpintesta R, Palatnik S, Rasia M. Erythrocyte aggregation in rheumatoid arthritis: Cell and plasma factor's role. Clin.Hemorheol.Microcirc. 2009, vol.41: 49-56.
23. Reggiori G, Occhipinti G, de Gasperi A, Vincent JL, Piagnerelli M. Early alterations of red blood cell rheology in critically ill patients. Crit Care Med. 2009, vol.37: 3041-3046.
24. Holme S. Current issues related to the quality of stored RBCs. Transfus.Apher.Sci. 2005, vol.33: 55-61.
25. Cicco G, Pirrelli A. Red blood cell (RBC) deformability, RBC aggregability and tissue oxygenation in hypertension. Clin.Hemorheol.Microcirc. 1999, vol.21: 169-177.
26. Yedgar S, Koshkaryev A, Barshtein G. The red blood cell in vascular occlusion. Pathophysiol. Haemost.Thromb. 2002, vol.32: 263-268.
27. Tomita M, Tanahashi N, Takeda H, Schiszler I, Osada T, Unekawa M, Suzuki N. Capillo-venous flow in the brain: significance of intravascular RBC aggregation for venous flow regulation. Clin.Hemorheol.Microcirc. 2006, vol.34: 51-57.
28. Mohandas N, Gallagher PG. Red cell membrane: past, present, and future. Blood. 2008, vol.112: 3939-3948.
29. Parthasarathi K, Lipowsky HH. Capillary recruitment in response to tissue hypoxia and its dependence on red blood cell deformability. Am.J.Physiol. 1999, vol.277: H2145-H2157.
30. Kaul DK, Koshkaryev A, Artmann G, Barshtein G, Yedgar S. Additive effect of red blood cell rigidity and adherence to endothelial cells in inducing vascular resistance. Am.J.Physiol Heart Circ.Physiol. 2008, vol.295: H1788-H1793.
31. Closse C, Dachary-Prigent J, Boisseau MR. Phosphatidylserine-related adhesion of human erythrocytes to vascular endothelium. Br.J.Haematol. 1999, vol.107: 300-302.
32. Manodori AB, Barabino GA, Lubin BH, Kuypers FA. Adherence of phosphatidylserine-exposing erythrocytes to endothelial matrix thrombospondin. Blood. 2000, vol.95: 1293-1300.
33. Fadok VA, Bratton DL, Rose DM, Pearson A, Ezekewitz RA, Henson PM. A receptor for phosphatidylserine-specific clearance of apoptotic cells. Nature. 2000, vol.405: 85-90.
34. Wu Y, Tibrewal N, Birge RB. Phosphatidylserine recognition by phagocytes: a view to a kill. Trends Cell Biol. 2006, vol.16: 189-197.
35. Schroit AJ, Madsen JW, Tanaka Y. In vivo recognition and clearance of red blood cells containing phosphatidylserine in their plasma membranes. J.Biol.Chem. 1985, vol.260: 5131-5138.
36. Machiedo GW, Zaets SB, Berezina TL, Xu DZ, Feketova E, Spolarics Z, Deitch EA. Trauma-hemorrhagic shock-induced red blood cell damage leads to decreased microcirculatory blood flow. Crit Care Med. 2009, vol.37: 1000-1010.
37. Mchedlishvili G. Disturbed blood flow structuring as critical factor of hemorheological disorders in microcirculation. Clin.Hemorheol.Microcirc. 1998, vol.19: 315-325.

Chapter 1
14
38. Den Uil CA, Klijn E, Lagrand WK, Brugts JJ, Ince C, Spronk PE, Simoons ML. The microcirculation in health and critical disease. Prog.Cardiovasc.Dis. 2008, vol.51: 161-170.
39. Ellis CG, Jagger J, Sharpe M. The microcirculation as a functional system. Crit Care. 2005, vol.9: S3-S8.
40. Hardeman MR, Dobbe JG, Ince C. The Laser-assisted Optical Rotational Cell Analyzer (LORCA) as red blood cell aggregometer. Clin.Hemorheol.Microcirc. 2001, vol.25: 1-11.
41. Hardeman MR, Ince C. Clinical potential of in vitro measured red cell deformability, a myth? Clin.Hemorheol.Microcirc. 1999, vol.21: 277-284.
42. Baskurt OK, Uyuklu M, Ulker P, Cengiz M, Nemeth N, Alexy T, Shin S, Hardeman MR, Meiselman HJ. Comparison of three instruments for measuring red blood cell aggregation. Clin.Hemorheol.Microcirc. 2009, vol.43: 283-298.
43. Baskurt OK, Hardeman MR, Uyuklu M, Ulker P, Cengiz M, Nemeth N, Shin S, Alexy T, Meiselman HJ. Comparison of three commercially available ektacytometers with different shearing geometries. Biorheology. 2009, vol.46: 251-264.
44. Connor J, Pak CC, Schroit AJ. Exposure of phosphatidylserine in the outer leaflet of human red blood cells. Relationship to cell density, cell age, and clearance by mononuclear cells. J.Biol.Chem.1994, vol.269:2399-4
45. Lang F, Gulbins E, Lerche H, Huber SM, Kempe DS, Foller M. Eryptosis, a window to systemic disease. Cell Physiol Biochem. 2008, vol.22: 373-380.
46. Antonelou MH, Kriebardis AG, Papassideri IS. Aging and death signalling in mature red cells: from basic science to transfusion practice. Blood Transfus. 2010, vol.8: S39-S47.
47. Waugh RE, Narla M, Jackson CW, Mueller TJ, Suzuki T, Dale GL. Rheologic properties of senescent erythrocytes: loss of surface area and volume with red blood cell age. Blood. 1992, vol.79: 1351-1358.
48. Hess JR. Red cell freezing and its impact on the supply chain. Transfus.Med. 2004, vol.14: 1-8.
49. Offner PJ. Age of blood: does it make a difference? Crit Care 2004, vol.8: S24-S26.
50. Koch CG, Li L, Sessler DI, Figueroa P, Hoeltge GA, Mihaljevic T, Blackstone EH. Duration of red-cell storage and complications after cardiac surgery. N.Engl.J.Med. 2008, vol.358: 1229-1239.
51. Weinberg JA, McGwin G, Jr., Marques MB, Cherry SA, III, Reiff DA, Kerby JD, Rue LW, III. Transfusions in the less severely injured: does age of transfused blood affect outcomes? J.Trauma. 2008, vol.65: 794-798.
52. Zallen G, Offner PJ, Moore EE, Blackwell J, Ciesla DJ, Gabriel J, Denny C, Silliman CC. Age of transfused blood is an independent risk factor for postinjury multiple organ failure. Am.J.Surg. 1999, vol.178: 570-572.
53. Purdy FR, Tweeddale MG, Merrick PM. Association of mortality with age of blood transfused in septic ICU patients. Can.J.Anaesth. 1997, vol.44: 1256-1261.
54. Vamvakas EC, Carven JH. Transfusion and postoperative pneumonia in coronary artery bypass graft surgery: effect of the length of storage of transfused red cells. Transfusion. 1999, vol.39: 701-710.
55. Keller ME, Jean R, LaMorte WW, Millham F, Hirsch E. Effects of age of transfused blood on length of stay in trauma patients: a preliminary report. J.Trauma. 2002, vol.53: 1023-1025.
56. Vlaar AP, Straat M, Juffermans NP. The relation between aged blood products and onset of transfusion-related acute lung injury. A review of pre-clinical data. Clin.Lab. 2011, vol.57: 267-272.

Introduction
15
57. Kakaiya R, Aronson CA, Julleis J. Whole blood collection and component processing, AABB Technical Manual 16th ed. Bethesda: AABB Press; 2008.
58. Council of Europe. Guide to the preparation, use and quality assurance of blood components, 14th edition. Strasbourg: Council of Europe publisher; 2008.
59. Van de Watering LM. Red cell storage and prognosis. Vox Sang. 2011, vol.100: 36-45.
60. Hess JR. Red cell storage. J.Proteomics. 2010, vol.73: 368-373.
61. Zimrin AB, Hess JR. Current issues relating to the transfusion of stored red blood cells. Vox Sang. 2009, vol.96: 93-103.
62. Kim-Shapiro DB, Lee J, Gladwin MT. Storage lesion: role of red blood cell breakdown. Transfusion. 2011, vol.51: 844-851.
63. Vlaar AP, Kulik W, Nieuwland R, Peters CP, Tool AT, van Bruggen R, Juffermans NP, de Korte D. Accumulation of bioactive lipids during storage of blood products is not cell but plasma derived and temperature dependent. Transfusion. 2011, vol.51: 2358-2366.
64. Relevy H, Koshkaryev A, Manny N, Yedgar S, Barshtein G. Blood banking-induced alteration of red blood cell flow properties. Transfusion. 2008, vol.48: 136-146.
65. Berezina TL, Zaets SB, Morgan C, Spillert CR, Kamiyama M, Spolarics Z, Deitch EA, Machiedo GW. Influence of storage on red blood cell rheological properties. J.Surg.Res. 2002, vol.102: 6-12.
66. Hovav T, Yedgar S, Manny N, Barshtein G. Alteration of red cell aggregability and shape during blood storage. Transfusion. 1999, vol.39: 277-281.
67. Izzo P, Manicone A, Spagnuolo A, Lauta VM, Di Pasquale A, Di Monte D. Erythrocytes stored in CPD SAG-mannitol: evaluation of their deformability. Clin.Hemorheol.Microcirc. 1999, vol.21: 335-339.
68. Kirkpatrick UJ, Adams RA, Lardi A, McCollum CN. Rheological properties and function of blood cells in stored bank blood and salvaged blood. Br.J.Haematol. 1998 , vol.101: 364-368.
69. Raat NJ, Berends F, Verhoeven AJ, de Korte D, Ince C. The age of stored red blood cell concentrates at the time of transfusion. Transfus.Med. 2005, vol.15: 419-423.
70. Corwin HL, Gettinger A, Pearl RG, Fink MP, Levy MM, Abraham E, MacIntyre NR, Shabot MM, Duh MS, Shapiro MJ. The CRIT Study: Anemia and blood transfusion in the critically ill--current clinical practice in the United States. Crit Care Med. 2004, vol.32: 39-52.
71. Fontaine MJ, Chung YT, Erhun F, Goodnough LT. Age of blood as a limitation for transfusion: potential impact on blood inventory and availability. Transfusion. 2010, vol.50: 2233-2239.
72. Lelubre C, Piagnerelli M, Vincent JL. Association between duration of storage of transfused red blood cells and morbidity and mortality in adult patients: myth or reality? Transfusion. 2009, vol.49: 1384-1394.
73. Van de Watering LM, Brand A. Effect of storage of red cells. Transfusion medicine and hemotherapy 2008, vol.35: 359-367.
74. Arnold DM. Bias in transfusion research: from study design to result reporting. Transfusion. 2011, vol.51: 237-240.
75. Van de Watering LM. Pitfalls in the current published observational literature on the effects of red blood cell storage. Transfusion. 2011, vol.51: 1847-1854.

Chapter 1
16
76. Middelburg RA, van de Watering LM, van der Bom JG. Blood transfusions: good or bad? Confounding by indication, an underestimated problem in clinical transfusion research. Transfusion. 2010, vol.50: 1181-1183.
77. Shanwell A, Kristiansson M, Remberger M, Ringden O. Generation of cytokines in red cell concentrates during storage is prevented by prestorage white cell reduction. Transfusion. 1997, vol.37: 678-684.
78. Glenister KM, Sparrow RL. Level of platelet-derived cytokines in leukoreduced red blood cells is influenced by the processing method and type of leukoreduction filter. Transfusion. 2010, vol.50: 185-189.
79. Willy C, Reithmeier W, Kuhlmann WD, Gerngross H, Flegel WA. Leukocyte depletion of red cell components prevents exposure of transfusion recipients to neutrophil elastase. Vox Sang. 2000,vol.78: 19-27.
80. Racek J, Herynkova R, Holecek V, Faltysova J, Krejcova I. What is the source of free radicals causing hemolysis in stored blood? Physiol Res. 2001, vol.50: 383-388.
81. Shapiro MJ. To filter blood or universal leukoreduction: what is the answer? Crit Care. 2004, vol.8:S27-S30.
82. Roddie PH, Turner ML, Williamson LM. Leucocyte depletion of blood components. Blood Rev. 2000, vol.14: 145-156.
83. Singh S, Kumar A. Leukocyte depletion for safe blood transfusion. Biotechnol.J. 2009, vol.4: 1140-1151.
84. Williamson LM. Leucocyte depletion of the blood supply - how will patients benefit? Br.J.Haematol. 2000, vol.110: 256-272.
85. Blajchman MA. The clinical benefits of the leukoreduction of blood products. J.Trauma.2006,.60:S83-S90.
86. Bratosin D, Leszczynski S, Sartiaux C, Fontaine O, Descamps J, Huart JJ, Poplineau J, Goudaliez F, Aminoff D, Montreuil J. Improved storage of erythrocytes by prior leukodepletion: flow cytometric evaluation of stored erythrocytes. Cytometry. 2001, vol.46: 351-356.
87. Phelan HA, Gonzalez RP, Patel HD, Caudill JB, Traylor RK, Yancey LR, Sperry JL, Friese RS, Nakonezny PA. Prestorage leukoreduction ameliorates the effects of aging on banked blood. J.Trauma. 2010, vol.69: 330-337.
88. Gyongyossy-Issa MI, Weiss SL, Sowemimo-Coker SO, Garcez RB, Devine DV. Prestorage leukoreduction and low-temperature filtration reduce hemolysis of stored red cell concentrates. Transfusion. 2005, vol.45: 90-96.
89. Anniss AM, Sparrow RL. Storage duration and white blood cell content of red blood cell (RBC) products increases adhesion of stored RBCs to endothelium under flow conditions. Transfusion. 2006, vol.46: 1561-1567.
90. Cardo LJ, Hmel P, Wilder D. Stored packed red blood cells contain a procoagulant phospholipid reducible by leukodepletion filters and washing. Transfus.Apher.Sci. 2008, vol.38: 141-147.
91. Bennett-Guerrero E, Veldman TH, Doctor A, Telen MJ, Ortel TL, Reid TS, Mulherin MA, Zhu H, Buck RD, Califf RM, et al. Evolution of adverse changes in stored RBCs. Proc.Natl.Acad.Sci. 2007, vol.104:17063-17068.
92. Baskurt OK, Meiselman HJ. Activated polymorphonuclear leukocytes affect red blood cell aggregability. J.Leukoc.Biol. 1998, vol.63: 89-93.

Introduction
17
93. Srour, Bilto YY, Juma M, Irhimeh MR. Exposure of human erythrocytes to oxygen radicals causes loss of deformability, increased osmotic fragility, lipid peroxidation and protein degradation. Clin.Hemorheol.Microcirc. 2000, vol.23: 13-21.
94. Blumberg N, Heal JM. Universal leukocyte reduction of blood transfusions. Clin.Infect.Dis. 2007, vol.45: 1014-1015.
95. Blumberg N, Sime PJ, Phipps RP. Leukoreduction of blood transfusions--there is such a thing as a free lunch. Crit Care Med. 2010, vol.38: 720-721.
96. Tsai AG, Cabrales P, Intaglietta M. Microvascular perfusion upon exchange transfusion with stored red blood cells in normovolemic anemic conditions. Transfusion. 2004, vol.44: 1626-1634.
97. Yuruk K, Almac E, Bezemer R, Goedhart P, De Mol B, Ince C. Blood transfusions recruit the microcirculation during cardiac surgery. Transfusion. 2011, vol.51: 961-967.
98. Walsh TS, McArdle F, McLellan SA, Maciver C, Maginnis M, Prescott RJ, McClelland DB. Does the storage time of transfused red blood cells influence regional or global indexes of tissue oxygenation in anemic critically ill patients? Crit Care Med. 2004, vol.32: 364-371.
99. Kiraly LN, Underwood S, Differding JA, Schreiber MA. Transfusion of aged packed red blood cells results in decreased tissue oxygenation in critically injured trauma patients. J.Trauma. 2009, vol.67: 29-32.
100. Corwin HL, Carson JL. Blood transfusion--when is more really less? N.Engl.J.Med. 2007,356: 1667-1669.
101. Varghese R, Myers ML. Blood conservation in cardiac surgery: let's get restrictive. Semin.Thorac.Cardiovasc.Surg. 2010, vol.22: 121-126.
102. Reesink HW, Engelfriet CP, Schennach H, Gassner C, Wendel S, Fontao-Wendel R, de Brito MA, Sistonen P, Matilainen J, Peyrard T, et al. Donors with a rare pheno (geno) type. Vox Sang. 2008, vol.95: 236-253.
103. Hess JR, Thomas MJ. Blood use in war and disaster: lessons from the past century. Transfusion. 2003, vol.43: 1622-1633.
104. Valeri CR, Ragno G. Cryopreservation of human blood products. Transfus.Apher.Sci. 2006, vol.34: 271-287.
105. Lelkens CC, Noorman F, Koning JG, Truijens de Lange R, Stekkinger PS, Bakker JC, Lagerberg JW, Brand A, Verhoeven AJ. Stability after thawing of RBCs frozen with the high- and low-glycerol method. Transfusion. 2003, vol.43: 157-164.
106. Cregan P, Donegan E, Gotelli G. Hemolytic transfusion reaction following transfusion of frozen and washed autologous red cells. Transfusion. 1991, vol.31: 172-175.
107. Bechdolt S, Schroeder LK, Samia C, Schmidt PJ. In vivo hemolysis of deglycerolized red blood cells. Arch.Pathol.Lab Med. 1986, vol.110: 344-345.
108. Silliman CC, Moore EE, Kelher MR, Khan SY, Gellar L, Elzi DJ. Identification of lipids that accumulate during the routine storage of prestorage leukoreduced red blood cells and cause acute lung injury. Transfusion. 2011, vol.51: 2549-2554.
109. Bandarenko N, Hay SN, Holmberg J, Whitley P, Taylor HL, Moroff G, Rose L, Kowalsky R, Brumit M, Rose M, et al. Extended storage of AS-1 and AS-3 leukoreduced red blood cells for 15 days after deglycerolization and resuspension in AS-3 using an automated closed system. Transfusion. 2004, vol.44: 1656-1662.

Chapter 1
18
110. Arnaud FG, Meryman HT. WBC reduction in cryopreserved RBC units. Transfusion. 2003, vol.43: 517-525.
111. Valeri CR, Ragno G. An approach to prevent the severe adverse events associated with transfusion of FDA-approved blood products. Transfus.Apher.Sci. 2010, vol.42: 223-233.
112. Valeri CR, Ragno G, Pivacek LE, Cassidy GP, Srey R, Hansson-Wicher M, Leavy ME. An experiment with glycerol-frozen red blood cells stored at -80°C for up to 37 years. Vox Sang. 2000, vol.79: 168-174.
113. Valeri CR, Srey R, Tilahun D, Ragno G. The in vitro quality of red blood cells frozen with 40 percent (wt/vol) glycerol at -80°C for 14 years, deglycerolized with the Haemonetics ACP 215, and stored at 4°C in additive solution-1 or additive solution-3 for up to 3 weeks. Transfusion.2004,vol.44:990-995.
114. Lecak J, Scott K, Young C, Hannon J, Acker JP. Evaluation of red blood cells stored at -80 °C in excess of 10 years. Transfusion. 2004, vol.44: 1306-1313.
115. Bohonek M, Petras M, Turek I, Urbanova J, Hradek T, Chmatal P, Staroprazska V, Kostirova J, Horcickova D, Duchkova S, et al. Quality evaluation of frozen apheresis red blood cell storage with 21-day postthaw storage in additive solution 3 and saline-adenine-glucose-mannitol: biochemical and chromium-51 recovery measures. Transfusion. 2010, vol.50: 1007-1013.
116. Valeri CR, Ragno G, Van HP, Rose L, Rose M, Egozy Y, Popovsky MA. Automation of the glycerolization of red blood cells with the high-separation bowl in the Haemonetics ACP 215 instrument. Transfusion. 2005, vol.45: 1621-1627.
117. Lagerberg JW, Truijens-de Lange R, de Korte D, Verhoeven AJ. Altered processing of thawed red cells to improve the in vitro quality during postthaw storage at 4°C. Transfusion. 2007, vol.47: 2242-2249.
118. Holovati JL, Wong KA, Webster JM, Acker JP. The effects of cryopreservation on red blood cell microvesiculation, phosphatidylserine externalization, and CD47 expression. Transfusion. 2008, vol.48: 1658-1668.
119. Wegner G, Kucera W. Changes in erythrocyte deformability in blood preservation. Z.Gesamte Inn.Med. 1987, vol.42: 575-581.
120. Chien S, Usami S, Rowe AW. Rheologic properties of erythrocytes preserved in liquid nitrogen. J.Lab Clin.Med. 1971, vol.78: 175-180.
121. Hardeman MR, Lagerberg JWM. Rheology of 10 year old frozen and thawed blood. Possible use of frozen RBCs as standard reference material in hemorheology. Biorheology. 2002, vol.39: 650-651.
122. Goolsby HA, Blanton JR, Cotter PZ, Prien SD. Preliminary trial: motility comparisons of a unique freezing technology (UFT) to liquid nitrogen mist methodology for cryopreservation of porcine spermatozoa. Reprod.Domest.Anim 2004, vol.39: 328-332.
123. Cotter PZ, Goolsby HA, Prien SD. Preliminary evaluation of a unique freezing technology for bovine spermatozoa cryopreservation. Reprod.Domest.Anim 2005, vol.40: 98-99.
124. Rinfret AP. Factors affecting the erythrocyte during rapid freezing and thawing. Ann.N.Y.Acad.Sci. 1960, vol.85: 576-594.
125. Fahy GM, Levy DI, Ali SE. Some emerging principles underlying the physical properties, biological actions, and utility of vitrification solutions. Cryobiology. 1987, vol.24: 196-213.
126. Fahy GM. Cryoprotectant toxicity neutralization. Cryobiology. 2010, vol.60: S45-S53.

Chapter 2
Use of hydroxyethyl starch for inducing red blood cell aggregation
Sandra Henkelman
Gerhard Rakhorst Henny C. van der Mei Henk J. Busscher
Accepted for publication in the journal of:
Clinical Hemorheology and Microcirculation
(Reproduced with permission of IOS press)

Chapter 2
20
Abstract The aggregation of human RBC remains of biological and clinical interest. Replacement of plasma proteins by polymers to induce RBC aggregation may help to unravel the fundamentals of the aggregation process. Two theories exist to explain the RBC aggregation mechanism: a depletion and a bridging theory. RBC aggregation induced by hydroxyethyl starch (HES) polymers increases with polymer size, which suggests that aggregation is induced via the bridging theory. In the present study, the electrophoretic mobility (EPM) was measured to investigate RBC aggregation induced by 200-kDa HES polymers. In addition, we evaluated if these polymers were useful for demonstrating aggregation differences between RBCs from healthy and type-1 diabetes mellitus (T1DM) subjects. Our results demonstrate that the EPM values of RBCs in 200-kDa HES solutions were less negative than could be predicted by the viscosity of the suspension, supporting the bridging theory. Furthermore, aggregation analysis using the LORCA demonstrated that 200-kDa HES solution enhanced RBC aggregation of healthy and diabetic subjects in a similar manner as standard 500-kDa dextran solutions. In conclusion: our data supports the bridging mechanism underlying 200-kDa HES induced RBC aggregation. In addition, both polymers are useful for demonstrating cellular induced aggregation differences between RBCs from healthy and T1DM subjects.

Polymer induced RBC aggregation
21
2.1. Introduction RBC aggregation is an important determinant of the flow behavior of blood. In regions with low shear rate, human RBCs form linear stacks of cells and multi-cellular aggregates that markedly enhance the blood viscosity and further reduce the blood flow rate.1 Under physiological conditions RBC aggregates are easily dispersed when the blood flow rate increases. However, under pathologic conditions stronger and larger aggregates may form that are more resistant to dispersion by the blood flow, therewith contributing to the occlusion of microvessels and hindering adequate tissue perfusion.2,3 Despite numerous studies, the physiological role of RBC aggregation still remains unclear. It has been demonstrated that different RBC aggregation tendencies can exist between healthy individuals.4 In addition, enhanced RBC aggregation is observed in a variety of diseases such as cardiovascular disease, diabetes mellitus, renal failure, sepsis, thalassemia and sickle cell disease.3,5,6 The RBC surface consists of a layer of proteins, called the glycocalyx, which bears a net negative charge. The ability of RBCs to aggregate is a balance between repulsive and attractive forces. On one hand RBC aggregates are dispersed by repulsive electrostatic forces between negatively charged RBCs as well as the prevailing shear forces generated by the blood flow. On the other hand the promotion of aggregation is induced by the presence of plasma proteins, especially fibrinogen, or by high molecular weight polymers.5,7 Usage of high molecular weight polymers to induce RBC aggregation gives more insight in the aggregation process because it will exclude aggregation differences that are induced by variations in plasma protein composition. Non-ionic polymers like dextran, polyethylene glycol and to a lesser extent hydroxyethyl starch (HES) have been investigated for their RBC aggregation inducing properties.8-14 Like dextran, HES polymers are used in plasma substitutes to treat for example hypovolemia during surgery.15,16 HES polymers are available in a broad range of molecular weights all with different tendencies to promote RBC aggregation. In this regard, only HES polymers with low aggregation tendency, such as the 130-kDa HES solutions, are currently preferred for infusion, whereas HES polymers with hyper-aggregating tendencies, such as 450-kDa HES solutions, are less favored in clinical practice.11,17,18 However, the 200-kDa HES polymer, which has intermediate RBC aggregation tendencies, could be useful as a pro-aggregant in RBC rheologic studies.

Chapter 2
22
At present there are two theories that explain the RBC aggregation mechanism induced by high molecular weight polymers.19 In the bridging theory, it is hypothesized that macromolecules adsorbed onto the RBC membrane form bridges between adjacent cell surfaces, which help overcome repulsive electrostatic forces between the RBCs. In the depletion theory, RBC aggregation occurs due to a lower concentration of macromolecules near the RBC surface compared to the suspending medium. This depletion of macromolecules near the cell surface leads to an osmotic gradient that causes fluid to move away from adjacent RBCs, which subsequently favors RBC aggregation. Previous data support the depletion theory as the mechanism of dextran induced RBC aggregation.13,20 The mechanism through which HES induces RBC aggregation has not yet been established. However, since stronger and larger RBC aggregates are formed with HES solutions of increasing molecular weight,18 an adsorption mediated aggregation mechanism is expected to exist for this polymer. This is in contrast with the depletion mechanism, where RBC aggregation has a nonlinear dependency on the molecular weight of the polymer.8 Electrophoresis has shown to be a useful tool for studying the aggregation behavior of RBCs.9,13,21,22 By measuring the electrophoretic mobility (EPM) of RBCs in polymer solutions, a better understanding of the underlying aggregation mechanism can be obtained. The EPM of RBCs in saline solutions demonstrates a negative value. In this regard a less negative EPM of RBCs in polymer solutions (i.e. a lower mobility of RBCs) suggests bridging mediated aggregation due to interactions of polymers with the RBC surface. In contrast, a more negative EPM value (i.e. a higher mobility of RBCs) is explained by depletion of polymers near the RBC surface.23 Differentiating between plasmatic and cellular factors leading to altered RBC aggregation is important from both a biological and clinical point of view. Usage of high molecular weight polymers will manifest RBC aggregation differences which are induced by cellular factors such as RBC shape, deformability and membrane surface properties. This intrinsic tendency of RBC to aggregate (i.e. RBC aggregability) is dependent on the type and the molecular weight of polymers in solution.4 In the past standard dextran polymers have been useful pro-aggregants in RBC rheologic studies.5,24 Like the standard 500-kDa dextran polymers, the 200-kDa HES polymers could be useful to manifest cellular induced aggregation differences between RBCs from healthy individuals and patients with a disease. Blood from type-1 diabetes mellitus (T1DM)

Polymer induced RBC aggregation
23
patients is especially suitable for aggregation testing. Particularly, because in these patients alterations in both plasma and RBC cellular properties are responsible for enhanced RBC aggregation tendencies.25 Nowadays, RBC aggregation can be studied using the LORCA. This technique allows RBC aggregation to be studied ex vivo either in whole blood or in polymer solutions. In this study, the EPM of RBCs from healthy volunteers was explored in 200-kDa HES and standard 500-kDa dextran solutions in order to elucidate the underlying aggregation mechanism. Furthermore, the LORCA was used to investigate if these polymers could discriminate cellular induced aggregation differences between RBCs from healthy volunteers and T1DM patients. 2.2. Materials and methods
Sample preparation Blood was collected from healthy volunteers and T1DM patients after informed consent and in accordance with University Medical Center Groningen protocols. Briefly, whole blood was anticoagulated with EDTA (1.5 mg/ml) and part of it was used in aggregation experiments within 1 hour after collection. To obtain RBCs, whole blood was centrifuged at 1100 x g for 12 min to remove the buffycoat and supernatant. The concentrated RBCs were washed twice with PBS (pH 7.4), and resuspended in 0.9% sodium chloride solution (NaCl; pH 4.5 or pH 7.4) to a final hematocrit (Hct) of 40 ± 1%. Aggregation experiments and EPM measurements with washed RBCs were performed within 4 h after collection. A 10% HES solution with a molecular weight of 200-kDa and a molar substitution of 0.5 was obtained from Fresenius, 's-Hertogenbosch, the Netherlands. Dextran fractions with a molecular weight of 500-kDa were obtained from Sigma-Aldrich, Steinheim, Germany. A 2% dextran solution (wt/vol) was prepared by dissolving this polymer into 0.9% NaCl solution with pH 7.4.
Electrophoretic mobility and viscosity measurements The speed of RBCs in an electrical field was determined in suspension using a Laser Zee Meter 501 equipped with image analysis options (PenKem, Bedford Hills, N.Y.), as has been described previously.26 Briefly, RBCs were suspended in either 0.9% NaCl pH 7.4,

Chapter 2
24
0.9% NaCl pH 4.5, 10% 200-kDa HES pH 4.5 or 2% 500-kDa dextran pH 7.4 solutions at a Hct of 0.05%. The electrophoresis chamber was filled with the RBC suspension and a voltage difference of 150V was applied over the chamber. EPM values were measured at 20°C in duplicate with RBCs obtained from four different healthy volunteers. EPM values were obtained from samples that contained a minimum of 35 RBCs per suspending solution. The viscosities of the suspending solutions were measured in duplicate at 20°C using a viscometer (DV-II+ Pro, Brookfield, USA). All EPM values are reported as measured and are not corrected for differences in viscosity between the various solutions.
Aggregation measurements RBC aggregation was assessed in vitro, by a laser-assisted optical rotational cell analyzer (LORCA; R&R Mechatronics, Zwaag, the Netherlands).27 Aggregation was tested with 1 ml of either whole blood or with RBCs suspended in 200-kDa HES or 500-kDa dextran solutions. In short, RBC suspensions were centrifuged for 1 min at 3500 x g and the supernatant was discarded. RBCs were resuspended in either 10% 200-kDa HES or 2% 500-kDa dextran solutions, to obtain a Hct between 35 ± 5%. The aggregation of the RBCs was monitored after complete disaggregation under increased shear stresses. Both the aggregation measuring procedure and the subsequent analyses were computer controlled. Aggregation of RBCs was expressed by the aggregation index (AI), where a larger AI reflects an increased ability to aggregate. The AI depends on the kinetics and extent of aggregation. The AI was determined after correcting the Hct in all the samples to a constant value of 45%. The kinetics of aggregation (T1/2) was expressed by the time necessary to induce 50% aggregation. All measurements were performed at 37°C.
Statistical analysis Statistical analysis was performed using statistical software (SPSS, version 16.0, SPSS Inc., Chicago, IL). Data were tested for normality with the Kolmgorov-Smirnov goodness-of-fit test. The EPM values were analyzed by using a repeated-measure ANOVA. Post hoc comparisons were performed to quantify differences between suspension solutions by using paired t-tests. Aggregation differences between RBCs from healthy and T1DM subjects were determined by using unpaired t-tests. Within subjects, paired t-tests were performed to show

Polymer induced RBC aggregation
25
differences with regard to whole blood. A Bonferroni correction was applied to correct for multiple comparisons. Differences are considered to be significant with a two-tailed p-value of less than 0.05. Results are presented as means ± SD.
2.3. Results
RBC electrophoretic mobility and viscosity The EPM of RBCs suspended in polymer-free saline solutions was not significantly affected by the pH. RBCs suspended in 200-kDa HES solutions showed significantly less negative EPM values as compared to RBCs suspended in polymer-free solutions (-0.04 vs -0.58 10-8m2V-1s-1). However, the EPM values of RBCs suspended in 200-kDa HES solutions were less negative, than would be expected based upon the inverse relation between suspension medium viscosity and EPM values found (Figure 2.1A).28 In contrast, RBCs suspended in 500-kDa dextran solutions demonstrated significantly more negative EPM values as compared to RBCs suspended in polymer-free solutions (-1.40 vs –0.74 10-8m2V-1s-1). However, these observed EPM values were considerable more negative than would be expected based upon the inverse relation between the suspending medium viscosity and the EPM values found (Figure 2.1B).
RBC aggregation Whole blood from T1DM subjects showed an increased tendency to aggregate as compared to that of healthy subjects (p < 0.05: Figure 2A). Moreover, the AI induced by 200-kDa HES solution is higher with RBCs from T1DM subjects than that of healthy subjects (p < 0.05). However, it was notable that the AI induced by 200-kDa HES polymers was similar to the AI induced by the 500-kDa dextran polymers. No significant differences in the aggregation half-time between whole blood from healthy and T1DM subjects (Figure 2B) were observed. The 200-kDa HES solution markedly lowered the T1/2 as compared to that observed with whole blood of healthy and T1DM subjects (p < 0.05), indicating a faster aggregation process with the use of this polymer. However, both polymers did not induce significant differences in T1/2 between RBCs from healthy and T1DM subjects.

Chapter 2
26
A
B
Figure 2.1. The electrophoretic mobility of human RBCs suspended in 200-kDa HES (A) or standard 500-kDa dextran (B) solutions as a function of their viscosity. The dashed line is the predicted relation between EPM and suspension viscosity assuming an inverse relation between these variables. Values are expressed as mean of eight experiments.

Polymer induced RBC aggregation
27
A
B
Figure 2.2. Aggregation indexes (A) and aggregation half-time (B) for RBCs obtained from healthy and T1DM human subjects in either autologous plasma or RBCs resuspended in polymer solutions. Values are expressed as the mean ± SD of ten different subjects. Significant changes are illustrated in the figure (p < 0.05); * significantly different from whole blood, † significantly different from its healthy counterpart.

Chapter 2
28
2.4. Discussion The found EPM values of healthy RBCs suspended in 200-kDa HES solution supports the bridging theory as the mechanism underlying HES induced RBC aggregation. Although 200-kDa HES and 500-kDa dextran polymers induce RBC aggregation via different mechanisms, we demonstrated that both polymers are useful pro-aggregants for manifesting cellular induced aggregation differences between RBCs from healthy and T1DM subjects. Aggregation of RBCs is a physiological phenomenon that has been studied for decades.5 Although RBC aggregation is an important determinant of blood viscosity in regions of low shear rate, the physiological role of this process still remains elusive. Nevertheless, enhanced RBC aggregation has been observed in various diseases.3,5,6 The use of standard polymer suspensions to induce RBC aggregation will give more insight in the fundamentals of the aggregation process. Currently, HES polymers are primarily used as plasma expanders.15,16 However, studies demonstrating that 200-kDa HES polymers could be useful pro-aggregants in rheologic studies were lacking. In the present study, the EPM values of RBCs suspended in 200-kDa HES solution were considerable less negative than the linear reciprocal viscosity change that is predicted by the Helmholtz–Smoluchowski equation.28 The EPM of RBCs in an electrical field is determined by the viscosity of the suspending fluid as well as the RBC surface charge density and the glycocalyx thickness.5,23,29 Given that HES polymers are nonionic, it is unlikely that these EPM changes are associated with alterations in RBC surface charge density. Instead, absorption of nonionic HES polymers onto the RBC surface may have attributed to structural changes within and near the RBC glycocalyx, explaining the less negative EPM values.30 Clearly, our EPM measurements underline the presence of a bridging mechanism between RBCs when using 200-kDa HES polymers. In contrast, dextran induced aggregation is characterized by an optimal aggregation tendency at a molecular weight of approximately 500-kDa.8 In this study, the EPM values of RBC in 500-kDa dextran solutions became more negative despite the higher viscosity of the bulk suspending medium. These findings are in agreement with a depletion interaction in which a lower viscosity near the RBC surface compared to the bulk suspending medium, enhances the EPM of the RBCs.19 A drawback with current depletion theories is that polymer absorption onto RBC membranes are often not taken into account due to experimental limitations.5,8,13 Nevertheless, a depletion theory does not require an absolute

Polymer induced RBC aggregation
29
lack of absorption on RBCs in order to initiate aggregation, but merely that the absorbed polymer level is less than the polymers present in the bulk suspending medium.5,31 Recently, Liu et al. have provided new insights for characterizing absorption of polymers onto RBCs.32 In their study tritium labeling was used to quantify the in vitro absorption of polymers onto the RBCs, while laser scanning confocal microscopy was used to locate the precise binding sites. Future aggregation studies that allow the implementation of an absorption interaction with these techniques are therefore warranted. In the present study, whole blood from T1DM human subjects showed an increased tendency to aggregate as opposed to whole blood from healthy subjects. It has been recognized that a disturbed glucose homeostasis in T1DM subjects results in elevated plasma fibrinogen concentrations and a reduced anionic RBC surface charge.25,33 The increased AI observed with whole blood from T1DM subjects is thus a result of changes in both plasma and RBC cellular properties. Usage of high molecular weight polymers, exclude aggregation differences induced by variations in plasma protein compositions. In this study, the 200-kDa HES and standard 500-kDa dextran solutions markedly enhanced the AI of RBCs as compared to whole blood. Nevertheless, both polymer solutions were clearly able to discriminate aggregation differences between RBCs from healthy and T1DM subjects. Our results therefore demonstrate that also 200-kDa HES polymers are useful in manifesting RBC aggregation differences which are of cellular origin. The parameter AI depends both on the kinetics (t1/2) and extent of aggregation. In this study no significant differences in t1/2 between healthy and T1DM subjects with either whole blood or polymer induced RBC aggregation could be observed. This indicated that aggregates are not necessarily formed faster but that the extent of aggregation is the determining factor when it comes to manifesting differences between RBCs from healthy and T1DM human subjects. The ability of RBCs to form aggregates in the presence of plasma proteins may play a pivotal role in maintaining hemodynamics. RBCs flow from the endothelial wall into the center of the blood vessel where they form aggregates. While physiological values of RBC aggregation direct leukocytes and possibly platelets from the axial core towards the vascular wall,34 intensified RBC aggregation may hinder or obstruct the blood flow in micro-vessels. More insight in the RBC aggregation process is therefore important from both a biological and clinical point of view.

Chapter 2
30
In the past, the depletion theory has been particularly favored as the mechanism of non-ionic polymer induced RBC aggregation. Although the 200-kDa HES solutions induced similar aggregation tendencies as the standard 500-kDa dextran solutions, our data supports the bridging theory as the mechanism underlying HES induced aggregation. Since RBCs from healthy volunteers show significant variations in terms of their aggregation tendency,4 maybe in vivo both aggregation mechanisms can exist depending on the protein composition of plasma. Furthermore, our results demonstrate that like the standard 500-kDa dextran solutions, the 200-kDa HES solutions are useful for manifesting cellular induced aggregation differences between RBCs from healthy and T1DM subjects.
References 1. Marossy A, Svorc P, Kron I, Gresova S. Hemorheology and circulation. Clin.Hemorheol.Microcirc. 2009,
vol 42: 239-258.
2. Mchedlishvili G, Varazashvili M, Gobejishvili L. Local RBC aggregation disturbing blood fluidity and causing stasis in microvessels. Clin.Hemorheol.Microcirc. 2002, vol 26: 99-106.
3. Yedgar S, Koshkaryev A, Barshtein G. The red blood cell in vascular occlusion. Pathophysiol. Haemost.Thromb. 2002, vol 32: 263-268.
4. Rampling MW, Meiselman HJ, Neu B, Baskurt OK. Influence of cell-specific factors on red blood cell aggregation. Biorheology. 2004, vol 41: 91-112.
5. Meiselman HJ. Red blood cell aggregation: 45 years being curious. Biorheology. 2009, vol 46: 1-19.
6. Reggiori G, Occhipinti G, de Gasperi A, Vincent JL, Piagnerelli M. Early alterations of red blood cell rheology in critically ill patients. Crit Care Med. 2009, vol 37: 3041-3046.
7. Ben-Ami R, Barshtein G, Mardi T, Deutch V, Elkayam O, Yedgar S, Berliner S. A synergistic effect of albumin and fibrinogen on immunoglobulin-induced red blood cell aggregation. Am.J.Physiol Heart Circ.Physiol. 2003, vol 285: H2663-H2669.
8. Neu B, Wenby R, Meiselman HJ. Effects of dextran molecular weight on red blood cell aggregation. Biophys.J. 2008, vol 95: 3059-3065.
9. Baskurt OK, Tugral E, Neu B, Meiselman HJ. Particle electrophoresis as a tool to understand the aggregation behavior of red blood cells. Electrophoresis 2002, 23: 2103-2109.
10. Pribush A, Zilberman-Kravits D, Meyerstein N. The mechanism of the dextran-induced red blood cell aggregation. Eur.Biophys.J. 2007, vol 36: 85-94.
11. Neff TA, Fischler L, Mark M, Stocker R, Reinhart WH. The influence of two different hydroxyethyl starch solutions (6% HES 130/0.4 and 200/0.5) on blood viscosity. Anesth.Analg. 2005, vol 100: 1773-1780.
12. Armstrong JK, Meiselman HJ, Fisher TC. Covalent binding of poly(ethylene glycol) (PEG) to the surface of red blood cells inhibits aggregation and reduces low shear blood viscosity. Am.J.Hematol. 1997, 56:26-28.

Polymer induced RBC aggregation
31
13. Rad S, Gao J, Meiselman HJ, Baskurt OK, Neu B. Depletion of high molecular weight dextran from the red cell surface measured by particle electrophoresis. Electrophoresis 2009, vol 30: 450-456.
14. Mosbah IB, Franco-Gou R, Abdennebi HB, Hernandez R, Escolar G, Saidane D, Rosello-Catafau J, Peralta C. Effects of polyethylene glycol and hydroxyethyl starch in University of Wisconsin preservation solution on human red blood cell aggregation and viscosity. Transplant.Proc. 2006, vol 38: 1229-1235.
15. Jungheinrich C, Neff TA. Pharmacokinetics of hydroxyethyl starch. Clin.Pharmacokinet. 2005, vol 44: 681-699.
16. Treib J, Baron JF, Grauer MT, Strauss RG. An international view of hydroxyethyl starches. Intensive Care Med. 1999, vol 25: 258-268.
17. Kozek-Langenecker SA, Jungheinrich C, Sauermann W, Van der Linden P. The effects of hydroxyethyl starch 130/0.4 (6%) on blood loss and use of blood products in major surgery: a pooled analysis of randomized clinical trials. Anesth.Analg. 2008, vol 107: 382-390.
18. Morariu AM, van der Plaats A, van Oeveren W, 'T Hart NA, Leuvenink HG, Graaff R, Ploeg RJ, Rakhorst G. Hyperaggregating effect of hydroxyethyl starch components and University of Wisconsin solution on human red blood cells: a risk of impaired graft perfusion in organ procurement? Transplantation. 2003, vol 76: 37-43.
19. Baumler H, Neu B, Donath E, Kiesewetter H. Basic phenomena of red blood cell rouleaux formation. Biorheology. 1999, vol 36: 439-442.
20. Neu B, Meiselman HJ. Depletion-mediated red blood cell aggregation in polymer solutions. Biophys.J. 2002, vol 83: 2482-2490.
21. Sabolovic D, Sestier C, Perrotin P, Guillet R, Tefit M, Boynard M. Covalent binding of polyethylene glycol to the surface of red blood cells as detected and followed up by cell electrophoresis and rheological methods. Electrophoresis. 2000, vol 21: 301-306.
22. Neu B, Armstrong JK, Fisher TC, Baumler H, Meiselman HJ. Electrophoretic mobility of human red blood cells coated with poly(ethylene glycol). Biorheology. 2001, vol 38: 389-403.
23. Baumler H, Donath E, Krabi A, Knippel W, Budde A, Kiesewetter H. Electrophoresis of human red blood cells and platelets. Evidence for depletion of dextran. Biorheology. 1996, vol 33: 333-351.
24. Baskurt OK, Bor-Kucukatay M, Yalcin O, Meiselman HJ. Aggregation behavior and electrophoretic mobility of red blood cells in various mammalian species. Biorheology. 2000, vol 37: 417-428.
25. Singh M, Shin S. Changes in erythrocyte aggregation and deformability in diabetes mellitus: a brief review. Indian J.Exp.Biol. 2009, vol 47: 7-15.
26. Van Merode AE, Duval JF, van der Mei HC, Busscher HJ, Krom BP. Increased adhesion of Enterococcus faecalis strains with bimodal electrophoretic mobility distributions. Colloids Surf.B Biointerfaces. 2008, vol 64: 302-306.
27. Hardeman MR, Dobbe JG, Ince C. The Laser-assisted Optical Rotational Cell Analyzer (LORCA) as red blood cell aggregometer. Clin.Hemorheol.Microcirc. 2001, vol 25: 1-11.
28. Mewis J, Wagner NJ. Colloidal suspension rheology. Cambridge: Cambridge university press; 2011.

Chapter 2
32
29. Jan KM, Chien S. Role of surface electric charge in red blood cell interactions. J.Gen.Physiol. 1973, vol 61: 638-654.
30. Neu B, Sowemimo-Coker SO, Meiselman HJ. Cell-cell affinity of senescent human erythrocytes. Biophys.J. 2003, vol 85: 75-84.
31. Van Oss CJ, Arnold K, Coakley WT. Depletion flocculation and depletion stabilization of erythrocytes. Cell Biophys. 1990, vol 17: 1-10.
32. Liu Z, Janzen J, Brooks DE. Adsorption of amphiphilic hyperbranched polyglycerol derivatives onto human red blood cells. Biomaterials. 2010, vol 31: 3364-3373.
33. Le Devehat C, Vimeux M, Khodabandehlou T. Blood rheology in patients with diabetes mellitus. Clin.Hemorheol.Microcirc. 2004, vol 30: 297-300.
34. Nash GB, Watts T, Thornton C, Barigou M. Red cell aggregation as a factor influencing margination and adhesion of leukocytes and platelets. Clin.Hemorheol.Microcirc. 2008, vol 39: 303-310.

Chapter 3
Red blood cell aggregation; an important phenomenon in damage control resuscitation?
Sandra Henkelman Michael Piagnerelli Gerhard Rakhorst

Chapter 3
34
Abstract To improve the survival of severely bleeding trauma patients, a damage control resuscitation strategy has been recommended. This strategy includes the early infusion of fresh frozen plasma (FFP), platelets and RBCs in a 1:1:1 unit ratio to control the bleeding and favor survival of these patients. Although lower FFP to RBC ratios have been linked to higher rates of mortality, these high ratios have been associated with adverse outcomes as well. The formation of RBC aggregates in regions with low shear rate could play a key role in these findings. Administration of FFP and thus fibrinogen is essential for coagulation. Yet, fibrinogen also promotes RBC aggregation. Although physiological levels of RBC aggregation support the hemostasis, promotion of aggregation could be disadvantageous in patients in which the RBC rheology is already compromised, as was observed in certain trauma states. Notably, enhanced RBC aggregation may hinder tissue perfusion and contribute to the occlusion of micro-vessels. We consider that RBC aggregation could play an important role in damage control resuscitation of severely injured trauma patients.

RBC aggregation in trauma patients
35
Uncontrolled bleeding is the leading cause of death in trauma patients. It was recognized that approximately 25% of severely injured trauma patients are coagulopathic upon admission and that these patients are three times more likely to die than those without it.1 This acute coagulopathy of trauma has been attributed to multiple factors such as loss, dilution and consumption of coagulation factors and platelets as well as to fibrinolysis, hypothermia and metabolic acidosis.2 To improve the survival of severely bleeding trauma patients, an early damage control resuscitation strategy has been recommended. This resuscitation approach prima1rily advocates limited crystalloid use, prevention and treatment of acidosis and hypothermia, as well as an early administration of fresh frozen plasma (FFP), platelets and RBCs in a 1:1:1 unit ratio.3 Although the optimal blood component ratio is still a matter of debate, the general consensus is that an early resuscitation approach with this high FFP to RBC ratio, controls the bleeding and potentially favors survival of severely bleeding trauma patients.4 Limited attention has been addressed, to the mechanism by which a high FFP to RBC ratio influences survival in these trauma patients. Recently, the improved survival has been linked to inhibition of vascular endothelial permeability and subsequently diminished interstitial edema.5 Yet the ability of RBCs to form aggregates in the presence of plasma proteins, especially fibrinogen, could also play a pivotal role in damage control resuscitation. The formation of RBC aggregates in regions with low shear rate is a physiological phenomenon that has been studied for decades. Although RBC aggregation is a major determinant of the whole blood viscosity at low shear rate, the physiological role of this process is still elusive. Under normal physiological conditions, RBC aggregates are easily dispersed by the rise in blood flow rate. However, under pathological conditions stronger and or larger RBC aggregates are formed which are more resistant to dispersion by shear forces. Enhanced RBC aggregation may impair the blood flow in the microcirculation and contribute to the occlusion of micro-vessels, which may induce local hypoxia and damage to endothelial cells.6,7 In this regard, the LORCA is a useful device, which allows RBC aggregation to be studied ex vivo. In cases of massive bleeding, fibrinogen is the first coagulation factor that reaches critically low levels. Administration of blood components in an 1:1:1 unit ratio will replenish depleted coagulation factors and platelets and minimize dilutional coagulopathy, as is the case when only RBCs or volume expanders will be administrated.8 Yet, in a trauma setting

Chapter 3
36
the use of FFP in massive bleedings has also been questioned. Adverse outcome such as increased incidence of nosocomial infections, multiple organ failure, lung injury and death have been linked to the usage of FFP.9 Studies have shown mixed results, but in general, high FFP to RBC ratios have been associated with adverse outcomes whereas low FFP to RBC ratios have been linked to increased rates of mortality.1,10 For many years, it has been recognized that RBCs can actively participate in clot formation by enhancing platelet de-granulation and by recruitment of additional platelets into the forming clot.11 The role of RBC aggregation in hemostasis has been given less attention. RBCs migrate from the endothelial wall into the center of the blood vessel where they form aggregates. RBC aggregates exclude leukocytes and possibly platelets from the axial core and direct them towards the vascular wall.12,13 This process is essential since leukocytes and platelets need to get into close contact with the damaged endothelium, in order to exert their function. RBC aggregation increases proportionally with fibrinogen levels.14 Administration of FFP, platelets and RBCs in a 1:1:1 unit ratio, a composition that approximates whole blood, could promote RBC aggregation. On the one hand promotion of aggregation would be beneficial for supporting hemostasis in severely injured trauma patients. Especially, since it has been recognized that people with leukocyte adherence deficiency suffer from recurrent bacterial infections and impaired wound healing and because it has been recognized that infections remain a concerning complication of combat-related injuries.15,16 On the other hand, promotion of aggregation could be detrimental to patients in which the RBC rheology is already compromised, as was observed in certain trauma states.17-19 In this regard, enhanced RBC aggregation could subsequently hamper tissue perfusion and contribute to the occlusion of micro-vessels. Enhanced RBC aggregation, which was also evident during long-term storage of non-leukoreduced RBC units, could furthermore explain the finding that blood component infusion was less effective than fresh whole blood in supporting hemostasis of trauma patients.3,20,21 The above mentioned data underline the potential importance of RBC aggregation in damage control resuscitation. Most studies regarding resuscitation practices are retrospective. Although these studies are limited inherently to their retrospective design, they do provide interesting hypotheses. Early FFP infusion is considered lifesaving in severely bleeding trauma patients. Yet, to determine the influence of high FFP to RBC ratios on promoting RBC aggregation in these patients, ex vivo aggregation testing will be necessary. In this regard, the LORCA could be

RBC aggregation in trauma patients
37
useful to demonstrate RBC aggregation tendencies after damage control resuscitation of severely bleeding trauma patients.
References
1. Nascimento B, Callum J, Rubenfeld G, Neto JB, Lin Y, Rizoli S. Clinical review: Fresh frozen plasma in massive bleedings, more questions than answers. Crit Care. 2010, vol.14: 202.
2. Murthi SB, Stansbury LG, Hess JR. Blood and coagulation support in trauma. Blood Rev. 2009, vol.23: 149-155.
3. Spinella PC, Holcomb JB. Resuscitation and transfusion principles for traumatic hemorrhagic shock. Blood Rev. 2009, vol.23: 231-240.
4. Ho AM, Dion PW, Yeung JH, Ng CS, Karmakar MK, Critchley LA, Rainer TH, Cheung CW, Tay BA. Fresh-frozen plasma transfusion strategy in trauma with massive and ongoing bleeding. Common (sense) and sensibility. Resuscitation. 2010, vol.81: 1079-81.
5. Pati S, Matijevic N, Doursout MF, Ko T, Cao Y, Deng X, Kozar RA, Hartwell E, Conyers J, Holcomb JB. Protective effects of fresh frozen plasma on vascular endothelial permeability, coagulation, and resuscitation after hemorrhagic shock are time dependent and diminish between days 0 and 5 after thaw. J.Trauma. 2010, vol.69: S55-S63.
6. Yedgar S, Koshkaryev A, Barshtein G. The red blood cell in vascular occlusion. Pathophysiol. Haemost.Thromb. 2002, vol.32:263-268.
7. Mchedlishvili G. Disturbed blood flow structuring as critical factor of hemorheological disorders in microcirculation. Clin.Hemorheol.Microcirc. 1998, vol.19: 315-325.
8. Fries D, Martini WZ. Role of fibrinogen in trauma-induced coagulopathy. Br.J.Anaesth. 2010,105: 116-121.
9. Spahn DR, Ganter MT. Towards early individual goal-directed coagulation management in trauma patients. Br.J.Anaesth. 2010, vol.105: 103-105.
10. Wafaisade A, Maegele M, Lefering R, Braun M, Peiniger S, Neugebauer E, Bouillon B. High plasma to red blood cell ratios are associated with lower mortality rates in patients receiving multiple transfusion (4≤ red blood cell units <10) during acute trauma resuscitation. J.Trauma. 2011, vol.70: 81-88.
11. Andrews DA, Low PS. Role of red blood cells in thrombosis. Curr.Opin.Hematol. 1999, vol.6: 76-82.
12. Nash GB, Watts T, Thornton C, Barigou M. Red cell aggregation as a factor influencing margination and adhesion of leukocytes and platelets. Clin.Hemorheol.Microcirc. 2008, vol.39: 303-310.
13. Pearson MJ, Lipowsky HH. Effect of fibrinogen on leukocyte margination and adhesion in postcapillary venules. Microcirculation. 2004, vol.11: 295-306.
14. Falco C, Vaya A, Simo M, Contreras T, Santaolaria M, Aznar J. Influence of fibrinogen levels on erythrocyte aggregation determined with the Myrenne aggregometer and the Sefam erythro-aggregometer. Clin.Hemorheol.Microcirc. 2005, vol.33: 145-151.

Chapter 3
38
15. Van Vliet DN, Brandsma AE, Hartwig NG. Leukocyte-adhesion deficiency: a rare disorder of inflammation. Ned.Tijdschr.Geneeskd. 2004, vol.148: 2496-2500.
16. Murray CK, Wilkins K, Molter NC, Li F, Yu L, Spott MA, Eastridge B, Blackbourne LH, Hospenthal DR. Infections complicating the care of combat casualties during operations Iraqi Freedom and Enduring Freedom. J.Trauma 2011, vol.71:S62-S73.
17. Machiedo GW, Zaets SB, Berezina TL, Xu DZ, Feketova E, Spolarics Z, Deitch EA. Trauma-hemorrhagic shock-induced red blood cell damage leads to decreased microcirculatory blood flow. Crit Care Med. 2009, vol.37: 1000-1010.
18. Tatarishvili J, Sordia T, Mchedlishvili G. Comparison of blood rheological changes in the microcirculation during experimental hemorrhagic and traumatic shock. Clin.Hemorheol.Microcirc. 2006, vol.35: 217-221.
19. Reggiori G, Occhipinti G, de Gasperi A, Vincent JL, Piagnerelli M. Early alterations of red blood cell rheology in critically ill patients. Crit Care Med. 2009, vol.37: 3041-3046.
20. Hovav T, Yedgar S, Manny N, Barshtein G. Alteration of red cell aggregability and shape during blood storage. Transfusion. 1999, vol.39: 277-281.
21. Phelan HA, Eastman AL, Aldy K, Carroll EA, Nakonezny PA, Jan T, Howard JL, Chen Y, Friese RS, Minei JP. Prestorage leukoreduction abrogates the detrimental effect of aging on packed red cells transfused after trauma: a prospective cohort study. Am.J.Surg. 2012, vol.203:198-204.

Chapter 4
Is red blood cell rheology preserved during routine blood bank storage?
Sandra Henkelman
Margriet J. Dijkstra-Tiekstra Janny de Wildt-Eggen Reindert Graaff Gerhard Rakhorst Willem van Oeveren
Transfusion 50: 941-948, 2010
(Reproduced with permission of John Wiley and Sons)

Chapter 4
40
Abstract RBCs refrigerated stored for more than 2 weeks at 4°C are currently considered of impaired quality. This opinion has among others been based on altered RBC rheologic properties (i.e. enhanced aggregability, reduced deformability and elevated EC interaction) observed during storage of non-leukoreduced RBC units. Nevertheless, with the implementation of leukoreduction the storage-induced lesions have considerably diminished. In this study, the aggregability and deformability of leukoreduced RBCs during routine blood bank storage were investigated. At the blood bank, ten leukoreduced RBC units were refrigerated stored in SAGM preservation solution for up to 7 weeks. RBCs were weekly tested for aggregability, deformability and other hematologic variables. The RBC aggregability was significantly reduced after the first week of storage but recovered during the following weeks. After 7 weeks of storage the aggregability was slightly reduced (from 46.9 to 44.3 AI; p < 0.05). During storage the osmotic fragility was not significantly enhanced and the deformability at a shear stress of 3.9 Pa was not significantly reduced. The deformability at a shear stress of 50 Pa was reduced (from 0.58 to 0.54 EI; p < 0.05) but remained within physiological values (0.53 ± 0.04). During 5 weeks of storage the ATP content was reduced by 54% whereas the MCV, pH and MCHC were minimally affected. We conclude that the rheologic properties of leukoreduced RBC units were well preserved during routine blood bank storage.

Rheologic properties of refrigerated stored RBCs
41
4.1. Introduction During refrigerated storage at 4°C, RBCs undergo physical and biochemical alterations collectively referred to as the storage lesion.1 Recent publications suggest that transfusion of long-term refrigerated stored RBCs are associated with adverse clinical outcome in critical ill, cardiac surgery, and trauma patients.2 The RBC rheology, that is, the ability to aggregate, deform and adhere to ECs, are important determinants of the blood flow and hence the oxygen delivery to the tissues.3,4 Aggregation takes place in the venous system where RBCs form linear stacks of cells or multi-cellular aggregates at low shear rates.5 Normally the increasing blood flow is sufficient to disperse these aggregates. However, under pathologic conditions stronger and larger aggregates may form, which are more resistant to dispersion by the blood flow. The RBC ability to deform due to applied forces makes these cells capable of passing the capillaries. High RBC deformability and a rapid recovery to the normal shape are therefore essential factors for maintaining tissue perfusion.6 The RBC deformability is also a major determinant of the posttransfusion survival, since less deformable cells will be sequestered and destroyed in the spleen.7 Elevated adherence of RBCs to ECs can reduce the blood flow in the microcirculation and activate ECs, contributing to the occlusion of micro-vessels.3, 8 Alterations in RBC rheology have been observed in a variety of diseases such as cardiovascular disease, hypertension, diabetes mellitus, renal failure, malaria, thalassemia and sickle cell disease.3,9,10 Transfusion of rheologic impaired RBCs may hinder or obstruct the blood flow in micro-vessels leading to impaired tissue perfusion, ischemia or infarction.3,4 Therefore, rheologic impaired RBCs may form a hemodynamic risk particularly in recipients with circulatory and /or cardiovascular disorders. Long-term refrigerated storage may alter the RBC rheologic properties and adversely influence transfusion outcome. In vitro studies with non-leukoreduced RBCs demonstrated enhanced aggregability, reduced RBC deformability and elevated adherence to ECs already after the second week of storage.11-14 Since 2002, leukoreduction of RBC units is a standard procedure in many European countries.15 Pre-storage leukofiltration reduces RBC damage caused by cytokines and enzymes derived from activated leukocytes.16,17 As a result leukoreduced RBC units demonstrate a lower degree of hemolysis, potassium leakage, osmotic fragility, and free radical production during storage.18,19 Interestingly, it was also demonstrated that activated

Chapter 4
42
leukocytes induce cellular changes that adversely affect the rheologic properties of the RBCs.20,21 The in vitro aggregability and deformability of leukoreduced RBCs during routine blood bank storage remains to be determined and may contribute to the ongoing discussion of safety of blood transfusions. In this study the aggregability, deformability and other hematologic variables of leukoreduced refrigerated stored RBC were investigated during a 7-week period.
4.2. Materials and Methods
Preparation and sampling of RBC units Blood (500ml ± 10%) was collected from ten volunteers donors at the Sanquin blood bank in a quadruple top and -bottom bag system (Composelect, Fresenius Hemocare, the Netherlands) containing 70 ml of CPD anticoagulant. After cooling under butandiol plates for at least 4 hours, whole blood was separated into plasma, buffycoat and RBCs using an automated blood processing device (Compomat G4, Fresenius HemoCare, the Netherlands). SAGM solution (110 ml) was transferred to the RBCs and in-line filtration was carried out to remove residual leukocytes. The resulting RBC suspension had a Hct of 45-60% and contained less than 106 leukocytes per unit. RBCs were refrigerated stored at 4 ± 2 °C for 7 weeks. RBCs were released for use on day 3 after donation, hereafter referred to as Time 0, which were the freshest RBCs routinely available for transfusion. Weekly, samples were aseptically withdrawn from the RBC units for analysis after gentle mixing by inversion.
Rheologic features RBC aggregability and deformability were monitored in vitro by the LORCA (R&R Mechatronics, Zwaag the Netherlands).22,23 Aggregation was induced by the addition of 10% HES (MW 200-kDa). Briefly, RBCs suspensions were centrifuged for 1 minute at 3500 x g and the supernatant was discarded. RBCs were resuspended in 10% HES 200-kDa solution (Fresenius, Bad Homburg, Germany). The Hct in all the samples was corrected to a constant value of 45%. Aggregability was tested with 1 ml of the RBC suspension. Aggregation of RBCs was monitored after disaggregating under increased shear stress. Both the aggregation measuring procedure and the subsequent analyses were computer

Rheologic properties of refrigerated stored RBCs
43
controlled. Aggregability of RBC was expressed by the AI, where a larger AI reflects an increased ability to aggregate. The deformability of stored RBCs was determined with RBC suspension diluted 1:100 in PBS, (pH 6.5), containing 5% polyvinylpyrrolidone (PVP; MW 360 kDa, Sigma-Aldrich, Germany) and with a viscosity of 30 mPa.sec. One ml of the latter RBC suspension was inserted into the LORCA and the RBC diffraction pattern was recorded at various shear stresses at 36.8 ± 0.2°C. The deformability of the RBCs, which is expressed by the EI, was determined by the LORCA from the size of the vertical (L) and horizontal (W) axes of the diffraction pattern according to the formula: EI = (L-W) / (L+W). An increased EI at a given shear stress indicates greater RBC deformability. A deformability curve was obtained by plotting the calculated values for EI versus the corresponding shear stress. The deformability at two shear stress values were examined more closely; the deformability at a shear stress of 3.9 Pa, which reflects the rigidity of the cell membrane, and the maximal deformability at shear stress of 50 Pa. Since the freshest available RBCs for transfusion were already 3 days old, the deformability was also performed with RBCs that were obtained within 2 hours after donation from healthy donors.
Osmotic fragility The osmotic fragility of RBCs, which reflects the membrane’s ability to maintain structural integrity, was determined by diluting RBCs in PBS solutions ranging from 0.90% to 0.35%. RBCs with a Hct level of 30 to 35% were diluted 1:100 in each PBS solution, mixed and incubated for 30 minutes at 4°C, followed by centrifugation for 12 minutes at 1100 x g. The free Hb in the supernatant was measured by a spectrophotometer (PowerWave 200 spectrophotometer, Bio-Tek Instruments, USA) The concentration of PBS necessary to induce 50% hemolysis defined the osmotic fragility index of the RBCs.18 With this method, a larger osmotic fragility index corresponds to more fragile cells.
Hemolysis Hemolysis as measured by the amount of free Hb present in the RBC suspensions was determined according to the method of Harboe.24 Briefly, cell supernatant was obtained by centrifugation of RBC units for 1 minute at 3500 x g. The supernatant was diluted 1:10 in 0.01% sodium carbonate in a flat-bottom 96-well microtiter plate and mixed for 30 minutes.

Chapter 4
44
The Hb concentration in the supernatant was determined with a spectrophotometer by measuring the optical density (OD) at 415 nm and correcting for the OD at 380 and 450 nm according to the formula OD = 2* (OD 415 nm) – (OD 380 nm) – (OD 450 nm). The hemolysis was expressed as a percentage of the total amount of Hb present in the RBC lysates.
Hematologic variables To determine the cellular ATP content, RBC samples were incubated for 30 minutes with 8% ice cold trichloroacetic acid in a ratio of 1:3. Samples were centrifuged for 1 minute at 3500 x g and the protein free supernatant was neutralized with 1.5 mol/L sodium carbonate. Aliquots were stored at –80°C for later batch analyses of ATP. The ATP content was determined with a commercially available enzyme assay (Roche Diagnostics, Germany). For detection of ATP, light emission was measured at 560 nm by an illuminometer (Fluostar Optima, BMG Labtech, Germany). The supernatant pH and the RBC MCV were determined with a blood gas analyzer (Rapidlab 860, Siemens, the Netherlands). Total Hb content and Hct were determined with a hematology analyzer (Sysmex K4500, Goffin Meyvis, the Netherlands). The internal viscosity of RBC, as reflected by the MCHC, was determined by dividing the Hb content by the Hct.
Statistical analysis Statistical analysis was performed using statistical software (SPSS, version 16.0, SPSS Inc., Chicago, IL). Data were tested for normality with the Kolmgorov-Smirnov goodness-of-fit test. For each variable a repeated measure analysis of variance was performed to identify subject by time profiles. Post-hoc comparisons were performed to quantify differences between Time 0 and stored RBC values, using paired t-tests. Differences are considered to be significant with a p value of less than 0.05. Results are presented as means ± SD.

Rheologic properties of refrigerated stored RBCs
45
4.3. Results
Rheologic features The RBC ability to aggregate, as represented by the AI, was reduced after the first week of storage (AI from 46.9 ± 2.4 % to 41.9 ± 3.6 %; p < 0.01; Figure 4.1). In the following weeks the aggregability recovered; after week 2, 3, and 4, the RBC AIs were 45.2 ± 4.9 %, 46.0 ± 3.8 % and respectively 45.8 ± 3.5 %, which was not significantly different from Time 0 (46.9 ± 2.4 %). After 5 and 7 weeks of storage the aggregability was significantly reduced (AI 44.4 ± 4.5 % and respectively 44.3 ± 2.2 %) compared to Time 0. The RBC deformation curve showed a typical s-shape over a shear stress range of 0.6-50 Pa for all different time points (Figure 4.2). The deformation curve of 5 week old RBCs was slightly higher at the low-shear-stress regions and somewhat lower at the high-shear-stress regions compared to Time 0 (Figures 4.2 and 4.3). After 7 weeks of storage (outdated RBCs) the deformability at the high-shear-stress regions was further diminished (Figures. 4.2 and 4.3B). The rigidity of the cell membrane, which is reflected by the deformability at a shear stress of 3.9 Pa, was fluctuating during 5 weeks of storage (Figure 4.3A). Ultimately, after 7 weeks of storage the deformability at this low shear stress was not significantly different from Time 0 (from 0.35 ± 0.01 to 0.35 ± 0.02 EI). The deformability at a shear stress of 50 Pa was reduced from week 1 of storage onward (from 0.58 ± 0.01 to 0.54 ± 0.01 EI) as can be seen in Figure 4.3B. This downward trend in deformability (p < 0.01) remained within the physiological range, as determined with fresh RBCs (0.53 ± 0.04 EI).
Osmotic fragility and hemolysis The osmotic fragility index, represented by the osmolarity at half-maximum hemolysis of the RBCs, was not significantly altered during 7 weeks of storage (0.47 ± 0.02 % PBS; Table 4.1) compared to Time 0. A gradual increase in hemolysis was observed throughout the storage period (Table 4.1). After 5 weeks of storage, the hemolysis (0.53 ± 0.24%) still remained below allowable levels (i.e., 0.8% in Europe and 1% in the United States),15, 25 despite the weekly removal of samples. Closer evaluation of individual RBC units demonstrated that after 5 and 7 weeks of storage, respectively, one and four out of ten samples contained hemolysis which

Chapter 4
46
exceeded 0.8%. After 7 weeks of storage one RBC unit even showed hemolysis exceeding 2%, accounting for the large SD. No bacterial contamination was found in this RBC unit.
Figure 4.1. Effect of storage on RBCs ability to aggregate. AI in percentage and corrected for Hct. Values are expressed as mean ± SD of ten RBC units. Significant difference from Time 0 are shown (* p < 0.05 and † p < 0.01).
Figure 4.2. Shear stress EI curves for Time 0 (■) and after 5 (-) and 7 weeks (▲) of storage. The shear stress value is plotted on the logarithmic axis. Data represent mean ± SD of ten RBC units

Rheologic properties of refrigerated stored RBCs
47
A
B
Figure 4.3. Deformability for two representative shear stress levels as a function of storage time. (A) EI at shear stress of 3.9 Pa. With the exception of week 7, all samples were significantly altered (p < 0.05) compared to Time 0. (B) EI at shear stress of 50 Pa. All samples were significantly reduced compared to Time 0 during 7 weeks of storage (p < 0.04). Data represent mean ± SD of ten RBC units.
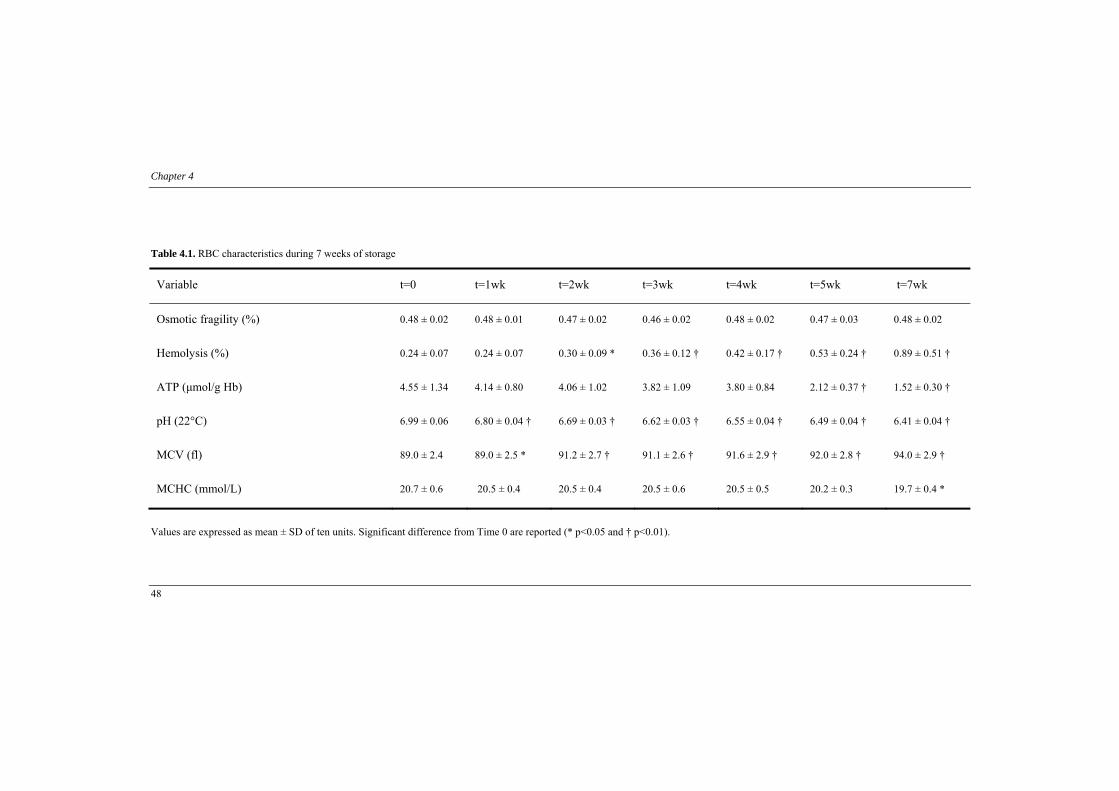
Chapter 4
48
Table 4.1. RBC characteristics during 7 weeks of storage
Variable t=0 t=1wk t=2wk t=3wk t=4wk t=5wk t=7wk
Osmotic fragility (%) 0.48 ± 0.02 0.48 ± 0.01 0.47 ± 0.02 0.46 ± 0.02 0.48 ± 0.02 0.47 ± 0.03 0.48 ± 0.02
Hemolysis (%) 0.24 ± 0.07 0.24 ± 0.07 0.30 ± 0.09 * 0.36 ± 0.12 † 0.42 ± 0.17 † 0.53 ± 0.24 † 0.89 ± 0.51 †
ATP (μmol/g Hb) 4.55 ± 1.34 4.14 ± 0.80 4.06 ± 1.02 3.82 ± 1.09 3.80 ± 0.84 2.12 ± 0.37 † 1.52 ± 0.30 †
pH (22°C) 6.99 ± 0.06 6.80 ± 0.04 † 6.69 ± 0.03 † 6.62 ± 0.03 † 6.55 ± 0.04 † 6.49 ± 0.04 † 6.41 ± 0.04 †
MCV (fl) 89.0 ± 2.4 89.0 ± 2.5 * 91.2 ± 2.7 † 91.1 ± 2.6 † 91.6 ± 2.9 † 92.0 ± 2.8 † 94.0 ± 2.9 †
MCHC (mmol/L) 20.7 ± 0.6 20.5 ± 0.4 20.5 ± 0.4 20.5 ± 0.6 20.5 ± 0.5 20.2 ± 0.3 19.7 ± 0.4 *
Values are expressed as mean ± SD of ten units. Significant difference from Time 0 are reported (* p<0.05 and † p<0.01).

Rheologic properties of refrigerated stored RBCs
49
Hematologic variables The RBC ATP content reduced during 7 weeks of storage (Table 4.1). Notably, after 5 weeks of storage the ATP content had declined with 54% compared to Time 0 (from 4.6 to 2.1 μmol/gHb; p < 0.01). After the first week of storage the PH of the RBC suspension and the MCV were significantly altered (Table 4.1). Ultimately, after 5 weeks of storage the pH was reduced with 7% while the MCV was enhanced by 3%. The MCHC was only significantly reduced after 7 weeks of storage compared to Time 0 (from 20.7 to 19.7 mmol/L).
4.4. Discussion This study was undertaken to explore the rheologic properties and quality of leukoreduced refrigerated stored RBC units. The RBC aggregability was significantly reduced at the first week of storage but recovered during the following weeks. RBC deformability at shear stress of 50 Pa was significantly reduced during 7 weeks of storage. However, values were still within physiological reference ranges. The osmotic fragility did not change significantly over time. Throughout 5 weeks of storage the ATP content had declined with 54% from initial levels, whereas MCV, PH and MCHC were affected to a lesser degree. During storage at 4°C RBCs undergo different biochemical and structural alterations that may adversely affect clinical outcome. Currently, RBCs are considered of impaired quality after the second week of storage.11,13 As a result, a more restrictive transfusion strategy is currently favored.26 Nonetheless, pre storage leukofiltration has significantly diminished storage-induced lesions.18,19 Additionally, it has been shown, that activated leukocytes can induce damage which adversely affects the rheologic properties of RBCs.20,21 Increased aggregability and diminished cell deformability during RBC storage have been previously observed. However, in those studies the RBC units had not been leukoreduced prior to storage.11-14, 27 In this study, RBC aggregability significantly decreased after 1 week of storage but recovered throughout the following weeks. At the end of storage, aggregability was slightly but significantly reduced. RBC aggregation is primarily dependent on the RBC surface characteristics and the composition of the suspension medium.3 Since the latter was standardized for all the samples, our results suggest that the observed differences are caused

Chapter 4
50
by alterations in cellular properties. In addition, our results show that RBCs are, to some extent, able to adapt to environmental changes during storage. RBC morphology can affect RBC aggregation. During storage at 4°C, RBC morphology shifts from discocyte towards echinocyte shape, leading to decreased aggregability.28,29 Our results show that the aggregability in SAGM solution was minimally reduced at the end of storage. Usage of other additive solutions, which also enhances the MCV during storage, are therefore expected to give a similar aggregation pattern. The clinical relevance of altered RBC aggregability, however, remains an ongoing debate.30 The RBC ability to deform depends mainly on the visco-elastic properties of the cytoskeleton, the intracellular viscosity of the RBC and the overall cell shape.31 In this study storage induced minor changes in the deformation curve at low shear stress of 3.9 Pa. The decreasing trend in deformability at a shear stress of 50 Pa has been interpreted as structural changes in the RBC that cannot be corrected by increasing shear force.32 However, we suggest that the gradual increase in MCV during storage may be responsible for the observed reduction in deformability, particularly since at high shear stress the cell volume becomes a limiting factor for the ability of RBCs to deform. Our data suggests that this effect plays a role for shear stresses above 5 Pa, as can been seen from the deformation curve. The increased RBC deformation at high shear stress after 5 weeks of storage compared to the preceding week has been observed by others and can only partly be explained by hemolysis of less deformable RBCs.11 The occurring deformability changes in this study were not likely biologically relevant because the reduction in deformation observed only at high shear stress was minimal and within physiological reference ranges. Furthermore, shear stresses of 3.9 Pa, which are predominantly found in the microcirculation,33 are clinically more relevant than those of 50 Pa. We showed that the osmotic fragility, which is determined by applying deforming stress from inside the RBC, was not significantly enhanced during 7 weeks of storage. In addition, the RBC intracellular viscosity, which is predominantly determined by the MCHC, did not yield significant changes until 7 weeks of storage. Taken together, these findings further substantiate that the observed reduction in deformability was caused primarily by morphology changes of the RBC. It has previously been shown, that RBC deformability was unaffected at pH values ranging between 6.4 and 7.7.34 In our study, the pH was still within these limits after 5 weeks of

Rheologic properties of refrigerated stored RBCs
51
storage, suggesting that the reduced RBC deformability was not caused by pH alterations in the storage solution. ATP as an energy source is important for the overall functioning of the cell. Loss of ATP is associated with more rigid cell membranes, loss of vasodilatation properties, exposure of PS on the outer leaflet of the RBC membrane, microvesiculation and decreased RBC viability.35-39 In our study the ATP content gradually reduced during storage. Although loss of ATP stiffens the RBC membrane due to calcium accumulation,40 the observed reduction in deformability at high shear stress was within physiological reference values, indicating that ATP loss during storage only marginally affected the RBC ability to deform. As proposed earlier, the RBC ATP content must be at least 2.7 µmol per gram Hb to have a 90 percent chance of acceptable in vivo survival (24-hr in vivo recovery of 75% or higher).41,42 The present study showed that after 5 weeks of storage the RBC ATP content was below this limit. Similar findings were observed for leukoreduced RBC that were stored for 6 weeks in SAGM solution.43 The in vivo viability of RBCs may be reduced by the low ATP content. However, it is not likely that this results in the proposed adverse clinical outcome, in particular, because the observed alterations in rheologic features were minimal and the hemolysis in the RBC units was still below the allowable limits (i.e. 0.8% in Europe and 1% in the United States).15, 25 Leukocytes can affect the RBC deformability even when they are significantly reduced, e.g. during the storage of buffycoat-depleted RBC units.44-46 Apparently, removing solely the buffycoat from the RBCs without performing leukofiltration, still resulted in 108 to 109
leukocytes per RBC units,15, 47 whereas nowadays less than 106 leukocytes per RBC unit are permitted. Our results demonstrate that cell deformability was minimally affected when the RBCs were leukofiltrated prior to storage. Recently, reduced deformability at shear stress of 3 and 30 Pa during the storage of leukoreduced RBCs has been observed,48 whereas others showed no reduction in RBC deformability at these shear stresses.49 This reduction in deformability has been linked to loss of Hb-bound nitric oxide, a variable that participates in controlling the RBC deformation.50 However, we also suggest that differences in storage procedures could explain this discrepancy in results. Particularly because the reduction in deformability at low and high shear stress was observed in the preservative solution AS-3, while in the SAGM preservation solution28 less changes in RBC deformability were observed.

Chapter 4
52
RBC adherence to ECs is mediated by PS exposure on the cell membrane. Additionally, PS expression triggers recognition by macrophages and subsequent clearance of RBCs from the circulation.51,52 PS expression was not determined in this study, since recent work demonstrated that leukoreduction significantly lowered the PS expression on the RBCs, resulting in negligible PS exposure after 5 weeks of storage48,53 and because leukoreduction reduces the progressive adherence of RBCs to ECs with storage time.54, 55 In the Netherlands approximately 37% of all the transfused RBC units are older than three weeks.56 The primary goal of blood transfusion is to deliver oxygen to the microcirculation with high-quality stored RBCs. During storage at 4°C, RBCs undergo different alterations that might influence the patient condition. However, leukoreduction has significantly reduced storage-induced lesions. Based on our findings, we postulate that the observed changes in RBC variables during refrigerated storage minimally affected the RBC ability to aggregate and deform, even after long-term storage. The rheologic properties of leukoreduced RBC units were well preserved during routine blood bank storage and not likely to contribute to adverse clinical outcome after transfusion.
References 1. Zimrin AB, Hess JR. Current issues relating to the transfusion of stored red blood cells. Vox Sanq. 2009,
vol.96: 93-103.
2. Lelubre C, Piagnerelli M, Vincent JL. Association between duration of storage of transfused red blood cells and morbidity and mortality in adult patients: myth or reality? Transfusion 2009, vol.49: 1384-1394
3. Yedgar S, Koshkaryev A, Barshtein G. The red blood cell in vascular occlusion. Pathophysiol Haemost Thromb. 2002, vol.32: 263-268.
4. Mchedlishvili G. Disturbed blood flow structuring as critical factor of hemorheological disorders in microcirculation. Clin Hemorheol Microcirc. 1998, vol.19: 315-325.
5. Barshtein G, Wajnblum D, Yedgar S. Kinetics of linear rouleaux formation studied by visual monitoring of red cell dynamic organization. Biophys J. 2000, vol.78: 2470-2474.
6. Parthasarathi K, Lipowsky HH. Capillary recruitment in response to tissue hypoxia and its dependence on red blood cell deformability. Am J Physiol. 1999, vol.277: H2145-H2157.
7. Mohandas N, Chasis JA. Red blood cell deformability, membrane material properties and shape: regulation by transmembrane, skeletal and cytosolic proteins and lipids. Semin Hematol. 1993, vol.30: 171-192.
8. Kaul DK, Koshkaryev A, Artmann G, Barshtein G, Yedgar S. Additive effect of red blood cell rigidity and adherence to endothelial cells in inducing vascular resistance. Am J Physiol Heart Circ Physiol. 2008, vol.295: H1788-H1793.

Rheologic properties of refrigerated stored RBCs
53
9. Rampling MW, Meiselman HJ, Neu B, Baskurt OK. Influence of cell-specific factors on red blood cell aggregation. Biorheology 2004, vol.41: 91-112.
10. Cicco G, Pirrelli A. Red blood cell (RBC) deformability, RBC aggregability and tissue oxygenation in hypertension. Clin Hemorheol Microcirc. 1999, vol.21: 169-177.
11. Berezina TL, Zaets SB, Morgan C, Spillert CR, Kamiyama M, Spolarics Z, Deitch EA, Machiedo GW. Influence of storage on red blood cell rheological properties. J Surg Res. 2002, vol.102: 6-12.
12. Hovav T, Yedgar S, Manny N, Barshtein G. Alteration of red cell aggregability and shape during blood storage. Transfusion 1999, vol.39: 277-281.
13. Relevy H, Koshkaryev A, Manny N, Yedgar S, Barshtein G. Blood banking-induced alteration of red blood cell flow properties. Transfusion 2008, vol.48: 136-146.
14. Izzo P, Manicone A, Spagnuolo A, Lauta VM, Di Pasquale A, Di Monte D. Erythrocytes stored in CPD SAG-mannitol: evaluation of their deformability. Clin Hemorheol Microcirc 1999, vol.21: 335-339.
15. Council of Europe. Guide to the preparation, use and quality assurance of blood components. 13th edition. 2007. Strasbourg, Council of Europe Publishing.
16. Bratosin D, Leszczynski S, Sartiaux C, Fontaine O, Descamps J, Huart JJ, Poplineau J, Goudaliez F, Aminoff D, Montreuil J. Improved storage of erythrocytes by prior leukodepletion: flow cytometric evaluation of stored erythrocytes. Cytometry 2001, vol.46: 351-356.
17. Shanwell A, Kristiansson M, Remberger M, Ringden O. Generation of cytokines in red cell concentrates during storage is prevented by prestorage white cell reduction. Transfusion 1997, vol.37: 678-684.
18. Gyongyossy-Issa MI, Weiss SL, Sowemimo-Coker SO, Garcez RB, Devine DV. Prestorage leukoreduction and low-temperature filtration reduce hemolysis of stored red cell concentrates. Transfusion 2005, vol.45: 90-96.
19. Racek J, Herynkova R, Holecek V, Faltysova J, Krejcova I. What is the source of free radicals causing hemolysis in stored blood? Physiol Res. 2001, vol.50: 383-388.
20. Baskurt OK, Meiselman HJ. Activated polymorphonuclear leukocytes affect red blood cell aggregability. J Leukoc Biol. 1998, vol.63: 89-93.
21. Baskurt OK, Temiz A, Meiselman HJ. Effect of superoxide anions on red blood cell rheologic properties. Free Radic Biol Med. 1998, vol.24: 102-110.
22. Hardeman MR, Ince C. Clinical potential of in vitro measured red cell deformability, a myth? Clin Hemorheol Microcirc. 1999, vol.21: 277-284.
23. Hardeman MR, Dobbe JG, Ince C. The Laser-assisted Optical Rotational Cell Analyzer (LORCA) as red blood cell aggregometer. Clin Hemorheol Microcirc. 2001, vol.25: 1-11.
24. Harboe M. A method for determination of hemoglobin in plasma by near-ultraviolet spectrophotometry. Scand J Clin Lab Invest. 1959, vol.11: 66-70.
25. Kakaiya R, Aronson CA, Julleis J. Whole blood collection and component processing, AABB Technical Manual 16th ed. Bethesda, MD.: AABB Press; 2008.

Chapter 4
54
26. Hill SR, Carless PA, Henry DA, Carson JL, Hebert PC, McClelland DB, Henderson KM. Transfusion thresholds and other strategies for guiding allogeneic red blood cell transfusion. Cochrane Database Syst Rev. 2002, vol.2: CD002042.
27. Kirkpatrick UJ, Adams RA, Lardi A, McCollum CN. Rheological properties and function of blood cells in stored bank blood and salvaged blood. Br J Haematol. 1998, vol.101: 364-368.
28. Zehnder L, Schulzki T, Goede JS, Hayes J, Reinhart WH. Erythrocyte storage in hypertonic (SAGM) or isotonic (PAGGSM) conservation medium: influence on cell properties. Vox Sang. 2008, vol.95: 280-287.
29. Singh A, Reinhart WH. The influence of fractions of abnormal erythrocytes on aggregation. Eur J Clin Invest. 1991, vol.21: 597-600.
30. Baskurt OK, Meiselman HJ. Hemodynamic effects of red blood cell aggregation. Indian J Exp Biol. 2007, vol.45: 25-31.
31. Mohandas N, Gallagher PG. Red cell membrane: past, present, and future. Blood 2008, vol.112: 3939-3948.
32. Condon MR, Kim JE, Deitch EA, Machiedo GW, Spolarics Z. Appearance of an erythrocyte population with decreased deformability and hemoglobin content following sepsis. Am J Physiol Heart Circ Physiol. 2003, vol.284: H2177-H2184.
33. Pries AR, Secomb TW. Microcirculatory network structures and models. Ann Biomed Eng. 2000, vol.28: 916-921.
34. Johnson RM. pH effects on red cell deformability. Blood Cells 1985, vol.11: 317-321.
35. Heaton WA. Evaluation of posttransfusion recovery and survival of transfused red cells. Transfus Med Rev 1992, vol.6: 153-169.
36. Hess JR. An update on solutions for red cell storage. Vox Sang. 2006, vol.91: 13-19.
37. Scott KL, Lecak J, Acker JP. Biopreservation of red blood cells: past, present, and future. Transfus Med Rev. 2005, vol.19: 127-142.
38. Verhoeven AJ, Hilarius PM, Dekkers DW, Lagerberg JW, de Korte D. Prolonged storage of red blood cells affects aminophospholipid translocase activity. Vox Sang. 2006, vol.91: 244-251.
39. Wan J, Ristenpart WD, Stone HA. Dynamics of shear-induced ATP release from red blood cells. Proc Natl Acad Sci. 2008, vol.105: 16432-16437.
40. Hogman CF, Meryman HT. Storage parameters affecting red blood cell survival and function after transfusion. Transfus Med Rev. 1999, vol.13: 275-296.
41. Van der Meer PF, Pietersz RN. The effect of plastic over wraps on storage measures of red cell concentrates. Vox Sang. 2007, vol.93: 176-178.
42. Stichting Sanquin bloedvoorziening. Richtlijn Bloedproducten. PT009.RL.SQ_002. 2005. Amsterdam.
43. Sollberger T, Walter R, Brand B, Contesse J, Meredith DO, Reinhart WH. Influence of prestorage leucocyte depletion and storage time on rheologic properties of erythrocyte concentrates. Vox Sang. 2002, vol.82: 191-197.
44. Van Bommel J, de Korte D, Lind A, Siegemund M, Trouwborst A, Verhoeven AJ, Ince C, Henny CP. The effect of the transfusion of stored RBCs on intestinal microvascular oxygenation in the rat. Transfusion 2001, vol.41: 1515-1523.

Rheologic properties of refrigerated stored RBCs
55
45. Nagaprasad V, Singh M. Sequential analysis of the influence of blood storage on aggregation, deformability and shape parameters of erythrocytes. Clin Hemorheol Microcirc. 1998, vol.18: 273-284.
46. Taleghani BM, Grossmann R, Waltenberger G, Geise W, Langer R, Henrich HA, Wiebecke D. Storage induced rheological and biochemical changes in erythrocyte concentrates with added solution and possible correlations. Beitr Infusionsther Transsfusionsmed. 1996, vol.33: 141-144.
47. Jensen LS, Kissmeyer-Nielsen P, Wolff B, Qvist N. Randomised comparison of leucocyte-depleted versus buffy-coat-poor blood transfusion and complications after colorectal surgery. Lancet 1996, vol.348: 841-845.
48. Bennett-Guerrero E, Veldman TH, Doctor A, Telen MJ, Ortel TL, Reid TS, Mulherin MA, Zhu H, Buck RD, Califf RM, McMahon, TJ. Evolution of adverse changes in stored RBCs. Proc Natl Acad Sci. 2007, vol.104: 17063-17068.
49. Raat NJ, Verhoeven AJ, Mik EG, Gouwerok CW, Verhaar R, Goedhart PT, de Korte D, Ince C. The effect of storage time of human red cells on intestinal microcirculatory oxygenation in a rat isovolemic exchange model. Crit Care Med. 2005, vol.33: 39-45.
50. Carvalho FA, Maria AV, Braz Nogueira JM, Guerra J, Martins-Silva J, Saldanha C. The relation between the erythrocyte nitric oxide and hemorheological parameters. Clin Hemorheol Microcirc. 2006, vol.35: 341-347.
51. Manodori AB, Barabino GA, Lubin BH, Kuypers FA. Adherence of phosphatidylserine-exposing erythrocytes to endothelial matrix thrombospondin. Blood 2000, vol.95: 1293-1300.
52. Schroit AJ, Madsen JW, Tanaka Y. In vivo recognition and clearance of red blood cells containing phosphatidylserine in their plasma membranes. J Biol Chem. 1985, vol.260: 5131-5138.
53. Cardo LJ, Hmel P, Wilder D. Stored packed red blood cells contain a procoagulant phospholipid reducible by leukodepletion filters and washing. Transfus Apher Sci. 2008, vol.38: 141-147.
54. Anniss AM, Sparrow RL. Storage duration and white blood cell content of red blood cell (RBC) products increases adhesion of stored RBCs to endothelium under flow conditions. Transfusion 2006, vol.46: 1561-1567.
55. Luk CS, Gray-Statchuk LA, Cepinkas G, Chin-Yee IH. WBC reduction reduces storage-associated RBC adhesion to human vascular endothelial cells under conditions of continuous flow in vitro. Transfusion 2003, vol.43: 151-156.
56. Raat NJ, Berends F, Verhoeven AJ, de Korte D, Ince C. The age of stored red blood cell concentrates at the time of transfusion. Transfus Med. 2005, vol.15: 419-423.


Chapter 5
Utilization of cryopreserved red blood cells in transfusion medicine
Sandra Henkelman Gerhard Rakhorst Reindert Graaff Johan W.M. Lagerberg
Submitted

Chapter 5
58
5.1. Introduction The ability to store RBCs outside the body has been regarded as a life-saving practice for many years.1 More recently, the usage of refrigerated stored RBCs in transfusion medicine has been under extensive evaluation. During refrigerated storage RBCs progressively deteriorates2 and infusion of prolonged stored RBCs has been linked to adverse clinical outcome in terms of postoperative infections, length of hospital stay and mortality.3-7 Although the majority of these studies are prone to selection bias due to a retrospective study design,8 concerns regarding the infusion of aged RBCs still remains and a restrictive transfusion strategy is currently being favored.9,10 The latter concerns have revived the interest in cryopreservation. Storage of RBCs at ultra-low temperatures halts the cellular metabolism and subsequently prevents the progressive cellular deterioration that has been linked to adverse clinical outcome. Initially, cryopreservation appeared a promising approach for maintaining RBCs viable for prolonged periods of time. Yet, the clinical applicability of cryopreserved RBCs (commonly known as frozen RBCs) was hampered by the expensive, time-consuming and less efficient nature of this preservation method.11,12 The subsequent unfamiliarity with regard to the quality of cryopreserved RBCs has also limited clinical usage. However, ongoing scientific and technological advancement has made cryopreserved RBCs more utilizable for clinical practice. This could be advantageous especially in situations where refrigerated stored RBCs are less desired. In this review the utilization of cryopreserved RBCs in modern transfusion practice will be discussed.
5.2. Cryopreservation methods Storage of RBCs at ultra-low temperatures ceases the biological activity of RBCs which enables them to be preserved for prolonged periods of time.13 In order to minimize freezing damage, cryoprotective additives are pivotal. In general, either high concentrations of cryoprotective additives or rapid freezing rates are necessary to prevent cell damage.14 Along the years different non-permeating and permeating additives for the cryopreservation of RBCs have been investigated. Non-permeating additives such as hydroxyethyl starch and polyvinylpyrrolidone, as well as a variety of glycols and sugars appeared promising because it was proposed that removal from thawed RBCs prior to transfusion was not

Usage of frozen RBCs
59
required.15-20 To date, the lack of data that support safe usage of non-permeating additives have prevented these cryoprotectants from being licensed.11,12 Conversely, the permeating additive glycerol is well known for its ability to protect RBCs at ultra-low temperatures. The concentration of glycerol that is necessary to protect the RBCs is dependent on the cooling rate and the storage temperature.14 At slow cooling rates, ice formation will occur extra-cellular. As ice forms, the solute content of the unfrozen fraction becomes more concentrated. The resulting osmotic imbalance causes fluid to move out of the RBC and intracellular dehydration occurs. However, at rapid cooling rates the RBC cytoplasm becomes super-cooled and intracellular ice formation occurs, which subsequently can lead to mechanical damage (Figure 5.1). Glycerol protects the RBCs by slowing the rate and extent of ice formation while minimizing cellular dehydration and solute effects during freezing.11
Figure 5.1. RBC changes in response to cooling rate. The snowflakes indicate the presence of ice crystals in the extra- and /or intracellular environment. Printed with permission of Scott et al.11

Chapter 5
60
To date, there are two freezing methods approved for the storage of RBCs.21,22 On the one hand, RBCs can be frozen rapidly (i.e. > 100°C/ min) by the low-glycerol method (LGM). With this method, RBCs are frozen with a final concentration of approximately 20% glycerol (wt/vol) at temperatures below -150°C. On the other hand, RBCs can be frozen slowly (i.e. ~1-3°C/ min) by the high-glycerol method (HGM). With this method, RBCs are frozen with a final concentration of approximately 40% (wt/vol) glycerol at temperatures between -65°C and -80°C. Overall, RBC preservation can be extended to at least ten years if the correct storage temperature is guaranteed. RBC units are preferentially thawed in a shaking water bath of about 36 to 42°C. The general consensus is that thawing should be done rapidly in order to prevent ice crystal growth (so called re-crystallization) upon warming.23 Once thawed, a deglycerolization washing procedure is performed to reduce the glycerol content in the RBC prior to infusion. This is necessary, since incomplete deglycerolized RBCs will swell and lyse upon infusion, resulting in hemolytic transfusion reactions and renal failure.24,25 The deglycerolization washing process causes to some extent osmotic stress to the RBC which results in cellular losses. Yet, the deglycerolization washing process is also advantageous in that it considerably reduces the amount of detrimental substances such as bioactive lipids, microparticles, cytokines, potassium and free Hb as well as leukocytes from the RBC unit.26-32 Hence, in the absence of leukofiltration, the deglycerolization process reduces the leukocyte count to a mean of 1 x 107 leukocytes per RBC storage unit.32,33 In addition, buffycoat depletion before cryopreservation further reduces the leukocytes count to a mean of 1-3 x 106 leukocytes per unit,32 which in some countries may even eliminate the need to leukofiltrate RBC units. It became also apparent that after post-thaw washing the immunogenicity of the residual leukocytes was reduced.34 So although frozen storage results in a RBC unit with a lower yield, the remaining RBCs contain less detrimental substances as compared to refrigerated stored RBCs.12 Altogether, transfusions of cryopreserved RBCs are associated with less febrile transfusion reactions, human leukocyte antigen alloimmunization as well as occurrences of TRALI and SIRS.12,35 If the glycerolization and deglycerolization of RBCs is performed in open systems, the post-thaw storage time is limited to 24-hours due to the potential risk of bacterial contamination. Yet, with the implementation of the ACP-215 device (Haemonetics, Braintree, MA), glycerol could be added and removed via an automated closed system, which minimized the risk of bacterial contamination.36 As a result, the post-thaw storage

Usage of frozen RBCs
61
time of RBCs could be extended to 7 days when stored in SAGM solution and to 14 days when stored in AS-3.31,37,38 Cryopreserved RBCs are less efficient due to the cellular losses that occur during the processing procedure. This cell loss was more pronounced in the HGM cryopreserved group (~ 10-20%) since these RBCs required more extensive washing. However, despite the higher yield of RBCs with the LGM method, it was recognized that HGM cryopreserved RBCs could tolerate wide fluctuations in temperature during freezing and were more stable during post-thaw storage.13,39 In addition, HGM cryopreserved RBCs did not require liquid nitrogen which eased storage and transportation conditions. Consequently, the HGM is currently the most applicable RBC freezing method in Europe and the United States.
5.3. Quality of cryopreserved RBCs Cryopreservation prolongs the longevity of RBCs. However, once thawed the shelf life of RBCs is limited. Cryopreserved RBCs have to meet certain guidelines (Table 5.1)21,22. Yet, these guidelines do not specifically reflect the ability of the RBCs to function after infusion. Cryopreservation subjects RBC to a range of chemical, thermal and mechanical forces, which might affect their oxygen delivering capacity after infusion. The quality of HGM cryopreserved RBCs is primarily dependent on the pre-freeze and post-thaw storage time, as well as on the anticoagulant and additive solution used.40 The duration of frozen storage per se minimally attributes to cellular damage.41,42 In order to limit storage induced lesions, refrigerated stored RBCs need to be frozen as soon as possible. According to the AABB, RBCs collected in CPDA-1 need to be frozen within 6 days, whereas in Europe the RBCs are preferably frozen within 7 days after collection.21,22 It is possible to freeze prolonged or outdated refrigerated stored RBCs, provided that the RBCs have been rejuvenated prior to freezing in order to restore the metabolic status of the cell (i.e. ATP, 2,3-DPG and Hb p50 levels).43-46 After deglycerolization even the outdated rejuvenated RBCs showed acceptable quality up to seven days of post-thaw storage in AS-3.36 In recent years more knowledge about the quality of HGM cryopreserved RBCs has become available. During frozen storage, the ATP and 2,3-DPG content is preserved. Yet, the length of pre-freezing storage time at 4°C was an important predictor of the ATP and

Chapter 5
62
2,3-DPG content after deglycerolization.36,42 The RBC ATP content is important for the overall functioning of the cell. Loss of ATP has been associated with rigid cell membranes, echinocyte shape change, enhanced cation permeability, exposure of phosphatidylserine on the RBC surface, microvesiculation, loss of vasodilatation control and decreased RBC viability.47-55 After deglycerolization the RBCs have a high ATP content when stored in AS-3 or SAGM additive solution. Nevertheless, during post-thaw storage the ATP content gradually declines. This decline was more prominent in AS-3 due to the diminished glycolytic activity induced by the lower pH of this storage solution.38 In the RBC binding of 2,3-DPG to the Hb induces a conformation state which will release oxygen from the Hb. In regions with low oxygen tension, oxygen is released from the Hb due to the high binding affinity of 2,3-DPG for deoxygenated Hb. Loss of 2,3-DPG will increase the affinity of oxygen to the Hb which may hamper the oxygen delivery to the tissues. Due to the low pH of the storage media a considerable loss of 2,3-DPG content was observed already after one week of refrigerated RBC storage.2 By limiting this pre-freezing storage time, higher 2,3-DPG values could be obtained post-thaw.56 Nevertheless, transfusing RBCs with low 2,3-DPG content appeared not to be detrimental to tissue oxygenation.57-59 Presumably, because the RBC 2,3-DPG content can be replenished hours following infusion,60-62 or because in hypoxic regions oxygen can still be released from the RBCs despite the low 2,3-DPG content. The ability of RBCs to adhere to the vascular endothelium is an important determinant of the flow behavior of blood and subsequently the oxygenation of the micro-vascular environment.63,64 Under physiological conditions, the adherence of RBCs to the vascular endothelium is negligible. Yet, structural changes in the RBC membrane may promote adherence to ECs and impair the microcirculatory blood flow. PS expression on the RBC surface mediates adherence of RBCs to ECs and might trigger RBC clearance from the circulation.65-67 In contrast, surface expression of CD-47, a marker of self, prevents RBCs from being engulfed by phagocytes.68 In general PS exposure and loss of CD-47 expression on the RBC surface as well as membrane microvesiculation are all apoptotic signals and therefore determining factors for the lifespan of the RBCs.69,70 HGM cryopreserved RBCs that were post-thaw stored in SAGM additive solution showed no significant changes in PS exposure, CD-47 expression and membrane microvesiculation when the pre-freezing storage time was limited to three days. However, surface expression of PS and microvesiculation was observed when longer pre-freezing storage times were used.56

Usage of frozen RBCs
63
Previously, it has been shown that the freeze-thaw process makes the RBC membrane permeable to cations, which result in RBC swelling and subsequently hemolysis.38,71,72 However, post-thaw storage of RBCs in AS-3 media, which contains the impermeable solute citrate, prevented cell swelling and limited the level of hemolysis during post-thaw storage.38 Altogether, it can be concluded that good post-thaw quality of HGM cryopreserved RBCs can be obtained when the appropriate storage conditions pre-freeze, frozen and post-thaw were used.
Table 5.1. Requirements of cryopreserved RBCs
Variable European Guidelines * AABB guidelines **
Hemolyse < 0.8 % < 1.0 %
Volume > 185 ml -
Hb content > 36 g/unit -
HCT 0.65-0.75 % -
Post-thaw recovery - ≥ 80 %
24-hour posttransfusion survival - ≥ 75 %
Leukocytes < 0.1 x109 cells/unit -
Osmolarity < 340 mOsm/l -
* European guidelines22 ** AABB guidelines21
5.4. Utilization of cryopreserved RBCs Currently, cryopreserved RBCs are primarily used for controlling an inventory in situations where the RBC availability is limited or unpredictable. Yet, cryopreserved RBCs are useful in a variety of clinical settings. The utilization of cryopreserved RBCs in transfusion medicine will be described in more detail below.

Chapter 5
64
Storage of rare RBCs Cryopreservation is currently used for preserving RBCs with rare blood phenotypes. In general, a blood group is regarded as rare if the RBC phenotype has a frequency of approximately 1 in 100-1000 or less in the general population.73 Refrigerated stored RBCs need to be as soon as possible, ideally within 6 to 7 days. However, cryopreservation of RBCs even beyond the regulated expiration date is still possible for exceedingly rare RBC phenotypes.11 Cryopreservation of rare RBCs is advantageous to patients for whom compatible blood is not instantly available. Over the years, numerous countries in Europe, America and Asia have set up frozen rare RBC banks.73-77 In Europe, most notably are the blood banks in Amsterdam, Birmingham and Paris, which house a large collection of cryopreserved rare RBC units. Most cryopreserved rare RBC units are for national use. This is because the RBC unit usually needs to be thawed in the donor blood center, imposing a time limit in which the RBCs must be transfused. Nevertheless, when no compatible blood can be found via the national blood banks, it is general practice to appeal to countries abroad. Although usage of cryopreserved rare RBCs is extremely costly and the international shipment is usually cumbersome, it has been lifesaving for a variety of patients.
Military blood bank In military combat massive blood loss is a major cause of death.78,79 Having RBCs available in the military theater at all times is therefore of vital importance. Previously, an inventory of RBCs was difficult to maintain in combat areas due to the unpredictable demand and the limited shelf life of refrigerated stored RBCs. Yet, cryopreserved RBCs are a valuable blood resource due to the prolonged storage time. Cryopreserved RBCs have been used in the military theater since the Vietnam war.80,81 Although back then the processing of cryopreserved RBCs was still in its early stages, it was already concluded that usage of cryopreserved RBCs in combat casualty care was technically feasible and clinically acceptable. Ongoing scientific and technological advancement have made cryopreserved RBCs become a more utilized blood product in modern military operations.13,82-84 Especially the Dutch army has been using cryopreserved RBCs in theater operations with great success.83,85,86 This was also demonstrated at the conflicts in Afghanistan, where already 1360 cryopreserved RBC units have been transfused by the Dutch military blood

Usage of frozen RBCs
65
bank without any shortages or transfusion reactions reported.87 Hence, the Dutch military blood bank only uses type O leukodepleted RBCs in the theater in order to improve the effectiveness and decrease the chance of clerical errors. In this regard, leukodepleted type O whole blood is processed into RBCs and frozen within 24 hours after donation. Additionally, the short pre-storage time reduces storage induced lesions to a minimum. Cryopreserved RBC units are transported to combat areas on dry ice. These RBCs have been frozen in polyvinyl chloride plastic (PVC) bags inside polyester plastic bags which were placed in rigid cardboard boxes. In the past, PVC bags that were stored at -80°C and subjected to transportation had a breakage incidence of approximately 6.7%.88 Due to the use of an additional vacuum sealed over-wrap bag, the breakage incidence of cryopreserved RBC units subjected to transportation is currently negligible.83 Subsequently, all in theater storage, thawing and washing procedures are performed in a temperature controlled blood bank container, designed by the Dutch army. Thawed and washed RBCs are ultimately preserved in AS-3 and can be refrigerated stored at 2-6 °C for a maximum of 14 days. Usage of cryopreserved RBCs in combat casualty care offers a better inventory control. This is because cryopreserved RBC units can be prepared either on demand or in advance, thereby providing a continuous RBC supply even when standard refrigerated stored RBCs cannot be replenished on time. Furthermore, cryopreserved RBCs may be advantageous to prolonged refrigerated stored RBCs. In combat hospitals, it is not uncommon to receive refrigerated stored RBC units with a mean storage age of 27 days on arrival.89,90 Compared to prolonged refrigerated stored RBCs, cryopreserved RBCs have less detrimental substances that can cause transfusion reactions. Nevertheless, randomized-controlled trials comparing transfusion of cryopreserved RBCs with refrigerated stored RBCs have still to be performed. Cryopreserved RBCs are undeniable more costly. Hence, a unit of cryopreserved RBCs cost approximately twice the amount of a refrigerated stored RBC unit. In addition, thawing and washing of cryopreserved RBC units requires skilled personnel and takes up about 70-120 minutes.12,82,83 Yet despite these disadvantages, stockpiling a frozen RBC inventory proved to be an efficient and safe blood resource in combat casualty care.

Chapter 5
66
Blood shortages Blood shortages due to natural or civil disasters as well as due to seasonal shortages can pose a major health challenge. However, in emergency blood management planning cryopreserved RBCs are rarely implemented. This is primarily due to the longer processing time of cryopreserved RBCs, making it difficult to prepare large quantities of RBC units within a given time frame. Cryopreserved RBCs are also less implemented during times of blood shortages, due to the improved emergency procedures of blood centers with regard to refrigerated stored RBCs.91-94 A main strategy of the emergency procedures is to mobilize stocks of refrigerated stored RBC through coordination with nearby blood centers. Hence, blood centers will usually have a 2-3 day supply of refrigerated stored RBC units on hand. This strategy could compensate for RBC shortages as long as the local stocks of the nearby blood centers are repleted appropriately. After a disaster the influx of blood donations is often increased because of the altruistic response of the public. Although this influx of blood donations could be used to replenish local stocks, this is true for only part of the donated blood. Notably, part of the donated blood is non-transfusible due to the higher reactive screening tests and sometimes inadequately processing procedures.95-99 In this regard, blood centers that do send refrigerated stored RBC units could face the risk of becoming under-supplied themselves. Recently, it was demonstrated that managing a frozen RBC reserve could be useful in emergencies scenarios.100 In this regard, cryopreserved RBCs would serve as a bridge-over supply during short term RBC deficits until support by the blood centers could be re-established. Despite the complexity and costs of implementing and maintaining a frozen RBC reserve, it was concluded that the benefits of self-sufficiency outweighs the disadvantages.
Autologous transfusion Cryopreserved RBCs have occasionally been used for preoperative autologous storage.101 In general, preoperative autologous RBC transfusion offers advantage above allogeneic RBC transfusions in that it prevents immunosuppression and infectious disease transmission, while it reduces postoperative infections and subsequently length of hospital stay.102,103 During the last couple of years preoperative autologous RBCs usage has been

Usage of frozen RBCs
67
questioned and its demand has declined. This is predominantly due to the improved safety of allogeneic RBCs as well as due to the organizational and logistic hurdles, the higher disposal rate and the more costly nature associated with autologous RBCs transfusions.103-
107 Additionally, the beneficial effect of preoperative autologous RBC donation has been compromised by the short time period between the last donation and the planned surgical procedure. As a result, patients often develop anemia before the surgery and are more likely to receive transfusion.105,106,108 In order to avoid anemia, RBCs need to be harvested months in advance of the expected use so that the Hb level of the patient can be restored. Yet, prolongation of the time period between the last donation and the surgical procedure is hampered by the short storage time of refrigerated stored RBCs. In contrast, cryopreservation enables storage of RBCs for years, which allow RBCs to be donated far in advance of the surgical procedure109 without affecting its quality. Usage of cryopreserved RBCs has an additional advantage in that less detrimental substances, that have accumulate during storage and which may affect transfusion outcome, are transfused. Although the transmission of infectious diseases is currently very low and the benefit of preoperative autologous RBC transfusions is questioned, new pathogens keep emerging110-
113 and with the implementation of cryopreservation, preoperative autologous RBC transfusions could become more attractive again.
Usage when refrigerated stored RBCs are less desired Cryopreserved RBCs could be advantageous in situations where refrigerated stored RBCs are less desired. This could be the case for patients who receive transfusions frequently, such as patients with thalassemia, sickle-cell disease or with certain types of glucose-6-phosphate dehydrogenase deficiency. This is because frequent transfusions with refrigerated stored RBCs are a risk of human leukocyte antigen alloimmunization. Hence, the development of antibodies against infused RBC surface antigens is of considerable medical importance because it will result in hemolytic transfusion reactions.114,115 In contrast, usage of cryopreserved RBCs reduces the risk of alloimmunization due to a reduction in the number of leukocytes by the deglycerolization washing procedure.12,13 Cryopreserved RBCs may also be helpful as part of the routine RBC inventory because of its advantages over prolonged refrigerated stored RBCs. As mentioned earlier, transfusion

Chapter 5
68
of prolonged refrigerated stored RBCs is associated with occurrences of TRALI and SIRS.12,35 Although the etiology of TRALI and SIRS remains incompletely understood, it is recognized that substances that accumulate in the supernatant of refrigerated stored RBCs are involved in the pathogenesis of these syndromes.116-120 Ultimately, these syndromes will lead to an increased hospitalization and subsequently higher burden on the healthcare costs. Cryopreserved RBCs on the other hand have less than 5% cytokines and biologically active substances in the supernatant, which make them the ideal blood product to prevent TRALI and SIRS.35,121 In line with this, cryopreserved RBCs are also useful for patients with immunoglobulin A (IgA) deficiency. These patients usually have undetectable IgA and high titer of class specific anti-IgA. Transfusion of only a small amount of blood can cause severe anaphylactic reactions due to the presence of IgA in plasma.40 Since cryopreserved RBCs are extensively washed, these blood products are especially recommended for patients with IgA deficiency.13,40,122
5.5. Conclusion In transfusion medicine the balance between the RBC availability and demand is variable. However due to the perishable nature of refrigerated stored RBCs, hospitals often maintain only a minimal RBC reserve to maximize the efficiency while minimizing the cost of wastage. This policy also means that hospitals will be more vulnerable to RBC shortages due to fluctuations in the RBC availability and or demand. Cryopreserved RBCs may be helpful as part of the routine refrigerated stored RBC inventory. Hence, having cryopreserved RBCs available in civilian blood banks and or hospitals could result in better blood management and patient care need. Today, cryopreserved RBCs are still infrequently implemented in transfusion medicine. This is mainly because of the expensive nature of this preservation method. Although, the higher costs of cryopreserved RBCs are of major concern, the cost difference with regard to refrigerated stored RBCs is often overrated. This is because the costs of treating and managing adverse events are not taken into account, indicating that the total cost of a refrigerated stored RBC unit would be substantial higher than currently is represented.123 Cryopreserved RBCs are also less efficient and more time-consuming, however ongoing scientific and technological advancement has made cryopreserved RBCs more utilizable for

Usage of frozen RBCs
69
clinical practice. Notably, more knowledge about the quality of cryopreserved RBCs could further expand its use in clinical practice. In the foreseeable future the overall use of cryopreserved RBCs could expand as a result of a change in RBC supply and demand, due to a shift of increasingly older patient population.124,125 For now, we showed that cryopreservation of RBCs is already useful in a variety of clinical settings. Especially, since cryopreserved RBCs are available, save, in compliance with European and US guidelines and can be used effectively.
References 1. Giangrande PL. The history of blood transfusion. Br.J.Haematol. 2000, vol.110: 758-767.
2. Bennett-Guerrero E, Veldman TH, Doctor A, Telen MJ, Ortel TL, Reid TS, Mulherin MA, Zhu H, Buck RD, Califf RM, et al. Evolution of adverse changes in stored RBCs. Proc.Natl.Acad.Sci. 2007, vol.104: 17063-17068.
3. Lelubre C, Piagnerelli M, Vincent JL. Association between duration of storage of transfused red blood cells and morbidity and mortality in adult patients: myth or reality? Transfusion. 2009, vol.49: 1384-1394.
4. Van de Watering LM, Brand A. Effect of storage of red cells. Transfusion medicine and hemotherapy. 2008;35: 359-367.
5. Zimrin AB, Hess JR. Current issues relating to the transfusion of stored red blood cells. Vox Sang. 2009, vol.96: 93-103.
6. Edgren G, Kamper-Jorgensen M, Eloranta S, Rostgaard K, Custer B, Ullum H, Murphy EL, Busch MP, Reilly M, Melbye M, et al. Duration of red blood cell storage and survival of transfused patients (CME). Transfusion. 2010, vol.50: 1185-1195.
7. Vamvakas EC. Meta-analysis of clinical studies of the purported deleterious effects of "old" (versus "fresh") red blood cells: are we at equipoise? Transfusion. 2010, vol.50: 600-10.
8. Van de Watering LM. Pitfalls in the current published observational literature on the effects of red blood cell storage. Transfusion. 2011, vol.51: 1847-1854.
9. Corwin HL, Carson JL. Blood transfusion, when is more really less? N.Engl.J.Med. 2007, vol.356: 1667-1669.
10. Varghese R, Myers ML. Blood conservation in cardiac surgery: let's get restrictive. Semin.Thorac.Cardiovasc.Surg. 2010, vol.22: 121-126.
11. Scott KL, Lecak J, Acker JP. Biopreservation of red blood cells: past, present, and future. Transfus.Med.Rev. 2005, vol.19: 127-142.
12. Hess JR. Red cell freezing and its impact on the supply chain. Transfus.Med. 2004, vol.14: 1-8.
13. Valeri CR, Ragno G. Cryopreservation of human blood products. Transfus.Apher.Sci. 2006, vol.34: 271-287.

Chapter 5
70
14. Rowe AW. Cryopreservation of red cells by freezing and vitrification - some recollections and predictions. Infusion therapy transfusion medicine. 2002, vol.29: 25-30.
15. Quan G, Zhang L, Guo Y, Liu M, Wang J, Wang Y, Dong B, Liu A, Zhang J, Han Y. Intracellular sugars improve survival of human red blood cells cryopreserved at -80 °C in the presence of polyvinyl pyrrolidone and human serum albumin. Cryo.Letters. 2007, vol.28: 95-108.
16. Boutron P, Arnaud F. Comparison of the cryoprotection of red blood cells by 1,2-propanediol and glycerol. Cryobiology. 1984, vol.21: 348-358.
17. Mehl P, Boutron P. Cryoprotection of red blood cells by 1,3-butanediol and 2,3-butanediol. Cryobiology. 1988, vol.25: 44-54.
18. Quan GB, Han Y, Liu MX, Gao F. Effects of pre-freeze incubation of human red blood cells with various sugars on postthaw recovery when using a dextran-rapid cooling protocol. Cryobiology. 2009, vol.59: 258-267.
19. Pellerin-Mendes C, Million L, Marchand-Arvier M, Labrude P, Vigneron C. In vitro study of the protective effect of trehalose and dextran during freezing of human red blood cells in liquid nitrogen. Cryobiology. 1997, vol.35: 173-186.
20. Horn EP, Sputtek A, Standl T, Rudolf B, Kuhnl P, Schulte Am EJ. Transfusion of autologous, hydroxyethyl starch-cryopreserved red blood cells. Anesth.Analg. 1997, vol.85: 739-745.
21. Kakaiya R, Aronson CA, Julleis J. Whole blood collection and component processing, AABB Technical Manual 16th ed. Bethesda. AABB Press; 2008.
22. Council of Europe. Guide to the preparation, use and quality assurance of blood components, 14th edition. Strasbourg: Council of Europe publisher; 2008.
23. Pegg DE. The relevance of ice crystal formation for the cryopreservation of tissues and organs. Cryobiology. 2010, vol.60: S36-S44.
24. Cregan P, Donegan E, Gotelli G. Hemolytic transfusion reaction following transfusion of frozen and washed autologous red cells. Transfusion. 1991, vol.31: 172-175.
25. Bechdolt S, Schroeder LK, Samia C, Schmidt PJ. In vivo hemolysis of deglycerolized red blood cells. Arch.Pathol.Lab Med. 1986, vol.110: 344-345.
26. Vlaar AP, Kulik W, Nieuwland R, Peters CP, Tool AT, van Bruggen R, Juffermans NP, de Korte D. Accumulation of bioactive lipids during storage of blood products is not cell but plasma derived and temperature dependent. Transfusion. 2011, vol.51: 2358-2366.
27. Silliman CC, Moore EE, Kelher MR, Khan SY, Gellar L, Elzi DJ. Identification of lipids that accumulate during the routine storage of prestorage leukoreduced red blood cells and cause acute lung injury. Transfusion. 2011, vol.51: 2549-2554.
28. Kim-Shapiro DB, Lee J, Gladwin MT. Storage lesion: role of red blood cell breakdown. Transfusion. 2011, vol.51: 844-851.
29. Shanwell A, Kristiansson M, Remberger M, Ringden O. Generation of cytokines in red cell concentrates during storage is prevented by prestorage white cell reduction. Transfusion. 1997, vol.37: 678-684.

Usage of frozen RBCs
71
30. Glenister KM, Sparrow RL. Level of platelet-derived cytokines in leukoreduced red blood cells is influenced by the processing method and type of leukoreduction filter. Transfusion. 2010, vol.50: 185-189.
31. Bandarenko N, Hay SN, Holmberg J, Whitley P, Taylor HL, Moroff G, Rose L, Kowalsky R, Brumit M, Rose M, et al. Extended storage of AS-1 and AS-3 leukoreduced red blood cells for 15 days after deglycerolization and resuspension in AS-3 using an automated closed system. Transfusion. 2004, vol.44: 1656-1662.
32. Arnaud FG, Meryman HT. WBC reduction in cryopreserved RBC units. Transfusion. 2003, vol.43: 517-525.
33. Valeri CR, Ragno G, Pivacek LE, Srey R, Hess JR, Lippert LE, Mettille F, Fahie R, O'Neill EM, Szymanski IO. A multicenter study of in vitro and in vivo values in human RBCs frozen with 40-percent (wt/vol) glycerol and stored after deglycerolization for 15 days at 4°C in AS-3: assessment of RBC processing in the ACP 215. Transfusion. 2001, vol.41: 933-939.
34. Farrugia A, Shea N, Knowles S, Holdsworth R, Piouronowski H, Portbury D, Romeo A. Cryopreservation of red blood cells: effect of freezing on red cell quality and residual lymphocyte immunogenicity. J.Clin.Pathol. 1993, vol.46: 742-745.
35. Valeri CR, Ragno G. An approach to prevent the severe adverse events associated with transfusion of FDA-approved blood products. Transfus.Apher.Sci. 2010, vol.42: 223-233.
36. Valeri CR, Ragno G, Van HP, Rose L, Rose M, Egozy Y, Popovsky MA. Automation of the glycerolization of red blood cells with the high-separation bowl in the Haemonetics ACP 215 instrument. Transfusion. 2005, vol.45: 1621-1627.
37. Bohonek M, Petras M, Turek I, Urbanova J, Hradek T, Chmatal P, Staroprazska V, Kostirova J, Horcickova D, Duchkova S, et al. Quality evaluation of frozen apheresis red blood cell storage with 21-day postthaw storage in additive solution 3 and saline-adenine-glucose-mannitol: biochemical and chromium-51 recovery measures. Transfusion. 2010, vol.50: 1007-1013.
38. Lagerberg JW, Truijens-de Lange R, de Korte D, Verhoeven AJ. Altered processing of thawed red cells to improve the in vitro quality during postthaw storage at 4°C. Transfusion. 2007, vol.47: 2242-2249.
39. Lelkens CC, Noorman F, Koning JG, Truijens de Lange R, Stekkinger PS, Bakker JC, Lagerberg JW, Brand A, Verhoeven AJ. Stability after thawing of RBCs frozen with the high- and low-glycerol method. Transfusion. 2003, vol.43: 157-164.
40. Klein HG, Anstee D. Mollison's Blood Transfusion in Clinical Medicine, 11th ed. Massachusetts: Blackwell Publishing; 2005.
41. Valeri CR, Ragno G, Pivacek LE, Cassidy GP, Srey R, Hansson-Wicher M, Leavy ME. An experiment with glycerol-frozen red blood cells stored at -80°C for up to 37 years. Vox Sang. 2000, vol.79: 168-174.
42. Lecak J, Scott K, Young C, Hannon J, Acker JP. Evaluation of red blood cells stored at -80°C in excess of 10 years. Transfusion. 2004, vol.44: 1306-1313.
43. Lockwood WB, Hudgens RW, Szymanski IO, Teno RA, Gray AD. Effects of rejuvenation and frozen storage on 42-day-old AS-3 RBCs. Transfusion. 2003, vol.43: 1527-1532.
44. Szymanski IO, Teno RA, Lockwood WB, Hudgens R, Johnson GS. Effect of rejuvenation and frozen storage on 42-day-old AS-1 RBCs. Transfusion. 2001, vol.41: 550-555.

Chapter 5
72
45. Ragno G, Robert VC. Salvaging of liquid-preserved O-positive and O-negative red blood cells by rejuvenation and freezing. Transfus.Apher.Sci. 2006, vol.35: 137-143.
46. Valeri CR, Pivacek LE, Cassidy GP, Ragno G. The survival, function, and hemolysis of human RBCs stored at 4°C in additive solution (AS-1, AS-3, or AS-5) for 42 days and then biochemically modified, frozen, thawed, washed, and stored at 4 °C in sodium chloride and glucose solution for 24 hours. Transfusion. 2000, vol.40:1341-1345.
47. Betz T, Lenz M, Joanny JF, Sykes C. ATP-dependent mechanics of red blood cells. Proc.Natl.Acad.Sci. 2009, vol.106:15320-15325.
48. Bucki R, Bachelot-Loza C, Zachowski A, Giraud F, Sulpice JC. Calcium induces phospholipid redistribution and microvesicle release in human erythrocyte membranes by independent pathways. Biochemistry. 1998, vol.37: 15383-15391.
49. Gov NS, Safran SA. Red blood cell membrane fluctuations and shape controlled by ATP-induced cytoskeletal defects. Biophys.J. 2005, vol.88: 1859-1874.
50. Gov NS. Active elastic network: cytoskeleton of the red blood cell. Phys.Rev.E.Stat.Nonlin.Soft.Matter Phys. 2007, vol.75: 011921.
51. Heaton WA. Evaluation of posttransfusion recovery and survival of transfused red cells. Transfus.Med.Rev. 1992, vol.6: 153-169.
52. Johnson RM. Membrane stress increases cation permeability in red cells. Biophys.J. 1994, vol.67: 1876-1881.
53. Park Y, Best CA, Auth T, Gov NS, Safran SA, Popescu G, Suresh S, Feld MS. Metabolic remodeling of the human red blood cell membrane. Proc.Natl.Acad.Sci. 2010, vol.107: 1289-1294.
54. Verhoeven AJ, Hilarius PM, Dekkers DW, Lagerberg JW, de Korte D. Prolonged storage of red blood cells affects aminophospholipid translocase activity. Vox Sang. 2006, vol.91: 244-251.
55. Wan J, Ristenpart WD, Stone HA. Dynamics of shear-induced ATP release from red blood cells. Proc.Natl.Acad.Sci. 2008, vol.105:16432-16437.
56. Holovati JL, Wong KA, Webster JM, Acker JP. The effects of cryopreservation on red blood cell microvesiculation, phosphatidylserine externalization, and CD47 expression. Transfusion. 2008, vol.48: 1658-1668.
57. Raat NJ, Verhoeven AJ, Mik EG, Gouwerok CW, Verhaar R, Goedhart PT, de Korte D, Ince C. The effect of storage time of human red cells on intestinal microcirculatory oxygenation in a rat isovolemic exchange model. Crit Care Med. 2005, vol.33: 39-45.
58. Ibrahim ED, McLellan SA, Walsh TS. Red blood cell 2,3-diphosphoglycerate concentration and in vivo P50 during early critical illness. Crit Care Med. 2005, vol.33: 2247-2252.
59. D'Almeida MS, Gray D, Martin C, Ellis CG, Chin-Yee IH. Effect of prophylactic transfusion of stored RBCs on oxygen reserve in response to acute isovolemic hemorrhage in a rodent model. Transfusion. 2001, vol.41: 950-956.
60. Heaton A, Keegan T, Holme S. In vivo regeneration of red cell 2,3-diphosphoglycerate following transfusion of DPG-depleted AS-1, AS-3 and CPDA-1 red cells. Br.J.Haematol. 1989, vol.71: 131-136.

Usage of frozen RBCs
73
61. Valeri CR, Collins FB. The physiologic effect of transfusing preserved red cells with low 2,3-diphosphoglycerate and high affinity for oxygen. Vox Sang. 1971, vol.20: 397-403.
62. Matthes G, Strunk S, Siems W, Grune T. Posttransfusional changes of 2,3-diphosphoglycerate and nucleotides in CPD-SAGM-preserved erythrocytes. Infusionsther.Transfusionsmed. 1993, vol.20: 89-92.
63. Yedgar S, Koshkaryev A, Barshtein G. The red blood cell in vascular occlusion. Pathophysiol.Haemost.Thromb. 2002, vol.32: 263-268.
64. Kaul DK, Koshkaryev A, Artmann G, Barshtein G, Yedgar S. Additive effect of red blood cell rigidity and adherence to endothelial cells in inducing vascular resistance. Am.J.Physiol Heart Circ.Physiol. 2008, vol.295:H1788-H1793.
65. Closse C, Dachary-Prigent J, Boisseau MR. Phosphatidylserine-related adhesion of human erythrocytes to vascular endothelium. Br.J.Haematol. 1999, vol.107: 300-302.
66. Manodori AB, Barabino GA, Lubin BH, Kuypers FA. Adherence of phosphatidylserine-exposing erythrocytes to endothelial matrix thrombospondin. Blood. 2000, vol.95: 1293-1300.
67. Wu Y, Tibrewal N, Birge RB. Phosphatidylserine recognition by phagocytes: a view to a kill. Trends Cell Biol. 2006, vol.16: 189-197.
68. Oldenborg PA, Zheleznyak A, Fang YF, Lagenaur CF, Gresham HD, Lindberg FP. Role of CD47 as a marker of self on red blood cells. Science. 2000, vol.288:2051-2054.
69. Lang F, Gulbins E, Lerche H, Huber SM, Kempe DS, Foller M. Eryptosis, a window to systemic disease. Cell Physiol Biochem. 2008, vol.22: 373-380.
70. Gardai SJ, Bratton DL, Ogden CA, Henson PM. Recognition ligands on apoptotic cells: a perspective. J.Leukoc.Biol. 2006, vol.79: 896-903.
71. Hess JR, Hill HR, Oliver CK, Lippert LE, Greenwalt TJ. The effect of two additive solutions on the postthaw storage of RBCs. Transfusion. 2001, vol.41: 923-927.
72. Valeri CR, Srey R, Tilahun D, Ragno G. The in vitro quality of red blood cells frozen with 40 percent (wt/vol) glycerol at -80°C for 14 years, deglycerolized with the Haemonetics ACP 215, and stored at 4°C in additive solution-1 or additive solution-3 for up to 3 weeks. Transfusion. 2004, vol.44: 990-995.
73. Reesink HW, Engelfriet CP, Schennach H, Gassner C, Wendel S, Fontao-Wendel R, de Brito MA, Sistonen P, Matilainen J, Peyrard T, et al. Donors with a rare pheno (geno) type. Vox Sang. 2008, vol.95: 236-253.
74. Woodfield G, Poole J, Nance ST, Daniels G. A review of the ISBT rare blood donor program. Immunohematology. 2004, vol.20: 244-248.
75. Peyrard T, Pham BN, Le Pennec PY, Rouger P. Transfusion of rare cryopreserved red blood cell units stored at -80°C: the French experience. Immunohematology. 2009, vol.25: 13-17.
76. Rijnen A. Zeldzaam bloed voor als het erop aankomt. Bloedbeeld 2004, vol.1: 18-19.
77. Morelati F, Arnaboldi P, Barocci F, Bodini U, Boiani E, Bresciani S, Cambie G, Cazzaniga G, Cocco E, Copeta A, et al. Strategies for the transfusion of subjects with complex red cell immunisation: the Bank of rare blood donors of the Region of Lombardy. Blood Transfus. 2007, vol.5: 217-226.

Chapter 5
74
78. Kelly JF, Ritenour AE, McLaughlin DF, Bagg KA, Apodaca AN, Mallak CT, Pearse L, Lawnick MM, Champion HR, Wade CE, et al. Injury severity and causes of death from Operation Iraqi Freedom and Operation Enduring Freedom: 2003-2004 versus 2006. J.Trauma. 2008, vol.64: S21-S26.
79. Martin M, Oh J, Currier H, Tai N, Beekley A, Eckert M, Holcomb J. An analysis of in-hospital deaths at a modern combat support hospital. J.Trauma. 2009, vol.66: S51-S60.
80. Moss GS, Valeri R, Brodine CE. Clinical experience with the use of frozen blood in combat casulties. The New England Journal of Medicine. 1968, vol.278: 748-752.
81. Hess JR, Thomas MJ. Blood use in war and disaster: lessons from the past century. Transfusion. 2003, vol.43: 1622-1633.
82. Neuhaus SJ, Wishaw K, Lelkens C. Australian experience with frozen blood products on military operations. Med.J.Aust. 2010, vol.192: 203-205.
83. Lelkens CC, Koning JG, de Korte D, Floot IB, Noorman F. Experiences with frozen blood products in the Netherlands military. Transfus.Apher.Sci. 2006, vol.34: 289-298.
84. Rentas FJ. Cryopreserved red blood cells: summary. J.Trauma. 2011, vol.70: S45-S46.
85. Holovati JL, Hannon JL, Gyongyossy-Issa MI, Acker JP. Blood preservation workshop: new and emerging trends in research and clinical practice. Transfus.Med.Rev. 2009, vol.23: 25-41.
86. Noorman F, Lelkens CCM. Hemovigilantie onder militaire operationele omstandigheden in Afghanistan. Tijdschrift voor bloedtransfusie. 2011, vol.1: 37-41.
87. Noorman F, Strelitski R, Lelkens CCM. Frozen -80°C red cells, plasma and platelets in combat casualty care. Transfusion. 2009, vol.49: 26A.
88. Valeri CR, Lane JP, Srey R, Ragno G. Incidence of breakage of human RBCs frozen with 40-percent wt/vol glycerol using two different methods for storage at -80°C. Transfusion. 2003, vol.43: 411-414.
89. Spinella PC, Perkins JG, Grathwohl KW, Repine T, Beekley AC, Sebesta J, Jenkins D, Azarow K, Holcomb JB. Risks associated with fresh whole blood and red blood cell transfusions in a combat support hospital. Crit Care Med. 2007, vol.35: 2576-2581.
90. Hess JR, Holcomb JB. Transfusion practice in military trauma. Transfus.Med. 2008, vol.18: 143-150.
91. Ramsey G. Frozen red blood cells: cold comfort in a disaster? Transfusion. 2008, vol.48: 2053-2055.
92. Joseph BG, Hendry C, Walsh TS. Red blood cell use outside the operating theater: a prospective observational study with modeling of potential blood conservation during severe blood shortages. Transfusion. 2009, vol.49: 2060-2069.
93. Kuruppu KK. Management of blood system in disasters. Biologicals. 2010, vol. 38: 87-90.
94. Zimrin AB, Hess JR. Planning for pandemic influenza: effect of a pandemic on the supply and demand for blood products in the United States. Transfusion. 2007, vol.47: 1071-1079.
95. Dodd RY, Orton SL, Notari EP, Stramer SL. Viral marker rates among blood donors before and after the terrorist attacks on the United States on September 11, 2001. Transfusion. 2002, vol.42: 1240-1241.
96. Linden JV, Davey RJ, Burch JW. The September 11, 2001 disaster and the New York blood supply. Transfusion. 2002, vol.42:1385-1387.

Usage of frozen RBCs
75
97. Glynn SA, Busch MP, Schreiber GB, Murphy EL, Wright DJ, Tu Y, Kleinman SH. Effect of a national disaster on blood supply and safety: the September 11 experience. JAMA. 2003, vol.289: 2246-2253.
98. Schmidt PJ. Blood and disaster-supply and demand. N.Engl.J.Med. 2002, vol.346: 617-620.
99. Mujeeb SA, Jaffery SH. Emergency blood transfusion services after the 2005 earthquake in Pakistan. Emerg.Med.J. 2007, vol.24: 22-24.
100. Erickson ML, Champion MH, Klein R, Ross RL, Neal ZM, Snyder EL. Management of blood shortages in a tertiary care academic medical center: the Yale-New Haven Hospital frozen blood reserve. Transfusion. 2008, vol.48: 2252-2263.
101. Fontaine MJ, Winters JL, Moore SB, McGregor CG, Santrach PJ. Frozen preoperative autologous blood donation for heart transplantation at the Mayo Clinic from 1988 to 1999. Transfusion. 2003, vol.43: 476-480.
102. Innerhofer P, Klingler A, Klimmer C, Fries D, Nussbaumer W. Risk for postoperative infection after transfusion of white blood cell-filtered allogeneic or autologous blood components in orthopedic patients undergoing primary arthroplasty. Transfusion. 2005, vol.45: 103-110.
103. Kretschmer V. Preoperative autologous blood donation, a confessor's' point of view. Transfus.Med.Hemother. 2006, vol.33: 330-335.
104. Alter HJ, Klein HG. The hazards of blood transfusion in historical perspective. Blood. 2008, vol.112: 2617-2626.
105. Brecher ME, Goodnough LT. The rise and fall of preoperative autologous blood donation. Transfusion. 2001, vol.41: 1459-1462.
106. Goodnough LT, Brecher ME, Kanter MH, AuBuchon JP. Transfusion medicine. Second of two parts--blood conservation. N.Engl.J.Med. 1999, vol.340: 525-533.
107. Muller MM, Seifried E. Do we still need preoperative autologous blood donation? It is time for a reappraisal. Transfus.Med.Hemother. 2006, vol.33: 336-347.
108. Singbartl G, Schreiber J, Singbartl K. Preoperative autologous blood donation versus intraoperative blood salvage: intraindividual analyses and modeling of efficacy in 1103 patients. Transfusion. 2009, vol.49: 2374-2383.
109. Ashenden M, Morkeberg J. Net haemoglobin increase from reinfusion of refrigerated vs. frozen red blood cells after autologous blood transfusions. Vox Sang. 2011, vol.101: 320-326.
110. Zou S, Stramer SL, Dodd RY. Donor Testing and Risk: Current Prevalence, Incidence, and Residual Risk of Transfusion-Transmissible Agents in US Allogeneic Donations. Transfus.Med.Rev. 2011, in press.
111. Blajchman MA, Vamvakas EC. The continuing risk of transfusion-transmitted infections. N.Engl.J.Med. 2006, vol.355: 1303-1305.
112. Stramer SL, Hollinger FB, Katz LM, Kleinman S, Metzel PS, Gregory KR, Dodd RY. Emerging infectious disease agents and their potential threat to transfusion safety. Transfusion. 2009, vol.49: 1S-29S.
113. Atreya C, Nakhasi H, Mied P, Epstein J, Hughes J, Gwinn M, Kleinman S, Dodd R, Stramer S, Walderhaug M, et al. FDA workshop on emerging infectious diseases: evaluating emerging infectious diseases (EIDs) for transfusion safety. Transfusion. 2011, vol.51: 1855-1871.

Chapter 5
76
114. Gupta R, Singh DK, Singh B, Rusia U. Alloimmunization to red cells in thalassemics: emerging problem and future strategies. Transfus.Apher.Sci. 2011, , vol.45: 167-170.
115. Zimring JC, Welniak L, Semple JW, Ness PM, Slichter SJ, Spitalnik SL. Current problems and future directions of transfusion-induced alloimmunization: summary of an NHLBI working group. Transfusion. 2011, vol.51: 435-441.
116. Biffl WL, Moore EE, Offner PJ, Ciesla DJ, Gonzalez RJ, Silliman CC. Plasma from aged stored red blood cells delays neutrophil apoptosis and primes for cytotoxicity: abrogation by poststorage washing but not prestorage leukoreduction. J.Trauma. 2001, vol.50: 426-431.
117. Dunne JR, Malone DL, Tracy JK, Napolitano LM. Allogenic blood transfusion in the first 24 hours after trauma is associated with increased systemic inflammatory response syndrome (SIRS) and death. Surg.Infect.(Larchmt.) 2004, vol.5: 395-404.
118. McFaul SJ, Corley JB, Mester CW, Nath J. Packed blood cells stored in AS-5 become proinflammatory during storage. Transfusion. 2009, vol.49: 1451-1460.
119. Vlaar AP, Hofstra JJ, Levi M, Kulik W, Nieuwland R, Tool AT, Schultz MJ, de Korte D, Juffermans NP. Supernatant of aged erythrocytes causes lung inflammation and coagulopathy in a "two-hit" in vivo syngeneic transfusion model. Anesthesiology. 2010, vol.113: 92-103.
120. Shaz BH, Stowell SR, Hillyer CD. Transfusion-related acute lung injury: from bedside to bench and back. Blood. 2011, vol.117: 1463-1471.
121. Valeri CR, Ragno G. The effects of preserved red blood cells on the severe adverse events observed in patients infused with hemoglobin based oxygen carriers. Artif.Cells Blood Substit.Immobil.Biotechnol. 2008, vol.36: 3-18.
122. Huggins C. Preparation and usefulness of frozen blood. Annu.Rev.Med. 1985, vol.36: 499-503.
123. Shander A, Hofmann A, Gombotz H, Theusinger OM, Spahn DR. Estimating the cost of blood: past, present, and future directions. Best.Pract.Res.Clin.Anaesthesiol. 2007, vol.21: 271-289.
124. Seifried E, Klueter H, Weidmann C, Staudenmaier T, Schrezenmeier H, Henschler R, Greinacher A, Mueller MM. How much blood is needed? Vox Sang. 2011, vol.100: 10-21.
125. Benjamin RJ, Whitaker BI. Boom or bust? Estimating blood demand and supply as the baby boomers age. Transfusion. 2011, vol.51: 670-673.

Chapter 6
The effects of cryopreservation on red blood cell rheologic properties
Sandra Henkelman Johan W.M. Lagerberg Reindert Graaff Gerhard Rakhorst Willem van Oeveren
Transfusion 50: 2393-2401, 2010
(Reproduced with permission of John Wiley and Sons)

Chapter 6
78
Abstract In transfusion medicine, cryopreserved RBCs are an alternative for refrigerated stored RBCs. Less is known about the rheologic properties of cryopreserved RBCs. In this study the aggregability and deformability of HGM cryopreserved RBCs that were post-thaw stored in SAGM solution were compared to that of refrigerated stored and fresh RBCs. Fresh RBCs were obtained from healthy volunteers while leukoreduced refrigerated stored and cryopreserved RBC units were obtained from the Sanquin Blood Bank. RBCs were tested for aggregability, deformability and various hematologic variables. The AI of cryopreserved RBCs was considerably reduced, compared to fresh and refrigerated stored RBCs. The EI of stored RBCs was significantly enhanced over a shear stress range of 2.0 to 50 Pa compared to fresh RBCs. No significant differences in EI between cryopreserved and 21- or 35-day refrigerated stored RBCs were observed. The osmotic fragility, hemolysis, MCV and MCHC of cryopreserved RBCs were markedly altered, compared to fresh and refrigerated stored RBCs (p < 0.05). The ATP content of cryopreserved RBCs was similar to fresh and 3-or 21-day refrigerated stored RBCs. These findings suggest that although cryopreserved RBCs are more fragile than fresh and refrigerated stored RBCs, the freeze-thaw-wash process did not did not adversely affect the aggregability, deformability or the ATP content of cryopreserved RBCs. Based on these rheologic properties, we conclude that cryopreserved RBCs are a valuable alternative to refrigerated stored RBCs.

Rheologic properties of frozen RBCs
79
6.1. Introduction In transfusion medicine RBCs are refrigerated stored for a maximum of 5 to 6 weeks before being discarded. Alternatively, cryopreservation enables storage of RBCs for years.1,2 Cryopreservation is currently a valuable approach for long-term storage of RBCs from donors with rare blood groups and for military deployment.3-5 However, stockpiling cryopreserved RBCs can also be beneficial in emergency or clinical situations, where the demand exceeds the supply of RBCs. At the moment, the shelf life of HGM cryopreserved RBCs has been approved for up to ten years.3,4 Usage of glycerol as a cryoprotectant requires a deglycerolization washing procedure post-thaw to prevent hemolytic transfusion reactions and renal failure after infusion.6-8 Usage of the ACP 215 device (Haemonetics, Braintree, MA) to glycerolize and deglycerolize RBC units has minimized the risk of bacterial contamination. As a result, the post-thaw storage time of RBCs has extended to 2 days in SAGM solution and to 14 days in AS-3 respectively.9 Current regulations require that cryopreserved RBCs have a post-thaw recovery of at least 80% and that the hemolysis in the RBC unit remains below allowable levels (i.e., 0.8% in Europe and 1% in the United States). Additionally, at least 75% of the cryopreserved RBCs should remain within the circulation 24 hour after infusion.3,4 Current regulations however, do not specifically address the quality of stored RBCs.10 The quality of HGM cryopreserved RBCs is primarily dependent on the pre-freeze and post-thaw storage time as well as on the anticoagulant and additive solution used. The duration of frozen storage per se, however, minimally attributes to cellular damage.1,2,11 Previously it was shown that cryopreserved RBCs have no significant loss of cellular ATP content or CD47 antigen expression on the external RBC membrane during 24 hours of post-thaw storage in AS-3. Moreover, when the pre-freezing storage time was limited to 3 days, no 2,3-DPG loss, microvesiculation or PS externalization was observed after 24 hours of post-thaw storage.1,12 Cryopreserved RBC units usually contain low potassium and free Hb content due to the removal of these extra cellular substances during washing. 9 The aggregability and deformability of cryopreserved RBCs remain to be elucidated. These rheologic properties are important determinants of the blood flow and hence the oxygenation of the tissues.13 In the venous system RBCs are able to form linear aggregates (so called Rouleaux) or more complex multi-cellular aggregates at low shear rates.14 Normally, the blood flow is sufficient to disperse these aggregates. However, under

Chapter 6
80
pathologic conditions stronger and larger aggregates may form that are more resistant to dispersion by the blood flow.15 In the microcirculation, the RBC ability to deform due to applied forces, make these cells capable of passing narrow capillaries. A high deformability of RBCs and a rapid recovery of the normal shape are essential factors for maintaining tissue perfusion and cell survival.16,17 Transfusion of rheologic impaired RBCs may hinder or obstruct the blood flow in the microcirculation, leading to reduced tissue perfusion, ischemia or infarction.13,18-20 In the Netherlands, leukoreduced RBCs are refrigerated stored in SAGM preservation solution for a maximum of 5 weeks. In chapter 4 we demonstrated that refrigerated storage minimally affected the RBC ability to aggregate and deform, even after 5 weeks of routine blood bank storage. Cryopreservation offers the advantage of storing RBCs for prolonged periods of time. However, once thawed the shelf life of RBCs is limited. In the current study, the aggregability, deformability and other hematologic variables of HGM cryopreserved RBCs that were post-thaw stored in SAGM solution for 2 days were assessed and compared to those of refrigerated stored and fresh RBCs.
6.2. Materials and Methods
RBC preparation and processing Human blood (50 ml ± 10 %) was collected after informed consent from ten healthy volunteers and anticoagulated with 7 ml of CPD. Functional measurements were performed with whole blood, whereas the rheologic features were determined with washed RBCs. Briefly, whole blood was washed by centrifugation at 1100 x g for 12 minutes and decanted three times with PBS pH 7.4. The final Hct of the RBC solution was set between 45% and 60 % and all measurements were performed within 3 hours after donation. Ten leukoreduced RBC units stored in SAGM solution were obtained from the Sanquin Blood Bank and refrigerated stored according to standard blood bank procedures as described in chapter 4. These RBC units, which were released for use on day 3 after donation, had a Hct of 45% to 60 % and contained fewer than 106 leukocytes per unit. In general, the average storage time of RBC units that are used in transfusion medicine is approximately 21 days.21,22 For analysis samples were aseptically withdrawn from the RBC units at 3, 21, and 35 days of refrigerated storage, reflecting short-, average- and long term stored RBCs, respectively.

Rheologic properties of frozen RBCs
81
Ten cryopreserved RBC units were obtained from the Sanquin blood bank and stored in polypropylene tubes at 2 to 6° for 48 hours. These RBC have been cryopreserved according to the HGM freezing method and post-thaw resuspended in SAGM solution as described previously.9 Briefly, leukoreduced refrigerated stored RBC units were centrifuged at 3200 x g for 5 minutes to remove the SAGM solution. Subsequently, the RBCs were glycerolized to a final concentration of 40% glycerol via the Haemonetics ACP-215 device.23 All glycerolized RBC units were frozen and stored at –80 ± 10°C in a mechanical freezer for 34.1 ± 20.5 months. Cryopreserved RBC units were thawed in a temperature-controlled water-bath of 40°C, until the units reached a temperature between 25°C and 30°C. Subsequently, thawed RBCs were deglycerolized via the Haemonetics ACP-215 device and resuspended in SAGM solution. The supernatant osmolarity of all cryopreserved units was below 400 mOsm/ kg H2O, indicating an efficient removal of glycerol.
Rheologic features RBC aggregability and deformability were monitored in vitro by the LORCA (R&R Mechatronics, Zwaag the Netherlands).24,25 Aggregation was induced by the addition of 10% HES (MW 200-kDa). Briefly, RBCs suspensions were centrifuged for 1 minute at 3500 x g and the supernatant was discarded. RBCs were resuspended in 10% HES 200-kDa solution (Fresenius, Bad Homburg, Germany) to obtain a Hct of 32 ± 6%. Aggregability was tested with 1 ml of the RBC suspension. RBC aggregation was monitored after complete disaggregation under increased shear stress. Both the aggregation measuring procedure and the subsequent analyses were computer controlled. Aggregability of RBCs was expressed by the AI, where a larger AI reflects an increased ability to aggregate. The AI was determined after correcting the Hct in all the samples to a constant value of 45%. The kinetics of aggregation (T½) was expressed by the time necessary to induce 50% aggregation. The deformability of RBCs was determined with RBC suspension that were diluted 1:100 in PBS (pH 6.5), containing 5% PVP (MW 360 kDa, Sigma-Aldrich, Germany) and with a viscosity of 30 mPa.sec. One ml of the latter RBC suspension was inserted into the LORCA and the RBC diffraction pattern was recorded at various shear stresses at 36.8 ± 0.2 °C. The deformability of the RBCs, which is expressed by the EI, was determined by the LORCA from the size of the vertical (L) and horizontal (W) axes of the diffraction pattern according

Chapter 6
82
to the formula: EI = (L-W) / (L+W). An increased EI at a given shear stress indicates greater RBC deformability. A deformability curve was obtained by plotting the calculated values for EI versus the corresponding shear stress. The deformability at two shear stress values were examined more closely; the deformability at a shear stress of 3.9 Pa, which reflects the rigidity of the cell membrane, and the maximal deformability at shear stress of 50 Pa.
Osmotic fragility The osmotic fragility of RBCs, which reflects the membrane’s ability to maintain structural integrity, was determined by diluting RBCs in PBS solutions ranging from 0.90% to 0.35%. RBCs with a Hct level of 30 to 35% were diluted 1:100 in each PBS solution, mixed and incubated for 30 minutes at 4°C, followed by centrifugation for 12 minutes at 1100 x g. The free Hb in the supernatant was measured by a spectrophotometer (PowerWave 200 spectrophotometer, Bio-Tek Instruments, USA). The concentration of PBS necessary to induce 50% hemolysis defined the osmotic fragility index of the RBCs.26 With this method, a larger osmotic fragility index corresponds to more fragile cells.
Hematologic variables The RBC MCV, the Hb concentration and the Hct were determined with a hematologic analyzer (Medonic CA 530-Oden, Sweden). The cytoplasmic viscosity of RBCs, which is determined by the MCHC,16,27 was calculated by dividing the Hb concentration by the Hct. The amount of free Hb in the RBC suspension was determined according to the Harboe method.28 In short, cell supernatant was obtained by centrifugation of RBC suspensions for 1 minute at 3500 x g. The supernatant was diluted 1:10 in 0.01% sodium carbonate in a flat-bottom 96-well microtiter plate and mixed for 30 minutes. The free Hb concentration in the supernatant was determined by measuring the OD at 415 nm and correcting for the OD at 380 and 450 nm (PowerWave 200 spectrophotometer, Bio-Tek Instruments, USA), according to the formula OD = 2* (OD 415 nm) – (OD 380 nm) – (OD 450 nm). The hemolysis was expressed as a percentage of the total amount of Hb present in the RBC lysates. To determine the RBC ATP content, RBC samples were incubated with 8% ice-cold trichloroacetic acid, in a ratio of 1:3, during a 30-minute period. The samples were

Rheologic properties of frozen RBCs
83
centrifuged for 1 minute at 3500 x g and the protein-free supernatant was neutralized with 1.5 mol/L sodium carbonate. Aliquots were stored at –80°C for later batch analyses of ATP. The ATP content was determined with a commercially available enzyme assay (Roche Diagnostics, Germany). For detection of ATP, light emission was measured at 560 nm by an illuminometer (Wallac 1420 Multilabel Counter, Perkin Elmer Life Sciences, Finland). Due to the detrimental influence of pre-freezing storage time on the RBC energy content,1,11 ATP was determined in RBC units with a pre-freezing storage time that did not exceeded 8 days.
Statistical analysis Statistical analysis was performed using statistical software (SPSS, version 16.0, SPSS Inc., Chicago, IL). Data were tested for normality with the Kolmgorov-Smirnov goodness-of-fit test. In the case of normally distributed data, differences between storage groups were demonstrated by using unpaired t-tests. Within storage groups, paired t-tests were performed to show differences over time. For none normally distributed data, the Mann-Whitney test was used to quantify differences between groups whereas the Wilcoxon signed ranks test was used to quantify differences within storage groups. Differences are considered to be significant with a two-tailed p value of less than 0.05. Results are presented as means ± SD.
6.3. Results
Rheologic features The RBC ability to aggregate, as represented by the AI, was significantly reduced in cryopreserved RBCs when compared to both fresh and refrigerated stored RBCs (Table 6.1). During storage, the AI of refrigerated stored RBCs was significantly reduced, whereas the AI of cryopreserved RBCs remained stable. The T½ of cryopreserved RBCs was significantly higher on Day 0, compared to refrigerated stored RBCs (Table 6.1). The latter observation indicated that aggregation formation immediately after deglycerolization was slightly slower. During storage, the T½ of refrigerated stored RBCs was significantly reduced in accordance with the AI, whereas the T½ of cryopreserved RBCs did not significantly change. It should be noted that the SD of both AI and T½ obtained with

Chapter 6
84
cryopreserved RBCs, were much larger than those of fresh and refrigerated stored RBCs. Probably the variability in (pre) freezing time may provide an explanation. The RBC deformation curve showed a typical sigmoid shape over a shear stress range of 0.6- 50 Pa for all the tested RBC groups (Figure 6.1). Yet, the deformation curves of cryopreserved and refrigerated stored RBCs were significantly elevated over the shear stress range of 2.0 Pa to 50 Pa compared to fresh RBCs. The deformability of RBCs at a shear stress of 3.9 Pa and 50 Pa was not significantly different between cryopreserved and 21- or 35-day refrigerated stored RBCs (Figure 6.2). During post-thaw storage, the RBC deformability at a shear stress of 3.9 Pa and 50 Pa did not yield significant changes. However, during refrigerated storage the RBC deformability at a shear stress of 3.9 Pa slightly increased (from 0.35 ± 0.01 to 0.38 ± 0.01 EI; p < 0.01), whereas the deformability at a shear stress of 50 Pa slightly decreased (from 0.58 ± 0.01 to 0.56 ± to 0.02; p < 0.01).
Table 6.1. Aggregation behavior of fresh, refrigerated stored and cryopreserved RBCs.
Measure Fresh Refrigerated storage Post-thaw storage
Day 3 Day 21 Day 35 Day 0 Day 2
AI (%) 45.3 ± 2.6 46.9 ± 2.4 46.0 ± 3.8 44.4 ± 4.5 † 36.0 ± 8.6 *† ‡§ 36.9 ± 8.0 *† ‡§
T ½ (sec) 1.3 ± 0.2 1.03 ± 0.3 * 0.86 ± 0.21 *† 0.91 ± 0.12 *† 2.4 ± 1.7 †‡§ 2.3 ± 2.4
Aggregation behavior was expressed as mean ± SD of ten units. Significant differences with a P- value < 0.05 were reported
* significantly different from fresh RBCs † significantly different from day 3 refrigerated stored RBCs ‡ significantly different from day 21 refrigerated stored RBCs § significantly different from day 35 refrigerated stored RBCs

Rheologic properties of frozen RBCs
85
Table 6.2. Hematologic variables of fresh, refrigerated stored and cryopreserved RBCs
Measure Fresh Refrigerated storage Post-thaw storage
Day 0 Day 3 Day 21 Day 35 Day 0 Day 2
MCV (fl) 89.1 ± 2.4 89.0 ± 2.4 91.1 ± 2.6 † 92.0 ± 2.8 *†‡ 104.2 ± 10.2 *†‡§ 106.4 ± 8.3 *†‡§
MCHC (mmol/L) 20.4 ± 0.4 20.7 ± 0.6 20.5 ± 0.6 20.2 ± 0.3 16.1 ± 0.6 *†‡§ 16.0 ± 0.8 *†‡§
Osmotic fragility (%) 0.45 ± 0.03 0.48 ± 0.02 0.46 ± 0.02 0.47 ± 0.02 0.66 ± 0.05 *†‡§ 0.67 ± 0.08 *†‡§
Hemolysis (%) 0.26 ± 0.04 0.24 ± 0.07 0.36 ± 0.12 *† 0.53 ± 0.24 *†‡ 0.57 ± 0.26 *†‡ 0.99 ± 0.28 *†‡§
ATP (μmol/gHb) 6.1 ± 3.1 4.6 ± 1.3 3.8 ± 1.1 2.1 ± 0.4 *†‡ 4.5 ± 2.2 § 4.0 ± 1.9 §
Values were expressed as mean ± SD of ten units. ATP values were determined from seven RBC units. Significant differences with a P-value less than 0.05 were reported * significantly different from fresh RBCs † significantly different from day 3 refrigerated stored RBCs ‡ significantly different from day 21 refrigerated stored RBCs § significantly different from day 35 refrigerated stored RBCs

Chapter 6
86
Figure 6.1. Shear stress EI curves for fresh, refrigerated stored and thawed deglycerolized RBCs. The shear stress value is plotted on logarithmic axis and data represent the mean ± SD of ten RBC units. The EI over a shear stress range of 2.0 Pa to 50 Pa of refrigerated stored and thawed deglycerolized RBCs was significantly elevated for all time points, compared to fresh RBCs (* p < 0.05). Fresh (▲); Day 3 liquid storage (■); Day 21 liquid storage (*); Day 35 liquid storage (--); Day 0 postthaw storage (Δ); Day 2 postthaw storage (♦).
Osmotic fragility The osmotic fragility index of cryopreserved RBCs, as represented by the osmolarity at half-maximum hemolysis, was markedly enhanced (p < 0.01) in comparison to both fresh and refrigerated stored RBCs (Table 6.2). These results indicate that cryopreserved RBCs are more fragile than fresh and refrigerated stored RBCs. During storage, the osmotic fragility index of cryopreserved and refrigerated stored RBCs did not change significantly.

Rheologic properties of frozen RBCs
87
A
B
Figure 6.2. Deformability for two representative shear stress levels as a function of storage time. (A) EI at a shear stress of 3.9 Pa. (B) EI at a shear stress of 50 Pa. Values are expressed as the mean ± SD of ten RBC units. Significant changes are illustrated in the figure (p < 0.05); * significantly different from fresh RBCs, † significantly different from day 3 refrigerated stored RBCs, ‡ significantly different from day 21 refrigerated stored RBCs

Chapter 6
88
Hematological variables Cryopreserved RBCs had a considerably higher MCV (p < 0.05) and a lower MCHC (p < 0.01) compared to fresh and refrigerated stored RBCs (Table 6.2). During storage, the MCV of refrigerated stored RBCs was significantly enhanced. At the end of storage, the MCV and MCHC of cryopreserved RBCs differed from refrigerated stored RBCs by 15.7 % and respectively 20.8 %. The hemolysis was generally higher in cryopreserved RBC units than in refrigerated stored RBC units (Table 6.2). Moreover, during storage the hemolysis increased with 1.7-fold in cryopreserved RBC units (p < 0.05) and with 2.2-fold in refrigerated stored RBC units (p < 0.01). As a result, after 2 days of post-thaw storage the hemolysis in the RBC unit slightly exceeded the allowable limits, whereas after 35 days of refrigerated storage the hemolysis in the RBC unit clearly remained below these limits (i.e., 0.8% in Europe and 1% in the United States).3,4 The ATP content of cryopreserved RBCs was similar to that of fresh and 3- or 21-day refrigerated stored RBCs (Table 6.2). During storage, the ATP content of cryopreserved and refrigerated stored RBCs reduced with 8.6 and 53.4% respectively. As a result, the ATP content at the end of refrigerated RBC storage was significantly different from that of fresh and cryopreserved RBCs. None of the aerobic and anaerobic blood cultures of cryopreserved RBC units showed evidence of bacterial contamination.
6.4. Discussion The RBC aggregability and deformability are important hemodynamic determinants. Alterations in RBC rheology, that is, enhanced RBC aggregability and reduced deformability, have been observed in a variety of pathological states such as sepsis, myocardial ischemia, renal failure, inflammation, diabetes mellitus, obesity, hypertension, sickle cell disease and malaria.15,19,20,29,30 Throughout the years, the rheologic properties of refrigerated stored RBCs have been extensively investigated.31-40 So far, the rheologic properties of cryopreserved RBCs have scarcely been investigated.41-43 The current study was undertaken to explore the rheologic features and hematologic variables of HGM cryopreserved RBCs and to compare this to conventional refrigerated stored and fresh RBCs.

Rheologic properties of frozen RBCs
89
In this study the aggregability of cryopreserved RBCs was markedly reduced, in comparison to that of refrigerated stored and fresh RBCs. The time necessary to induce aggregation was also slightly prolonged after deglycerolization, compared to refrigerated stored RBCs. In circulating blood, this may result in less effective transport of RBCs in the microcirculation. The ability of RBCs to aggregate is dependent on cellular properties and the composition of the suspension medium.20 In our study the suspension medium was standardized for all samples, indicating that the observed differences were caused by alterations in cellular properties only. Cell morphology is a major determinant of RBC aggregability. Cell swelling suppresses cell contact and subsequently Rouleaux formation.44,45 Previously, it has been shown that the freeze-thaw (wash) process made the RBC membrane permeable to cations and hence induced cell swelling.9,46 Our study supports these results, as the MCV of cryopreserved RBCs markedly exceeded the MCV of fresh and refrigerated stored RBCs. This gain in RBC volume, as obtained by the freeze-thaw-wash process, could be responsible for the observed reduction in aggregability. Although the clinical relevance of altered RBC aggregability is an ongoing debate,47 RBC swelling during storage is probably irrelevant, as long as it is reversible upon transfusion. In vitro data obtained from refrigerated stored RBCs support the hypothesis that RBC swelling can be reversed in plasma.40 The RBC ability to deform depends on the cytoplasmic viscosity of the cell, which is reflected by the MCHC, as well as on the overall cell shape and the viscoelastic properties of the cytoskeleton.16 Our data showed no significant difference in the deformability between cryopreserved and long-term refrigerated stored RBCs. The deformation curves of both cryopreserved and refrigerated stored RBCs were, however, significantly elevated as compared to fresh RBCs. At a shear stress of 3.9 Pa an increasing deformability of refrigerated stored RBC and an increased deformability of cryopreserved RBC, with respect to fresh RBCs, was observed during storage, despite an enhanced MCV. At a shear stress of 50 Pa the deformability during refrigerated storage reduced, but was still elevated, whereas the deformability of cryopreserved RBCs was elevated and comparable to long-term refrigerated stored RBCs. These observations indicated that the RBC deformability in the low shear stress regions was less affected by changes in MCV. The deformability at a shear stress of 3.9 Pa, a shear stress which is predominantly found in the microcirculation,48 is thus a direct reflection of the rigidity of the cell membrane. Altogether, the increased MCV and the resultant lowered MCHC value of cryopreserved RBCs did not seem to adversely

Chapter 6
90
influence the RBC flexibility, because the deformability of cryopreserved RBCs was still higher than the deformability of fresh RBCs. The improved flexibility of 21- and 35-day refrigerated stored as well as that of cryopreserved RBCs, could be explained by di(2-ethylhexyl)phthalate (DEHP) leaking from the PVC storage bag. This plasticizer, which is added to the PVC to impart flexibility, is known to improve RBC storage by suppressing hemolysis, microvesiculation and morphology changes.49-53 It was also shown that the presence of DEHP improved the flexibility of RBCs during long-term refrigerated storage.54 We hypothesize that DEHP enhances the viscoelastic properties of the cytoskeleton, explaining the improved deformability of long-term refrigerated stored and cryopreserved RBCs, despite the gain in cell volume. The observation that the deformability at a shear stress of 3.9 Pa of both fresh and 3-day refrigerated stored RBCs was significantly lower than long-term refrigerated stored and cryopreserved RBCs further substantiates this hypothesis. In the past, the RBC flexibility has been investigated after subjection of RBCs to subzero temperatures.41-43 Nevertheless, in those studies the RBC storage conditions were not representative for current clinical standards, explaining the discrepancy between our results. The osmotic fragility was significantly increased in cryopreserved RBCs compared to fresh and refrigerated stored RBCs. This indicates that the freeze-thaw-wash process made the RBCs more fragile. Moreover, the hemolysis was also significantly higher during post-thaw storage. At the end of storage, the hemolysis in cryopreserved RBC units exceeded the allowable limits. Previous studies demonstrated, however, that hemolysis during 48 hours of post-thaw storage remained clearly within the allowable limits.5,9,23 In our study, polypropylene containers were used for storage of thawed RBCs. It cannot be ruled out that the lower gas permeability or the absence of DEHP plasticizer of these containers, was responsible for the increased hemolysis observed in our experiments.55 ATP as an energy source is important for the overall functioning of the RBC. Loss of ATP is associated with more rigid cell membranes, echinocyte shape change, enhanced cation permeability, loss of vasodilatation control, exposure of phosphatidylserine on the external RBC membrane, microvesiculation, and decreased RBC viability.56-64 In the past it was shown that the RBC ATP content must be at least 2.7 µmol per gram Hb to have a 90 percent chance of acceptable in vivo survival (24-hr in vivo recovery of 75% or higher). 65,66 In our study, the ATP content of 35-day refrigerated stored RBCs was slightly below this limit, whereas the ATP content of cryopreserved RBCs remained close to those of fresh

Rheologic properties of frozen RBCs
91
RBCs. These results are in line with current findings, which showed a minimal loss of ATP in cryopreserved RBCs.9,12,23 More recently, it was indicated that the RBC ATP content plays a prominent role in restoring oxygen deficits in the microcirculation.67,68 Transfusion of cryopreserved RBCs with physiological ATP content would therefore favor the RBC viability and the oxygen delivery to the tissues. In transfusion medicine, cryopreserved RBCs are a valuable blood resource for controlling an inventory in situations where the RBC availability is limited or unpredictable. This is the case for storage of RBC with rare blood types or for usage in military settings and occasionally during civil disasters.5,69,70 Routine usage of cryopreserved RBCs in transfusion medicine is limited due to the more expensive, time consuming and less efficient nature of this preservation method. Consequently, the unfamiliarity with regard to the quality of cryopreserved RBCs has further limited clinical usage over the years. Yet, cryopreserved RBCs showed satisfactory in vitro quality and posttransfusion in vivo survival,1,9,12,71 Improving the RBC freezing technology with preservation of the current quality, could ultimately make cryopreserved RBCs more utilizable for modern transfusion medicine. Our data demonstrate that although cryopreserved RBCs are more fragile than refrigerated stored and fresh RBCs, the freeze-thaw-wash process did not adversely affect the ATP content or the aggregability and deformability of cryopreserved RBCs. From a rheologic point of view, we concluded that cryopreserved RBCs are a valuable alternative to refrigerated stored RBCs for usage in transfusion medicine.
References 1. Lecak J, Scott K, Young C, Hannon J, Acker JP. Evaluation of red blood cells stored at -80°C in excess of 10
years. Transfusion 2004, vol.44: 1306-1313.
2. Valeri CR, Ragno G, Pivacek LE, Cassidy GP, Srey R, Hansson-Wicher M, Leavy ME. An experiment with glycerol-frozen red blood cells stored at -80 °C for up to 37 years. Vox Sang. 2000, vol.79: 168-174.
3. Council of Europe. Guide to the preparation, use and quality assurance of blood components, 14th edition. Strasbourg: Council of Europe publisher; 2008.
4. Kakaiya R, Aronson CA, Julleis J. Whole blood collection and component processing, AABB Technical Manual 16th ed. Bethesda, MD.: AABB Press; 2008.
5. Lelkens CC, Koning JG, de Korte D, Floot IB, Noorman F. Experiences with frozen blood products in the Netherlands military. Transfus.Apher.Sci. 2006, vol.34: 289-298.

Chapter 6
92
6. Bechdolt S, Schroeder LK, Samia C, Schmidt PJ. In vivo hemolysis of deglycerolized red blood cells. Arch.Pathol.Lab Med. 1986, vol.110: 344-345.
7. Cregan P, Donegan E, Gotelli G. Hemolytic transfusion reaction following transfusion of frozen and washed autologous red cells. Transfusion 1991, vol.31: 172-175.
8. Klein HG, Anstee D. Mollison's Blood Transfusion in Clinical Medicine, 11th ed. Massachusetts: Blackwell Publishing; 2005.
9. Lagerberg JW, Truijens-de Lange R, de Korte D, Verhoeven AJ. Altered processing of thawed red cells to improve the in vitro quality during postthaw storage at 4 °C. Transfusion 2007, vol.47: 2242-2249.
10. Hogman CF, Meryman HT. Red blood cells intended for transfusion: quality criteria revisited. Transfusion 2006, vol.46: 137-142.
11. Valeri CR, Ragno G, Van HP, Rose L, Rose M, Egozy Y, Popovsky MA. Automation of the glycerolization of red blood cells with the high-separation bowl in the Haemonetics ACP 215 instrument. Transfusion 2005, vol.45: 1621-1627.
12. Holovati JL, Wong KA, Webster JM, Acker JP. The effects of cryopreservation on red blood cell microvesiculation, phosphatidylserine externalization, and CD47 expression. Transfusion 2008, vol.48: 1658-1668.
13. Marossy A, Svorc P, Kron I, Gresova S. Hemorheology and circulation. Clin.Hemorheol.Microcirc. 2009, vol.42: 239-258.
14. Bishop JJ, Popel AS, Intaglietta M, Johnson PC. Rheological effects of red blood cell aggregation in the venous network: a review of recent studies. Biorheology 2001, vol.38: 263-274.
15. Rampling MW, Meiselman HJ, Neu B, Baskurt OK. Influence of cell-specific factors on red blood cell aggregation. Biorheology 2004, vol.41: 91-112.
16. Mohandas N, Gallagher PG. Red cell membrane: past, present, and future. Blood 2008, vol.112: 3939-3948.
17. Parthasarathi K, Lipowsky HH. Capillary recruitment in response to tissue hypoxia and its dependence on red blood cell deformability. Am.J.Physiol 1999, vol.277: H2145-H2157.
18. Kaul DK, Koshkaryev A, Artmann G, Barshtein G, Yedgar S. Additive effect of red blood cell rigidity and adherence to endothelial cells in inducing vascular resistance. Am.J.Physiol Heart Circ.Physiol. 2008, vol.295: H1788-H1793.
19. Mchedlishvili G. Disturbed blood flow structuring as critical factor of hemorheological disorders in microcirculation. Clin.Hemorheol.Microcirc. 1998, vol.19: 315-325.
20. Yedgar S, Koshkaryev A, Barshtein G. The red blood cell in vascular occlusion. Pathophysiol. Haemost.Thromb. 2002, vol.32: 263-268.
21. Raat NJ, Berends F, Verhoeven AJ, de Korte D, Ince C. The age of stored red blood cell concentrates at the time of transfusion. Transfus.Med. 2005, vol.15: 419-423.
22. Corwin HL, Gettinger A, Pearl RG, Fink MP, Levy MM, Abraham E, MacIntyre NR, Shabot MM, Duh MS, Shapiro MJ. The CRIT Study: Anemia and blood transfusion in the critically ill--current clinical practice in the United States. Crit Care Med. 2004, vol.32: 39-52.

Rheologic properties of frozen RBCs
93
23. Lelkens CC, Noorman F, Koning JG, Truijens- de Lange R, Stekkinger PS, Bakker JC, Lagerberg JW, Brand A, Verhoeven AJ. Stability after thawing of RBCs frozen with the high- and low-glycerol method. Transfusion 2003, vol.43: 157-164.
24. Hardeman MR, Ince C. Clinical potential of in vitro measured red cell deformability, a myth? Clin.Hemorheol.Microcirc. 1999, vol.21: 277-284.
25. Hardeman MR, Dobbe JG, Ince C. The Laser-assisted Optical Rotational Cell Analyzer (LORCA) as red blood cell aggregometer. Clin.Hemorheol.Microcirc. 2001, vol.25: 1-11.
26. Gyongyossy-Issa MI, Weiss SL, Sowemimo-Coker SO, Garcez RB, Devine DV. Prestorage leukoreduction and low-temperature filtration reduce hemolysis of stored red cell concentrates. Transfusion 2005, vol.45: 90-96.
27. Shiga T, Maeda N, Kon K. Erythrocyte rheology. Crit Rev.Oncol.Hematol. 1990, vol.10: 9-48.
28. Harboe M. A method for determination of hemoglobin in plasma by near-ultraviolet spectrophotometry. Scand.J.Clin.Lab Invest 1959, vol.11: 66-70.
29. Reggiori G, Occhipinti G, de Gasperi A, Vincent JL, Piagnerelli M. Early alterations of red blood cell rheology in critically ill patients. Crit Care Med. 2009, vol.37: 3041-3060.
30. Banerjee R, Nageshwari K, Puniyani RR. The diagnostic relevance of red cell rigidity. Clin.Hemorheol.Microcirc. 1998, vol.19: 21-24.
31. Bennett-Guerrero E, Veldman TH, Doctor A, Telen MJ, Ortel TL, Reid TS, Mulherin MA, Zhu H, Buck RD, Califf RM, et al. Evolution of adverse changes in stored RBCs. Proc.Natl.Acad.Sci 2007, vol.104: 17063-17068.
32. Berezina TL, Zaets SB, Morgan C, Spillert CR, Kamiyama M, Spolarics Z, Deitch EA, Machiedo GW. Influence of storage on red blood cell rheological properties. J.Surg.Res. 2002, vol.102: 6-12.
33. Hovav T, Yedgar S, Manny N, Barshtein G. Alteration of red cell aggregability and shape during blood storage. Transfusion 1999, vol.39: 277-281.
34. Izzo P, Manicone A, Spagnuolo A, Lauta VM, Di Pasquale A, Di Monte D. Erythrocytes stored in CPD SAG-mannitol: evaluation of their deformability. Clin.Hemorheol.Microcirc. 1999, vol.21: 335-339.
35. Kirkpatrick UJ, Adams RA, Lardi A, McCollum CN. Rheological properties and function of blood cells in stored bank blood and salvaged blood. Br.J.Haematol. 1998, vol.101: 364-368.
36. Nagaprasad V, Singh M. Sequential analysis of the influence of blood storage on aggregation, deformability and shape parameters of erythrocytes. Clin.Hemorheol.Microcirc. 1998, vol.18: 273-284.
37. Relevy H, Koshkaryev A, Manny N, Yedgar S, Barshtein G. Blood banking-induced alteration of red blood cell flow properties. Transfusion 2008, vol.48: 136-146.
38. Taleghani BM, Grossmann R, Waltenberger G, Geise W, Langer R, Henrich HA, Wiebecke D. Storage induced rheological and biochemical changes in erythrocyte concentrates with added solution and possible correlations. Beitr Infusionsther Transsfusionsmed 1996, vol.33: 141-144.
39. Taleghani BM, Langer R, Grossmann R, Opitz A, Halbsguth U, Buchheister A, Schuler S, Bachthaler R, Wiebecke D. Improvement of the biochemical rheological quality of whole blood and red cell concentrates

Chapter 6
94
obtained by leukocyte depletion before storage. Anasthesiol.Intensivmed.Notfallmed.Schmerzther. 2001, vol.36: S11-S19.
40. Zehnder L, Schulzki T, Goede JS, Hayes J, Reinhart WH. Erythrocyte storage in hypertonic (SAGM) or isotonic (PAGGSM) conservation medium: influence on cell properties. Vox Sang. 2008, vol.95: 280-287.
41. Wegner G, Kucera W. Changes in erythrocyte deformability in blood preservation. Z.Gesamte Inn.Med. 1987, vol.42: 575-581.
42. Chien S, Usami S, Rowe AW. Rheologic properties of erythrocytes preserved in liquid nitrogen. J.Lab Clin.Med. 1971, vol.78: 175-180.
43. Mantzavinos D, Bailey AI, Rampling MW. Flash freezing of erythrocyte suspensions. Biorheology 1997, vol.34: 73-83.
44. Reinhart WH, Singh-Marchetti M, Straub PW. The influence of erythrocyte shape on suspension viscosities. Eur.J.Clin.Invest.1992, vol.22: 38-44.
45. Cicha I, Suzuki Y, Tateishi N, Maeda N. Changes of RBC aggregation in oxygenation-deoxygenation: pH dependency and cell morphology. Am.J.Physiol Heart Circ.Physiol 2003, vol.284: H2335-H2342.
46. Kofanova OA, Zemlyanskikh NG, Ivanova L, Bernhardt I. Changes in the intracellular Ca2+ content in human red blood cells in the presence of glycerol. Bioelectrochemistry. 2008, vol.73: 151-154.
47. Baskurt OK, Meiselman HJ. Hemodynamic effects of red blood cell aggregation. Indian J.Exp.Biol. 2007, vol.45: 25-31.
48. Pries AR, Secomb TW. Microcirculatory network structures and models. Ann.Biomed.Eng 2000, vol.28: 916-921.
49. Deepa Devi KV, Manoj Kumar V, Arun P, Santhosh A, Padmakumaran Nair KG, Lakshmi LR, Kurup PA. Increased lipid peroxidation of erythrocytes in blood stored in polyvinyl chloride blood storage bags plasticized with di-(2-ethyl hexyl) phthalate and effect of antioxidants. Vox Sang. 1998, vol.75: 198-204.
50. Draper CJ, Greenwalt TJ, Dumaswala UJ. Biochemical and structural changes in RBCs stored with different plasticizers: the role of hexanol. Transfusion 2002, vol.42: 830-835.
51. Estep TN, Pedersen RA, Miller TJ, Stupar KR. Characterization of erythrocyte quality during the refrigerated storage of whole blood containing di-(2-ethylhexyl) phthalate. Blood 1984, vol.64: 1270-1276.
52. Hill HR, Oliver CK, Lippert LE, Greenwalt TJ, Hess JR. The effects of polyvinyl chloride and polyolefin blood bags on red blood cells stored in a new additive solution. Vox Sang. 2001, vol.81: 161-166.
53. Rock G, Tocchi M, Ganz PR, Tackaberry ES. Incorporation of plasticizer into red cells during storage. Transfusion 1984, vol.24: 493-498.
54. Labow RS, Card RT, Rock G. The effect of the plasticizer di(2-ethylhexyl)phthalate on red cell deformability. Blood 1987, vol.70: 319-323.
55. Sowemimo-Coker SO. Red blood cell hemolysis during processing. Transfus.Med.Rev. 2002, vol.16: 46-60.
56. Betz T, Lenz M, Joanny JF, Sykes C. ATP-dependent mechanics of red blood cells. Proc.Natl.Acad.Sci. 2009, vol.106: 15320-15325.

Rheologic properties of frozen RBCs
95
57. Bucki R, Bachelot-Loza C, Zachowski A, Giraud F, Sulpice JC. Calcium induces phospholipid redistribution and microvesicle release in human erythrocyte membranes by independent pathways. Biochemistry 1998, vol.37: 15383-15391.
58. Gov NS, Safran SA. Red blood cell membrane fluctuations and shape controlled by ATP-induced cytoskeletal defects. Biophys.J. 2005, vol.88: 1859-1874.
59. Gov NS. Active elastic network: cytoskeleton of the red blood cell. Phys.Rev.E.Stat.Nonlin.Soft.Matter Phys. 2007, vol.75:011921.
60. Heaton WA. Evaluation of posttransfusion recovery and survival of transfused red cells. Transfus.Med.Rev. 1992, vol.6: 153-169.
61. Johnson RM. Membrane stress increases cation permeability in red cells. Biophys.J. 1994, vol.67:1876-1881.
62. Park Y, Best CA, Auth T, Gov NS, Safran SA, Popescu G, Suresh S, Feld MS. Metabolic remodeling of the human red blood cell membrane. Proc.Natl.Acad.Sci. 2010, vol.107: 1289-1294.
63. Verhoeven AJ, Hilarius PM, Dekkers DW, Lagerberg JW, de Korte D. Prolonged storage of red blood cells affects aminophospholipid translocase activity. Vox Sang. 2006, vol.91: 244-251.
64. Wan J, Ristenpart WD, Stone HA. Dynamics of shear-induced ATP release from red blood cells. Proc.Natl.Acad.Sci.2008, vol.105: 16432-16437.
65. Stichting Sanquin bloedvoorziening. Richtlijn Bloedproducten. PT009.RL.SQ_002. Amsterdam: 2005.
66. van der Meer PF, Pietersz RN. The effect of plastic overwraps on storage measures of red cell concentrates. Vox Sang. 2007, vol.93: 176-178.
67. Raat NJ, Verhoeven AJ, Mik EG, Gouwerok CW, Verhaar R, Goedhart PT, de Korte D, Ince C. The effect of storage time of human red cells on intestinal microcirculatory oxygenation in a rat isovolemic exchange model. Crit Care Med. 2005, vol.33: 39-45.
68. Raat NJ, Hilarius PM, Johannes T, de Korte D, Ince C, Verhoeven AJ. Rejuvenation of stored human red blood cells reverses the renal microvascular oxygenation deficit in an isovolemic transfusion model in rats. Transfusion 2009, vol.49: 427-434.
69. Hess JR. Red cell freezing and its impact on the supply chain. Transfus.Med. 2004, vol.14: 1-8.
70. Zimrin AB, Hess JR. Planning for pandemic influenza: effect of a pandemic on the supply and demand for blood products in the United States. Transfusion 2007, vol.47: 1071-1079.
71. Valeri CR, Ragno G, Pivacek LE, Srey R, Hess JR, Lippert LE, Mettille F, Fahie R, O'Neill EM, Szymanski IO. A multicenter study of in vitro and in vivo values in human RBCs frozen with 40-percent (wt/vol)
glycerol and stored after deglycerolization for 15 days at 4°C in AS-3: assessment of RBC processing in the ACP 215. Transfusion 2001, vol.41: 933-939.


Chapter 7
A new freezing technique allows storage of red blood cells with 20% glycerol at -80°C
Sandra Henkelman Willem van Oeveren John Blanton Reindert Graaff Gerhard Rakhorst
Submitted

Chapter 7
98
Abstract Freezing RBCs with 40% glycerol at -80°C allow long-term storage of RBCs. However, due to the high glycerol content, up to 20% of the RBCs are lost during the addition and removal of glycerol. A liquid Bio-freezer with a rapid heat exchange could be suitable to preserve RBCs with lower glycerol concentrations. In this study, the quality and in vivo survival of RBCs cryopreserved in a liquid Bio-freezer were investigated. Human and rat RBCs were cryopreserved with 20 or 40% glycerol (Bio-20 or Bio-40) in the Bio-freezer at -25°C or with 20% glycerol, followed by storage at -80°C in a mechanical freezer (Combi-20). Cryopreserved human RBCs were tested in vitro for deformability, hemolysis and various other hematologic variables. The posttransfusion survival was determined in rats with chromium-51 labeled RBCs. After deglycerolization, Bio-20 cryopreserved RBCs demonstrated 0.80 ± 0.3 % hemolysis and reduced EI, compared to fresh RBCs (p < 0.05). Bio-40 cryopreserved RBCs maintained the EI but the 48-hour posttransfusion survival values were lower than those of fresh RBCs (p < 0.05). Combi-20 cryopreserved RBCs showed 0.50 ± 0.1% hemolysis and the MCV, EI and the 48-hour posttransfusion survival values were not significant different from fresh RBCs. The temperature of the Bio-freezer was not cold enough to maintain the RBC quality during storage, neither with 20% nor with 40% glycerol. However, combining the Bio-freezer with -80°C storage enabled cryopreservation of RBCs with 20% glycerol, while maintaining the RBC integrity, deformability and high 48-hour posttransfusion survival values. This new freezing method could contribute to an increased use of cryopreservation in transfusion medicine.

Quality and viability of RBCs frozen in a Bio-freezer
99
7.1. Introduction In transfusion medicine, cryopreservation has been particularly useful for long-term storage of RBCs with rare phenotypes and to stock RBCs for military deployment.1-4 In addition, stockpiling frozen RBCs could be beneficial in emergency or clinical situations, where the demand exceeds the supply of RBCs.5-7 Currently, the HGM is the most applied RBC freezing method in Europe and the United States. This method allows preservation of RBCs with a final concentration of approximately 40% (wt/vol) glycerol at temperatures between -65 and -80°C for up to ten years.1,2 Usage of glycerol as a cryoprotectant requires an intensive deglycerolization washing procedure post-thaw in order to reduce the glycerol concentration within the cell. International guidelines require that cryopreserved RBCs have a post-thaw recovery of at least 80% and that the hemolysis in the RBC unit remains below allowable levels (i.e., 0.8% in Europe and 1% in the United States). Additionally, at least 75% of the cryopreserved RBCs should remain within the circulation 24 hour after infusion.1,2 The RBC viability, as defined by the posttransfusion survival of RBCs, is often determined with Chromium-51 (51CR) labeled RBCs. In the Netherlands this method is not allowed in humans because of the radiation exposure. However, murine transfusion models have been proven useful for assessing the in-vivo survival of short term preserved RBCs.8-11 In transfusion medicine, routine usage of cryopreserved RBCs is still limited, due to the more expensive and labor intensive nature of this preservation method. Also HGM frozen storage is less efficient due to the freeze-thaw-wash related cell loss of approximately 10 to 20 %.12-14 A freezing method which would allow RBCs to be frozen with lower glycerol concentrations could reduce the cell loss and subsequently make cryopreserved RBCs more applicable for routine clinical usage. Currently, a new liquid Bio-freezer (Supachill, Lubbock, USA) has become available which enables preservation of sperm cells at temperatures of -25°C to-32°C with a high post-thaw quality.15,16 This new Bio-freezer consists of a reservoir of fluid with a high heat absorption capacity. Sperm samples are immersed in the fluid and heat is rapidly dissipated from the sample, which limits the freezing damage. The rate of heat release during freezing is also an important contributor to the post-thaw quality of RBCs.17 Traditional freezing of RBCs at -25°C in a mechanical freezer, normally requires high glycerol concentrations to minimize ice formation.18 However, high concentrations of cryoprotectants can exert detrimental

Chapter 7
100
effects on the cell.19 The rapid heat exchange of the liquid Bio-freezer, might allow RBCs to be frozen with lower glycerol concentrations at higher temperatures. The latter could reduce the osmotic stress and subsequently the freeze-thaw-wash related cell loss. This characteristic could make the liquid Bio-freezer interesting for the cryopreservation of RBCs. The deformability of the cell membrane is a major determinant of the RBC posttransfusion survival, because less deformable cells are removed from the circulation via the spleen.20-22 In addition, a high RBC deformability and a rapid recovery to the normal shape are essential for capillary passage and subsequently to maintain adequate tissue perfusion.23 In chapter 6 we demonstrated that the freeze-thaw-wash process did not adversely affect the deformability of HGM cryopreserved RBCs. Previously, it was also demonstrated that cryopreserved RBCs showed satisfactory 24-hour posttransfusion survival, even after 7 days of post-thaw storage in SAGM solution.24 The objective of this study was to assess whether the liquid Bio-freezer could be suitable for the preservation of RBCs, especially with low glycerol concentrations. The RBC deformability and other in vitro quality measures were determined with human RBCs whereas a rat transfusions model was used to test the in vivo survival of preserved rat RBCs.
7.2. Materials and methods
RBC collection and processing Human whole blood was obtained from eight healthy volunteers, after informed consent and in accordance with University Medical Center Groningen protocols. Briefly, whole blood (54 ± 2 ml) was anticoagulated with 14% CPD and centrifuged at 1100 x g for 12 minutes, to remove the buffycoat and supernatant. The concentrated RBCs were washed twice with PBS (pH 7.4), after which they were resuspended in PBS to a final Hct value of 50 ± 10 %. This volume allowed the preparation of identical samples in the glycerolization procedure. The animal experiments to study in vivo RBC survival were approved by the ethical committee for animal research of the University Medical Center Groningen. Animal care and handling was performed in accordance with Dutch guidelines for the care of laboratory animals. Rats were anaesthetized with 2.5% isoflurane and 0.8 L oxygen per minute. A total of twenty six male inbred Lewis rats (LEW/Hantm Hsd: Harlan, the Netherlands), with

Quality and viability of RBCs frozen in a Bio-freezer
101
a body weight of 270 ± 23g, were included in this study. Twenty rats were used for transfusion experiments, whereas six rats were used to collect the required amount of blood. Briefly, whole blood (10 ± 1 ml) was obtained by cardiac puncture of anaesthetized rats. Rat blood was collected into 14% CPD and processed similar as described above for human blood. After blood withdrawal, the rats were euthanized.
Glycerolization Human and rat RBCs were glycerolized at room temperature to obtain a final concentration of 20% or 40% glycerol. Briefly, an equal amount of standard 57% (wt/vol) glycerol mixture (S.p.A. Laboratorio Farmacologico, Italy) was added to the RBCs,25 to achieve a final concentration of approximately 40% (wt/vol) glycerol. A 25% (wt/vol) glycerol mixture, containing 50mM sodium phosphate and 46mM sodium gluconate, was added in a ratio of 5:1 to the RBCs to achieve a final concentration of approximately 20% (wt/vol) glycerol. To estimate the glycerol concentration within the RBCs,26 the supernatant osmolarity of the RBC suspension was measured with a cryoscopic osmometer (Osmomat 030-D, Gonotec, Germany).
RBC preservation The liquid Bio-freezer consists of a reservoir of circulating fluid which is set at a temperature of -25°C. RBC suspensions are immersed in the fluid allowing heat to be rapidly dissipated from the sample. A mechanical freezer was used to store RBCs at temperatures of -80°C. Human and rat RBCs were frozen for 6 weeks and respectively 48 hours according to different protocols (Table 7.1). In short, human and rat RBCs with a Hct of 50 ± 10% were incubated with the appropriate glycerol mixture for 15 minutes. RBC suspensions were frozen in 20 ml syringes (Omnifix, Braun, Germany) within 2 hours after donation. In protocol Combi-20, human RBCs were frozen for 24-hours in the liquid Bio-freezer prior to storage at –80°C, while rat RBCs were frozen for 2-hours in the liquid Bio-freezer prior to storage at –80°C. To mimic routine refrigerated blood bank storage, washed rat RBCs were diluted 2:1 in SAGM solution (Fresenius Hemocare, Germany) and stored at 4 ± 2 °C. RBC samples were withdrawn from the bag one week after storage for determination of the

Chapter 7
102
posttransfusion survival. The RBCs were washed and resuspended in PBS to a Hct of 30-35% and radioactive labeled, as will described further on.
Table 7.1. Freezing protocols for human and rat RBCs
Freezing protocol
Bio-20 Combi-20 Bio-40
Final glycerol % (wt/vol) 20% Glycerol 20% Glycerol 40% Glycerol
Freezing -25°C liquid Bio-freezer -25°C liquid Bio-freezer -25°C liquid Bio-freezer
Storage -25°C liquid Bio-freezer -80°C mechanical freezer -25°C liquid Bio-freezer
Deglycerolization Human and rat RBC suspensions were thawed at room temperature for 40 minutes with gentle inversion of the syringes every 10 minutes. Thawed RBC suspensions were manually deglycerolized by washing with NaCl solutions of decreasing osmolarity. Briefly, 2 ml of thawed Bio-20 or Combi-20 RBC suspension were diluted with 0.5 ml of 8% NaCl, followed by dilutions with 2.5 ml, 3 ml and 16 ml of 0.9% NaCl, with a 3 minute incubation period between each step. After centrifugation for 12 minutes at 1100 x g the supernatant was discarded. Human RBCs were resuspended in PBS to a final Hct of 40-45%, while rat RBCs were resuspended to a final Hct of 30-35%. Thawed Bio-40 RBC suspensions were treated similar as above, with the exception of using 12% instead of 8% NaCl in the first washing step. In all thawed deglycerolized samples, the supernatant osmolarity was measured to ensure an efficient removal of glycerol. Functionality testing with human and rat RBCs were performed within 2- and respectively 4 hours after deglycerolization.

Quality and viability of RBCs frozen in a Bio-freezer
103
Hematologic variables The RBC MCV and the total Hb concentration were determined with a hematologic analyzer (Medonic CA 530-Oden, Sweden). The Hct was manually determined using a micro-centrifuge (Mikro-20, Hettich, Germany). The MCHC, which is a primary determinant of cytoplasmic viscosity,27,28 was calculated by dividing the Hb concentration by the Hct. Hemolysis as measured by the amount of free Hb present in human RBC suspensions,29 was determined after glycerolization, thawing and deglycerolization respectively. Briefly, cell supernatant was obtained by centrifugation of RBC suspensions for 1 minute at 3500 x g. The supernatant was diluted 1:10 in 0.01% sodium carbonate in a flat-bottom 96-well microtiter plate and mixed for 30 minutes. The free Hb concentration in the supernatant was determined by measuring the OD at 415 nm and correcting for the OD at 380 and 450 nm (PowerWave 200 spectrophotometer, Bio-Tek Instruments, USA), according to the formula OD = 2* (OD 415 nm) – (OD 380 nm) – (OD 450 nm). Hemolysis was expressed as a percentage of the total amount of Hb present in the RBC lysates. RBC deformability RBC deformability was monitored in vitro by the LORCA (R&R Mechatronics, Zwaag the Netherlands).30 Deformability was determined with fresh and thawed deglycerolized human RBC suspensions diluted 1:100 in PBS, pH 6.5, containing 5% polyvinylpyrrolidone (MW 360 kDa, Sigma-Aldrich, Germany) and with a viscosity of 30 mPa.sec. About 1 ml of the PVP suspension was transferred into the LORCA device. The diffraction pattern of the RBCs was recorded at increasing shear stresses at a temperature range of 36.7°C – 37°C. The deformability of the RBCs, which is expressed by the elongation index (EI), was determined by the LORCA from the size of the vertical (L) and horizontal (W) axes of the diffraction pattern according to the formula: EI = (L-W) / (L+W). An increased EI at a given shear stress indicates greater RBC deformability. The deformability at two shear stress values were examined more closely; the deformability at a shear stress of 3.9 Pa, reflecting the rigidity of the cell membrane, and the maximal deformability at shear stress of 50 Pa.

Chapter 7
104
Assessment of RBC survival by chromium-51 labeling Fresh, refrigerated stored and thawed deglycerolized rat RBC suspensions with a Hct of 30 -35 % were labeled with 51CR according to standard protocols.31 The in vitro label decay was determined by measuring the radioactivity of 51CR labeled fresh RBCs stored for up to one week at 4°C. The posttransfusion survival per storage group was determined by infusing five rats intravenously with each 0.5 ml of 3.0 µCi/ml 51CR labeled RBC solution. The 51CR radioactivity was measured in blood samples obtained from the rat tail vein after 15 and 20 minutes, 24- and 48 hours and 1 week after infusion with a gamma counter (Compugamma 1282 CS; LKB Wallac, Finland). The survival of RBCs after infusion, was determined by the 51CR single-label method after correction for label decay.32 Statistics Statistical analysis was performed using statistical software (SPSS, version 16.0, SPSS Inc., Chicago, IL). Data were tested for normality with the Kolmogorov-Smirnov goodness-of-fit test. For variables used in the in vitro tests, a repeated measure analysis of variance was performed to identify subject by storage group profiles. Post-hoc multiple comparisons were performed to quantify differences between the storage groups using paired t-tests. For variables used in the in vivo test, a one way analysis of variance was performed to compare groups. Post-hoc multiple comparisons were performed to quantify differences between the groups using a Turkey HSD test. For all post-hoc tests a Bonferroni correction was applied to correct for multiple comparisons. Differences are considered to be significant with a two-tailed p value of less than 0.05. Results are presented as means ± SD. 7.3. Results
In vitro measurement of human RBCs After glycerolization the hemolysis for Bio-20, Combi-20 and Bio-40 cryopreserved RBCs were respectively 0.28 ± 0.1 %; 0.29 ± 0.1% and 0.41 ± 0.1%. The hemolysis was more pronounced when a higher glycerol concentration was used (p < 0.01). After thaw the hemolysis for Bio-20, Combi-20 and Bio-40 cryopreserved RBCs were respectively 0.96 ± 0.5 %; 0.77 ± 0.2 % and 1.05 ± 0.1 % and did not significantly differ from each other. After deglycerolization (Table 7.2) the lowest hemolysis was observed with protocol Combi-20
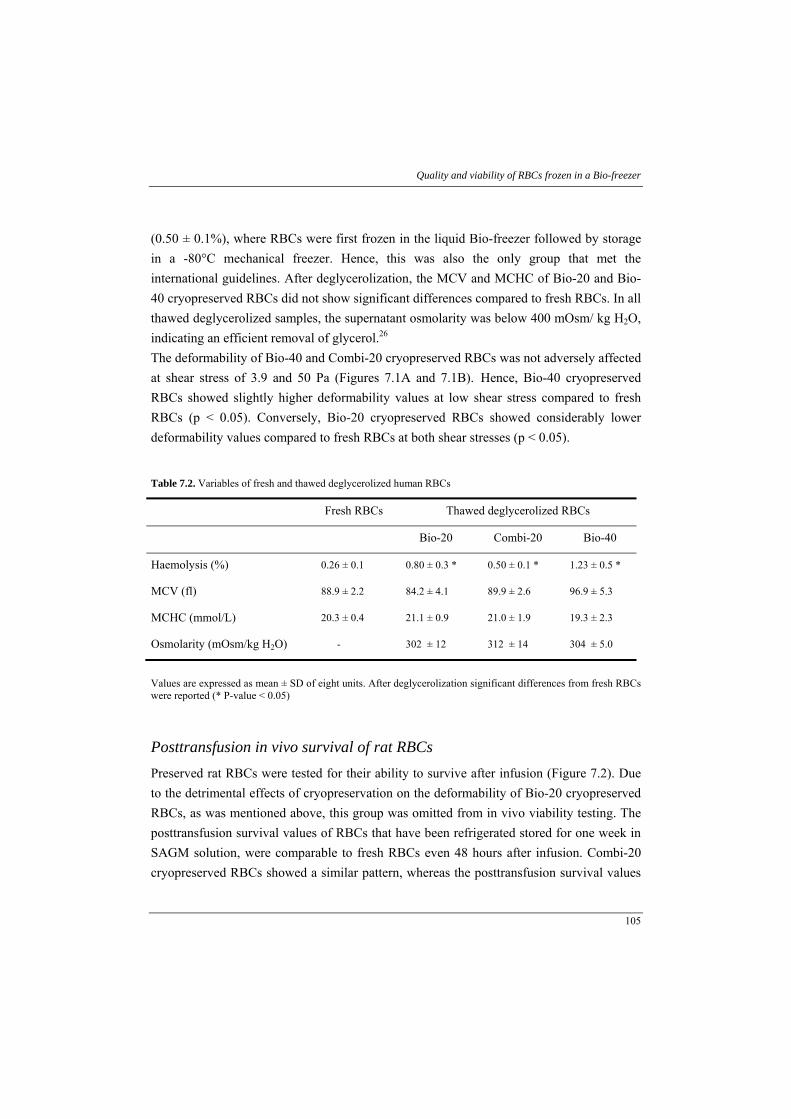
Quality and viability of RBCs frozen in a Bio-freezer
105
(0.50 ± 0.1%), where RBCs were first frozen in the liquid Bio-freezer followed by storage in a -80°C mechanical freezer. Hence, this was also the only group that met the international guidelines. After deglycerolization, the MCV and MCHC of Bio-20 and Bio-40 cryopreserved RBCs did not show significant differences compared to fresh RBCs. In all thawed deglycerolized samples, the supernatant osmolarity was below 400 mOsm/ kg H2O, indicating an efficient removal of glycerol.26 The deformability of Bio-40 and Combi-20 cryopreserved RBCs was not adversely affected at shear stress of 3.9 and 50 Pa (Figures 7.1A and 7.1B). Hence, Bio-40 cryopreserved RBCs showed slightly higher deformability values at low shear stress compared to fresh RBCs (p < 0.05). Conversely, Bio-20 cryopreserved RBCs showed considerably lower deformability values compared to fresh RBCs at both shear stresses (p < 0.05).
Table 7.2. Variables of fresh and thawed deglycerolized human RBCs
Fresh RBCs Thawed deglycerolized RBCs
Bio-20 Combi-20 Bio-40
Haemolysis (%) 0.26 ± 0.1 0.80 ± 0.3 * 0.50 ± 0.1 * 1.23 ± 0.5 *
MCV (fl) 88.9 ± 2.2 84.2 ± 4.1 89.9 ± 2.6 96.9 ± 5.3
MCHC (mmol/L) 20.3 ± 0.4 21.1 ± 0.9 21.0 ± 1.9 19.3 ± 2.3
Osmolarity (mOsm/kg H2O) - 302 ± 12 312 ± 14 304 ± 5.0
Values are expressed as mean ± SD of eight units. After deglycerolization significant differences from fresh RBCs were reported (* P-value < 0.05)
Posttransfusion in vivo survival of rat RBCs Preserved rat RBCs were tested for their ability to survive after infusion (Figure 7.2). Due to the detrimental effects of cryopreservation on the deformability of Bio-20 cryopreserved RBCs, as was mentioned above, this group was omitted from in vivo viability testing. The posttransfusion survival values of RBCs that have been refrigerated stored for one week in SAGM solution, were comparable to fresh RBCs even 48 hours after infusion. Combi-20 cryopreserved RBCs showed a similar pattern, whereas the posttransfusion survival values
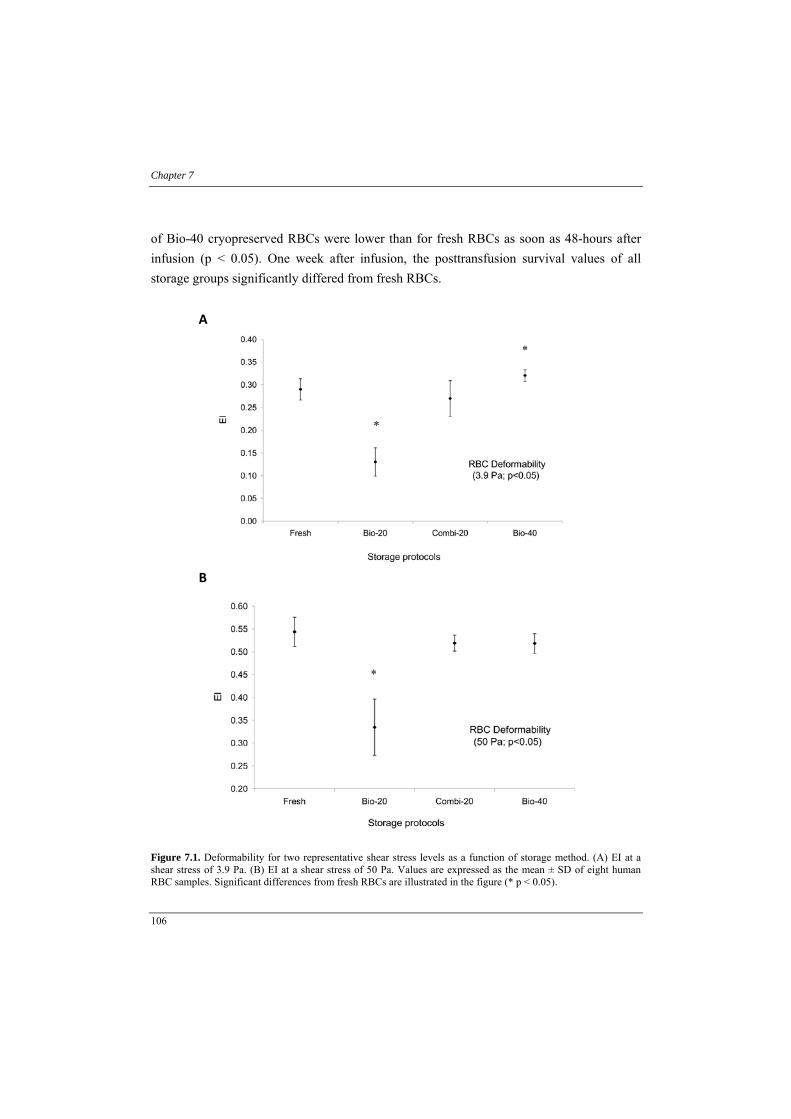
Chapter 7
106
of Bio-40 cryopreserved RBCs were lower than for fresh RBCs as soon as 48-hours after infusion (p < 0.05). One week after infusion, the posttransfusion survival values of all storage groups significantly differed from fresh RBCs.
A
B
Figure 7.1. Deformability for two representative shear stress levels as a function of storage method. (A) EI at a shear stress of 3.9 Pa. (B) EI at a shear stress of 50 Pa. Values are expressed as the mean ± SD of eight human RBC samples. Significant differences from fresh RBCs are illustrated in the figure (* p < 0.05).

Quality and viability of RBCs frozen in a Bio-freezer
107
Figure 7.2. The posttransfusion survival (%) of preserved rat RBCs using the 51CR single-label procedure. Survival percentages are expressed as the mean ± SD in five inbred rats. Significant changes are illustrated in the figure (p < 0.05); * significantly different from fresh RBCs 48 hours after infusion, † significantly different from fresh RBCs 168 hours after infusion.
7.4. Discussion During freezing and thawing, RBCs undergo alterations which may hamper their ability to survive and function after transfusion. Cellular damage of HGM cryopreserved RBCs has been attributed to cellular dehydration and extra-cellular ice formation, as well as to osmotic stress obtained during the glycerolization and deglycerolization process.13,33-35 New freezing techniques that improve the efficiency of the freeze-thaw-wash process could make cryopreserved RBCs more applicable for routine clinical usage. In this study, the quality and functionality of RBCs cryopreserved in a new liquid Bio-freezer were investigated. Addition and removal of glycerol can be detrimental to the RBCs. In our study, glycerolization with 20% glycerol maintained the integrity of the RBCs, whereas usage of

Chapter 7
108
40% glycerol slightly increased the hemolysis. The freezing process per se, did not result in much cellular losses, since after thaw the hemolysis in the freezing protocols was only slightly increased. After deglycerolization, low hemolysis was observed with RBCs which were cryopreserved with 20% glycerol in the liquid Bio-freezer. Yet, only the RBCs that were frozen in the liquid Bio-freezer and stored at -80°C (Combi-20), demonstrated hemolysis values that met international guidelines (e.g. 0.8% in Europe and 1% in the United States).1,2 Even though we observed low hemolysis values after thaw, our preservation methods could have adversely affected the hemolysis. Due to the small sample size, RBCs mixtures were frozen in syringes instead of PVC bags. It has been demonstrated that the material of the freezing container can influence the hemolysis post-thaw.36 Changing to PVC bags for human RBCs could possibly further reduce hemolysis due to the more rapid heat exchange and or the reduction in mechanical force experienced during freezing and thawing. The ability of RBCs to deform is an important determinant of the flow behavior of blood. RBC deformability depends on the viscosity of the cytoplasm, which is reflected by the MCHC, as well as on the overall cell shape and the viscoelastic properties of the cytoskeleton.28 Long-term preservation of RBCs with 20% glycerol in the liquid Bio-freezer considerably lowered the deformability compared to fresh and the other frozen storage groups. Since the MCV and MCHC of these RBCs were not significantly different, alterations in the RBC cytoskeleton were presumably responsible for the observed changes. It is therefore likely that the lower deformability of these RBCs will not be reversed upon infusion and subsequently results in removal from the circulation. Altogether, our results thus indicated that a concentration of 20% glycerol was not sufficient to protect the flexibility of RBCs cryopreserved at -25°C, despite the rapid heat exchange of the liquid Bio-freezer. Either lowering the storage temperature (Combi-20) or raising the glycerol concentration (Bio-40), maintained the deformability of RBCs cryopreserved in the liquid Bio-freezer. Hence, it appeared that at -25°C, the detrimental effects become more evident at lower glycerol concentrations. The slightly higher deformability of RBCs cryopreserved with 40% glycerol in the liquid Bio-freezer versus fresh RBCs, was only present at a shear stress of 3.9 Pa. It is likely that the enhanced MCV of some RBC samples may have counteracted this difference at a shear stress of 50 Pa. Particularly, since the RBC deformability in the high shear stress regions are more influenced by changes in cell volume.

Quality and viability of RBCs frozen in a Bio-freezer
109
The survival of RBCs after infusion, depends on the morphological and metabolic status of the RBC, as well as on the protein and lipid composition of the RBC membrane.37-43 In the present study rat RBCs, which have been refrigerated stored for one week in SAGM solution, demonstrated high 24-hour posttransfusion survival which was consistent with earlier findings.8,9 In addition, the posttransfusion survival of the latter group was even comparable to fresh RBCs 48-hours after infusion. However, one week after infusion, the posttransfusion survival of refrigerated stored rat RBCs was lower than that of fresh RBCs. Although one week of refrigerated storage is often not considered to be detrimental to human RBCs, the latter results indicate that the more accelerated aging process of rat RBCs, adversely affect the in vivo RBC viability on the long-term.8,9 Frozen storage of rat RBCs with 40% glycerol (Bio-40) in the liquid Bio-freezer, demonstrated posttransfusion survival values which were below the international limit of 75%. Yet, the deformability of human RBCs was not adversely affected with the Bio-40 protocol. Since changes in morphological and metabolic state of the RBCs would have affected the RBC deformability,44-46 we hypothesize that the low posttransfusion survival values could be the result of alterations in the protein and lipid composition of the RBC membrane. Combi-20 cryopreserved RBCs demonstrated posttransfusion survival values which were comparable to fresh and refrigerated stored RBCs even 48-hours after infusion. This indicates that the rapid heat exchange of the liquid Bio-freezer may be favorable for freezing the RBCs. However, in order to maintain a high degree of viability, storage of RBCs at lower temperatures was required. The use of only 20% glycerol, could be beneficial in reducing the osmotic stress associated with the addition and removal of glycerol which result in cellular losses.12,13,35 One week after infusion, the survival of all preserved RBCs was lower than that of fresh RBCs. Although current guidelines focus on the 24-hour posttransfusion RBC survival, maintaining a high in vivo survival for longer time periods may be beneficial to the patient outcome. This is a first study, which evaluates the feasibility of the liquid Bio-freezer for human RBC preservation. Nevertheless, further studies are needed to establish if Combi-20 cryopreserved RBCs, further satisfy international guidelines and demonstrate acceptable quality during prolonged post-thaw storage. In transfusion medicine, cryopreservation enables RBCs to be stockpiled for years.47-49 Nevertheless, current freezing techniques still limit the routine clinical usage of cryopreserved RBCs. Our data demonstrated that the liquid Bio-freezer alone was not

Chapter 7
110
sufficient to preserve the RBC deformability or maintain high in vivo RBC survival. However, usage of the liquid Bio-freezer in combination with the -80°C mechanical freezer, enabled preservation of RBCs with low glycerol concentrations, while maintaining the RBC fragility, deformability and high 48-hour posttransfusion survival values. The liquid Bio-freezer could become a valuable tool for the cryopreservation of RBCs.
Acknowledgements We gratefully thank Supachill and Haemoscan for their financial support. Furthermore, we thank Michel de Vries from the department of Nuclear Medicine and Molecular Imaging at the University Medical Center Groningen for his assistance with the 51CR-labeling procedures.

Quality and viability of RBCs frozen in a Bio-freezer
111
References 1. Council of Europe. Guide to the preparation, use and quality assurance of blood components, 14th edition.
Strasbourg: Council of Europe publisher; 2008.
2. Kakaiya R, Aronson CA, Julleis J. Whole blood collection and component processing, AABB Technical Manual 16th ed. Bethesda, MD.: AABB Press; 2008.
3. Lelkens CC, Koning JG, de Korte D, Floot IB, Noorman F. Experiences with frozen blood products in the Netherlands military. Transfus Apher Sci. 2006, vol.34: 289-298.
4. Peyrard T, Pham BN, Le Pennec PY, Rouger P. Transfusion of rare cryopreserved red blood cell units stored at -80 °C: the French experience. Immunohematology. 2009, vol.25: 13-17.
5. Zimrin AB, Hess JR. Planning for pandemic influenza: effect of a pandemic on the supply and demand for blood products in the United States. Transfusion. 2007, vol.47: 1071-1079.
6. Tian F, Wang J, Huang Y, Guo X, Yun Z, Yang T, Nelson K, Schreiber GB, Ness P, Shan H. Psychological and behavioral impacts of the 2008 China earthquake on blood donors. Vox Sang. 2010, vol.99: 142-148.
7. Schmidt PJ. Blood and disaster-supply and demand. N Engl J Med. 2002, vol.346: 617-620.
8. d'Almeida MS, Jagger J, Duggan M, White M, Ellis C, Chin-Yee IH. A comparison of biochemical and functional alterations of rat and human erythrocytes stored in CPDA-1 for 29 days: implications for animal models of transfusion. Transfus Med. 2000, vol.10: 291-303.
9. Gilson CR, Kraus TS, Hod EA, Hendrickson JE, Spitalnik SL, Hillyer CD, Shaz BH, Zimring JC. A novel mouse model of red blood cell storage and posttransfusion in vivo survival. Transfusion. 2009, vol.49: 1546-1553.
10. Hall CE, Nash JB, Hall O. Erythrocyte survival and blood volume in the rat as determined by labeling the red cells with Cr 51. Am J Physiol. 1957, vol.190: 327-329.
11. Hod EA, Arinsburg SA, Francis RO, Hendrickson JE, Zimring JC, Spitalnik SL. Use of mouse models to study the mechanisms and consequences of RBC clearance. Vox Sang. 2010, vol.99: 99-111.
12. Valeri CR, Ragno G, Pivacek LE, Srey R, Hess JR, Lippert LE, Mettille F, Fahie R, O'Neill EM, Szymanski IO. A multicenter study of in vitro and in vivo values in human RBCs frozen with 40-percent (wt/vol) glycerol and stored after deglycerolization for 15 days at 4 °C in AS-3: assessment of RBC processing in the ACP 215. Transfusion. 2001, vol.41: 933-939.
13. Lelkens CC, Noorman F, Koning JG, Truijens de Lange R, Stekkinger PS, Bakker JC, Lagerberg JW, Brand A, Verhoeven AJ. Stability after thawing of RBCs frozen with the high- and low-glycerol method. Transfusion. 2003, vol.43: 157-164.
14. Hess JR. Red cell freezing and its impact on the supply chain. Transfus Med. 2004, vol.14: 1-8.
15. Goolsby HA, Blanton JR, Cotter PZ, Prien SD. Preliminary trial: motility comparisons of a unique freezing technology (UFT) to liquid nitrogen mist methodology for cryopreservation of porcine spermatozoa. Reprod Domest Anim. 2004, vol.39: 328-332.
16. Cotter PZ, Goolsby HA, Prien SD. Preliminary evaluation of a unique freezing technology for bovine spermatozoa cryopreservation. Reprod Domest Anim. 2005, vol.40: 98-99.

Chapter 7
112
17. Rinfret AP. Factors affecting the erythrocyte during rapid freezing and thawing. Ann NY Acad Sci. 1960, vol.85: 576-594.
18. Fahy GM, Levy DI, Ali SE. Some emerging principles underlying the physical properties, biological actions, and utility of vitrification solutions. Cryobiology. 1987, vol.24: 196-213.
19. Fahy GM. Cryoprotectant toxicity neutralization. Cryobiology. 2010, vol.60: S45-53.
20. Simchon S, Jan KM, Chien S. Influence of reduced red cell deformability on regional blood flow. Am J Physiol. 1987, vol.253: H898-H903.
21. Marossy A, Svorc P, Kron I, Gresova S. Hemorheology and circulation. Clin Hemorheol Microcirc. 2009, vol.42: 239-258.
22. Baskurt OK. The role of spleen in suppressing the rheological alterations in circulating blood. Clin Hemorheol Microcirc. 1999, vol.20: 181-188.
23. Parthasarathi K, Lipowsky HH. Capillary recruitment in response to tissue hypoxia and its dependence on red blood cell deformability. Am J Physiol. 1999, vol.277: H2145-H2157.
24. Bohonek M, Petras M, Turek I, Urbanova J, Hradek T, Chmatal P, Staroprazska V, Kostirova J, Horcickova D, Duchkova S, et al. Quality evaluation of frozen apheresis red blood cell storage with 21-day postthaw storage in additive solution 3 and saline-adenine-glucose-mannitol: biochemical and chromium-51 recovery measures. Transfusion. 2010, vol.50: 1007-1013.
25. Valeri CR. Simplification of the methods for adding and removing glycerol during freeze-preservation of human red blood cells with the high or low glycerol methods: biochemical modification prior to freezing. Transfusion. 1975, vol.15: 195-218.
26. Valeri C.R, Ragno G. Use of supernatant osmolality and supernatant refraction to assess the glycerol concentration in glycerolized and deglycerolized previously frozen RBC. Transfus Apher Sci. 2007, vol.36: 133-137.
27. Shiga T, Maeda N, Kon K. Erythrocyte rheology. Crit Rev Oncol Hematol. 1990, vol.10: 9-48.
28. Mohandas N, Gallagher PG. Red cell membrane: past, present, and future. Blood. 2008, vol.112: 3939-3948.
29. Harboe M. A method for determination of hemoglobin in plasma by near-ultraviolet spectrophotometry. Scand J Clin Lab Invest. 1959, vol.11: 66-70.
30. Hardeman MR, Ince C. Clinical potential of in vitro measured red cell deformability, a myth? Clin Hemorheol Microcirc. 1999, vol.21: 277-284.
31. Commissie kwaliteitsbevordering van de Nederlandse vereniging voor nucleaire geneeskunde. Aanbevelingen nucleaire geneeskunde, 4th ed. Delft: Eburon; 2000.
32. Moroff G, Sohmer PR, Button LN. Proposed standardization of methods for determining the 24-hour survival of stored red cells. Transfusion. 1984, vol.24: 109-114.
33. Scott KL, Lecak J, Acker JP. Biopreservation of red blood cells: past, present, and future. Transfus Med Rev. 2005, vol.19: 127-142.
34. Pegg DE. The relevance of ice crystal formation for the cryopreservation of tissues and organs. Cryobiology. 2010, vol.60: S36-44.

Quality and viability of RBCs frozen in a Bio-freezer
113
35. Song YS, Moon S, Hulli L, Hasan SK, Kayaalp E, Demirci U. Microfluidics for cryopreservation. Lab Chip. 2009, vol.9: 1874-1881.
36. Hornblower M, Meryman HT. Influence of the container material on the hemolysis of glycerolized red cells after freezing and thawing. Cryobiology. 1974, vol.11: 317-323.
37. Schroit AJ, Madsen JW, Tanaka Y. In vivo recognition and clearance of red blood cells containing phosphatidylserine in their plasma membranes. J Biol Chem. 1985, vol.260: 5131-5138.
38. Jain SK, Mohandas N, Clark MR, Shohet SB. The effect of malonyldialdehyde, a product of lipid peroxidation, on the deformability, dehydration and 51Cr-survival of erythrocytes. Br J Haematol. 1983, vol.53: 247-55.
39. Hogman CF, de Verdier CH, Ericson A, Hedlund K, Sandhagen B. Studies on the mechanism of human red cell loss of viability during storage at +4 °C in vitro. I. Cell shape and total adenylate concentration as determinant factors for posttransfusion survival. Vox Sang. 1985, vol.48: 257-268.
40. Heaton WA. Evaluation of posttransfusion recovery and survival of transfused red cells. Transfus Med Rev. 1992, vol.6: 153-169.
41. Head DJ, Lee ZE, Swallah MM, Avent ND. Ligation of CD47 mediates phosphatidylserine expression on erythrocytes and a concomitant loss of viability in vitro. Br J Haematol. 2005, vol. 130: 788-790.
42. Bessos H, Seghatchian J. Red cell storage lesion: the potential impact of storage-induced CD47 decline on immunomodulation and the survival of leucofiltered red cells. Transfus Apher Sci. 2005, vol.32: 227-232.
43. Aminoff D, Anderson J, Dabich L, Gathmann WD. Sialic acid content of erythrocytes in normal individuals and patients with certain hematologic disorders. Am J Hematol. 1980, vol.9: 381-389.
44. Chien S. Red cell deformability and its relevance to blood flow. Annu Rev Physiol. 1987, vol.49:177-192.
45. Gov NS. Active elastic network: cytoskeleton of the red blood cell. Phys Rev E Stat Nonlin Soft Matter Phys. 2007, vol.75: 011921.
46. Park Y, Best CA, Auth T, Gov NS, Safran SA, Popescu G, Suresh S, Feld MS. Metabolic remodeling of the human red blood cell membrane. Proc Natl Acad Sci. 2010, vol.107: 1289-1294.
47. Valeri CR, Ragno G, Pivacek LE, Cassidy GP, Srey R, Hansson-Wicher M, Leavy ME. An experiment with glycerol-frozen red blood cells stored at -80 °C for up to 37 years. Vox Sang. 2000, vol.79: 168-174.
48. Valeri CR, Srey R, Tilahun D, Ragno G. The in vitro quality of red blood cells frozen with 40 percent (wt/vol) glycerol at -80 °C for 14 years, deglycerolized with the Haemonetics ACP 215, and stored at 4 °C in additive solution-1 or additive solution-3 for up to 3 weeks. Transfusion. 2004, vol.44:990-995.
49. Lecak J, Scott K, Young C, Hannon J, Acker JP. Evaluation of red blood cells stored at -80 °C in excess of 10 years. Transfusion. 2004, vol.44: 1306-1313


Chapter 8
General discussion

Chapter 8
116
8.1. General Discussion The ability to store RBCs outside the body has resulted in a worldwide network of blood banks that collect blood and distributed it when needed. RBCs are frequently life-saving in many circumstances. Yet, over the last few years concerns regarding the quality of long-term refrigerated stored RBCs have arisen. The RBC rheologic properties for example, have been repeatedly documented to be impaired during prolonged refrigerated storage. Nevertheless, RBC storage conditions have continuously improved and the majority of these rheologic studies were either outdated or not adequately corrected for known confounding factors. New studies that demonstrated the RBC rheologic properties during contemporary refrigerated storage were therefore desirable. A well-recognized problem of refrigerated storage is that due to the limited shelf life, RBCs continuously need to be replenished to balance the inventory. In an effort to circumvent storage induced lesions and thus to prolong the shelf life of RBCs, cryopreservation seems to be a promising method to preserve RBC. Particularly, since in recent years research has focused on optimizing the use of cryopreserved RBCs for clinical practice. This thesis aimed to gain a better understanding of the RBC quality from a rheologic perspective in transfusion medicine, as well as to explore the utilization of cryopreservation for long-term storage of RBC.
8.2. RBC rheology The ability of RBCs to deform and aggregate are key determinants of the flow behavior of blood. The deformability enables RBCs to adapt their size to minimize flow resistance as well as to pass narrow capillaries. In contrast, RBC aggregation increases the flow resistance by enhancing whole blood viscosity at low shear rate, albeit that the physiological role of RBC aggregation is still elusive.1 A better understanding of RBC aggregation my help to guide hemorheologic studies and therapeutic approaches in pathophysiology. In general, considerable variations in RBC aggregation tendencies exists between healthy individuals.2 Interestingly, RBCs also exhibit different aggregation tendencies among mammalian species.3 Yet, these interspecies differences are not solely ascribed to variations in plasma composition because differences continue to exist even when RBC are washed and re-suspended in a standard medium of

General discussion
117
high molecular weight polymers. Replacement of plasma proteins by standard polymers to induce RBC aggregation may help to unravel the fundamentals of the aggregation process. At present, a depletion and a bridging theory have been proposed to explain the RBC aggregation mechanism induced by high molecular weight polymers. Neu et al.4-7support the depletion theory underlying 500-kDa dextran induced RBC aggregation. In contrast, our data support the bridging theory as the mechanism underlying 200-kDa HES induced RBC aggregation, which is in line with the formation of stronger and larger RBC aggregates with HES polymers of increasing molecular weight.8 Despite these differences in aggregation mechanisms, the 200-kDa HES and the 500-Kda dextran polymer solutions are useful to manifest cellular induced aggregation differences between healthy individuals and T1DM patients. Disturbed rheologic properties can be detrimental to the flow dynamics and subsequently the oxygen delivery in the microcirculation. Reduced deformation and enhanced aggregation of RBCs have been observed in a variety of chronic diseases. However, in some acute situations, such as during damage control of severely bleeding trauma patients, enhanced RBC aggregation could also play a pivotal role. Presumably, adoption of a liberal FFP to RBC ratio could promote RBC aggregation. This could be disadvantageous in trauma patients in which the RBC rheology is already compromised. In this regard, the LORCA (R&R Mechatronics, Zwaag, the Netherlands) could be useful to demonstrate RBC aggregation tendencies of trauma patients who undergo damage control resuscitation. In addition, analysis of the RBC cellular factors that affect the RBC aggregability3 could further improve our understanding of altered aggregation tendencies in disease.
8.3. Refrigerated storage of RBCs The rheologic properties of prolonged refrigerated stored RBCs have been repeatedly documented to be impaired (i.e. enhanced aggregation, reduced deformation and elevated endothelial cell interaction).9-15 Yet, these studies analyzed the rheologic properties of non-leukofiltrated refrigerated stored RBCs, which is not representative for European blood banking practice. More recently, it was demonstrated that leukoreduction minimizes storage associated adhesiveness of RBCs to endothelial cells.16-18 Our data showed that although the aggregability of leukoreduced refrigerated stored RBCs was slightly altered over time, an enhanced RBC aggregability was clearly not observed. In addition, we found that the

Chapter 8
118
reduction in RBC deformability, which was only observed at high shear stress, was still within physiological ranges. Previously, it was demonstrated that the phosphorylation of cytoskeleton protein 4.1, which was powered by ATP, played an important role in regulating the RBC deformability.19,20 Although we demonstrated that the RBC ATP content became progressively depleted during refrigerated storage, we did not observe a reduction in the RBC deformability at low shear stress. This could suggest that DEHP leaching from the PVC storage bag has counteracted the storage induced deterioration in flexibility21,22 or that the total adenine phosphate pool, which consist of ATP, ADP and AMP, has not depleted yet.23,24 We also consider that the sphericity of the RBCs, induced by the progressive ATP depletion,25,26 could play a fundamental in role in our observations. Particularly, because at high shear stress the cell volume becomes a limiting factor for the RBCs to deform and because RBC swelling suppresses cell contact and subsequently aggregation formation.27,28 Due to the high osmotic content of plasma, RBC swelling is most likely to be reversed after infusion.29 Based on these and recent findings we concluded that the rheologic properties of leukoreduced refrigerated stored RBC are not likely to contribute to adverse clinical outcomes after transfusion. ATP does not only play a role in the deformability of the RBCs, it is fundamental for the overall functioning of the cell. We observed considerable low ATP values after 5 weeks of refrigerated RBC storage. Although a low ATP content can be replenished within 48 hours following infusion of RBCs,30 in some patient groups, such as critical ill, an immediate restoration of the oxygen delivery may be desired. Recently, Spinella et al proposed that transfusion of 1 to 2 RBC units of increased storage age was sufficient to compromise clinical outcome in trauma patients.31 Nonetheless, this retrospective study was further limited by the lack of discrimination between leukoreduced and non-leukoreduced RBCs within each storage group. Despite these limitations, it is possible that there is a storage limit beyond which RBC transfusion may be less beneficial or can even compromise the clinical outcome of critical ill patients.32-34 Especially, if a low ATP content hinders the RBCs to properly function. Yet the clinical consequences of prolonged stored RBCs are poorly understood and well-designed randomized controlled trials are currently lacking.35-42 Implementation of procedures that improve the RBC quality rather than extending the maximal storage duration of refrigerated RBC storage are therefore desirable.43 In this regard, a new storage solution (i.e PAGGGM) was developed that maintains high ATP and 2,3-DPG content of RBCs and prevents microvesicle formation during 5 weeks of

General discussion
119
refrigerated storage.44,45 Other quality determinants, including the RBC rheologic properties will still need to be determined. Alternatively, storage of RBCs at ultra-low subzero temperatures halts the biochemical processes in the cell which may circumvent storage induced lesions. From this perspective, cryopreservation could be a promising approach to prolong the lifespan of RBCs.
8.4. Cryopreservation of RBCs Currently, the HGM is the most widely applied freezing method for long-term storage of RBCs. Cryopreserved RBCs have to meet certain guidelines,46,47 yet as in the case of refrigerated stored RBCs, these guidelines do not reflect the RBCs ability to function after infusion. Over the years more knowledge about the quality of HGM cryopreserved RBCs has become available. Data about the rheologic properties of cryopreserved RBCs were however scarce. In this regard, we investigated the deformability and aggregability of HGM cryopreserved RBCs that were post-thaw stored in SAGM additive solution. In our study the deformability of cryopreserved RBCs was higher than that of fresh RBCs but similar to prolonged refrigerated stored RBCs. Fresh RBCs were not leukofiltrated as opposed to the stored RBCs. Although the presence of leukocytes could have been detrimental to the flexibility of RBCs, it is unlikely that this caused the lower deformability of fresh RBCs. Especially, because their contact time with fresh RBCs was only minimal. We hypothesized that DEHP leaching from the PVC storage bag has enhanced the flexibility of the RBCs, explaining the improved deformability of both refrigerated stored and cryopreserved RBCs over that of fresh RBCs. Unfortunately, we did not have the infrastructure to study DEPH leaching per se. Surprisingly, the deformability of cryopreserved and long-term refrigerated stored RBCs was similar despite their differences in MCV. Clearly, additional studies are warranted to elucidate the precise role of RBC volume in limiting the flexibility of the RBCs. We also observed a higher osmotic fragility of cryopreserved RBCs that were post-thaw stored in SAGM additive solution, compared to fresh and refrigerated stored RBCs. The increasing cell volume could have made the RBCs more prone to hemolysis upon concomitant swelling. Especially, since no alterations in osmotic fragility have been observed with cryopreserved RBCs that were post-thaw stored in AS-3 or CPDA-1

Chapter 8
120
solution.48 Presumably, the absence of citrate, an impermeable anion, in the SAGM additive solution allows for more substantial cell swelling.49,50 We demonstrated that cryopreserved RBCs were less able to aggregate then fresh and refrigerated stored RBCs. This was not surprisingly in the light of the enhanced RBC volume after deglycerolization. Although enhanced RBC aggregation has been commonly observed in various diseases, less is known about the effect of diminished RBC aggregation. Previously, it has been hypothesized that low RBC aggregation is a pathogenic co-factor in endothelial activation during hemodilution,51. However, it was recognized that hemodilution per se induced a hypercoagulable state.52 One could argue that infusion of RBCs with low aggregation tendencies may be less beneficial in directing leukocytes and possibly platelets to a damaged vessel wall.53 On the other hand, infusion of RBCs with lower aggregation tendencies could also be beneficial to patients with microcirculatory disorders.54,55 Nevertheless, the high 24-hour in vivo survival of cryopreserved RBCs indirectly suggests that RBC swelling and subsequently the lower aggregability tendencies of cryopreserved RBCs are of less physiologic importance.56 Infusion of cryopreserved RBCs has potential advantages over that of refrigerated stored RBCs. We demonstrated that the ATP content of cryopreserved RBCs was similar to that of fresh RBCs. Indicating that cryopreserved RBCs could have superior oxygen delivering capacity compared to that of prolonged refrigerated stored RBCs.57 Also, due to the deglycerolization washing procedure less biological active substances will be present in the RBC unit that may cause transfusion related reactions in the recipient.58 In this regard, cryopreserved RBCs could be useful in situations where refrigerated stored RBCs are less desired. This could be the case for chronically transfused patients. In addition, cryopreserved RBCs could also be useful in order to minimize occurrences of TRALI and SIRS. Nevertheless, cryopreserved RBCs are primarily used for controlling an inventory of RBCs with rare blood phenotypes and for military conflicts. Occasionally, cryopreserved RBCs are used during temporarily blood shortages. Cryopreserved RBCs are more expensive, time-consuming and less efficient than refrigerated stored RBCs. However, among the years cryopreserved RBCs have become more utilizable. Implementation of the automated ACP-215 device (Haemonetics, Braintree, MA) for example has simplified the washing process and prolonged the postthaw storage time of RBCs in AS-3 to 14 days. This development has eased the clinical

General discussion
121
applicability of cryopreserved RBCs particularly, because RBCs could now be thawed and washed in advance. The developments of new techniques that could make cryopreserved RBCs more utilizable for transfusion medicine are still ongoing. Implementation of a liquid Bio-freezer with a rapid heat exchange for example could make cryopreserved RBCs more interesting for clinical usage. Although our results documenting the quality of RBCs cryopreserved in the liquid Bio-freezer demonstrated that the temperature of the Bio-freezer was not sufficient to preserve the RBC deformability or maintain high in vivo RBC survival, combining the bio-freezer with -80°C storage enabled preservation of RBCs with only 20% glycerol while maintaining good in vitro and in vivo viability. In this context, the lower glycerol content could be favorable since it potentially reduces the osmotic stress during the glycerolization and deglycerolization process that causes cellular losses. Additional studies will be needed to determine the RBC quality during prolonged post-thaw storage.
8.5. Conclusion Refrigerated storage induces a variety of cellular changes that could hamper the RBCs to properly function after infusion. In this regard, the rheologic properties are important quality determinants, because alterations in the rheologic properties can be detrimental to the flow dynamics and subsequently the oxygen delivery in the microcirculation. This thesis demonstrates that the rheologic properties of leukoreduced RBCs were well preserved during refrigerated storage and not likely to contribute to adverse clinical outcome after infusion. Cryopreservation of RBCs has been practiced to counteract storage induced lesions and subsequently to extend the shelf life of preserved RBCs. This thesis furthermore demonstrates that the freeze-thaw-wash process was not detrimental to the aggregability and deformability of HGM cryopreserved RBCs. RBC cryopreservation is still infrequently used. However, ongoing scientific and technological advancement has eased the clinical applicability of cryopreserved RBCs which could extend their utilization in clinical practice. As we progress towards a more aging society the demand for RBC transfusions will inevitably increase.59,60 and cryopreserved RBCs could become a more routinely used blood resource. Especially, since cryopreserved RBCs are available, safe, in compliance with international regulations and guidelines and can be used effectively.61-63

Chapter 8
122
References 1. Meiselman HJ. Red blood cell aggregation: 45 years being curious. Biorheology. 2009, vol.46: 1-19.
2. Rampling MW, Meiselman HJ, Neu B, Baskurt OK. Influence of cell-specific factors on red blood cell aggregation. Biorheology. 2004, vol.41: 91-112.
3. Baskurt OK, Meiselman HJ. Lessons from comparative hemorheology studies. Clin.Hemorheol.Microcirc. 2010, vol.45: 101-108.
4. Neu B, Meiselman HJ. Depletion-mediated red blood cell aggregation in polymer solutions. Biophys.J. 2002, vol.83: 2482-2490.
5. Neu B, Wenby R, Meiselman HJ. Effects of dextran molecular weight on red blood cell aggregation. Biophys.J. 2008, vol.95: 3059-3065.
6. Rad S, Gao J, Meiselman HJ, Baskurt OK, Neu B. Depletion of high molecular weight dextran from the red cell surface measured by particle electrophoresis. Electrophoresis. 2009, vol.30: 450-456.
7. Baskurt OK, Tugral E, Neu B, Meiselman HJ. Particle electrophoresis as a tool to understand the aggregation behavior of red blood cells. Electrophoresis. 2002, vol.23: 2103-2109.
8. Morariu AM, van der Plaats A, van Oeveren W, 'T Hart NA, Leuvenink HG, Graaff R, Ploeg RJ, Rakhorst G. Hyperaggregating effect of hydroxyethyl starch components and University of Wisconsin solution on human red blood cells: a risk of impaired graft perfusion in organ procurement? Transplantation. 2003, vol.76: 37-43.
9. Berezina TL, Zaets SB, Morgan C, Spillert CR, Kamiyama M, Spolarics Z, Deitch EA, Machiedo GW. Influence of storage on red blood cell rheological properties. J.Surg.Res. 2002, vol.102: 6-12.
10. Hovav T, Yedgar S, Manny N, Barshtein G. Alteration of red cell aggregability and shape during blood storage. Transfusion. 1999, vol.39: 277-281.
11. Relevy H, Koshkaryev A, Manny N, Yedgar S, Barshtein G. Blood banking-induced alteration of red blood cell flow properties. Transfusion. 2008, vol.48: 136-146.
12. Izzo P, Manicone A, Spagnuolo A, Lauta VM, Di Pasquale A, Di Monte D. Erythrocytes stored in CPD SAG-mannitol: evaluation of their deformability. Clin.Hemorheol.Microcirc. 1999, vol.21: 335-339.
13. Godin C, Caprani A. Effect of blood storage on erythrocyte/wall interactions: implications for surface charge and rigidity. Eur.Biophys.J. 1997, vol.26: 175-182.
14. Kirkpatrick UJ, Adams RA, Lardi A, McCollum CN. Rheological properties and function of blood cells in stored bank blood and salvaged blood. Br.J.Haematol. 1998 May;101(2):364-368.
15. Nagaprasad V, Singh M. Sequential analysis of the influence of blood storage on aggregation, deformability and shape parameters of erythrocytes. Clin.Hemorheol.Microcirc. 1998, vol. 18: 273-284.
16. Anniss AM, Sparrow RL. Storage duration and white blood cell content of red blood cell (RBC) products increases adhesion of stored RBCs to endothelium under flow conditions. Transfusion. 2006, vol.46: 1561-1567.
17. Luk CS, Gray-Statchuk LA, Cepinkas G, Chin-Yee IH. WBC reduction reduces storage-associated RBC adhesion to human vascular endothelial cells under conditions of continuous flow in vitro. Transfusion. 2003, vol.43: 151-156.

General discussion
123
18. Chin-Yee IH, Gray-Statchuk L, Milkovich S, Ellis CG. Transfusion of stored red blood cells adhere in the rat microvasculature. Transfusion. 2009, vol.49: 2304-2310.
19. Park Y, Best CA, Auth T, Gov NS, Safran SA, Popescu G, Suresh S, Feld MS. Metabolic remodeling of the human red blood cell membrane. Proc.Natl.Acad.Sci. 2010, vol.107: 1289-1294.
20. Betz T, Lenz M, Joanny JF, Sykes C. ATP-dependent mechanics of red blood cells. Proc.Natl.Acad.Sci. 2009, vol.106: 15320-15325.
21. Labow RS, Card RT, Rock G. The effect of the plasticizer di(2-ethylhexyl)phthalate on red cell deformability. Blood. 1987, vol.70: 319-323.
22. Shaz BH, Grima K, Hillyer CD. 2-(Diethylhexyl)phthalate in blood bags: is this a public health issue? Transfusion. 2011, vol.51: 2510-2517.
23. Hogman CF, de Verdier CH, Ericson A, Hedlund K, Sandhagen B. Studies on the mechanism of human red cell loss of viability during storage at 4°C in vitro. I. Cell shape and total adenylate concentration as determinant factors for posttransfusion survival. Vox Sang. 1985, vol.48: 257-268.
24. Ataullakhanov FI, Vitvitsky VM. What determines the intracellular ATP concentration. Biosci.Rep. 2002, vol.22: 501-511.
25. Lang KS, Lang PA, Bauer C, Duranton C, Wieder T, Huber SM, Lang F. Mechanisms of suicidal erythrocyte death. Cell Physiol Biochem. 2005, vol.15: 195-202.
26. Chien S. Red cell deformability and its relevance to blood flow. Annu.Rev.Physiol. 1987, vol.49: 177-192.
27. Cicha I, Suzuki Y, Tateishi N, Maeda N. Changes of RBC aggregation in oxygenation-deoxygenation: pH dependency and cell morphology. Am.J.Physiol Heart Circ.Physiol. 2003, vol.284: H2335-H2342.
28. Reinhart WH, Singh-Marchetti M, Straub PW. The influence of erythrocyte shape on suspension viscosities. Eur.J.Clin.Invest. 1992, vol.22: 38-44.
29. Zehnder L, Schulzki T, Goede JS, Hayes J, Reinhart WH. Erythrocyte storage in hypertonic (SAGM) or isotonic (PAGGSM) conservation medium: influence on cell properties. Vox Sang. 2008, vol.95: 280-287.
30. Valeri CR, Ragno G. The effects of preserved red blood cells on the severe adverse events observed in patients infused with hemoglobin based oxygen carriers. Artif.Cells Blood Substit.Immobil.Biotechnol. 2008, vol.36: 3-18.
31. Spinella PC, Carroll CL, Staff I, Gross R, Mc QJ, Keibel L, Wade CE, Holcomb JB. Duration of red blood cell storage is associated with increased incidence of deep vein thrombosis and in hospital mortality in patients with traumatic injuries. Crit Care. 2009, vol.13: R151.
32. Gauvin F, Spinella PC, Lacroix J, Choker G, Ducruet T, Karam O, Hebert PC, Hutchison JS, Hume HA, Tucci M. Association between length of storage of transfused red blood cells and multiple organ dysfunction syndrome in pediatric intensive care patients. Transfusion. 2010, vol.50: 1902-1913.
33. Pettila V, Westbrook AJ, Nichol AD, Bailey MJ, Wood EM, Syres G, Phillips LE, Street A, French C, Murray L, et al. Age of red blood cells and mortality in the critically ill. Crit Care. 2011, vol.15: R116.
34. Kiraly LN, Underwood S, Differding JA, Schreiber MA. Transfusion of aged packed red blood cells results in decreased tissue oxygenation in critically injured trauma patients. J.Trauma. 2009, vol.67: 29-32.

Chapter 8
124
35. Ho J, Sibbald WJ, Chin-Yee IH. Effects of storage on efficacy of red cell transfusion: when is it not safe? Crit Care Med. 2003, vol.31: S687-S697.
36. Lelubre C, Piagnerelli M, Vincent JL. Association between duration of storage of transfused red blood cells and morbidity and mortality in adult patients: myth or reality? Transfusion. 2009, vol.49: 1384-1394.
37. Zimrin AB, Hess JR. Current issues relating to the transfusion of stored red blood cells. Vox Sang. 2009, vol.96: 93-103.
38. Van de Watering LM, Brand A. Effect of storage of red cells. Transfusion medicine and hemotherapy. 2008, vol.35: 359-367.
39. Vamvakas EC. Meta-analysis of clinical studies of the purported deleterious effects of "old" (versus "fresh") red blood cells: are we at equipoise? Transfusion. 2010, vol.50: 600-610.
40. Edgren G, Kamper-Jorgensen M, Eloranta S, Rostgaard K, Custer B, Ullum H, Murphy EL, Busch MP, Reilly M, Melbye M, et al. Duration of red blood cell storage and survival of transfused patients (CME). Transfusion. 2010, vol.50: 1185-1195.
41. Steiner ME, Stowell C. Does red blood cell storage affect clinical outcome? When in doubt, do the experiment. Transfusion. 2009, vol.49: 1286-1290.
42. Carson JL, Reynolds RC, Klein HG. Bad bad blood? Crit Care Med. 2008, vol.36: 2707-2708.
43. Hogman CF, Meryman HT. Red blood cells intended for transfusion: quality criteria revisited. Transfusion. 2006, vol.46: 137-142.
44. Burger P, Korsten H, de Korte D, Rombout E, van Bruggen R, Verhoeven AJ. An improved red blood cell additive solution maintains 2,3-diphosphoglycerate and adenosine triphosphate levels by an enhancing effect on phosphofructokinase activity during cold storage. Transfusion. 2010, vol.50: 2386-2392
45. De Korte D, Kleine M, Korsten HG, Verhoeven AJ. Prolonged maintenance of 2,3-diphosphoglycerate acid and adenosine triphosphate in red blood cells during storage. Transfusion. 2008, vol.48: 1081-1089.
46. Kakaiya R, Aronson CA, Julleis J. Whole blood collection and component processing, AABB Technical Manual, 16th ed. Bethesda, MD.: AABB Press; 2008.
47. Council of Europe. Guide to the preparation, use and quality assurance of blood components, 14th edition. Strasbourg: Council of Europe publisher; 2008.
48. Lecak J, Scott K, Young C, Hannon J, Acker JP. Evaluation of red blood cells stored at -80°C in excess of 10 years. Transfusion. 2004, vol.44: 1306-1313.
49. Hess JR. An update on solutions for red cell storage. Vox Sang. 2006, vol.91: 13-19.
50. Lagerberg JW, Truijens-de Lange R, de Korte D, Verhoeven AJ. Altered processing of thawed red cells to improve the in vitro quality during postthaw storage at 4°C. Transfusion. 2007, vol.47: 2242-2249.
51. Morariu AM, Maathuis MH, Asgeirsdottir SA, Leuvenink HG, Boonstra PW, van Oeveren W, Ploeg RJ, Molema I, Rakhorst G. Acute isovolemic hemodilution triggers proinflammatory and procoagulatory endothelial activation in vital organs: role of erythrocyte aggregation. Microcirculation. 2006, vol.13: 397-409.
52. Ruttmann TG. Haemodilution enhances coagulation. Br.J.Anaesth. 2002, vol.88: 470-472.

General discussion
125
53. Nash GB, Watts T, Thornton C, Barigou M. Red cell aggregation as a factor influencing margination and adhesion of leukocytes and platelets. Clin.Hemorheol.Microcirc. 2008, vol.39: 303-310.
54. Blaha M, Rencova E, Blaha V, Maly R, Blazek M, Studnicka J, Andrys C, Fatorova I, Filip S, Kasparova M, et al. The importance of rheological parameters in the therapy of microcirculatory disorders. Clin.Hemorheol.Microcirc. 2009, vol.42: 37-46.
55. Kirschkamp T, Schmid-Schonbein H, Weinberger A, Smeets R. Effects of fibrinogen and alpha2-macroglobulin and their apheretic elimination on general blood rheology and rheological characteristics of red blood cell aggregates. Ther.Apher.Dial. 2008, vol.12: 360-367.
56. Bohonek M, Petras M, Turek I, Urbanova J, Hradek T, Chmatal P, Staroprazska V, Kostirova J, Horcickova D, Duchkova S, et al. Quality evaluation of frozen apheresis red blood cell storage with 21-day postthaw storage in additive solution 3 and saline-adenine-glucose-mannitol: biochemical and chromium-51 recovery measures. Transfusion. 2010, vol.50: 1007-1013.
57. Raat NJ, Verhoeven AJ, Mik EG, Gouwerok CW, Verhaar R, Goedhart PT, de Korte D, Ince C. The effect of storage time of human red cells on intestinal microcirculatory oxygenation in a rat isovolemic exchange model. Crit Care Med. 2005, vol.33: 39-45.
58. Sparrow RL. Red blood cell storage and transfusion-related immunomodulation. Blood Transfus. 2010, Suppl 3: S26-S30.
59. Seifried E, Klueter H, Weidmann C, Staudenmaier T, Schrezenmeier H, Henschler R, Greinacher A, Mueller MM. How much blood is needed? Vox Sang. 2011, vol.100: 10-21.
60. Benjamin RJ, Whitaker BI. Boom or bust? Estimating blood demand and supply as the baby boomers age. Transfusion. 2011, vol.51: 670-673.
61. Valeri CR, Ragno G. An approach to prevent the severe adverse events associated with transfusion of FDA-approved blood products. Transfus.Apher.Sci. 2010, vol.42: 223-233.
62. Holovati JL, Hannon JL, Gyongyossy-Issa MI, Acker JP. Blood preservation workshop: new and emerging trends in research and clinical practice. Transfus.Med.Rev. 2009, vol.23: 25-41.
63. Lelkens CC, Koning JG, de Korte D, Floot IB, Noorman F. Experiences with frozen blood products in the Netherlands military. Transfus.Apher.Sci. 2006, vol.34: 289-298.


Summary

Summary
128
RBC transfusion practice has been widely applied in clinical settings to compensate for excessive blood loss as well as to improve the oxygen carrying capacity. Although the infusion of refrigerated stored RBCs has been considered a life-saving practice for years, concerns about the efficacy and safety of prolonged stored RBCs are currently emerging. This is because RBCs biochemical processes are not completely suppressed during refrigerated storage. Consequently, the RBCs undergo various cellular and biochemical changes that could hamper the RBC to adequately function after infusion. Yet, evidence of a significant detrimental clinical effect associated with the infusion of prolonged stored RBCs is still inconclusive. The rheologic properties are important determinants of the quality of RBCs. Particularly because impaired RBC rheologic properties, which are enhanced aggregability, reduced deformability and elevated adherence to endothelial cells, proposes a circulatory risk by hindering adequate tissue perfusion and contributing to ischemia or even infarction in the micro-vascular environment. Chapter 1 reviews ex vivo RBC preservation and emphasizes the importance of the RBC rheologic properties and in particular those that can be measured by the LORCA. Alternatively, the utilization of cryopreservation for long-term storage of RBCs is discussed. The general objectives of this thesis were to gain a better understanding of the RBC quality from a rheologic perspective in transfusion medicine, as well as to explore the utilization of cryopreserved RBCs for routine clinical practice. The first part of this thesis mainly focuses on the RBC aggregation process and the rheologic properties of refrigerated stored RBCs. RBC aggregation has been studied for decades in healthy and diseased subjects. Despite these studies, the underlying mechanism and the physiological role are still elusive. Replacement of plasma proteins by standard polymers to induce RBC aggregation helps to unravel the fundamentals of the aggregation process. In chapter 2 the usefulness of 200-kDa HES polymers to induce RBC aggregation was investigated. The EPM of RBCs in 200-kDa HES solution was measured to get more insight into the RBC aggregation mechanism. The measured EPM values of RBCs in HES solutions were less negative than could be predicted by the suspension viscosity, which supports the bridging theory as the mechanism underlying 200-kDa HES induced RBC aggregation. These findings are in line with previous observations in which stronger and larger RBC aggregates are formed with HES solutions of increasing molecular weight. Furthermore, the LORCA was used to demonstrate aggregation differences between RBCs from healthy individuals and patients with a disease. We demonstrate that like the standard

Summary
129
500-kDa dextran polymers, the 200-kDa HES polymers are able to discriminate cellular induced aggregation differences between RBCs from healthy and T1DM subjects. Altogether, our results demonstrate that 200-kDa HES polymers are useful as a pro-aggregant in RBC rheologic studies. Enhanced RBC aggregation has been observed in a variety of chronic diseases, among which T1DM disease. Yet, RBC aggregation could also play a pivotal role in certain acute situations, such as during damage control resuscitation of severely bleeding trauma patients. Chapter 3 discusses the potential importance of RBC aggregation in supporting hemostasis after blood component infusion in severely bleeding trauma patients. In general, RBCs aggregates exclude leukocytes and possibly platelets from the axial core and direct them towards the vascular wall. This process is essential, since leukocytes and platelets need to get into close contact with the damaged endothelium, in order to exert their function. Adoption of a liberal policy with regard to FFP to RBC infusion could promote RBC aggregation. Although physiological levels of RBC aggregation would be beneficial for supporting hemostasis, promotion of aggregation could be detrimental to patients in which the RBC rheology is already compromised, as was observed in certain trauma states. Future studies with the LORCA will be helpful to elucidate the role of RBC aggregation in damage control resuscitation of severely injured trauma patients. A better insight of the RBC rheologic properties during ex vivo preservation may improve transfusion outcome. Earlier studies have shown that the RBC rheologic properties become impaired as soon as the second week of refrigerated storage. Yet, most of these studies were not representative for modern RBC storage due to the absence of leukofiltration prior to storage. In chapter 4 the RBC rheologic features and other hematologic variables of leukoreduced RBCs were studied during seven weeks of refrigerated blood bank storage. Our data shows, that although the aggregability of leukoreduced refrigerated stored RBCs was slightly altered over time, an enhanced aggregability was clearly not observed. In addition, we found that the reduction in RBC deformability, which was only observed at high shear stress, was still within physiological ranges. We did observe that the ATP content became progressively depleted during refrigerated storage, whereas the MCV, pH and MCHC were affected to a lesser degree. We postulate, that the observed changes in RBC variables during refrigerated storage minimally affected the RBC ability to aggregate and deform, even after prolonged refrigerated storage. Based on these and recent findings,

Summary
130
we conclude that the rheologic properties of leukoreduced RBC units were well preserved during routine blood bank storage. In order to circumvent storage induced lesion and thus to extend the shelf-life of RBCs, cryopreservation of RBCs has become a feasible alternative. In Chapter 5 cryopreservation of RBC and in particular its utilization in modern transfusion practice is discussed. In the past, the clinical applicability of cryopreserved RBCs was hampered by the expensive, less efficient and more time consuming nature of this preservation method. Yet, the subsequent unfamiliarity with regard to the quality of cryopreserved RBCs has further limited clinical usage. In this regard, the rheologic features and various hematologic variables of HGM cryopreserved RBCs were investigated in chapter 6. We showed that the aggregability of cryopreserved RBCs was reduced, compared to fresh and refrigerated stored RBCs. The deformability of stored RBCs was enhanced compared to fresh RBCs, but no significant differences in deformability between cryopreserved and 21- or 35-day refrigerated stored RBCs was observed. We also show that the osmotic fragility, hemolysis, MCV and MCHC of cryopreserved RBCs were altered, compared to fresh and refrigerated stored RBCs, but that the ATP content of cryopreserved RBCs remained close to those of fresh RBCs. We demonstrated that although cryopreserved RBCs were more fragile than refrigerated stored and fresh RBCs, the HGM procedure did not adversely affect the ATP content or the aggregability and deformability of cryopreserved RBCs. Therefore, cryopreserved RBCs could become a more utilized blood resource in clinical settings. To date however, utilization of cryopreserved RBCs is primarily restricted to controlling an inventory in situations where the RBC availability is limited or unpredictable. Such is the case for storage of RBC with rare blood groups or for usage in military conflicts. In this regard, the feasibility of a new Bio-freezer was investigated in chapter 7, as a way to improve the clinical applicability of cryopreserved RBCs. Human and rat RBCs were preserved with different glycerol concentrations in the Bio-freezer at -25°C or in combination with storage at -80°C in a mechanical freezer. Cryopreserved human RBCs were tested in vitro for deformability and various hematologic variables. The posttransfusion survival was determined in rats with chromium-51 labeled preserved rat RBCs. Our data demonstrated that the liquid Bio-freezer alone was not sufficient to preserve the RBC deformability or to maintain high in vivo RBC survival. However, usage of the liquid Bio-freezer in combination with the -80°C mechanical freezer, enabled preservation of RBCs with 20% glycerol, while maintaining the RBC integrity,

Summary
131
deformability and high 48-hour posttransfusion survival values. The use of only 20% glycerol could be beneficial in reducing the osmotic stress that is associated with the glycerolization and deglycerolization procedure and which causes cellular losses. We therefore consider that the liquid Bio-freezer could become a valuable tool for the cryopreservation of RBCs. Ultimately important findings in this thesis were discussed in a broader perspective in chapter 8.


Samenvatting

Samenvatting
134
In Nederland ontvangen jaarlijks vele mensen een bloedtransfusie. De rode bloedcellen (RBCs) worden toegediend om bloedarmoede, veroorzaakt door ernstig bloedverlies of een probleem in de bloedaanmaak, te corrigeren. De belangrijkste functie van de RBCs is het vervoeren van zuurstof vanuit de longen naar de lichaamsweefsels. De RBCs hebben in de circulatie een levensduur van 120 dagen. RBCs die in de koelkast bewaard worden hebben echter een aanzienlijke kortere levensduur. Afhankelijk van de bewaarvloeistof worden RBCs in de bloedbank momenteel 5 á 6 weken bij 2-6°C bewaard. Bij deze temperaturen worden de biochemische processen in de RBC verlaagd maar niet volledig onderdrukt. Hierdoor treden er tijdens het bewaren veranderingen op in de eigenschappen van de RBCs, die de functionaliteit na infusie zouden kunnen beïnvloeden. Er zijn diverse studies gepubliceerd die veronderstellen dat de infusie van oudere RBCs de gezondheidstoestand van de patiënt negatief kunnen beïnvloeden. Echter, de meeste van deze studies zijn retrospectief van aard en kunnen dus dan ook geen causaal verband leggen tussen de bewaarduur van RBCs en de uitkomst van de patiënt op het toegediende bloed. Het is dus van essentieel belang dat er meer inzicht in de kwaliteit van de RBCs gedurende het bewaren bij 2-6°C verkregen wordt. De reologie van de RBCs speelt hierbij een belangrijke rol, met name omdat veranderingen in de reologische eigenschappen van de RBCs (verhoogde aggregatie, lagere deformabiliteit en een toegenomen binding aan endotheelcellen) de bloedtoevoer en dus de oxygenatie in de microcirculatie kunnen belemmeren of zelfs kunnen blokkeren. In hoofdstuk 1 wordt een literatuuroverzicht gegeven over de preservatie van RBCs en de cellulaire veranderingen die hierbij optreden. De reologie van de RBC staat hierin centraal, en dan in het bijzonder de aggregatie en deformabiliteit van RBCs, aangezien deze eigenschappen met behulp van de laser-assisted optical red blood cell analyzer (LORCA) gemeten kunnen worden. Het doel van dit proefschrift was om RBCs te onderzoeken vanuit een reologisch perspectief om zo meer inzicht te verkrijgen in de kwaliteit van gepreserveerde RBCs. Daarnaast werd het gebruik van ingevroren RBCs voor transfusie doeleinden onderzocht. Het eerste gedeelte van dit proefschrift legt zowel de nadruk op de aggregatie van RBCs onder fysiologische en pathologische omstandigheden als wel op de reologie van RBCs die bij 2-6°C bewaard worden. Aggregatie van RBC (ook wel Rouleaux vorming genoemd) vindt plaats in gebieden waar de stroomsnelheid van het bloed laag of gestagneerd is. Naarmate de stroomsnelheid van het bloed weer toeneemt zullen, onder fysiologische omstandigheden, de RBC aggregaten

Samenvatting
135
uiteenvallen. Echter, onder sommige pathologische omstandigheden kunnen er grotere en sterkere aggregaten gevormd worden die niet zo gemakkelijk uiteenvallen en zo de zuurstof afgifte in de microcirculatie zouden kunnen bemoeilijken. Tot op heden is er nog veel onduidelijkheid over de fysiologische functie en het mechanisme dat ten grondslag ligt aan RBC aggregatie. Het vervangen van plasma-eiwitten door standaard polymeer oplossingen om de aggregatie van RBCs te induceren zal meer inzicht verschaffen in dit fenomeen. In hoofdstuk 2 wordt het gebruik van 200-kDa HES polymeren in RBC aggregatie studies onderzocht. Hierbij werd de electroforetische mobiliteit van RBCs gemeten om meer inzicht te verschaffen in het onderliggende aggregatie proces. De electroforetische mobiliteit van RBCs in een 200-kDa HES oplossing bleek minder negatief te zijn dan aan de hand van de toegenomen viscositeit verwacht zou worden. Dit impliceert dat 200-kDa HES polymeren de aggregatie van RBCs induceren via een absorptie mechanisme. Deze uitkomst sluit aan bij bevindingen van anderen die aantoonden dat de mate van RBC aggregatie afhankelijk is van het molecuul gewicht van HES polymeren. Ook werd de LORCA ingezet om aggregatie verschillen tussen RBCs van gezonde en zieke mensen op te sporen. We toonden aan dat naast de standaard 500-kDa dextran polymeren ook de 200-kDa HES polymeren in staat waren om aggregatie verschillen tussen RBCs van gezonde en type-1 diabetes mellitus patiënten (T1DM) te onderscheiden. Kort samengevat kan gesteld worden dat 200-kDa HES polymeren nuttig zijn om als pro-aggreganten in reologische studies gebruikt te worden. Een verhoogde aggregatie van RBCs wordt aangetroffen in diverse chronische aandoeningen, waaronder T1DM. Echter, RBC aggregatie zou ook een belangrijke rol kunnen spelen in acute situaties, zoals bij trauma patiënten met ernstig bloedverlies. Hoofdstuk 3 beschrijft de potentiële rol van RBC aggregatie na infusie van bloed componenten in hevig bloedende trauma patiënten. Onder fysiologische omstandigheden vind de aggregatie van RBCs plaats in de aanwezigheid van plasma-eiwitten. Deze RBC aggregaten worden voornamelijk gevormd in het midden van het bloedvat waardoor leukocyten en mogelijk bloed plaatjes naar de wand van het bloedvat gedreven worden. Dit proces is essentieel aangezien leukocyten en bloedplaatjes in nauw contact dienen te komen met het endotheel om effectief te kunnen zijn bij het bestrijden van een infectie en het bevorderen van de bloedstolling. Momenteel wordt voor trauma patiënten met ernstig bloedverlies de infusie van RBC, bloedplaatjes en ingevroren plasma in een 1:1:1 verhouding aanbevolen. Infusie van ingevroren plasma (en dus van fibrinogeen) zou de

Samenvatting
136
aggregatie van RBCs kunnen promoten. Zoals gezegd kan RBC aggregatie een belangrijke rol spelen in het ondersteunen van de hemostase omdat leukocyten en mogelijk bloedplaatjes zo makkelijker in de nabijheid van het beschadigde endotheel terecht komen. Echter, een verhoogde aggregatie kan ook nadelig zijn in bepaalde trauma patiënten waarvan de RBC reologie al verstoord is. De LORCA zou uitkomst kunnen bieden om te bepalen of in deze trauma patiënten de infusie van bloedcomponenten in een 1:1:1 verhouding invloed heeft op het aggregatie proces van RBCs. De reologische eigenschappen van RBCs zijn belangrijke kwaliteitsvariabelen in de transfusiegeneeskunde. Meer inzicht in de reologische veranderingen tijdens RBC preservatie zou de transfusie uitkomst kunnen beïnvloeden. Er zijn diverse studies die aantonen dat de reologische eigenschappen van RBCs die bij 2-6°C bewaard worden, al na enkele weken verslechterd zijn. Echter, de meeste van deze studies zijn verouderd omdat er RBCs getest werden die niet gefilterd waren van hun leukocyten. In hoofdstuk 4 worden daarom de reologische eigenschappen en diverse hematologische variabelen van leukogefiltreerde RBCs onderzocht die gedurende 7 weken bij 2-6°C bewaard worden. We toonden aan dat de bewaarde RBCs geen verhoogde aggregatie vertoonden. Wel namen we een afname waar in de RBC deformabiliteit gedurende het bewaren. Deze afname in RBC deformabiliteit viel echter binnen de fysiologische waarden en werd alleen onder hoge shear stress condities waargenomen. Het ATP gehalte van de RBC nam wel aanzienlijk af tijdens het bewaren. De MCV, pH en MCHC daarentegen veranderde minder gedurende 7 weken. Kort samengevat kan gesteld worden dat er tijdens het bewaren bij 2-6°C veranderingen optreden in de eigenschappen van de RBCs. Deze veranderingen hebben echter geen nadelige uitwerkingen op het vermogen van de RBC om te kunnen aggregeren en vervormen. Aan de hand van deze en recente bevindingen van anderen kunnen we concluderen dat de reologische eigenschappen van RBCs goed behouden blijven bij 2-6°C. Een manier om de veranderingen die tijdens het koud bewaren optreden tegen te gaan is om de bewaartemperatuur zodanig te verlagen dat de biochemische processen in de RBC volledig onderdrukt worden. Cryopreservatie is een methode om RBCs langdurig te kunnen preserveren. In het tweede gedeelte van dit proefschrift wordt daarom de nadruk gelegd op ingevroren RBCs. Hierbij wordt in hoofdstuk 5 de cryopreservatie van RBCs en het gebruik ervan in de transfusiegeneeskunde besproken. Ondanks het feit dat ingevroren RBCs decennia lang bewaard kunnen worden en zo de beschikbaarheid van RBCs aanzienlijk kunnen vergroten, is het gebruik van dit bloedproduct in de transfusie-

Samenvatting
137
geneeskunde beperkt. Dit komt voornamelijk door de hogere kostprijs, de langere verwerkingstijd en de lagere opbrengst die aan het invriezen van RBCs verbonden zijn. Ook de beperkte kennis omtrent de kwaliteit van ingevroren RBC heeft het gebruik van dit bloedproduct in de kliniek beperkt. Momenteel worden RBCs voornamelijk ingevroren voor militaire doeleinden en om zeldzame bloedgroepen langdurig te bewaren. De laatste jaren is er echter steeds meer onderzoek verricht naar de kwaliteit van ingevroren RBCs. De reologische eigenschappen van ingevroren RBCs waren echter nog nauwelijks onderzocht. In hoofdstuk 6 worden daarom de reologische eigenschappen en hematologische variabelen van ingevroren RBCs onderzocht. We toonden aan dat de aggregatie van ingevroren RBCs lager was dan die van verse en in de koeling (2-6°C) bewaarde RBCs. De deformabiliteit van ingevroren RBCs daarentegen was hoger dan die van verse RBCs maar vergelijkbaar met de deformabiliteit van RBCs die 21 of 35 dagen in de koeling bewaard werden. De osmotische flexibiliteit, hemolyse, MCV en MCHC van ingevroren RBCs was veranderd vergeleken met verse en in de koeling bewaarde RBCs. Het ATP gehalte van ingevroren en verse RBCs was echter vergelijkbaar. We concludeerden dat ondanks dat ingevroren RBCs fragieler waren dan verse en in de koeling bewaarde RBCs, dit geen nadelige uitwerking had op de aggregatie, deformabiliteit en het ATP gehalte van ingevroren RBCs. Meer kennis omtrent de kwaliteit van ingevroren RBC zou het gebruik in de kliniek kunnen bevorderen. Ook het gebruik van nieuwe vriestechnieken zou hierbij een rol kunnen spelen. In dit opzicht werd in hoofdstuk 7 het gebruik van een nieuwe Bio-vriezer (Supachill) getest om RBCs in te vriezen. Humane en ratten RBCs werden met verschillende glycerol concentraties in de Bio-vriezer bij -25°C ingevroren of in combinatie met een mechanische vriezer bij -80°C opgeslagen. Van de humane RBCs werden de deformabiliteit en diverse hematologische variabelen getest. De ratten RBCs werden gebruikt om de overlevingsduur van ingevroren RBCs na infusie te bepalen. We toonden aan dat RBCs die in de Bio-vriezer ingevroren en bewaard waren, ofwel een lagere deformabiliteit, dan wel een verminderde overleving 48 uur na infusie vertoonden. Het gebruik van de Bio-vriezer in combinatie met een standaard -80°C vriezer, maakt het mogelijk om RBCs met slechts 20% glycerol in te vriezen terwijl de RBC integriteit, deformabiliteit en de overlevingsduur tot en met 48 uur na infusie behouden werd. Het gebruik van een glycerol concentratie van slechts 20% kan voordelig zijn bij het invriezen van RBCs omdat dit de osmotische stress en dus de

Samenvatting
138
hemolyse van de vriesprocedure zou kunnen verminderen. We veronderstellen dat de Bio-vriezer een belangrijke rol zou kunnen gaan spelen in de preservatie van RBCs. De belangrijkste bevinding van dit proefschrift worden in een breder perspectief besproken in hoofdstuk 8.

Acknowledgements

Acknowledgements
140
Eindelijk is het zo ver, “het boekje” is af.
Gedurende de afgelopen jaren is er veel gebeurd en er zijn diverse mensen die ik toch zeer dankbaar ben. Allereerst wil ik Supachill en Haemoscan bedanken voor hun financiële bijdrage waardoor dit proefschrift mede mogelijk gemaakt werd. Ook wil ik ook de leescommissie bedanken voor hun tijd en inzet.
Daarnaast Gerhard. Beste Gerhard, het is niet altijd even makkelijk geweest voor ons beiden maar ik ben je zeer dankbaar voor je advies en begeleiding. Vooral op het eind bleek het of we het steeds vaker eens waren. Het was een nuttig leerproces voor ons beiden.
Reindert, gedurende mijn gehele AIO periode stond je deur altijd open voor raad en daad, maar zeker ook voor gezelligheid. Dat heb ik super gewaardeerd. Ook heb ik veel geleerd van je kritische blik met betrekking tot het schrijven van artikelen. Bedankt hiervoor en blijf zoals je bent!
Beste Johan. Bedankt voor al je wijsheid, tijd en gezelligheid. Ik heb veel van je geleerd. Het was altijd een waar genoegen om richting Amsterdam te reizen om nieuwe sampels op te halen of om nog het een en ander door te nemen. Thankxxx
Margriet en Janny. Ook jullie bedankt voor jullie raad en tijd. Mede dankzij jullie kritische blik is het eerste artikel goed tot uiting gekomen.
Hans, soms moet je wel eens gedacht hebben: daar komt ze weer. Dat krijg je als je statistiek zo goed kunt uitleggen. Je maakte altijd tijd ondanks dat je agenda zelf super vol zat. En na een half uur bij jouw geweest te zijn werd statistiek ineens hèèel simpel. Bedankt dat je wat rust in de chaos verzorgde.
Michel, ook jij bedankt voor je medewerking en inzet bij het labelen. Je vond altijd wel even tijd voor me ondanks je eigen werkzaamheden.
Lieve mensen van het CDL: waar zouden we zijn zonder jullie. Bedankt voor het meedenken, assisteren en de gezelligheid. Jullie zijn een fijne club om mee te werken.
Ook op de afdeling zijn er diverse mensen die ik wil bedanken. Allereerst wil ik de vele bloeddonoren bedanken voor hun bijdrage. Zonder jullie geen onderzoek: super bedankt!!

Acknowledgements
141
Henny en Betsy, bedankt voor jullie tijd, inzet en meedenken met mijn proeven. Jullie waren erg geduldig en altijd bereid om me te helpen. Dit heeft geresulteerd in een mooi artikel, mede dankzij jullie.
Bastiaan en Prashant. Ook jullie deur stond altijd open. Ik heb daar af en toe gebruik van gemaakt. Bedankt hiervoor en voor alle gezelligheid. Het is motiverend om wetenschappers zoals jullie tegen te komen.
Marten, Marieke, Arina, Stefan, Adam, Anieska en Deepak. Bedankt voor jullie gezelligheid, conversaties en de lekkere borrels. Vooral de spontane vrijdag middag borrels waren super!
Also many thanks to my Indonesian friends: Dear Punto, when you came over it was always very nice (especially the Indonesian food). The congress in Amsterdam was special en we had lots of fun! It was a pleasure of having you here in the department.
Dear Shanti, You rock girl!! It was great having you here in our department. I hope your new job will be great as well, you deserve it!. I want to especially thank for your beautiful drawings in this thesis.
Beste Ingrid. Ook jij bedankt voor al je tijd en energie en dan met name met betrekking tot de lay-out: het is een super mooi geheel geworden.
Lieve Babs, thanks voor al je gezelligheid en advies. Zoals jij is er maar een. Laat je niet gek maken!!
Lieve Ed, waar zou ik zijn zonder jou. Niet alleen voor alle blond gerelateerde computer vraagjes maar zeker ook voor de snoepjes. Thanks voor making my life sweeter!!
Lieve Paranimfen: jullie zijn allen speciaal, daarom heb ik er ook vier.
Ward, je bent een bijzonder persoon. Laat je niets anders wijs maken. Soms iets meer in jezelf geloven en dan zie je dat je alles kunt bereiken wat je wilt! Bedankt voor je positieve mentaliteit en je gezelligheid.
Gerwin: je handigheid met computers werd vaak door mij benut. Daar komt bij dat we dezelfde mentaliteit en humor hebben. Bedankt dat je de laatste jaren mijn roommate was. Het was in ieder geval een stuk gezelliger met jouw erbij.

Acknowledgements
142
Oana, Ook voor jou was het niet altijd even gemakkelijk, een nieuwe cultuur, het werken op het lab. Je was echter altijd bereid te helpen, en we hebben veel aan elkaar gehad. Bedankt voor je gezelligheid.
Lieve Anita, door je gezellige babbel waren we niet altijd even productief. Echter het was erg gezellig en gelukkig ben jij ook zo’n snoepert. Jouw tijd komt nog wel. Gewoon nog even volhouden.
Alle dierbare Calameriaantjes: in het bijzonder Ghis en Peer, Henk en Manon, Marjolein, Wietse, Rob en Ciske. Bedankt voor jullie gezelligheid. Vooral de borrels in de Singelier, de gezellige etentjes, de duikkampjes en de Egypte vakanties hebben me weer nieuwe energie gegeven om er tegen aan te kunnen. Wanneer gaan we weer??
Alle lieve vriendjes en vriendinnetjes die mij de afgelopen jaren iets minder hebben kunnen zien: El, Syl, Simo en Ramona. Thanks dat jullie er waren en vanaf nu wordt er weer tijd gemaakt voor leuke dingen.
Ritsert, bedankt dat je de tijd neemt om me op het juiste pad te brengen. Een uur met jouw geeft enorm veel motivatie. Thanks
Ed, Michell, Jackie en Robert ook jullie bedankt voor alle steun, raad en gezelligheid. Ook de borrels (ja ja ik hou van borrels), etentjes en ski vakanties werden zeer gewaardeerd. Mam en Pap, bedankt dat jullie er altijd voor me waren. Mede dankzij jullie opvoeding is dit alles mogelijk geweest. Ik ben trots op jullie.
Last but not least: lieve Stephen, ze zeggen dat je niet in je eentje promoveert. Daar heb jij het nodige van ondervonden. Een vriendin die vele uren maakt en slapeloze nachten als gevolg. Het was zeker ook niet gemakkelijk voor jou: bedankt voor je oneindige geduld en dat je er altijd voor me was: Tá tú an solas na gréine de mo shaol agus is breá liom tú. Dikke kus San