universidade federal do cearÁ faculdade de medicina … · 2019-01-21 · universidade federal do...
TRANSCRIPT

UNIVERSIDADE FEDERAL DO CEARÁ
FACULDADE DE MEDICINA
DEPARTAMENTO DE PATOLOGIA E MEDICINA LEGAL
PROGRAMA DE PÓS-GRADUAÇÃO EM MICROBIOLOGIA MÉDICA
FELIPE RODRIGUES MAGALHÃES DE AGUIAR
BIOFILMES DO COMPLEXO Sporothrix schenckii: FORMAÇÃO E
SENSIBILIDADE IN VITRO
FORTALEZA
2016

1
FELIPE RODRIGUES MAGALHÃES DE AGUIAR
BIOFILMES DO COMPLEXO Sporothrix schenckii: FORMAÇÃO E
SENSIBILIDADE IN VITRO
Dissertação apresentada ao Programa de
Pós-Graduação em Microbiologia
Médica da Universidade Federal do
Ceará, como requisito parcial à obtenção
do título de mestre em Microbiologia
Médica. Área de concentração:
Micologia Médica.
Orientadora: Profª Drª. Raimunda Sâmia
Nogueira Brilhante
FORTALEZA
2016

2
______________________________________________________________________
_____
Página reservada para ficha catalográfica que deve ser confeccionada após apresentação
e alterações sugeridas pela banca examinadora.
Para solicitar a ficha catalográfica de seu trabalho, acesse o site: www.biblioteca.ufc.br,
clique no banner Catalogação na Publicação (Solicitação de ficha catalográfica)
______________________________________________________________________
_____

3
FELIPE RODRIGUES MAGALHÃES DE AGUIAR
BIOFILMES DO COMPLEXO Sporothrix schenckii: FORMAÇÃO E
SENSIBILIDADE IN VITRO
Dissertação apresentada ao Programa de
Pós-Graduação em Microbiologia
Médica da Universidade Federal do
Ceará, como requisito parcial à obtenção
do título de mestre em Microbiologia
Médica. Área de concentração:
Micologia Médica.
Aprovada em: ___/___/______.
BANCA EXAMINADORA
________________________________________
Profª. Drª. Raimunda Sâmia Nogueira Brilhante (Orientadora)
Universidade Federal do Ceará (UFC)
_________________________________________
Profª. Drª. Flávia Almeida Santos
Universidade Federal do Ceará (UFC)
_________________________________________
Profª. Drª. Adriana de Queiroz Pinheiro
Universidade Estadual do Ceará (UECE)

4
A Deus, Mestre e Senhor do Universo;
À minha avó, Sônia Magalhães;
Aos meus pais, Marcos e Ana Paula;
Ao meu irmão, Rafael Cortez.

5
AGRADECIMENTO
A Deus, por me proporcionar a oportunidade ímpar de escrever essas linhas;
À minha avó Sônia Magalhães, a quem eu jamais cansarei de agradecer,
pois foi a grande responsável por tudo de melhor que já aconteceu na minha vida;
Aos meus pais, Ana Paula Cortez e Marcos Magalhães, que foram o
caminho escolhido por Deus para me dar a vida;
Ao meu irmão, Rafael Cortez, que tem um coração generoso como poucos;
À toda a minha família, meus amores, Vovó Vanda, Vô Hermenegildo,
Nelsa, tios (Fernando, Adelaide, Gláucia, Messias, Neném, Fernanda, Érika, Alexandre,
Germana, André, Socorro e Elias) e primos (Lia, Leo, Ruth, Carolina, Daniel, Mariana,
Victor, Bia, João Victor, Iolanda, Yuri, Yan, Júlia);
À Profª. Drª. Sâmia Brilhante, pelo “acreditar”, pelo voto de confiança e
pelo acompanhamento nessa jornada tão difícil, mas tão gratificante. Por sua paciência,
por suas orientações e por querer o melhor de cada um dos seus alunos. Fui muito feliz
no dia que pude tê-la como orientadora, pois já sabia que meu período de mestrado seria
tutorado por uma pessoa de enormes capacidades.
Aos participantes da banca qualificadora, Prof. Dr. Manoel Paiva de Araújo
Neto e Draª Délia Jéssica Astete Medrano, assim como da banca de defesa, Profª Drª
Flávia Almeida Santos e Profª Drª Adriana Queiroz Pinheiro, pelo tempo,
disponibilidade, valiosas colaborações e sugestões.
À Lucilene Queiroz, minha eterna companheira de bancada, que viveu do
meu lado todas as angústias, surpresas, tristezas e alegrias que só o Sporothrix pôde nos
proporcionar. Saiba, amiga, que você tem grande parte nessa dissertação;
Aos melhores amigos do mundo: Leo Moraes (sem palavras para expressar
o quanto você foi importante nessa vitória!), Thiago Alberine, Alisson Leno, Carlinhos
de Carvalho, Jontinhas, Ranilson Alves, Diogo Evangelista, Samuel Estevam, Juninho,
João Marques, Amanda Militão, Felipe Sobral, Isabella Militão, Neiva, Luciana
Moraes, Cássio, Vivi Araújo, Marquinhos Araújo, Rubens Filho, Ianna Andrade,
Cristiano Rodrigues, Kelton Bantim e tantos outros que sempre estiveram e estão
comigo nos desafios da vida e que, durante dois anos, viveram o mestrado junto
comigo, assim como Cristiane, Aurenívea, Naruytto, Isabely, Nadja, Leonardo, Dudu,
Kéllen, Clarissa, Alana, Marcelo, Cesinha, Márcia, Carol, Mércia, Fernanda, Cláudia,

6
Yelena, Carlysandra e os demais (que graças a Deus são muitos), pelo apoio e
compreensão de sempre;
Aos colegas do Centro Especializado em Micologia Médica (CEMM),
Aline, David, Erisvaldo, Giovanna, Lucas, Gláucia, Jônathas, Rosana, Jamille, Tony,
Jaime, Edmilson, Lana, Lívia, Kleybson, Ewerton, Vandbergue, Jônatas, Patrícia,
Raquel, Yago, Ana Luiza, Gleiciane, Raíssa, Géssica, Chryster, Thalita, Juliana, Jônatas
e Terezinha (que eu “amo de paixão” desde o primeiro dia!), por toda a ajuda e
paciência durante o mestrado. Assim como a mais “Mãecretária” do PPGMM, Carol,
que sempre ajudou no que foi possível e impossível.
Aos Professores Marcos Fábio Gadelha, Rossana Cordeiro, Júlio Sidrim,
Cibele Barreto, Fernanda Edna, Lidiany Rodrigues, Lilia Maria, Luís, pela generosidade
e vontade de me fazer aprender muito além da Microbiologia, não somente através da
transferência de conhecimento, mas do amadurecimento do olhar crítico, da percepção
do meio e da vida como um todo.
À Dra. Silviane e Profª Débora Castelo Branco, pelo conhecimento e pelas
horas e horas de microscopia, sem importar se durante o dia ou entrando à noite.
Ao LIME, à Central Analítica da UFC e ao CEMM como um todo por
disponibilizarem seus profissionais e equipamentos para a realização dessa dissertação.
A todos os funcionários do bloco da Biomedicina, porteiros, auxiliares de
serviços gerais, etc., que proporcionaram um ambiente saudável para o desenvolvimento
desse estudo;
Ao Professor Zoilo Pires de Camargo, assim como todos os colaboradores
de seu laboratório na Universidade Federal de São Paulo, por gentilmente terem cedido
as cepas utilizadas neste estudo;
À Universidade Federal do Ceará, instituição que me acolheu desde a
graduação e que eu tenho o maior orgulho do mundo em fazer parte;
À CAPES, pelo aporte financeiro que tanto foi útil durante esses anos;

7
“Talvez não tenha conseguido fazer o
melhor, mas lutei para que o melhor
fosse feito. Não sou o que deveria ser,
mas, Graças a Deus, não sou o que era
antes.”
Marthin Luther King

8
RESUMO
A esporotricose é uma micose subcutânea cosmopolita, encontrada, principalmente, em
áreas tropicais e subtropicais da América Latina, sendo causada por espécies do
complexo Sporothrix schenckii. O objetivo deste estudo foi demonstrar a capacidade de
fungos do complexo S. schenckii de produzir biofilmes in vitro e determinar o perfil
desses biofilmes frente às drogas antifúngicas clássicas. Foram utilizadas 4 espécies do
complexo Sporothrix schenckii: S. brasiliensis (n=10), S. schenckii sensu stricto (n=2),
S. globosa (n=2) e S. mexicana (n=4). A formação dos biofilmes foi feita através da
transferência de inóculos de 2 x 105 conídios/mL para placas de microdiluição, que
foram incubadas a 25 °C por 5 dias. Após cada dia de incubação, realizaram-se ensaios
de XTT e Cristal violeta, para se determinar a atividade metabólica e biomassa dos
biofilmes, respectivamente. Para os ensaios de sensibilidade, três concentrações
diferentes (MIC planctônico, 10 x MIC e 50 x MIC) dos antifúngicos anfotericina B
(AMB), caspofungina (CAS), cetoconazol (KTC), voriconazol (VRC) e fluconazol
(FLC) foram transferidas para os poços com biofilmes, que foram incubados por 72
horas a 25 °C. O perfil dos biofilmes frente às drogas antifúngicas foi determinado
através de ensaios de XTT e cristal violeta. Biofilmes foram produzidos em lamínulas
de Thermanox™ para visualização através de microscopia óptica com coloração de
vermelho-congo, microscopia eletrônica de varredura e microscopia confocal.
Verificou-se que todas as espécies do complexo S. schenkii são fortes formadoras de
biofilme, não havendo diferença de formação entre as espécies. A absorbância dos
biofilmes após exposição às drogas demonstrou que a biomassa e a atividade metabólica
foram reduzidas de forma significativa (p<0,05), principalmente, em S. brasiliensis. Os
antifúngicos testados mostraram redução na atividade metabólica e biomassa,
principalmente, em concentrações 50 vezes maiores que os MICs planctônicos,
demonstrando o incremento protetivo que a forma séssil possui frente à forma
planctônica.
Palavras-chave: Dimorfismo. Resistência. Antimicóticos.

9
ABSTRACT
Sporotrichosis is a cosmopolitan subcutaneous mycosis, found mainly in tropical and
subtropical areas of Latin America, caused by species of Sporothrix schenckii complex.
The aim of this study was to demonstrate the capacity of Sporothrix schenckii complex
to produce in vitro biofilms and to determine the profile of these biofilms against to
classical antifungal drugs. Four species from Sporothrix schenckii complex were used:
S. brasiliensis (n=10), S. schenckii sensu stricto (n=2), S. globosa (n=2) and S.
mexicana (n=4). The formation of biofilms was performed by transferring inoculum of
2 x 105 conidia/mL for microdilution plates, which were incubated at 25 °C for 5 days.
After each day of incubation, XTT and Crystal Violet assays were made to determine
the metabolic activity and biomass of biofilm, respectively. For susceptibility tests,
three different concentrations (planktonic MIC, 10 x MIC and 50 x MIC) of antifungals
amphotericin B (AMB), caspofungin (CAS), ketoconazole (KTC), voriconazole (VRC)
and fluconazole (FLC) were transferred to biofilm wells, which were incubated for 72
hours at 25 °C. The biofilms profile then antifungal drugs presence was determined by
the XTT and crystal violet assays. Biofilms were produced in Thermanox™ coverslips
for viewing by optical microscopy with congo-red staining, scanning electron
microscopy and confocal microscopy. It was found that all species from S. schenckii
complex are strong biofilm-forming, with no difference formation between them.
Absorbance of biofilm after drug exposure showed that biomass and metabolic activity
was significantly reduced (p<0.05), mainly in S. brasiliensis. Antifungal drugs tested
showed metabolic activity and biomass reduction, especially at concentrations 50 times
higher than the planktonic MICs, demonstrating the protective increase in sessile form
when compared for planktonic form.
Keywords: Dimorphism. Sporothrix. Antimycotics.

10
LISTA DE ILUSTRAÇÕES
Figura 1 – Alguns casos reportados de esporotricose no Peru, Estados Unidos da
América e Austrália após a classificação do agente etiológico no gênero
Sporothrix......................................................................................................... 19
Figura 2 – A. Esporotricose cutâneo-fixa-fixa: lesão desenvolvida no local de
inoculação. B. Esporotricose cutâneo-linfática: Lesões nódulo-ulcerativas
surgem ao longo dos vasos linfáticos, a partir da lesão inicial......................... 22
Figura 3 – Sporothrix spp. em sua fase filamentosa. A. Macromorfologia das cepas
melanizadas (escuras) e não-melanizadas (brancas). B. Hifas e conídios em
forma de “flor de margarida” (destacados por setas). C. Conídios de
Sporothrix brasiliensis em microscopia eletrônica de varredura –Ampliação
10999X............................................................................................................. 25
Figura 4 – Distribuição mundial das espécies S. brasiliensis, S. globosa e Sporothrix s.
str. (Clades IIa e IIb)......................................................................................... 27
Figura 5 – Etapas de formação dos biofilmes microbianos............................................... 35
Figura 6 – A. Macromorfologia de colônias filamentosas de Sporothrix schenckii s.
str., melanizadas, em ágar batata dextrose, incubadas a 25 °C. B.
Macromorfologia de colônias de Sporothrix schenckii s. str. em ágar BHI
suplementado com 5% de sangue de carneiro, incubadas a 37 °C................. 37
Figura 7 – Esquema de metodologia para a formação de biofilmes filamentosos de
espécies do complexo S. schenckii.................................................................. 48
Figura 8 – Esquema de metodologia para a avaliação da cinética de biofilmes
filamentosos de espécies do complexo S. schenckii através de ensaios de
biomassa (cristal violeta) e atividade metabólica (XTT).................................. 50
Figura 9 – Esquema de metodologia para ensaio de sensibilidade antifúngica em
espécies do complexo S. schenckii, em suas fases filamentosas, por
microdiluição em caldo..................................................................................... 52
Figura 10 – Esquema de metodologia para ensaio de atividade metabólica e biomassa de
espécies do complexo S. schenckii, em suas fases filamentosas, frente aos
antifúngicos....................................................................................................... 53
Figura 11 – Fluxograma dos ensaios utilizados na metodologia do estudo......................... 54
Figura 12 – Atividade metabólica (A) e biomassa (B) de biofilmes de Sporothrix spp. de

11
acordo com o tempo de maturação em ensaios de XTT e cristal violeta,
respectivamente (n=18)....................................................................................
55
Figura 13 – Microscopia óptica de biofilmes maduros de Sporothrix globosa, corados
através da técnica com corante Vermelho-Congo. A. Matriz extracelular
densa (fundo vermelho), com hifas (setas pretas) e conídios (setas brancas);
B. Canais de água dentro da rede de hifas (setas). Ampliação de 400x........... 57
Figura 14 – Imagens de CLSM de biofilmes maduros de Sporothrix brasiliensis. A.
Hifas (setas pretas) e conídios (setas brancas) formando uma estrutura densa
de biofilme. B. Visão tridimensional do biofilme. Ampliação de 400X.
Imagens adquiridas com laser 488 nm e 561nm para a detecção de SYTO9 e
iodeto de propídio, que identificam células vivas e mortas, respectivamente.. 59
Figura 15 – Imagens de microscopia eletrônica de varredura de biofilmes de S.
brasiliensis formados em lamínulas de Thermanox™ após 24, 48, 72, 96,
120 e 144 horas de incubação (A - F, respectivamente) - Ampliação:
2000X (A - E) e 4032X (F). A. Células aderidas à superfície da lâmina. B.
Início da filamentação pós-adesão. C. Formação de redes de hifas e
conídios. D. Espessamento do biofilme com detecção de produção de
matriz. E. Biofilme maduro com presença de conídios, hifas, matriz e
canais de água (identificados por setas). F. Canais de água no biofilme de
S. brasiliensis (identificados por setas)...........................................................
61

12
LISTA DE TABELAS
Tabela 1 – Origem e local de isolamento das cepas de espécies do complexo Sporothrix
schenckii utilizadas neste estudo...................................................................... 47
Tabela 2 – Classificação dos biofilmes das espécies com complexo Sporothrix
schenckii........................................................................................................... 56
Tabela 3 – Concentração inibitória mínima (MIC) e concentração fungicida mínima
(MFC) de isolados planctônicos do complexo Sporothrix schenckii............... 63
Tabela 4 – Atividade metabólica dos biofilmes de Sporothrix spp. após exposição às
drogas antifúngicas (492 nm) (p<0,05)............................................................ 65
Tabela 5 – Biomassa dos biofilmes de Sporothrix spp. após exposição às drogas
antifúngicas (590 nm) (p<0,05)........................................................................ 65

13
LISTA DE ABREVIATURAS E SIGLAS
3D Terceira dimensão
AMB Anfotericina B
BHI Brain Heart Infusion
CAS Caspofungina
CEM Concentração Mínima Eficaz
CEMM Centro Especializado em Micologia Médica
CFM Concentração Fungicida Mínima
CIM Concentração Inibitória Mínima
CLSI Clinical & Laboratory Standards Institute
CLSM Confocal Laser Scanning Microscopy
DO Densidade óptica
EUA Estados Unidos da América
FLC Fluconazol
h hora
HIV Human Immunodeficiency Virus – Vírus da Imunodeficiência
Humana
HMDS Hexametildisilazano
ITS Internal Transcribed Spacer
KTC Cetoconazol
MALDI-TOF Matrix Assisted Laser Desorption/Ionization - Time of Flight
MEV Microscopia Eletrônica de Varredura
MG Média geométrica
min minuto
MOPS 3-[N-morfolino]-propano-ácido sulfônico
PBS phosphate buffered saline
PCR Polymerase Chain Reaction – Reação em Cadeia da Polimerase
PDA Potato Dextrose Agar – Agar Batata Dextrose
pH Potencial Hidrogeniônico
RPMI Roswell Park Memorial Institute
UFC Unidades Formadoras de Colônias
VRC Voriconazol
XTT 2,3-Bis-(2-methoxy-4-nitro-5-sulfophenyl)-2H-tetrazolium-5-
carboxanilide salt

14
LISTA DE SÍMBOLOS
% Porcentagem
°C Graus Celsius
µg Micrograma
µL Microlitro
µm Micrômetro
kDa Quilodalton
kV Quilovolt
mL Mililitro
mM Milimolar
nm Nanômetro
™ Trademark (Marca Registrada)

15
SUMÁRIO
1 REVISÃO BIBLIOGRÁFICA .............................................................................. 17
1.1 Esporotricose ........................................................................................................... 17
1.1.1 Histórico..................................................................................................................... 17
1.1.2 Epidemiologia............................................................................................................ 19
1.1.3 Patogenia................................................................................................................... 21
1.2 Agentes etiológicos – Sporothrix spp....................................................................... 22
1.2.1 Taxonomia................................................................................................................. 22
1.2.2 Morfologia.................................................................................................................. 24
1.2.3 Distribuição mundial do complexo Sporothrix schenckii........................................ 26
1.2.4 Fatores de Virulência................................................................................................ 28
1.2.5 Biofilmes..................................................................................................................... 32
1.3 Diagnóstico laboratorial da esporotricose.............................................................. 36
1.4 Sensibilidade aos antifúngicos e tratamentos propostos....................................... 38
1.4.1 Iodeto de potássio....................................................................................................... 39
1.4.2 Compostos azólicos.................................................................................................... 40
1.4.3 Anfotericina B............................................................................................................ 42
1.4.4 Caspofungina............................................................................................................. 43
1.4.5 Termoterapia.............................................................................................................. 44
2 HIPÓTESES.............................................................................................................. 45
3 OBJETIVO GERAL................................................................................................ 45
3.1 Objetivos específicos................................................................................................. 45
4 MATERIAIS E MÉTODOS.................................................................................... 46
4.1 Microrganismos......................................................................................................... 46
4.2 Formação do biofilme filamentoso de Sporothrix spp. para testes de
sensibilidade............................................................................................................... 47
4.3 Cinética de crescimento dos biofilmes..................................................................... 48
4.4 Morfologia e estrutura dos biofilmes de Sporothrix spp........................................ 50
4.5 Sensibilidade da forma planctônica........................................................................ 51
4.6 Atividade metabólica e biomassa dos biofilmes frente aos antifúngicos............. 53
4.7 Análises estatísticas................................................................................................... 53
5 RESULTADOS......................................................................................................... 54
5.1 Desenvolvimento e cinética do biofilme.................................................................. 54

16
5.2 Morfologia e estrutura dos biofilmes de Sporothrix spp....................................... 56
5.3 Sensibilidade planctônica......................................................................................... 62
5.4 Atividade metabólica e biomassa dos biofilmes frente aos antifúngicos.............. 64
6 DISCUSSÃO............................................................................................................. 66
7 REFERÊNCIAS BIBLIOGRÁFICAS.................................................................... 75
ANEXO A.................................................................................................................. 95

17
1 REVISÃO BIBLIOGRÁFICA
1.1 Esporotricose
1.1.1 Histórico
Em 1898, no Johns Hopkins Hospital em Baltimore-EUA, o médico americano
Benjamin Robinson Schenck isolou um fungo proveniente de nódulos ulcerados nos
dedos e antebraço de um paciente adulto de 36 anos de idade e encaminhou ao
patologista e micologista Erwin Frink Smith, que identificou o fungo como uma espécie
pertencente ao gênero Sporotrichum, sendo este o primeiro caso relatado do agente
etiológico da esporotricose (SCHENCK, 1898; TÉLLEZ et al., 2014), uma micose
subcutânea, que apresenta-se, geralmente, na forma sub-aguda ou crônica. Já em 1900,
dois anos após o primeiro relato, um segundo caso suspeito de esporotricose foi
visualizado em Chicago por Hektoen e Perkins. Uma criança do sexo masculino foi
atendida com uma lesão no dedo causada por um martelo. Sem necessidade de
tratamento, esta lesão regrediu espontaneamente e, pela primeira vez, o agente
etiológico foi classificado como Sporothrix schenckii, sua atual denominação
(HEKTON & PERKINS, 1900). Outros possíveis casos de esporotricose foram
relatados por Linck em 1809 e por Lutz em 1889, porém, estes pesquisadores não
obtiveram sucesso no isolamento do fungo para confirmação (KWON-CHUNG &
BENNETT, 1992).
Cerca de uma década depois, novos avanços no entendimento sobre a
esporotricose foram descritos no Brasil. Em 1907, os pesquisadores Adolf Lutz e
Alfonso Splendore reportaram, pela primeira vez, a possibilidade de cultivo de S.
schenckii em sua fase leveduriforme in vitro e a infecção natural por este
microrganismo em ratos (LUTZ & SPLENDORE, 1907), levando alguns pesquisadores
a cogitarem a hipótese de transmissão da esporotricose através da mordedura destes
roedores (PUPO, 1917).
Surtos em países como a África do Sul têm sido relatados desde os primeiros
anos do século XX (PIJPER & PULLINGER, 1927). Em Witwatersrand, uma cadeia de
montanhas sul-africanas, um surto de esporotricose em trabalhadores de minas de ouro
foi registrado entre os anos de 1938 e 1947, chegando a aproximadamente 3300
mineiros doentes, que tiveram traumas na pele por lascas de madeira contaminada das

18
vigas de sustentação por onde, provavelmente, o fungo foi inoculado e ocorreu o início
da infecção (LURIE, 1963; GOVENDER et al., 2015).
Apesar dos avanços, somente décadas após a identificação do fungo causador da
esporotricose, S. schenckii foi classificado corretamente, baseado em observações feitas
por Carmichael em 1962, que verificou diferenças significativas na esporulação dos
conídios entre espécies do gênero Sporotrichum e fungos isolados em casos de
esporotricose (CARMICHAEL, 1962).
Surtos com menor quantidade de pacientes também foram registrados em vários
estados norte-americanos, como em 1988, onde casos começaram a aparecer em agentes
de reflorestamento em Nova Iorque e Illinois e depois foram identificados em 15
diferentes estados, sempre em trabalhadores que estiveram em contato com esfagno, um
musgo bastante utilizado no cultivo de orquídeas e bonsais, adquirido de um mesmo
distribuidor (DIXON et al., 1991) e em uma estação militar de Oklahoma entre 1992 e
1993 (DOOLEY et al., 1997). Países como Austrália (FEENEY et al., 2007; DHINGRA
et al., 2015), China (SONG et al., 2013) e Venezuela (CAMACHO et al., 2015) também
apresentaram casos de esporotricose humana.
Na Austrália, nas proximidades de Darwin, Territórios do Norte, nove casos de
esporotricose foram confirmados entre julho de 2014 e março de 2015. Todos os
pacientes tiveram atividades de jardinagem, seja por lazer ou como atividade laboral.
Além disso, tiveram contato com feno que havia sido molhado pela chuva, situação
extremamente propícia para o crescimento de fungos do gênero Sporothrix
(MCGUINNESS et al., 2016).
Na Europa, o maior surto relatado ocorreu no início do século XX, onde um
grande número de pessoas adoeceu de esporotricose na França. Porém, houve um
decréscimo no número de casos nas décadas seguintes, fazendo com que a Europa não
seja um continente comumente afetado (CHAKRABARTI et al., 2015), exceto por
casos esporádicos, como os relatados na Itália em 2008 (CRISEO et al., 2008) e na
França em 2009 (MAGAND et al., 2009).
Na América do Sul, continente com muitas áreas endêmicas de esporotricose,
Ramírez-Soto (2015) reuniu vários estudos sobre esta micose no Peru, contabilizando

19
em sua pesquisa um total de 1503 casos entre 1985 e 2012 e demonstrando uma região
com altos índices de infecção, Abancay, que contabilizou mais de 1300 pacientes
diagnosticados nesses 27 anos.
Figura 1. Alguns casos reportados de esporotricose no Peru, Estados Unidos da América e Austrália após
a classificação do agente etiológico no gênero Sporothrix.
Fonte: CARMICHAEL, 1962; DIXON et al., 1991; RAMÍREZ-SOTO, 2015; MCGUINNESS et al.,
2016
1.1.2 Epidemiologia
O aparecimento de casos de esporotricose e outras micoses emergentes e re-
emergentes em lugares tão diferentes entre si pode estar intimamente ligado à evolução
das sociedades. Hoje, o comércio intenso promovido pela globalização, o fluxo de
migrantes entre países e continentes, além de mudanças biogeográficas podem favorecer
o deslocamento dos microrganismos de áreas endêmicas para zonas onde,
anteriormente, eram pouco ou nunca descritos. O exemplo citado anteriormente de
esfagno contaminado distribuído por vários estados norte-americanos, relatado por
Dixon et al. (1991) reforça essa idéia.
A esporotricose é uma doença cosmopolita, com uma grande quantidade de
relatos em regiões de clima tropical, subtropical ou temperado, havendo áreas de
hiperendemicidade. O agente etiológico tem predileção por áreas quentes e com alta
umidade, fazendo com que determinados países como Brasil, Peru, Uruguai, México,
Estados Unidos, França, Índia, China, Japão, Austrália, tenham casos confirmados
(MAGAND et al., 2009; BORGES et al., 2013; MAHAJAN, 2014; ZHOU, et al., 2014;
CHAKRABARTI et al., 2015; MCGUINNESS et al., 2016).

20
A esporotricose é uma doença que acomete ambos os sexos e todas as idades,
porém, os pacientes mais acometidos são do gênero masculino e em plena idade laboral,
pois, como a doença é adquirida, majoritariamente, através de inoculação traumática e o
complexo S. schenckii é um conjunto de fungos saprofíticos, os principais acometidos
são trabalhadores do campo como agricultores, jardineiros, floristas, madeireiros,
fazendeiros e caçadores, cuja atividade é, predominantemente, ocupada por homens,
jovens e adultos (MAHAJAN, 2014). Apesar da grande quantidade de trabalhadores do
campo acometidos por essa doença, novos ramos laborais como médicos veterinários
que cuidam de animais doentes e pesquisadores que trabalham diretamente com
espécies do complexo S. schenckii foram incluídos no grupo de atividades de risco
(REED et al., 1993; YEGNESWARAN et al., 2009). Vários surtos de esporotricose,
relacionados a essas atividades laborais, já foram descritos por pesquisadores em
diversos continentes (TÉLLEZ et al., 2014).
No Brasil, a esporotricose é um caso urgente de saúde pública principalmente no
Estado do Rio de Janeiro, onde, entre 1997 e 2007, 1848 casos de esporotricose humana
foram registrados nos serviços de saúde. Destes casos, 1289 foram registrados somente
nos anos entre 2004 e 2007, demonstrando o enorme crescimento da doença nos últimos
anos. Esta epidemia no Rio de Janeiro apresentou uma peculiaridade que a difere das
outras relatadas ao redor do mundo: as mulheres foram mais acometidas, com uma
razão de 2:1. Provavelmente, este fenômeno está relacionado à transmissão zoonótica
por gatos infectados, não tendo relação com a atividade laboral de campo, onde os
homens são mais acometidos (SILVA et al., 2012). Esta hipótese ganha força quando
são considerados os milhares de casos de esporotricose felina registrados neste estado
(PEREIRA et al., 2014).
Na Região Nordeste do Brasil, poucos casos foram relatados e o aparecimento
da doença é raro, tendo um caso de esporotricose canina relatado em 2006 na cidade de
Mossoró, estado do Rio Grande do Norte (FILGUEIRA, 2009) e um caso de
esporotricose felina em 2009 na cidade de Itaporanga, estado da Paraíba (NUNES et al.,
2011). No Ceará, até o presente momento, não existem casos relatados de esporotricose,
seja humana ou animal.

21
É importante salientar que não somente o homem e os gatos são acometidos pela
esporotricose, mas outros mamíferos como cães, tatus e cavalos podem apresentar
manifestações da doença (HAY & MORRIS-JONES, 2008; ROMEO & CRISEO,
2013).
1.1.3 Patogenia
Enquanto micoses como a cromoblastomicose, feohifomicose, micetomas,
zigomicoses, hialohifomicoses e doença de Jorge Lobo têm prevalência em áreas mais
restritas, a esporotricose, causada pelo complexo Sporothrix schenckii, é a micose
subcutânea mais relatada ao redor do mundo (MAHAJAN, 2014).
A esporotricose é uma doença subcutânea crônica e granulomatosa de humanos
e mamíferos, com distribuição mundial e presente, principalmente, em regiões tropicais
e subtropicais, relacionada, geralmente, à inoculação traumática, que, muitas vezes,
pode acontecer com espinhos ou farpas contaminadas pelos agentes etiológicos,
espécies do complexo Sporothrix schenckii (RODRIGUES et al., 2013a; ROMEO &
CRISEO, 2013), os únicos fungos dimórficos causadores de micose subcutânea
primária, diferentemente da histoplasmose ou coccidioidomicose, causadas por
Histoplasma spp. e Coccidioides spp., que são micoses profundas (HOGAN et al.,
1996).
As manifestações clínicas da esporotricose estão classificadas como formas
cutânea fixa, cutâneo-linfática, disseminada e extracutânea. Apesar de ser considerada
uma doença subcutânea e suas lesões serem visualizadas na pele ou nas adjacências de
vasos linfáticos (sua forma mais comum), a esporotricose pode apresentar-se
disseminada a outros órgãos, como os pulmões (forma pulmonar, quando os conídios
são inalados) ou, mais raramente, ter disseminação sistêmica (AUNG et al., 2013;
ALMEIDA-PAES et al., 2014). Porém, fatores do hospedeiro ou do microrganismo
influenciam na mudança do perfil da doença como o status imunológico do hospedeiro,
virulência da cepa inoculada, dentre outros (BARROS et al., 2011). Alguns desses casos
de esporotricose disseminada foram reportados por alguns autores a partir do advento da
aids, como esporotricose cerebral, ocular, endocárdica ou com comprometimento
praticamente total da pele, havendo, em alguns lugares, um aumento do número de
casos em pacientes HIV-positivos, em detrimento de outras doenças como

22
histoplasmose, paracoccidioidomicose e criptococose (SILVA-VERGARA et al., 2012;
FREITAS et al., 2014; PAIXÃO et al., 2015).
Figura 2. A. Esporotricose cutâneo-fixa-fixa: lesão desenvolvida no local de inoculação. B. Esporotricose
cutâneo-linfática: Lesões nódulo-ulcerativas surgem ao longo dos vasos linfáticos, a partir da lesão
inicial.
Fonte: MAHAJAN, 2014.
1.2 Agentes etiológicos - Sporothrix spp.
1.2.1 Taxonomia
Na classificação atual, S. schenckii pertence ao reino Fungi, divisão Ascomycota,
ordem Ophiostomatales, família Ophiostomataceae, gênero Sporothrix e é naturalmente
encontrado em substratos de solo rico em plantas em decomposição, excretas de animais
e celulose, com pH entre 3,5 a 9,4, temperatura próxima a 31 °C e umidade relativa do
ar acima de 92% (LOPES-BEZERRA et al., 2006; TÉLLEZ et al., 2014).
Ao longo de muitas décadas, classificaram-se taxonomicamente os agentes
etiológicos da esporotricose como sendo uma única espécie, denominada Sporothrix
schenckii. Porém, em 2007, baseada em análises fenotípicas e genotípicas, uma nova
classificação foi adotada, descrevendo-se seis novas espécies para o complexo
Sporothrix schenckii: (1) S. globosa, (2) S. brasiliensis, (3) S. mexicana, (4) S. luriei, (5)
S. schenckii sensu stricto e (6) S. albicans, esta última sem relato de causar
esporotricose humana ou animal. Esta nova forma de se classificar foi baseada em
observações macroscópicas (crescimento a 30 °C, 35 °C, 37 °C), morfologia dos
conídios, assimilação de sacarose e rafinose, sequência do gene nuclear da calmodulina
e MALDI-TOF (MENDOZA et al., 2005; MARIMON et al., 2007; OLIVEIRA et al.,
2014). As novas espécies descritas possuem diferenças significativas em relação a
A B

23
cariótipo, fatores de virulência relacionados a polimorfismos genéticos e distribuição
mundial (FERNANDES et al., 2013; RODRIGUES et al., 2013b; SASAKI et al., 2014),
o que implica em uma diferença de resposta contra os principais antifúngicos utilizados
na esporotricose (MARIMON et al,. 2007).
Estudos filogenéticos como o de Teixeira et al. (2014) mostram que espécies
como S. schenckii s. str. e S. brasiliensis possuem cerca de 97,5 % de sequências
genéticas idênticas entre si e possuem 10293 e 9091 genes codificadores de proteínas,
respectivamente, bem similares a um outro fungo, patógeno de plantas, chamado
Grosmannia clavigera. Nas espécies do gênero Sporothrix, percebeu-se a falta de genes
codificadores de liases polissacarídicas, responsáveis por problemas causados em
vegetais, podendo-se supor que esses fungos sofreram um processo adaptativo que os
fez deixar de serem patógenos vegetais para infectarem e causarem doenças em animais.
Além disso, análises filogenéticas demonstraram que S. schenckii s. str. e S. brasiliensis
iniciaram sua divergência de especiação “recentemente”, a cerca de 3,8 a 4,9 milhões de
anos.
Através de estudos baseados em análises do gene da calmodulina e região ITS
do cromossomo fúngico pode-se diferenciar as espécies do complexo S. schenckii. O
perfil eletroforético do cariótipo de cepas de S. brasiliensis já demonstraram que esta
espécies possui menor variabilidade genética que S. schenckii s. str., por exemplo.
Variações no comprimento dos cromossomos são comuns em espécies fúngicas, porém,
muitas vezes, os mecanismos envolvidos em polimorfismos não estão bem esclarecidos
e podem ter correlação com recombinações em sequências repetidas, quebras e
reparações cromossômicas, recombinações entre regiões subteloméricas ou a perda de
cromossomos que contém sequências não necessárias ao crescimento. Os polimorfismos
cromossômicos podem estar intimamente ligados às diferenças encontradas entre as
espécies do complexo S. schenckii em relação à virulência (SASAKI et al, 2014).
Com o avanço das técnicas de identificação e classificação de microrganismos, a
questão taxonômica das espécies vem mudando constantemente, com diversas
proposições para a criação de novas espécies, inclusive no gênero Sporothrix spp., com
a inclusão da espécie Sporothrix chilensis sp. nov., identificada em casos de

24
onicomicose no Chile através de análises multigênicas (RODRIGUES et al., 2016) e
também as espécies S. stylites, S. humicola e S. lignivora (DE MEYER et al., 2008)
1.2.2 Morfologia
O complexo Sporothrix schenckii tem característica termodimórfica, ou seja,
apresentam-se na fase leveduriforme quando em temperatura de 37 °C ou filamentosa
quando se encontram em temperatura entre 25 °C e 30 °C, além de serem saprófitos e
geofílicos (BARROS et al., 2011; RODRIGUES et al., 2013a; SASAKI et al., 2014).
Em laboratório, quando em crescimento em agar Sabouraud dextrose (o meio de cultura
preferencial para seu crescimento) a 25 °C, as colônias de Sporothrix spp. apresentam-
se como colônias planas, úmidas e coloração entre o branco e o marfim. Porém, com o
passar dos dias, as colônias podem adquirir cor marrom ou negra e aspecto rugoso.
Microscopicamente, estes fungos possuem hifas delgadas, hialinas, septadas e
ramificadas, medindo de 1,5 a 2,0 µm de espessura, com pequenos conídios medindo
entre 2,0 a 6,0 µm, piriformes de paredes finas, hialinos, dispostos em cachos que
lembram “flores de margarida” nas pontas dos conidióforos. Com o passar do tempo, os
conídios apresentam-se com paredes mais espessas, ovais ou esféricos e com coloração
negra, o que os dá aspecto dematiáceo, uma característica clássica das espécies do
complexo S. schenckii (TRAVASSOS & LLOYD, 1980; MARQUES-EVANGELISTA
et al., 2010).
Suas colônias leveduriformes, quando crescidas em laboratório em ágar BHI
acrescido ou não de sangue de carneiro e incubadas a 37 °C, possuem,
macroscopicamente, coloração entre branca e creme, superfície irregular, úmida e
aparência cremosa. Ao observá-las ao microscópio, as células leveduriformes podem ser
ovais ou esféricas, com 2,0 a 6,0 µm de diâmetro, ou em formato de charuto, geralmente
com brotamentos ligados a uma célula central (MARQUES-EVANGELISTA et al.,
2010).

25
Figura 3. Sporothrix spp. em sua fase filamentosa. A. Macromorfologia das cepas melanizadas (escuras)
e não-melanizadas (brancas). B. Hifas e conídios em forma de “flor de margarida” (destacados por setas).
C. Conídios de Sporothrix brasiliensis em microscopia eletrônica de varredura – Magnitude 10999X.
Fonte: CEMM, 2016.
A
B
C

26
1.2.3 Distribuição mundial do complexo Sporothrix schenckii
Durante mais de um século, o agente causador da esporotricose foi conhecido
como uma única espécie do gênero Sporothrix. Esta afirmativa ainda era considerada
uma verdade completa até 2007, quando se percebeu através de estudos fenotípicos,
genotípicos e metabólicos que, na realidade, os agentes etiológicos faziam parte deste
gênero, mas eram espécies diferentes, que foram agrupadas em um complexo. Se as
características fenotípicas se mostram muito parecidas, as características genotípicas são
bem distintas entre si, resultando em grandes modificações onde, antes, se imaginava
haver unidade absoluta (MARIMON et al., 2007).
Atualmente, devem-se levar em consideração diversos aspectos relacionados à
epidemiologia das espécies do complexo Sporothrix schenckii, como as diferenças de
distribuição destas, as capacidades de cada uma causar a esporotricose, seus fatores de
virulência e as formas principais de transmissão relacionadas a cada espécie. Dentre as
espécies deste complexo, S. brasiliensis (associada, principalmente, às infecções
animais e transmissão zoonótica), S. schenckii s. str., S. globosa (associada à
transmissão ambiental) e S. mexicana são as espécies mais encontradas nos pacientes
acometidos (GREMIÃO et al., 2015).
Como citado anteriormente, a predileção do gênero Sporothrix por solos ricos
em substratos, principalmente de plantas em decomposição e excretas, além de
ambientes com altíssima umidade relativa do ar, faz com que esses microrganismos
tendam a ser encontrados em regiões cujo clima seja favorável, relacionando a presença
deste gênero, em especial, a regiões úmidas e de clima tropical ou microambientes que
tragam essas características, como celeiros e minas, apesar do gênero Sporothrix ser
encontrado em quase todos os continentes (MAHAJAN, 2014; SASAKI et al., 2014;
TÉLLEZ et al., 2014).

27
Figura 4. Distribuição mundial das espécies S. brasiliensis, S. globosa e Sporothrix s. str. (Clades IIa e IIb).
Fonte: ZHOU et al., 2013; CHAKRABARTI et al., 2015.

28
1.2.4 Fatores de Virulência
Para ter a capacidade de infectar diversos nichos no hospedeiro, os
microrganismos podem apresentar diversos tipos de adaptações, alguns desses sendo
conhecidos fatores de virulência. Esses fatores podem auxiliar na entrada do
microrganismo para regiões específicas do hospedeiro ou até mesmo inibir a ação do
sistema imunitário (NICHOLLS et al., 2011; SILVA et al., 2011). A interação entre os
microrganismos e outros organismos presentes no ambiente pode colaborar para que os
microrganismos adquiram determinadas estratégias de sobrevivência. Estas estratégias
são os fatores de virulência, que interferem na patogenia de determinada infecção,
quando o microrganismo encontra um hospedeiro animal (CASADEVALL, 2012).
Incluídos na teoria acima relatada, os fatores de virulência de S. schenckii
também devem estar relacionados com sua interação com o ambiente. Steenbergen et al.
(2004) sugerem que, quando células leveduriformes de S. schenckii são ingeridas por
uma ameba do solo de vida livre denominada Acanthamoeba castellanii, são capazes
não somente de sobreviver, mas de matar o protozoário e utilizá-lo como nutriente.
Candida albicans e outros fungos que não possuem o solo como habitat não possuem
essa capacidade quando em contato com o mesmo protozoário.
Diversos estudos demonstram os fatores clássicos de virulência das espécies do
complexo Sporothrix schenckii e as diferenças relacionadas em cada uma delas. S.
brasiliensis e S. schenckii s. str. foram consideradas nesses estudos como as espécies
mais virulentas, enquanto S. globosa e S. mexicana não demonstraram ou demonstraram
pouca virulência em modelo murino (ARRIGALA-MONCRIEFF et al., 2009;
FERNÁNDEZ-SILVA et al., 2012; FERNANDES et al., 2013 RODRIGUES et al.,
2014).
Sporothrix spp. possui a capacidade de resistir a condições ambientais
relativamente difíceis, mostrando versatilidade para sobreviver nessas situações, como
temperaturas muito baixas por vários anos e grandes pressões osmóticas (MENDOZA et
al., 2005). Além disso, demonstrou-se que cepas de Sporothrix spp. respondem de
forma diferente a incidências de radiações, como raios ultra-violeta ou radiação gama,

29
podendo manter sua viabilidade, mesmo com variações morfológicas causadas por essas
radiações (TORRES-GUERRER & ARENAS-LÓPEZ, 1998; LACERDA et al., 2011).
Além disso, com o desenrolar da evolução, Sporothrix spp., como diversos
microrganismos, desenvolveu formas de captar íons ferro, imprescindíveis para seu
desenvolvimento, como a produção de sideróforos, moléculas capazes de quelar íons
ferro formando um complexo, mantendo a quantidade necessária deste íon mesmo em
condições desfavoráveis, seja no ambiente ou no hospedeiro (PÉREZ-SANCHÉZ et al.,
2010).
Alguns fatores de virulência já foram descritos para as espécies do complexo
Sporothrix schenckii, como dimorfismo (GAUTHIER & KLEIN, 2008), produção de
melanina (ALMEIDA-PAES, 2009) e enzimas hidrolíticas, termotolerância
(ALMEIDA-PAES et al., 2015), adesinas (TEIXEIRA et al., 2009), proteinase (DA
ROSA et al., 2009) e catalase (WANG et al., 2008).
Algumas espécies fúngicas de importância médica e veterinária têm uma
característica peculiar e importante: possuem a capacidade de modificar sua morfologia
de acordo, principalmente, com a temperatura do ambiente que estão inseridos. Esses
fungos são chamados de dimórficos e crescem na forma de levedura quando a 37 °C
(forma parasitária) e como fungo filamentoso quando entre 25 a 25 °C (forma
infectante) (SASAKI et al., 2014). Alguns exemplos são Blastomyces dermatitidis,
Coccidioides immitis, Coccidioides posadasii, Histoplasma capsulatum var.
capsulatum, Histoplasma capsulatum var. duboisii, Paracoccidioides lutzii,
Paracoccidioides brasiliensis, Penicillium marneffei e Sporothrix spp. (HOGAN et al.,
1996; TÉLLEZ et al., 2014).
Estudos com fungos dimórficos, como H. capsulatum, mostram que a reversão
da fase filamentosa para a fase leveduriforme é um passo imprescindível para o
estabelecimento da doença no hospedeiro (MEDOFF et al., 1987; KLEIN & TEBBETS,
2007), provavelmente devido a uma maior proteção encontrada pelo microrganismo
contra o sistema imunológico. Este fator deve ser, também, de extrema importância para
os outros fungos com essa característica. Blastomyces dermatitidis, outro fungo
dimórfico, quando na fase leveduriforme, é muito grande para ser fagocitado por
neutrófilos polimorfonucleares, diferente de seus pequenos conídios, que sofrem a ação

30
tantos destes neutrófilos quanto de monócitos presentes em sangue periférico (DRUTZ
& FREY, 1985; KLEIN & TEBBETS, 2007). Coccidioides posadasii, fungo dimórfico
encontrado no Nordeste brasileiro, também possui a capacidade de evadir-se do sistema
imune. Os neutrófilos polimorfonucleares conseguem fagocitar somente múltiplos
endósporos deste fungo, que possuem entre 2 e 7 µm, mas não são capazes de fagocitar
estruturas maiores, como suas esférulas, com tamanhos maiores que 11 µm (LEE et al.,
2015).
Diversos mecanismos já foram estudados para revelar os caminhos utilizados
pelos fungos dimórficos para alcançarem a reversão entre as fases morfológicas, mas,
alguns em especial, demonstraram sucesso em seus resultados. Segundo Valentín-
Berríos et al. (2009), um gene de subunidade α (40,9 kDa) de proteína G, denominado
SSG-2, presente em Sporothrix schenckii, participa ativamente do dimorfismo desse
fungo através de interação com fosfolipase A2 citosólica.
Descobertas genéticas vêm ajudando os pesquisadores a compreender cada vez
mais quais genes estão diretamente ligados à capacidade de transição entre fases. Para S.
schenckii, um gene denominado sscmk1 capaz de decodificar a produção de uma
proteína-quinase dependente de Ca2+
/calmodulina (CaMK) parece ter fundamental
importância nesse processo, pois, quando experimentos inibem esta proteína-quinase, a
reversão da fase leveduriformes para filamentosa é anulada (VALLE-AVILES et al.,
2007; VALENTIN-BERRIOS et al., 2009). Além disso, quando a há a supressão da
expressão do gene sscmk1, as leveduras de S. schenckii não conseguem crescer em
temperaturas próximas a 35 °C e mostram que o fungo tem uma perda de
termotolerância (RODRIGUEZ-CABAN et al., 2011).
Outras proteínas também podem estar relacionadas à reversão de fases, como
demonstrou estudos de Hou et al. (2013), que demonstraram que uma histidina-quinase
híbrida é possivelmente reguladora de dimorfismo, pois, em S. schenckii, quando
estudada através de PCR em tempo real, há uma maior expressividade na fase
leveduriforme em detrimento da fase filamentosa.
Os pigmentos de melanina, produzidos por espécies do complexo S. schenckii,
são polímeros com multi-funções, com estrutura molecular diversa, de coloração negra
ou marrom escura, encontrados em toda a natureza e produzidos por uma imensa

31
variedade de organismos, incluindo bactérias, outros fungos, vegetais e animais através
de polimerização oxidativa de compostos fenólicos e indólicos (TABORDA et al.,
2008; ALMEIDA-PAES et al., 2012).
Como dito anteriormente, uma grande diversidade de fungos é produtora de
melanina e esta substância é considerada um importante fator na sobrevivência dos
microrganismos que a produzem, seja na natureza, protegendo-os contra a radiação
ultravioleta ou no hospedeiro, onde encontrarão condições também desfavoráveis em
alguns aspectos, como o ataque do sistema imunológico (JACOBSEN, 2000; LIU &
NIZET, 2009).
A melanina pode ser produzida pelos sistemas orgânicos por várias vias. S.
schenckii possui algumas vias conhecidas, como a produção desse pigmento derivado
da dihidrofenilalanina (L-DOPA), do 1,8-dihidroxinaftaleno (DHN) e também a
produção de piomelanina, sintetizada na presença de L-tirosina (ALMEIDA-PAES et
al., 2012).
A parede celular é uma estrutura de extrema importância para os fungos. Para
um patógeno por excelência, a capacidade de adesão e colonização de tecidos de um
hospedeiro é uma etapa crucial no processo infectante e a parede celular é um
protagonista nesse sentido, tendo as modificações estruturais e de composição como o
maior mecanismo de adaptação para invadir os microambientes do hospedeiro
(SHEPARDSON & CRAMER, 2013).
Alguns “padrões moleculares” em agentes patogênicos, conhecidos como
“PAMPs”, como polímeros de carboidratos e proteínas, estão presentes na parede
celular e, ao se ligarem aos receptores em células imunológicas, desencadeiam a
resposta imune (LATGÉ, 2010). Porém, essa estrutura celular vai muito além de um
sinalizador imunitário. Moléculas de adesão estão presentes na superfície da parece
celular e medeiam um interação com componentes da matriz extracelular e células do
hospedeiro. (TRONCHIN et al., 2008).
Um dos principais componentes da parede celular de Sporothrix spp. é um
glicopeptídeo chamado peptídeo-ramnomanana (57% de manose, 33,5% de ramnose e
14,2% de proteína) (LLOYD & BITOON, 1971; LOPES-BEZERRA, 2011).

32
Uma glicoproteína antigênica de 60 kDa (Gp60) foi encontrada somente na
parede celular das leveduras de Sporothrix schenckii, enquanto uma glicoproteína,
também antigênica, de 70 kDa (Gp70) foi encontrada em ambas as morfologias e
mostrou-se a mais prevalente. Além de estimular a resposta imune, a glicoproteína
Gp70 mostra fortes evidências de fazer parte, como adesina, da interação entre o fungo
e a fibronectina presente na matriz extracelular (RUIZ-BACA et al., 2009; TEIXEIRA
et al., 2009; RUIZ-BACA et al., 2011).
Além disso, S. schenckii tem a capacidade de produzir vesículas que medem
entre 51 e 100 nm, com membrana fosfolipídica que, provavelmente, estão relacionadas
com a transferência de substâncias como moléculas periplasmáticas e componentes de
pigmento entre a membrana plasmática e o espaço extracelular. Essas membranas
podem também estar relacionadas com muitos aspectos de virulência desse patógeno,
pois, a secreção de moléculas fosfolipídicas ou proteínas responsivas a stress ambiental,
ligadas à patogênese, à arquitetura da parece celular, à fisiologia fúngica, dentre outras
funções (ALBUQUERQUE et al., 2008).
Outros estudos mostram que muitas substâncias presentes na parede celular de
espécies do complexo S. schenckii podem ser responsáveis pela virulência das cepas,
como a maior presença de ramnomana em relação à manose na parece de conídios mais
virulentos (FERNANDES et al., 2000), a formação de ergosterol peróxido, que
auxiliaria na evasão do fungo às espécies reativas de oxigênio da fagocitose (SGARBI
et al., 1997) e ao ácido siálico presente na superfície de S. schenckii ,que pode estar
relacionado à proteção da célula contra a fagocitose promovida pelo sistema
imunológico do hospedeiro (ALVIANO et al., 1999).
1.2.5 Biofilmes
Nas últimas décadas estamos vivendo um grande problema de saúde pública
relacionado às doenças infecciosas, sobretudo em pacientes imunocomprometidos. A
formação de biofilmes microbianos é um fator de extrema importância no entendimento
das infecções, pois estão, muitas vezes, intimamente associados a dispositivos médico-
hospitalares de grande utilização, como cateteres, tubos endotraqueais, válvulas
cardíacas, marca-passos, entre outros (ROBERTS, 2013).

33
Os biofilmes, formados por células denominadas sésseis, fazem parte de uma
descoberta relativamente recente para a microbiologia. Eles são considerados
comunidades microbianas que podem estar aderidos às superfícies bióticas ou abióticas,
envolvidos por uma matriz polimérica extracelular, com um fenótipo divergente das
células planctônicas em relação a diversos fatores genéticos ou morfológicos, incluindo
processos de crescimento, metabolismo, transcrição gênica e resistência a drogas
antimicrobianas. Esses biofilmes têm a finalidade de proteger as comunidades
microbianas dos fatores ambientais adversos, que podem comprometer a viabilidade
celular, como pH, desidratação, substâncias químicas, dentre outras (RAMAGE et al.,
2006; TOURNU & VAN DIJCK, 2012).
Em estudos com biofilmes bacterianos, verificou-se que a matriz extracelular
encontrada era composta por quatro componentes básicos: exopolissacarídeos, DNA
extracelular, diversos tipos de proteínas e vesículas, que auxiliavam esses biofilmes a
burlarem o sistema imunológico do hospedeiro, diminuindo sua capacidade de
fagocitose, retendo substâncias antimicrobianas produzidas pelo hospedeiro e, muitas
vezes, promovendo a produção, pelas células bacterianas, de fatores que limitam a
capacidade oxidativa e não-oxidativa das células fagocitárias (ALLEWELL, 2016;
GUNN et al., 2016).
Porém, as conclusões que diversos estudos sobre biofilmes microbianos
obtiveram em relação à composição da matriz dos biofilmes é que a mesma depende de
muitos fatores, que incluem desde diferenças entre biofilmes procariontes e eucariontes
ao microambiente onde o biofilme está sendo formado (DESAI et al., 2014;
ALLEWELL, 2016). Biofilmes produzidos in vivo e em algumas condições in vitro por
Aspergillus fumigatus possuem como componentes de matriz: galactomanana, α-1,3
glucano, monossacarídeos, polióis, melanina e proteínas, bem diferentes da composição
de biofilmes bacterianos e de outras espécies fúngicas, como Candida spp. (AL-
FATANI & DOUGLAS, 2006; BEUVAIS & LATGÉ, 2015; GUNN et al., 2016).
Para a formação da comunidade fúngica em biofilme, algumas etapas são
necessárias. A primeira é a adesão dos conídios ou blastoconídios em uma superfície,
seja ela biótica ou abiótica. Após a adesão, ocorre um processo de intensa filamentação
(para fungos filamentosos ou fungos dimórficos nessa fase) e multiplicação, que pode

34
resultar em uma rede densa unida a uma matriz exopolissacarídica (AL-FATANI &
DOUGLAS, 2006).
Essa transição entre as fases planctônicas e sésseis durante a formação do
biofilme envolve diversos sistemas celulares, que podem ser metabólicos ou
fisiológicos, com modificações fenotípicas advindas de sinalizações intracelulares
responsivas a estímulos ambientais (VALENTINI & FILLOUX, 2016).
Como dito anteriormente, os biofilmes de diversas espécies microbianas podem
ser encontrados aderidos a muitos dispositivos utilizados na medicina e estes possuem
capacidade de resistência aos processos químicos e físicos de desinfecção e
esterilização, mostrando-se, com isso, um problema médico importante (TAFF et al.,
2012; ROBERTS, 2013). Após o processo de maturação do biofilme, este se torna mais
resistente também a ação de agentes antimicrobianos e fatores imunitários do
hospedeiro, quando comparado às células planctônicas, pois a complexa arquitetura do
biofilme, sua matriz, o aumento na expressão das bombas de efluxo de drogas e a
plasticidade metabólica aumentam sua resistência (FINKEL & MITCHELL, 2011;
FANNING & MITCHELL, 2012).
A composição dos biofilmes da cada espécie é bastante variável. Espécies de um
mesmo gênero podem formar biofilmes com composições bem distintas entre si, o que
pode modificar, inclusive, a resistência que cada microrganismo possui às adversidades.
Como exemplo, temos os biofilmes de espécies do gênero Candida. Enquanto o
biofilme formado por Candida albicans possui alta concentração de glicose em sua
matriz, o biofilme de Candida tropicalis tem como principal componente a hexosamina
(AL-FATANI & DOUGLAS, 2006).
Alguns estudos, como o de CHATZIMOSCHOU et al. (2011) verificaram
claramente a capacidade dos biofilmes de fornecerem uma intrínseca resistência aos
agentes antimicrobianos. Neste estudo, biofilmes de espécies de Candida foram
submetidos a testes com equinocandinas e triazóis, que se mostraram menos eficazes
quando em comparação à ação em células planctônicas.
Muitos desses mecanismos de resistência são fontes de estudo importantes para a
comparação entre a as formas planctônicas e sésseis dos microrganismos. A resistência

35
antifúngica em biofilmes pode ser explicada devido a mudanças genéticas irreversíveis
causadas pela longa exposição dos fungos a esses componentes. Essas mudanças podem
causar a diferenciação de alvos moleculares anteriormente importantes para a atividade
antifúngica, a presença de bombas de efluxo, limitada difusão das drogas no interior do
biofilme, aumento de tolerância e densidade celular (barreira física), fatores essenciais
na demonstração da maior resistência dos biofilmes frente aos antifúngicos quando
comparadas às formas planctônicas (NIIMI et al., 2010).
Os biofilmes, como dito anteriormente, são comunidades microbianas, altamente
organizadas, possuidores de uma arquitetura que permite a difusão de nutrientes e o
expurgo de substância nociva e, quando maduros, são formados por micro-colônias e
canais de água, que auxiliam essas perfusões. A capacidade de comunicação entre as
células formadoras dá-se através de um processo denominado “quorum-sensing”, onde
há a secreção de sinalizadores moleculares dependentes da quantidade populacional do
biofilme, sendo considerado um dos reguladores centrais de formação do biofilme (DE
BEER et al., 1994; MILLER & BASSLER, 2001; PERUMAL et al., 2007).
Figura 5. Etapas de formação dos biofilmes microbianos.
O processo de sinalização por “quorum-sensing” é composto por um peptídeo
indutor, uma histidina-quinase (que é ativada pelo peptídeo indutor) e um regulador de
resposta, uma proteína que, quando fosforilada pela quinase, liga-se a quatro promotores
no cromossoma. Esses promotores estão envolvidos em processos essenciais para a
formação e dispersão do biofilme e os processos de infecção, como a produção de
exotoxinas e exoenzimas, através de interações proteína-RNA (ALLEWELL, 2016;
KAVANAUGH & HORSWILL, 2016).
Por todos os motivos já elencados, pesquisadores de todo o mundo trabalham
para conhecer melhor as características inerentes dos biofilmes das diversas espécies

36
fúngicas, sejam elas leveduriformes, como o gênero Candida spp. (AL-FATANI &
DOUGLAS, 2006), filamentosas, como o gênero Aspergillus spp. (BEAUVAIS &
LATGÉ, 2015) ou dimórficas, como os gêneros Coccidioides spp. (DAVIS et al., 2002)
e Histoplasma spp. (BRILHANTE et al., 2015) assim como a capacidade de resistência
desses biofilmes aos antifúngicos. Deste modo, o estudo sobre os aspectos biológicos
dos biofilmes produzidos pelas espécies do complexo S. schenckii torna-se necessária
para a aquisição de conhecimento sobre essa forma de vida do fungo, já que relatos na
literatura sobre esses biofilmes do gênero Sporothrix são escassos (SÁNCHEZ-
HERRERA et al., 2014).
1.3 Diagnóstico laboratorial da esporotricose
Nos casos suspeitos de esporotricose, a coleta de amostras para a análise
laboratorial é imprescindível. Por ter a forma cutânea como a mais prevalente em todo o
mundo, grande parte da coleta de secreções e tecidos ulcerados provenientes das lesões
é feita com swabs estéreis, que devem ser encaminhados imediatamente para as análises
histológicas e micológicas.
O exame direto pode se mostrar uma forma de auxiliar no diagnóstico, revelando
a presença de leveduras ovaladas ou em “forma de charuto” com 3 a 5 µm de diâmetro,
em exsudatos ou tecidos. Porém, mesmo micologistas experientes podem encontrar
dificuldades devido à pequena concentração de microrganismos nestes espécimes
clínicos (KAUFFMAN et al., 2007).
O diagnóstico sorológico não se mostrou eficaz e a cultura do microrganismo
continua sendo o padrão-ouro para o diagnóstico da esporotricose. É um método de
identificação barato, simples e seguro, onde, em torno de 5 a 8 dias, já se pode
confirmar o agente causador e fornecer o diagnóstico. Os espécimes clínicos
provenientes do paciente que podem ser utilizados para a cultura podem ser os mais
diversos, levando-se em consideração o tipo de acometimento, como secreções ou
raspados de lesões, escarro, líquido sinovial e líquor. Eles devem ser semeados e
incubados a 25 °C, para que seja visualizada a forma filamentosa do complexo S.
schenckii. Porém, para que o diagnóstico seja definitivo, a conversão da fase
filamentosa para a fase leveduriforme deve ser realizada (KAUFFMAN et al., 2007).

37
Figura 6. A. Macromorfologia de colônias filamentosas de Sporothrix schenckii s. str., melanizadas, em
ágar batata dextrose, incubadas a 25 °C. B. Macromorfologia de colônias de Sporothrix schenckii s. str.
em ágar BHI suplementado com 5% de sangue de carneiro, incubadas a 37 °C.
Fonte: CEMM, 2016.
Muitas doenças apresentam manifestações clínicas que podem ser confundidas
com a esporotricose, o que pode dificultar o diagnóstico clínico. Por isso, o diagnóstico
diferencial entre a esporotricose e doenças como a nocardiose, leishmaniose e infecções
atípicas por micobactérias deve ser considerado (SMEGO et al., 1999; DOWNEY et al.,
2015).
Atualmente, após vários estudos que demonstram as diferenças entre as espécies
do complexo S. schenckii relacionadas à sensibilidade às drogas antifúngicas, virulência
e distribuição geográfica das mesmas, a importância de se ter um diagnóstico correto de
esporotricose com a identificação do agente etiológico em nível de espécie se torna cada
vez maior (FERNANDES et al., 2013). A distribuição geográfica das espécies deve ser
levada em consideração, já que espécies como S. schenckii s. str. e S. globosa são
cosmopolitas e responsáveis, principalmente, por infecções por via ambiente, através de
traumas ocasionados por espinhos e galhos contaminados, e S. brasiliensis, a mais
virulenta, é encontrada somente no Brasil e responsável, principalmente, pela grande
epidemia de esporotricose no Rio de Janeiro, através da transmissão zoonótica por
mordidas ou arranhaduras de animais infectados (GREMIÃO et al, 2015).
A B

38
Em decorrência desta necessidade, o diagnóstico molecular torna-se uma
ferramenta importante na identificação precisa do agente etiológico da esporotricose em
cada paciente, para que haja a precocidade diagnóstica e o tratamento adequado,
principalmente em áreas endêmicas.
Em estudo conduzido por Rodrigues et al., (2015), verificou-se a viabilidade da
utilização de marcadores moleculares para o diagnóstico da esporotricose. Foram
utilizados ensaios de Polymerase Chain Reaction (PCR) com primers espécie-
específicos que mostraram-se eficientes e rápidos na identificação das espécies do
gênero Sporothrix, após cerca de 6,5 horas, bem abaixo da média de 21 dias que os
exames micológicos necessitam. Esses pesquisadores demonstraram que essa técnica é
considerada sensível e confiável, além de ser de fácil procedimento e baixo custo.
1.4 Sensibilidade aos antifúngicos e tratamentos propostos
Assim como em várias outras micoses, a esporotricose possui, geralmente, um
longo período de tratamento, sendo necessários de três a seis meses para a cura da
doença. Esse é um fator preponderante em problemas de adesão ao tratamento por parte
dos pacientes, fazendo com que os medicamentos sejam utilizados de maneira indevida
(ou descontinuados) e não ocorra a melhora na condição de saúde (KAUFFMAN,
2000).
Alguns medicamentos, como o iodeto de potássio e o itraconazol, são clássicos
no tratamento da esporotricose e já vêm sendo utilizados há décadas, principalmente
contra as formas cutâneas. Já a anfotericina B é geralmente utilizada em manifestações
clínicas sistêmicas ou extracutâneas. Já a terbinafina pode ser utilizada em
manifestações cutâneas, porém, são menos prescritos na prática clínica do que os
medicamentos azólicos, apesar de estudos demonstrarem que, juntamente com o
itraconazol, praticamente não há aquisição de resistência pelos microrganismos
(MAHAJAN, 2014; ALMEIDA-PAES et al., 2016).

39
1.4.1 Iodeto de Potássio
Quando os profissionais de saúde estudam os possíveis tratamentos que deverão
ser indicados pra cada doença, muitos fatores devem ser levados em consideração. Qual
o agente etiológico, sintomatologia, condições físicas e psicológicas do paciente, dentre
muitos outros. Mas, além dos fatores biológicos, outro é de extrema importância: as
condições sócio-econômicas do paciente.
Neste sentido, no tratamento da esporotricose, o motivo socioeconômico
também deve ser levado em consideração, já que se trata de uma doença que acomete,
principalmente, populações menos favorecidas, como trabalhadores rurais, floristas,
mineiros, dentre outros (MAHAJAN, 2014).
Conhece-se o iodo como útil no tratamento de doenças desde o início do século
XIX, quando era bastante utilizado contra problemas tireoidianos. Anos depois, novos
tratamentos utilizando o iodo foram surgindo e esta substância passou a participar do
arsenal terapêutico contra eritemas inflamatórios, Síndrome de Sweet, pioderma
gangrenoso, além de doenças infecciosas, como a zigomicose e a esporotricose (HORIO
et al., 1983, STERLING & HEYMANN, 2000).
A solução saturada de iodeto de potássio ainda é uma opção viável para o
tratamento da esporotricose, principalmente em países em desenvolvimento, em
algumas de suas formas clínicas (cutânea fixa e cutâneo-linfática), devido sua boa
relação custo-benefício, facilidade de obtenção e relativa segurança. Possui alta
solubilidade em água, é higroscópico e fotossensível (STERLING & HEYMANN,
2000; HASSAN & KEEN, 2012).
O iodo tem baixa absorção através da pele e, quando utilizado por via oral, o
iodeto de potássio é convertido a iodo, que é transportado para a tireóide para ser
armazenado. Porém, nem todo o iodo consegue ser armazenado e a quantidade
excedente é eliminada principalmente através da urina, fezes, saliva, suor e leite, sendo
este último, motivo para evitar a utilização por lactantes. Também atravessa a barreira
placentária, sendo contra-indicado em gestantes (STERLING & HEYMANN, 2000;
KAUFFMAN et al., 2007), já que pode levar a criança a desenvolver hipotireoidismo
neonatal, tireomegalia, obstrução respiratória fetal e parto prolongado (COSTA et al.,

40
2011). Além disso, não deve ser utilizado por pacientes com tireoidopatias conhecidas e
alergia ao iodo (MAHAJAN, 2014).
É contra-indicado também em doenças infecciosas com acometimento sistêmico,
onde órgãos internos foram afetados, como nos casos de esporotricose cutâneo-
disseminada ou extra-cutânea, pois há a possibilidade de diminuição da atividade
imunológica nesses pacientes (KAUFFMAN et al., 2007).
Poucos estudos de sensibilidade “in vitro” com iodeto de potássio contra cepas
de Sporothrix spp. têm sido relatados na literatura, porém, estudos diversos vêm
mostrando como se comporta a esporotricose quando os pacientes são tratados com
doses de iodeto de potássio. Macedo et al. (2015) demonstraram que doses deste
fármaco foram administradas a 102 pacientes com esporotricose e a taxa de cura foi de
77,4%.
1.4.2 Compostos azólicos
Após décadas da utilização do iodeto de potássio no tratamento da esporotricose
(e de maneira empírica), a evolução das pesquisas farmacológicas e da indústria
farmacêutica mundial fez surgirem novos fármacos para o tratamento desta e de outras
doenças infecciosas. Os compostos azólicos, surgidos na segunda metade do século XX
e com grande foco a partir dos anos 80, foram uma grande esperança para várias
micoses, incluindo a esporotricose, que, atualmente, tem o itraconazol como droga de
escolha para a maioria de suas manifestações clínicas (YAMADA et al., 2011).
Os fármacos azólicos são divididos em imidazóis (cetoconazol, miconazol, etc) e
triazóis (itraconazol, voriconazol, fluconazol, etc), e têm seu mecanismo de ação
baseado na inibição da biossíntese do ergosterol (MAHAJAN, 2014).
O ergosterol é um esterol biologicamente importante por vários motivos. É o
principal componente da membrana celular dos fungos e de alguns protozoários,
exercendo função comparada ao colesterol presente na membrana citoplasmática dos
animais. Por não estar presente na membrana dos animais, o ergosterol tornou-se um
dos alvos para o desenvolvimento de antifúngicos seletivos, que não interferissem na
membrana celular destes (MAERTENS, 2004; MAHAJAN, 2014).

41
O principal efeito dos medicamentos azólicos sobre os fungos está na inibição
reversível e não-competitiva de um sistema microssomal dependente da Citocromo
P450, a enzima esterol 14-α-desmetilase. Essa enzima bloqueia desmetilação do átomo
de carbono da posição 14 da molécula de lanosterol, culminando na depleção de
ergosterol e no acúmulo de 14-α-metilesteróis. Essa modificação gera uma instabilidade
na membrana celular, modificando a permeabilidade, a atividade de enzimas como
ATPase e enzimas transportadoras e a síntese da quitina (VANDEN BOSSCHE et al.,
1995; MAERTENS, 2004; CARRILLO-MUÑOZ et al., 2006).
Porém, apesar de todas essas características e dos avanços obtidos para a
terapêutica antifúngica, problemas de não-seletividade de alvos terapêuticos também
geram alguns problemas, como a inibição cruzada da biossíntese de compostos nos
mamíferos por enzimas dependentes da P450, gerando problemas de toxicidade
(CARRILLO-MUÑOZ et al., 2006).
O itraconazol, atualmente, é o principal fármaco utilizado no tratamento das
formas cutâneas e linfo-cutâneas da esporotricose. Porém, mesmo tendo-se bem
definido o papel dos fármacos azólicos nesses casos, sempre é importante um screening
das cepas relacionadas e processos epidêmicos, principalmente para evitar falhas no
tratamento ou recidivas. Estes screenings já demonstraram que o itraconazol e o
posaconazol, em ensaios in vitro, por exemplo, possuem atividade melhor contra cepas
de S. brasiliensis e S. schenckii s. str., quando comparadas às cepas de S. mexicana e S.
globosa. Esses resultados demonstram direta correlação entre a diferença genética e o
perfil de sensibilidade das espécies (RODRIGUES et al., 2014).
Muitas interações medicamentosas também podem ser encontradas, já que, como
citado anteriormente, os fármacos azólicos são inibidores do sistema enzimático P-450
e, desta forma, a administração conjunta com drogas metabolizadas por este sistema,
como varfarina, carbamazepina, fenobarbital, astemizol, rifampicina, midazolam,
sinvastatina, hipoglicemiantes orais, etc, podendo causar toxicidade grave (MAHAJAN,
2014).
Assim como praticamente todos os fármacos antimicrobianos, os azólicos
também sofrem com a problemática da resistência microbiana adquirida. Antes dos anos
90, era muito difícil verificarmos cepas com resistência adquirida aos azólicos. Porém,

42
atualmente, a resistência vem se tornando um problema de saúde pública,
principalmente, pela utilização inadequada de medicamentos como, por exemplo, o
fluconazol (MAERTENS, 2004).
Em estudos com Candida albicans, Maccallum et al. (2010) verificaram que um
dos principais mecanismos de resistência aos fármacos azólicos é devido a modificações
nas enzimas-alvo e na regulação de transportadores desses fármacos. Esses mecanismos,
muitas vezes, estão combinados, não agindo isoladamente em algumas cepas. Essas
descobertas foram possíveis graças à manipulação genética, verificando que essas cepas
conseguem regular genes codificadores de transportadores celulares.
Esses quadros de resistência também vêm aparecendo em estudos com espécies
do complexo Sporothrix schenckii. Rodrigues et al. (2014), em estudo produzido no
Brasil, verificaram a presença de uma cepa de Sporothrix schenckii stricto sensu
advinda de amostra humana com resistência cruzada aos polienos e todos os azólicos
testados in vitro (fluconazol, itraconazol, voriconazol e posaconazol).
Em ensaios de sensibilidade in vitro, alguns estudos demonstram que esses
fármacos possuem atividade antifúngica contra cepas do complexo Sporothrix schenckii
bem variável. Oliveira et al., (2011) demonstraram que o cetoconazol, um dos fármacos
azólicos mais tradicionais, mostrou MICs entre 0,06 a 2 µg/mL quando em contato com
cepas de S. brasiliensis e S. schenckii s. str., enquanto o fluconazol, neste mesmo
estudo, demonstrou MICs para essas cepas entre 16 a 128 µg/mL. Já em ensaios com
posaconazol e itraconazol in vivo demonstraram que estes fármacos foram bastante
eficientes na diminuição da carga fúngica e no tratamento da esporotricose em
camundongos infectados por Sporothrix spp. (ANTUNES et al., 2009; MARIO et al.,
2015).
1.4.3 Anfotericina B
A anfotericina B é um dos antifúngicos mais antigos disponíveis para o
tratamento de infecções micóticas. Faz parte da família dos polienos e faz parte da
prática clínica desde a década de 50 (MATHEW & NATH, 2009).
É um agente de amplo espectro com eficácia demonstrada contra grande parte
dos agentes causadores de micoses sistêmicas, como Paracoccidioides spp.,

43
Histoplasma spp., Coccidioides spp. e Sporothrix spp. Age ligando-se ao ergosterol ou
outros esteróis da membrana fúngica sensível, componentes estruturais essenciais. Essa
ligação provoca a formação de poros aquosos e não aquosos, fazendo com que a
permeabilidade dessa membrana seja alterada e componentes intracelulares como
proteínas e cátions monovalentes (sódio, potássio) e bivalentes sejam perdidos. Isso leva
a perda do potencial da membrana celular, resultando na morte do microrganismo
(MOHR et al., 2008; MATHEW & NATH, 2009).
Atualmente, somente formulações parenterais e tópicas de anfotericina B estão
disponíveis para utilização. Porém, devido a grande quantidade de efeitos adversos
causados por sua administração, novas formulações parenterais foram introduzidas na
prática médica, como formulações complexadas com lipídios, dispersões coloidais e a
anfotericina B lipossomal, que diminui esses efeitos após administração (LANIADO-
LABORÍN & CABRALES-VARGAS, 2009; CHANDRASEKAR, 2011; MAHAJAN
et al., 2015).
Diversos estudos relatam diferenças entre a sensibilidade de cepas e espécies do
complexo S. schenckii frente à Anfotericina B. Rodrigues et al., (2014) verificaram que
este fármaco possui MICs entre 1 a 8 µg/mL contra S. brasiliensis, enquanto contra S.
mexicana os MICs variam entre 8 µg/mL a valores maiores que 16 µg/mL.
Estudo produzido por Ishida et al., (2015) avaliando a sensibilidade de cepas de
S. brasiliensis causadoras de esporotricose disseminada em modelo murino frente à
anfotericina B desoxicolato demonstrou que esse fármaco, quando utilizado por via
parenteral após 7 dias da infecção, se mostrou efetivo em diminuir a carga fúngica nos
modelos infectados, porém, quando o mesmo fármaco atua em conjunto com o
itraconazol administrado por via oral, encontra-se melhor opção terapêutica.
1.4.4 Caspofungina
A caspofungina foi o primeiro fármaco da classe das equinocandinas a ser
licenciado no mundo. É um fármaco semi-sintético derivado da pneumocandina B0,
isolado do fungo Glarea lozoyensis, que age inibindo a síntese de β-(1,3)-D-glucana,
um componente importante da parede celular fúngica. Com o decréscimo desse
componente, causado pela ação do fármaco, ocorre um estresse osmótico, vacuolização

44
e consequente lise e morte celular. A síntese dessa glucana é obtida através de uma
enzima não presente nas células dos mamíferos, denominada β-(1,3)-D-glucana
sintetase, o que a torna um alvo importante para o estudo das propriedades moleculares
antifúngicas (WALSH et al., 2008).
Porém, a caspofungina possui um espectro de ação antifúngica restrito quando
comparado a outros antifúngicos, sendo utilizado, principalmente, contra espécies dos
gêneros Candida e Aspergillus. A caspofungina não se mostrou muito eficaz contra
cepas de espécies do complexo S. schenckii, tendo menor atividade que outros
antifúngicos como itraconazol, cetoconazol, terbinafina e anfotericina B (OLIVEIRA et
al., 2011; BRILHANTE et al., 2016).
1.4.5 Termoterapia
Uma antiga forma de tratamento não medicamentosa que pode ser utilizada em
casos não complicados de esporotricose é a termoterapia. A maioria das espécies do
complexo S. schenckii é sensível e não resistem a temperaturas mais altas, acima de
38,5 °C. A aplicação de calor (42 °C a 43 °C) através de compressas quentes ou
radiação infravermelha nas lesões cutâneas por algumas semanas mostra-se um
tratamento relativamente eficaz, que pode ser ainda mais eficaz quando utilizado em
conjunto com fármacos azólicos ou iodeto de potássio, que, porém, não devem ser
administrados à gestantes (KAUFFMAN et al., 2007; COSTA et al., 2011).
Ainda em estudo de Costa et al. (2011) realizado em um hospital na cidade do
Rio de Janeiro, Brasil, entre 1997 e 2009, 171 pacientes foram admitidos com
esporotricose, sendo, destes, 5 gestantes, o que mostra ser uma doença relativamente
incomum nessa condição. Porém, a cautela deve ser redobrada em relação ao tratamento
que deve ser adotado, pois, as drogas mais utilizadas, atualmente, contra essa doença,
são contra-indicadas em mulheres grávidas, tendo a termoterapia como uma opção
viável. Os derivados azólicos como o cetoconazol, itraconazol e fluconazol devem ser
evitados durante a gravidez, pois possuem razoável potencial teratogênico e
embriotóxico (MOUDGAL & SOBEL, 2003).
Diante do exposto, percebemos que a esporotricose tem afetado a população de
alguns estados brasileiros de maneira preocupante, tornando-se um sério problema de

45
saúde pública, principalmente pela cultura de criação de animais domésticos, dentre eles
os felinos, fonte potencial de transmissão desta doença. Além disso, a formação de
biofilmes microbianos tem demonstrado um aumento no limiar de sensibilidade de
bactérias e fungos frente aos antimicrobianos, dificultando as terapias propostas e
trazendo prognósticos desfavoráveis a muitos pacientes.
Portanto, é necessário o aprofundamento de estudos sobre biofilmes fúngicos e
como os mesmos se comportam frente aos antifúngicos utilizados na prática clínica, a
fim de obter-se um melhoramento nos protocolos de tratamento através do
entendimento da relação do funcionamento destes biofilmes.
2 HIPÓTESES
a) Os biofilmes formados pelas espécies do complexo S. schenckii apresentam
diferenças de metabolismo e biomassa intra-espécie sobre o mesmo
substrato;
b) Os biofilmes das espécies mais virulentas (S. brasiliensis e S. schenckii s.
str.) serão mais espessos e complexos que os das espécies menos virulentas
(S. mexicana e S. globosa);
c) O biofilme de Sporothrix spp. encontra-se organizado de forma a diminuir a
atividade dos antifúngicos contra os microrganismos, quando comparado à
sensibilidade das mesmas cepas em forma planctônica.
3 OBJETIVO GERAL
Conhecer se quatro diferentes espécies do complexo Sporothrix schenckii (S.
schenckii s. str., S. brasiliensis, S. mexicana e S. globosa) são capazes de produzir
biofilmes, in vitro, padronizá-los e determinar seus perfis de sensibilidade frente aos
antifúngicos clássicos.
3.1 Objetivos específicos
3.1.1 Verificar a formação de biofilme, in vitro, por cepas de S. schenckii s. str., S.
brasiliensis, S. mexicana e S. globosa filamentosas;
3.1.2 Determinar a cinética de crescimento, in vitro, dos biofilmes de S. schenckii

46
s. str., S. brasiliensis, S. mexicana e S. globosa filamentosas de acordo com
sua atividade metabólica;
3.1.3 Determinar a cinética de crescimento, in vitro, dos biofilmes de S. schenckii
s. str., S. brasiliensis, S. mexicana e S. globosa filamentosas de acordo com
sua biomassa produzida;
3.1.4 Visualizar a estrutura dos biofilmes de Sporothrix spp. por microscopia
óptica;
3.1.5 Visualizar a estrutura tridimensional dos biofilmes de Sporothrix spp. por
microscopia eletrônica de varredura e por microscopia confocal.
3.1.6 Determinar a concentração inibitória mínima (CIM) e concentração
fungicida mínima (CFM), in vitro, dos antifúngicos azólicos clássicos,
anfotericina B e caspofungina ante as cepas de Sporothrix spp. planctônicas
na fase filamentosa;
3.1.7 Determinar a atividade, in vitro, de concentrações ascendentes dos
antifúngicos azólicos clássicos, anfotericina B e caspofungina ante o
biofilme das cepas de Sporothrix spp. na fase filamentosa.
.
4 MATERIAIS E MÉTODOS
4.1 Microrganismos
Neste estudo, foram utilizadas 18 cepas de espécies do complexo Sporothrix
schenckii na fase filamentosa: 10 cepas de S. brasiliensis, 2 cepas de S. schenckii s. str.,
2 cepas de S. globosa e 4 cepas de S. mexicana. O número maior de cepas da espécie S.
brasiliensis leva em consideração a espécie mais encontrada no Brasil, onde este estudo
foi realizado.
Os isolados pertencem à micoteca do Centro Especializado em Micologia
Médica (CEMM), da Universidade Federal do Ceará (UFC) - Brasil e foram
armazenadas em solução salina a 4 °C. Antes dos experimentos, todos os isolados foram
plaqueados em ágar batata dextrose (PDA) (Himedia, Índia), incubados a 25 °C durante
5 dias e mantidos em fase filamentosa. Todos os experimentos foram realizados no
CEMM e na Central Analítica da UFC. Em todos os experimentos, como controle
positivo, foi utilizada a cepa de Candida parapsilosis ATCC 22019. Como controle
negativo, o procedimento foi repetido, porém, sem a presença de inóculo.

47
Tabela 1. Origem e local de isolamento das cepas de espécies do complexo Sporothrix schenckii
utilizadas neste estudo.
Cepa Espécie Origem Isolamento
CEMM 05-3-050 S. brasiliensis Gato BRASIL
CEMM 05-3-052 S. brasiliensis Cão BRASIL
CEMM 05-3-053 S. brasiliensis Gato BRASIL
CEMM 05-3-054 S. brasiliensis Gato BRASIL
CEMM 05-3-055 S. brasiliensis Gato BRASIL
CEMM 05-3-056 S. brasiliensis Gato BRASIL
CEMM 05-3-057 S. brasiliensis Gato BRASIL
CEMM 05-3-058 S. brasiliensis Gato BRASIL
CEMM 05-3-075 S. brasiliensis Gato BRASIL
CEMM 05-3-078 S. brasiliensis Gato BRASIL
CEMM 05-3-090 S. schenckii s. str. Humano BRASIL
CEMM 05-3-100 S. mexicana Vegetal MÉXICO
CEMM 05-4-001 S. mexicana Solo MÉXICO
CEMM 05-4-002 S. schenckii s. str. Solo BRASIL
CEMM 05-3-004 S. globosa Humano BRASIL
CEMM 05-3-005 S. globosa Humano BRASIL
CEMM 05-3-008 S. mexicana Indeterminada BRASIL
CEMM 05-3-009 S. mexicana Indeterminada BRASIL
4.2 Formação do biofilme filamentoso de Sporothrix spp. para testes de
sensibilidade
A formação de biofilmes filamentosos foi realizada como descrito por Cordeiro
et al. (2015c), com algumas modificações. Todos os isolados (n=18) foram previamente
cultivados em PDA por 5 dias a 25 °C. Duzentos microlitros de inóculos com 2 x 104, 2
x 105 e 2 x 10
6 conídios/mL foram transferidos para poços de placas de microdiluição
para a formação dos biofilmes e foram analisados com 1, 8 e 24 horas de adesão.
Através de análises espectrofotométricas, foram encontradas as melhores condições para
a formação dos biofilmes quando utilizado um inóculo com 2 x 105 conídios/mL, com a
adesão de 24 horas. As células foram suspensas com 0,9 % de salina estéril para se obter
uma concentração de inóculo de 2 x 105 conídios/mL (1,0 McFarland). Alíquotas de
1000 µL de cada inóculo foram transferidas para placas de poliestireno de 24 poços
contendo lamínulas de Thermanox™ (Thermo Fisher Scientific, New York, NY) e
incubadas a 25 °C durante 24 horas, sob condições estáticas, para a etapa de pré-adesão.

48
Depois, o sobrenadante de cada poço foi cuidadosamente removido e foi adicionado
1000 µL de meio RPMI 1640 (Sigma, Alemanha) tamponado a pH 7,0 com MOPS
0,165 M. As placas foram incubadas durante 5 dias a 25 °C, em condições estáticas,
para a formação e maturação dos biofilmes e realização dos ensaios posteriores.
Figura 7. Esquema de metodologia para a formação de biofilmes filamentosos de espécies do complexo
S. schenckii.
4.3 Cinética de crescimento dos biofilmes
Seguindo a metodologia de formação dos biofilmes descrita acima, a cinética de
crescimento foi avaliada e mensurada após diferentes tempos de formação (24, 48, 72,
96, 120 e 144 horas). As culturas fúngicas foram crescidas em PDA incubadas a 25 °C
por 5 dias. Os biofilmes foram cultivados em meio RPMI, pH 7,0 a 25 °C em condições
estáticas. Depois de cada tempo de formação, os sobrenadantes foram descartados e as
placas foram lavadas duas vezes com PBS/Tween estéril para remover as células não
aderidas. A biomassa do biofilme foi avaliada com o método de coloração com cristal
violeta e a sua viabilidade metabólica foi avaliada pelo ensaio de XTT [2,3-Bis-(2-
methoxy-4-nitro-5-sulfophenyl)-2H-tetrazolium-5-carboxanilide salt], de acordo com
Pitangui et al. (2012) e Cordeiro et al. (2015c). Para ensaio de XTT, foram realizados
testes utilizando soluções de XTT (1 mg/mL em PBS) e menadiona (1 mM em etanol).
Os biofilmes foram formados como descrito acima em placas de 96 poços separadas em
diferentes tempos (24, 48, 72, 96, 120 e 144 horas). Em cada tempo de formação, 50 µL
de solução de XTT mais 4 µL de solução de menadiona foram adicionados a todos os
poços que contém biofilme e de controle negativo. Após 3 h de tempo de incubação a
25 °C, no escuro, a mudança de cor foi medida em espectrofotômetro a 492 nm. O

49
ensaio de cristal violeta foi efetuado tal como descrito por Brilhante et al. (2015), com
modificações. As placas foram incubadas a 25 °C e, em cada tempo de formação, os
poços referentes a esses tempos foram lavados três vezes com solução salina a 0,9%
para remover as células não aderidas. Posteriormente, os poços foram lavados com 100
µL de metanol 100% e o sobrenadante foi aspirado. Uma alíquota de 100 µL de cristal
violeta 0,3% foi adicionada a cada poço. Depois de 20 min a 25 °C, a solução de
corante foi aspirada e os poços foram lavados duas vezes com 200 µL de água destilada
estéril. Finalmente, foram adicionados 150 µL de ácido acético 33% aos poços corados
e deixados durante 30 segundos. Após este período, o volume foi transferido para outra
placa, que foi imediatamente lida num espectrofotômetro a 590 nm. O valor de cut-off
para a densidade óptica (DOc) para o ensaio foi definida como três desvios padrão
acima da média DO590 nm do controle negativo. Os experimentos foram realizados em
triplicata.
Para estabelecer uma visão geral de formação de biofilme por espécies do
complexo Sporothrix schenckii, foi utilizada a classificação proposta por Cordeiro et al.
(2015a) como uma maneira de se relacionar valores para a formação de biofilme. Para
avaliar a capacidade formadora de biofilme das diferentes cepas de diferentes espécies
do complexo S. schenckii, os valores de densidade óptica (DO) dos poços não
inoculados com os microrganismos foram usados como controle negativo, definindose
o valor de corte (DOC) para o teste como sendo três desvios-padrão acima da média de
DO do controle negativo a 590 nm. No final, as cepas foram classificadas como não
formadoras (DO ≤ DOc), fracas formadoras (DOc < DO ≤ 2 × DOc), formadora
moderada (2 × DOc < DO ≤ 4 × DOc), e fortes formadoras (4 × DOc < DO). Os
resultados obtidos são representativos de três experimentos independentes, realizados
em triplicata.

50
Figura 8. Esquema de metodologia para a avaliação da cinética de biofilmes filamentosos de espécies do
complexo S. schenckii através de ensaios de biomassa (cristal violeta) e atividade metabólica (XTT).
4.4 Morfologia e estrutura dos biofilmes de Sporothrix spp.
Microscopia óptica. Para a visualização microscópica do biofilme por
microscopia óptica foi utilizada a metodologia por coloração com Vermelho-Congo. A
superfície de uma lamínula de Thermanox™ em poços de placas de poliestireno após a
formação do biofilme maduro foi coberta com cloreto de cetilpiridínio a 10 mM e
deixou-se secar naturalmente, no ar, durante 30 minutos. Em seguida, a superfície foi
corada durante 15 minutos com uma mistura 2:1 de solução aquosa saturada de
Vermelho-Congo contendo 10% (v/v) de solução de Tween 80. Após lavagem com PBS
1X, a lamínula foi corada com uma solução 10% (v/v) de Ziehl carbolfucsina durante 6
minutos. Em seguida, a lamínula de Thermanox™ foi lavada novamente com PBS 1X e
deixada para secar a 37 °C (ALLISON & SUTHERLAND, 1984). As análises de
Thermanox™ foram feitas através de microscópio óptico, modelo OLYMPUS BX41 e
as imagens foram captadas pela câmara OLYMPUS DP71.
Microscopia confocal a laser (CLSM). Os biofilmes formados foram
analisados por microscopia confocal, como descrito por DiBonaventura et al. (2006)
com modificações. Eles foram formados como descrito acima em lamínulas de
Thermanox™. Após o período de incubação, os biofilmes foram lavados com PBS e
corados utilizando corante LIVE/DEAD™ fluorescente (Invitrogen, EUA). Os
biofilmes formados foram analisadas em microscópio confocal Nykon C2, a 488 nm,
para a detecção de corante fluorescente SYTO 9, que identifica as células vivas e a 561
nm para identificação das células mortas por detecção de iodeto de propídio. Várias

51
secções foram obtidos no plano xy com intervalos de 1 µm ao longo do eixo z.
Reconstruções tridimensionais dos biofilmes foram obtidos pelo software residente. As
imagens foram processadas em software Photoshop (Adobe Systems, San Jose, EUA)
Microscopia eletrônica de varredura (MEV). Para obter informações sobre a
morfologia e estrutura dos biofilmes durante a formação e após a maturação (24, 48, 72,
96, 120 e 144 h), utilizaram-se imagens produzidas por microscopia eletrônica de
varredura. Os biofilmes foram formados em Thermanox™ tal como descrito acima e
analisados de acordo com Wen-Ling et al. (2009) e Cordeiro et al. (2015c), com
modificações. Após incubação, o sobrenadante foi removido e as placas foram lavadas
com tampão cacodilato (0,15 M) por duas vezes subsequentes. Os biofilmes foram,
então, cobertos com glutaraldeído (2,5% em tampão cacodilato de sódio 0,15 M com
azul de alcian 0,1 %) e incubou-se a 4 °C overnight. Após esse processo, os biofilmes
foram lavados duas vezes com tampão cacodilato durante 5 minutos e os biofilmes nas
lamínulas de Thermanox™ foram desidratados numa série ascendente de etanol (50, 70,
80, 95 e 100 %) por duas vezes, durante 10 minutos cada. Os biofilmes foram secos a
25 °C e cobertos com hexametildisilazano (HMDS) (Polysciences Europa, Alemanha)
durante 30 minutos. Depois disso, o HMDS foi removido e biofilmes foram secos
durante a noite no dessecador. As lâminas foram cobertas com 10 nm de ouro (Emitech
Q150T) e observadas com um microscópio eletrônico de varredura FEI Inspect S50 no
modo de alto vácuo a 15 kV. As imagens foram processadas no software Photoshop
(Adobe Systems, San Jose, Califórnia), sem nenhuma manipulação de imagem.
4.5 Sensibilidade da forma planctônica
Os ensaios de microdiluição em caldo foram baseados de acordo com Clinical &
Laboratory Standards Institute (CLSI) M38-A2 (2008) com algumas modificações. O
inóculo foi preparado como descrito por Brilhante et al. (2014). As cepas na fase
filamentosa foram preparadas a partir de culturas em PDA, incubadas durante 6 - 7 dias
a 25 °C. Solução salina estéril foi adicionada a cada cultura de fungos e raspagens
foram feitas a partir da superfície do micélio. Em seguida, cada suspensão fúngica foi
ajustada a 2,0 na escala de McFarland. As suspensões foram diluídas na proporção de
1:10 com meio RPMI para se obter inóculos com concentração final de 1-5 x 105
UFC/ml para Sporothrix spp. Este estudo avaliou a concentração inibitória mínima

52
(MIC) para quatro agentes antifúngicos: anfotericina B (AMB) (Sigma Chemical
Corporation, EUA), voriconazol (VRC) (Sigma), fluconazol (FLC) (Sigma),
cetoconazol (KTC) (Sigma); e a concentração efetiva mínima (MEC) para a
caspofungina (CAS) (Sigma). As concentrações finais de drogas variaram de 0,03-16
µg/mL para AMB, VRC, KTC e CAS e 0,12-64 µg/mL para FLC. Para o ensaio de
sensibilidade para a forma filamentosa, as placas foram incubadas a 25 °C e lidas após
72 h. Todos os isolados foram testados em duplicata. Para AMB e VRC, o MIC foi
definido como a concentração mais baixa de droga que inibiu a 100 % de crescimento
fúngico; para KTC e FLC, o MIC foi definido como a concentração mais baixa de droga
que inibia 50 % do crescimento de fungos; para CAS, a MEC foi definida como a
menor concentração de droga que leva ao crescimento de formas pequenas,
arredondadas, com compactação de hifas, em comparação com o crescimento de hifas
observado no controle de crescimento nos poços. Candida parapsilosis ATCC 22019
foi incluída como controle de qualidade. Após a leitura do MIC, alíquotas de 100 µL a
partir de poços que não apresentaram crescimento de fungos visíveis foram transferidos
para tubos contendo PDA, que foram incubados durante 7 dias a 25-25 °C. A
concentração fungicida mínima (MFC) foi definida como a concentração mais baixa
capaz de inibir o crescimento das colônias fúngicas.
Figura 9. Esquema de metodologia para ensaio de sensibilidade antifúngica em espécies do complexo S.
schenckii, em suas fases filamentosas, por microdiluição em caldo.

53
4.6 Atividade metabólica e biomassa dos biofilmes frente aos antifúngicos
Para determinar a resistência dos biofilmes filamentosos às drogas testadas, os
biofilmes foram formados em placas de poliestireno de 96 poços como descrito
anteriormente no item 4.2. Após 5 dias de incubação a 25 °C, o sobrenadante de cada
poço foi aspirado, descartado e, pouco depois, foram adicionados 200 µL de cada
concentração a ser testada (MIC da forma planctônicas, 10xMIC e 50xMIC – ou MEC,
em relação à caspofungina) de cada fármaco (AMB, VRC, FLC, KTC e CAS) em três
poços, respectivamente, cada poço com uma concentração diferente. Os testes foram
feitos em duplicata. As placas foram incubadas durante 72 h a 25 °C, de forma estática,
e, após este período, foram submetidas a testes de quantificação da biomassa, utilizando
a metodologia de cristal violeta e atividade metabólica, utilizando o ensaio com XTT,
tal como descrito anteriormente no item 4.3. Quando obtivemos como resultado um
valor de MIC ou MEC não mensurável, ou seja, acima da maior concentração testada,
esta maior concentração foi considerada como valor de MIC ou MEC para os cálculos
dos testes de sensibilidade dos biofilmes.
Figura 10. Esquema de metodologia para ensaio de atividade metabólica e biomassa de espécies do
complexo S. schenckii, em suas fases filamentosas, frente aos antifúngicos.
4.7 Análises estatísticas
Para a comparação dos dados de absorbância (Densidade óptica - “DO”) intra-
espécie para as drogas testadas, foi utilizada a ANOVA, seguida do teste de
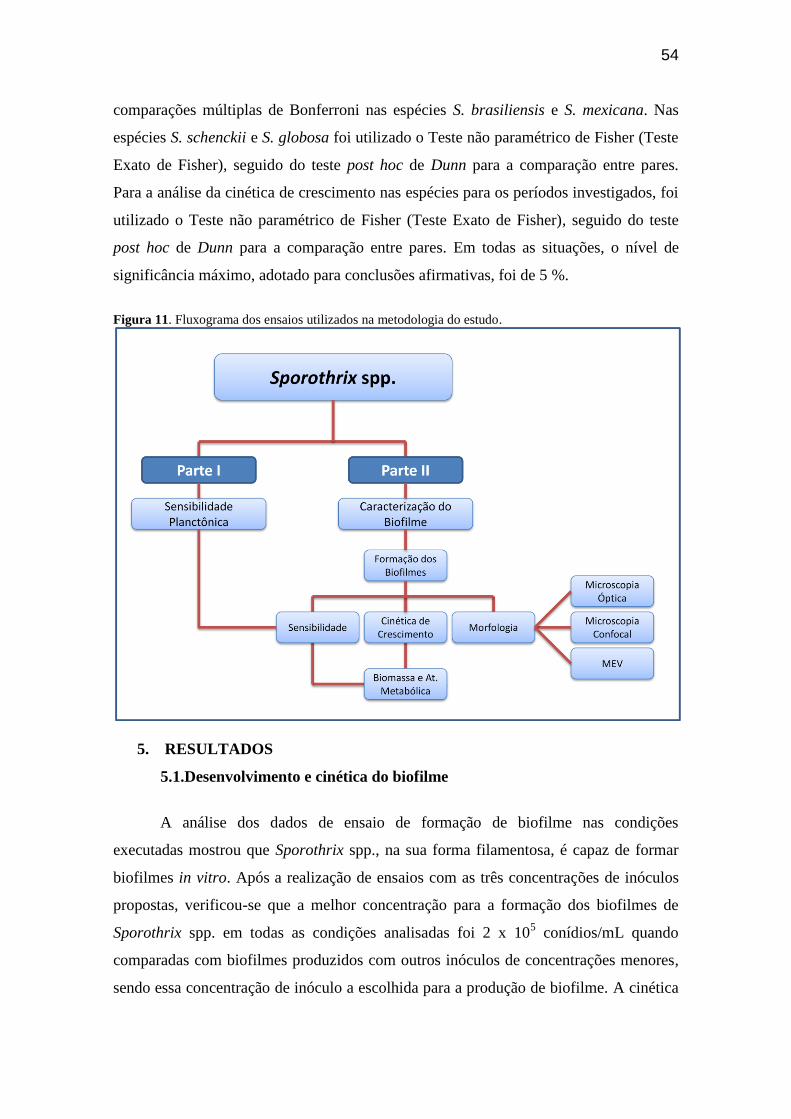
54
comparações múltiplas de Bonferroni nas espécies S. brasiliensis e S. mexicana. Nas
espécies S. schenckii e S. globosa foi utilizado o Teste não paramétrico de Fisher (Teste
Exato de Fisher), seguido do teste post hoc de Dunn para a comparação entre pares.
Para a análise da cinética de crescimento nas espécies para os períodos investigados, foi
utilizado o Teste não paramétrico de Fisher (Teste Exato de Fisher), seguido do teste
post hoc de Dunn para a comparação entre pares. Em todas as situações, o nível de
significância máximo, adotado para conclusões afirmativas, foi de 5 %.
Figura 11. Fluxograma dos ensaios utilizados na metodologia do estudo.
5. RESULTADOS
5.1.Desenvolvimento e cinética do biofilme
A análise dos dados de ensaio de formação de biofilme nas condições
executadas mostrou que Sporothrix spp., na sua forma filamentosa, é capaz de formar
biofilmes in vitro. Após a realização de ensaios com as três concentrações de inóculos
propostas, verificou-se que a melhor concentração para a formação dos biofilmes de
Sporothrix spp. em todas as condições analisadas foi 2 x 105 conídios/mL quando
comparadas com biofilmes produzidos com outros inóculos de concentrações menores,
sendo essa concentração de inóculo a escolhida para a produção de biofilme. A cinética
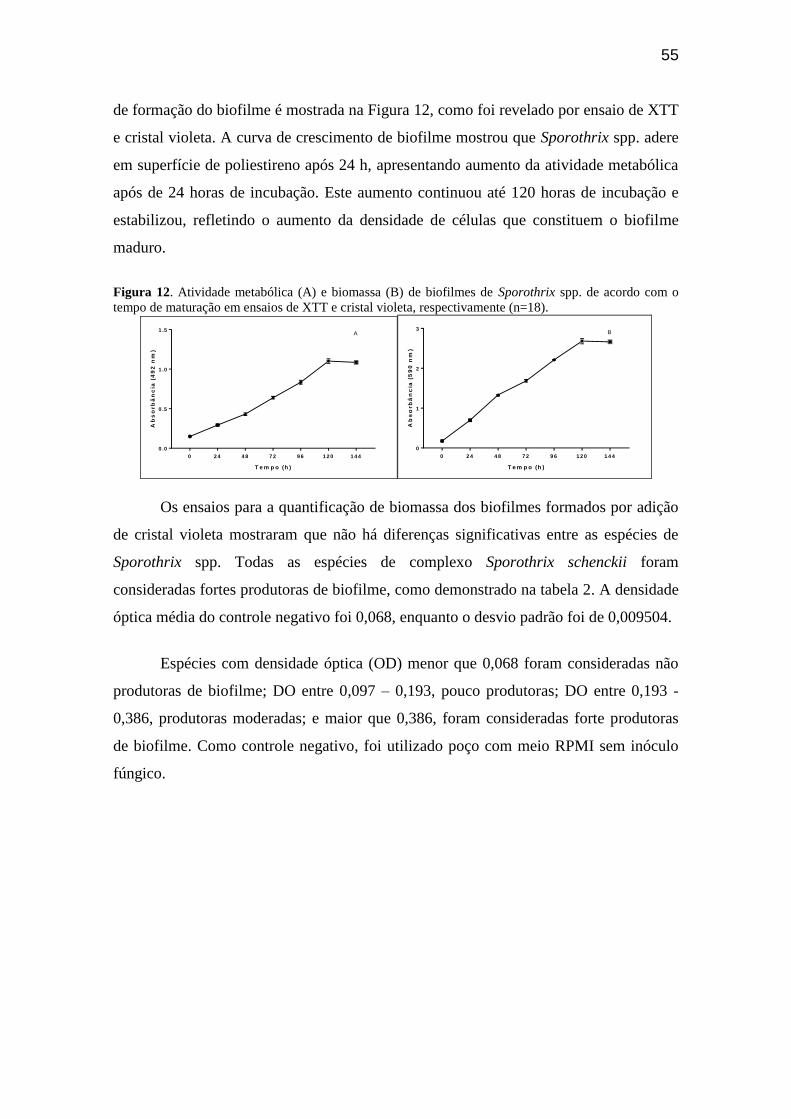
55
de formação do biofilme é mostrada na Figura 12, como foi revelado por ensaio de XTT
e cristal violeta. A curva de crescimento de biofilme mostrou que Sporothrix spp. adere
em superfície de poliestireno após 24 h, apresentando aumento da atividade metabólica
após de 24 horas de incubação. Este aumento continuou até 120 horas de incubação e
estabilizou, refletindo o aumento da densidade de células que constituem o biofilme
maduro.
Figura 12. Atividade metabólica (A) e biomassa (B) de biofilmes de Sporothrix spp. de acordo com o
tempo de maturação em ensaios de XTT e cristal violeta, respectivamente (n=18).
0 2 4 4 8 7 2 9 6 1 2 0 1 4 4
0 .0
0 .5
1 .0
1 .5
T e m p o (h )
Ab
so
rb
ân
cia
(4
92
nm
)
A
0 2 4 4 8 7 2 9 6 1 2 0 1 4 4
0
1
2
3
T e m p o (h )
Ab
so
rb
ân
cia
(5
90
nm
)
B
Os ensaios para a quantificação de biomassa dos biofilmes formados por adição
de cristal violeta mostraram que não há diferenças significativas entre as espécies de
Sporothrix spp. Todas as espécies de complexo Sporothrix schenckii foram
consideradas fortes produtoras de biofilme, como demonstrado na tabela 2. A densidade
óptica média do controle negativo foi 0,068, enquanto o desvio padrão foi de 0,009504.
Espécies com densidade óptica (OD) menor que 0,068 foram consideradas não
produtoras de biofilme; DO entre 0,097 – 0,193, pouco produtoras; DO entre 0,193 -
0,386, produtoras moderadas; e maior que 0,386, foram consideradas forte produtoras
de biofilme. Como controle negativo, foi utilizado poço com meio RPMI sem inóculo
fúngico.
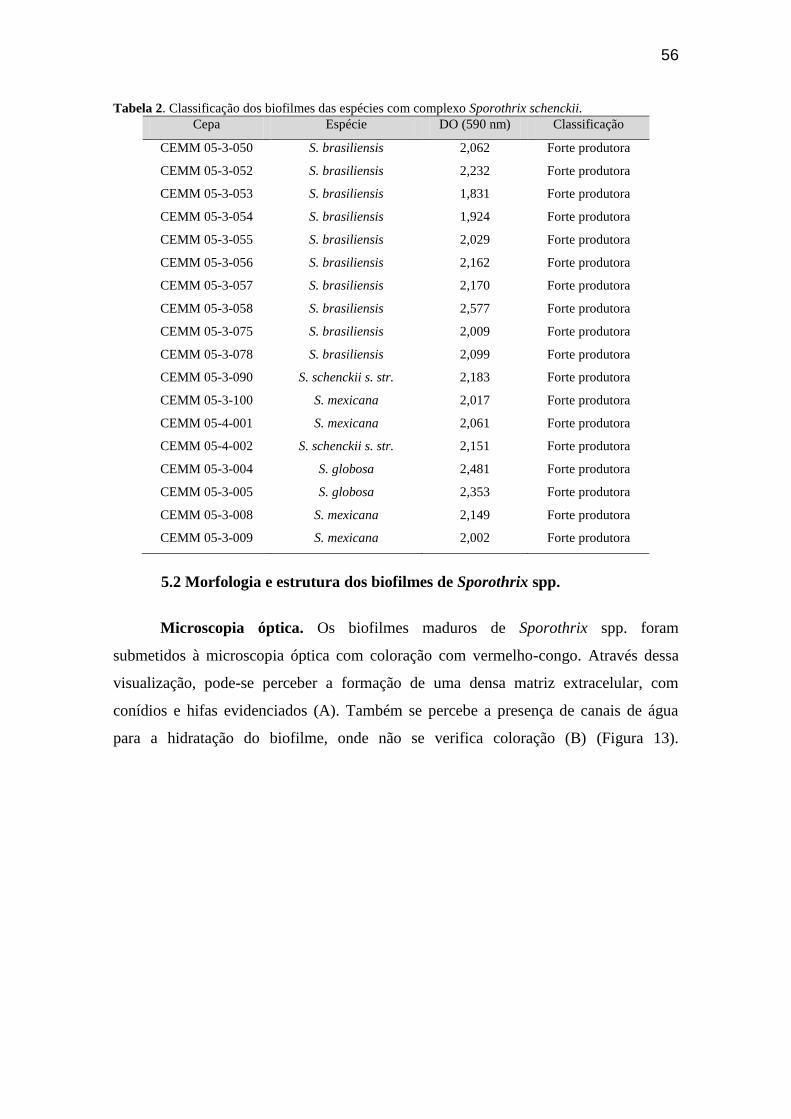
56
Tabela 2. Classificação dos biofilmes das espécies com complexo Sporothrix schenckii.
Cepa Espécie DO (590 nm) Classificação
CEMM 05-3-050 S. brasiliensis 2,062 Forte produtora
CEMM 05-3-052 S. brasiliensis 2,232 Forte produtora
CEMM 05-3-053 S. brasiliensis 1,831 Forte produtora
CEMM 05-3-054 S. brasiliensis 1,924 Forte produtora
CEMM 05-3-055 S. brasiliensis 2,029 Forte produtora
CEMM 05-3-056 S. brasiliensis 2,162 Forte produtora
CEMM 05-3-057 S. brasiliensis 2,170 Forte produtora
CEMM 05-3-058 S. brasiliensis 2,577 Forte produtora
CEMM 05-3-075 S. brasiliensis 2,009 Forte produtora
CEMM 05-3-078 S. brasiliensis 2,099 Forte produtora
CEMM 05-3-090 S. schenckii s. str. 2,183 Forte produtora
CEMM 05-3-100 S. mexicana 2,017 Forte produtora
CEMM 05-4-001 S. mexicana 2,061 Forte produtora
CEMM 05-4-002 S. schenckii s. str. 2,151 Forte produtora
CEMM 05-3-004 S. globosa 2,481 Forte produtora
CEMM 05-3-005 S. globosa 2,353 Forte produtora
CEMM 05-3-008 S. mexicana 2,149 Forte produtora
CEMM 05-3-009 S. mexicana 2,002 Forte produtora
5.2 Morfologia e estrutura dos biofilmes de Sporothrix spp.
Microscopia óptica. Os biofilmes maduros de Sporothrix spp. foram
submetidos à microscopia óptica com coloração com vermelho-congo. Através dessa
visualização, pode-se perceber a formação de uma densa matriz extracelular, com
conídios e hifas evidenciados (A). Também se percebe a presença de canais de água
para a hidratação do biofilme, onde não se verifica coloração (B) (Figura 13).

57
Figura 13. Microscopia óptica de biofilmes maduros de Sporothrix globosa, corados através da técnica com corante Vermelho-Congo. A. Matriz extracelular densa (fundo
vermelho), com hifas (setas pretas) e conídios (setas brancas); B. Canais de água dentro da rede de hifas (setas). Ampliação de 400x.

58
Microscopia confocal. Através da microscopia confocal, os biofilmes do
complexo S. schenckii puderam ser visualizados em perspectiva de terceira dimensão
(3D), demonstrando a diferença entre células vivas e mortas, identificadas através das
cores verdes e vermelhas, respectivamente. Esses biofilmes maduros possuem um maior
número de células vivas em detrimento às células mortas, demonstrando que a atividade
metabólica dos biofilmes permanece alta após a maturação, como demonstrado na
figura 14.

59
Figura 14. Imagens de CLSM de biofilmes maduros de Sporothrix brasiliensis. A. Hifas (setas pretas) e conídios (setas brancas) formando uma estrutura densa de biofilme.
B. Visão tridimensional do biofilme. Ampliação de 400X. Imagens adquiridas com laser 488 nm e 561nm para a detecção de SYTO9 e iodeto de propídio, que identificam
células vivas e mortas, respectivamente.

60
Microscopia eletrônica de Varredura (MEV). Os biofilmes maduros de
Sporothrix spp. mostraram, nas condições avaliadas, em microscopia eletrônica de
varredura, que são formados por uma massa densa e heterogênea de conídios e hifas.
Após 24 horas de incubação, as células aderidas à superfície da lamínula de
Thermanox™ são evidentes e, após 48 horas de incubação, percebe-se o início do
processo de filamentação. A rede de conídios e hifas começa a ser visualizada após 72
horas, prosseguindo o processo de amadurecimento após 96 e 120 horas, quando já se
percebe a formação de canais de água para hidratação do biofilme. Após 120 horas de
incubação, o biofilme maduro pode ser visualizado, percebendo-se a presença de uma
matriz extracelular e canais de água bem definidos, como demonstrado na figura 15.

61
Figura 15. Imagens de microscopia eletrônica de varredura de biofilmes de S. brasiliensis formados em lamínulas de Thermanox™ após 24, 48, 72, 96, 120 e 144 horas de
incubação (A - F, respectivamente) - Ampliação: 2000X (A - E) e 4032X (F). A. Células aderidas à superfície da lâmina. B. Início da filamentação pós-adesão. C. Formação
de redes de hifas e conídios. D. Espessamento do biofilme com detecção de produção de matriz. E. Biofilme maduro com presença de conídios, hifas, matriz e canais de água
(identificados por setas). F. Canais de água no biofilme de S. brasiliensis (identificados por setas).

62
5.3 Sensibilidade planctônica
Os resultados dos ensaios in vitro de sensibilidade dos antifúngicos contra as
cepas estudadas de quatro espécies de complexo S. schenckii estão mostrados na Tabela
3. Verificou-se que, para cepas de S. brasiliensis, os resultados de MIC variaram entre
0, 25 µg/mL < MIC < 8,0 µg/mL para AMB, 8,0 a >16 µg/mL para CAS, 0,25 µg/mL <
MIC < 8,0 µg/mL para KTC e >16 e >64 µg/mL para VRC e FLC, respectivamente.
Para cepas de S. schenckii s. str., os MICs foram 0,25 µg/mL para AMB, >16 µg/mL
para CAS e VRC, 8,0 µg/mL para KTC e >64 µg/mL para FLC. Sporothrix globosa
obteve valores de MICs de 0,5 µg/mL para AMB, >16 µg/mL para CAS e VRC, >64
µg/mL para FLC e entre 2,0 µg/mL < MIC < 4,0 µg/mL para KTC. Já S. mexicana
demonstrou MICs variando entre 1,0 µg/mL < MIC < 8,0 µg/mL para AMB, 8,0 a >16
µg/mL para CAS, 2,0 µg/mL < MIC < 16,0 µg/mL para KTC, ≥ 16 µg/mL para VRC e
>64 µg/mL para FLC.

63
Tabela 3. Concentração inibitória mínima (MIC) e concentração fungicida mínima (MFC) de isolados planctônicos do complexo Sporothrix schenckii.
Espécies
AMB (µg/mL)
CAS (µg/mL)
VRC (µg/mL)
FLC (µg/mL)
KTC (µg/mL)
MIC
CEM
MIC
MIC
MIC
Média MG MFC Média MG MFC Média MG MFC Média MG MFC Média MG MFC
S. brasiliensis (n=10)
50%
8 - >16 >16
2 – 16 6,498
64 - >64 >64
0,0625 – 1 0,233
80%
>16 >16
4 - >16 11,314
>64 >64
0,25 – 1 0,5
100%
0,25 – 1 0,660 0,25 – 1
>16 >16 >16
8 - >16 12,126 >16
>64 >64 >64
1 - 4 1,625 2 – 8
S. schenckii s. str. (n=2)
50%
16 16
4 – 8 5,657
32 – 64 45,255
0,5 – 1 0,707
80%
>16 >16
16 - >16 >16
64 - >64 64
1 1
100%
0,25 0,25 0,5
>16 >16 >16
>16 16 >16
>64 >64 >64
1 - 2 1,414 2.
S. globosa (n=2)
50%
16 - >16 >16
8 8
16 – 32 22,627
0,5 0,5
80%
>16 >16
16 16
32 - 64 45,255
1 – 2 1,414
100%
0,5 0,5 0,5
>16 >16 >16
>16 16 >16
>64 >64 >64
2 - 4 2,828 2 - 4
S. mexicana (n=4)
50%
>16 >16
0,5 - >16 4,757
32 - >64 53,817
0,5 – 2 1
80%
>16 >16
4 - >16 9,514
64 - >64 64
1 - 4 1,682
100% 1 – 8 2,378 8 >16 >16 >16 16 - >16 >16 16,0 - >16 64 - >64 >64 >64 2 - 16 4,757 4 - 16
AMB, anfotericina B; CAS, caspofungina; VRC, voriconazol; FLC, fluconazol; KTC, cetoconazol. 50%, 80% e 100%: porcentagem de inibição visual de crescimento por poço; MG: média geométrica; MFC: Concentração fungicida mínima; MIC: Concentração inibitória mínima; CEM: Concentração mínima eficaz

64
5.4 Atividade metabólica e biomassa dos biofilmes frente aos antifúngicos
Neste estudo, a atividade metabólica do biofilme de S. brasiliensis teve uma
redução considerada significativa (p<0.05) quando comparada ao crescimento controle
para todas as concentrações de AMB e KTC e para 10xMIC e 50xMIC de VCR e CAS.
Já para S. schenckii s. str. e S. globosa, a redução foi significativa (p<0.05) para as
concentrações de 50xMIC de AMB, FLC, KTC e CAS. A última espécie analisada, S.
mexicana, mostrou redução significativa (p<0.05) para as concentrações de 50xMIC de
todas as drogas analisadas, além de 10xMIC para KTC e CAS. A atividade metabólica
in vitro dos biofilmes do complexo S. schenckii contra AMB, CAS VRC, FLC e KTC
estão demonstradas da tabela 4.
Quando relacionamos a diferença entre a biomassa dos biofilmes que sofreram
ação de antifúngicos e o crescimento controle, percebemos que para S. brasiliensis
houve redução significativa (p<0.05) em todas as concentrações de FLC e CAS,
50xMIC de KTC e 10xMIC e 50xMIC de AMB. Para S. schenckii s. str, a redução
significativa (p<0.05) foi encontrada para 50xMIC de AMB e CAS. Somente as
concentrações de 50xMIC de todas as drogas testadas foram significativas (p<0.05) para
S. globosa. Cepas de S. mexicana apresentaram redução de biomassa em todas as
concentrações de VOR e KTC e para 50xMIC de AMB e CAS (p<0.05). A biomassa in
vitro dos biofilmes do complexo S. schenckii contra AMB, CAS VRC, FLC e KTC
estão demonstradas na tabela 5.

65
Tabela 4.Atividade metabólica dos biofilmes de Sporothrix spp. após exposição às drogas antifúngicas (492 nm) (p<0,05).
Espécies Controle** AMB
VRC***
FLC***
KTC
CAS***
MIC 10xMIC 50xMIC
MIC 10xMIC 50xMIC
MIC 10xMIC 50xMIC
MIC 10xMIC 50xMIC
MEC 10xMEC 50xMEC
S.brasiliensis
(n=10) 0,9180
0,8696* 0,8221* 0,7540*
0,9351 0,8853* 0,8509*
0,9211 0,9091 0,8895
0,8825* 0,8353* 0,7880*
0,9153 0,8915* 0,8628*
S. schenckii s.
str. (n=2) 0,9338
0,9150 0,8873 0,8558*
0,9260 0,9155 0,9020
0,9170 0,8848 0,8643*
0,9078 0,8855 0,8678*
0,9265 0,9108 0,8970*
S. globosa
(n=2) 0,9303
0,9060 0,8235 0,7893*
0,9883 0,9525 0,9003
0,9110 0,8893 0,8443*
0,8900 0,8550 0,8240*
0,9195 0,8935 0,8590*
S. mexicana
(n=4) 0,8149
0,8093 0,7813 0,7271*
0,8130 0,7908 0,7643*
0,8131 0,7974 0,7601*
0,7921 0,7830* 0,7450*
0,8065 0,7904* 0,7713*
* Redução significativa em relação ao controle
** Crescimento do biofilme sem antifúngico *** Para valores de MICs ou MECs planctônicos acima das concentrações testadas, utilizou-se o valor da maior concentração testada.
Tabela 5. Biomassa dos biofilmes de Sporothrix spp. após exposição às drogas antifúngicas (590 nm) (p<0,05).
Espécies Controle** AMB
VRC***
FLC***
KTC
CAS***
MIC 10xMIC 50xMIC
MIC 10xMIC 50xMIC
MIC 10xMIC 50xMIC
MIC 10xMIC 50xMIC
MEC 10xMEC 50xMEC
S.brasiliensis
(n=10) 2,180 2,163 2,111* 2,059* 2,191 2,189 2,183 2,163* 2,151* 2,131* 2,173 2,170 2,105* 2,164* 2,140* 2,120*
S. schenckii s. str.
(n=2) 2,252 2,222 2,190 2,159* 2,253 2,225 2,193 2,268 2,261 2,230 2,246 2,194 2,156 2,247 2,228 2,212*
S. globosa (n=2) 2,498 2,500 2,457 2,418* 2,607 2,566 2,528* 2,511 2,436 2,402* 2,444 2,384 2,348* 2,479 2,469 2,431*
S. mexicana
(n=4) 2,133 2,126 2,113 2,086* 2,108* 2,083* 2,064* 2,133 2,125 2,115 2,107* 2,079* 2,053* 2,134 2,121 2,106*
* Redução significativa em relação ao controle ** Crescimento do biofilme sem antifúngico
*** Para valores de MICs ou MECs planctônicos acima das concentrações testadas, utilizou-se o valor da maior concentração testada.

66
6 DISCUSSÃO
Quatro espécies do complexo Sporothrix schenckii são relacionadas aos casos de
esporotricose humana e animal relatados em todo o mundo. Dentre as espécies deste
complexo, S. brasiliensis (associada, principalmente, às infecções animais e transmissão
zoonótica), S. schenckii sensu stricto, S. globosa (associada à transmissão ambiental) e
S. mexicana são as espécies mais encontradas nestes pacientes (GREMIÃO et al., 2015).
Os biofilmes são comunidades microbianas que podem aderir-se às superfícies
bióticas ou abióticas, envolvidas por uma matriz polimérica extracelular, com um
fenótipo divergente das células planctônicas em vários fatores genéticos e morfológicos,
incluindo o crescimento e os processos de metabolismo, liberação de endotoxinas,
muramil peptídeos, polissacarídeos e DNA, produção de enzimas, transcrição de genes e
resistência a medicamentos antimicrobianos e mediadores específicos da resposta imune
dos hospedeiros (CAPPELLI et al., 2007; PIERCE et al., 2008), fatores importantes nos
processos infecciosos. Estes biofilmes são destinados a proteger as comunidades
microbianas de fatores ambientais adversos que podem comprometer a viabilidade das
células, como, por exemplo, pH, desidratação, substâncias químicas, entre outros (DI
BONAVENTURA et al., 2006; TOURNU; VAN DIJCK, 2012; MITCHELL, 2016).
Conhecer os mecanismos envolvidos na formação, maturação e reprodução desses
biofilmes é importante para a aquisição de formas de controle dos mesmos.
Neste estudo, exploramos a capacidade de espécies do complexo S. schenckii de
formarem biofilmes em placas de poliestireno, avaliando parâmetros como a produção
de biomassa através dos ensaios com cristal violeta e a atividade metabólica através de
ensaios com XTT. Além disso, os biofilmes foram visualizados por microscopia óptica,

67
confocal e eletrônica de varredura, com informações sobre suas formações estruturais e
ultra-estruturais.
Demonstramos que as espécies do complexo Sporothrix schenckii possuem a
capacidade de formar biofilmes em sua fase filamentosa, encontrada na natureza, assim
como outros autores já relataram que várias espécies fúngicas, sejam elas
leveduriformes, como o gênero Candida spp. (AL-FATANI; DOUGLAS, 2006),
filamentosos, como o gênero Aspergillus spp. (BEAUVAIS; LATGÉ, 2015) e Fusarium
spp. (SENGUPTA et al., 2012) ou dimórficos, como os gêneros Coccidioides spp.
(DAVIS et al., 2002), Histoplasma spp. (BRILHANTE et al., 2015) e Paracoccidioides
spp. (SARDI et al., 2015) também possuem, além da capacidade de resistência desses
biofilmes aos antifúngicos.
Esses biofilmes tiveram a melhor capacidade de crescimento em placas de
poliestireno com inóculo de 2 x 105 conídios/mL e tempo de adesão de 24 horas, em
detrimento das outras concentrações e tempos de adesão, pois, quando utilizou-se a
concentração de 2 x 104 conídios/mL, não houve boa capacidade de adesão mesmo após
24 horas, restando poucas células e o biofilme não formava sua rede característica
mesmo após vários dias de incubação. Ao utilizar-se a concentração de 2 x 106
conídios/mL, percebeu-se uma equivalência de crescimento com a concentração testada
imediatamente menor, tanto relacionada à adesão celular medida através de
espectrofotômetro quanto pela visualização da rede de hifas e conídios através de
microscopia eletrônica de varredura. Os ensaios de tempo de adesão com todas as
concentrações de inóculos testadas mostraram que, para a melhor formação de biofilmes
de Sporothrix spp., há a necessidade de contato entre o inóculo e a superfície de

68
poliestireno por, pelo menos, 24 horas, não havendo boa formação de biofilme em
tempos de adesão menores (dados não mostrados).
Assim como outros biofilmes microbianos, os biofilmes formados in vitro por
espécies do complexo S. schenckii seguem padrões de formação que incluem a adesão à
superfície após 24 horas de incubação, a multiplicação celular e a maturação do
biofilme. Com o passar das horas, há um lento crescimento do metabolismo do
biofilme, assim como a biomassa produzida, quando comparado a outros biofilmes
microbianos até que, após 120 horas, é atingido seu ápice de crescimento e a
manutenção das taxas descritas, como foi demonstrado no presente estudo, semelhante
ao encontrado por Sardi et al. (2015) e Brilhante et al. (2015) para P. brasiliensis e H.
capsulatum, respectivamente.
A atividade metabólica determinada por ensaio com XTT é uma metodologia
com alta precisão, pois possui como mecanismo a redução deste sal de tetrazólio em seu
produto colorido, chamado formazan. Esta reação somente acontece na presença de
desidrogenases mitocondriais, ou seja, somente células vivas têm a capacidade de
produzir essa reação. Este ensaio é importante tanto para avaliar a cinética de
crescimento do biofilme quanto à atividade exercida por drogas no metabolismo deste
(PITANGUI et al., 2012; CORDEIRO et al., 2015c).
Através da microscopia eletrônica de varredura pudemos perceber que a
formação dos biofilmes de Sporothrix spp. se dá de forma altamente organizada, assim
como demonstrado com biofilmes de H. capsulatum (PITANGUI et al., 2012). Porém,
diferentemente deste outro fungo dimórfico, o processo de adesão das células de
Sporothrix spp. não é rápido. Enquanto as células de H. capsulatum possuem um tempo
de adesão na superfície de poliestireno de 7 horas, Sporothrix spp. necessita de 24 horas

69
para uma adesão significativa. Durante o período de maturação (24 a 72 horas para H.
capsulatum e 120 horas para Sporothrix spp.), o incremento de matriz extracelular e a
estruturação do biofilme são similares entre os dois fungos, havendo uma aderência
persistente à superfície abiótica.
Em estudo de cinética de crescimento e morfologia de biofilme de leveduras de
P. brasiliensis (Pb18 – cepa utilizada em muitos estudos de virulência), Sardi et al.
(2015) obtiveram resultados bem próximos aos encontrados em nosso estudo. Apesar do
tempo de adesão ser idêntico ao encontrado para H. capsulatum (7 horas), o período de
crescimento e maturação segue ao encontrado para Sporothrix spp., tendo uma
formação consistente próximo a 144 horas de maturação. Entretanto, para que o
biofilme de P. brasiliensis fosse formado, os autores incubaram a 37 °C com 5% de
presença de CO2.
Os biofilmes de Sporothrix spp. após a maturação, apresentam-se como uma
rede heterogênea, ou um aglomerado, de hifas e conídios imersos em matriz,
intercalados com uma grande quantidade de canais de água, por onde o líquido presente
flui. Possui estrutura com grande quantidade de filamentos, organizada, com múltiplas
camadas, que formam uma arquitetura de barreira, protegendo o fungo contra ataques
externos. Nossos achados corroboram Sánchez-Herrera et al. (2014), que demonstraram
essa capacidade estrutural nos biofilmes de S. schenckii, formados por carboidratos
totais que vão sendo incorporados ao biofilme no decorrer do seu desenvolvimento.
Para se investigar a atividade dos antifúngicos contra os biofilmes de Sporothrix
spp., foi necessária, inicialmente, a determinação do MIC planctônico da fase
filamentosa. Diversos estudos em todos os continentes demonstram divergências no

70
perfil de sensibilidade destas espécies em sua forma planctônica a muitos antifúngicos
utilizados na prática clínica, sendo verificados, em alguns casos, altos valores de MIC.
Nossos achados relacionados à atividade in vitro dos antifúngicos contra a forma
planctônica de Sporothrix spp. estão de acordo com muitos trabalhos previamente
realizados, como os resultados obtidos por Brilhante et al. (2016) com cepas de S.
brasiliensis provenientes de gatos com esporotricose. Os MICs são variáveis de acordo
com o antifúngico e com a espécie estudada. Verificou-se, por exemplo, que AMB,
CAS e KTC apresentam valores mais altos de MIC ou MEC planctônicos para S.
mexicana, quando comparados às outras espécies estudadas. Esse achado corrobora
outros autores que têm reportado tolerância desta espécie a antifúngicos (BRILHANTE
at al., 2014; RODRIGUES et al., 2014). Os valores de MEC encontrados para a
caspofungina contra células planctônicas (8 - >16 µg/mL) em nosso estudo corroboram
outros estudos que demonstram a sensibilidade de cepas de S. brasiliensis e S. schenckii
s. str. a este antifúngico (OLIVEIRA et al., 2011).
Verificamos que AMB, CAS e KTC apresentaram valores mais altos de MIC
planctônico para S. mexicana quando comparados às outras espécies estudadas,
corroborando com outros autores que têm reportado determinada tolerância desta
espécie a muitas drogas atualmente utilizadas (BRILHANTE et al., 2014; RODRIGUES
et al., 2014). Os valores de MIC encontrados para a caspofungina contra células
planctônicas (8 - >16 µg/mL) corroboram com os achados por Oliveira et al. (2011),
que demonstraram a sensibilidade de cepas de S. brasiliensis e S. schenckii str. sensu
contra este antifúngico.
S. globosa apresentou MICs planctônicos para AMB próximos aos encontrados
em S. brasiliensis, porém, quanto aos MICs para FLC e KTC, foram menores que as

71
outras espécies estudadas. Rodrigues et al. (2014) verificaram que S. globosa
apresentou MICs para AMB também próximos aos de S. brasiliensis, porém, algumas
cepas apresentaram MICs maiores. Já Oliveira et al. (2015) encontraram MICs de KTC
para S. globosa menores que para S. schenckii str. sensu. Nossos resultados também
corroboram com os encontrados por Khao et al. (2015), com MICs de FLC menores
para S. globosa que para outras espécies estudadas.
Em outro estudo de avaliação de sensibilidade entre fármacos azólicos e espécies
do complexo S. schenckii com suas formas planctônicas, os MICs para o cetoconazol
contra cepas de S. brasiliensis, S. schenckii s. str., S. globosa, S. mexicana foram 0,25 –
0,50 µg/mL, 0,06 – 2,0 µg/mL, 2,0 µg/mL, 4,0 µg/mL, respectivamente. Já para o
voriconazol, com as mesmas cepas, os valores foram 1,0 – 8,0 µg/mL, 1,0 – 16,0
µg/mL, 16,0 µg/mL, 16,0 µg/mL, respectivamente (OLIVEIRA, et al., 2015). Em
nossas análises, todos os MICs para KTC e VRC foram maiores que os encontrados por
esses pesquisadores, exceto o MIC de KTC para S. brasiliensis, que se mostrou o
mesmo nos dois estudos. Isso demonstra a variedade de sensibilidade entre as cepas
encontradas de todas as espécies do complexo Sporothrix schenckii.
Fatores como a diferença entre MICs encontrados em vários estudos e valores
altos para esses dados, em células planctônicas, despertam uma necessidade científica
de promover estudos de sensibilidade que considerem as formas sésseis destes
microrganismos, pois, apesar de evidências da presença de biofilmes microbianos na
natureza e em hospedeiros infectados serem uma realidade, o conhecimento nesta seara
em relação às espécies do gênero Sporothrix é limitado e pouco discutido (DI
BONAVENTURA et al., 2006; ITURRIETA-GONZÁLEZ et al., 2014; SÁNCHEZ-
HERRERA et al., 2014; BRILHANTE et al., 2015a).

72
Os resultados encontrados para a atividade metabólica e a biomassa in vitro dos
biofilmes maduros após a exposição aos antifúngicos analisados mostraram que há
diferenças entre as espécies do complexo S. schenckii para cada droga e para cada
concentração utilizada, reproduzindo o que acontece com os testes de sensibilidade para
células planctônicas. Podemos perceber que a atividade metabólica e a biomassa de
todas as espécies tiveram uma redução significativa quando utilizamos a concentração
de 50xMIC de AMB e CAS. Já VRC mostrou redução nas atividades metabólicas dos
biofilmes de S. brasiliensis (10xMIC e 50xMIC) e S. mexicana (50xMIC) e redução na
biomassa de S. globosa (50xMIC) e S. mexicana (MIC, 10xMIC e 50xMIC).
Já FLC, mostrou redução significativa no metabolismo dos biofilmes de S.
schenckii s. str., S. globosa e S. mexicana (50xMIC). Porém, em relação à redução na
biomassa, FLC foi efetivo contra S. globosa (MICx50) e S. brasiliensis em todas as
concentrações estudadas. Cetoconazol reduziu significativamente o metabolismo dos
biofilmes de S. brasiliensis em todas as concentrações, de S. schenckii s. str. e S.
globosa (MICx50) e S. mexicana (MICx10 e MICx50). Não houve redução significativa
de biomassa nos biofilmes de S. schenckii s. str.
Observou-se atividade anti-biofilme quando utilizadas os MICs planctônicos em
algumas drogas, porém, essa atividade foi mais pronunciada nas concentrações 50 x
MIC de AMB e 50 x MEC de CAS. Vale destacar que achado semelhante foi observado
pesquisa com drogas antifúngicas e farnesol contra os biofilmes do fungo dimórfico H.
capsulatum var. capsulatum, onde as taxas de inibição do biofilme contra essas drogas
foram achadas preponderantemente em concentrações acima dos MICs planctônicos
entre 50 e 100 vezes (BRILHANTE et al., 2015). Esses achados em biofilmes de fungos
dimórficos, incluindo de Sporothrix spp. corroboram as pesquisas com biofilme de

73
Candida albicans, que demonstraram que estas estruturas estão implicadas na menor
responsividade a essas drogas, que também não inibiram a dispersão celular proveniente
do biofilme (SELEEM et al., 2016).
Chatzimoschou et al. (2011) demontraram em seu estudo a dificuldade para se
combater as espécies microbianas quando as mesmas estão sob a forma de biofilmes,
nesse caso, as do gênero Candida spp., as leveduras mais encontradas em infecções
fúngicas. Foram testados triazóis e equinocandinas de forma simultânea, avaliando o
sinergismo contra os biofilmes. Porém, os resultados se mostraram desanimadores,
sendo indiferentes contra as células sésseis.
Davis et al. (2002), ao estudarem falha terapêutica em paciente com meningite
por Coccidioides immitis, corroboraram estes estudos que demonstram a dificuldade de
combater biofilmes microbianos. Foi verificado que este fungo é capaz de produzir
biofilmes em dispositivo médico ventrículo-peritoneal e que estes biofilmes foram
responsáveis pela resistência ao fluconazol utilizado no tratamento, além de permitir a
persistência do fungo causando a infecção por quatro anos.
As diferentes estratégias utilizadas pelos fungos quando em biofilmes para
burlarem a ação das drogas antimicrobianas podem incluir a ação de bombas de efluxo,
a baixa expressão de alvos moleculares utilizados pelos fármacos para exercerem sua
ação, mudanças na fisiologia geral das células, alta densidade celular e a presença de
matriz extracelular (RAMAGE et al., 2012). Deste modo, através dos ensaios para
avaliar a produção de biomassa, pudemos verificar no presente estudo que o gênero
Sporothrix possui a capacidade de formar fortes biofilmes e têm alta densidade celular,
o que pode influenciar nos resultados de baixa resposividade destes biofilmes contra as
concentrações utilizadas, mesmo que altas, dos antifúngicos testados.

74
Além disso, os biofilmes formados pelo gênero Sporothrix possuem altas taxas
de atividade metabólica, avaliados por ensaios com XTT, mesmo após a maturação.
Essa atividade metabólica demonstra que a produção de enzimas e outros componentes
químicos também continua com taxas elevadas, o que pode favorecer os mecanismos de
desativação molecular dos fármacos antimicrobianos por proteases, hidrolases, dentre
outras.
Estas diferenças reforçam a importância da identificação das espécies do
complexo S. schenckii e seus perfis de sensibilidade para que seja feita uma avaliação
dos tratamentos propostos para a esporotricose na prática clínica, já que muitos estudos
já demonstram a capacidade de outros fungos produzirem biofilmes no curso de
infecções em modelos in vivo, como Aspergillus spp. e Candida albicans (SELEEM et
al., 2016; WILLIAMS et al., 2016).
Em resumo, os resultados obtidos por este trabalho mostraram que as quatro
espécies do complexo Sporothrix schenckii causadoras de esporotricose, S. brasiliensis,
S. schenckii s. str., S. globosa e S. mexicana possuem a capacidade de produzirem, in
vitro, robustos biofilmes, tendo sua maturação entre 24 e 120 horas. A caracterização
estrutural destes biofilmes mostra também a similaridade que eles possuem em relação a
outros biofilmes fúngicos, principalmente em relação a outros fungos dimórficos. Além
disso, demonstra que estes biofilmes possuem uma capacidade de proteção dos
microrganismos contra drogas antifúngicas bastante utilizadas na clínica médica, como
anfotericina B, voriconazol, fluconazol, cetoconazol e caspofungina e isso traz a
necessidade do aprofundamento do estudo dos biofilmes do gênero Sporothrix, com
conhecimento das suas composições, expressão gênica, assim como a responsividade
dos mesmos a diversos outros antimicrobianos.

75
7 REFERÊNCIAS BIBLIOGRÁFICAS
ALBUQUERQUE, P.C.; NAKAYASU, E.S.; RODRIGUES, M.L.; FRASES, S.;
CASADEVALL, A.; ZANCOPE-OLIVEIRA, R.M.; ALMEIDA, I.C.;
NOSANCHUK, J.D. Vesicular transport in Histoplasma capsulatum: an effective
mechanism for trans-cell wall transfer of proteins and lipids in ascomycetes. Cell
Microbiol, v. 10, n.8, p.1695-1710, 2008.
AL-FATANI, M.A.; DOUGLAS, L.J. Biofilm matrix of Candida albicans and
Candida tropicalis: chemical composition and role in drug resistance. J Med
Microbiol. v. 55, n. Pt8, p. 999-1008, 2006.
ALLEWELL, N.M. Introduction to biofilms thematic minireviews series. J Biol
Chem, v. 291, n. 24, p. 12527-12528, 2016.
ALLISON, D.G.; SUTHERLAND, I.W.; A staining techinique for attached bacteria
and its correlation to extracellular carbohydrate production. J Microbiol Methods, v.
2, p. 93-99, 1984.
ALMEIDA-PAES, R.; FRASES, S.; FIALHO MONTEIRO, P.C.; GUTIERREZ-
GALHARDO, M.C.; ZANCOPÉ-OLIVEIRA, R.M.; NOSANCHUK, J.D. Growth
conditions influence melanization of brazilian clinical Sporothrix schenckii isolates.
Microbes Infect. v. 11, n. 5, p. 554-562, 2009.
ALMEIDA-PAES, R.; FRASES, S.; ARAÚJO, G.S.; OLIVEIRA, E.M.E.;
GERFEN, G.J.; NOSANCHUK, J.D.; ZANCOPÉ-OLIVEIRA, R.M. Biosyntesis
and functions of a melanoid pigment produced by species of Sporothrix complex in
the presence of L-tyrosine. Appl Env Microbiol, v. 78, n. 24, p. 8623-86-30, 2012.
ALMEIDA-PAES, R.; OLIVEIRA, M.M.; FREITAS, D.F.; VALLE, A.C.;
ZANCOPÉ-OLIVEIRA, R.M.; GUTIERREZ-GALHARDO, M.C. Sporotrichosis in
Rio de Janeiro, Brazil: Sporothrix brasiliensis in associated with a typical clinical
presentations. PLoS Neglected Tropical Diseases, v. 8, n. 9, e3094, 2014.
ALMEIDA-PAES, R.; OLIVEIRA, L.C.; OLIVEIRA, M.M.E.; GUTIERREZ-
GALHARDO, M.C.; NOSANCHUK, J.D.; ZANCOPÉ-OLIVEIRA, R.M.

76
Phenotypic characteristics associated with virulence of clinical isolates from the
Sporothrix complex. Biomed Res Int, v. 2015, 2015.
ALMEIDA-PAES, R.; OLIVEIRA, M.M.; FREITAS, D.F.; VALLE, A.C.;
GUTIERREZ-GALHARDO, M.C.; ZANCOPÉ-OLIVEIRA, R.M. Refractory
Sporotrichosis due to Sporothrix brasiliensis in humans appears to be unrelated to in
vivo resistance. Med Mycol, pii: myw103, 2016.
ALVIANO, C.S.; TRAVASSOS, L.R.; SCHAUER, R. Sialic acids in fungi: a
minireview. Glycoconj J, v. 16, n.9, p.545-554, 1999.
ANTUNES, T.A.; NOBRE, M.O.; FARIA, R.O.; MEINERZ, A.R.; MARTINS,
A.A.; CLEFF, M.B.; FERNANDES, C.G.; MEIRELES, M.C. Experimental
cutaneous Sporotrichosis: in vivo evaluation of itraconazole and terbinafine. Rev Soc
Bras Me Trop, v. 42, n. 6, p. 706-710, 2009.
ARRILLAGA-MONCRIEFF, I.; CAPILLA, J.; MAYAYO, E.; MARIMON, R.;
MARINÉ, M.; GENÉ, J.; CANO, J.; GUARRO, J. Different virulence levels of the
species of Sporothrix in a murine model. Clin Microbiol Infect, v. 15, n. 7, p. 651-
655, 2009.
AUNG, A.K.; THE, B.M.; MCGRATH, C.; THOMPSON, P.J. Pulmonary
sporotrichosis: case series and systematic analysis of literature on clinico-
radiological patterns and management outcomes. Med Mycol, v. 51, n. 5, p. 534-
544, 2013.
BARROS, M.B.; DE ALMEIDA PAES, R; SCHUBACH, A.O. Sporothrix
schenckii and sporotrichosis. Clin Microbiol Rev, v. 24, n. 4, p. 633-654, 2011.
BEAUVAIS, A.; LATGÉ, J.P. Aspergillus biofilm in vitro and in vivo. Microbial
Spectr. v. 3, n. 4, 2015.
BORGES, T.S.; ROSSI, C.N.; FEDULLO, J.D.; TABORDA, C.P.; LARSSON,
C.E. Isolation of Sporothrix schenckii from the claws of domestic cats (indoor and
outdoor) and in captivity in São Paulo (Brazil). Mycopathologia, v. 176, n. 1-2, p.
129-137, 2013.

77
BRILHANTE, R.S.N.; MALAQUIAS, A.D.; CAETANO, E.P.; CASTELO-
BRANCO, DdeS.; LIMA, R.A.; MARQUES, F.J.; SILVA, N.F.; ALENCAR, L.P.;
MONTEIRO, A.J.; CAMARGO, Z.P.; BANDEIRA, T de J.; RODRIGUES, A.M.;
CORDEIRO, RdeA.; MOREIRA, J.L.; SIDRIM, J.J.; ROCHA, M.F. In vitro
inhibitory effect of miltefosine against strains of Histoplasma capsulatum var.
capsulatum and Sporothrix spp. Med Mycol, v. 52, n. 3, p. 320-325, 2014.
BRILHANTE, R.N.S.; LIMA, R.A.C.; MARQUES, F.J.F.; SILVA, N.F.;
CAETANO, E.P.; CASTELO-BRANCO, D.S.C.; BANDEIRA, T.J.P.G.;
MOREIRA, J.L.B.; CORDEIRO, R.A.; MONTEIRO, A.J.; CAMARGO, Z.P.;
SIDRIM, J.J.C.; ROCHA, M.F.G. Histoplasma capsulatum in planktonic and
biofilm forms: in vitro susceptibility to amphotericin B, itraconazole and farnesol. J
Med Microbiol. v. 64, n. Pt4, p. 394-399, 2015.
BRILHANTE, R.S.N.; RODRIGUES, A.M.; SIDRIM, J.J.C.; ROCHA, M.F.G.;
PEREIRA, S.A.; GREMIÃO, I.D.F.; SCHUBACH, T.M.P.; DE CAMARGO, Z.P.
In vitro susceptibility of antifungal drugs against Sporothrix brasiliensis recovered
from cats with sporotrichosis in Brasil. Med Mycol, v. 54, n. 3, p. 275-279, 2016.
CAMACHO, E.; LÉON-NAVARRO, I.; RODRÍGUEZ-BRITO, S.; MENDOZA,
M.; NIÑO-VEGA, G.A. Molecular epidemiology of human sporotrichosis in
Venezuela revels high frequency of Sporothrix globosa. BMC Infec Dis, v.15, n.94,
2015.
CAPPELLI, G.; RICARDI, M.; RAVERA, F.; LIGABUE, G.; BALLESTRI, M.;
BONUCCHI, D.; BONDI, M. Biofilm on artificial surfaces. Contrib Nephrol, v.
154, p. 61-71, 2007.
CARMICHAEL, J.W. Chrysosporium and some other aleuriosporic hyphomycetes.
Canad J Bot, v.40, p. 1137-1181, 1962.
CARRILLO-MUÑOZ, A.J.; GIUSIANO, G.; EZKURRA, P.A.; QINDÓS, G.
Antifungal agents: Mode of action in yeasts cells. Rev Esp Quimioterap, v.19, n.2,
p.130-139, 2006.

78
CASADEVALL, A. Amoeba provide insight into the origin of virulence in
pathogenic fungi. Adv Exp Med Biol. v. 710, p. 1-10, 2012.
CHAKRABARTI, A.; BONIFAZ, A.; GUTIERREZ-GALHARDO, M.C.;
MOCHIZUKI, T.; LI, S. Global epidemiology of sporotrichosis. Med Mycol, v.53,
p.3-14, 2015.
CHANDRASEKAR, P. Management of invasive fungal infections: a role for
polyenes. J Antimicrob Chemother, v. 66, p. 457-465, 2011.
CHATZIMOSCHOU, A.; KATRAGKOU, S.; SIMITSOPOULOU, M.;
ANTACHOPOULOS, C.; GEORGIADOU, E.; WALSH, T.J.; ROILIDES, E.
Activies of triazole-echinocandin combinations against Candida species in biofilms
and as planktonic cells. Antimicrobial Agents and Chemotherapy, v. 55, n. 5, p.
1968-1974, 2011.
CLSI, Reference Method for Broth Dilution Antifungal Susceptibility Testing of
Filamentous Fungi. 2nd
edition, Clinical and Laboratory Standards Institute, 2008.
CORDEIRO, R.A.; OLIVEIRA, J.S.; CASTELO-BRANCO, D.S.C.M.;
TEIXEIRA, C.E.C.; MARQUES, F.J.F.; BITTENCOURT, P.V.; CARVALHO,
V.L.; BANDEIRA, T.J.P.G.; BRILHANTE, R.S.N.; MOREIRA, J.L.B.; PEREIRA-
NETO, W.A.; SIDRIM, J.J.C.; ROCHA, M.F.G.; Candida tropicalis isolates
obtained from veterinary sources show resistance to azoles and produce virulence
factors. Med Mycol, v. 53, n. 2, p. 145-152, 2015a.
CORDEIRO, R.A.; SERPA, R.; ALEXANDRE, C.F.U.; MARQUES, F.J.F.;
MELO, C.V.S.; FRANCO, J.S.; EVANGELISTA, A.J.J.; CAMARGO, Z.P.;
BRILHANTE, R.S.N.; ROCHA, M.F.G.; MOREIRA, .J.L.B.; BANDEIRA,
T.J.P.G.; SIDRIM, J.J.C. Trichosporon inkin biofilms produce extracellular

79
proteases and exhibit resistance to antifungals. J Med Microbiol, v. 64, n. 11, p.
1277-1286, 2015b.
CORDEIRO, R.A.; SERPA, R.; MARQUES, F.J.; DE MELO, C.V.;
EVANGELISTA, A.J.; MOTA, V.F.; BRILHANTE, R.S.; BANDEIRA, TdeJ.;
ROCHA, M.F.; SIDRIM, J.J. Inhibitory activity of isoniazide and ethionamide
against Cryptococcus biofilms. Can J Microbiol, v. 61, n. 11, p. 827-836, 2015c.
COSTA, R.O.; AZULAY-ABULAFIA, L.; NEVES, M.L.P.; BERNARDES-
ENGEMANN, A.R.; BENVENUTO, F.; LOPES-BEZERRA, L.M. Sporotrichosis
in pregnancy: case reports of 5 patients in a zoonotic epidemic in Rio de Janeiro,
Brazil. An Bras Dermatol, v.86, n.5, p.995-998, 2011.
CRISEO, G.; MALARA, G.; ROMEO, O.; PUGLISI GUERRA, A.
Lynphocutaneous sporotrichosis in na immunocompetent patient: a case report from
extreme southern Italy. Mycopathologia, v. 166, n.3, p.159-162, 2008.
DA ROSA, D.; GEZUELE, E.; CALEGARI, L.; GOÑI, F. Excretion-secretion
products and proteases from live Sporothrix schenckii yeast phase: immunological
detection and cleavage of human IgG. Rev Inst Med Trop São Paulo, v. 51, n. 1, p.
1-7, 2009.
DAVIS, L.E.; COOK, G.; COSTERTON, J.W. Biofilm in ventriculo-peritoneal
shunt tubing as a cause of treatment failure in coccidioidal meningitis. Emerg Infect
Dis. v. 8, n. 4, p. 376-379, 2002.
DE BEER, P.; STOODLEY, P.; LEWANDOWSKY, Z. Liquid flow in
heterogeneous biofilms. Biotech and Bioeng. V. 44, n. 5, p. 636-641, 1994.
DE MEYER, E.M.; SUMMERBELL, R.C.; MOHARRAM, A.M.; DE HOOG,
G.S.; VISMER, H.F.; WINGFIELD, M.J. Taxonomy and phylogeny of new wood-
and soil-inhabiting Sporothrix species in the Ophiostoma stenoceras - Sporothrix
schenckii complex. Mycologia. v.100, n. 4, p.647-661, 2008.

80
DESAI, J.V.; MITCHELL, A.P.; ANDES, D.R. Fungal biofilms, drug resistance,
and recurrent infection. Cold Spring Harb Perspect Med, v. 4, n. 10, 2014.
DHINGRA, D.; DURREHEIN, D.; PORIGNEAUX, P. Sporotrichosis outbreak and
moudly hay in NSW. Aust Farm Physician, v.44, n.4, p. 217-221, 2015.
DI BONAVENTURA, G.; POMPILIO, A.; PICCIANI, C.; IEZZI, M.;
D’ANTONIO, D.; PICCOLOMINI, R. Biofilm formation by the emerging fungal
pathogen Trichosporon asahii: development, architecture and antifungal resistance.
Antimicrob Agents Chemother, v. 50, n. 10, p. 3269-3276, 2006.
DIXON, D.M.; SALKIN, I.F.; DUNCAN, R.A.; HURD, N.J.; HAINES, J.H.;
KEMNA, M.E.; COLES, F.B. Isolation and Characterization of Sporothrix
schenckii from Clinical and Enviromental Sources Associated with Largest U.S.
Epidemic of Sporotrichosis. J of Clin Microbiol, v.29, n.6, p.1106-1113, 1991.
DOOLEY, D.P.; BOSTIC, P.S.; BECKIUS, M.L. Spook house sporotrichosis. A
point-source outbreak of sporotrichosis associated with hay bale props in a
Halloween haunted-house. Arch Int Med, v.157, n.16, p.1885-1887, 1997.
DOWNEY, C.; NAVAJAS, L.; ANDINO, R.; VERA-KELLET, C.; MANRÍQUEZ,
J.J. Sporotrichoid-like tuberculosis: an unusual presentation of cutaneous
tuberculosis in an immunocompetent patient. Rev Chil Infectol, v. 32, n. 2, p. 234-
237, 2015.
DRUTZ, D.J.; FREY, C.L. Intracellular and extracellular defenses of human
phagocytes against Blastomyces dermatitidis conidia and yeasts. J Lab Clin Med,
v.105, n. 6, p.737-750, 1985.
FANNING, S.; MITCHELL, A.P. Fungal biofilms. PLoS Pathogens, v. 8, n. 4,
e1002585, 2012.
FEENEY, K.T.; ARTHUR, I.H.; WHITTLE, A.J.; ALTMAN, S.A.; SPEERS, D.J.
Outbreak of Sporotrichosis, Western Australia. Emerg Infec Dis, v.13, n.8, p.1228-
1231, 2007.

81
FERNANDES, G.F.; DOS SANTOS, P.O.; RODRIGUES, A.M.; SASAKI, A.A.;
BURGUER, E.; DE CAMARGO, Z.P. Characterization of virulence profile, protein
secretion and immunogenicity of different Sporothrix schenckii sensu stricto isolates
compared with S. globosa and S. brasiliensis species. Virulence, v. 4, n. 3, p. 241-
249, 2013.
FERNANDES, K.S.S.; COELHO, A.L.J.; BEZERRA, L.M.L.; BARJA-FIDALGO,
C. Virulence of Sporothrix schenckii conidia and yeast cells, and their susceptibility
to nitric oxide. Immunology, v.101, n.4, p.563-569, 2000.
FERNÁNDEZ-SILVA, F.; CAPILLA, J.; MAYAYO, E.; GUARRO, J. Virulence
of Sporothrix luriei in a Murine Model of Disseminated Infection. Mycopathologia,
v. 173, n. 4, p. 245-249, 2012.
FILGUEIRA, K.D. Esporotricose na espécie canina: relato de um caso na cidade de
Mossoró, RN. Cien Anim Bras, v.10, n.2, p.673-677, 2009.
FINKEL, J.S.; MITCHELL, A.P. Genetic control of Candida albicans biofilm
development. Nat Rev Microbiol, v. 9, n. 2, p. 109-118, 2011.
FREITAS, D.F.S.; VALLE, A.C.F.; SILVA, M.B.T.; CAMPOS, D.P.; LYRA,
M.R.; SOUZA, R.V.; VELOSO, V.G.; ZANCOPÉ-OLIVEIRA, R.M.; BASTOS,
F.I.; GALHARDO, M.C.G. Sporotrichosis: An emerging neglected opportunistic
infection in HIV-infected patients in Rio de Janeiro, Brazil. PLoS Neg Trop Dis, v.8,
n.8, 2014.
GAUTHIER, G.; KLEIN, B.S. Insights into fungal morphogenesis and immune
evasion: fungal conidia, when situated in mammalian lungs, may switch from mold
to pathogenic yeasts or spore-forming spherules. Microbe Wash DC, v. 3, n. 9, p.
416-423, 2008.
GOVENDER, N.P.; MAPHANGA, T.G.; ZULU, T.G.; PATEL, J.; WALAZA, S.;
JACOBS, C.; EBONWU, J.I.; NTULI, S.; NAICKER, S.D.; THOMAS, J. An
Outbreak of Lymphocutaneous Sporotrichosis among Mine-Workes in South Africa.
PLoS Negl Trop Dis, v.9, n.9, 2015.

82
GREMIÃO, I. D.; MENEZES, R. C.; SCHUBACH, T. M.; FIGUEIREDO, A. B.;
CAVALCANTI, M.C.; PEREIRA, S. A. Feline sporotrichosis: epidemiological and
clinical aspects. Med Mycol, v. 53, n. 1, p. 15-21, 2015.
GUNN, J.S.; BAKALETZ, L.O.; WOZNIAK, D.J. What’s on the outside matters:
the role of the extracellular polymeric substance of Gram-negative biofilms in
evading host immunity and as a target for therapeutic intervention. J Biol Chem, v.
291, n. 24, p. 12538-12546, 2016.
HASSAN, I; KENN, A. Potassium iodide in Dermatology. Ind J Dermatol Venereol
Leprol, v. 78, n.3, p.390-393, 2012.
HAY, R.J.; MORRIS-JONES, R. Outbreaks of sporotrichosis. Curr Opin Infect Dis,
v. 21, n. 2, p. 119-121, 2008.
HEKTON, L.; PERKINS, C.F. Refractory subcutaneous abscess caused by
Sporothrix schenckii: a new pathogenic fungus. J Exp Med, v.5, p. 77-91, 1900.
HIRUMA, M.; KAGAWA, S. Ultrastructure of Sporothrix scheckii treated with
iodine-potassium solution. Mycopathologia, v.97, n.2, p.121-127, 1987.
HOGAN, L.H.; KLEIN, B.; LEVITZ, S.M. Virulence factors of medically
important fungi. Clin Microbiol Rev, v.9, n.4, p.469-488, 1996.
HORIO, T.; DANNO, K.; OKAMOTO, H.; MIYACHI, Y.; IMAMURA, S.
Potassium iodide in erythema nodosum and other erythematous dermatoses. J Am
Acad Dermatol, v.9, n.1, p.77-81, 1983.
HOU, B.; ZHANG, Z.; ZHENG, F.; LIU, X. Molecular cloning, characterization
and differential expression of DRK1 in Sporothrix schenckii. Int J Mol Med, v.31,
n.1, p. 99-104, 2013.
ISHIDA, K.; DE CASTRO, R.A.; BORBA DOS SANTOS, L.P.; QUINTELLA,
L.P.; LOPES-BEZERRA, L.M.; ROZENTAL, S. Amphotericin B, alone or
followed by itraconazole therapy, is effective in the control of experimental

83
disseminated sporotrichosis by Sporothrix brasiliensis. Med Mycol, v. 53, n. 1, p.
34-41, 2015.
ITURRIETA-GONZÁLEZ, I.A.; PADOVAN, A.C.B.; BIZERRA, F.C.; HAHN,
R.C.; COLOMBO, A.L. Multiple species oh Trichosporon produce biofilms highly
resistant to triazoles ans amphotericin B. PLoS One, v. 9, n. 10, 2014.
JACOBSEN, E.S. Pathogenic roles for fungal melanins. Clin Microbiol Rev. v. 13,
n. 4, p. 708-717, 2000.
KAUFFMAN, C.A. Sporotrichosis. Clin Infect Dis, v. 30, p. 684-687, 2000.
KAUFFMAN, C.A.; BUSTAMANTE, B.; CHAPMAN, S.W.; PAPPAS, P.G.
Clinical practice guidelines for the management os Sporotrichosis: 2007 Update by
the Infectious Diseases Society of America. Clin Infect Dis, v.45, p.1255-1265,
2007.
KAVANAUGH, J.S.; HORSWILL, A.R. Impact of environmental cues on
Staphilococcal quorum sensing and biofilm development. J Biol Chem, v. 291, n.
24, p. 12556-12564, 2016.
KHAO, M.; ZHOU, X.; LIU, T.; YANG, Z. Morphological and phisiological
comparison of taxa comprising the Sporothrix schenckii complex. J Zhejiang Univ,
v. 16, n. 11, p. 940-947, 2015.
KLEIN, B.S.; TEBBETS, B. Dimorphism and virulence in fungi. Curr Opin
Microbiol, v. 10, n. 4, p. 314-319, 2007.
KWON-CHUNG, K.J.; BENNETT, J.E.; Sporotrichosis. Medical Mycology
Pennsylvania, p. 707-729, 1992.
LACERDA, C.M.; MARTINS, E.M.; DE RESENDE, M.A.; DE ANDRADE, A.S.
Gamma radiation effects on Sporothrix schenckii yeast cells. Mycophatologia,
v.171, n.6, p.395-401, 2011.
LANIADO-LABORÍN, R.; CABRALES-VARGAS, M.N.; Amphotericin B: side
effects and toxicity. Rev Iberoam Micol, v. 26, n. 4, p. 223-227, 2009.

84
LATGÉ, J.P. Tasting the fungal cell wall. Cell Microbiol, v.12, n.7, p.863-872,
2010.
LEE, C.Y.; THOMPSON III, G.R.; HASTEY, C.J.; HODGE, G.C.; LUNETTA,
J.M.; PAPPAGIANIS, D.; HEINRICH, V. Coccidioides endospores and spherules
draw strong chemotactic, adhesive, and phagocytic responses by individual human
neutrophils. PLoS One, v. 10, n. 6, 2015.
LIU, G.Y.; NIZET, V. Color me bad: microbial pigments as virulence factors.
Trends Microbiol. v. 17, p. 406-413, 2009.
LLOYD, K.O.; BITOON, M.A. Isolation and purification of a peptide-
rhamnomannan from the yeast form of Sporothrix schenckii. Structural and
immunochemical studies. J. Immunol, v.107, n.3, p.663-671, 1971.
LOPES-BEZERRA, L.M.; SCHUBACH, A.; COSTA, R.O. Sporothrix schenckii
and Sporotrichosis. An Acad Bras Cienc, v.78, n.2, p.293-308, 2006.
LOPES-BEZERRA, L.M. Sporothix schenckii cell wall peptidorhamnomannans.
Front Microbiol, v. 2, 2011.
LURIE, H.I. Five unusual cases of Sporotrichosis from South Africa showing
lesions in muscles, bones and viscera. Brit J of Surg, v.50, n.224, p.585-591, 1963.
LUTZ, A.; SPLENDORE, A. Sobre uma mycose observada em homens e ratos. Ver
Med S Paulo, v.21, p.433-450, 1907.
MACCALLUM, D.M.; COSTE, A.; ISCHER, F.; JACOBSEN, M.D.; ODDS, F.C.;
SANGLARD, D. Genetic dissection of azole resistant mechanism in Candida
albicans and their validation in a mouse model of disseminated infection.
Antimicrob Agents Chemother, v.54. n.4, p.1476-1483, 2010.
MACEDO, P.M.; LOPES-BEZERRA, L.M.; BERNARDES-ENGEMANN, A.R.;
OROFINO-COSTA, R. New posology of potassium iodide for the treatment of
cutaneous sporotrichosis: study of efficacy and safety in 102 patients. J Eur Acad
Dermatol Venereol, v. 29, n. 4, p. 719-724, 2015.

85
MAERTENS, J. History of the development of azoles derivates. Clin Microbiol
Infect, v.10, n.1, p.1-10, 2004.
MAGAND, F.; PERROT, J.L.; CAMBAZARD, F.; RABERIN, M.H.; LABEILLE,
B. Autochthonous cutaneous Sporotrichosis in France. Ann Dermatol Venereol,
v.136, n.3, p.273-275, 2009.
MAHAJAN, V.K. Sporotrichosis: an overview and therapeutical options. Dermat
Res Pract, v. 2014, 13p, 2014.
MAHAJAN, V.K.; MEHTA, K.S.; CHAUHAN, P.S.; GUPTA, R.S.; RAWAT, R.
Fixed cutaneous sporotrichosis treated with topical amphotericin B in an immune
suppressed patient. Med Mycol Case Rep, v. 7, p. 23-25, 2015.
MARIMON, R.; CANO, J.; GENÉ, J.; SUTTON, D.A.; KAWASAKI, M.;
GUARRO, J. Sporothrix brasiliensis, S. globosa, and S. mexicana, three new
Sporothrix species of clinical interest. J Clin Microbiol, v. 45, n. 10, p. 3198-3206,
2007.
MARIO, D.N.; GUARRO, J.; SANTURIO, J.M.; ALVES, S.H.; CAPILLA, J. In
Vitro and In Vivo Efficacy of Amphotericin B Combined with Posaconazole against
Experimental Disseminated Sporotrichosis. Antimicrob Ag Chemother, v. 59, n. 8, p.
5018-5021, 2015.
MARQUES-EVANGELISTA, O.M.; ALMEIDA-PAES, R.; DE MEDEIROS-
MUNIZ, M.; DE LIMA-BARROS, B.M.; GUTIERREZ-GALHARDO, C.M.;
ZANCOPÉ-OLIVEIRA, R.M. Sporotrichosis caused by Sporothrix globosa in Rio
de Janeiro, Brazil: case report. Mycopathologia, v.169, n.5, p.359-363, 2010.
MATHEW, B.P.; NATH, M. Recent approaches to antifungal therapy for invasive
mycosis. Chem Med Chem, v. 4, n. 3, p. 310-323, 2009.
MCGUINNESS, S.L.; BOYD, R.; KIDD, S.; MCLEOD, C.; KRAUSE, V.L.;
RAUPH, A.P. Epidemiological investigation of an outbreak of cutaneos
Sporotrichosis, Northern Territory, Australia. BMC Infect Dis, v.16, n.16, 2016.

86
MEDOFF, G.; KOBAYASHI, G.S.; PAINTER, A.; TRAVIS, S. Morphogenesis
and pathogenicity of Histoplasma capsulatum. Infect Immun, v.55, n.6, p.1355-
1358, 1987.
MENDOZA, M.; ALVARADO, P.; DÍAZ DE TORRES, E.; LUCENA, L.; DE
ALBORNOZ, M.C. Comportamiento fisiológico y de sensibilidad in vitro de
aislamientos de Sporothrix schenckii mantenidos 18 años por dos métodos de
preservación. Rev Iberoam Micol, v. 22, n. 3, p. 151-156, 2005.
MILLER, M.B.; BASSLER, B.L. Quorum sensing in bacteria. Ann Review
Microbiol. v. 55, p. 165-199, 2001.
MITCHELL, K.F.; ZARNOWSKI, R.; ANDES, D.R.; The Extracellular Matrix of
Fungal Biofilms. Adv Exp Med Biol, v. 931, p. 21-35, 2016.
MOHR, J.; JOHNSON, M.; COOPER, T.; LEWIS, J.S.; OSTROSKY-ZEICHNET,
L. Current options in antifungal pharmacotherapy. Pharmacotherapy, v. 28, n. 5, p.
614-645, 2008.
MOUDGAL, V.V.; SOBEL, J.D. Antifungal drugs in pregnancy: a review. Expert
Opin Drug Saf, v. 2, n. 5, p. 475-483, 2003.
NICHOLLS, S.; MACCALLUN, D.M.; KAFFARNIK, F.A.; SELWAY, L.; PECK,
S.C.; BROWN, A.J. Activation of the heat shock transcription factor Hsf1 is
essential for the fungal virulence of the fungal pathogen Candida albicans. Fungal
Genet Biol, v. 48, n. 3, p. 297-305, 2011.
NIIMI, M.; FIRTH, N.A.; CANNON, R.D. Antifungal drug resistence of oral fungi.
Odontology. V. 98, n. 1, p. 15-25, 2010.
NUNES, G.D.L.; CARNEIRO, R.S.; FILGUEIRA, F.G.F.; FERNANDES, T.H.T.
Esporotricose felina no município de Itaporanga. Arq Cien Vet Zoo, v.14, n.2, p.157-
161, 2011.
OLIVEIRA, D.C.; LOPES, P.G.M.; SPADER, T.B.; MAHL, C.D.; TRONCO-
ALVES, G.R.; LARA, V.M.; SANTURIO, J.M.; ALVES, S.H. Antifungal

87
susceptibilities of Sporothrix albicans, S. brasiliensis, and S. luriei of the S.
schenckii complex identified in Brazil. J Clin Microbiol, v. 49, n. 8, p. 3047-3049,
2011.
OLIVEIRA, M.M.; SANTOS, C.; SAMPAIO, P.; ROMEO, O.; ALMEIDA-PAES,
R.; PAIS, C.; LIMA, N.; ZANCOPÉ-OLIVEIRA, R.M. Development and
optimization of a new MALDI-TOF protocol for identification of
the Sporothrix species complex. Res Microbiol, vol. 166, n. 2, p. 102-110, 2014.
OLIVEIRA, D.C.; LORETO, E.S.; MARIO, D.A.N.; LOPES, P.G.M.; NEVES,
L.V.; ROCHA, M.P.; SANTURIO, J.M.; ALVES, S.H. Sporothrix schenckii
complex: susceptibilities to combined antifungal agents and characterization of
enzymatic profiles. Rev Inst Med Trop Sao Paulo, v. 57, n. 4, p. 289-294, 2015.
PAIXÃO, A.G.; GALHARDO, M.C.; ALMEIDA-PAES, R.; NUNES, E.P.;
GONÇALVES, M.L.; CHEQUER, G.L.; LAMAS CDA, C.; The difficult
management of disseminated Sporothrix brasiliensis in a patient with advanced aids.
AIDS Res Ther, v.12, n.16, 2015.
PEREIRA, S.A.; GREMIÃO, I.D.; KITADA, A.A.; BOECHAT, J.S.; VIANA,
P.G.; SCHUBACH, T.M. The epidemiological scenario of feline sporotrichosis in
Rio de Janeiro, State of Rio de Janeiro, Brazil. Rev Soc Bras Med Trop, v.47, n.3,
p.392-393, 2014.
PÉREZ-SANCHEZ, L.; GONZÁLEZ, E.; CÓLON-LORENZO, E.E.;
GONZÁLEZ-VELAZQUEZ, W.; GONZÁLEZ-MÉNDEZ, R.; RODRÍGUEZ-DEL
VALLE, N.; Interaction of the heterotrimeric G protein alpha subunit SSG-1 of
Sporothrix schenckii with proteins related to stress response and fungal
pathogenicity using a yeast two-hybrid assay. bmc Microbiol, v. 10, n. 317, 2010.
PERUMAL, P.; MEKALA, S.; CHAFFIN, W.L. Role of cell density in antifungal
drug resistence in Candida albicans biofilms. Antimicrob Agents and
Chemotherapy. v. 51, n. 7, p. 2454-2463, 2007.
PIERCE, C.G.; UPPULURI, P.; TRISTAN, A.R.; WORMLEY, F.L.; MOWAT, E.;
RAMAGE, G.; LOPEZ-RIBOT, J.L. A simple and reproducible 96-well plate-based

88
method for the formation of fungal biofilms and its application to antifungal
susceptibility testing. Nat Protoc, v. 3, n. 9, p. 1494-1500, 2008.
PITANGUI, N.S.; SARDI, J.C.O.; SILVA, J.F.; BENADUCCI, T.; MORAES DA
SILVA, R.A.; RODRÍGUEZ-ARELLANES, G.; TAYLOR, M.L.; MENDES-
GIANNINI, M.J.S.; FUSCO-ALMEIDA, A.M. Adhesion of Histoplasma
capsulatum to pneumocytes and biofilm formation on an abiotic surface. Biofouling,
v. 28, n. 7, p. 711-718, 2012.
PJIPER, A.; PULLINGER, B.D. An outbreak of Sporotrichosis among South
African native miners. The Lancet, v.210, n.5435, p.914-916, 1927.
PUPO, J.A. Frequência da Sporotrichose em São Paulo. Annaes Paulista de Med e
Cirurgia, v. 8, n. 3, p. 53-68, 1917.
RAMAGE, G.; MARTÍNEZ, J. P.; LÓPEZ-RIBOT, J. L. Candida biofilms on
implanted biomaterials: a clinically significant problem. FEMS Yeast Res, v. 6, n. 7,
p. 979-986, 2006.
RAMAGE, G.; RAJENDRAN, R.; SHERRY, L.; WILLIAMS, C. Fungal biofilm
resistance. Int J Microbiol, v. 2012, 2012.
RAMÍREZ-SOTO, M.C. Sporotrichosis: The Story of an endemic region in Peru
over 28 years (1985 to 2012). PLoS ONE, v.10, n.6, 2015.
REED, K.D.; MOORE, F.M.; GEIGER, G.E.; STEMPER, M.E. Zoonotic
Transmission of Sporotrichosis: Case Report and Review. Clin Infect Dis, v.16,
p.384-387, 1993.
ROBERTS, C.G. The role of biofilms in reprocessing medical devices. Am J Infect
Control, v. 41, n. 5, p. S77-80, 2013.
RODRIGUES, A.M.; CRUZ CHOAPPA, R.; FERNANDES, G.F.; DE HOOG,
G.S.; DE CAMARGO, Z.P. Sporothrix chilensis sp. nov. (Ascomycota:
Ophiostomatales), a soil-borne agent of human sporotrichosis with mild-pathogenic
potential to mammals. Fungal Biol, v.120. n. 2, p. 246-264, 2016.

89
RODRIGUES, A.M.; DE HOOG, S.; DE CARMARGO, Z.P. Emergence of
pathogenicity in the Sporothrix schenckii complex. Med Mycol, v. 51, n. 4, p. 405-
412, 2013a.
RODRIGUES, A.M.; DE HOOG, S.; DE CARMARGO, Z.P. Molecular diagnosis
oh pathogenic Sporothrix species. PLoS Negl Trop Dis, v. 9, n. 12, 2015.
RODRIGUES, A.M.; DE HOOG, G.S.; PIRES, D.C.; BRILHANTE, R.S.N.;
SIDRIM, J.J.C.; GADELHA, M.F.; COLOMBO, A.L.; DE CAMARGO, Z.P.
Genetic diversity and antifungal susceptibility profiles in causative agents of
Sporotrichosis. BMC Infect Dis, v.14, n.219, 2014.
RODRIGUES, A.M.; DE MELO TEIXEIRA, M.; DE HOOG, G.S.; SCHUBACH,
T.M.; PEREIRA, S.A.; FERNANDES, G.F.; BEZERRA, L.M.; FELIPE, M.S.; DE
CAMARGO, Z.P. Phylogenetic analysis reveals a high prevalence of Sporothrix
brasiliensis in feline sporotrichosis outbreaks. PLoS Negl Trop Dis, v. 7, n. 6,
e2281, 2013b.
RODRIGUEZ-CABAN, J.; GONZALEZ-VELAZQUEZ, W.; PÉREZ-SÁNCHEZ,
L.; GONZALEZ-MENDEZ, R.; RODRIGUEZ-DEL VALLE, N.
Calcium/calmodulin kinase1 and its relation to thermotolerance and HSP90 in
Sporothrix schenckii: an RNAi and yeast two-hybrid study. BMC Microbiol, v.11,
n.162, 2011.
ROMEO, O.; CRISEO, G. What lies beyond genetic diversity in Sporothrix
schenckii species complex? New insights into virulence profiles, immunogenicity
and protein secretion in S. schenckii sensu stricto isolates. Virulence, v. 4, n. 3, p.
203-206, 2013.
RUIZ-BACA, E. TORIELLO, C.; PEREZ-TORRES, A.; SABANERO-LOPEZ, M.;
VILLAGOMEZ-CASTRO, J.C.; LOPEZ-ROMERO, E. Isolation and some
properties of a glycoprotein of 70 kDa (Gp70) from the cell wall of Sporothrix
schenckii involved in fungal adherence to dermal extracellular matrix. Med Mycol,
v.47, v.2, p.185-96, 2009.

90
RUIZ-BACA, E.; MORA-MONTES, H.M.; LÓPEZ-ROMERO, E.; TORIELLO,
C.; MOJICA-MARÍN, V.; URTIZ-ESTRADA, N.; 2D-immunoblotting analysis of
Sporothrix schenckii cell wall. Mem Inst Oswaldo Cruz, v.106, n.2, p.248-250,
2011.
SÁNCHEZ-HERRERA, R.; FLORES-VILLAVICENCIO, L.L.; PADILLA-
GUERRERO, I.; BARBOSA-SABANERO, G.; SABANERO-LÓPEZ, M.
Formación de biopelículas en el hongo patógeno Sporothrix schenckii: desarrollo,
arquitectura y características bioquímicas. Acta Universitaria, v.24, n. NE-2, p. 11-
15, 2014.
SARDI, J.C.O.; PITANGUI, N.S.; VOLTAN, A.R.; BRAZ, J.D.; MACHADO,
M.P.; FUSCO ALMEIDA, A.M.; GIANNINI, M.J.S.M. In vitro Paracoccidioides
brasiliensis biofilm and gene expression of adhesins and hydrolytic enzymes.
Virulence, v. 6, n. 6, p. 642-651, 2015.
SASAKI, A.A.; FERNANDES, G.F.; RODRIGUES, A.M.; LIMA, F.M.; MARINI,
M.M.; DOS S FEITOSA, L.; DE MELO TEIXEIRA, M.; FELIPE, M.S.; DA
SILVEIRA, J.F.; DE CAMARGO, Z.P. Chromosomal polymorphism in the
Sporothrix schenckii complex. PLoS ONE, v. 9, n. 1, e86819, 2014.
SCHENCK, B.R. On refractory subcutaneous abcess caused by a fungus possibly
related to the Sporotricha. Bull. Johns Hopkins Hosp. v.9, p. 286-290, 1898.
SELEEM, D.; BENSO, B.; NOGUTI, J.; PARDI, V.; MURATA, R.M. In vitro and
in vivo antifungal activity of Linchochalcone-A against Candida albicans biofilms.
PLoS One, v. 11, n. 6, 2016.
SENGUPTA, J.; SAHA, S.; KHETAN, A.; SARKAR, S.K.; MANDAL, S.M.
Effects of lactoferricin B against keratitis-associated fungal biofilms. J Infect
Chemother, v. 18, n. 5, p. 698-703, 2012.
SGARBI, D.B.G.; SILVA, A.J.R.; CARLOS, I.Z.; SILVA, C.L.; ANGLUSTER, J.;
ALVIANO, C.S. Isolation of ergosterol peroxide and its reversion to ergosterol in
pathogenic fungus Sporothrix schenckii. Mycopathologia, v.139, n.1, p.9-14, 1997.

91
SHEPARDSON, K.M.; CRAMER, R.A. Fungal cell wall dynamics and infection
site microenvironments: signal integration and infection outcome. Curr Opin
Microbiol, v. 16, n. 4, p. 385-390, 2013.
SILVA, S.; NEGRI, M.; HENRIQUES, M.; OLIVEIRA, R.; WILLIAMS, D.W.;
AZEREDO, J. Adherence and biofilm formation on non-Candida albicans Candida
species. Trends Microbiol, v. 19, n. 5, p. 241-247, 2011.
SILVA, M.B.T.; COSTA, M.M.M.; TORRES, C.C.S.; GALHARDO, M.C.G.;
VALLE, A.C.F.; MAGALHÃES, M.A.F.M.; SABROZA, P.C; OLIVEIRA, R.M.;
Urban sporotrichosis: a negleted epidemic in Rio de Janeiro, Brazil. Cad Saude
Publ, v.28, n.10, p.1867-1880, 2012.
SILVA-VERGARA, M.L.; DE CAMARGO, Z.P.; SILVA, P.F.; ABDALLA, M.R.;
SQARBIERI, R.N.; RODRIGUES, A.M.; DOS SANTOS, K.C.; BARATA, C.H.;
FERREIRA-PAIM, K. Disseminated Sporothrix brasiliensis infection with
endocardial and ocular involvemente in na HIV-infected patient. Am J Trop Med
Hyg, v.86, n.3, p.477-480, 2012.
SMEGO, R.A. Jr.; CASTIGLIA, M.; ASPERILLA, M.O. Lymphocutaneous
syndrome: a review of non-Sporothrix causes. Medicine (Baltimore), v. 78, n. 1, p.
38-63, 1999.
SONG, Y., LI, S.S.; ZHONG, S.X.; LIU, Y.Y.; YAO, L.; HUO, S.S. Report of 457
sporotrichosis cases from Jilin province, northeast China, a serious endemic region.
Journ Eur Acad Dermatol Venereol, v.27. n.3, 313-318, 2013.
STEENBERGEN , J.N.; NOSANCHUK, J.D.; MALLIARIS, S.D.;
CASADEVALL, A. Interaction of Blastomyces dermatitidis, Sporothrix schenckii
and Histoplasma capsulatum with Acanthamoeba castellanii. Infect Immun. v. 72, n.
6, 3478-3488, 2004.
STERLING, J.B.; HEYMANN, W.R. Potassium iodide in dermatology: a 19th
century drug for the 21st century uses, pharmacology, adverse effects and
contraindications. J Am Acad Dermatol, v.43, n.4, p.691-697, 2000.

92
TABORDA, C.P.; SILVA, M.B.; NOSANCHUK, J.D.; TRAVASSOS, L.R.
Melanin as a virulence factor of Paracoccidioides brasiliensis and other dimorphic
pathogenic fungi: a minireview. Mycopathologia. v. 165, n.4-5, p. 331-339, 2008.
TAFF, H.T; NETT, J.E.; ZARNOWSKY, R.; ROSS, K.M.; SANCHEZ, H.; CAIN,
M.T.; HAMAKER, J.; MITCHELL, A.P., ANDES, D.R. A Candida biofilm-
induced pathway for matrix glucan delivery: implications for drug resistance. PLoS
Pathog, v. 8, n. 8, e1002848, 2012.
TEIXEIRA, P.A.; DE CASTRO, R.A.; NASCIMENTO, R.C.; TRONCHIN, G.;
TORRES, A.P.; LAZÉRA, M.; DE ALMEIDA, S.R.; BOUCHARA, J.P.;
LOUREIRO Y PENHA, C.V.; LOPES-BEZERRA, L.M.; Cell surface expression of
adhesins for fibronectin correlates with virulence in Sporothrix schenckii.
Microbiology, v. 155, n. Pt11, 2009.
TEIXEIRA, M.M.; ALMEIDA, L.G.P.; KUBITSCHEK-BARREIRA, P.; ALVES,
F.L.; KIOSHIMA, E.S.; ABADIO, A.K.R.; FERNANDES, L.; DERENGOWSKI,
L.S.; FERREIRA, K.S.; SOUZA, R.C.; RUIZ, J.C.; ANDRADE, N.C.; PAES, H.C.;
NICOLA, A.M.; ALBUQUERQUE, P.; GERBER, A.L.; MARTINS, V.P.;
PECONICK, L.D.F.; NETO, A.V.; CHAUCANEZ, C.B.; SILVA, P.A.; CUNHA,
O.L.; OLIVEIRA, F.F.M.; SANTOS, T.C.; BARROS, A.L.N.; SOARES, M.A.;
OLIVEIRA, L.M.; MARINI, M.M.; VILLALOBOS-DUNO, H.; CUNHA, M.M.L.;
DE HOOG, S.; SILVEIRA, J.F.; HENRISSAT, B.; NIÑO-VEGA, G.; CISALPINO,
P.S.; MORA-MONTES, H.M.; ALMEIDA, S.R.; STAJICH, J.E.; LOPES-
BEZERRA, L.M.; VASCONCELOS, A.T.R.; FELIPE, M.S.S. Comparative
genomics of the major fungal agents of human and animal Sporotrichosis:
Sporothrix schenckii and Sporothrix brasiliensis. BMC Genomics, v. 15, n. 943,
2014.
TÉLLEZ, M.D.; BATISTA-DUHARTE, A.; PORTUONDO, D.; QUINELLO, C.;
BONNE-HERNÁNDEZ, R.; CARLOS, I.Z. Sporothrix schenckii complex biology:
environment and fungal pathogenicity. Microbiology, v. 160, n. Pt11, p. 2352-2365,
2014.

93
TORRES-GUERRER, H.; ARENAS-LÓPEZ, G. UV irradiation induced high
frequency of colonial variants of altered morphology in Sporothrix schenckii. Med
Mycol, v.36, n.2, p.81-87, 1998.
TORRES-MENDONZA, B.M.; VÁSQUEZ-VALLS, E.; GONZÁLEZ-
MENDONZA, A. Effect of potassium iodide on the immune response in the
sporotrichosis. Rev Iberoam Micol, v.14, n.3, p.98-100, 1997.
TOURNU, H.; VAN DIJCK, P. Candida biofilms and the host: models and new
concepts for erradication. Int J Microbiol, v. 2012, 16p., 2012.
TRAVASSOS, L.R.; LLOYD, K.O. Sporothrix schenckii and related species of
Ceratocystis. Microbiol Rev, v.44, n.4, p.683-721, 1980.
TRONCHIN, G.; PIHET, M.; LOPES-BEZERRA, L.M.; BOUCHARA, J.O.
Adherence mechanisms in human pathogenic fungi. Med Mycol, v.46, n.8, p.749-
772, 2008.
VALENTÍN-BERRÍOS, S.; GONZÁLEZ-VELÁZQUEZ, W.; PÉREZ-SÁNCHEZ,
L.; GONZÁLEZ-MÉNDEZ, R.; RODRIGUEZ-DEL VALLE, N. Cytosolic
phospholipase A2: a member of the signalling pathway of new G protein α subunit
in Sporothrix schenckii. BMC Microbiol, v.9, n.100, 2009.
VALENTINI, M.; FILLOUX, A. Biofilms and c-di-GMP signaling: lessons from
Pseudomonas aeruginosa and other bacteria. J Biol Chem, v. 291, n. 24, p. 12547-
12555, 2016.
VALLE-AVILES, L.; VALENTIN-BERRIOS, S.; GONZÁLEZ-MENDEZ, R.R.;
RODRIGUEZ-DEL VALLE, N. Functional genetic and bioinformatic
characterization of a calcium/calmodulin kinase gene in Sporothrix schenckii. BMC
Microbiol, v.7, n.107, 2007.
VANDEN BOSSCHE, H.; KOYMANS, L.; MOEREELS, H. P450 inhibitors of use
in medical treatment: focus on mechanism of action. Pharmac Ther, v.67, n.1, p.79-
100, 1995.

94
WALSH, T.J.; ANAISSIE, E.J.; DENNING, D.W.; HERBRECHT, R.;
KONTOYIANNIS, D.P.; MARR, K.A.; MORRISON, V.A.; SEGAL, B.H.;
STEINBACH, W.J.; STEVENS, D.A.; VAN BURIK, J.A.; WINGARD, J.R.;
PATTERSON, T.F. Treatment of Aspergillosis: Clinical Practice Guidelines of
Infectious Diseases Society of America. Clin Infect Dis, v.46, n.3, p.327-60, 2008.
WANG, X.H.; LI, R.Y.; CAO, C.W.; WANG, Y.H.; LIU, W.; QIAO, J.J.; LI, Y.Z.
Differential mRNA expression of Sporothrix schenckii catalase gene in two growth
phases and effects factors. Chin Med J (Engl), v. 121, n. 20, p. 2100-2102, 2008.
WEN-LING, W.; RONG-YA, Y.; JUN-HONG, A.O. Uncommon characteristics of
the structure and development of Trichosporon asahii. Chin Med Journ, v. 122, n.
15, p. 1806-1810, 2009.
WILLIAMS, C.; RAJENDRAN, R.; RAMAGE, G. Aspergillus biofilms in human
disease. Adv Exp Med Biol, 2016.
YAMADA, K.; ZAITZ, C.; FRAMIL, V.M.S.; MURAMATU, L.H. Tratamento da
esporotricose cutânea com SSKI: experiência de 24 anos no estado de São Paulo,
Brasil. Rev Inst Med Trop, v. 53, n.2, p.89-93, 2011.
YEGNESWARAN, P.P.; SRIPATHI, H.; BAIRY, I.; LONIKAR, V.; RAO, R.;
PRABHU, S. Zoonotic Sporotrichosis of lymphocutaneous type in a man acquired
from a domesticated feline source: report of a first case in southern Karnataka,
India. Int J Dermatol, v.48, n.11, p.1198-1200, 2009.
ZHAO, M.; ZHOU, X.; LIU, T.; YANG, Z. Morphological and physiological
comparison of taxa comprising the Sporothrix schenckii complex. J Zhejiang Univ
Sci B, v. 16, n. 11, 2015.
ZHOU, X.; RODRIGUES, A.M.; FENG, P.; DE HOOG, G.S. Global ITS diversity
in the Sporothrix schenckii complex. Fungal Diversity, v. 66, n. 1, p. 153-165, 2014.

95
ANEXO A – PUBLICAÇÃO ORIGINADA A PARTIR DA DISSERTAÇÃO
Antifungal susceptibility of Sporothrix spp. in biofilm form
Raimunda Sâmia Nogueira Brilhante¹*
Felipe Rodrigues Magalhães de Aguiar¹
Maria Lucilene Queiroz da Silva¹
Jonathas Sales de Oliveira¹
Zoilo Pires de Camargo3
Anderson Messias Rodrigues3
Vandbergue Santos Pereira¹
Rosana Serpa¹
Débora Castelo Branco de Sousa Colares¹
Edmilson Emanuel Monteiro Correia¹
Waldemiro Aquino Pereira Neto¹
Rossana de Aguiar Cordeiro¹
Marcos Fábio Gadelha Rocha1,2
José Júlio Costa Sidrim¹.
¹Specialized Medical Mycology Center, Postgraduate Program in Medical Microbiology, Department of
Pathology and Legal Medicine, Federal University of Ceará, Fortaleza, Ceará, Brazil.
²College of Veterinary Medicine, Postgraduate Program in Veterinary Sciences, State University of
Ceará, Fortaleza, Ceará, Brazil.
3Department of Microbiology, Immunology and Parasitology, Cellular Biology Division, Federal
University of São Paulo, São Paulo, Brazil
RESUMO
A esporotricose é a micose subcutânea cosmopolita mais comumente encontrada em áreas tropicais e
subtropicais da América Latina, sendo causada por espécies do complexo Sporothrix schenckii. As
espécies relacionadas à esporotricose são S. brasiliensis, S. schenckii sensu stricto, S. globosa e S.
mexicana. O objetivo deste estudo foi avaliar a sensibilidade dos biofilmes com complexo Sporothrix
schenckii frente às drogas antifúngicas. Para tanto, inicialmente investigou-se a capacidade de fungos
do complexo S. schenckii produzir biofilmes in vitro, e, a seguir, determinou-se a influência da exposição
a concentrações crescentes (MIC da forma planctônica, 10xMIC e 50xMIC) de anfotericina B (AMB),
caspofungina (CAS), cetoconazol (KTC), voriconazol (VRC) e fluconazol (FLC) na biomassa e atividade
metabólica destes biofilmes, mensurada através de ensaios de cristal violeta e XTT, respectivamente.
Após a formação dos biofilmes do complexo S. schenckii, os mesmos foram levados à microscopia óptica
para visualização através da coloração por vermelho-congo, assim como para microscopia eletrônica de
varredura e microscopia confocal. Através das análises de absorbância da biomassa produzida pelos

96
biofilmes maduros, os mesmos foram classificados como não formadores, fracos ou fortes formadores de
biofilmes, de acordo com a quantidade de biomassa produzida. Após a transferência do inóculo fúngico
para poços de placas de poliestireno, incubou-se por 24 horas a 25 °C para a adesão das células à
superfície, passo inicial para a formação dos biofilmes. Depois de 24 horas, o sobrenadante foi retirado e
acrescentou-se meio RPMI 1640 e incubou-se por 120 horas para a maturação do biofilme. Após esse
período, a biomassa foi lida em espectrofotômetro e verificou-se que todas as espécies do complexo S.
schenkii são fortes formadoras de biofilme, não havendo diferença de formação entre as espécies. Os
biofilmes foram analisados por microscopia e percebeu-se a presença de uma densa rede de hifas e
conídios, entrelaçados entre si e imersos em matriz. A presença de canais de água, por onde os fluidos
transportam as substâncias necessárias e produzidas pelas células sésseis. A absorbância dos biofilmes de
cepas de S. brasiliensis (n=10), S. schenckii sensu stricto (n=2), S. globosa (n=2) e S. mexicana (n=4)
após exposição às drogas demonstrou que a AMB e CAS em concentrações de 50xMIC foram capazes de
reduzir significativamente a biomassa e a atividade metabólica dos biofilmes de todas as espécies e KTC,
na mesma concentração, somente não mostrou redução na biomassa do biofilme S. schenckii s. str. A
concentração encontrada para os MICs planctônicos somente conseguiu reduzir a biomassa dos biofilmes
de S. mexicana com VRC e KTC e S. brasiliensis com FLC e CAS, assim como a atividade metabólica
somente dos biofilmes de S. brasiliensis com AMB e KTC. Os antifúngicos utilizados nos testes de
sensibilidade demonstraram redução na atividade metabólica dos biofilmes somente em concentrações
bem acima dos MICs encontrados nas células planctônicas, demonstrando que, entre as mesmas espécies,
há diferença positiva na proteção que o biofilme demonstra contra as drogas antifúngicas em relação à
forma planctônica, mesmo em concentrações mais altas.