universidade estadual de londrina · do trabalho. À profª. drª. daniele sartori, por toda a...
TRANSCRIPT

FERNANDA APARECIDA PIRES FAZION
EFEITO DO GENE cry SOBRE A CAPACIDADE DE
GERMINAÇÃO, CRESCIMENTO E ESPORULAÇÃO DE
Bacillus thuringiensis INFECTANDO LARVAS DE
Anticarsia gemmatalis
Londrina
2013
Universidade Estadual de Londrina

FERNANDA APARECIDA PIRES FAZION
EFEITO DO GENE cry SOBRE A CAPACIDADE DE
GERMINAÇÃO, CRESCIMENTO E ESPORULAÇÃO DE
Bacillus thuringiensis INFECTANDO LARVAS DE
Anticarsia gemmatalis
Defesa apresentada ao Programa de Pós–
Graduação em Genética e Biologia
Molecular, da Universidade Estadual de
Londrina, como requisito para a obtenção do
título de mestre.
Orientadora: Profª. Drª. Gislayne Fernandes
Lemes Trindade Vilas-Bôas.
Londrina
2013

FERNANDA APARECIDA PIRES FAZION
EFEITO DO GENE cry SOBRE A CAPACIDADE DE
GERMINAÇÃO, CRESCIMENTO E ESPORULAÇÃO DE
Bacillus thuringiensis INFECTANDO LARVAS DE
Anticarsia gemmatalis
Defesa apresentada ao Programa de Pós–
Graduação em Genética e Biologia
Molecular, da Universidade Estadual de
Londrina, como requisito para a obtenção do
título de mestre.
BANCA EXAMINADORA
____________________________________
Profª. Drª. Gislayne Fernandes Lemes
Trindade Vilas-Bôas
Universidade Estadual de Londrina
__________________________________
Prof. Dr. Daniel Ricardo Sosa-Gomez
Empresa Brasileira de Pesquisa
Agropecuária
__________________________________
Prof.ª Dr.ª Gisele Maria de Andrade de
Nobrega
Universidade Estadual de Londrina
Londrina, 28 de fevereiro de 2013.

DEDICATÓRIA
À Deus
Aos Meus Pais,
Américo José Fazion
Cleusa Maria Netto Pires Fazion
Às Minhas Irmãs,
Mariangela Pires Fazion
Heloisa Pires Fazion
À Deus, acima de tudo, por me abençoar
nessa caminhada, me ajudando a superar
todos os desafios. Por me proporcionar uma
vida maravilhosa, ao lado de pessoas
incríveis que me ajudam e que me fazem
superar cada dificuldade. Aos meus
familiares, por tudo que me ensinaram e
principalmente pelo apoio incondicional nos
momentos mais difíceis dessa caminhada.
Devo essa conquista à eles, que não me
deixaram desistir dos objetivos.

AGRADECIMENTOS ESPECIAIS
À Profª. Drª. Gislayne Trindade Vilas-Bôas &
Prof. Dr. Laurival Antonio Vilas-Boas
Pela atenção, apoio e por todo conhecimento que me proporcionaram,
me apoiando e ajudando sempre que dúvidas surgiam. Pelos conselhos e principalmente
por todo conhecimento que adquiri nesses anos, agradeço a paciência das imensas
conversas sobre esse grupo fascinante de bactérias e também pelas conversas pessoais,
por sempre entenderem quando estava triste ou precisando de um incentivo, sou muito
grata a tudo.

AGRADECIMENTOS
Ao Programa de Pós-graduação em Genética e Biologia Molecular da
Universidade Estadual de Londrina, pela formação profissional e pessoal.
À Universidade Estadual de Londrina, pelo acolhimento nesses
últimos anos. Ao Prof. Dr. Daniel Ricardo Sosa-Gomez e Prof.ª Dr.ª Gisele Maria de
Andrade de Nobrega, membros da banca examinadora, pela importante contribuição e
disponibilidade em avaliar este trabalho.
Ao Prof. Dr. Túlio Oliveira de Carvalho por toda ajuda com as
análises estatísticas e apoio na interpretação dos resultados.
À EMBRAPA, especialmente ao Prof. Dr. Daniel Ricardo Sosa-
Gomez, juntamente com os técnicos Fábio e Ivanilda, todos com uma simpatia incrível
me proporcionaram estrutura material, conselhos e suporte intelectual para a realização
do trabalho.
À Profª. Drª. Daniele Sartori, por toda a ajuda e contribuição para
melhora deste trabalho.
À Profª. Drª. Ana Lúcia Dias com seu trabalho muito competente na
coordenação da Pós-graduação em Genética e Biologia Molecular me apoiou em um
momento difícil, com uma conversa e um abraço aconchegantes.
Aos meus pais e irmãs, que sempre com um carinho enorme, me
acolheram e me incentivaram com um amor incondicional. Sem essas pessoas
maravilhosas não teria conseguido finalizar essa etapa. Por todas as idas na UEL nos
finais de semana, por entenderem meu trabalho e nunca me deixarem desanimar.
À minha amiga de mestrado Ana Paula que sempre me apoiou e
contribuiu muito com conselhos e principalmente incentivos para que este trabalho

fosse finallizado. Apesar de todos os “perrengues” que passamos esses anos, ela sempre
estava presente pra ouvir e acalmar minhas inseguranças. Não posso esquecer do
divertimento nas aulas de francês, envolvendo o futuro marido João.
Ao meu amigo Gabriel, que sempre me deu conselhos muito bons, e
sempre foi muito preocupado com meus momentos difíceis, agradeço à ele por me
ajudar a superar problemas pessoais que surgiram nessa caminhada.
À queridíssima “Roseane”, com quem me divirto muito, com as
dormidas de crachá e com cassinos, com os almoços às 10:30 e com idas no laboratório
em horários fora do expediente. Sempre me ouvindo a qualquer hora, me dando
conselhos, mesmo que pelo telefone, sempre me compreende e me ajuda a superar cada
desafio que surge, me dizendo palavras de carinho e aconchego, nas quais me sinto
melhor pra continuar meu caminho.
Ao meu amigo Thiago Vidotto, com junto com a Josi me
proporcionou momentos muito alegres e teve disposição para me presentear com fotos
maravilhosas das minhas queridas lagartas.
À Amanda, vulgo João, que sempre está de bom humor e fazendo as
mulheres do lab gastarem no eBay, mas estou totalmente agradecida por toda a ajuda e
preocupação que a Amanda dedicou para que nosso trabalho, por todas as lagartas
maceradas, diluídas e inoculadas que enfrentamos juntas. Por todo apoio emocional que
ela me proporcionou, me animando sempre que as coisas ficavam difíceis.
À mulherada do lab, com quem adoro conviver e que me animam a
cada dia e me proporcionam muitas risadas. À Lully que é sempre animada e tem
conselhos sensatos pra me apoiar. A Pri que está começando no lab, sendo sempre
simpática e prestativa, sem falar nos deliciosos cookies. À querida Fer, que é uma
pessoa maravilhosa, que sempre torceu por mim e me incentivou com conversas ótimas
que me fizeram sentir conforto e vontade de continuar. À Jana e Kelly, sempre queridas
e ajudando a manter o lab muito agradável.

Ao Andrézinho, o melhor músico, sempre que estava presente o lab
tudo era muito mais animado, com as “melhores músicas” (sertanejas), e com as
conversas mais engraçadas. Ao André Nadal, com seu inglês que me ajudou muito e
com as análises de bioestastística. Ao Shang, que mesmo por ao estar mais no lab
ajudou muito com as conversas mirabolantes e assuntos inesperados.
À Edna pelas conversas e incentivos, pelas palavras de carinho e
sabedoria que me proporcionou nesses anos.
Ao técnico João Godoy pelos momentos de descontração nesses anos.
À querida amiga Sueli, secretária do Programa de Mestrado em
Genética e Biologia Molecular, por toda disposição em sempre nos ajudar e nos lembrar
de todos os prazos possíveis.
À CAPES pela concessão da bolsa de mestrado.
À Deus, por definitivamente tudo. Sempre estando ao meu lado e me
dando cada dia mais força e coragem para encarar as dificuldades.

FAZION, Fernanda Aparecida Pires; VILAS-BÔAS, Gislayne Fernandes Lemes
Trindade. EFEITO DO GENE cry SOBRE A CAPACIDADE DE GERMINAÇÃO,
CRESCIMENTO E ESPORULAÇÃO DE Bacillus thuringiensis INFECTANDO
LARVAS DE Anticarsia gemmatalis. 2013. 79. Defesa (Programa de Pós–Graduação em
Genética e Biologia Molecular) - Universidade Estadual de Londrina, Londrina, 2012.
RESUMO
A sistemática bacteriana não possui até hoje consenso quanto ao conceito de espécie, o
que tem gerado discussões sobre a classificação de diferentes espécies, como as
estudadas dentro do grupo Bacillus cereus sensu lato, que inclui além de Bacillus
anthracis, B. cereus sensu stricto e Bacillus thuringiensis. Essas duas últimas espécies
compartilham um grande nível de similaridade genética e são fenotipicamente idênticas.
A diferenciação destas espécies é realizada por um gene plasmidial, denominado gene
cry. Esse gene codifica um cristal paraesporal (composto por proteínas Cry) que confere
a atividade entomopatogênica. Uma das formas de dispersão desses plasmídeos é pelo
processo de conjugação. Essa troca de material genético entre as espécies de bactérias
dificulta a classificação dessas espécies. Com o objetivo de avaliar a influência do
plasmídeo pHT73 e do gene cry1Ac sobre o comportamento de linhagens de B.
thuringiensis, larvas de 2º ínstar de Anticarsia gemmatalis, foram alimentadas com
dietas contendo culturas esporuladas dessa espécie bacteriana. Em seguida, cada larva
foi avaliada quanto à germinação de esporos, multiplicação e sobrevivência das
linhagens selvagens, B. thuringiensis subsp. thuringiensis 407-1; B. thuringiensis
subsp. kurstaki KT0 pHT73-EmR e B. thuringiensis subsp. kurstaki KT0
pHT73Δcry1Ac TetR, e linhagens exconjugantes B. thuringiensis subsp. thuringiensis
407-1 pHT73-EmR e B. thuringiensis subsp. thuringiensis 407-1 pHT73Δcry1Ac Tet
R,
sendo que cada experimento foi realizado com 10 repetições. As avaliações foram
realizadas em diversos tempos, incluindo um tempo antes da morte, t0, t24 e t48 horas
após a morte das larvas. As análises estatísticas evidenciaram que linhagens contendo o
plasmídeo pHT73-EmR se multiplicam em uma taxa maior que linhagens contendo o
plasmídeo pHT73Δcry1Ac TetR, demonstrando que o gene cry confere uma vantagem
nas linhagens que o abrigam. As linhagens que possuem o plasmídeo pHT73Δcry1Ac
TetR
se multiplicam a uma taxa maior que linhagens que não apresentam nenhum dos
dois plasmídeos estudados. Portanto, as linhagens que possuem o plasmídeo pHT73-
EmR
são mais adaptadas ao ambiente larval quando comparadas às linhagens que
carregam o plasmídeo pHT73Δcry1Ac TetR
ou as que não possuem nenhum dos dois
plasmídeos. As linhagens que abrigam o plasmídeo pHT73Δcry1Ac TetR
possuem uma
vantagem em relação as linhagens que não contem nenhum dos dois plasmídeos, com
isso pode-se inferir que existem outros genes plasmidiais que influenciam no
comportamento de linhagens desse grupo bacteriano. Estudos posteriores poderão
evidenciar se essa influência do plasmídeo pHT73 e do gene cry1Ac sobre o
comportamento de linhagens de B. thuringiensis é espécie específico ou se essa
influência ocorre também em linhagens de B. cereus, além de apontar o papel desses
genes e sua importância no comportamento bacteriano.
Palavras-chave: Bacillus cereus sensu lato. Comportamento. Conjugação. Plasmídeo.

FAZION, Fernanda Aparecida Pires; VILAS-BÔAS, Gislayne Fernandes Lemes
Trindade. EFFECT OF cry GENE IN BEHAVIOR OF Bacillus thuringiensis
STRAINS IN Anticarsia gemmatalis LARVAE. 2013. 70. Defesa (Programa de Pós–
Graduação em Genética e Biologia Molecular) - Universidade Estadual de Londrina,
Londrina, 2012.
ABSTRACT
The bacterial systematic has so far not consensus about species concept, which has
generated discussions on the classification of different species, such as those studied
within the group Bacillus cereus sensu lato, which includes mainly Bacillus anthracis,
B. cereus sensu stricto and Bacillus thuringiensis. These latter two species share a high
level of chromosomal similarity and are phenotypically identical. The differentiation of
these species is carried by plasmid genes, denominated cry genes. These genes encode a
paraesporal crystal (composed by Cry proteins) that confers entomopathogenic activity.
The dispersion of these plasmids occurs by conjugation process. This exchange of
genetic material between bacterial species complicates their classification. Aiming to
evaluate the behavior of B. thuringiensis strains, second instar Anticarsia gemmatalis
larvae were fed with diets containing sporulated cultures of this bacterial species. Thus,
each larva was assessed for spore germination, proliferation and survival of the strains
B. thuringiensis subsp. thuringiensis 407-1; B. thuringiensis subsp. kurstaki KT0
pHT73-EmR and B. thuringiensis subsp. kurstaki KT0 pHT73Δcry1Ac Tet
R, and
exconjugants strains, B. thuringiensis subsp. thuringiensis 407-1 pHT73-EmR and B.
thuringiensis subsp. thuringiensis 407-1 pHT73Δcry1Ac TetR, and each experiment was
performed with 10 repetitions. The evaluations were done at different times, including
one time before death, t0, t24 and t48 hours after the death of the larvae. Statistical
analyzes showed that strains containing plasmid-pHT73-EmR multiply at a higher rate
than strains containing plasmid pHT73Δcry1Ac TetR, demonstrating that the cry gene
confers an advantage in strains that harbor. Strains that containing the plasmid
pHT73Δcry1Ac TetR multiply at a higher rate than strains that have none of the two
plasmids studied. Therefore, strains containing the plasmid pHT73-EmR are more
adapted to the larval environment when compared to strains that carry the plasmid
pHT73Δcry1Ac TetR or those without any of the two plasmids. Strains containing the
plasmid pHT73Δcry1Ac TetR have are more adapted than strains that do not contains
any of the two plasmids. Therefore, it can be inferred that there are other plasmid genes
that influence the behavior of this group of bacterial strains. Further studies may reveal
whether the influence of the pHT73 plasmid and of the cry1Acgene about the behavior
of B. thuringiensis strains is species specific or whether this effect also occurs in strains
of B. cereus, besides indicating the role of these genes and their importance in bacterial
behavior.
Key words: Bacillus cereus sensu lato. Behavior. Conjugation. Plasmid.

LISTA DE FIGURAS
Figura 1 – A ecologia complexa de B. thuringiensis. Diagrama dos locais em que B.
thuringiensis tem sido isolado e, onde, pode ocorrer multiplicação celular. Possíveis
locais de multiplicação incluem o solo, a rizosfera, o filoplano e outros tecidos das
plantas, dentro de insetos vivos ou mortos, ou dentro de outros invertebrados. B.
thuringiensis é mostrado na forma vegetativa (retângulos vermelhos) e na forma de
esporos (elipses amarelas) e cristais (losangos rosas) .................................................... 1
Figura 2 – Modo de ação das proteínas Cry. (1) Solubilização e ativação da toxina; (2)
ligação dos monômeros da toxina no primeiro receptor induzindo a mudança na
conformação da toxina (3) formação do oligômero; (4) ligação do oligômero de toxina
no segundo receptor; (5) inserção do oligômero da toxina na membrana e formação do
poro .................................................................................................................................. 3
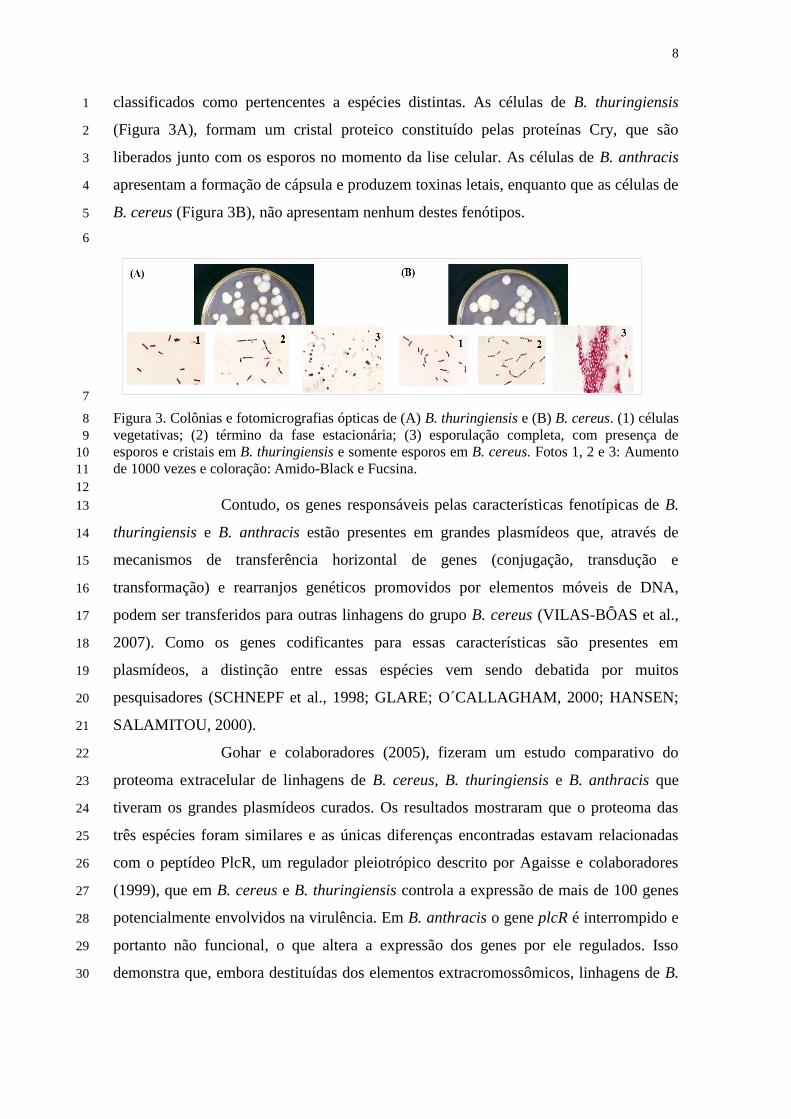
Figura 3 – Colônias e fotomicrografias ópticas de (A) B. thuringiensis e (B) B. cereus.
(1) células vegetativas; (2) término da fase estacionária; (3) esporulação completa, com
presença de esporos e cristais em B. thuringiensis e somente esporos em B. cereus ..... 8
Figura 4 – Eletromicrografia de varredura do processo de conjugação em bactérias
Gram-positivas, compreendendo células doadoras (linhagem AND801) e células
receptoras (linhagem GBJ001) ...................................................................................... 11
Figura 5 – Representação esquemática dos modelos de retromobilização. (A) Modelo
de transferência bidirecional: transferência do plasmídeo conjugativo para a célula
receptora ocorre simultaneamente com a transferência do plasmídeo mobilizável para a
doadora por apenas um evento de conjugação. (B) Modelo de transferência
unidirecional: o primeiro evento conjugativo permite a transferência do plasmídeo
conjugativo para a receptora, enquanto que um segundo evento permite a transferência
do plasmídeo mobilizável para a doadora. Elipse grande: plasmídeo conjugativo;
Círculo pequeno: plasmídeo mobilizável; linha fina: célula receptora; linha grossa:
célula doadora; setas brancas: indicação da transferência dos plasmídeos e setas pretas:
resultado da transferência .............................................................................................. 13
Figura 6 – Representação esquemática de conjugação biparental e triparental.
Conjugação refere-se à transferência de plasmídeos conjugativos, enquanto a
mobilização e retromobilização refere-se à transferência de plasmídeos mobilizáveis.
(A) Conjugação biparental: conjugação para a receptora (1); (B) Mobilização biparental:

conjugação para a receptora (1), mobilização para a receptora (2), e transferência de
ambos os plasmídeos para a receptora (3). (C) Retromobilização biparental: conjugação
para a receptora (1) e retromobilização para a doadora (2). (D) Encontro triparental:
conjugação para a receptora (1) e para a linhagem auxiliar (2), mobilização para a
receptora (3), transferência de ambos os plasmídeos para a receptora (4), e
retromobilização para a doadora (5). Elipse grande: plasmídeo conjugativo; Círculo
pequeno: plasmídeo mobilizável; linha fina: célula receptora; linha grossa: célula
doadora; setas brancas: indicação da transferência dos plasmídeos; Seta branca em
formato de “T”: transferência de ambos os tipos de plasmídeos e setas pretas: resultado
da transferência .............................................................................................................. 15

LISTA DE ABREVIATURAS, SÍMBOLOS E UNIDADES
°C Graus Celsius
δ-endotoxinas Delta-endotoxinas
μg Micrograma
µL Microlitro
µm Micrômetro
AFLP Polimorfismo de Comprimentos de
Fragmentos Amplificados
ATP Adenosina Trifosfato
BP Bacto-peptona
cm Centímetro
cry Gene codificador da proteína Cry
cry1Ac Gene codificador da proteína Cry1Ac
presente no plasmídeo conjugativo
pHT73
Cry Proteína com atividade
entomopatogênica produzida por
linhagens de B. thuringiensis
CytK Citotoxina – K
DNA Ácido desoxirribonucleico
D. O. Densidade óptica
EmR
Linhagem resistente à eritromicina
h Hora
HBL Hemolisina BL
His Histidina
kb Kilobase
LB Luria-Bertani
MGEs Elementos genéticos móveis
mL Mililitro
MLEE Eletroforese de aloenzimas
MPF Encontro formador de poro
NHE Enterotoxina não hemolítica
ICEs Elementos integrativos conjugativos
IMES Elementos integrativos e mobilizáveis
oriT Origem de transferência
PFGE Eletroforese em campo pulsado
pH Potencial hidrogeniônico
pHT73 - EmR
Plasmídeo conjugativo contendo o gene
cry intacto
pHT73Δcry1Ac TetR
Plasmídeo conjugativo com o gene cry

deletado
plcR Gene codificador de um regulador
pleiotrópico
RifR
Linhagem resistente à rifampicina
RNA Ácido ribonucleico
rRNA RNA ribossomal
Ser Serina
SmR
Linhagem resistente à estreptomicina
ssDNA Fita simples de DNA
t Tempo
T4CP Proteína de acoplamento do tipo IV
T4SS Sistema de secreção do Tipo IV
TetR
Linhagem resistente à tetraciclina
Tyr Tirosina
UFC Unidade formadora de colônia

SUMÁRIO
1. FUNDAMENTAÇÃO TEÓRICA ......................................................................... 01
1.1 CARACTERÍSTICAS GERAIS E TAXONOMIA DO GRUPO Bacillus cereus ..................... 01
1.1.1 B. thuringiensis .................................................................................................... 01
1.1.2 B. cereus ............................................................................................................... 04
1.1.3 Taxonomia do Grupo do B. cereus ...................................................................... 06
1.2 CONJUGAÇÃO BACTERIANA ...................................................................................... 10
1.2.1 Características dos Plasmídeos e do Processo de Conjugação ............................. 12
1.2.2 Conjugação em B. thuringiensis ........................................................................... 15
1.3 ECOLOGIA DE B. thuringiensis .................................................................................. 17
2 OBJETIVOS ............................................................................................................. 23
2.1 OBJETIVOS GERAIS .................................................................................................. 23
2.2 OBJETIVOS ESPECÍFICOS ......................................................................................... 23
REFERÊNCIAS ........................................................................................................... 24
3 ARTIGO .................................................................................................................. 37
3.1 RESUMO ................................................................................................................... 38
3.2 INTRODUÇÃO............................................................................................................. 39
3.3 MATERIAL E MÉTODOS ............................................................................................. 41
3.3.1 Linhagens Utilizadas ............................................................................................ 41
3.3.2 Características dos Experimentos de Conjugação ............................................... 41
3.3.3 Conjugação em Suporte Sólido ............................................................................ 41
3.3.4 Produção da suspensão de esporos-cristais .......................................................... 42
3.3.5 Comportamento de Linhagens de Bacillus thuringiensis e Bacillus cereus em
Larvas de A. gemmatalis................................................................................................ 42
3.3.5 Análise dos Resultados ........................................................................................ 44
3.4 RESULTADOS ........................................................................................................... 45
3.5 DISCUSSÃO .............................................................................................................. 46
REFERÊNCIAS ........................................................................................................... 50

ANEXOS .......................................................................................................................53
ANEXO A – Propriedades das linhagens empregadas neste estudo ............................. 54
ANEXO B – Etapas de montagem do experimento. (A) Visão geral da bandeja plástica
com os discos foliares; (B) A. gemmatalis sendo inserida individualmente no poço; (C)
Final da montagem vedação da bandeja plástica com uma camada plástica
...................................................................................................................................... 55
ANEXO C – Fotomicroscopia das linhagens (A) B. thuringiensis subsp. kurstaki KT0
pHT73-EmR evidenciando os cristais bipiramidais (c) e esporos (e) e (B) linhagem B.
thuringiensis subsp. kurstaki KT0 pHT73Δcry1Ac TetR, contendo apenas esporos (e).
Aumento de 1000 vezes e coloração: Amido-Black e Fucsina
...................................................................................................................................... 56
ANEXO D – Demonstração das etapas de infecção pela linhagem B. thuringiensis
subsp. kurstaki KT0 pHT73-EmRe exconjugantes com o plasmídeo pHT73-Em
R em A.
gemmatalis. (A) antes do tratamento; (B) logo após morte (t0); (C) 12 horas após a
morte (t12) .................................................................................................................... 57
ANEXO E – Desenvolvimento de linhagens de B. thuringiensis em cadáveres de A.
gemmatalis. Somente as linhagens B. thuringiensis subsp. kurstaki KT0 pHT73-EmR
e
B. thuringiensis subsp. thuringiensis 407-1 pHT73-EmR são tóxicas para as larvas.
Larvas alimentadas com a suspensão contendo outras linhagens foram mortas
mecanicamente. Tempos 0, 24 e 48 representam o tempo após a morte larval. Os
resultados apresentados são médias de três experimentos independentes. Barra de
desvio-padrão são mostradas (p<0,05). Amostras sem tratamento de calor;
Amostras com tratamento de calor (80 ºC por 20 min)
...................................................................................................................................... 58
ANEXO F - Modelo de comportamento temporal de linhagens de B. thuringiensis em
cadáveres de A. gemmatalis. Somente as linhagens B. thuringiensis subsp. kurstaki
KT0 pHT73-EmR
e B. thuringiensis subsp. thuringiensis 407-1 pHT73-EmR são
tóxicas para as larvas. Valores plotados correspondem aos anteriores ao tratamento de
calor. Tempos 0, 24 e 48 representam o tempo após a morte larval. A) Valores
obsevados; Valores médios;
Linha de tendência da linhagem B. thuringiensis
subsp. kurstaki KT0 pHT73-EmR
(inclinação da reta: 1,3285); Valores observados;
ppValores médios; Linha de tendência da linhagem da linhagem B. thuringiensis
subsp. kurstaki KT0 pHT73Δcry1Ac TetR
(inclinação da reta: 0,6598); B) Valores

observados; Valores médios;
Linha de tendência da linhagem B. thuringiensis
subsp. thuringiensis 407-1 (inclinação da reta: 3,047); Valores observados;
pppValores médios; Linha de tendência da linhagem B. thuringiensis subsp.
thuringiensis 407-1 pHT73-EmR
(inclinação da reta: 4,198); Valores observados;
pppValores médios;
Linha de tendência das médias da linhagem B. thuringiensis
subsp. thuringiensis 407-1 pHT73Δcry1Ac TetR;
(inclinação da reta: 3,043);59

1
1. FUNDAMENTAÇÃO TEÓRICA 1
2
1.1. Características gerais e taxonomia do grupo Bacillus cereus 3
4
1.1.1 Bacillus thuringiensis 5
6
B. thuringiensis destaca-se, entre as linhagens pertencentes ao grupo B. 7
cereus, por ser a bactéria mais conhecida e estudada do grupo (VILAS-BÔAS et al., 8
2007). Trata-se de um bacilo Gram-positivo, aeróbio, anaeróbio facultativo, 9
quimioheterotrófico e mesófilo com temperatura ótima de crescimento de 30 ºC ± 2 º C. 10
As células possuem dimensões de 1,0 - 1,2 µm de largura por 3,0 - 5,0 µm de 11
comprimento, são capazes de se movimentarem por meio de flagelos peritríquios, são 12
resistentes a antibióticos β-lactâmicos, como ampicilina e penicilina. B. thuringiensis é 13
cosmopolita, sendo encontrado desde amostras de solo (MORAES; CAPALBO, 1986; 14
OHBA; AIZAWA, 1986; MARTIN; TRAVERS, 1989; APTOSOGLOU; 15
SIVROPOULOU; KOLIAIS, 1997; VILAS-BÔAS; LEMOS, 2004), filoplano (SMITH; 16
COUCHE, 1991; DAMGAARD et al., 1998), em insetos vivos ou mortos 17
(BERNHARD et al., 1997; CHAUFAUX et al., 1997; CHILCOTT; WIGLEY, 1993) e 18
até em grãos estocados (MEADOWS et al., 1992) (Figura 1). 19
20
Figura 1. A ecologia complexa de B. thuringiensis. Diagrama dos locais em que B. thuringiensis 21
tem sido isolado e, onde, pode ocorrer multiplicação celular. Possíveis locais de multiplicação 22
incluem o solo, a rizosfera, o filoplano e outros tecidos das plantas, dentro de insetos vivos ou 23
mortos, ou dentro de outros invertebrados. B. thuringiensis é mostrado na forma vegetativa 24
(retângulos vermelhos) e na forma de esporos (elipses amarelas) e cristais (losangos rosas) 25
(Fonte: RAYMOND, 2010a). 26
27

2
A característica que distingue B. thuringiensis de outros bacilos, como 1
B. cereus, é que, ao desenvolver o ciclo de esporulação durante a fase estacionária, 2
juntamente com um esporo elipsoidal, há a formação de proteínas, denominadas Cry, 3
que se juntam e cristalizam ainda dentro da célula vegetativa, formando os cristais 4
proteicos (CRICKMORE et al., 1998; YAMAMOTO; DEAN, 2000). Esses cristais 5
garantem o caráter entomopatogênico às linhagens de B. thuringiensis (GLARE; 6
O´CALLAGHAM, 2000). Essas proteínas são codificadas pelos genes cry, localizados 7
principalmente em grandes plasmídeos (40-200 MDa), porém podem estar presentes no 8
cromossomo bacteriano, ou ainda nos dois locais (GONZÁLES; BROWN; CARLTON, 9
1982; SANCHIS et al., 1998). 10
Uma ou várias cópias de um mesmo gene ou de diferentes genes cry 11
podem ser encontradas em uma mesma linhagem bacteriana, sendo que os produtos 12
poderão formar somente um ou diferentes cristais. A linhagem B. thuringiensis kurstaki 13
HD73 contém apenas o gene cry1Ac localizado num plasmídeo de 50 MDa 14
(LERECLUS et al., 1993). A linhagem aizawai 7.29, ao contrário, contem cinco genes 15
cry, quatro localizados no cromossomo e um num plasmídeo de 45 MDa (SANCHIS et 16
al., 1998), enquanto que a linhagem israelenses que também apresenta quatro genes 17
codificadores de proteínas Cry, apresenta todos os genes localizados num único 18
plasmídeo de 72 MDa (GONZÁLEZ; CARLTON, 1984; BOURGOUIN et al., 1988; 19
LERECLUS et al., 1989). A localização preferencial desses genes em plasmídeos 20
conjugativos e a frequente associação a elementos genéticos móveis promovem a 21
grande diversidade de linhagens pela combinação dos diferentes genes (KRONSTAD; 22
WHITELEY, 1984; LERECLUS et al., 1984). Essa combinação resulta na síntese de 23
cristais proteicos com diferentes tipos de proteínas Cry e com perfis de toxicidade 24
distintos (ARANTES; VILAS-BÔAS; VILAS-BÔAS, 2002; PIGOTT; ELLAR, 2007). 25
A maioria dos genes cry é expressa na fase de esporulação, portanto o 26
principal modo de regulação é através dos fatores sigma (σ) específicos dessa fase. 27
Outro modo de regulação é independente da esporulação (AGAISSE; LERECLUS, 28
1995), com a expressão desses genes ocorrendo durante a fase de crescimento 29
vegetativo (VALADARES-INGLIS et al., 1998). Tanto quando a expressão dos genes 30
cry ocorre na fase de esporulação, tanto quando a expressão ocorre durante crescimento 31
vegetativo, a proteína Cry sempre é produto do metabolismo secundário (ARANTES; 32
VILAS-BÔAS; VILAS-BÔAS, 2002). 33
A Figura 2 demonstra as diferentes etapas do modo de ação das 34

3
proteínas Cry. Inicialmente, os cristais ingeridos pelo inseto susceptível são 1
solubilizados liberando monômeros de proteínas Cry na forma de protoxinas, que 2
através de proteases presentes no intestino do inseto alvo, são clivadas e passam para 3
sua forma ativa. Estas, em seguida, atravessam a membrana peritrófica e ligam-se a 4
receptores específicos presentes na porção apical das microvilosidades das células 5
localizadas no intestino médio do inseto, conduzindo à formação de poros ou canais 6
iônicos e consequentemente a lise das células epiteliais, levando o inseto à morte. Além 7
disso, os esporos ingeridos juntamente com os cristais podem germinar e as células 8
vegetativas se multiplicarem causando septicemia no inseto hospedeiro (SCHNEPF et 9
al., 1998). 10
11
Figura 2. Modo de ação das proteínas Cry. (1) Solubilização e ativação da toxina; (2) ligação 12
dos monômeros da toxina no primeiro receptor induzindo a mudança na conformação da toxina 13
(3) formação do oligômero; (4) ligação do oligômero de toxina no segundo receptor; (5) 14
inserção do oligômero da toxina na membrana epitelial do intestino anterior, médio ou posterior 15
e formação do poro (Fonte: BRAVO; GILL; SOBERÓN, 2007). 16
17
O intestino dos insetos susceptíveis as toxinas de B. thuringiensis, 18
geralmente possuem pH elevado, o que impede a germinação dos esporos ingeridos do 19
patógeno. Porém com a ação das toxinas que destroem a parede do intestino, o pH é 20
reduzido permitindo a germinação dos esporos. O cadáver do inseto serve como fonte 21
de alimento para o crescimento vegetativo da bactéria (COPPING; MENN, 2000). 22
Os cristais proteicos são tóxicos, principalmente para insetos das 23
ordens Lepidoptera, Diptera, Coleoptera, Hymenoptera, Homoptera, Dictyoptera, 24
Orthoptera, Mallophaga, além de nematóides (Strongylida, Tylenchida), protozoários 25
(Diplomonadida) e ácaros (Acari) (FEITELSON; PAYNE; KIM, 1992; SCHNEPF et 26
al., 1998), sendo esses específicos, pois diferentes toxinas ligam-se a diferentes 27
receptores e com intensidade diferentes nas diversas espécies de inseto (COPPING; 28
MENN, 2000). Essas toxinas não apresentam qualquer dano aos mamíferos, aves, 29
anfíbios, répteis ou outros vertebrados, devido ao modo de ação das mesmas, o qual é 30
específico para as condições presentes no aparelho digestivo de insetos (SCHNEPF et 31
al., 1998). 32

4
A utilização de produtos a base de B. thuringiensis no controle de 1
insetos pragas da agricultura e vetores de doenças não é recente. Os primeiros produtos 2
começaram a ser comercializados em 1938. Em 1950, houve aumento na produção e na 3
comercialização dos produtos à base de B. thuringiensis, através da implementação de 4
novas tecnologias como o uso de fermentadores e com a descoberta de novas 5
subespécies de B. thuringiensis, o que ampliou o número de insetos-alvo controlados 6
BRAR et al., 2006; CAPALBO; VILAS-BÔAS; ARANTES, 2004). 7
A principal característica que diferencia os produtos à base de B. 8
thuringiensis de outros produtos comercialmente disponíveis é a atividade 9
entomopatogênica restrita, ou seja, a maioria dos produtos à base de B. thuringiensis 10
apresenta espectro de ação somente a um ou poucos insetos. Além disso, os produtos 11
são inócuos a outros animais, além de não contaminar o ambiente e não favorecer a 12
rápida seleção de insetos resistentes, o que acontece frequentemente quando se utiliza 13
produtos agroquímicos sintéticos (ARANTES; VILAS-BÔAS; VILAS-BÔAS, 2002). 14
Estas características, entre outras, permitem que os produtos à base de 15
B. thuringiensis possuam um custo menor tanto no desenvolvimento, quanto no registro. 16
Por exemplo, o custo de desenvolvimento de um bioinseticida à base de B. thuringiensis 17
subspécie israelensis, chega a ser 1/40 do custo de um novo pesticida químico sintético 18
(SCHNEPF et al., 1998). Além do mais, pode-se ressaltar que a utilização de 19
bioinseticidas à base de B. thuringiensis, permite integrar estratégias de manejo no 20
controle de insetos praga, preservando seus inimigos naturais. 21
22
1.1.2 Bacillus cereus 23
24
B. cereus é uma bactéria em forma de bastonete, Gram-positiva, 25
formadora de esporos, móvel pela presença de flagelos peritríquios, aeróbia, anaeróbia 26
facultativa, mesófila com temperatura de crescimento variando de 10-48 ºC, mas com 27
crescimento ótimo de 28-35 ºC (GRANUM; BAIRD-PARKER, 2000). Linhagens 28
psicotróficas podem crescer entre 4-5 ºC, sendo que o crescimento e a produção de 29
enterotoxinas foram observados em farelo de arroz após 24 dias a 4 ºC. (JENSON; 30
MOIR, 1997). B. cereus degrada glucose como fonte de carbono, e não degrada manitol, 31
xilose ou arabinose (ADAMS; MOSS, 1995), é resistente a ampicilina, apresenta 32
atividade hemolítica e suas colônias possuem aspecto irregular em meio de cultura 33
sólido (VILAS-BÔAS; PERUCA; ARANTES, 2007). É uma bactéria amplamente 34

5
distribuída no ambiente como solo e vegetação, sendo considerada como saprófita de 1
solo e encontrada também em alimentos, como os de origem vegetal, animal e produtos 2
lácteos (GRANUM; BAIRD-PARKER, 2000). 3
Em 1949, B. cereus foi reconhecido como patógeno de alimentos 4
(HAUGE, 1950), desde então inúmeras linhagens dessa bactéria, foram 5
reconhecidamente envolvidas em casos de intoxicação gastrointestinal, causados pela 6
ingestão de alimentos, produzindo diarreias e vômitos (KOTIRANTA et al., 2000). 7
Alimentos contaminados apresentam diferenças na textura e no sabor devido à 8
multiplicação das células vegetativas ou produção de toxinas (GRANUM; LUND, 9
1997). O processo de infecção alimentar pode começar poucas horas após a ingestão do 10
alimento contaminado, podendo causar vômito pela produção de toxinas eméticas 11
durante o crescimento de células vegetativas no alimento, e/ou diarreia pela produção de 12
toxinas diarreicas durante o crescimento vegetativo no intestino do hospedeiro 13
(KRAMER; GILBERT, 1989; DROBNIEWSKI, 1993; GRANUM, 1994; AGATA et 14
al., 1995; GRANUM; LUND, 1997; WIJNANDS; DUFRENNE; VAN LEUSDEN, 15
2002, ARNESEN et al., 2008, SENESI; GHELARDI, 2010). 16
Essa natureza oportunista de B. cereus como patógeno é relacionada 17
com sua capacidade de produzir inúmeros fatores de virulência não específicos, 18
incluindo fosfolipases, hemolisinas e enterotoxinas (DROBNIEWSKI, 1993, VILAS-19
BÔAS et al., 2012). A síndrome diarreica é causada por enterotoxinas, onde destacam-20
se a CytK (citotoxina-K) (LUND et al., 2000) e os complexos HBL (hemolisina BL) 21
(BEECHER; SCHOENI; WONG, 1995) e NHE (enterotoxina não hemolítica) 22
(GRANUM et al., 1994). A cereulide é a principal toxina responsável pela síndrome 23
emética, sendo resistente ao pH e ao calor (GRANUM; LUND, 1997). 24
Segundo Heyndrickx e Scheldeman (2002), diversas são as razões que 25
tornam o B. cereus um importante agente de contaminação na indústria de alimentos. I) 26
Os esporos estão em todos os lugares, o que torna praticamente impossível impedir sua 27
presença em alimentos crus e em ingredientes. Também pela ampla ocorrência na 28
natureza, contaminações após tratamentos de calor podem ocorrer facilmente quando 29
deficiências ocorrem na maquinaria ou nos materiais utilizados para embalagem. II) 30
Tratamentos de calor como a pasteurização (um mínimo de 71,7 °C durante 15 31
segundos) são eficientes para eliminar células vegetativas, mas não esporos. Esporos 32
sobreviventes em produtos tratados com calor chegam a cerca de 95% (MEER et al., 33
1991), podem germinar tendo pouca ou nenhuma competição com células vegetativas 34

6
de bactérias Gram-negativas. III) Esporos de diversas espécies têm características 1
adesivas, o que facilita sua fixação às superfícies de encanamentos e equipamentos de 2
processamentos com a posterior formação de biofilmes. IV) A tolerância que esporos e 3
células vegetativas podem apresentar às condições ou tratamentos que geralmente são 4
considerados seguros (baixas temperaturas e baixo pH). 5
Considerável ênfase tem sido dada à capacidade de linhagens de B. 6
cereus causarem contaminação de alimentos, por isso, a maior parte das publicações 7
envolvendo esta bactéria está relacionada a métodos de detecção rápida dessa espécie 8
em alimentos, assim como a identificação de genes que codifiquem para toxinas. No 9
entanto, esta bactéria tem também importância como espécie modelo do grupo B. 10
cereus, sendo utilizada em estudos taxonômicos que envolvem além do B. cereus, 11
outras espécies de grande importância, como o B. thuringiensis. 12
13
1.1.3 Taxonomia do Grupo do B. cereus 14
15
Atualmente existem 259 espécies descritas no gênero Bacillus (COHN, 16
1872; EUZÉBY, 2012). Porém, a dificuldade de estabelecer um consenso sobre a 17
definição da unidade fundamental da diversidade biológica, a espécie, tem gerado 18
inconvenientes acerca da classificação de micro-organismos. A definição do taxa 19
espécie abrange organismos posicionados numa escala evolutiva maior, sendo difícil 20
aplicar esse conceito em populações bacterianas. 21
A maior dificuldade da classificação bacteriana é devido a 22
transferência horizontal de genes, que mesmo quando ocorre em baixa frequência na 23
natureza (COHAN, 1994), permite a troca de material genético entre espécies 24
extremamente distantes (LINZ et al., 2000; SMITH; DOWSON; SPRATT, 1991). 25
Estima-se que cerca de 5 a 15% dos genes de um genoma bacteriano típico tenham sido 26
adquiridos de outras espécies através da transferência de segmentos homólogos ou 27
heterólogos, incluindo elementos transponíveis nos plasmídeos ou fagos (MAJEWSKI; 28
COHAN, 1999; OCHMAN; LAWRENCE; GROISMAN, 2000). 29
Anteriormente à disponibilidade dos testes moleculares, a classificação 30
bacteriana era realizada através de características fenotípicas e testes bioquímicos, o que 31
possibilitou a classificação dos organismos, considerando os graus de similaridade entre 32
os mesmos. No início dos anos 1970, os bacteriologistas começaram a utilizar dados 33
moleculares para a classificação taxonômica, o que tornou a sistemática bacteriana mais 34

7
acessível, incluindo bactérias não cultiváveis (COHAN, 2002). Dentre essas técnicas 1
inclui-se a hibridização DNA-DNA, na qual Johnson (1973) determinou que espécies 2
diferentes deveriam compartilhar um nível de similaridade inferior a 70% no genoma 3
completo. Outra técnica inovadora para a classificação foi a variação nucleotídica nos 4
genes de rRNA 16S, onde Stackebrandt e Goebel (1994) encontraram que linhagens 5
com mais de 3% de divergência deveriam ser classificadas como espécies diferentes, e 6
linhagens com menos de 3% de divergência nos genes de rRNA 16S poderiam ou não 7
ser consideradas como membros de uma mesma espécie. Como no caso de B. cereus e 8
B. thuringiensis que apresentam apenas 1% de divergência, e ainda são classificadas 9
como espécies diferentes. 10
O termo Grupo do B. cereus é um termo não taxonômico e 11
compreende, além de B. cereus sensu stricto, B. anthracis, B. weihenstephanensis, B. 12
mycoides, B. thuringiensis e B. pseudomycoides. As três primeiras espécies possuem 13
impactos negativos na atividade humana, sendo B. cereus e B. anthracis consideradas 14
patógenos de mamíferos, incluindo o homem. Da mesma forma, linhagens de B. 15
weihenstephanensis se multiplicam em baixas temperaturas, sendo um risco para 16
conservação de alimentos congelados (MAYR et al., 1999). Linhagens de B. mycoides 17
podem melhorar o crescimento de plantas (PETERSEN et al., 1995) enquanto que 18
linhagens de B. thuringiensis são utilizadas como inseticida biológico em várias partes 19
do mundo (SCHNEPF et al.,1998). A última espécie, B. pseudomycoides, tem sido 20
descrita como responsável por infecções em humanos como endoftalmites 21
(NAKAMURA et al., 1998). No entanto, apesar de não ser taxonômico, o agrupamento 22
destas espécies é legítimo e baseado na similaridade entre as sequências do gene do 23
rRNA 16S destas espécies. 24
Atualmente, segundo a sequência do gene 16S, o gênero Bacillus pode 25
ser dividido em seis grupos, denominado Bacillus RNA grupo 1 ao 6, sendo que as 26
espécies do grupo do B. cereus ficam agrupadas na subdivisão 3 do grupo 1 27
(STACKEBRANDT; SWIDERSKI, 2002). No entanto, a extensa similaridade entre as 28
sequências do gene de rRNA 16S apresentada por linhagens destas espécies é uma das 29
principais razões que justificam as discussões taxonômicas envolvendo este grupo de 30
bactérias (VILAS-BÔAS et al., 2007). 31
As células vegetativas e os esporos das três espécies, vistos ao 32
microscópio óptico, são extremamente semelhantes, entretanto, foi devido às diferenças 33
fenotípicas que B. thuringiensis, B. anthracis e B. cereus, foram inicialmente 34
8
classificados como pertencentes a espécies distintas. As células de B. thuringiensis 1
(Figura 3A), formam um cristal proteico constituído pelas proteínas Cry, que são 2
liberados junto com os esporos no momento da lise celular. As células de B. anthracis 3
apresentam a formação de cápsula e produzem toxinas letais, enquanto que as células de 4
B. cereus (Figura 3B), não apresentam nenhum destes fenótipos. 5
6
7
Figura 3. Colônias e fotomicrografias ópticas de (A) B. thuringiensis e (B) B. cereus. (1) células 8
vegetativas; (2) término da fase estacionária; (3) esporulação completa, com presença de 9
esporos e cristais em B. thuringiensis e somente esporos em B. cereus. Fotos 1, 2 e 3: Aumento 10
de 1000 vezes e coloração: Amido-Black e Fucsina. 11
12
Contudo, os genes responsáveis pelas características fenotípicas de B. 13
thuringiensis e B. anthracis estão presentes em grandes plasmídeos que, através de 14
mecanismos de transferência horizontal de genes (conjugação, transdução e 15
transformação) e rearranjos genéticos promovidos por elementos móveis de DNA, 16
podem ser transferidos para outras linhagens do grupo B. cereus (VILAS-BÔAS et al., 17
2007). Como os genes codificantes para essas características são presentes em 18
plasmídeos, a distinção entre essas espécies vem sendo debatida por muitos 19
pesquisadores (SCHNEPF et al., 1998; GLARE; O´CALLAGHAM, 2000; HANSEN; 20
SALAMITOU, 2000). 21
Gohar e colaboradores (2005), fizeram um estudo comparativo do 22
proteoma extracelular de linhagens de B. cereus, B. thuringiensis e B. anthracis que 23
tiveram os grandes plasmídeos curados. Os resultados mostraram que o proteoma das 24
três espécies foram similares e as únicas diferenças encontradas estavam relacionadas 25
com o peptídeo PlcR, um regulador pleiotrópico descrito por Agaisse e colaboradores 26
(1999), que em B. cereus e B. thuringiensis controla a expressão de mais de 100 genes 27
potencialmente envolvidos na virulência. Em B. anthracis o gene plcR é interrompido e 28
portanto não funcional, o que altera a expressão dos genes por ele regulados. Isso 29
demonstra que, embora destituídas dos elementos extracromossômicos, linhagens de B. 30

9
anthracis podem ser diferenciadas de B. cereus e B. thuringiensis por mudanças na 1
expressão gênica, como posteriormente relatado por Mignot e colaboradores (2001). 2
O polimorfismo genético apresentado pelas linhagens de B. cereus e B. 3
thuringiensis e a extensiva similaridade de seus conjuntos gênicos são os principais 4
fatores que complicam a resolução da questão taxonômica entre essas duas espécies. 5
Linhagens acristalíferas de B. thuringiensis são indistinguíveis de B. cereus 6
(LYSENKO, 1983), vários estudos iniciais analisando diferentes características, como 7
bioquímicas e análises de DNA (KANEKO; NOZAKI; AIZAWA, 1978; BAUMANN, 8
1984; LOGAN; BERELEY, 1984; PRIEST, 1988) indicam que essas espécies devam 9
ser consideradas apenas uma. Um dos primeiros estudos para a investigação das 10
relações taxonômicas entre as duas espécies foi conduzido por Carlson e colaboradores 11
(1994) usando eletroforese em campo pulsado (PFGE) e eletroforese de aloenzimas 12
(MLEE). Um alto grau de variabilidade genética foi observado dentro e entre as duas 13
espécies, não sendo possível agrupar separadamente cada espécie, tendo sido sugerido o 14
agrupamento de B. cereus e B. thuringiensis como única espécie. 15
Outros estudos usando MLEE e a análise de sequências de genes 16
cromossômicos apresentaram alto grau de similaridade genética e indicaram a falta de 17
diferenciação entre B. thuringiensis e B. cereus sensu stricto e, assim, os autores 18
sugeriram que essas espécies devem ser consideradas como membros de uma espécie 19
designada B. cereus sensu lato (CARLSON, CAUGANT; KOLSTØ, 1994; 20
HELGASON et al., 1998, 2000a, 2000b; DAFFONCHIO et al., 2000; BAVYKIN et al,. 21
2004). Ticknor e colaboradores (2001), baseados em resultados de polimorfismo de 22
comprimento de fragmentos amplificados (AFLP), MLEE e dados de sequências de 23
DNAr 16S mostraram que B. cereus e B. thuringiensis são altamente polimórficos e que 24
uma simples análise de um número limitado de linhagens não é suficiente para 25
caracterizar essas espécies. 26
Outros pesquisadores apoiam a separação das duas espécies. Vilas-27
Boas et al. (2002) analisaram pela técnica de MLEE e atividade hemolítica, populações 28
simpátricas coletadas de diferentes amostras de solo. Encontraram que populações de 29
uma mesma espécie de Bacillus (B. thuringiensis ou B. cereus) foram mais similares 30
geneticamente entre si do que com as populações da outra espécie de Bacillus. Além 31
disso, foi evidenciada uma maior taxa de fluxo gênico entre as linhagens da mesma 32
espécie. Porém, mesmo em baixas frequências, ocorreram trocas genéticas 33
interespecíficas (entre B. thuringiensis e B. cereus). Cherif e colaboradores (2003) 34

10
utilizaram rep-RCR para analisar agrupamentos de linhagens pertencentes ao grupo do 1
B. cereus. Os resultados mostraram que linhagens de B. thuringiensis exibiram um 2
agrupamento bem diferente das outras linhagens de B. cereus, sugerindo que estas 3
espécies podem ser consideradas como unidades taxonômicas separadas. 4
As análises, utilizando outras ferramentas genéticas, trazem resultados 5
controversos quanto à taxonomia de B. cereus e B. thuringiensis, com trabalhos que ora 6
distinguem as três espécies de Bacillus como unidades taxonômicas distintas, ora como 7
linhagens de uma mesma espécie. Desta forma, tem se mostrado relevante, acrescentar 8
aos estudos genéticos, estudos relacionados à ecologia das linhagens do grupo B. 9
cereus. A soma destas duas áreas pode, contribuir para a definição da taxonomia destas 10
espécies. 11
12
1.2 Conjugação Bacteriana 13
14
O processo de conjugação foi descoberto por Lederberg e Tatum em 15
1946, porém foi descrito em detalhes por Lederberg em 1986 (LEDERBERG, 1986). 16
Esse processo foi definido como a transferência do material genético de uma linhagem 17
bacteriana para outra, sendo dependente de contato celular entre os membros de duas 18
linhagens bacterianas (DAVIS, 1950). Essa descoberta foi considerada o mais 19
importante avanço na história da bacteriologia até aquele momento. Atualmente 20
sabemos que o sistema de conjugação envolve a transferência do DNA entre uma ampla 21
gama de gêneros de bactérias, e em alguns casos, de bactérias para fungos e células de 22
plantas (BATTISTI; GREEN; THORNE, 1985; OULTRAM; YOUNG, 1985; 23
KOEHLER; THORNE, 1987; BEIJERSBERGEN et al., 1992; HAYMAN; BOLEN, 24
1993; MAHMOOD et al., 1996; BERTOLLA; SIMONET, 1999; HEINEMANN, 1999; 25
CHRISTIE; VOGEL, 2000; CHUMAKOV, 2000; CHEN, 2002; BROOTHAERTS et 26
al., 2005). 27
A partir do contato entre as células doadoras e receptoras são 28
formados os exconjugantes ou transconjugantes, que são capazes de realizar outros 29
processos de conjugação. Assim, a conjugação é um importante componente da 30
evolução bacteriana. 31
O sistema de conjugação de bactérias Gram-positivas difere do 32
sistema de Gram-negativas por não formar o pili, estrutura que proporciona o contato 33
entre células em bactérias Gram-negativas. Em bactérias Gram-positivas são formados 34

11
agregados de células em culturas com duas linhagens (Figura 4), os quais não são 1
formados em monoculturas (ANDRUP et al., 1996). 2
3
Figura 4. Eletromicrografia de varredura do processo de conjugação em bactérias Gram-4
positivas, compreendendo células doadoras (linhagem AND801) e células receptoras (linhagem 5
GBJ001) (Fonte: ANDRUP et al., 1996). 6
7
A questão central do processo de conjugação é como o DNA é 8
transferido entre as células envolvidas nesse contato. A conjugação é um conjunto de 9
eventos, que envolvem desde o processamento do DNA até a transferência propriamente 10
dita. O primeiro evento é gerar uma cópia de fita simples do DNA (ssDNA), para isso 11
uma relaxase se liga a origem de transferência do plasmídeo (oriT) e cliva a fita de 12
DNA que será destinada a transferência, essa fita é denominada fita-T. A relaxase 13
juntamente com uma ou mais proteína(s) auxiliar(es) formam um complexo 14
multiproteico denominado relaxossomo, o qual permanece ligado a extremidade 5' da 15
fita-T, resultando na formação do complexo relaxase-fita-T (GROHMANN; MUTH; 16
ESPINOSA, 2003; CHEN; CHRISTIE; DUBNAU, 2005; GARCILLÁN-BARCIA; 17
FRANCIA; DE LA CRUZ, 2009). 18
O próximo passo é o transporte intercelular, que é realizado pelo 19
sistema de secreção tipo IV (T4SS), também conhecido como mating pair formation 20
(MPF). O sistema de secreção tipo IV, ocorre dentro de uma ampla gama de 21
organismos, incluindo bactérias Gram-positivas, Gram-negativas e algumas archaea 22
(ZECHNER et al., 2012). O grupo do B. cereus sensu lato utiliza esse sistema para 23
efetuar a conjugação (GROHMANN; MUTH; ESPINOSA, 2003). 24

12
O aparato de T4SS de conjugação é um complexo multiproteico que é 1
codificado por genes plasmidiais, envolvidos no transporte do DNA (GROHMANN; 2
MUTH; ESPINOSA, 2003). A relaxase deve permanecer ligada a T-strand para 3
sinalizar o reconhecimento do substrato. Dessa maneira o sistema de conjugação é 4
considerado um sistema de transporte de proteína, que tem evoluído a capacidade de 5
translocar as relaxases e consequentemente moléculas de DNA à ela ligadas (CHEN; 6
CHRISTIE; DUBNAU, 2005). 7
O DNA é bombeado ativamente para dentro da célula receptora pela 8
proteína de acoplamento do tipo IV (T4CP) (LLOSA et al., 2002; CHRISTIE, 2004). 9
T4CP é uma proteína integral de membrana homohexamérica pertencente à família de 10
ATPases (FIRTH; IPPEN-IHLER; SKURRAY, 1996; MONCALIÁN et al., 1999; 11
TATO et al., 2005). Essa proteína é responsável por conectar o processamento e 12
translocação do DNA entre as células, sendo que a hidrólise de ATP é a energia 13
necessária para bombear a molécula de DNA através do canal central da mesma 14
(GOMIS-RÜTH, 2006). 15
16
1.2.1 Características dos Plasmídeos e do Processo de Conjugação 17
18
Os plasmídeos podem ser classificados de acordo com os genes que 19
possuem, sendo esses relacionados com o aparato conjugativo, podendo ser 20
classificados pela habilidade de mobilização. Plasmídeos conjugativos, também 21
denominados auto-transmissíveis, codificam a maquinaria proteica completa para a 22
transferência conjugal. Esses plasmídeos carregam os genes tra e codificam seu próprio 23
conjunto de proteínas do complexo MPF ou T4SS que permite a montagem e 24
funcionamento do canal na membrana para a conjugação. Geralmente são grandes, com 25
pelo menos 30 kb, porém com baixo número de cópias. Plasmídeos mobilizáveis 26
contem o mínimo conjunto de genes que permitem a transferência conjugativa, porém 27
somente quando existem funções conjugativas adicionais. Esses geralmente contém 28
somente a oriT, o gene da relaxase e uma ou mais proteínas acessórias e não possuem as 29
funções requeridas para a formação do poro de conjugação. Possuem até 15 kb, sendo 30
considerados plasmídeos pequenos, porém em um alto número de cópias 31
(GARCILLÁN-BARCIA; FRANCIA; DE LA CRUZ, 2009). 32

13
Um terceiro tipo, denominado plasmídeo não mobilizável, não possui 1
a região MOB nem o sítio oriT, porém consegue se tornar mobilizável pela 2
recombinação com plasmídeos conjugativos (ANDRUP et al., 1996). 3
A transferência do DNA não é unidirecional, podendo ocorrer o 4
processo de retromobilização, descrito por Ankenbauer (1997). Nesse processo o DNA 5
da célula receptora é capturado pela célula doadora que abriga o plasmídeo conjugativo. 6
A transferência de plasmídeos mobilizáveis da célula receptora para 7
célula doadora (retromobilização) pode ser explicada de duas maneiras. Existe o modelo 8
de transferência bidirecional, também conhecido como modelo de um passo, no qual 9
ocorre um evento simples de conjugação, nesse processo o DNA flui livremente e 10
simultaneamente entre as linhagens doadora e receptora (Figura 5A). O outro modelo é 11
unidirecional, conhecido por modelo de dois passos, onde duas etapas de transferência 12
unidirecional ocorrem. A primeira transfere o plasmídeo conjugativo para célula 13
receptora e após a célula doadora transfere o plasmídeo mobilizável para a célula 14
doadora (Figura 5B) (TOP et al., 1992). Estudos em bactérias Gram-negativas sugerem 15
esse último modelo (HEINEMANN; ANKENBAUER 1993; HEINEMANN, SCOTT; 16
WILLIAMS,1996; SIA; KUEHNER; FIGURSKI, 1996) . 17
18
Figura 5. Representação esquemática dos modelos de retromobilização. (A) Modelo de 19
transferência bidirecional: transferência do plasmídeo conjugativo para a célula receptora ocorre 20
simultaneamente com a transferência do plasmídeo mobilizável para a doadora por apenas um 21
evento de conjugação. (B) Modelo de transferência unidirecional: o primeiro evento conjugativo 22
permite a transferência do plasmídeo conjugativo para a receptora, enquanto que um segundo 23
evento permite a transferência do plasmídeo mobilizável para a doadora. Elipse grande: 24
plasmídeo conjugativo; Círculo pequeno: plasmídeo mobilizável; linha fina: célula receptora; 25
linha grossa: célula doadora; setas brancas: indicação da transferência dos plasmídeos e setas 26
pretas: resultado da transferência (Fonte: TIMMERY et al., 2009). 27
28
Os sistemas de conjugação podem variar sendo classificados de 29
acordo com a utilização de duas ou três linhagens, conjugação biparental e triparental, 30
respectivamente. A conjugação biparental (Figura 6A e C) envolve a transferência de 31
plasmídeos conjugativos da célula doadora para a célula receptora, onde pode ocorrer a 32

14
mobilização e a retromobilização de plasmídeos. Nesse processo vários eventos podem 1
ocorrer, a transferência de plasmídeos conjugativos da doadora para receptora; 2
mobilização de plasmídeos mobilizáveis da doadora para receptora; a transferência de 3
ambos os plasmídeos para a receptora (Figura 6B) e a retromobilização de plasmídeos 4
mobilizáveis da receptora para doadora. 5
O sistema de conjugação triparental transfere plasmídeos conjugativos 6
da doadora para a receptora, com isso a retromobilização de plasmídeos da receptora 7
para a doadora pode ocorrer. Nesse evento de conjugação pode ocorrer a transferência 8
de plasmídeos conjugativos, a mobilização e retromobilização de plasmídeos. 9
Recentemente foram descritos dois sistemas diferentes, um utilizando 10
uma linhagem doadora, uma receptora e uma auxiliar, esta pode transferir plasmídeos 11
mobilizáveis e não-mobilizáveis. Nesse sistema três eventos podem ocorrer, a 12
mobilização do plasmídeo da linhagem auxiliar para a receptora; transferência de ambos 13
os plasmídeos para a linhagem receptora e retromobilização do plasmídeo da linhagem 14
auxiliar para a linhagem doadora (Figura 6D) (TIMMERY et al., 2009). O segundo, 15
descrito por Santos et al. (2010) analisa se existe transferência preferencial do 16
plasmídeo conjugativo, utilizando três linhagens; duas receptoras e uma doadora, esse 17
sistema é denominado conjugação com duas receptoras. 18
19

15
Figura 6. Representação esquemática de conjugação biparental e triparental. Conjugação refere-1
se à transferência de plasmídeos conjugativos, enquanto a mobilização e retromobilização 2
refere-se à transferência de plasmídeos mobilizáveis. (A) Conjugação biparental: conjugação 3
para a receptora (1); (B) Mobilização biparental: conjugação para a receptora (1), mobilização 4
para a receptora (2), e transferência de ambos os plasmídeos para a receptora (3). (C) 5
Retromobilização biparental: conjugação para a receptora (1) e retromobilização para a doadora 6
(2). (D) Encontro triparental: conjugação para a receptora (1) e para a linhagem auxiliar (2), 7
mobilização para a receptora (3), transferência de ambos os plasmídeos para a receptora (4), e 8
retromobilização para a doadora (5). Elipse grande: plasmídeo conjugativo; Círculo pequeno: 9
plasmídeo mobilizável; linha fina: célula receptora; linha grossa: célula doadora; setas brancas: 10
indicação da transferência dos plasmídeos; Seta branca em formato de “T”: transferência de 11
ambos os tipos de plasmídeos e setas pretas: resultado da transferência (Fonte: TIMMERY et 12
al., 2009). 13
14
A transferência de plasmídeos, tanto por conjugação, mobilização ou 15
até mesmo por transformação demonstra a importância da transferência gênica na 16
evolução dos genomas bacterianos, como no caso estudado do grupo do B. cereus sensu 17
lato. 18
1.2.2 Conjugação em B. thuringiensis 19
20
Genes cry são contidos em grandes plasmídeos (GONZÁLEZ; 21
DULMAGE; CARLTON, 1981; KRONSTAD; SCHNEPF; WHITELEY, 1983; 22
GONZÁLEZ; CARLTON, 1984), por isso que o interesse de pesquisadores tem sido 23
voltado principalmente a eles (GONZÁLEZ; CARLTON, 1980; BATTISTI; GREEN; 24
THORNE, 1985; REDDY et al., 1987; JENSEN et al., 1996; WILCKS et al., 1998). 25
Porém, linhagens de B. thuringiensis geralmente exibem um conjunto complexo de 26
plasmídeos, podendo chegar a 17 plasmídeos com tamanhos entre 2 – 600 kb 27
(LERECLUS et al.. 1982; McDOWELL; MANN, 1991; BERRY et al., 2002; HAN et 28
al., 2006; KASHYAP; AMLA, 2007; AMADIO et al., 2009; ZHONG et al., 2011). 29
González e Carlton (1984) demonstraram a primeira ocorrência de 30
conjugação entre linhagens de B. thuringiensis, pela qual confirmaram a localização dos 31
genes cry em plasmídeos. Outros experimentos demonstraram a possível ocorrência de 32
conjugação entre linhagens de B. thuringiensis (Cry+, doadora) com linhagens de B. 33
cereus (Cry-, receptora), sendo que as linhagens exconjugantes Cry
- passaram a produzir 34
a proteína cristal (GONZÁLEZ; BROWN; CARLTON, 1982). 35
Anteriormente plasmídeos conjugativos não apresentavam marcas 36
seletivas, assim evidência de transferência era realizada por plasmídeos não-37
conjugativos que continham genes para a resistência a antibióticos. O plasmídeo pBC16 38
foi o mais utilizado, esse plasmídeo foi isolado de B. cereus, e carrega genes que 39

16
codificam resistência para antibióticos. O uso desse plasmídeo permitiu caracterizar 1
plasmídeos como pXO11, pXO12, pXO13, pXO14, pXO15 e pXO16, isolados de 2
diferentes subspécies de B. thuringiensis, capazes de promover a transferência de 3
pequenos plasmídeos mobilizáveis dentro da variedade de receptoras incluindo 4
linhagens B. thuringiensis, B. cereus e B. anthracis (BATTISTI; GREEN; THORNE 5
1985; REDDY et al., 1987). Andrup e colaboradores (1993) conduziram estudos para 6
monitorar a transferência entre linhagens de B. thuringiensis subsp. israelenses, 7
utilizando a resistência a antibióticos codificada por genes contidos em pequenos 8
plasmídeos mobilizáveis (pCB16 e pAND006). 9
Durante a conjugação notou-se a presença de agregados entre as 10
linhagens doadora e receptora, durante o crescimento exponencial. Isso permitiu a 11
diferenciação de dois fenótipos, Agr+ que formam agregados quando combinadas com 12
as linhagens Agr- (ANDRUP, et al., 1996). 13
A base genética dos sistemas de agregados de B. thuringiensis subsp. 14
israelensis, foi localizada no plasmídeo pXO16 (JENSEN et al., 1995). Esse plasmídeo 15
foi mobilizado tanto por replicação em círculo rolante quanto por plasmídeos baseados 16
na origem de replicação teta (ANDRUP et al., 1996). 17
O plasmídeo conjugativo pHT73 de B. thuringiensis subsp. kurstaki 18
KT0, foi o primeiro plasmídeo conjugativo marcado com um gene que confere 19
resistência a antibiótico (eritromicina); permitindo monitorar a frequência de 20
conjugação sem necessitar de plasmídeos mobilizáveis (VILAS-BÔAS et al., 1998). 21
Os estudos anteriormente citados demonstraram apenas a transferência 22
unilateral utilizando linhagens de B. thuringiensis. Entretanto Timmery et al. (2009) 23
demonstrou a capacidade de retromobilização através do plasmídeo conjugativo pXO16 24
de B. thuringiensis subsp. israelenses. Neste estudo foram comparados a habilidade de 25
conjugação e mobilização desses plasmídeos, usando plasmídeos mobilizáveis e 26
elementos não-mobilizáveis que perderam o gene mob e o sítio oriT. 27
Experimentos de cinética mostraram que a retromobilização foi 28
atrasada quando comparada com a conjugação de pXO16, sugerindo que a 29
retromobilização de pXO16 ocorreu por transferências sucessivas, como no segundo 30
modelo que é sugerido para bactérias Gram-negativas. 31
O uso de variáveis e métodos alternativos para transferência de DNA 32
entre células permitiram a detecção da conjugação, mobilização e retromobilização. 33
Diversos trabalhos in vitro usando meio líquido, meio sólido, amostras de solo, água e 34

17
alimentos, combinados com ensaios in vivo em larvas de inseto, e com o 1
sequenciamento de grandes plasmídeos, gerou um aumento no interesse do papel dos 2
plasmídeos na biologia de B. thuringiensis. Assim, a transferência gênica horizontal, 3
tanto dentro quanto entre espécies, tem sido frequentemente utilizada na investigação da 4
posição taxonômica de B. thuringiensis e de outras espécies do grupo do B. cereus. 5
6
1.3 Ecologia de B. thuringiensis 7
8
Estudos de abundância, diversidade e distribuição de isolados do 9
grupo do B. cereus sensu lato buscam compreender questões ligadas à evolução e as 10
relações ecológicas desse grupo, como exemplo o comportamento dessas bactérias no 11
solo, água e em insetos. Uma questão essencial é a classificação dessa bactéria em taxa 12
diferentes ou como uma única espécie, sendo que B. thuringiensis representaria uma 13
subespécie de B. cereus (VILAS-BÔAS; SANTOS, 2012). 14
As características fenotípicas específicas dessas espécies estão 15
presentes em megaplasmídeos conjugativos. A transferência horizontal desses 16
plasmídeos foi descrita para uma variedade de linhagens receptoras que pertencem ao 17
grupo do B. cereus. Isso levou a um aumento nos trabalhos in vitro e in vivo, 18
conduzidos em condições controladas em amostras de solo, insetos, água, alimentos que 19
permitem o acesso ao comportamento dessas bactérias envolvendo germinação de 20
esporos, formação de esporos e persistência, bem como a multiplicação de células 21
vegetativas e conjugação (VILAS-BÔAS; SANTOS, 2012). 22
O ciclo infeccioso de B. thuringiensis tem sido estudado há anos; a 23
primeira evidência de germinação dos esporos, esporulação e produção de toxinas de B. 24
thuringiensis foi realizada por Aly, Mulla e Federici (1985) em larvas mortas de 25
mosquitos. Jarrett e Stephenson (1990) infectaram larvas com linhagens de B. 26
thuringiensis e verificaram que larvas de insetos mortos seriam um meio ideal para a 27
troca de material genético, obtendo frequência de 100% na conjugação. Com isso foi 28
evidenciada a ocorrência de troca de material genético na natureza e novas questões 29
surgiram sobre o nicho ecológico de B. thuringiensis e o impacto da transferência 30
horizontal de genes no ambiente 31
A avaliação da conjugação em meio de cultura, microcosmos de solo e 32
larvas de lepidópteros infectados, Vilas-Bôas et al. (1998) verificaram através do 33
plasmídeo pTH73, que a larva de inseto é o ambiente mais favorável para a ocorrência 34

18
de conjugação, com frequência de 10-1
exconjugantes/receptora. Furlaneto et al. (2000), 1
avaliaram a sobrevivência e a transferência conjugal entre linhagens de B. thuringiensis 2
em ambiente aquático. A baixa frequência de conjugação detectada permitiu inferir que 3
esse ambiente não é propício para a multiplicação das células vegetativas, permitindo a 4
rápida formação dos esporos. 5
Suzuki e colaboradores, (2004), avaliaram a especificidade das toxinas 6
e a especificidade de hospedeiros. A primeira se refere a habilidade da toxina produzida 7
por linhagens específicas de B. thuringiensis atingir insetos específicos e a segunda se 8
refere a habilidade de linhagens específicas de B. thuringiensis colonizarem espécies 9
específicas de larvas de insetos. Resultados mostraram que linhagens de B. thuringiensis 10
sem especificidade para matar o inseto podem se multiplicar e transferir material 11
genético no inseto morto mecanicamente, indicando que não há correlação entre a 12
especificidade da toxina e a especificidade do hospedeiro. Portanto, linhagens de B. 13
thuringiensis podem colonizar insetos não susceptíveis desde que os insetos sejam 14
mortos mecanicamente. Resultados similares foram descritos por Thomas et al. (2001; 15
2002) em larvas de insetos não susceptíveis, onde baixas taxas de transferência conjugal 16
foram detectadas e a multiplicação de células vegetativas também foi prejudicada nessa 17
condição. 18
Guidi e colaboradores (2011) avaliaram a distribuição espacial de 19
esporos de B. thuringiensis subsp. israelensis em um ambiente úmido, onde ocorreu 20
aplicação de produtos a base de B. thuringiensis desde 1988. Foi verificado o 21
decréscimo dos esporos perto de 96%, mostrando que o solo natural geralmente não 22
suporta a germinação de esporos ou mesmo a multiplicação de células vegetativas. A 23
ocorrência do processo de conjugação em amostras de solo é também limitada por 24
condições especiais, como de suplementação com nutrientes, valores neutros de pH e 25
esterilização, para a ocorrência de germinação de esporos e a multiplicação de células 26
vegetativas. 27
As diversas condições ambientais utilizadas para avaliar a ocorrência 28
de transferência conjugal envolvendo linhagens de B. thuringiensis demonstraram que o 29
principal fator limitante é a capacidade de permitir a germinação dos esporos ou 30
simplesmente a presença de células viáveis, embora pareça que a multiplicação dessas 31
células não influenciam drasticamente esse evento (SANTOS et al., 2010). 32
Santos e colaboradores (2010) avaliaram, em B. thuringiensis, a 33
ocorrência de transferência horizontal triparental preferencial (uma doadora e duas 34

19
receptoras) e a germinação e multiplicação de linhagens de B. thuringiensis e B. cereus 1
em larvas mortas de Bombyx mori. A linhagem B. thuringiensis KT0 não mostrou 2
preferência para troca genética dentro de linhagens receptoras de B. thuringiensis e B. 3
cereus. Com isso o inseto hospedeiro pode ser considerado como uma fonte de 4
nutrientes, um meio de transmissão, e como um reservatório natural em que as bactérias 5
do grupo B. cereus podem ser submetidas à recombinação genética através de diferentes 6
mecanismos de adaptação. Estudos demonstram a grande habilidade de disseminação de 7
plasmídeos entre diferentes espécies do grupo do B. cereus sensu lato (JARRETT; 8
STEPHENSON, 1990; VILAS-BOAS et al., 1998; SANTOS et al., 2010). Devido ao 9
mecanismo de conjugação os plasmídeos podem ser considerados como uma entidade 10
biológica autônoma, capaz de se transferir de uma célula bacteriana pra outra, assim um 11
plasmídeo como o pHT73 de B. thuringiensis não poderia ser considerado específico 12
para essa espécie bacteriana. Quando condições ambientais são desfavoráveis, a célula 13
bacteriana pode eliminar elementos extracromossômicos e com isso obter vantagem 14
seletiva. Dessa maneira são as trocas genéticas que fazem com que os plasmídeos sejam 15
mantidos nas populações bacterianas, com isso esses podem ser mantidos na população 16
através do mecanismo de conjugação. 17
Santos e colaboradores (2010) demonstraram que a conjugação entre 18
linhagens de B. thuringiensis e B. cereus ocorre de maneira randômica, assim não 19
ocorre preferência na transferência de material genético entre essas duas espécies 20
bacterianas. Entretanto, resultados de germinação e multiplicação, no mesmo trabalho, 21
mostraram que linhagens de B. thuringiensis germinaram e multiplicaram mais 22
eficientemente que linhagens de B. cereus nas larvas de inseto. Esses resultados 23
mostraram que não existe nenhuma associação positiva entre a eficiência da 24
multiplicação bacteriana com a capacidade de conjugação em insetos infectados nas 25
condições testadas, ou seja, a multiplicação de células vegetativas não é essencial para a 26
ocorrência de conjugação entre linhagens de B. thuringiensis ou entre B. thuringiensis e 27
B. cereus em insetos mortos. 28
Experimentos de conjugação também foram conduzidos em alimentos, 29
devido ao B. cereus ser uma espécie conhecida pelas infecções gastrointestinais. Van 30
der Auwera et al. (2007) avaliaram o comportamento da conjugação de linhagens de B. 31
thuringiensis em meio Luria-Bertani (LB), leite e pudim de arroz usando dois 32
plasmídeos conjugativos, pXO16 e pAW63, bem como o plasmídeos mobilizável 33
pC194, em encontros bi e triparentais. Altas frequências de conjugação foram 34

20
observadas no leite, quando comparados com o meio LB. Além disso, quando linhagens 1
de B. cereus foram utilizadas como doadoras do plasmídeo pXO16 (isolado de B. 2
thuringiensis) para outra linhagem de B. cereus, a razão de conjugação foi em torno de 3
10-1
exconjugante/receptora, sendo similares com os valores observados na conjugação 4
com linhagens de B. thuringiensis em larvas de insetos. 5
Muitas evidências indicaram que B. thuringiensis e B. cereus exibem 6
uma variação de comportamento em diferentes ambientes. Essas diferenças são 7
relatadas como a alta taxa de conjugação ambos dentro e entre essas espécies, 8
germinação dos esporos e multiplicação das células vegetativas. Entretanto muitos 9
aspectos da biologia dessas espécies permanecem desconhecidos. Wilcks e 10
colaboradores em 2008 demonstraram que linhagens de B. thuringiensis são capazes de 11
estabelecer o ciclo de vida completo no trato digestório de ratos, incluindo germinação 12
de esporos, multiplicação de células vegetativas e transferência do plasmídeo pXO16. 13
Bizzarri e Bishop (2008) obtiveram o isolamento de células vegetativas do filoplano, 14
indicando novamente a possibilidade do ciclo de vida de B. thuringiensis ocorrer em 15
outros ambientes e não somente em cadáveres de insetos. 16
A maioria dos ambientes onde B. thuringiensis é encontrado pode ser 17
considerado reservatório de esporos, mas ocasionalmente células vegetativas podem 18
ocorrer em qualquer um desses ambientes, uma vez que eles fornecem condições ideais 19
como nutrientes, pH, temperatura e umidade. Entretanto, o ambiente onde células de B. 20
thuringiensis foram encontradas em melhores condições para multiplicação foram 21
larvas de insetos mortas, que podem ser consideradas como nicho ecológico dessa 22
espécie bacteriana (SALAMITOU et al., 2000; TAKATSUKA; KUNIMI, 2000; 23
THOMAS et al., 2000, 2001, 2002; YUAN et al., 2007; SANTOS et al., 2010; 24
RAYMOND et al., 2012). Consequentemente, nesse ambiente provavelmente ocorre a 25
evolução do genoma de B. thuringiensis através de diferentes mecanismos como as 26
trocas genéticas. 27
Outra linha de pesquisa desenvolvida por Raymond e colaboradores 28
investigou a estrutura genética populacional e ecologia do grupo do B. cereus. Raymond 29
et al., (2010a) encontraram o que muitos trabalhos anteriores tentavam, verificar a 30
relação da comunidade bacteriana com os insetos. Acreditava-se que em solo de 31
florestas iria existir uma comunidade bacteriana mais diversa que em um campo aberto, 32
devido a diversidade de espécies de insetos. Raymond et al., (2010a) observaram que 33
em solos existe um genótipo bem adaptado de B. thuringiensis, encontrado como 34

21
predominante nas amostras de comunidades do grupo do B. cereus, esse genótipo 1
correspondia ao da espécie B. thuringiensis subsp.kurstaki HD-1, linhagem utilizada no 2
bioinseticida DiPel. 3
Análises mostraram que essa predominância não é devido a aplicações 4
contínuas de bioinseticida, pois ocorreu um decaimento drástico de esporos após 10 dias 5
a partir da aplicação, então concluiu-se que esse genótipo está bem adaptado ao 6
ambiente e possui os genes comuns codificadores das proteínas Cry, Cry1Ab e Cry1Ac. 7
Alguns estudos observaram resistência de alguns insetos a essas duas proteínas Cry 8
(WIRTH; GEORGHIOU; FEDERICI, 1997; FERRÉ; VAN RIE, 2002). Isso é devido, 9
portanto não a aplicação de bioinseticida, mas sim a adaptação desse genótipo 10
bacteriano no ambiente. 11
Raymond et al., (2010) em experimentos com microcosmos, em 12
filoplano, concluíram que tanto a pulverização de bioinseticida quanto a introdução de 13
insetos no ambiente, alteram a constituição da comunidade bacteriana. A aplicação do 14
bioinseticida é indetectável após 28 dias da aplicação, tendo um efeito transiente na 15
comunidade bacteriana. Já insetos hospedeiros aumentam a densidade bacteriana e a 16
proporção de linhagens que expressam as proteínas Cry, pois as bactérias proliferam 17
dentro dos hospedeiros. 18
Raymond e colaboradores (2012) evidenciaram a cooperação entre 19
bactérias do grupo do B. cereus sensu lato. Experimentos em microcosmos foram 20
realizados com linhagens com e sem o gene cry, sendo que as linhagens sem o gene 21
foram obtidas por cura plasmidial através da linhagem com o gene. No início da 22
infecção foi recuperada a mesma quantidade de bactérias sem e com o gene cry, porém 23
após a infecção a linhagem sem o gene cry se multiplicou melhor e ficou com maior 24
densidade bacteriana que a linhagem com o gene. Porém foi evidenciado que esse 25
evento é de cooperação entre as linhagens e não de competição, pois a multiplicação de 26
bactérias é benéfica para a espécie, onde as linhagens não produtoras da proteína Cry 27
exploram os benefícios gerados pelas produtoras (acesso aos nutrientes do hospedeiro). 28
O aumento na densidade de bactérias sem o gene cry pode ser explicado pela “carga” 29
que os plasmídeos, carregados pela linhagem com o gene, geram. A multiplicação da 30
bactéria com o plasmídeo é mais lenta, enquanto que as bactérias que não carregam o 31
plasmídeo, se multiplicam mais rapidamente. 32
A falta de epizootias por B. thuringiensis pode ser explicada por essa 33
multiplicação das linhagens sem o gene cry, pois para a ocorrência da transferência 34

22
direta, requerida para uma epidemia, bactérias produtoras de cristal devem existir em 1
alta quantidade, o que não ocorre nas infecções naturais, onde as não produtoras se 2
multiplicam melhor (RAYMOND, 2012). Outra evidência para a não ocorrência de 3
epizootias é que ao morrer pela ação das proteínas Cry o inseto é depositado no solo, 4
seu reservatório natural. Assim, para que ocorra a transferência para outros hospedeiros 5
que estejam nas folhas, esporos de B. thuringiensis devem germinar, e ocorrer um novo 6
ciclo para que as proteínas Cry sejam produzidas e infectem novamente os insetos 7
hospedeiros (RAYMOND, 2010b). 8
As características fenotípicas das espécies bacterianas do grupo do B. 9
cereus sensu lato estão contidas em plasmídeos conjugativos. Entretanto, plasmídeos 10
são muitas vezes vistos como apoio de elementos genéticos na evolução do genoma 11
destas bactérias e esta é uma forma injusta de se referir a um grupo genético tão 12
importante. Portanto, é preciso entender melhor esses plasmídeos, realizando o 13
sequenciamento e anotação, conhecer tanto o seu papel na bactéria e como as condições 14
favoráveis para sua dispersão, bem como as células vegetativas contendo diferentes 15
plasmídeos interagindo com diferentes ambientes. A natureza é dinâmica e as mudanças 16
ocorrem gradualmente no curso da evolução. Com isso outros estudos devem ser 17
realizados para uma melhor compreensão da ecologia de B. thuringiensis (VILAS-18
BÔAS; SANTOS, 2012). 19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34

23
2. OBJETIVOS
2.1 GERAL
Verificar a influência da presença dos plasmídeos pHT73-EmR
e 1
pHT73Δcry1Ac TetR no comportamento de linhagens de B. thuringiensis em larvas de 2
A. gemmatalis sobre as habilidades de germinação de esporos, multiplicação de células 3
vegetativas e esporulação. 4
5
6
2.2 ESPECÍFICOS 7
8
1- Obter linhagens exconjugantes de B. thuringiensis contendo o 9
plasmídeo pHT73-EmR que possui o gene cry intacto e um gene de resistência ao 10
antibiótico eritromicina; 11
2- Obter linhagens exconjugantes de B. thuringiensis contendo o 12
plasmídeo pHT73Δcry1Ac TetR
no qual o gene cry foi substituído por um gene de 13
resistência ao antibiótico tetraciclina; 14
3- Determinar a capacidade de germinação, crescimento e esporulação 15
de linhagens selvagens de B. thuringiensis em larvas de A. gemmatalis; 16
4- Determinar o comportamento de linhagens exconjugantes de B. 17
thuringiensis contendo o plasmídeo pHT73-EmR
com o gene cry ativo e o plasmídeo 18
pHT73Δcry1Ac TetR sem o gene cry em larvas de A. gemmatalis, verificando a 19
influência dessas linhagens; 20
5- Fazer um estudo comparativo de germinação, crescimento e 21
esporulação entre as linhagens de B. thuringiensis contendo o plasmídeo pHT73-EmR
22
com o gene cry ativo, ou plasmídeo pHT73Δcry1Ac TetR sem o gene cry ou nenhum 23
dos dois plasmídeos. 24
25
26
27
28
29
30

24
REFERÊNCIAS 1
2
ADAMS, M. R.; MOSS, M. O. Food Microbiology. Royal Society of Chemistry, 3
Cambridge, p. 244-250, 1995. 4
5
AGAISSE, H.; LERECLUS, D. How does B. thuringiensis produce so much 6
insecticidal crystal protein? Journal of Bacteriology. v. 177, n. 21, p. 6027-6032, 1995. 7
8
AGAISSE, H.; GOMINET, M.; ØKSTAD, O. A.; KOLSTØ, A. B.; LERECLUS, D. 9
PlcR is a pleiotropic regulator of extracellular virulence factor gene expression 10
in Bacillus thuringiensis. Molecular Microbiology, v. 32, p. 1043–1053, 1999. 11
12
AGATA, N.; OHTA, M.; MORI, M.; ISOBE, M. A novel dodecadedepsipeptide, 13
cerulide, is an emetic toxin of Bacillus cereus. FEMS Microbiology Letters, v. 129, n. 14
1, p. 17-20, 1995. 15
16
ALY, C.; MULLA, M. S.; FEDERICI, B. A. Sporulation and toxin production by 17
Bacillus thuringiensis var. israelensis in cadavers of mosquito larvae. Journal of 18
Invertebrate Pathology.v. 46, n. 3, p. 251–258, 1985. 19
20
AMADIO, A. F.; BENINTENDE, G. B.; ZANDOMENI, R. O. Complete sequence of 21
three plasmids from Bacillus thuringiensis INTA-FR7-4 environmental isolate and 22
comparison with related plasmids from the Bacillus cereus group. Plasmid. v. 62, n. 3, 23
p. 172–182, 2009. 24
25
ANDRUP, L.; DAMGAARD, J.; WASSERMANN, K. Mobilization of small plasmids 26
in Bacillus thuringiensis subsp. israelensis is accompanied by specific aggregation. 27
Journal of Bacteriology. v. 175, n. 20, p. 6530–6536, 1993. 28
29
ANDRUP, L.; JØRGENSEN, O.; WILCKS, A.; SMIDT, L.; JENSEN, G. B. 30
Mobilization of ‘‘Nonmobilizable’’ Plasmids by the Aggregation-Mediated. Plasmid, v. 31
36, n. 2, p. 75-85, 1996. 32
33
ANKENBAUER, R. G. Reassessing forty years of genetic doctrine: retrotransfer and 34
conjugation.Genetics. v. 145, n. 3, p.543–549, 1997. 35
36
APTOSOGLOU, S.G.; SIVROPOULOU, A.; KOLIAIS, S.I. Plasmid patterns of 37
Bacillus thuringiensis strains and isolates. Microbios. v. 91, n. 368-369, p. 203 – 214, 38
1997. 39
40
ARANTES, O. M. N.; VILAS-BÔAS, L. A.; VILAS-BÔAS, G. F. L. T. Bacillus 41
thuringiensis: estratégias no controle biológico. In: SERAFINE, L.A; BARROS, N. M; 42
AZEVEDO, J. L. (Org.). Biotecnologia: avanços na agricultura e na agroindústria. 43
Caxias do Sul: Agropecuária. p. 269-293, 2002. 44
45
ARNESEN, L. P. S.; FAGERLUND, A.; GRANUM, P. E. From soil to gut: Bacillus 46
cereus and its food poisoning toxins. FEMS Microbiology Reviews. v. 32, n. 4, p. 579-47
606, 2008. 48
49

25
BATTISTI, L.; GREEN, B.; THORNE, C. Mating system for transfer of plasmids 1
among Bacillus anthracis, Bacillus cereus, and Bacillus thuringiensis. Journal of 2
Bacteriology. v. 162, n. 2, p. 543–550, 1985. 3
4
BAUMANN, L.; OKAMOTO K.; UNTERMAN B. M.; LYNCH M. J.; BAUMAN, P. 5
Phenotypic characterization of Bacillus thuringiensis and Bacillus cereus. Journal of 6
Invertebrate Pathology. v. 44, n. 3, p. 329-341, 1984. 7
8
BAVYKIN, S.G.; LYSOV, Y.P.; ZAKHARIEV, V.; KELLY, J. J.; JACKMAN, J.; 9
STAHL, D.A.; CHERNI, A.; 2004. Journal Clinical of Microbiology. 42, 3711 – 373, 10
2004. 11
12
BEECHER, D. J., SCHOENI, J. L.; WONG, A. C. Enterotoxic activity of hemolysin 13
BL from Bacillus cereus. Infection and Immunity. v. 63, n. 11, p. 4423–4428, 1995. 14
15
BEIJERSBERGEN, A.; DEN DULK-RAS, A.; SCHILPEROORT, R. A.; 16
HOOYKAAS, P. J. J. Conjugative transfer by the virulence system of Agrobacterium 17
tumefaciens. Science. v. 256, n. 5061, p. 1324–1327, 1992. 18
19
BERNHARD, K.; JARRET, P.; MEADOWS, M.; BUTT, J.; ELLIS, D. J.; ROBERTS, 20
G. M.; PAULI, S.; RODGERS, P.; BURGES, H. D. Natural isolates of Bacillus 21
thuringiensis worldwide distribution, characterization and activity against insect pests. 22
Journal of Invertebrate Pathology. v. 70, p. 59 - 68, 1997. 23
24
BERRY, C.; O’ NEIL, S.; BEN-DOV, E.; JONES, A. F.; MURPHY, L.; QUAIL, M. 25
A.; HOLDEN, M. T. G.; HARRIS, D.; ZARITSKY, A.; PARKHILL, J. Complete 26
sequence and organization of pBtoxis, the toxin-coding plasmid of Bacillus 27
thuringiensis subsp. israelensis. Applied and Environmental Microbiology. v. 68, n. 28
10, p. 5082–5095, 2002. 29
30
BERTOLLA, F.; SIMONET, P. Horizontal gene transfers in the environment: natural 31
transformation as a putative process for gene transfers between transgenic plants and 32
microorganisms. Research in Microbiology. v. 150, n. 6, p. 375–384, 1999. 33
34
BIZZARRI, M. F.; BISHOP, A. H. The ecology of Bacillus thuringiensis on the 35
Phylloplane: colonization from soil, plasmid transfer, and interaction with larvae of 36
Pieris brassicae. Microbial Ecology. v. 56, n. 1, p. 133–139, 2008. 37
38
BOURGOUIN, C.; DELÉCLUSE, A.; RIBIER, J.; KLIER, A.; RAPOPORT, G. A 39
Bacillus thuringiensis subsp. israelensis gene encoding a 125-kilodalton larvicidal 40
polypeptide is associated with inverted repeat sequences. Journal Bacteriology. v. 170, 41
n.8, p. 3575-3583, 1988. 42
43
BRAR, S. K.; VERMA, M.; TYAGI, R. D.; VALÉRO J. R. Recent advances in 44
downstream processing and formulations of Bacillus thuringiensis based biopesticides. 45
Process Biochemistry. v. 41, n. 2, p. 323-342, 2006. 46
47
BRAVO, A.; GILL, S. S.; SOBERÓN, M. Mode of action of B. thuringiensis Cry and 48
Cyt toxins and their potential for insect control. Toxicon. v. 49, n. 4, p. 423-435, 2007. 49
50

26
BROOTHAERTS, W.; MITCHELL, H. J.; WEIR, B.; KAINES, S.; SMITH, L. M.; 1
YANG, W.; MAYER, J. E.; ROA-RODRIGUEZ, C.; JEFFERSON, R. A. Gene transfer 2
to plants by diverse species of bacteria. Nature. v. 433, n. 7026, p. 629–633, 2005. 3
4
CAPALBO, D. M. F.; VILAS-BÔAS, G. T.; ARANTES, O. M. N. Bacillus 5
thuringiensis: formulações e plantas transgênicas. In: BORÉM, A. (Ed.). Biotecnologia 6
e meio ambiente. Viçosa: Folha de Viçosa, 2004. p. 309-350. 7
8
CARLSON, C. R.; CAUGANT D. A.;KOLSTØ, A. B. Genotypic Diversity among 9
Bacillus cereus and Bacillus thuringiensis Strains. Applied and Environmental 10
Microbiology. v. 60, n. 6, p. 1719-1725, 1994. 11
12
CHAUFAUX, J.; MARCHAL, M.; GILOIS, N.; JEHANNO, I.; BUISSON, C. 13
Recherche de souches naturelles du Bacillus thuringiensis dans différents biotopes, à 14
travers le monde. Canadian Journal Microbiology. v. 43, p. 337 - 343, 1997. 15
16
CHEN, L.; CHEN, Y.; WOOD, D. W.; NESTER, E. W. A new type IV secretion 17
system promotes conjugal transfer in Agrobacterium tumefaciens. Journal of 18
Bacteriology. v. 184, n. 17, p. 4838–4845, 2002. 19
20
CHEN, I.; CHRISTIE, P. J.; DUBNAU, D. The Ins and Outs of DNA transfer in 21
bacteria. Science. v. 310, n. 5753, v. 1456–1460, 2005. 22
23
CHERIF, A.; BRUSETTI, L.; BORIN, S.; RIZZI, A.; BOUDABOUS, A.; HYAMI-24
HORANI, H. DAFFONCHIO, D. Genetic relationshio in the “Bacillus cereus group” 25
by rep-PCR fingerprint and sequencing of Bacillus anthracis- specific rep-PCR 26
fragment. Journal Applied Microbiology. v. 94, p. 1108-1119, 2003. 27
28
CHILCOTT, C. N.; WIGLEY, P. J. Isolation and toxicity of B. thuringiensis from soil 29
and insect habitats in New Zealand. Journal of Invertebrate Pathology. v. 61, n. 3, p. 30
244-247, 1993. 31
32
CHRISTIE, P. J.; VOGEL, J. P. Bacterial type IV secretion: conjugation systems 33
adapted to deliver effector molecules to host cells. Trends in Microbiology. v. 8, n. 8, 34
p. 354–360, 2000. 35
36
CHRISTIE, P. J. Type IV secretion: the Agrobacterium virB/D4 and related conjugation 37
systems. Biochimica et Biophysica Acta. v.1694, n. 1-3, p. 219-234, 2004. 38
39
CHUMAKOV, M. I. Transfer of genetic information from agrobacteria to bacterial and 40
plant cells: membrane and supramembrane structures involved in transfer. Membrane 41
and Cell Biology. v. 14, n. 3, p. 309–331, 2000. 42
43
COPPING, L. G.; MENN, J. J. Review biopesticides: a review of their action, 44
applications and efficacy. Pest Management Science. v. 56, n. 5, p. 651-676, 2000. 45
46
COHN, F. Ueber die Bakterien, die kleinsten lebenden Wesen. Sammlung 47
Gemeinverständlicher Wissenschattlicher Vorträge, Carl Habel, Berlin, 1872. 48
49
COHAN, F. M. The effects of rare but promiscuous genetic exchange on evolutionary 50

27
divergence in prokaryotes. The American Naturalist. v. 143, n. 6, p. 965–86, 1994. 1
2
COHAN, F. M. What are bacterial species? Annual Reviews of Microbiology. v. 56, p. 3
457-487, 2002. 4
5
CRICKMORE, N.; ZEIGLER, D.R.; FEITELSON, J.; SCHNEPF, E.; VAN RIE, J.; 6
LERECUS, D.; BAUM, J. and DEAN, D.H. Revision of the nomenclature for the 7
Bacillus thuringiensis pesticidal crystal proteins. Microbiology and Molecular Biology 8
Reviews. v. 62, nº. 3, p. 807-813, 1998. 9
10
DAFFONCHIO, D.; CHERIF, A.; BORIN, S. Homoduplex and heteroduplex 11
polymorphisms of the amplified ribosomal 16S–23S internal transcribed spacers 12
describe genetic relationships in the “Bacillus cereus group.” Applied and 13
Environmental Microbiology. v. 66, n. 12, p. 5460–5468, 2000. 14
15
DAMGAARD, P.H.; ABDEL-HAMEED, A.; EILENBERG, J.; SMITS, P.H. Natural 16
occurrence of Bacillus thuringiensis on grass foliage. World Journal of Microbiology 17
& Biotechnology. v. 14, p. 239-242, 1998. 18
19
DAVIS, B. D. Nonfiltrability of the agents of recombination in Escherichia coli. 20
Journal of Bacteriology. v. 60, n. 4, p. 507-508, 1950. 21
22
DROBNIEWSKI, F. A. Bacillus cereus and related species. Clinical Microbiology 23
Reviews. v. 6, n. 4, p. 324-338, 1993. 24
25
EUZÉBY, J. P. List of Prokaryotic names with Standing in Nomenclature – Genus 26
Bacillus. Disponível em: < http://www.bacterio.cict.fr/b/bacillus.html > Acesso em: 04 27
de setembro de 2012. 28
29
FEITELSON, J. S.; PAYNE, J.; KIM, L. Bacillus thuringiensis: insects and beyond. 30
Bio/Technology, v.10, p.271-275, 1992. 31
32
FIRTH, N.; IPPEN-IHLER, K.; SKURRAY, R. A. Structure and function of the F 33
factor and mechanism of conjugation. In: Escherichia coli and Salmonella: Cellular 34
and Molecular Biology. Edited by NEIDHART, F. C.; CURTISS, III R.; 35
INGRAHAM, J. L.; LIN, E. C. C.; LOW, K. B.; MAGASANIK, B.; REZNIKOFF, W. 36
S.; RILEY, M.; SCHAECHTER, M.; UMBARGER, H. C. American Society for 37
Microbiology. p. 2377-2401, 1996. 38
39
FURLANETO, L. M.; SARIDAKIS, H. O.; ARANTES, O. M. N. Survival and 40
conjugal transfer between Bacillus thuringiensis strains in aquatic environment. 41
Brazilian Journal of Microbiology. v. 31, n. 4, p. 233-238, 2000. 42
43
FERRÉ, J.; VAN RIE, J. Biochemistry and Genetics of Insect Resistance to Bacillus 44
thuringiensis. Annual Review of Entomology. v. 47, p. 501-533, 2002. 45
46
GARCILLÁN-BARCIA, M. P.; FRANCIA, M. V.; DE LA CRUZ, F. The diversity of 47
conjugative relaxases and its application in plasmid classication. FEMS Microbiology 48
Reviews. v. 33, n. 3, p. 657-687, 2009. 49
50

28
GLARE, T. R.; O´CALLAGHAN, M. B. thuringiensis: biology, ecology and safety. 1
Chichester: John Wiley, 350 p., 2000. 2
3
GOHAR, M.; GILOIS, N.; GRAVELINE, R.; GARREAU, C.; SANCHIS, V.; 4
LERECLUS, D. A comparative study of Bacillus cereus, Bacillus thuringiensis and 5
Bacillus anthracis extracellular proteomes. Proteomics. v. 5, p. 3696–3711, 2005. 6
7
GOMIS-RÜTH, F. X.; COLL, M. Cut and move: protein machinery for DNA 8
processing in bacterial conjugation. Current Opinion in Structural Biology. v. 16, n. 6, 9
p. 744-752, 2006. 10
11
GONZÁLEZ, J. M. Jr.; CARLTON, B. C. Patterns of plasmid DNA in crystalliferous 12
and acrystalliferous strains of Bacillus thuringiensis. Plasmid. v. 3, n. 1, p. 92–98, 13
1980. 14
15
GONZÁLEZ, J. M. Jr., DULMAGE, H. T.; CARLTON, B. C. Correlation between 16
specific plasmids and delta-endotoxin production in Bacillus thuringiensis. Plasmid. v. 17
5, n. 3, p. 352–365, 1981. 18
19
GONZÁLES, J. M. Jr.; BROWN, B. S.; CARLTON, B. C. Transfer of B. thuringiensis 20
plasmids coding for d-endotoxin among strains of B. thuringiensis and B. cereus. 21
Proceedings of the National Academic of Science. v. 79, n. 10, p. 6951-6955, 1982. 22
23
GONZÁLEZ, J. M. Jr.; CARLTON, B. C. A large transmissible plasmid is required for 24
crystal toxin production in Bacillus thuringiensis variety israelensis. Plasmid. v. 11, n. 25
1, p. 28-38, 1984. 26
27
GRANUM, P. E. B. cereus and its toxins. Journal of Applied Microbiology. v. 76, n. 28
S 23, p. 61S-66S, 1994. 29
30
GRANUM, P. E.; LUND T. Bacillus cereus and its food poisoning toxins. FEMS 31
Microbiology Letters. v. 157, n. 2, p. 223–228, 1997. 32
33
GRANUM, P.E.; BAIRD-PARKER, A.C. Bacillus spp. In: The Microbiological 34
Quality and Safety of Food. Edited by LUND, B. M.; BAIRD-PARKER, T. C.; 35
GOULD, G. W. Aspen Publishers, Maryland. pp. 1029 – 1039, 2000. 36
37
GROHMANN, E.; MUTH, G.; ESPINOSA, M. Conjugative plasmid transfer in Gram-38
positive bacteria. Microbiology and Molecular Biology Reviews. v. 67, n. 2 , p. 277–39
301, 2003. 40
41
GUIDI, V.; PATOCCHI, N.; LÜTHY, P.; TONOLLA, M. Distribution of Bacillus 42
thuringiensis subsp. israelensis in soil of a Swiss Wetland reserve after 22 years of 43
mosquito control. Applied and Environmental Microbiology. v. 77, n. 11, p. 3663–44
3668, 2011. 45
46
HAN, C. S.; XIE, G.; CHALLACOMBE, J. F.; ALTHERR, M. R.; BHOTIKA, S. S.; 47
BROWN, N.; BRUCE, D.; CAMPBELL, C. S.; CAMPBELL, M. L.; CHEN, J.; 48
CHERTKOV, O.; CLELAND, C.; DIMITRIJEVIC, M.; DOGGETT, N. A.; 49

29
FAWCETT, J. J.; GLAVINA, T.; GOODWIN, L. A.; GREEN, L. D.; HILL, K. K.; 1
HITCHCOCK, P.; JACKSON, P. J.; KEIM, P.; KEWALRAMANI, A. R.; 2
LONGMIRE, J.; LUCAS, S.; MALFATTI, S.; McMURRY, K.; MEINCKE, L. J.; 3
MISRA, M.; MOSEMAN, B. L.; MUNDT, M.; MUNK, A. C.; OKINAKA, R. T.; 4
PARSON-QUINTANA, B.; REILLY, L. P.; RICHARDSON, P.; ROBINSON, D. L.; 5
RUBIN, E.; SAUNDERS, E.; TAPIA, R.; TESMER, J. G.; THAYER, N.; 6
THOMPSON, L. S.; TICE, H.; TICKNOR, L. O.; WILLS, P. L.; BRETTIN, T. S.; 7
GILNA, P. Pathogenomic sequence analysis of Bacillus cereus and Bacillus 8
thuringiensis isolates closely related to Bacillus anthracis. Journal of Bacteriology. v. 9
188, n. 9, p. 3382–3390, 2006. 10
11
HANSEN, B. M.; SALAMITOU, S. Virulence of B. thuringiensis. In: CHARLES, J. F.; 12
DELÉCLUSE, A.; NIELSEN-LE ROUX, C. (Ed.). Entomopathogenic bacteria: from 13
laboratory to field application. Netherlands: Kluwer Academic publishers, p. 41-64, 14
2000. 15
16
HAUGE, S. Matforgiftninger fremkalt av Bacillus cereus. Nordisk Hygienisk 17
Tidskrift. v. 31, n. 6, p. 189-206, 1950. 18
19
HAYMAN, G. T.; BOLEN, P. L. Movement of shuttle plasmids from Escherichia coli 20
into yeasts other than Saccharomyces cerevisiae using trans-kingdom conjugation. 21
Plasmid. v. 30, n. 3, p. 251–257, 1993. 22
23
HEINEMANN, J. A.; ANKENBAUER, R. G. Retrotransfer in Escherichia coli 24
conjugation: bidirectional exchange or de novo mating? Journal of Bacteriology. v. 25
175, n. 3, p. 583–588, 1993. 26
27
HEINEMANN, J. A.; SCOTT, H. E.; WILLIAMS, M. Doing the conjugative two-step: 28
evidence of recipient autonomy in retrotransfer. Genetics. v. 143, n. 3, p. 1425–1435, 29
1996. 30
31
HEINEMANN, J. A. Genetic evidence of protein transfer during bacterial conjugation. 32
Plasmid. v. 41, n. 3, p. 240–247, 1999. 33
34
HELGASON, E., D. A.; CAUGANT, M. M.; LECADET, Y.; CHEN, J.; MAHILLON, 35
A.; LÖVGREN, I.; HEGNA, K.; KVALOY, A. B.; KOLSTØ. Genetic diversity of 36
Bacillus cereus/Bacillus thuringiensis isolates from natural sources. Current 37
Microbiology. v. 37, p. 80–87, 1998. 38
39
HELGASON, E.; CAUGANT, D. A.; OLSEN, I.; KOLSTØ, A. B. Genetic structure of 40
population of Bacillus cereus and B. thuringiensis isolates associated with periodontitis 41
and other human infections. Journal of Clinical Microbiology. v. 38, n. 4, p. 1615-42
1622, 2000a. 43
44
HELGASON, E.; ØKSTAD, P. A.; CAUGANT, D. A.; JOHANSEN, H. A.; FOUET, 45
A.; MOCK, M.; HEGNA, I.; KØLSTO, A. B. Bacillus anthracis, Bacillus cereus, 46
Bacillus thuringiensis - One species on the basis of genetic evidence. Applied and 47
Environmental Microbiology. v. 66, n. 6, p. 2627-2630, 2000b. 48
49

30
HEYNDRICKX, M.; SCHELDEMAN, P. Bacilli associated with spoilage in dairy 1
products and ther food. In Applications and Systematics of Bacillus and Relatives, 2
Edited by Berkeley, R. C. W.; Heyndrickx, M.; Logan, N. A.; De Vos, P. Oxford: 3
Blackwell Science. p. 65–82, 2002. 4
5
JARRETT, P.; STEPHENSON, M. Plasmid transfer between strains of Bacillus 6
thuringiensis infecting Galleria mellonella and Spodoptera littoralis. Applied and 7
Environmental Microbiology. v. 56, p. 1608–1614, 1990. 8
9
JENSON, I.; MOIR, C. J. Bacillus cereus and other Bacillus species. In: HOCKING, 10
A. D.; ARNOLD, G.; JENSON, I.; NEWTON, K.; SUTHERLAND, P. 11
(ed.), Foodborne Microorganisms of Public Health Significance, 5th ed. Australian 12
Institute of Food Science and Technology Inc., Sydney, Australia, 1997. 13
14
JENSEN, G.B.; WILCKS, A.; PETERSEN, S. S.; DAMGAARD, J.; BAUM, J.A.; 15
ANDRUP, L. The genetic basis of the aggregation system in Bacillus thuringiensis 16
subsp. israelensis is located on the large conjugative plasmid pXO16. Journal 17
Bacteriology. v. 177, p. 2914–2917, 1995. 18
19
JENSEN, G. B.; ANDRUP, L.; WILCKS, A.; SMIDT, L.; POULSEN, O. M. The 20
aggregation-mediated conjugation system of Bacillus thuringiensis subsp. israelensis: 21
host range and kinetics. Current Microbiology. v. 33, n. 4, p. 228–236, 1996. 22
23
JOHNSON, J. L. Use of nucleic-acid homologies in the taxonomy of anaerobic bacteria. 24
International Journal of Systematic and Evolutionary Microbiology. v. 23, n. 4, p. 25
308– 315, 1973. 26
27
KANEKO, T.; NOZAKI, T.; AIZAWA, K. Deoxyribonucleic acid relatedness between 28
Bacillus anthracis, Bacillus cereus and Bacillus thuringiensis. Microbiology 29
Immunology 22:639-641, 1978. 30
31
KASHYAP, S.; AMLA, D. V. Characterisation of Bacillus thuringiensis kurstaki strains 32
by toxicity, plasmid profiles and numerical analysis of their cryIA genes. African 33
Journal of Biotechnology. v. 6, n. 15, p. 1821–1827, 2007. 34
35
KOEHLER, T. M.; THORNE, C. B. Bacillus subtilis (natto) plasmid pLS20 mediates 36
interspecies plasmid transfer. Journal of Bacteriology. v. 169, n. 11, p. 5271–5278, 37
1987. 38
39
KOTIRANTA, A. Epidemiology and pathogenesis of Bacillus cereus infections. 40
Microbes and Infection, v.2, nº 2, p.189-198, 2000. 41
42
KRAMER, J. M.; GILBERT, R. J. B. cereus and other Bacillus species. In: Foodborne 43
Bacterial Pathogens (DOYLE, M. P., Ed.), p. 21-70. Marcel Dekker, New York, 1989. 44
45
KRONSTAD, J. W.; SCHNEPF,H. E.; WHITELEY, H. R. Diversity of locations for 46
Bacillus thuringiensis crystal protein genes. Journal of Bacteriololy. v. 154, n. 1, p. 47
419–428, 1983. 48
49

31
KRONSTAD, W.; WHITELEY, H. R. Inverted repeat sequences flank a Bacillus 1
thuringiensis crystal protein gene. Journal Bacteriology. v.160, n.1, p.95-102, 1984. 2
3
LEDERBERG, J. Forty years of genetic recombination in bacteria. Nature. v. 324, n. 4
6098, p. 627–628, 1986. 5
6
LERECLUS, D. LECADET, M. M.; RIBIER, J.; DEDONDER, R. Molecular 7
relationships among plasmids of Bacillus thuringiensis: conserved sequences through 8
11 crystalliferous strains. Molecular and General Genetics. v. 186, n. 3, p. 391–398 , 9
1982. 10
11
LERECLUS D.; RIBIER, J.; KLIER, A.; MENOU, G.; LECADET, M. M. A 12
transposon-like structure related to the delta-endotoxin gene of Bacillus thuringiensis. 13
EMBO Journal. v.3, n.11, p.2561-2567, 1984. 14
15
LERECLUS, D.; BOURGOIN, C. LECADET, M. M.; KLIER, A.; RAPOPORT, G. 16
Role, structure and molecular organization of the genes coding for the parasporal delta-17
toxins of Bacillus thuringiensis. In: SMITH, I.; SLEPECKY, R. A., SETLOW, P. (Ed.). 18
Regulation of prokaryotic development. Washington: American Society for 19
Microbiology. p. 255-276, 1989. 20
21
LERECLUS, D.; DELECLUSE, A.; LECADET, M. M. Diversity of Bacillus 22
thuringiensis toxins and genes. In: ENTWISTLE, P. F.; CORY, J. S.; BAILEY, M. J.; 23
HIGGS, S. Bacillus thuringiensis an environmental biopesticide: theory and practice. 24
Chichester : J. Wiley E Sons, 1993. p.37-70. 25
26
LINZ, B.; SCHENKER, M.; ZHU, P.; ACHTMAN, M. Frequent interspecific genetic 27
exchange between commensal Neisseriae and Neisseria meningitides. Molecular 28
Microbiology. v. 36, n. 5, p. 1049-1058, 2000. 29
30
LLOSA, M.; GOMIS-RUTH, F. X.; COLL, M.; de la CRUZ, F. Bacterial conjugation: 31
a two-step mechanism for DNA transport. Molecular Microbiology. v. 45, n. 1, p. 1–8, 32
2002. 33
34
LOGAN, N. A.; BERELEY R. C. W. Identification of Bacillus strains using the API 35
system. Journal of General Microbiology. v. 130, n. 7, p. 1871-1882, 1984. 36
37
LUND, T., DE BUYSER, M. L.; GRANUM, P. E. A new cytotoxin from Bacillus 38
cereus that may cause necrotic enteritis. Molecular Microbiology. v. 38, n. 2, p. 254–39
261, 2000. 40
41
LYSENKO, O. Bacillus thuringiensis: evolution of a taxonomic conception. Journal of 42
Invertebrate Pathology. v. 42, n. 3, p. 295-298, 1983. 43
44
McDOWELL, D. G.; MANN, N. H. Characterization and sequence analysis of a small 45
plasmid from Bacillus thuringiensis var. kurstaki strain HD1-DIPEL. Plasmid. v. 25, n. 46
2, p. 113-120, 1991. 47
48

32
MAHMOOD, A.; KIMURA, T.; TAKENAKA, M.; YOSHIDA, K. The construction of 1
novel mobilizable YAC plasmids and their behavior during trans-kingdom conjugation 2
between bacteria and yeasts. Genetic Analysis. v. 13, n. 2 ,p. 25–31, 1996. 3
MAJEWSKI, J.; COHAN, F. M. Adapt globally, act locally: the effect of selective 4
sweeps on bacterial sequence diversity. Genetics. v.152, n. 4, p.1459-1474, 1999. 5
6
MARTIN, P. A. W.; TRAVERS, R. S. Worldwide abundance and distribution of B. 7
thuringiensis isolates. Applied and Environmental Microbiology. v. 55, n. 10, p. 8
2437-2442, 1989. 9
10
MAYR, E. What is a species, and what is not? Philosophy of Science. v. 63, n. 2, p. 11
262-277, 1996. 12
13
MAYR, R.; EPPERT, I.; SCHERER, S. Incidence and identification of psychrotrophic 14
(7 °C-tolerant) Bacillus spp. in German HTST pasteurized milk. Milchwissenschaft. v. 15
54, p. 26–30, 1999. 16
17
MEADOWS, M. P.; ELLIS, D. J.; BUTT, J.; JARRET, P.; BURGES, H. D.; 18
Distribution, frequency, and diversity of B. thuringiensis in animal feed mill. Applied 19
and Environmental Microbiology. v. 59, n. 4, p. 1344-1350, 1992. 20
21
MEER, R. R.; BAKER, J.; BODYFELT F. W.; GRIFFITHS, M. W. Psychrotrophic 22
Bacillus spp in fluid milk products: A review. Journal of Food Protection, v. 54, n. 23
12, p. 969-979, 1991. 24
25
MIGNOT, E.; LIN, L.; ROGERS, W.; HONDA, Y.; QIU, X.; LIN, X.; OKUN, M.; 26
HOHJOH, H.; MIKI, T.; HSU, S.H. Complex HLA-DR and DQ interactions confer risk 27
for narcolepsy-cataplexy in three ethnic groups. The American Jounal Human 28
Genetics. v. 68, p. 686–699, 2001. 29
30
MONCALIÁN, G.; CABEZÓN, E.; ALKORTA, I.; VALLE, M.; MORO, 31
F.; VALPUESTA, J. M.; GOÑI, F. M.; de La CRUZ, F. Characterization of ATP and 32
DNA binding activities of TrwB, the coupling protein essential in plasmid R388 33
conjugation. Journal Biology Chemical. v. 274, n. 51, p. 36117-36124, 1999. 34
35
MORAES, I. O.; CAPALBO, D. M. F. Produção de bactérias entomopatogênicas. In: 36
ALVES, S. B. (Ed.). Controle microbiano de insetos. Piracicaba: Manole. p. 127-170, 37
1986. 38
39
NAKAMURA, L.K. Bacillus pseudomycoides sp. nov. International Journal of 40
Systematic Bacteriology. v. 48, n. p. 1031–1034, 1998. 41
42
OCHMAN, H.; LAWRENCE J. G.; GROISMAN, E. A. Lateral gene transfer and the 43
nature of bacterial innovation. Nature. v. 405, p. 299 - 304, 2000. 44
45
OHBA, M.; AIZAWA, K. Distribution of Bacillus thuringiensis in soils of Japan. 46
Journal of Invertebrate Pathology. v.47, n. 3, p. 277-282, 1986. 47
48

33
OULTRAM, J. D.; YOUNG, M. Conjugal transfer of plasmic pAMb1 from 1
Streptococcus lactis and Bacillus subtilis to Clostridium acetobutylicum. FEMS 2
Microbiology Letters. v. 27, n. 1, p. 129–134, 1985. 3
4
PETERSEN, D.J.; SHISHIDO, M.; BRIAN HOLL, F.; CHANWAY, C.P. Use of 5
species- and strain-specific PCR primers for identification of conifer root-associated 6
Bacillus spp. FEMS Microbiology Letters. v. 133, n. 1-2, p. 71–76, 1995. 7
8
PIGOTT, C.R.; ELLAR, D.J. Role of Receptors in Bacillus thuringiensis Crystal Toxin 9
Activity. Microbiology and Molecular Biology Reviews. v. 71, n. 2, p. 255-281, 2007. 10
11
PRIEST, F. G.; GOODFELLOW M.; TODD, C. A numerical classification of the genus 12
Bacillus. Journal of General Microbiology. v. 134, n. 7, p. 1847-1882, 1988. 13
14
RAYMOND, B.; WYRES, K. L.; SHEPPARD, S. K.; ELLIS, R. E.; BONSALL, M. B. 15
Environmental Factors Determining the Epidemiology and Population Genetic Structure 16
of the Bacillus cereus Group in the Field. PLoS Pathogens. v. 26, n. 6(5), e1000905, 17
2010a. 18
19
RAYMOND, B.; JOHNSTON, P. R.; NIELSEN-LEROUX, C.; LERECLUS, D.; 20
CRICKMORE, N. Bacillus thuringiensis: an impotent pathogen? Trends in 21
Microbiology. v. 18, n. 5, p. 183-232, 2010b. 22
23
RAYMOND, B.; WEST, S. A.; GRIFFIN, A. S.; BONSALL, M. B. The Dynamics of 24
Cooperative Bacterial Virulence in the Field. Science. v. 337, n. 6090, p. 85-88, 2012. 25
26
REDDY, A.; BATTIST, L.; THORNE, C.B. Identification of self-transmissible 27
plasmids in four Bacillus thuringiensis subspecies. Journal Bacteriology. v. 169, p. 28
5263–5270, 1987. 29
30
SALAMITOU, S.; RAMISSE, F.; BREHÉLIN, M.; BOURGUET, D.; GILOIS, 31
N.; GOMINET, M.; HERNANDEZ, E.; LERECLUS, D. The plcR regulon is involved 32
in the opportunistic properties of Bacillus thuringiensis and Bacillus cereus in mice and 33
insects. Microbiology. v. 146, n. 11, p. 2825-2832, 2000. 34
35
SANCHIS, V.; LERECLUS, D.; MENOU, G.; CHAUFAUX, J.; LECADET, M. M. 36
Multiplicity of d-endotoxinas genes with different insecticidal specificities in B. 37
thuringiensis aizawai 7.29. Molecular Microbilogy. v. 2, n. 2, p. 393-404, 1998. 38
39
SANTOS, C.; VILAS-BÔAS, G. T.; LERECLUS, D.; SUZUKI, M. T.; ANGELO, E. 40
A.; ARANTES, O. M. N. Conjugal transfer between Bacillus thuringiensis and Bacillus 41
cereus strains is not directly correlated with growth of recipient strains. Journal of 42
Invertebrate Pathology. v. 105, n. 2, p.171–175, 2010. 43
44
SCHNEPF, E.; CRICKMORE, N.; VAN RIE, J.; LERECLUS, D.; BAUM, J.; 45
FEITELSON, J.; ZEIGLER, D. R.; DEAN, D. H. B. thuringiensis and its pesticidal 46
crystal proteins. Microbiology and Molecular Biology Reviews. v. 62, n. 3, p. 775-47
806, 1998. 48
49

34
SENESI, S.; GHELARDI E. Production, secretion and biological activity of Bacillus 1
cereus enterotoxins. Toxins. v. 2, n. 7, p. 1690-1703, 2010. 2
3
SIA, E. A.; KUEHNER, D. M.; FIGURSKI, D. H. Mechanism of retrotransfer in 4
conjugation: prior transfer of the conjugative plasmid is required. Journal of 5
Bacteriology. v. 178, n. 5, p. 1457–1464, 1996. 6
7
SMITH, R. A.; COUCHE, G. A. The phylloplane as a source of B. thuringiensis 8
variants. Applied and Environmental Microbiology. v. 57, n. 1, p. 311-315, 1991. 9
10
SMITH, J. M.; DOWSON, C. G.; SPRATT, B. G. Localized sex in bacteria. Nature, v. 11
349, p. 29-31, 1991. 12
13
STACKEBRANDT, E.; GOEBEL, B. M. 1994. Taxonomic note: a place for 14
DNA:DNA reassociation and 16S rRNA sequence analysis in the present species 15
definition in bacteriology. International Journal of Systematic and Evolutionary 16
Microbiology. v. 44, n. 4, p. 846– 849, 1994. 17
18
STACKEBRANDT, E.; SWIDERSKI, J. From phylogeny to systematics: the dissection 19
of the genus Bacillus. In: Berkeley, R.; Heyndrickx, M.; Logan, N.; De Vos, P. 20
Applications and systematics of Bacillus and relatives. p. 8-22, 2002. 21
22
SUZUKI, M. T.; LERECLUS, D. ARANTES, O. M. Fate os B. thuringiensis strains in 23
different insect larvae. Canadian Journal of Microbiology. v. 50, n. 11, p. 973-975, 24
2004. 25
26
TAKATSUKA, J.; KUNIMI, Y. Intestinal bacteria affect growth of Bacillus 27
thuringiensis in larvae of the oriental tea tortrix, Homona magnanima diakonoff 28
(Lepidoptera: tortricidae). Journal of Invertebrate Pathology. v. 73, n.3, p. 222-29
226, 2000. 30
31
TATO, I.; ZUNZUNEGUI, S.; de la CRUZ, F.; CABEZÓN, E. TrwB, the coupling 32
protein involved in DNA transport during bacterial conjugation, is a DNA-dependent 33
ATPase. Proceeding of the National Academy of Sciences of the United States of 34
America. v. 102, n. 23, p. 8156-8161, 2005. 35
36
THOMAS, D. J. I.; MORGAN, J. A. W.; WHIPPS, J. M.; SAUNDERS, J. R. Plasmid 37
transfer between the Bacillus thuringiensis subspecies kurstaki and tenebrionis in 38
laboratory culture and soil and in Lepidopteran and Coleopteran larvae. Applied and 39
Environmental Microbiology. v. 66, p. 118–124, 2000. 40
41
THOMAS, D. J. I.; MORGAN, J. A. W.; WHIPPS, J. M.; SAUNDERS, J. R. Plasmid 42
transfer between the Bacillus thuringiensis subspecies. israelensis strains in laboratory 43
culture, river water, and dipteran larvae. Applied and Environmental Microbiology. 44
v. 67, p. 330–338, 2001. 45
46
THOMAS, D. J. I.; MORGAN, J. A. W.; WHIPPS, J. M.; SAUNDERS, J. R. Transfer 47
of plasmid pBC16 between Bacillus thuringiensis strains in non-susceptible larval. 48
FEMS Microbiology. v. 40, p.181–190, 2002. 49

35
1
TICKNOR, L. O.; KØLSTO, A.; HILL, K. K.; KEIM, P.; LAKER, M. T.; TONKS, M.; 2
JACKSON, P. J. Fluorescent Amplified Fragment Length Polymorphism Analysis of 3
Norwegian Bacillus cereus and Bacillus thuringiensis soil isolates. Applied and 4
Environmental Microbiology, Washington, v.67, nº 10, p.4.863-4.873, 2001. 5
6
TIMMERY, S.; MODRIE, P.; MINET, O.; MAHILLON, J. Plasmid Capture by the 7
Bacillus thuringiensis Conjugative Plasmid pXO16. Journal of Bacteriology. v. 191, n. 8
7, p. 2197–2205, 2009. 9
10
TOP, E. M.; VANROLLEGHEM, P.; MERGEAY, M.; VERSTRAETE, W. 11
Determination of the mechanism of retrotransfer by mechanistic mathematical 12
modeling. Journal of Bacteriology. v. 174, n. 18, p. 5953-5960, 1992. 13
14
VALADARES-INGLIS, M. C. C.; SOUZA, M. T.; SHILER, W. Engenharia genética 15
de microrganismos agentes de controle biológico. In: MELO, I. S.; AZEVEDO, J. L. 16
(Ed.). Controle biológico. EMBRAPA: Jaguariúna, p. 102-225, 1998. 17
18
VAN DER AUWERA, G.A.; TIMMERY, S.; HOTON, F.; MAHILLON, J. Plasmid 19
exchanges among members of the Bacillus cereus group in foodstuffs. International 20
Journal of Food Microbiology. v. 113, p. 164–172, 2007. 21
22
VILAS-BÔAS, G. F. L. T.; VILAS-BÔAS, L. A.; LERECLUS, D.; ARANTES, O. M. 23
N. Bacillus thuringiensis conjugation under environmental conditions. FEMS 24
Microbiology. v. 25, p. 369–374, 1998. 25
26
VILAS-BÔAS, G.; SANCHIS, V.; LERECLUS, D.; LEMOS, M.; BOURGUET, D. 27
Genetic differentiation between sympatric populations of Bacillus cereus and Bacillus 28
thuringiensis. Applied and Environmental Microbiology. v. 68, p. 1414–1424, 2002. 29
30
VILAS-BÔAS, G. T.; LEMOS, M. V. F. Diversity of cry genes and genetic 31
characterization of B. thuringiensis isolated from Brazil. Canadian Journal of 32
Microbiology. v. 50, n. 8, p. 605-613, 2004. 33
34
VILAS-BÔAS, G. T.; PERUCA, A. P. S.; ARANTES, O. M. N. Biology and taxonomy 35
of Bacillus cereus, Bacillus anthracis, and Bacillus thuringiensis. Canadian Journal of 36
Microbiology. v. 53, n. 6, p. 673-687, 2007. 37
38
VILAS-BÔAS, G, T. SANTOS, C. A. Conjugation in B. thuringiensis: Insights into the 39
Plasmids Exchange Process. In: B. thuringiensis Biotechnology. SANSINENEA, E. 40
(ed.) Springer, 2012, p. 159-174. 41
42
YAMAMOTO, T.; DEAN, D. H. Insecticidal proteins produced by bacterial pathogenic 43
to agriculturas pests. In: CHARLES, J. F.; DELÉCLUSE, A.; NIELSEN-LE ROUX, C. 44
(Ed.). Entomopathogenic bacteria: from laboratory to fiels application. Netherlands; 45
Kluwer Academic Publichers, p. 81-100, 2000. 46
47
YUAN, Y. M.; HU, X. M.; LIU, H. Z.; HANSEN, B. M.; YAN, J. P.; YUAN, Z. M. 48
Kinetics of plasmid transfer among Bacillus cereus group strains within lepidopteran 49
larvae. Archives of Microbiology. v. 187, n. 6, p. 425–431, 2007. 50

36
1
WIJNANDS, L. M.; DUFRENNE, J. B.; VAN LEUSDEN, F. M. Characterization of 2
Bacillus cereus. Disponível em: <http://www.rivm.nl/bibliotheek/rapporten/250912002. 3
pdf>. Acesso em 28 de setembro de 2012, 2002. 4
5
WILCKS, A.; JAYASWAL, N.; LERECLUS, D.; ANDRUP, L. Characterization of 6
plasmid pAW63, a second self-transmissible plasmid in Bacillus thuringiensis subsp. 7
kurstaki HD73. Microbiology. v. 144, p. 1263–1270, 1998. 8
9
WILCKS, A.; SMIDT, L.; BAHL, M. I.; HANSEN, B. M.; ANDRUP, L.; 10
HENDRIKSEN, N. B.; LICHT, T. R. Germination and conjugation of Bacillus 11
thuringiensissubsp. israelensis in the intestine of gnotobiotic rats. Journal Applied of 12
Microbiology. v. 104, n. 5, p. 1252–1259, 2008. 13
14
WIRTH, M. C.; GEORGHIOU, G. P.; FEDERICI, B. A. CytA enables CryIV 15
endotoxins of Bacillus thuringiensis to overcome high levels of CryIV resistance in the 16
mosquito, Culex quinquefasciatus. Agricultural Sciences. v. 94, n. 20, p. 10536-10540, 17
1997. 18
19
ZECHNER, E. L.; LANG, S.; SCHILDBACH, J. F. Assembly and mechanisms of 20
bacterial type IV secretion machines. Biological Science. v. 367, n. 1592, p. 1073-1087, 21
2012. 22
23
ZHONG, C.; PENG, D.; YE, W.; CHAI, L.; QI, J.; YU, Z.; RUAN, L.; SUN, M. 24
Determination of Plasmid Copy Number Reveals the Total Plasmid DNA Amount Is 25
Greater than the Chromosomal DNA Amount in Bacillus thuringiensis YBT-1520. 26
PLoS ONE. v. 6, n. 1, e16025, 2011. 27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43

37
1
2
3
4
5
6
7
8
9
10
3. ARTIGO 11
12
EFEITO DO GENE cry SOBRE A CAPACIDADE DE 13
GERMINAÇÃO, CRESCIMENTO E ESPORULAÇÃO DE 14
Bacillus thuringiensis INFECTANDO LARVAS DE 15
Anticarsia gemmatalis 16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31

38
1
3. 1 RESUMO 2
3
A sistemática bacteriana não possui até hoje consenso quanto ao conceito de espécie, o 4
que tem gerado discussões sobre a classificação de diferentes espécies, como as 5
estudadas dentro do grupo Bacillus cereus lato sensu, que inclui além de Bacillus 6
anthracis, B. cereus stricto sensu e Bacillus thuringiensis. Essas duas últimas espécies 7
compartilham um grande nível de similaridade cromossômica e são fenotipicamente 8
idênticas. A diferenciação destas espécies é realizada por um gene plasmidial, 9
denominado gene cry. Esse gene codifica um cristal paraesporal (composto por 10
proteínas Cry) que confere a atividade entomopatogênica. Uma das formas de dispersão 11
desses plasmídeos é pelo processo de conjugação. Essa troca de material genético entre 12
as espécies de bactérias dificulta a classificação dessas espécies. Com o objetivo de 13
avaliar a influência do plasmídeo pHT73 e do gene cry1Ac sobre o comportamento de 14
linhagens de B. thuringiensis, larvas de 2º ínstar de Anticarsia gemmatalis, foram 15
alimentadas com dietas contendo culturas esporuladas dessa espécie bacteriana. Em 16
seguida, cada larva foi avaliada quanto à germinação de esporos, multiplicação e 17
sobrevivência das linhagens selvagens, B. thuringiensis subsp. thuringiensis 407-1; B. 18
thuringiensis subsp. kurstaki KT0 pHT73-EmR e B. thuringiensis subsp. kurstaki KT0 19
pHT73Δcry1Ac TetR, e linhagens exconjugantes B. thuringiensis subsp. thuringiensis 20
407-1 pHT73-EmR e B. thuringiensis subsp. thuringiensis 407-1 pHT73Δcry1Ac Tet
R, 21
sendo que cada experimento foi realizado com 10 repetições. As avaliações foram 22
realizadas em diversos tempos, incluindo um tempo antes da morte, t0, t24 e t48 horas 23
após a morte das larvas. As análises estatísticas evidenciaram que linhagens contendo o 24
plasmídeo pHT73-EmR se multiplicam em uma taxa maior que linhagens contendo o 25
plasmídeo pHT73Δcry1Ac TetR, demonstrando que o gene cry confere uma vantagem 26
nas linhagens que o abrigam. As linhagens que possuem o plasmídeo pHT73Δcry1Ac 27
TetR
se multiplicam a uma taxa maior que linhagens que não apresentam nenhum dos 28
dois plasmídeos estudados. Portanto, as linhagens que possuem o plasmídeo pHT73-29
EmR
são mais adaptadas ao ambiente larval quando comparadas às linhagens que 30
carregam o plasmídeo pHT73Δcry1Ac TetR
ou as que não possuem nenhum dos dois 31
plasmídeos. As linhagens que possuem o plasmídeo pHT73Δcry1Ac TetR
possuem uma 32
vantagem em relação as linhagens que não possuem nenhum dos dois plasmídeos, com 33
isso pode-se inferir que existem outros genes plasmidiais que influenciam no 34
comportamento de linhagens desse grupo bacteriano. Estudos posteriores poderão 35
evidenciar se essa influência do plasmídeo pHT73 e do gene cry1Ac sobre o 36
comportamento de linhagens de B. thuringiensis é espécie específico ou se essa 37
influência ocorre também em linhagens de B. cereus, além de apontar o papel desses 38
genes e sua importância no comportamento bacteriano. 39
40
Palavras-chave: Bacillus cereus sensu lato. Conjugação. Plasmídeo. 41
42
43
44

39
3.2 INTRODUÇÃO
O grupo do Bacillus cereus sensu lato inclui seis espécies bacterianas 1
destacando-se, Bacillus thuringiensis, Bacillus cereus e Bacillus anthracis. B. 2
thuringiensis é uma bactéria de importância no controle biológico de insetos praga de 3
lavoura e vetores de doenças. Sua atividade entomopatogênica é devida à ação das 4
proteínas Cry (SCHNEPF et al., 1998), cujos genes estão, geralmente, presentes em 5
grandes plasmídeos conjugativos. Alguns autores acreditam que a presença destes 6
plasmídeos e, consequentemente, a atividade entomopatogênica, permite que linhagens 7
de B. thuringiensis possam colonizar os cadáveres de diferentes espécies de insetos, 8
sendo este, o provável nicho da bactéria no ambiente (ALY; MULLA; FEDERICI, 9
1985; JARRETT; STEPHENSON, 1990; TAKATSUKA; KUNIMI, 2000; THOMAS et 10
al., 2000, 2001, 2002; SUZUKI et al., 2004; RAYMOND et al., 2010a, b). 11
B. cereus, assim como B. thuringiensis, é uma bactéria frequentemente 12
isolada a partir de amostras ambientais, incluindo solo, água e materiais de origem 13
vegetal e animal. No entanto, algumas linhagens de B. cereus causam contaminação de 14
alimentos sendo responsáveis por eventos de intoxicação gastrointestinal, produzindo 15
diarreias e vômitos. Além disso, linhagens de B. cereus tem sido apontadas como 16
responsáveis em casos de infecções locais ou sistêmicas (CALLEGAN; GILMORE; 17
GREGORY, 2007), o que aponta a espécie como patógeno oportunista. 18
B. thuringiensis e B. cereus compartilham um grande nível de 19
similaridade genética e são fenotipicamente idênticas, excetuando a produção de cristais 20
proteicos em B. thuringiensis. No entanto, o fato dos genes codificantes para a formação 21
desses cristais estarem frequentemente localizados em grandes plasmídeos conjugativos, 22
tem gerado discussões acerca da classificação como duas espécies separadas ou como 23
uma única espécie (HELGASON et al., 2000; VILAS-BÔAS et al., 2002; CHERIF et 24
al., 2003; VILAS-BÔAS et al., 2007), uma vez que a transferência de material genético 25
por conjugação entre linhagens de B. thuringiensis e entre linhagens de B. thuringiensis 26
e B. cereus, transferindo plasmídeos carregando genes cry, tem sido descrita 27
frequentemente (GONZÁLES; BROWN; CARLTON, 1982; JARRET; STEPHENSON 28
1990; ANDRUP; DAMGAARD; WASSERMANN, 1993; JENSEN et al., 1995; 29
VILAS-BÔAS et al., 1998, 2000; WILCKS et al., 1998; THOMAS et al., 2000, 2001, 30
2002; KLIER et al., 1983; LERECLUS et al., 1983; BATTISTI; GREEN; THORNE, 31

40
1985; REDDY et al., 1987; HU et al., 2004, 2005; VAN DER AUWERA et al., 2007; 1
YUAN et al., 2007). 2
Assim, uma linha de pesquisa importante, visa investigar as diferenças 3
de comportamento entre estas espécies em diferentes tipos de ambientes como solo, 4
cadáveres de insetos e alimentos (VILAS-BÔAS, 2000; SUZUKI, LERECLUS; 5
ARANTES, 2004; SANTOS, 2010; RAYMOND, 2012). Em condições favoráveis os 6
esporos de B. thuringiensis e B. cereus presentes no ambiente podem germinar, 7
seguindo-se a formação e multiplicação das células vegetativas, troca de material 8
genético por conjugação e posterior esporulação. No entanto, estudos têm mostrado que 9
os esporos de B. thuringiensis não podem germinar em amostras de solo esterilizado 10
(VILAS-BÔAS et al., 2000) e em ambientes aquáticos (FURLANETO et al., 2000) 11
devido a escassez de nutrientes e oxigênio. Diversos estudos foram realizados em 12
cadáveres de insetos para avaliar a germinação de esporos, multiplicação de células 13
vegetativas, produção de fatores de virulência e conjugação dos plasmídeos contendo 14
genes cry (JARRETT e STEPHENSON, 1990; VILAS-BÔAS et al., 1998; WILCKS et 15
al., 2008; YUAN et al., 2007; SANTOS et al., 2010). 16
Estudos demonstram a grande habilidade de disseminação de 17
plasmídeos entre diferentes espécies do grupo do B. cereus sensu lato (JARRETT; 18
STEPHENSON, 1990; VILAS-BOAS et al., 1998; SANTOS et al., 2010). Devido ao 19
mecanismo de conjugação os plasmídeos podem ser considerados como uma entidade 20
biológica autônoma, capaz de se transferir de uma célula bacteriana pra outra, assim um 21
plasmídeo como o pHT73 de B. thuringiensis não poderia ser considerado específico 22
para essa espécie bacteriana. Quando condições ambientais são desfavoráveis, a célula 23
bacteriana pode eliminar elementos extracromossômicos e com isso obter vantagem 24
seletiva. Dessa maneira são as trocas genéticas que fazem com que os plasmídeos sejam 25
mantidos nas populações bacterianas. 26
Santos e colaboradores (2010) demonstraram que a conjugação entre 27
linhagens de B. thuringiensis e B. cereus ocorre de maneira randômica, assim não 28
ocorre preferência na transferência de material genético entre essas duas espécies 29
bacterianas. No mesmo trabalho os autores evidenciaram que linhagens de B. 30
thuringiensis apresentaram maior germinação de esporos e multiplicação de células 31
vegetativas que linhagens de B. cereus contendo o plasmídeo com o gene cry. No 32
entanto, a comparação das habilidades de germinação de esporos e multiplicação de 33
células vegetativas de linhagens bacterianas contendo o plasmídeo com o gene cry com 34

41
as habilidades de linhagens sem o plasmídeo, demonstrou maior eficiência no 1
desenvolvimento nas linhagens contendo o plasmídeo. Portanto, esse estudo visa 2
verificar se as alterações verificadas no comportamento destas linhagens são decorrentes 3
à expressão do gene cry que está contido no plasmídeo ou a expressão de outro genes 4
plasmidiais ou da interação desses genes com o cromossomo. 5
6
3.3 MATERIAL E MÉTODOS 7
8
3.3.1 Linhagens utilizadas 9
10
As características fenotípicas das linhagens de B. thuringiensis 11
utilizadas neste estudo estão listadas na Tabela 1. B. thuringiensis subsp. kurstaki KT0 12
pHT73-EmR apresenta um plasmídeo natural (pHT73-Em
R) de 75 kb, contendo o gene 13
cry1Ac, no qual foi inserido um gene de resistência à eritromicina (ermC) (VILAS-14
BÔAS et al., 1998). B. thuringiensis subsp. kurstaki KT0 pHT73Δcry1Ac TetR contém o 15
mesmo plasmídeo, no entanto, o gene cry1Ac foi substituído por um gene de resistência 16
à tetraciclina (tetR). 17
18
3.3.2 Características dos experimentos de conjugação 19
20
As linhagens B. thuringiensis subsp. kurstaki KT0 pHT73-EmR e B. 21
thuringiensis subsp. kurstaki KT0 pHT73Δcry1Ac TetR foram usadas como doadoras 22
(Tabela 1). A linhagem B. thuringiensis subsp. thuringiensis 407-1 foi empregada como 23
receptora nos processos de conjugação. Os experimentos foram realizados com 24
repetições variadas, sendo que valores muito discrepantes, devido a erros experimentais, 25
foram eliminados. A presença dos plasmídeos pHT73-EmR e pHT73Δcry1Ac-Tet
R nos 26
exconjugantes foi confirmada através da produção de proteína cristal e teste de 27
resistência. 28
29
3.3.3 Conjugação em suporte sólido 30
31
Os experimentos de conjugação em suporte sólido foram conduzidos 32
pelo método descrito por Santos et al., (2010). As linhagens doadora e receptora foram 33

42
inoculadas individualmente em 5,0 mL de meio Luria-Bertani (LB) líquido, com os 1
antibióticos apropriados, e incubadas a 30 ºC por 15 h. Em seguida, 0,1 mL, de cada 2
linhagem, separadamente, foi transferido para 10 mL de meio LB pré-aquecido com 3
antibiótico e cultivado até D.O.600nm ~ 1,0. Igual concentração de linhagem doadora e 4
receptora (0,5 mL de cada linhagem) foi colocada sobre um filtro de nitrocelulose 5
(Millipore HVLP 0,45 µM), e este colocado sobre um disco de papel de filtro 6
depositado dentro de uma placa de Petri sem meio de cultura. Após a absorção do meio 7
de cultivo pelo papel de filtro, a membrana de nitrocelulose foi transferida para uma 8
placa de Petri contendo meio LB sem antibiótico, seguindo uma incubação a 30 ºC por 9
24 h. 10
As células que se multiplicaram sobre o filtro de nitrocelulose foram 11
ressuspendidas em 1,0 mL de solução salina e inoculadas em placas contendo meio LB 12
suplementado com os antibióticos adequados. Após 24 horas foi verificado o 13
aparecimento de colônias exconjugantes, as quais foram estocadas em banco seco e 14
glicerol 25%, após foram armazenadas adequadamente e utilizadas nos estudos in vivo 15
em larvas de Anticarsia gemmatalis Hübner, 1918, (Erebidae: Lepidoptera). 16
17
3.3.4 Produção da suspensão de esporos-cristais 18
19
Linhagens de B. thuringiensis estocadas em papel de filtro contendo 20
esporos e/ou cristais foram recuperadas após imersão dos papéis em 1 ml de água 21
destilada esterilizada seguida de breve agitação. Placas de Petri contendo meio Bacto-22
Peptona (BP) (LECADET et al., 1980) foram inoculadas com 0,2 ml da suspensão 23
bacteriana, sobre o meio de cultura com alça de Drygalski. 24
As placas foram incubadas em estufa a 30 0C até completa 25
esporulação, a qual foi monitorada em microscópio óptico, sendo que as lâminas 26
contendo uma suspensão da cultura são coradas por 70 segundos com Amido Black, 27
posteriormente são enxaguadas e coradas por 20 segundos com Fucsina (Figura 1). 28
Após quatro dias de incubação, as placas foram ressuspendidas em 1,5 mL de água 29
destilada esterilizada. 30
31
3.3.5 Comportamento de linhagens de Bacillus thuringiensis em 32
larvas de A. gemmatalis 33
34

43
Estudou-se o comportamento das linhagens contidas na Tabela 1 em 1
lagartas de 3º instar de A. gemmatalis, criadas em dieta com tetraciclina. Os testes foram 2
conduzidos usando seleção randômica das larvas com aproximadamente o mesmo 3
tamanho (sexo indefinido). As lagartas foram cedidas pela Empresa Brasileira de 4
Pesquisa Agropecuária (EMBRAPA-SOJA), situada na cidade de Londrina/PR, Brasil. 5
Nos experimentos foram utilizadas culturas esporuladas de todas as linhagens. As 6
culturas bacterianas foram tratadas com calor (80 °C, 20 minutos), visando a eliminação 7
de células vegetativas e a persistência de esporos. 8
Os experimentos foram realizados em folhas de Glycine max 9
(Linnaeus). As folhas foram coletadas, lavadas em solução de hipoclorito de sódio a 10
5%, enxaguadas em água destilada e secas em papel filtro. Posteriormente, foram 11
cortados discos foliares com um centímetro de diâmetro, os quais foram mergulhados 12
em solução de TWEEN 0,001% e receberam uma suspensão com 1011
esporos das 13
linhagens Cry- de B. thuringiensis (B. thuringiensis subsp. kurstaki KT0 14
pHT73Δcry1Ac TetR; B. thuringiensis subsp. thuringiensis 407-1; B. thuringiensis 15
subsp. thuringiensis 407-1 pHT73Δcry1Ac TetR) ou suspensão de esporos e cristais das 16
linhagens Cry+ de B. thuringiensis (B. thuringiensis subsp. kurstaki KT0 pHT73-Em
R e 17
B. thuringiensis subsp. thuringiensis 407-1 pHT73-EmR). 18
Para cada linhagem de B. thuringiensis citada anteriormente foram 19
utilizadas 40 lagartas de 2º instar de A. gemmatalis. Os experimentos foram realizados 20
em bandejas plásticas retangulares (42 x 19,5 cm) contendo 128 poços (Figura 2). As 21
lagartas foram colocadas individualmente em cada poço contendo um disco de folha de 22
soja com a suspensão de esporos ou esporos e cristais das linhagens utilizadas. As 23
bandejas plásticas foram colocadas em estufa com temperatura controlada (25-27 ºC), 24
fotoperíodo (16/8 h claro/escuro) e com 80% de umidade relativa. 25
Após 24 horas de incubação, um novo disco de folha de soja, sem 26
suspensão bacteriana, foi inserido em cada poço, para permitir a oferta de alimento às 27
lagartas por todo o experimento. Nesse momento, além de ser ofertado novo disco de 28
folha de soja, um total de 10 lagartas foram processadas imediatamente, para cada 29
linhagem bacteriana, visando a quantificação de células vegetativas e esporos em 30
lagartas vivas. Esse tempo foi denominado Tempo 1, ou tempo anterior a morte das 31
lagartas, sendo este 24 horas após o início do experimento. 32
Lagartas que foram alimentadas inicialmente com folhas contendo 33
linhagens produtoras de cristais foram incubadas até a morte, ocorrida pela ação 34

44
entomopatogênica da bactéria. Lagartas alimentadas inicialmente com folhas contendo 1
linhagens sem atividade entomopatogênica (Cry-) foram mortas mecanicamente, 2
simultaneamente à morte das lagartas pela atividade entomopatogênica. Nesse tempo, 3
denominado Tempo 2, ou tempo de morte das lagartas, para cada linhagem, um lote de 4
10 lagartas foi processado imediatamente, e outros dois lotes de 10 lagartas foram 5
armazenados em tubos de microcentrífuga, os quais foram incubados a 30 ºC por 24 6
horas (Tempo 3) ou 48 horas (Tempo 4) e, então, processados. 7
Dessa forma, para cada linhagem, o processamento das lagartas para 8
quantificação de células vegetativas e esporos bacterianos foi realizado em quatro lotes 9
(Tempo 1, anterior a morte das lagartas, Tempo 2, tempo de morte das lagartas 10
(aproximadamente 48 horas após o início do experimento), Tempo 3, 24 horas após a 11
morte das lagartas e Tempo 4, 48 horas após a morte das lagartas), com 10 lagartas por 12
lote, totalizando 40 lagartas. 13
Cada experimento foi realizado com um conjunto de linhagens. Assim, 14
as duas linhagens B. thuringiensis subsp. kurstaki KT0 foram avaliadas sempre na 15
mesma bateria experimental. Da mesma forma, os experimentos com as três linhagens 16
B. thuringiensis subsp. thuringiensis 407-1 sempre foram realizados na mesma bateria 17
experimental. Durante cada experimento foi realizado um controle negativo contendo 18
10 lagartas. Dessa forma, B. thuringiensis subsp. kurstaki KT0 pHT73Δcry1Ac TetR e 19
B. thuringiensis subsp. kurstaki KT0 pHT73-EmR, possuem 40 lagartas cada e 10 20
lagartas como controle negativo e B. thuringiensis subsp. thuringiensis 407-1; B. 21
thuringiensis subsp. thuringiensis 407-1 pHT73-EmR e B. thuringiensis subsp. 22
thuringiensis 407-1 pHT73Δcry1Ac TetR, possuem 40 lagartas cada e 10 lagartas como 23
controle negativo. 24
Para a quantificação do número de unidades formadoras de colônias 25
(UFC) e esporos bacterianos, em cada tempo, as lagartas foram maceradas em 1 mL de 26
solução salina 0,85% em tubos de microcentrífuga de 2,0 mL, seguindo-se diluição em 27
série em solução salina. Cada diluição foi inoculada em placas contendo meio LB 28
suplementado com os antibióticos apropriados para cada linhagem (100 µg mL-1
de 29
eritromicina; 200 µg mL-1
de estreptomicina e 10 µg mL-1
de tetraciclina), antes e após 30
tratamento de calor (80 ºC por 20 min). As placas foram incubadas a 30 ºC por cerca de 31
16 horas até a formação de colônias. 32
33
3.3.6 Análise dos resultados 34

45
1
A análise de variância (teste de Tukey) em p <0,05, realizada pelo 2
programa Instat, foi utilizada para estabelecer se existe diferença significativa na 3
germinação de esporos, multiplicação das células vegetativas e esporulação das 4
linhagens de B. thuringiensis em larvas de A. gemmatalis. Para a comparação entre o 5
comportamento das diferentes linhagens foi utilizado um modelo binomial negativo. 6
7
3.4 RESULTADOS 8
9
Foram obtidos exconjugantes da linhagem B. thuringiensis subsp. 10
thuringiensis 407-1 com duas linhagens doadoras, B. thuringiensis subsp. kurstaki 11
KT0 pHT 73-EmR
e B. thuringiensis subsp. kurstaki KT0 pHT73Δcry1Ac Tet
R, 12
através da incubação conjunta em suporte sólido de células vegetativas de ambas 13
linhagens. Assim, um total de 5 linhagens foi utilizado nos experimentos de estudo do 14
comportamento das linhagens de B. thuringiensis em larvas de A. gemmatalis. 15
As linhagens com atividade entomopatogênica levaram o inseto à 16
morte aproximadamente 48 horas após o início do experimento, o que foi considerado 17
como t0 nesse estudo. As demais avaliações foram realizadas, 24 horas após o início do 18
experimento (tempo antes da morte) e com 24 e 48 horas após a morte do inseto (Figura 19
3). 20
As diversas linhagens bacterianas apresentaram diferentes 21
comportamentos em larvas de A. gemmatalis (Figura 4). A linhagem selvagem B. 22
thuringiensis subsp. kurstaki KT0 pHT73-EmR (Figura 4A) no tempo antes da morte 23
apresentou 106 esporos no interior da lagarta, não ocorrendo germinação nesse tempo; a 24
germinação ocorreu entre o t0 e t24 (p>0,05), sendo esta foi completa no t24. A 25
multiplicação das células vegetativas ocorreu entre os tempos t0 e t24 (p>0,05), seguindo 26
para um início de esporulação entre os tempos t24 e t48 (p>0,05). 27
A linhagem B. thuringiensis subsp. kurstaki KT0 pHT73Δcry1Ac 28
TetR (Figura 4B) no tempo antes da morte possuía 10
5 esporos no interior da lagarta, 29
sem ocorrer germinação dos esporos nesse tempo. Entre os tempos antes da morte e t0 30
ocorreu a germinação dos esporos (p>0,05), sendo que esta foi completa no t0 (neste 31
tempo não encontrou-se esporos). Não ocorreu multiplicação das células vegetativas 32
significativa em nenhum dos tempos analisados. 33
A linhagem B. thuringiensis subsp. thuringiensis 407-1 (Figura 4C), 34

46
não apresentou completa germinação dos esporos, a germinação parcial ocorreu entre t0 1
e t24 (p>0,05). Ocorreu multiplicação das células vegetativas entre t24 e t48 (p>0,05), 2
porém aproximadamente 102 das formas presentes permaneceram na forma de esporos 3
no decorrer do experimento. 4
B. thuringiensis var. thuringiensis 407-1 pHT73-EmR (Figura 4D) 5
apresentou germinação dos esporos no tempo antes da morte, porém em t0 ocorreu 6
esporulação em aproximadamente 105, a multiplicação da células vegetativas ocorreu 7
entre t0 e t24 (p>0,05). B. thuringiensis subsp. thuringiensis 407-1 pHT73Δcry1Ac-TetR 8
(Figura 4E) apresentou completa germinação dos esporos no tempo antes da morte, 9
ocorreu multiplicação das células vegetativas entre t0 e t48 (p>0,05) e entre t24 e t48 10
(p>0,05) ocorreu início de esporulação. 11
12
3.5 DISCUSSÃO 13
14
O desenvolvimento de espécies do grupo do B. cereus sensu lato em 15
insetos hospedeiros tem sido estudado por muitos pesquisadores, tentando compreender 16
o dinâmico comportamento desse grupo bacteriano (SALAMITOU et al., 2000; 17
TAKATSUKA; KUNIMI, 2000; THOMAS et al., 2000, 2001, 2002; YUAN et al., 18
2007; SANTOS et al., 2010; RAYMOND et al., 2012). 19
A linhagem selvagem B. thuringiensis subsp. kurstaki KT0 pHT73-20
EmR mostrou completa germinação dos esporos no t24, como descrito por Santos e 21
colaboradores (2010) onde essa mesma linhagem provocou a morte do inseto 22
hospedeiro e foi capaz de colonizar larvas de B. mori, apresentando alta multiplicação 23
de células vegetativas. A linhagem B. thuringiensis subsp. kurstaki KT0 contendo o 24
mesmo plasmídeo com o gene cry deletado (pHT73Δcry1Ac TetR) não conseguiu se 25
desenvolver no interior da lagarta, apresentando somente a germinação dos esporos, não 26
ocorrendo multiplicação das células vegetativas e posterior esporulação como a 27
linhagem B. thuringiensis subsp. kurstaki KT0 pHT73-EmR. 28
Através dos tempos citados pode-se observar a dinâmica do 29
comportamento de linhagens de B. thuringiensis em larvas de insetos. Os cristais 30
proteicos ao serem ingeridos, causaram danos na membrana epitelial do intestino larval, 31
com isso os esporos puderam invadir a hemolinfa e então germinar; consequentemente 32
pôde ocorrer multiplicação das células vegetativas. Com base nesses conceitos foram 33
realizadas análises de contagem no tempo antes da morte das largartas, para avaliar se 34

47
existe correlação entre o comportamento das linhagens e a morte do inseto, ou seja, se a 1
germinação e/ou multiplicação das linhagens é influenciada pela morte do inseto. 2
Através dessa análise pôde-se avaliar se a germinação ocorre em larvas de insetos antes 3
da morte. Escolheu-se o modelo binomial negativo utilizado para comparar o 4
comportamento das diferentes linhagens no interior do inseto hospedeiro, pois, este 5
permite incorporar o excesso de variância nos dados experimentais. 6
O modelo binomial negativo permitiu a comparação entre essas duas 7
linhagens, analisando dados anteriores ao tratamento de calor, ou seja, considerando o 8
número total da população bacteriana. Esse modelo permitiu concluir que o ambiente 9
larval foi mais favorável a linhagem B. thuringiensis subsp. kurstaki KT0 pHT73-EmR 10
no decorrer do tempo quando comparada a linhagem B. thuringiensis subsp. kurstaki 11
KT0 pHT73Δcry1Ac TetR (Figura 5A). Verificou-se que a linhagem B. thuringiensis 12
subsp. kurstaki KT0 pHT73-EmR
apresentou crescimento exponencial com taxa 1,3285 13
e a linhagem e KT0 pHT73Δcry1Ac TetR apresentou um crescimento menor, com taxa 14
de 0,6598, mostrando que o crescimento da linhagem B. thuringiensis subsp. kurstaki 15
KT0 pHT73-EmR
foi significativamente maior que o da linhagem KT0 pHT73Δcry1Ac 16
TetR
(Figura 5A). 17
Para as mesmas duas linhagens testou-se o efeito do calor no t0, onde 18
dados obtidos antes do tratamento de calor foram contrastados com dados obtidos 19
posteriores ao tratamento de calor. As diferenças para ambas as linhagens foram 20
significativas, ou seja, o calor tem efeito estatisticamente igual nas duas espécies, sendo 21
eficaz na eliminação de células vegetativas. Verificou-se o efeito do calor no tempo 22
antes da morte, evidenciando para ambas as espécies que não houve diferenças 23
estatísticas, com isso pode-se concluir que a população bacteriana total encontrava-se na 24
forma de esporos no tempo antes da morte, não ocorrendo germinação dos esporos 25
nesse tempo. 26
As análises das linhagens B. thuringiensis subsp. thuringiensis 407-1; 27
B. thuringiensis subsp. thuringiensis 407-1 pHT73-EmR e B. thuringiensis subsp. 28
thuringiensis 407-1 pHT73Δcry1Ac TetR utilizando dados antes do calor permitiram a 29
comparação do comportamento dessas linhagens no decorrer do tempo. As bactérias B. 30
thuringiensis subsp. thuringiensis 407-1; B. thuringiensis subsp. thuringiensis 407-1 31
pHT73Δcry1Ac TetR apresentam um crescimento exponencial com taxas 32
estatisticamente iguais (3,047 e 3,043, respectivamente) sendo que a bactéria B. 33
thuringiensis subsp. thuringiensis 407-1 pHT73-EmR apresentou crescimento 34

48
significativamente maior (4,198) (Figura 5B). 1
Para as mesmas três linhagens testou-se o efeito do calor no t0, onde 2
dados antes do tratamento de calor foram contrastados com dados posteriores ao 3
tratamento de calor. Com os resultados pode-se observar que o calor tem efeito mais 4
pronunciado nas bactérias B. thuringiensis subsp. thuringiensis 407-1 pHT73-EmR e B. 5
thuringiensis subsp. thuringiensis 407-1 pHT73Δcry1Ac TetR, portanto essas linhagens 6
possuíam um maior número de células vegetativas que a linhagem B. thuringiensis 7
subsp. thuringiensis 407-1. Análises estatísticas testando o efeito do calor, no tempo 8
antes da morte, verificaram que a população de esporos foi maior nas bactérias B. 9
thuringiensis subsp. thuringiensis 407-1 e B. thuringiensis subsp. thuringiensis 407-1 10
pHT73Δcry1Ac TetR, com isso evidencia-se que linhagem B. thuringiensis subsp. 11
thuringiensis 407-1 pHT73-EmR possui uma quantidade maior de células vegetativas 12
no tempo antes da morte. As diferenças para ambas as linhagens foram significativas, 13
ou seja, o calor tem efeito estatisticamente igual nas duas espécies. 14
Através do experimento constatou-se que as bactérias que continham 15
o plasmídeo pTH73-EmR, se multiplicaram em uma taxa maior que as bactérias com o 16
plasmídeo pHT73Δcry1Ac TetR, ou aquelas que não possuíam o plasmídeo (selvagem). 17
Portanto o plasmídeo pTH73-EmR
conferiu vantagem as bactérias que o possuíam no 18
ambiente larval, pois este foi mais favorável a essas bactérias. A análise das bactérias B. 19
thuringiensis subsp. thuringiensis 407-1; B. thuringiensis subsp. thuringiensis 407-1 20
pTH73-EmR e B. thuringiensis subsp. thuringiensis 407-1 pHT73Δcry1Ac Tet
R, 21
permitiu concluir que o gene cry, contido no plasmídeo pTH73-EmR, também 22
proporciona vantagem para as bactérias que o abrigam, pois a linhagem B. 23
thuringiensis subsp. thuringiensis 407-1 pTH73-EmR multiplicou-se de maneira mais 24
eficiente no interior do inseto do que a linhagem B. thuringiensis subsp. thuringiensis 25
407-1 pHT73Δcry1Ac TetR que possui o gene cry deletado, constatando assim a 26
vantagem do gene cry. Santos e colaboradores (2010) avaliaram o comportamento da 27
linhagem B. thuringiensis subsp. thuringiensis 407-1 selvagem, em larvas de 2º instar 28
de B. mori e obtiveram multiplicação das células vegetativas em altas taxas, a linhagem 29
exconjugante com o plasmídeo pHT73-EmR
não foi avaliada. 30
No entanto, estudos realizados por outros autores, como Raymond e 31
colaboradores (2012) e Takatsuka e Kunimi, (2000), relatam que linhagens sem 32
plasmídeos se multiplicam mais rapidamente que as linhagens com grandes plasmídeos 33
como o pHT73-EmR, assim as linhagens pertencentes a microbiota intestinal podem se 34

49
multiplicar mais rápido e gerar uma interação antagonística, impossibilitando o 1
desenvolvimento das linhagens com grandes plasmídeos. 2
Além disso, Suzuki e colaboradores (2004) concluíram que a 3
habilidade de linhagens de B. thuringiensis crescerem e esporularem em cadáveres de 4
insetos é variável. Esses autores observaram que linhagens de B. thuringiensis são 5
capazes de matar o inseto, mas nem sempre se multiplicam nos cadáveres. Algumas 6
linhagens sem especificidade para matar um hospedeiro foram capazes de se multiplicar 7
quando estes foram mortos mecanicamente. Linhagens acristalíferas de B. thuringiensis 8
var. israelenses mantém a habilidade para multiplicar em larvas de Diptera (SUZUKI et 9
al., 2004). Thomas et al. (2000) descreveram o mesmo fenômeno no coleóptera 10
Phaedon cochleriae com a linhagem B. thuringiensis var. tenebrionis. 11
Estudos posteriores poderão evidenciar se a influência do plasmídeo 12
pHT73 e do gene cry1Ac sobre o comportamento de linhagens de B. thuringiensis é 13
espécie específico ou se a influência ocorre também em linhagens de B. cereus. O 14
trabalho terá continuidade com análises de outras bactérias do Grupo do B. cereus, 15
incluindo linhagens selvagens de B. cereus sensu stricto, contendo o plasmídeo pHT73-16
EmR ou o plasmídeo pHT73Δcry1Ac Tet
R, para a confirmação dos resultados obtidos 17
em linhagens de B. thuringiensis e uma maior amplitude do papel desses plasmídeos ou 18
do gene cry nesse grupo bacteriano. Em acréscimo, os resultados obtidos também 19
devem levar a caracterização de genes plasmídicos envolvidos no desenvolvimento de 20
bactérias em insetos. Os resultados obtidos permitirão um aprimoramento nos 21
conhecimentos acerca de evolução das bactérias do grupo do B. cereus. No conjunto, 22
os resultados poderão fornecer dados essenciais sobre a evolução de bactérias do grupo 23
B. cereus, como a compreensão do papel de plasmídeos conjuntivos e de seus genes no 24
comportamento de linhagens do grupo do B. cereus em ecossistemas naturais, como o 25
cadáver de invertebrados. 26
27
28
29
30
31
32
33
34

50
1
2
REFERÊNCIAS 3
4
ALY, C.; MULLA, M. S.; FEDERICI, B. A. Sporulation and toxin production by 5
Bacillus thuringiensis var. israelensis in cadavers of mosquito larvae. Journal of 6
Invertebrate Pathology.v. 46, n. 3, p. 251–258, 1985. 7
8
ANDRUP, L.; DAMGAARD, J.; WASSERMANN, K. Mobilization of small plasmids 9
in Bacillus thuringiensis subsp. israelensis is accompanied by specific aggregation. 10
Journal of Bacteriology. v. 175, n. 20, p. 6530–6536, 1993. 11
12
BATTISTI, L.; GREEN, B.; THORNE, C. Mating system for transfer of plasmids 13
among Bacillus anthracis, Bacillus cereus, and Bacillus thuringiensis. Journal of 14
Bacteriology. v. 162, n. 2, p. 543–550, 1985. 15
16
CALLEGAN, M. C.; GILMORE, M. S.; GREGORY, M. Bacterial endophthalmitis: 17
therapeutic challenges and host-pathogen interactions. Progress in Retinal and Eye 18
Research. v. 26, p.189–203, 2007. 19
20
CHERIF, A.; BRUSETTI, L.; BORIN, S.; RIZZI, A.; BOUDABOUS, A.; HYAMI-21
HORANI, H. DAFFONCHIO, D. Genetic relationshio in the “Bacillus cereus group” 22
by rep-PCR fingerprint and sequencing of Bacillus anthracis- specific rep-PCR 23
fragment. Journal Applied Microbiology. v. 94, p. 1108-1119, 2003. 24
25
FURLANETO, L. M.; SARIDAKIS, H. O.; ARANTES, O. M. N. Survival and 26
conjugal transfer between Bacillus thuringiensis strains in aquatic environment. 27
Brazilian Journal of Microbiology. v. 31, n. 4, p. 233-238, 2000. 28
29
GONZÁLES, J. M. Jr.; BROWN, B. S.; CARLTON, B. C. Transfer of B. thuringiensis 30
plasmids coding for d-endotoxin among strains of B. thuringiensis and B. cereus. 31
Proceedings of the National Academic of Science. v. 79, n. 10, p. 6951-6955, 1982. 32
33
HELGASON, E.; ØKSTAD, P. A.; CAUGANT, D. A.; JOHANSEN, H. A.; FOUET, 34
A.; MOCK, M.; HEGNA, I.; KØLSTO, A. B. Bacillus anthracis, Bacillus cereus, 35
Bacillus thuringiensis - One species on the basis of genetic evidence. Applied and 36
Environmental Microbiology. v. 66, n. 6, p. 2627-2630, 2000. 37
38
HU, X.; HANSEN, B.M.; EILENBERG, J.; HENDRIKSEN, N. B.; SMIDT, L.; 39
YUAN, Z.; JENSEN, G.B. Conjugative transfer, stability and expression of a plasmid 40
encoding a cry1Ac gene in Bacillus cereus group strains. FEMS Microbiology. v. 231, 41
p. 45–52, 2004. 42
43
JARRETT, P.; STEPHENSON, M. Plasmid transfer between strains of Bacillus 44
thuringiensis infecting Galleria mellonella and Spodoptera littoralis. Applied and 45
Environmental Microbiology. v. 56, p. 1608–1614, 1990. 46
47

51
JENSEN, G.B.; WILCKS, A.; PETERSEN, S. S.; DAMGAARD, J.; BAUM, J.A.; 1
ANDRUP, L. The genetic basis of the aggregation system in Bacillus thuringiensis 2
subsp. israelensis is located on the large conjugative plasmid pXO16. Journal 3
Bacteriology. v. 177, p. 2914–2917, 1995. 4
5
KLIER, A.; BOURGOUIN, C .; RAPOPORT, G. Mating between Bacillus subtilis and 6
Bacillus thuringiensis and transfer of cloned crystal genes. Molecular and General 7
Genetics, v. 191, p. 257-262, 1983. 8
9
LERECLUS, D.; MENOU, G.; LECADET, M. M. Isolation of a DNA sequence related 10
to several plasmids from B. thuringiensis after a mating involving the Streptoccus 11
faecalis plasmid pAMb1. Molecular Genetics e Genomics. v. 191, p. 307–313, 1983. 12
13
RAYMOND, B.; WYRES, K. L.; SHEPPARD, S. K.; ELLIS, R. E.; BONSALL, M. B. 14
Environmental Factors Determining the Epidemiology and Population Genetic Structure 15
of the Bacillus cereus Group in the Field. PLoS Pathogens. v. 26, n. 6(5), e1000905, 16
2010a. 17
18
RAYMOND, B.; JOHNSTON, P. R.; NIELSEN-LEROUX, C.; LERECLUS, D.; 19
CRICKMORE, N. Bacillus thuringiensis: an impotent pathogen? Trends in 20
Microbiology. v. 18, n. 5, p. 183-232, 2010b. 21
22
RAYMOND, B.; WEST, S. A.; GRIFFIN, A. S.; BONSALL, M. B. The Dynamics of 23
Cooperative Bacterial Virulence in the Field. Science. v. 337, n. 6090, p. 85-88, 2012. 24
25
REDDY, A.; BATTIST, L.; THORNE, C.B. Identification of self-transmissible 26
plasmids in four Bacillus thuringiensis subspecies. Journal Bacteriology. v. 169, p. 27
5263–5270, 1987. 28
29
SALAMITOU, S.; RAMISSE, F.; BREHÉLIN, M.; BOURGUET, D.; GILOIS, 30
N.; GOMINET, M.; HERNANDEZ, E.; LERECLUS, D. The plcR regulon is involved 31
in the opportunistic properties of Bacillus thuringiensis and Bacillus cereus in mice and 32
insects. Microbiology. v. 146, n. 11, p. 2825-2832, 2000. 33
34
SANTOS, C.; VILAS-BÔAS, G. T.; LERECLUS, D.; SUZUKI, M. T.; ANGELO, E. 35
A.; ARANTES, O. M. N. Conjugal transfer between Bacillus thuringiensis and Bacillus 36
cereus strains is not directly correlated with growth of recipient strains. Journal of 37
Invertebrate Pathology. v. 105, n. 2, p.171–175, 2010. 38
39
SCHNEPF, E.; CRICKMORE, N.; VAN RIE, J.; LERECLUS, D.; BAUM, J.; 40
FEITELSON, J.; ZEIGLER, D. R.; DEAN, D. H. B. thuringiensis and its pesticidal 41
crystal proteins. Microbiology and Molecular Biology Reviews. v. 62, n. 3, p. 775-42
806, 1998. 43
44
SUZUKI, M. T.; LERECLUS, D. ARANTES, O. M. Fate os B. thuringiensis strains in 45
different insect larvae. Canadian Journal of Microbiology. v. 50, n. 11, p. 973-975, 46
2004. 47
48
TAKATSUKA, J.; KUNIMI, Y. Intestinal bacteria affect growth of Bacillus 49
thuringiensis in larvae of the oriental tea tortrix, Homona magnanima diakonoff 50

52
(Lepidoptera: tortricidae). Journal of Invertebrate Pathology. v. 73, n.3, p. 222-1
226, 2000. 2
3
THOMAS, D. J. I.; MORGAN, J. A. W.; WHIPPS, J. M.; SAUNDERS, J. R. Plasmid 4
transfer between the Bacillus thuringiensis subspecies kurstaki and tenebrionis in 5
laboratory culture and soil and in Lepidopteran and Coleopteran larvae. Applied and 6
Environmental Microbiology. v. 66, p. 118–124, 2000. 7
8
THOMAS, D. J. I.; MORGAN, J. A. W.; WHIPPS, J. M.; SAUNDERS, J. R. Plasmid 9
transfer between the Bacillus thuringiensis subspecies. israelensis strains in laboratory 10
culture, river water, and dipteran larvae. Applied and Environmental Microbiology. 11
v. 67, p. 330–338, 2001. 12
13
THOMAS, D. J. I.; MORGAN, J. A. W.; WHIPPS, J. M.; SAUNDERS, J. R. Transfer 14
of plasmid pBC16 between Bacillus thuringiensis strains in non-susceptible larval. 15
FEMS Microbiology. v. 40, p.181–190, 2002. 16
17
VAN DER AUWERA, G.A.; TIMMERY, S.; HOTON, F.; MAHILLON, J. Plasmid 18
exchanges among members of the Bacillus cereus group in foodstuffs. International 19
Journal of Food Microbiology. v. 113, p. 164–172, 2007. 20
21
VILAS-BÔAS, G. F. L. T.; VILAS-BÔAS, L. A.; LERECLUS, D.; ARANTES, O. M. 22
N. Bacillus thuringiensis conjugation under environmental conditions. FEMS 23
Microbiology. v. 25, p. 369–374, 1998. 24
25
VILAS-BÔAS, L. A.; VILAS-BÔAS, G. F. L. T.; SARIDAKIS, H. O.; LEMOS, 26
M.V.F.; LERECLUS, D.; ARANTES, O. M. N. Survival and conjugation of Bacillus 27
thuringiensis in a soil microcosm. FEMS Microbiology Ecology, v.31, n. 3, p.255-259, 28
2000. 29
30
VILAS-BÔAS, G.; SANCHIS, V.; LERECLUS, D.; LEMOS, M.; BOURGUET, D. 31
Genetic differentiation between sympatric populations of Bacillus cereus and Bacillus 32
thuringiensis. Applied and Environmental Microbiology. v. 68, p. 1414–1424, 2002. 33
34
VILAS-BÔAS, G. T.; PERUCA, A. P. S.; ARANTES, O. M. N. Biology and taxonomy 35
of Bacillus cereus, Bacillus anthracis, and Bacillus thuringiensis. Canadian Journal of 36
Microbiology. v. 53, n. 6, p. 673-687, 2007. 37
38
WILCKS, A.; JAYASWAL, N.; LERECLUS, D.; ANDRUP, L. Characterization of 39
plasmid pAW63, a second self-transmissible plasmid in Bacillus thuringiensis subsp. 40
kurstaki HD73. Microbiology. v. 144, p. 1263–1270, 1998. 41
42
WILCKS, A.; SMIDT, L.; BAHL, M. I.; HANSEN, B. M.; ANDRUP, L.; 43
HENDRIKSEN, N. B.; LICHT, T. R. Germination and conjugation of Bacillus 44
thuringiensissubsp. israelensis in the intestine of gnotobiotic rats. Journal Applied of 45
Microbiology. v. 104, n. 5, p. 1252–1259, 2008. 46
47
YUAN, Y. M.; HU, X. M.; LIU, H. Z.; HANSEN, B. M.; YAN, J. P.; YUAN, Z. M. 48
Kinetics of plasmid transfer among Bacillus cereus group strains within lepidopteran 49
larvae. Archives of Microbiology. v. 187, n. 6, p. 425–431, 2007. 50

53
ANEXOS
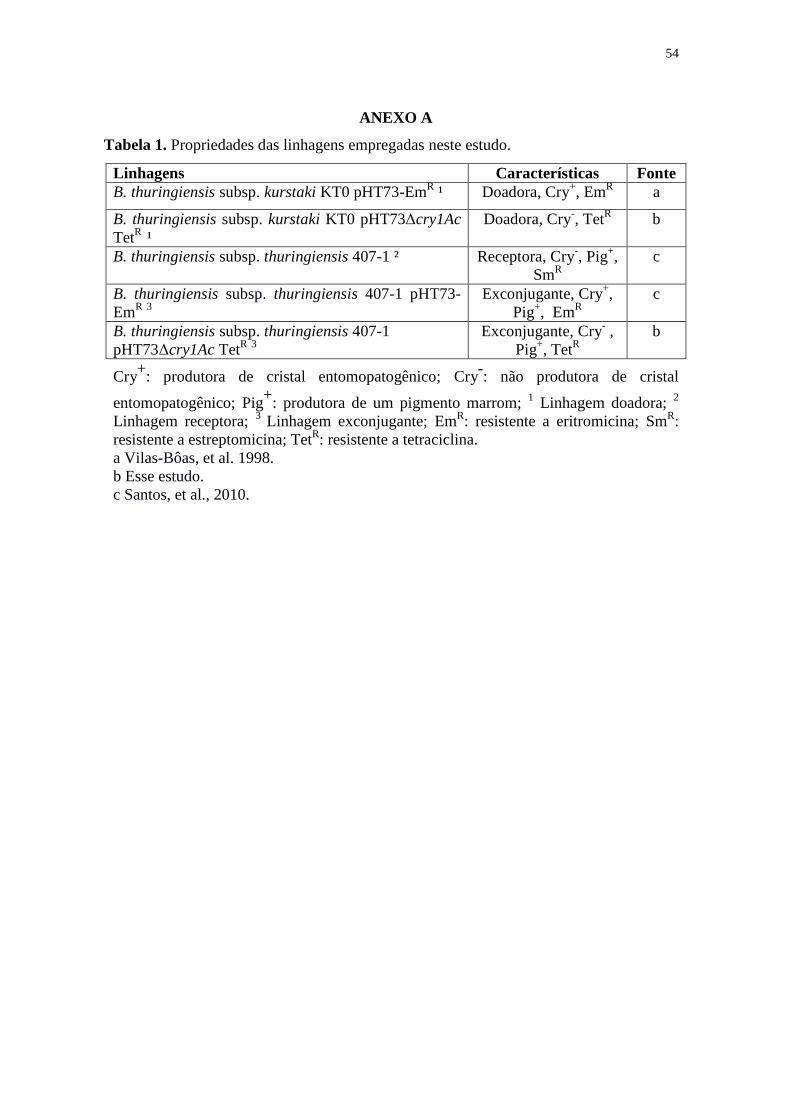
54
ANEXO A
Tabela 1. Propriedades das linhagens empregadas neste estudo.
Linhagens Características Fonte
B. thuringiensis subsp. kurstaki KT0 pHT73-EmR ¹ Doadora, Cry
+, Em
R a
B. thuringiensis subsp. kurstaki KT0 pHT73Δcry1Ac
TetR
¹
Doadora, Cry-, Tet
R b
B. thuringiensis subsp. thuringiensis 407-1 ² Receptora, Cry-, Pig
+,
SmR
c
B. thuringiensis subsp. thuringiensis 407-1 pHT73-
EmR 3
Exconjugante, Cry+,
Pig+, Em
R
c
B. thuringiensis subsp. thuringiensis 407-1
pHT73Δcry1Ac TetR 3
Exconjugante, Cry- ,
Pig+, Tet
R
b
Cry+
: produtora de cristal entomopatogênico; Cry-: não produtora de cristal
entomopatogênico; Pig+
: produtora de um pigmento marrom; 1 Linhagem doadora;
2
Linhagem receptora; 3
Linhagem exconjugante; EmR: resistente a eritromicina; Sm
R:
resistente a estreptomicina; TetR: resistente a tetraciclina.
a Vilas-Bôas, et al. 1998.
b Esse estudo.
c Santos, et al., 2010.
55
ANEXO B
Figura 1. Fotomicroscopia das linhagens (A) B. thuringiensis subsp. kurstaki KT0 pHT73-EmR
evidenciando os cristais bipiramidais (c) e esporos (e) e (B) linhagem B. thuringiensis subsp.
kurstaki KT0 pHT73Δcry1Ac TetR, contendo apenas esporos (e). Aumento de 1000 vezes e
coloração: Amido-Black e Fucsina.

56
ANEXO C
Figura 2. Etapas de montagem do experimento. (A) Visão geral da bandeja plástica com os
discos foliares; (B) Lagarta de A. gemmatalis sendo inserida individualmente no poço; (C) Final
da montagem do experimento com a vedação da bandeja plástica com um selo adesivo a camada
plástico.

57
ANEXO D
Figura 3. Etapas de infecção pela linhagem B. thuringiensis subsp. kurstaki KT0
pHT73-EmR e exconjugantes com o plasmídeo pHT73-Em
R em A. gemmatalis. (A)
antes do tratamento; (B) logo após morte (t0); (C) 12 horas após a morte (t12). Seta:
início da infecção de B. thuringiensis no intestino médio de A. gemmatalis.
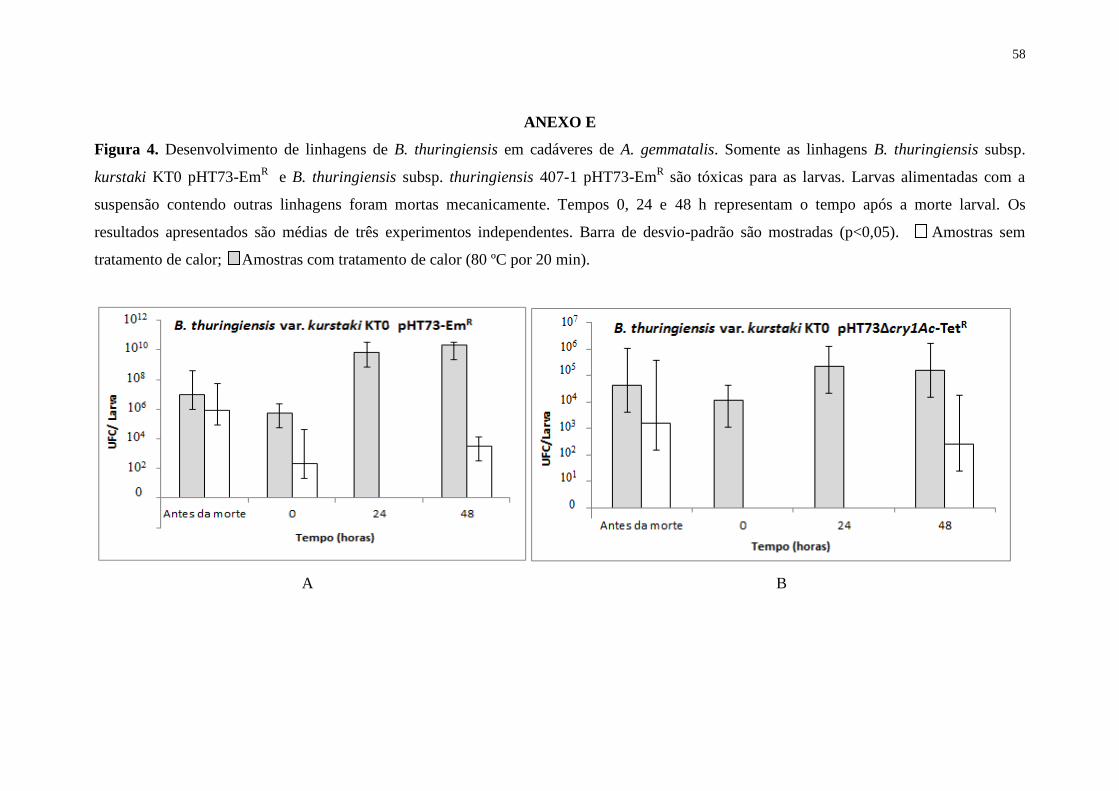
58
ANEXO E
Figura 4. Desenvolvimento de linhagens de B. thuringiensis em cadáveres de A. gemmatalis. Somente as linhagens B. thuringiensis subsp.
kurstaki KT0 pHT73-EmR
e B. thuringiensis subsp. thuringiensis 407-1 pHT73-EmR são tóxicas para as larvas. Larvas alimentadas com a
suspensão contendo outras linhagens foram mortas mecanicamente. Tempos 0, 24 e 48 h representam o tempo após a morte larval. Os
resultados apresentados são médias de três experimentos independentes. Barra de desvio-padrão são mostradas (p<0,05). Amostras sem
tratamento de calor; Amostras com tratamento de calor (80 ºC por 20 min).
A B
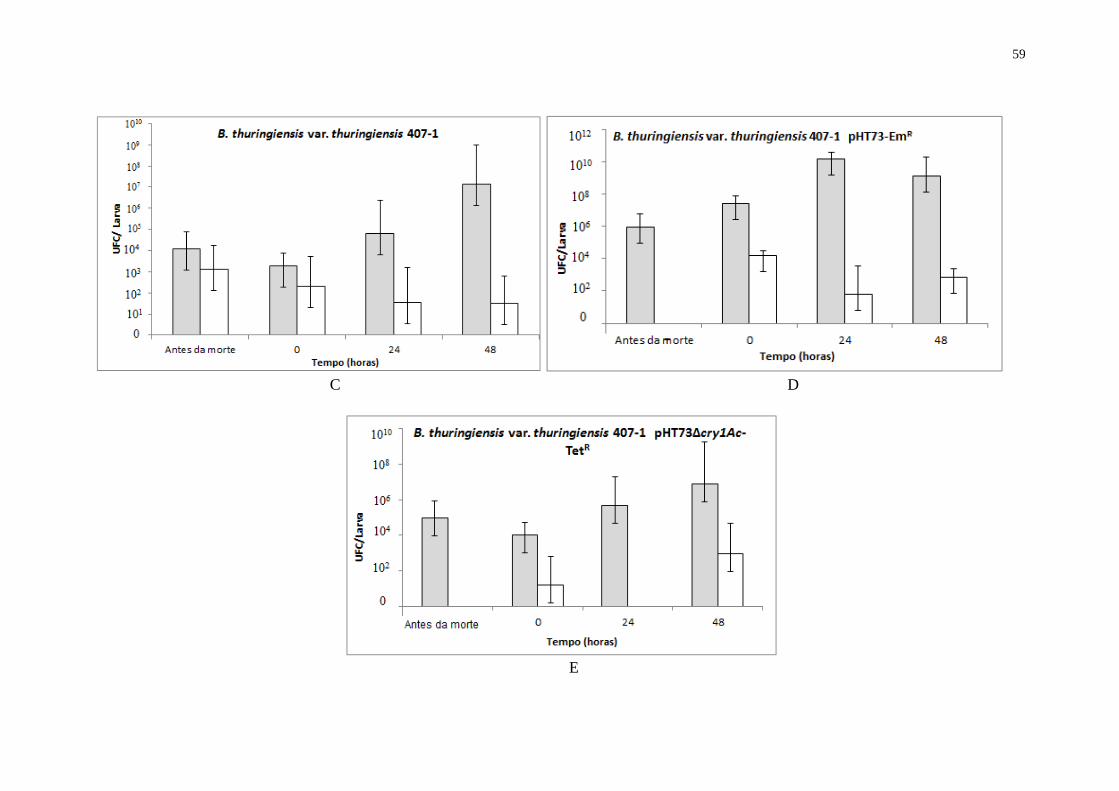
59
C D
E

60
ANEXO F
Figura 5. Modelo de comportamento temporal de linhagens de B. thuringiensis em cadáveres de A. gemmatalis. Somente as linhagens B.
thuringiensis subsp. kurstaki KT0 pHT73-EmR
e B. thuringiensis subsp. thuringiensis 407-1 pHT73-EmR são tóxicas para as larvas. Valores
plotados correspondem aos anteriores ao tratamento de calor. Tempos 0, 24 e 48 representam o tempo após a morte larval. A) Valores
obsevados; Valores médios;
Linha de tendência da linhagem B. thuringiensis subsp. kurstaki KT0 pHT73-EmR
(inclinação da reta:
1,3285); Valores observados; Valores médios; Linha de tendência da linhagem da linhagem B. thuringiensis subsp. kurstaki KT0
pHT73Δcry1Ac TetR
(inclinação da reta: 0,6598); B) Valores observados; Valores médios;
Linha de tendência da linhagem B.
thuringiensis subsp. thuringiensis 407-1 (inclinação da reta: 3,047); Valores observados; Valores médios; Linha de tendência da
linhagem B. thuringiensis subsp. thuringiensis 407-1 pHT73-EmR
(inclinação da reta: 4,198); Valores observados;
Valores médios;
pppLinha de tendência das médias da linhagem B. thuringiensis subsp. thuringiensis 407-1 pHT73Δcry1Ac TetR; (inclinação da reta: 3,043);
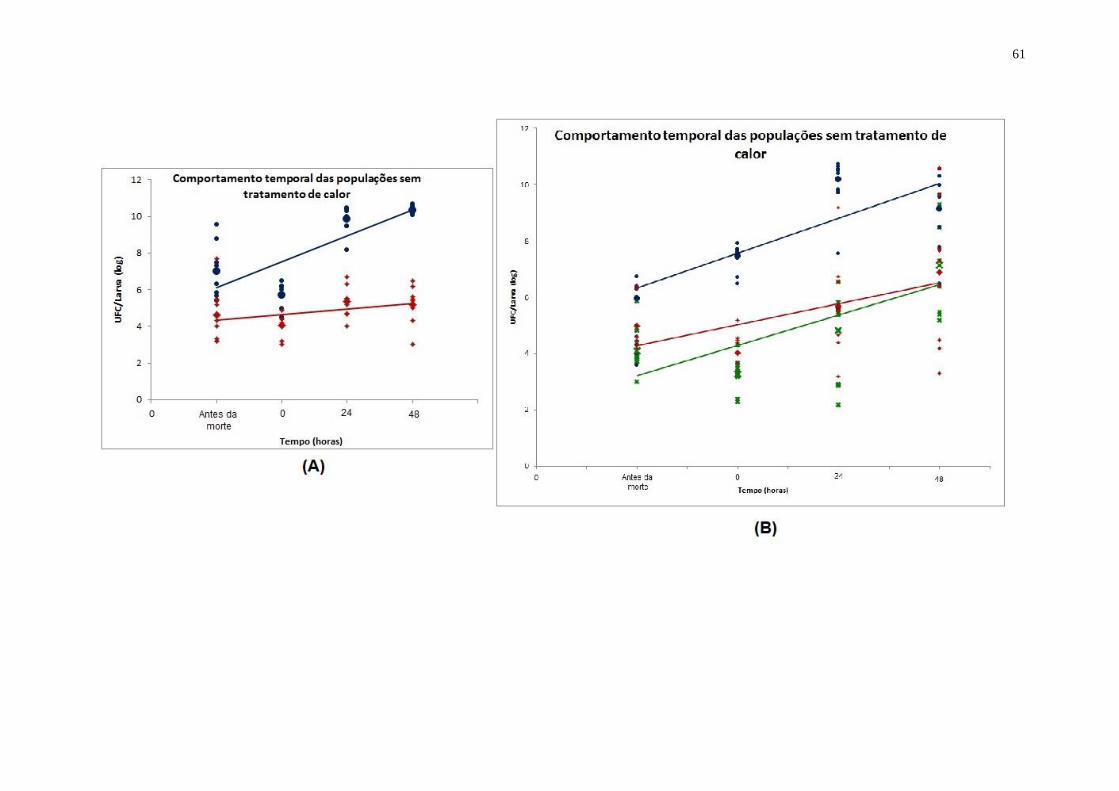
61