universidade de mogi das cruzes ana cristina …livros01.livrosgratis.com.br/cp019929.pdf ·...
TRANSCRIPT

UNIVERSIDADE DE MOGI DAS CRUZES
ANA CRISTINA SANCHES FLUMIGNAN
AVALIAÇÃO DE EXTRATOS BRUTOS NA REDUÇÃO ENZIMÁTICA DO NITRATO PARA FINS ANALÍTICOS
MOGI DAS CRUZES, SP
2006

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

UNIVERSIDADE DE MOGI DAS CRUZES
ANA CRISTINA SANCHES FLUMIGNAN
AVALIAÇÃO DE EXTRATOS BRUTOS NA REDUÇÃO ENZIMÁTICA DO NITRATO PARA FINS ANALÍTICOS
Dissertação apresentada ao programa de Pós-Graduação da Universidade de Mogi das Cruzes como parte dos requisitos para a obtenção do grau de Mestre em biotecnologia Área de Concentração: Exatas
Orientador: Prof. Dr. André Fernando de Oliveira
Mogi das Cruzes, SP
2006

FINANCIAMENTO: FAPESP, FAEP E DAEE/FAEP


DEDICATÓRIA
Dedico este trabalho ao Divino Mestre Jesus que em todos os
momentos tem sido sempre meu melhor amigo e companheiro.

AGRADECIMENTOS
Agradeço a todos os meus amigos e familiares pelo apoio, pela
amizade e pela preocupação.
Agradeço muito a minha mãe pelo exemplo de humildade,
simplicidade, bondade e força constante.
Agradeço meu pai por me proporcionar as oportunidades de
estudar, trabalhar e na realização de meus objetivos.
Agradeço ao meu companheiro João pela paciência, carinho,
esforço, dedicação e ao seu amor.
Agradeço ao Vô Zé, Vó Antônia e a Madrinha Belica (in
memorian) pela educação, amor e carinho dispensados.
Agradeço a Vó Clarice e ao Padrinho Roberto pelo amor e
auxílio por toda a vida.
Agradeço aos meus irmãos Zeca e Angelica pela união e
companheirismo.
Agradeço a Neila e Pedro por estarem sempre presentes,
incentivando e por vossa ajuda quando aqui cheguei.
Agradeço a Vilma A.Guimarães e a João F. Machado por me
cederem seus dias de folga, sem eles não teria tempo para concluir as
cinéticas.
Agradeço em especial a minha grande amiga Josi pela sua
ajuda inesgotável em todos os sentidos.
Agradeço de coração a verdadeira amizade que encontrei nesta
pessoa maravilhosa, que considero como meu irmão: Alexandre Correia de
Lima.
Agradeço do fundo da minha alma tudo que todos do
laboratório fizeram por mim (e não foi pouco): Alexandre Amâncio, Jonas,
Mônica, Débora, Sebastian, Aline Calixto, Aline Abbib, Hosney e Sérgio.
Agradeço a Professora Elisa e ao Professor Nelson pela

16

RESUMO Nitrato e nitrito são íons incolores, inodoros altamente solúveis em água e fracamente retidos no solo. Uma vez na água, o nitrato, devido a sua alta estabilidade e solubilidade, além de não formar espécies voláteis em equilíbrio-ácido base, permanece na água até a sua utilização por plantas e outros microrganismos. Por estes motivos, muitos estudos usam o nitrato como indicador potencial de poluição da água. Com o excesso de nitrato em ambiente aquático, as plantas aquáticas e algas podem se reproduzir excessivamente (eutrofização), caso haja disponibilidade de outros nutrientes (principalmente fósforo) e, eventualmente ocorrer uma grande diminuição do oxigênio dissolvido e causar a morte de espécies aeróbias, assim como impedir a realização da fotossíntese por algas e plantas submersas devido ao bloqueio da luz solar. Ao ser ingerido, o nitrato é rapidamente absorvido pelo trato gastrintestinal superior e poderá ser reduzido a nitrito devido à ação de nitrato redutases bacterianas, que poderão estar presentes na cavidade oral, estômago, intestino delgado, intestino grosso e bexiga. No sangue humano o nitrito ao chegar na corrente sanguínea pode causar metahemoglobinemia, ou seja, o íon ferroso da hemoglobina fica suscetível à oxidação pelo nitrito para a forma férrica que é instável para o transporte de oxigênio. Outro risco potencial a saúde é a formação do N-nitroso carcinogênico (também conhecido como nitrosaminas ou nitrosamidas) obtidos da reação entre amidas ou aminas secundárias ou terciárias. Por estes motivos existe a necessidade de um versátil, rápido e econômico ensaio para nitrato que permita a determinação precisa do mesmo para diversas amostras como água, alimentos e fluidos corporais humanos. Dentre os diversos métodos conhecidos para a determinação de nitrato a estratégia mais utilizada consiste na sua redução para nitrito e posterior determinação deste último, mediante, por exemplo, a reação de Griess. Geralmente cádmio metálico é o agente redutor preferido porque fornece uma redução quantitativa de nitrato para nitrito com desprezível redução de nitrito para espécies inferiores. Contudo, este redutor apresenta algumas desvantagens sendo a principal delas a geração de resíduos tóxicos. O presente trabalho teve como objetivo a avaliação de extratos brutos como fontes enzimáticas alternativas de baixo custo, para a redução do nitrato em amostras de água, com fins analíticos. Os testes foram divididos em três grupos de acordo com a fonte de nitrato redutase: o uso de cepas ou espécies distintas de fungos do gênero Fusarium, o uso de apenas um tipo de cepa de Escherichia coli e algumas fontes de origem vegetal como alface, gengibre e broto de feijão. Após a reação enzimática do nitrato com nitrato redutase o nitrito foi determinado com auxílio da reação de Griess. Quando se utilizou nitrato redutase de broto de feijão o método apresentou boa linearidade, baixa turbidez, boa eficiência (73%) e muitas vantagens quando comparados com as fontes de Nar microbiológicas e demais fontes vegetais: o preparo é rápido e muito fácil, tempo de reação curto, os equipamentos utilizados são simples, não descarta metais pesados, é inócuo para os analistas.
Palavra-chave: Nitrato redutase, determinação de nitrato, nitrito.

ABSTRACT Nitrate and nitrite are colorless, inodorous ions, highly soluble in water and weakly retained in the soil. Once in water, nitrate, because of its high stability and solubitity, besides not forming volatile species in acid-base equilibrium, it remains in water until its use by plants and other microorganisms. For these reasons, plenty of studies use the nitrate as a potential indicator of the water pollution. With the excess of nitrate in aquatic environment, aquatic plants and seaweed can reproduce in excess (eutrophication), in case there is availability of other nutrients (especially phosphorus) and, occasionally occur a large diminish of dissolved oxygen and cause death of aerobic species, as well as to block the accomplishment of photosynthesis by seaweeds and submerged plants because of the block of the solar light. Upon being swallowed, the nitrate is rapidly absorbed by the tract gastrointestinal upper and can be reduced to nitrite because of the action nitrate reductases bacterial, which can be present in the oral cavity, stomach, small intestine, large intestine and bladder. In the human blood, upon arriving in the blood flux the nitrite can cause methemoglobinemia, i.e., the ferrous ion of hemoglobin gets susceptible to the oxygenation by nitrite to the ferric form, which is unstable to the transportation of oxygen. Another potential risk to health is the formation of N-nitroso carcinogenic (also known as nitrosamines or nitrosamides) obtained from the reaction between amides or secondary or tertiary amines. For these reasons there is the necessity of a versatile, fast and economic essay to nitrate which allows the precise determination of it for several samples as water, food and human corporal fluids. Among the several methods known to the determination of nitrate, the most used strategy consists in its reduction to nitrite and later determination of this last one, through, for instance, the reaction of Griess. Usually metallic cadmium is the preferred reduction agent because it supplies a quantitative reduction of nitrate to nitrite with despicable reduction of nitrite to inferior species. However, this reduction agent shows some disadvantages, and the main one is the formation of toxic residues. The present work had as objective the evaluation of rough extracts as alternative enzymatic sources of low cost, for the reduction of nitrate in samples of water, with analytical ends. The tests were divided into three groups, according to the source of nitrate reductase: the use of strain or distinct species of fungi of the kind Fusarium, the use of just one kind of strain of Escherichia coli and some sources of vegetal origin as lettuce, ginger and beans sprout. After the enzymatic reaction of the nitrate with nitrate reductase the nitrite was determined with the auxiliary of the reaction of Griess. When nitrate reductase of beans sprout was used the method showed good linearity, low dimness, good efficiency (73%) and lots of advantages when compared to the sources of microbiological nitrate reductase and others vegetal sources: the preparation is fast and very easy, time of reaction is short, the used equipments are simple, it does not discard heavy metals, it is innocuous to the analysts.
Keywords: Determination of nitrate, nitrate reductase, nitrite.

LISTA DE ILUSTRAÇÕES
Figura 1 – Estruturas químicas dos reagentes usados na reação de Griess 22
Figura 2 – Mecanismo para as etapas de diazotação. . . . . . . . . . . . . . . . . . 23
Figura 3 – Espectro de absorção na região do visível do composto
produzido na etapa de acoplamento na reação de Griess. . . . . . 23
Figura 4 – Interação dos doadores e aceptores de elétrons com nitrato 28
Figura 5 – Diferentes espécies de Fusarium após 7 dias de crescimento. . 31
Figura 6 – Discos de micélio - ágar crescendo em meio de cultura líquido. 32
Figura 7 – Suspensão enzimática antes da filtração. . . . . . . . . . . . . . . . . . . 32
Figura 8 – Avaliação dos extratos de F.cacus e F.oxysporum como fontes
enzimáticas. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 43
Figura 9 – Avaliação da atividade da nitrato redutase com meio reacional
com água ou tampão. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 44
Figura 10 – Avaliação da biomassa da espécie cactus como fonte
de nitrato redutase em diferentes meios. . . . . . . . . . . . . . . . . . 45
Figura 11– Avaliação da atividade da nitrato redutase proveniente
da espécie cactus com diferentes proporções de biomassa. . . 46
Figura 12 – Influência de ácido ascórbico e glicose na atividade da nitrato
redutase proveniente da biomassa da espécie cactus . . . . . . . . 47
Figura 13 – Espectro evidenciando uma banda a 260 nm característica do
NAD+.. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 48
Figura 14 – Estudo do efeito da concentração de vitamina C no
sobrenadante. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 49

Figura 15 – Estudo do efeito da concentração de vitamina C no meio
ressuspendido. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 50
Figura 16 – Avaliação da atividade da Nar em meio reacional com
presença de sulfito de sódio. . . . . . . . . . . . . . . . . . . . . . . . . . . 51
Figura 17– Avaliação da atividade da Nar em meio reacional com
ausência de oxigênio. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 52
Figura 18– Diversas concentrações de nitrato em meio em anaerobiose. . . 53
Figura 19 – Avaliação da atividade da nitrato redutase proveniente da
cepa n˚ 69. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 54
Figura 20 – Teste utilizando Escherichia coli como fonte de Nar. . . . . . . . 55
Figura 21 – Curva analítica da concentração de nitrito formado e a
concentração de nitrato adicionado. . . . . . . . . . . . . . . . . . . . . 56
Figura 22 – Produção de Nar em condições anaeróbicas de crescimento 57
Figura 23 – Teste utilizando broto de feijão como fonte de Nar. . . . . . . . . 58
Figura 24 – Utilizando o protetor polivinilpirrolidona (PVP-VA). . . . . . . . 59
Figura 25 – Utilizando o protetor PVP-VA com diferentes concentrações
de nitrato. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 60
Figura 26 – Utilizando diferentes concentrações do protetor PVP-VA. . . . 61
Figura 27 – Utilizando uma concentração de 0,04 g/mL do protetor
PVP-VA em diferentes concentrações de nitrato. . . . . . . . . . 62
Figura 28 – Curva analítica da concentração de nitrito formado em função
da concentração de nitrato adicionado em tecido vegetal. . . . 63
Figura 29 – Avaliação da atividade da Nar em diferentes concentrações
de nitrato e pHs. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 64

Figura 30 – Avaliação do poder tamponante. . . . . . . . . . . . . . . . . . . . . . . . 65
Figura 31 – Avaliação do poder tamponante, considerando os valores de
eficiência máxima para as concentrações de fosfato no meio
reacional. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 66

SUMÁRIO
1 INTRODUÇÃO. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 12
1.1 CARACTERÍSTICAS GERAIS DO NITRATO E NITRITO. . . . . 12
1.2 ASPECTOS AMBIENTAIS. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 12
1.3 ASPECTOS TOXICOLÓGICOS. . . . . . . . . . . . . . . . . . . . . . . . . . . 14
1.4 ASPECTOS DE LEGISLAÇÃO. . . . . . . . . . . . . . . . . . . . . . . . . . . . 17
1.5 INTERESSE NA DETERMINAÇÃO DE NITRATO. . . . . . . . . . . . 17
1.6 ASPECTOS ANALÍTICOS NA DETERMINAÇÃO DE NITRATO. 18
2 OBJETIVOS. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 30
3 MÉTODO. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 31
3.1 CONDIÇÕES REACIONAIS. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 31
3.1.1TESTE UTILIZANDO FUSARIUM COMO FONTE DE NITRATO REDUTASE. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
31
3.1.2 TESTE UTILIZANDO ESCHERICHIA COLI COMO FONTE DE NITRATO RESUTASE. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
34
3.1.3 TESTE UTILIZANDO VIGNA RADIATA L. COMO FONTE DE NITRATO REDUTASE. . . . . . . . . . . . . . . . . . . . . . . . . . . . .
35
3.2 MATERIAIS. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 36
3.3 PREPARO DAS SOLUÇÕES. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 37
3.4 REAGENTES. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 40
3.5 EQUIPAMENTOS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 41
4 RESULTADOS. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 42
5 CONCLUSÕES E ETAPAS FUTURAS. . . . . . . . . . . . . . . . . . . . . . . . 67
REFERÊNCIAS. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 69

12
1 INTRODUÇÃO
1.1 CARACTERÍSTICAS GERAIS DO NITRATO E NITRITO
O íon nitrato é a base conjugada do ácido nítrico (HNO3) ácido forte (pKa = -1,37) que
se dissocia em água formando íons nitrato. O íon nitrito por sua vez é a base conjugada do
ácido nitroso (HNO2), ácido fraco (pKa = 3,37).(ANDRADE, 2004)
Nitrato e nitrito são íons incolores (o nitrato apresenta um máximo em 220 nm),
inodoros, altamente solúveis em água e fracamente retidos no solo.(NOLLET, 2000)
Enquanto nitrato é muito estável, o nitrito é altamente reativo e estão distribuídos
naturalmente em alimentos vegetais e animais.(FOX, 1983)
1.2 ASPECTOS AMBIENTAIS
Concentrações de nitrito em sistemas aquáticos geralmente são encontradas em valores
abaixo da concentração de 0,5 mg N/L devido a sua tendência a ser oxidado para nitrato.
Altas concentrações de nitrito podem estar indicando poluição de água em estuários ou
proximidades levando à redução da atividade microbiana. A poluição de corpos d’água por
fontes de nitrogênio usualmente começa com a introdução de fezes, urina, animais mortos ou
plantas. Por exemplo, os primeiros contêm quantidades apreciáveis de nitrogênio incorporado
aos compostos orgânicos, enquanto que a urina que contém nitrogênio na forma de uréia. A
ação bacteriana sob a matéria orgânica resulta em degradação e liberação de amônia que
poderá ser oxidada aerobicamente a nitrito por bactérias como as
Nitrossomonas.(ANDRADE, 20004)
NH3 + 2H2O → NO2 + 7H+ + 6e

13
que, por sua vez se converte a nitrato pela ação de bactérias como Nitrobacter:
NO2 + H2O NO3¯ + 2H+ + 2e
Por outro lado, as concentrações de nitrato em águas profundas dos mares e
reservatórios naturais de água não poluída estão, usualmente, abaixo de 0,05 mg N/L. Em
reservatórios rasos e superfícies de córregos a concentração pode variar desde valores
menores que 0,1 mg N/L até 20 mg N/L dependendo, por exemplo, do tipo de solo e do uso
da terra nas proximidades dos reservatórios.
Quando o nitrato está presente nas águas, devido a sua alta estabilidade e solubilidade,
além do fato de não formar espécies voláteis em equilíbrios ácido-base, ele permanece na
água até sua utilização por plantas e outros microrganismos. Por estes motivos, muitos
estudos usam o nitrato como indicador potencial de poluição da água. A contaminação do
meio ambiente pelo nitrato pode ocorrer principalmente através:
•
•
•
•
Uso intensivo de fertilizantes químicos nitrogenados e de compostagem, uma vez que
o rendimento da agricultura depende muito do suprimento de nitrato no
solo.(BORCHEDING, 2000; ZHU, 2006)
As águas de chuva e de irrigação ao atingir o solo podem solubilizar e transportar
nitrato e nitrito solubilizados até reservatórios de água.
Descartes de efluente doméstico municipal e industrial, sistemas de descartes e
detritos, uma vez que esses contêm uma grande quantidade de nitrogênio orgânico e amônia.
Após um período de tempo, estas espécies em ambiente aeróbio podem ser transformadas
para nitrato.
Alta população de gado em pequenas áreas com grande produção de esterco.
Com o excesso de nitrato em ambiente aquático, as plantas aquáticas e algas podem se
reproduzir excessivamente (eutrofização), caso haja disponibilidade de outros nutrientes
(principalmente fósforo) e, eventualmente ocorrer uma grande diminuição do oxigênio
dissolvido e causar a morte de espécies aeróbias, assim como impedir a realização da
fotossíntese por algas e plantas submersas devido ao bloqueio da luz solar.(ANDRADE,2004)

14
Outros efeitos negativos da eutrofização são: desenvolvimento excessivo e prejudicial
de algas, provocando alterações profundas da biota, com a substituição de espécies de peixes
e outros organismos. Quando essas algas começam a morrer há decomposição de matéria
orgânica, causando um aumento no consumo do oxigênio dissolvido e anóxia. Ainda,
degradação da qualidade da água, com alterações de composição, cor, turbidez, transparência,
liberação de gases e produção de maus odores, levando a prejuízos consideráveis para o
tratamento para água de abastecimento, irrigação e para aproveitamentos hidroelétricos, como
prejuízos para recreação, turismo e paisagismo, além do aumento da evaporação, elevação de
nível e entraves para o escoamento das águas, condições propícias para a criação de
mosquitos, larvas e outros vetores. Mas, a preocupação principal é com a produção de
substâncias tóxicas, as hepatotoxinas e neurotoxinas.(AZEVEDO NETO,1988)
De maneira geral, os nutrientes que devem ser removidos ou ter suas cargas reduzidas
nos efluentes são nitrogênio e o fósforo, pois são considerados os principais limitantes ou
controladores da produtividade primária.
1.3 ASPECTOS TOXICOLÓGICOS
Do ponto de vista fisiológico, ao ser ingerido, o nitrato é rapidamente absorvido pelo
trato gastrintestinal superior e poderá ser reduzido a nitrito devido à ação de Nars (nitrato
redutases) bacterianas, que poderão estar presentes na cavidade oral, estômago, intestino
delgado, intestino grosso e bexiga.(WARD, 2005; LUNDBERG, 2004)
Nitratos e nitritos além de estarem presentes em diversas concentrações em diversos
vegetais (Tabela 1) também são utilizados na indústria alimentícia como aditivos intencionais
na tecnologia de produtos cárneos devido à capacidade do nitrito de combinar-se com a
mioglobina presente na carne, originando o pigmento óxidonitricomioglobina, pigmento este
responsável pela cor avermelhada da carne curada.

15
Tabela 1: Concentrações de nitrato e nitrito presente em alguns vegetais.
Vegetal NO3-/ ppm NO2
- / ppm Repolho 784 0,5
Pepino 24 0,5
Berinjela 302 0,5
Alface 1.100 0,4
Cenoura 72 0,6
Espinafre 2.200 0,7
Brócolis 464 1,0
Couve-Flor 254 1,1
O nitrato de potássio é utilizado na elaboração de queijos já que inibe o crescimento de
bactérias do tipo Clostridium butyricum, presentes no leite e que produzem fermentações
indesejáveis, podendo comprometer a qualidade do produto.
Tanto nitrato como nitrito têm sido utilizados como protetores contra Clostridium
botulinum, mas estudos revelam que esta proteção é insuficiente quando há abundância de
esporos resistentes.(TURRA, 1999)
O nitrito ao chegar na corrente sanguínea pode causar metahemoglobinemia, ou seja, o
íon ferroso da hemoglobina fica suscetível à oxidação pelo nitrito para a forma férrica que é
instável para o transporte de oxigênio onde a oxihemoglobina passa a ser chamada de
metahemoglobina.(COSS, 2004; YANG, 2005; SILVA, 1998; FANQUIN, 2005)
Apenas uma quantidade muito pequena de metahemoglobina se encontra presente no
sangue normal (até 2%), pois o eritrócito (hemácia) possui um sistema efetivo para reduzir o
Fe3+ hemínico de volta ao estado Fe2+ chamado de sistema NADH-citocromo b5
metahemoglobina redutase. Ele consiste de NADH produzido pela glicólise, uma
flavoproteína denominada citocromo b5 redutase (também conhecida como metahemoglobina
redutase) e citocromo b5 que é então regenerado pela ação do citocromo b5
redutase.(MURRAY, 1998)
Existem duas situações em que hemácias produzem metahemoglobinemias:
- Metahemoglobinemia hereditária: existe uma deficiência da atividade da
metahemoglobina redutase que é transmitida de forma autossômica recessiva;

16
- Metahemoglobinemia adquirida: em que a metahemoglobina é causada devido à
ingestão de drogas ou substâncias químicas como, por exemplo, o nitrato que poderá ser
reduzido a nitrito.(LORENZI, 1999)
A atenção maior deve ser voltada para os indivíduos mais suscetíveis que são:
- Crianças lactentes de até três meses de idade, pois a atividade da NADH-citocromo b5
redutase é de aproximadamente de 50-60% da atividade encontrada em adultos. Do mesmo
modo o pH intestinal de crianças é altamente suficiente para contribuir para o crescimento de
organismos intestinais que são particularmente eficientes na conversão do nitrato ingerido
para nitrito.(ZEMAN, 2002)
- Indivíduos com acidez gástrica reduzida que resulta na proliferação de espécies
bacterianas capazes de reduzir nitrato para nitrito.(GEFFNER, 1981)
- Mulheres grávidas, uma vez que o nitrito é conhecido por causar aborto espontâneo e
ser teratogênico.
Os sinais mais evidentes ocorrem quando acima de 10 % da hemoglobina se encontra na
forma de metahemoglobina que são descolorações azuladas da pele e mucosas (cianose), já
acima de 30% os sintomas são fadiga, dor de cabeça, taquicardia e vertigem. Acima de 55% a
60 % a oxigenação dos tecidos fica inadequada podendo resultar em dispnéia, acidose,
arritmias, paralisias, coma e convulsões. A morte pode acontecer a níveis acima de 70 %
(FURLANI, 2006)
O diagnostico pode ser suspeitado quando além dos sinais e sintomas uma amostra de
sangue está com a coloração marrom e não avermelhada quando exposta ao ar.(FINAN, 1988)
Devido ao aspecto não convencional das membranas e mucosas a metahemoglobinemia
costuma ser referida como doença do sangue azul e quando ocorre em crianças síndrome do
bebê azul.
Os casos de intoxicação estão geralmente relacionados com a ingestão de água contendo
mais de 100mg/L de NO3-. Também tem sido observado prejuízo na função da tireóide,
decréscimo na alimentação e interferência no metabolismo das vitaminas A e E .(SANTOS,
2005)
Outro risco potencial a saúde é a formação do N-nitroso carcinogênico (também
conhecido como nitrosaminas ou nitrosamidas) obtidos da reação entre amidas ou aminas
secundárias ou terciárias.(KIANG, 1978)
As nitrosaminas (ou nitrosamidas) estão presentes em produtos alimentícios,
farmacêuticos, amostras ambientais (água, solo, ar...), pesticidas herbicidas, cosméticos, etc.

17
As nitrosaminas são um dos carcinógenos mais potentes e versáteis, produzindo
tumorações em muitas espécies e praticamente em todos os órgãos do corpo. São rapidamente
metabolizadas e em indivíduos suscetíveis apenas uma dose pode produzir tumorações, sendo
considerado o nível de exposição tolerável pelo homem para nitrosaminas mais voláteis é de
5 a 10 µg/Kg .(SANCHES, 2003)
Mesmo evitando o consumo de produtos contendo nitrosaminas, estudos in vitro,
envolvendo nitrito e aminas secundárias, têm mostrado que o suco gástrico de mamíferos
prova ser um excelente meio para a produção de derivados do nitroso.(DOMBROWSKI,
1972)
Embora os vegetais sejam ricos em nitrato, visto que a planta absorve nitrato como
fonte de nitrogênio para seu crescimento, os índices epidemiológicos do risco de câncer são
menores devido a fatores protetores como α-tocoferol e ácido ascórbico presentes nestes
alimentos (são conhecidos como inibidores da formação de compostos N-nitroso).
1.4 ASPECTOS DE LEGISLAÇÃO
Segundo a Agência de Proteção Ambiental dos Estados Unidos o nível máximo
considerado de contaminação para nitrato em água ingerida é 0,16 mmol/L, sendo
recomendada pela Organização Mundial de Saúde a ingestão diária é de 0,003 mg de
nitrato/kg de peso corporal.(BORCHERDING, 2000)
1.5 INTERESSE NA DETERMINAÇÃO DE NITRATO
O interesse na quantificação de nitrato em fluidos biológicos tem aumentado
significativamente desde a elucidação das propriedades do óxido nítrico (NO) em diagnóstico
clínico.(GRANGER, 1996)
O NO é um produto da L-arginina na presença da NO sintetase (E.C 1.14.13.39) em
diferentes tipos celulares.

18
Muitos estudos quantitativos têm usado medidas de nitrato e nitrito (já que são produtos
estáveis da oxidação de NO) em sistemas biológicos e culturas celulares como índice da
produção de NO uma vez que a determinação direta do NO é difícil por ele ser um radical
livre liberado pelas células em concentrações na ordem de pmol/L e nmol/L e tem um curto
tempo de vida.(SCHIMIDT,1995; LI, 2000)
Além do mais, NO é relativamente instável em presença de oxigênio e é rapidamente e
espontaneamente auto-oxidado em uma das fases do gás para produzir uma variedade de
óxidos de nitrogênio em solução aquosa.(GRISHAM, 1996)
Clinicamente os níveis de nitrato aumentam no soro após septsemia e gastrenterite. No
fluido cerebroespinhal, os níveis de nitrato e nitrito aumentam após meningite e Síndrome de
Parkinson. Crianças com síndrome nefrótica apresentam teores elevados de nitrato na urina.
Na urina de pacientes com artrite reumatóide os valores de nitrato e creatina aumentam três
vezes em relação aos níveis normais, e duas vezes em pacientes com infecção gastroentérica.(
FRIEDBERG, 1997; DEMBNY, 1998)
Por estes motivos existe a necessidade de um versátil, rápido e econômico ensaio para
nitrato que permita a determinação precisa do mesmo para diversas amostras como água,
alimentos e fluidos corporais humanos.
1.6 ASPECTOS ANALÍTICOS NA DETERMINAÇÃO DE NITRATO
São inúmeros os métodos para determinação e monitoramento de nitrato em água,
fluidos corporais humanos, plantas e solo e podem ser divididos em duas categorias:
A) Direto:
Com detecção direta, no UV (220 nm), ou com uso de eletrodo íon seletivo, ou por
derivatização, nitração de acido salicílico, nitração de fenóis, cromatografia a gás após

19
derivatização, cromatografia de íons, eletroforese capilar direta e indireta, polarografia. Os
métodos diretos podem ser:
A1) Métodos diretos sem derivatização
A2) Métodos diretos com derivatização
B) Indireto - Redução de espécies do nitrogênio
Com redução a nitrito e detecção espectrofotométrica, eletroquímica,
quimioluminescência. Com redução para amônia e detecção espectrofotométrica,
potenciométrica, condutimétrica. Ou ainda, com redução a óxidos gasosos com detecção de
quimioluminescência. A redução poderá ser:
B1) Redução heterogênea do nitrato
B2) Redução homogênea para nitrito
B3) Redução fotoquímica
B4) Redução enzimática
A.1- Métodos Diretos Sem Derivatização
A medida direta de nitrato, em 220 nm, como método padrão de análise não requer
nenhum reagente adicional e é adequado para amostras com baixo teor de matéria orgânica,
água natural não contaminada e água potável. Devido à matéria orgânica dissolvida também
absorver em 220 nm e nitrato não absorver a 275 nm, uma segunda medida pode ser usada
para corrigir o valor de nitrato.
Assim, dentre os interferentes a matéria orgânica dissolvida é a principal, embora
surfactantes, nitrito, e Cr (VI) também interfiram nessa determinação, além de vários íons
inorgânicos não encontrados em quantidades apreciáveis em água natural.
O eletrodo íon-seletivo para nitrato é um sensor potenciométrico com uma faixa
analítica usual de 10-5 e 10-1 mol/L (0,14 a 1400 mg NO3- N/L). O eletrodo mais comum é
baseado em um par iônico formado com Aliquat 336S, dissolvido em membranas de PVC. O
baixo limite de detecção é determinado por uma pequena, mas finita partição do par-iônico na
solução.

20
Dentre os interferentes, os íons cloreto e bicarbonato apresentam coeficientes de
seletividade maiores que 0,1 e 0,05, respectivamente. Íons que são potencialmente
interferentes, mas que não ocorrem normalmente em água potável são o NO2-, CN-, S2-, Br-, I-,
ClO3- e ClO4
-. Embora os eletrodos funcionem satisfatoriamente em tampão entre pH 3 e 9
são usualmente observados valores discrepantes, de maneira que este sensor, é usado de modo
semi-quantitativo em medidas em águas de rios e estações de tratamento de
efluentes.(CLESCERI,1998)
A.2- Métodos Diretos com Derivatização
O método de nitração do ácido salicílico, também conhecido como método do salicilato
é usado para determinação de nitrato não apenas em águas, mas também em extratos de solos
e plantas. O método se baseia na formação, em meio ácido e com aquecimento, dos íons
nitrônio (NO2+), que reagem com salicilato, em meio básico formando principalmente um
composto nitrobenzóico de cor amarela. Como o nitrogênio no nitrato não tem capacidade
eletrofílica, o elemento precisa ser ativado a nitrônio, que é um eletrófilo forte. O
procedimento envolve aquecimento da amostra até a secura, pois a água precisa ser removida
para conversão do NO3- a NO2
+. Portanto, o método é trabalhoso, demorado, e limita o
número de amostras a serem analisadas, particularmente quando se utilizam grandes volumes
de amostras de água.(FERREIRA, 2006)
B. Indireto
A determinação indireta do nitrato apresenta vantagens em relação à direta: maior
sensibilidade, maior precisão e melhor seletividade, ou seja, é menos sujeita à interferência de
outros íons.(MANTOVANI, 2005)
Dentro destes métodos, entretanto, convém destacar que a redução de nitrato para nitrito
é a principal estratégia para determinação desse íon.
Essa redução pode ser acompanhada por adição de um agente redutor homogêneo ou
heterogêneo ou através de reduções fotoquímicas ou ainda por reações enzimáticas.

21
B1)Redução Heterogênea do nitrato.
Os agentes redutores freqüentemente usados em estado sólido são: liga de cádmio, ou
cádmio metálico granulado recoberto com cobre ou ligas de zinco, adicionado diretamente na
amostra ou passando a amostra através de uma coluna contendo o agente redutor. O uso de
sistemas de injeção em fluxo tem facilitado bastante o procedimento.
Geralmente ele é o reagente preferido porque fornece uma redução quantitativa de
nitrato para nitrito com desprezível redução do nitrito para oxidação de espécies
inferiores.(ANDERSON, 1979)
NO3- + Cd 0 + 2H+ pH 7,.5 NO2
- + Cd 2+ + H2O (PATTON, 2002)
Tampão imidazol
Algumas desvantagens da redução cobre-cádmio é a degradação da superfície redutora
causada por uma gradual perda de cobre e de cádmio, a precipitação de outras espécies
presentes em amostras de água na superfície, a perda de carga na coluna devido à
compactação de partículas e a adsorção de certos componentes, principalmente matéria
orgânica. Para evitar a precipitação do Cd 2+ formado, cloreto de amônio ou EDTA são
adicionados à amostra, embora a degradação do agente redutor persista e a coluna deve ser
periodicamente regenerada para recuperar a camada da superfície metálica, até que seja
finalmente substituída pelo consumo do reagente.
A principal desvantagem deste método é a geração de resíduos tóxicos de Cd 2+.
Quando ocorre exposição ao cádmio os efeitos agudos consistem em edema e irritação. A
toxicidade crônica afeta os rins. Ocorre indução da síntese de metalotioneína, uma proteína de
ligação de metal, no fígado e nos rins. Quando este mecanismo de defesa é superado, o
cádmio provoca lesão dos túbulos contornados proximais, causando proteinúria. Ele pode
causar também cardiopatia, doença pulmonar crônica obstrutiva e infertilidade
feminina.(COTRAN, 2000)
Há uma crescente conscientização da toxicidade do cádmio, entretanto ele continua
sendo usado com preocupação por analistas. Os custos associados com os processos de
descartes aquáticos contendo cádmio e seu correto descarte também tem aumentado.

22
Estes fatores levam a explorar mais alternativos agentes redutores para a rotina na
determinação de nitrato.
O zinco metálico tem sido utilizado sozinho ou combinado com íons magnésio, para
promover essa redução. O método requer um estrito controle da temperatura e tempo de
reação para evitar a redução do nitrogênio para um estado de oxidação menor que III.
Reduções quantitativas não são obtidas e os melhores rendimentos reportados estão entre 85-
90%, em meio alcalino.
O nitrito formado é, em geral, monitorado espectrofotometricamente usando a reação de
Griess, qualquer que seja o agente redutor utilizado.
Princípio do método de Griess:
A reação de Griess é baseada na formação de um diazocomposto. Inicialmente, o nitrito
reage com a sulfanilamida (Figura 1a) em meio ácido (reação de diazotação) e o produto
reage com n-(1-naftil)-etilenodiamina (NNED – Figura 1b) formando o diazocomposto
(Figura 2).
(a)
(b) NH2
NH2
O OSNH2
NH
Figura 1 - Estruturas químicas dos reagentes usados na reação de Griess. (a) sulfanilamida;
(b) N-(1-Naftil) etilenodiamina.

23
dimetilamina
Figura 2 - Mecanismo para as etapas de diazotação (A) e acoplamento(B) da reação de Griess para determinação de nitrito.
diazocomposto
O espectro do diazocomposto obtido é apresentado na Figura 3. O máximo em 540
nm, que apresenta uma absortividade molar de 3,4. 104 L/(mol. cm) é usualmente utilizado
para a obtenção da curva analítica.
Figura 3 - Espectro de absorção na região do visível do composto produzido na reação de
Griess, obtido com solução padrão de nitrito de sódio 0,33 mg N/L. B2) Redução Homogênea para nitrito.

24
A redução do nitrato por hidrazina em meio alcalino, catalisada pelo íon cobre.
Diferentes métodos baseados neste princípio têm sido desenvolvido e adaptados para
diferentes tipos de água. A principal limitação é provavelmente a grande dependência da taxa
de conversão com a temperatura, que pode levar a uma pobre reprodutibilidade. Além disso, o
método apresenta interferências principalmente por cátions, tais como Mg2+, Ca2+ e Fe3+, pois
eles podem precipitar hidróxidos em meio alcalino. Essas interferências têm sido eliminadas
removendo esses cátions com resina catiônica ou por precipitação em meio alcalino e
filtração, ou mesmo trabalhando em altas temperaturas, quando o tempo para a determinação
é diminuído. A adição de agentes complexantes raramente é utilizada para evitar a inibição do
efeito catalítico do cobre. Esta inibição é causada por complexantes naturais, tais como os
ácidos húmicos ou fúlvicos, presentes em matrizes de águas de rios e lagos e de solo. Essa
interferência pode ser minimizada pela adição de íon zinco a amostra, para competir por esses
ligantes, ou, equivalentemente, com o aumento da concentração do cobre (II). Em alguns
casos isto é possível adicionando um agente complexante para remover a espécie interferente
específica sem afetar a reação catalítica permitindo a determinação de nitrato em água do mar
com limite de detecção de 0,003 mgN/L.
B3) Redução Fotoquímica
A irradiação da amostra com ultravioleta produz a redução do nitrato para nitrito por
fotólise deste ânion. A redução fotoquímica envolve um ou mais estados de excitação através
de diferentes caminhos. A predominância de uma reação sobre as outras e conseqüentemente,
o rendimento da fotólise, depende do pH, da concentração inicial de nitrato, da intensidade e
do comprimento de onda da radiação eletromagnética utilizada. A fotólise pela luz solar
ocorre naturalmente em ambiente aquático e pode ser reproduzida em laboratório pelo uso de
lâmpada de mercúrio. Alguns autores reportam uma eficiência de redução de 93 % para
nitrato após a irradiação da amostra por 8 minutos em pH alcalino e predominância da
emissão em 254 nm.(NOLLET, 2000)

25
Em outro método, o íon nitrato pode também ser determinado por quimiluminescência
após formação fotoquímica de espécies oxidantes, como peroxinitrito, que causam a oxidação
de luminol, responsável pelo efeito luminescente.(CAVACCHIOLI, 2003)
B4) Redução Enzimática
Neste método utilizam-se enzimas específicas na conversão do nitrato para nitrito, que
são chamadas de nitrato redutases. Estas enzimas estão amplamente distribuídas na natureza e
são produzidas por uma variedade de animais, plantas e microrganismos incluindo
fungos.(XUEIJUANG, 2006)
Nestes organismos, a redução do nitrato pode ocorrer por duas razões, devido as duas
principais rotas metabólicas:
♦ Assimilação do nitrato : neste caso o nitrogênio é utilizado como fonte para o
crescimento do organismo ( biossíntese);
♦ Respiração do nitrato (Dissimilação do nitrato) : ocorre geração de energia metabólica
com o nitrato como aceptor de elétrons uma vez que ele será utilizado como oxidante na
cadeia respiratória sob condições anaeróbias.(OLIVEIRA, 2005; MARTINEZ-ESPINOSA,
2001; KHAN, 2006)
Baseado nessas funções, as nitrato-redutases podem ser classificadas em duas classes
distintas:
I)Nitrato-redutase Assimilatória
São encontradas em plantas verdes, algas, fungos, leveduras, e algumas bactérias que
participam da redução do nitrato a amônia que será incorporada aos compostos
orgânicos.(HOFF, 1992)
A purificação e caracterização, em 1953, da piridina nucleotídeo nitrato redutase em
Neurospora e folhas de soja, representou a primeira e definitiva examinação reportada da
nitrato redutase assimilatória.(BOYER, 1963). Existem kits comerciais para determinação de

26
nitrato contendo nitrato redutase purificada, mas o custo costuma ser elevado (em torno de
100 dólares duas unidades as nitrato redutase – NADH dependente).
Dentre as fontes de nitrato redutase assimilatória, pode-se citar: Zea mays (milho),
Aspergilus niger (fungo filamentoso), Candida utillis (levedura), Neurospora crassa
(nitratooxidorredutase EC 1.6.6.3), Anabaena cylindrica (cianobacteria dependente de
ferredoxina), Eclothiohadospira shaposnikovii (cianobactéria fotossintética), Azobacter
vinelandii (flavodoxina dependente), Azobacter chrooccum (bactéria fixadora de nitrogênio),
Klebsiella pneumoniae, Rodobacter capsulatus e Haloferax mediterranei (NADH
dependentes).(MIKAMI, 1984)
As nitrato redutases ferredoxina dependentes ocorrem em procarióticos e têm como
função alternativa fisiológica nos anaeróbicos. Outros exemplos:
-Chlorella vulgaris (NADH dependente, EC. 1.6.6.1) alga
unicelular.(SOLOMONSON,1975; GEWTIZ, 1981)
-Nicotiana pumblaginifolia (embora a nitrato redutase esteja presente em níveis
relativamente baixos em tecidos de plantas superiores na Nicotiana é particularmente muito
baixa)
Na Trema guineensis atividade nas folhas é de 11.9 µmol NO2 gf. wt-1h-1, embora nas
raízes a atividade seja reduzida.(POONNACHIT, 2004)
Onde o transporte do nitrato para o meio intracelular é mediado pela permease do
nitrato e após a sua assimilação ocorre redução seqüencial do nitrato para nitrito, pela ação da
nitrato redutase e do nitrito para amônia, pela ação da nitrito redutase, que será utilizada para
biossíntese celular.(CHOW, 2004; LILLO, 2004)
I) Nitrato-redutase Dissimilatória
É encontrada em algumas bactérias, não tem sido encontrada em plantas e uma pequena
parte de sua estrutura é parecida com a nitrato redutase assimilatória.(HOFF, 1992)
Em alguns organismos, essas enzimas têm a capacidade de usar o nitrato como aceptor
terminal de elétrons no lugar do oxigênio. Devido a esta fisiológica e enzimológica
similaridade a respiração aeróbica, esse processo é usualmente chamado de respiração do
nitrato ou dissimilatória redução do nitrato.(UNKLES, 2004)

27
A primeira demonstração da respiração do nitrato em bactérias não denitrificantes foi
feita quando observaram que E.coli não cresceu anaerobicamente em um meio contendo
lactato como fonte de carbono a menos que um aceptor de elétrons como o nitrato fosse
colocado no meio.(HOFF, 1992)
Alguns exemplos de fontes de nitrato redutase respiratória são as Paracoccus
denitrificans, Pseudomonas aeruginosas, Pseudomonas stuzeri.(BORCHERDING, 2000). Na
E. coli (EC 1.7.99.4) a nitrato redutase está ligada a membrana e pode ser induzida em
grandes quantidades por crescimento anaeróbico em presença de nitrato.(MACGREGOR,
1974) A enzima apresenta uma habilidade de translocar prótons, pois ocupa uma localização
transmembranica na membrana citoplasmática.(MORPETH, 1985). Na Klebsiella aerogenes a
enzima também está ligada à membrana.(VAN`TRIET, 1975)
As nitrato redutases-respiratória ligadas à membrana são geralmente proteínas
integrais da membrana com o sítio ativo localizado na face citoplasmática da membrana
citoplasmática.(JEPSON, 2004)
A estrutura da nitrato redutase
De modo geral nitrato redutase apresenta um grande complexo multirredox que possui
duas subunidades idênticas (homodímero, podendo também se apresentar na forma de
homotetrâmero), de subunidades de peso molecular de aproximadamente 95 a 140 kDa e 50
kDa respectivamente, cada uma composta por três grupos prostéticos em domínios separados.
Nas extremidades c-terminal localiza-se o domínio FAD, mais ao centro o domínio Heme e na
extremidade terminal o molibdênio numa estequiometria de 1:1: 1. Cada cofator domínio
constitui de um elemento estrutural autônomo e igualmente isolado que reserva esta atividade
parcial. O molibdênio é responsável pela redução do nitrato, sendo também operado em
presença de doadores de elétrons sintéticos.(PEREIRA, 2006; SOLOMONSON, 1990;
MOUREAUX, 1989,FERREYRA, 2004; RIDLEY, 2006)
De maneira simples, uma unidade da nitrato redutase pode ser entendida de acordo com
a figura 4:

28
Fe (CN)63-
Cyt c MV
NAD(P)H
NO3-
MoFAD HEMENAD(P)+
NO2-
Figura 4 - Interação dos doadores e aceptores de elétrons com nitrato redutase.(BARBIER, 2005)
De acordo com o doador de elétron podemos dividir a nitrato redutase em dois grupos:
1) Diaforase: grupo de enzimas que usa NAD (P)H como doador de elétrons e inclui
NADH, dentre eles, a Ferricianida nitrato redutase (NADH: Fr), a NADH: citocromo c
redutase (NADH: CR) e a NADH: diclorofenolindol redutase (NADH: DR).
A NADH: FR requer FAD, enquanto que a NADH: CR E NADH: DR requerem FAD e
heme como grupos prostéticos.
2) Redutoras do nitrato, tais como a Reduzida flavina nitrato redutase (FH: NR),
Reduzida metil viologênio (MV: NR), Reduzida azul de bromofenol (BB: NR).A FH: NR e
BB: NR requerem apenas Mo.A Nar na maioria dos organismos se apresenta na forma de
homodímero, mas pode se apresentar na forma de Homotetrâmero como por exemplo na
Chlorella. sp (SOLOMONSON, 1990) ou Heterodímero (Klebsiella pnenumoniae,
Rhodobacter capsulatus (MARTINEZ-ESPINOSA, 2001), Haloferax denitrificans) e
Heterotrimétrica (na Haloferax volcanii).
Localização da nitrato redutase
A localização da nitrato redutase pode variar muito, dependo das necessidades
fisiológicas do organismo.
Como por exemplo, nas plantas existe uma necessidade eficiente para transferência do
nitrito formado pela nitrato redutase para os cloroplastos onde será reduzido a amônia pela
nitrito redutase, portando a mesma se encontrará no citosol, mesmo porque o NADH é

29
produzido no citosol. A nitrato redutase tem sido encontrada em diversos compartimentos
celulares, tais como em cloroplastos na Moraphidium braunii (alga verde), no plasmelema e
membrana tonoplástica das células miceliais de Neurospora crassa, assim como no
citoplasma do mesófilo celular em folhas de Zea mays (milho).
Portanto diferentes fontes de nitrato redutase podem se utilizadas no método da
redução enzimática, sendo a utilização de extratos brutos considerada mais vantajosa quando
comparada com outros métodos, pois:
- É uma fonte inesgotável de enzimas e não necessita de cofatores (os quais apresentam
custo comercial elevado).
- A nitrato redutase apresenta tempo de reação rápida, maior eficiência e especificidade
requerendo menor energia.(CAMPBELL, 1999)
- A análise é mais seletiva para nitrato;
- Apresenta menor interferência ou falso positivo;
- As condições de reação são simples;
- Não adiciona reagentes pesados;
- O custo da análise é baixo;(CAMPBELL, 2002)

30
2 OBJETIVOS
2.1 OBJETIVOS GERAIS
O presente trabalho teve como objetivo geral avaliar extratos brutos na redução
enzimática do nitrato para fins analíticos.
2.2 OBJETIVOS ESPECÍFICOS
Os objetivos específicos compreenderam em:
- Avaliar diferentes fontes de nitrato redutase visando seu uso como extrato bruto para
determinação de nitrato.
- Utilizar reagentes não carcinogênicos, de fácil descarte e de baixo custo.

31
3 MÉTODO
Todas as soluções foram preparadas com água tipo 1 ( 18,18 MΩ.cm ), obtida em um
purificador de água ultra System – Gehaka.
3.1 CONDIÇÕES REACIONAIS
Os testes foram divididos em três grupos de acordo com a fonte de nitrato redutase: o
uso de cepas ou espécies distintas de fungos do gênero Fusarium, o uso de apenas um tipo de
cepa de Escherichia coli e utilizando fontes de origem vegetal como alface, gengibre e Vigna
radiata L. conhecida como broto de feijão.(LIMA, 2004)
3.1.1 TESTE UTILIZANDO FUSARIUM COMO FONTE DE NITRATO REDUTASE
(FONTE FÚNGICA)
Foram testadas as seguintes fontes de fungos para a biossíntese de nitrato redutase:
Fusarium cactus, Fusarium oxysporum (cepa 55.1), Fusarium oxysporum (cepa 07),
Fusarium oxysporum (cepa 69). Essas foram cultivadas em meio de cultura sólido e mantidas
a temperatura de 28 °C por 7 dias em estufa BOD Q – 315D ( QUIMIS ®), Figura 5.
Figura 5 - Diferentes espécies de Fusarium após 7 dias de crescimento.

32
Destes foram repicados 3 discos de micélio – ágar de 5 mm de diâmetro e transferidos
para Erlenmeyer contendo 100 mL de meio de cultura líquido, e incubados a 28 °C por 12
dias em estufa BOD Q – 315D ( QUIMIS ®) a fim de produzir biomassa necessária para os
ensaios seguintes, Figura 6.
Figura 6 - Discos de micélio - ágar crescendo em meio de cultura líquido.
Neste grupo, os testes foram divididos em duas partes: o uso do extrato sem a presença
da biomassa (ou seja, os filamentos dos fungos) e outro contendo a biomassa.
3.1.1.1 EXTRATO DE FUSARIUM COMO FONTE ENZIMÁTICA
O extrato foi pesado e transferido para um erlenmeyer onde água destilada previamente
autoclavada foi adicionada na proporção de 100 mL de água para 10 g de extrato. Este extrato
ficou suspenso em água por 3 dias a 28 °C em estufa para a liberação de nitrato redutase para
o meio extracelular.
Figura 7 - Suspensão enzimática antes da filtração.

33
O material é filtrado em filtro de papel comum para eliminar o excesso de células que
podem deixar o meio turvo. Em seguida parte é filtrada em membrana de 0,45 µm para a
retenção de células.
Quantidades da solução de nitrato de sódio 315 mg N/L (estoque) foram adicionadas ao
extrato e após homogeneização iniciou a cinética enzimática.
Foram coletadas alíquotas de 1 mL do extrato com nitrato de sódio adicionado ao longo
do tempo (de hora em hora) sendo que ao término de cada tempo a alíquota coletada foi
mantida refrigerada ( 3 °C) para interromper a reação enzimática.
Em seguida foi feita a determinação do nitrito formado pela ação da nitrato redutase
através do Método de Griess, com auxílio de um espectrofotômetro Diode - Array Agilent
8354 A.
3.1.1.2 BIOMASSA DE FUSARIUM COMO FONTE ENZIMÁTICA
A biomassa foi pesada, triturada e transferida para Erlenmeyer onde água destilada ou
tampão foi adicionado na proporção de 10 g de biomassa para 100 mL de água ou diferentes
tipos de tampão
Quantidades da solução de nitrato de sódio (estoque) foram adicionadas ao extrato e
após homogeneização iniciou a cinética enzimática.
Foram realizados vários testes com a biomassa como fonte enzimática.
3.1.1.2.1 AVALIAÇÃO DA ATIVIDADE DA NITRATO REDUTASE
Para verificar a atividade da nitrato redutase em diferentes meios reacionais foram
utilizados os tampões fosfato 100 mmol/L a pH 6,0 e 7,2, tampão acetato a pH 5,0 e água.
Avaliou-se também a atividade da nitato redutase na presença de ácido ascórbico e glicose.
Foi adicionado 0,03 % (m/V) de glicose e diversas concentrações (0,1, 0,5 e 1% m/V) de
ácido ascórbico (vitamina C) em tampão fosfato 100 mmol/L pH 6,0. Após a homogeneização
o material foi centrifugado, sendo o precipitado ressuspendido em tampão fosfato 100
mmol/L pH 6,0 e adicionou-se solução de nitrato (estoque) ao precipitado e ao sobrenadante.

34
A atividade da nitrato redutase na presença de sulfito de sódio foi estudada. Ao meio
reacional (tampão fosfato 100 mmol/L pH 6,0) foi adicionado 0,1 % de sulfito de sódio. Para
a remoção do oxigênio dissolvido no meio reacional, preparado conforme descrito
anteriormente, borbulhou-se gás nitrogênio no meio reacional.
3.1.2 ESCHERICHIA COLI COMO FONTE DE NITRATO REDUTASE (FONTE
BACTERIANA)
3.1.2.1 Produção de nitrato redutase em condições aeróbicas de crescimento microbiano
e cinética enzimática
As colônias de Escherichia coli foram cultivadas em meio de cultura sólido. Foram
incubadas a 28 °C em meio aeróbio por 24 horas. Em seguida, com a alça bacteriológica, as
colônias foram transferidas e suspensas para o meio de cultura líquido . Foram incubadas a 28
°C em meio aeróbio por 3 dias na ausência de nitrato.
O meio líquido foi centrifugado e o precipitado foi ressuspendido com solução tampão
fosfato 100 mmol/L pH 6,0. Diferentes concentrações de nitrato foram adicionadas e após
homogeneização iniciou – se a cinética.
3.1.2.2 Produção de nitrato redutase em condições anaeróbicas de crescimento
microbiana e cinética enzimática
As colônias de Escherichia coli foram cultivadas em meio de cultura sólido. Foram
incubadas a 28 °C em meio aeróbio por 24 horas. Em seguida, com a alça bacteriológica, as
colônias foram transferidas e suspensas para o meio de cultura líquido, saturado previamente
com gás nitrogênio, a fim de se obter o crescimento bacteriano em meio anaeróbio.
Permaneceram a 28 °C por 3 dias na ausência de nitrato.
O meio líquido foi centrifugado e o precipitado foi ressuspendido com solução tampão
fosfato 100 mmol/L pH 6,0. Diferentes concentrações de nitrato foram adicionadas e após

35
homogeneização iniciou – se a cinética. Em seguida gás nitrogênio foi borbulhado no meio
reacional para expulsão do oxigênio por saturação para que a cinética prosseguisse em
anaerobiose.
3.1.3 TESTE UTILIZANDO VIGNA RADIATA L. COMO FONTE DE NITRATO
REDUTASE (FONTE VEGETAL)
As porções frescas de broto de feijão foram adquiridas diariamente no comércio local.
3.1.3.1 Avaliação da atividade da nitrato redutase com meio reacional com tampão
fosfato 100 mmol/L pH 6,0.
Uma porção de broto de feijão foi pesada e as partes foram picadas em tamanhos
uniformes. Adicionou-se 50 mL de tampão fosfato 100 mmol/L pH 6,0 e solução de nitrato
de sódio (estoque).
Para se ter certeza de que o nitrito detectado no meio reacional é produto da reação
enzimática da nitrato redutase com nitrato e não um produto liberado pelas células vegetais,
um branco foi feito colocando-se todos os reagentes exceto a solução de nitrato.
3.1.3.2 Avaliação da atividade da nitrato redutase utilizando o protetor
polivinilpirrolidona (PVP-VA)
Diferentes quantidades de PVP-VA foram adicionadas para que as concentrações sejam:
0,01 g/mL, 0,02 g/mL, 0,04 g/mL, 0,08 g/mL e 0,16 g/mL e quantidades da solução de nitrato
de sódio (estoque) foram adicionadas a fim de se obter as seguintes concentrações finais de
nitrato: 15 mgN/L, 10 mgN/L e 5 mgN/L.
3.1.3.3 Avaliação da atividade da nitrato redutase utilizando uma concentração de
0,04g/mL do protetor PVP-VA em diferentes pHs e concentrações de fosfato.

36
Para verificar o poder tamponante, diferentes concentrações de fosfato (50,100 e 200
mmol/L) em diferentes pHs (5,5; 6,0; 6,2; 7,0) foram requeridas a uma concentração fixa de
nitrato (15 mgN/L).
3.2 MATERIAIS
3.2.1 Fontes vegetais utilizadas
- Broto de feijão;
- Alface;
- Gengibre.
3.2.2 Obtenção das cepas utilizadas nos ensaios
Foram utilizadas para biossíntese de nitrato redutase as seguintes fontes obtidas da
coleção de cultura do laboratório de genética molecular da Escola Superior de Agronomia
Luis de Queiroz (ESALQ-USP) Piracicaba-SP:
- Fusarium oxysporum (cepas n° 07, n° 55.1 e n° 69);
-Fusarium cactus.
-Escherichia coli (DAUFPE 221)
3.2.3 Preparo dos meios de cultura
3.2.3.1 Meio de cultura sólido para Fusarium:

37
Pesou-se 0,5 g de extrato de levedura, 2 g de extrato de malte e 2 g de ágar e dissolveu-
se em 100 mL de água destilada. Autoclavou-se o meio (Vertical mo 415- FANEN) a 121°C
por 15 minutos e distribuiu-se em placas de Petri (20 mL) e após solidificação adicionou-se
ao mesmo um disco de micélio – ágar (este preparo foi realizado em câmara de fluxo
laminar).
3.2.3.2 Meio de cultura líquido para Fusarium:
Pesou-se 0,5 g de extrato de levedura, 2 g de extrato de malte e dissolve-se em 100 mL
de água. Autoclavou-se o meio foi e adicionou-se ao mesmo 3 discos de micélio - ágar.
3.2.3.3 Meio de cultura sólido para Escherichia coli
Pesou-se 4 g de triptona soja ágar e dissolveu-se em 100 mL de água. Autoclavou-se a
121°C por 15 minutos e distribuiu-se em placas de Petri (20 mL) e após solidificação colônias
de Escherichia coli inoculou-se ao meio (este preparo foi realizado em câmara de fluxo
laminar).
3.2.3.4 Meio de cultura líquido para Escherichia coli
Pesou-se 3 g de triptona soja caldo e dissolveu-se em 100 mL de água. Autoclavou-se o
meio a 121°C por 15 minutos e ao meio inoculou-se colônias provenientes do meio de cultura
sólido.
3.3 PREPARO DAS SOLUÇÕES

38
3.3.1 Reagente Espectrofotométrico para nitrito
Pesou-se 4 g de sulfanilamida (C6H8O2N2S) e 0,1 g de N-(1- naftil) - etilenodiamino
dihidrocloridrico e dissolveu-se em cerca de 30 mL água e 10 mL de ácido fosfórico
concentrado, em seguida completou-se o volume para 100 mL no balão volumétrico.
3.3.2 Solução de nitrato de sódio 315 mgN/L (estoque)
Pesou-se 0,1912 g de nitrato de sódio e dissolveu-se em cerca de 50 mL de água e em
seguida completou-se o volume para 100 mL no balão volumétrico
3.3.3 Tampão fosfato
3.3.3.1 Tampão fosfato 200 mmol/L
Pesou-se 2,756 g de NaH2PO4.H2O e dissolveu-se em água. Os valores de pH avaliados
foram: 5,5; 6,0; 6,2; 7,0; ajustados com NaOH ou HCl. Após ajuste do valor de pH
completou-se o volume para 100 mL no balão volumétrico.
3.3.3.2 Tampão fosfato 100 mmol/L
Pesou-se 1,3799 g de NaH2PO4.H2O e dissolveu-se com água, até aproximadamente 100
mL. Os valores de pH avaliados foram: 5,0; 5,5; 6,0; 6,2; 7,0; 7,2 e 8,0; ajustados com NaOH

39
ou HCl. Após ajuste do valor de pH completou-se o volume para 100 mL no balão
volumétrico.
3.3.3.3 Tampão fosfato 50 mmol/L
Pesou-se 0,6899 g de NaH2PO4.H2O e dissolveu-se em água. Os valores de pH
avaliados foram: 5,5; 6,0; 6,2; 7,0; o valor de pH foi ajustado com NaOH ou HCl. Após ajuste
do valor de pH completou-se o volume para 100 mL no balão volumétrico.
3.3.3.4 Tampão acetato 100 mmol/L pH = 5,0
Massa de 0,8203 g de CH3COONa foi dissolvida em água, adicionando 83 uL de HCl
concentrado (12 mol/L), ajustado o pH para 5,0 completou-se para 100 mL no balão
volumétrico.
3.3.4 Curva analítica para nitrito na faixa de concentração de 0,020 a 0,50 mg N/L
Pesou-se 0,9861 g de NaNO2 e dissolveu-se em água, completando o volume para
1000 mL no balão volumétrico (solução estoque 200 mgN/L).
Aos balões volumétricos de 100 mL foram adicionadas alíquotas, da solução estoque
200 mgN/L, de 100; 250; 500; 1000; 2000; 2500 uL sendo o volume completado com água.
Desta forma, foram obtidas soluções de concentrações 0; 020; 0,050; 0,10; 0,20; 0,40; 0,50
mgN/L, utilizadas na construção da curva analítica.
3.3.5 Curva analítica para amônia na faixa de concentração de 0,050 a 1,50 mg N/L
Pesou-se 3,819 g de NH4Cl e dissolveu-se em água, completando-se para 1000 mL no
balão volumétrico (solução estoque 1000 mgN/L).

40
Adicionou-se 10 mL dessa solução estoque 1000 mgN/L a um balão volumétrico e
completou-se para 100 mL com água, a fim de obter-se uma solução com concentração final
de 100 mgN/L de NH4Cl.
Aos balões volumétricos de 100 mL adicionou-se alíquotas, da solução estoque 100
mgN/L, de 50; 100; 250; 500; 1000; 1500 uL e completou-se o volume com água. Dessa
forma, obteve-se soluções de concentrações 0; 0,050; 0,10; 0,25; 0,50; 1,0; 1,5 mgN/L
utilizadas na construção da curva analítica.
3.3.6 Determinação de nitrito
A determinação de nitrito foi realizada com auxílio da reação de Griess. Uma alíquota
de 1 mL, retirada dos meios reacionais, foi misturada com 1 mL de reagente
espectrofotométrico, homogeneizada e mantida em banho-maria a 25˚C por 30 minutos. A
medida da absorbância foi realizada em 540 nm em um espectrofotômetro Agilent 8453 A. A
linha base foi corrigida nos seguintes comprimentos de onda: 540, 427, 445, 620, 648 e 685
nm.
3.4 REAGENTES
Os reagentes utilizados no desenvolvimento e aplicação do método para a
determinação de nitrato foram:
- Sulfanilamida (Synth)
- N-(1- naftil) - etilenodiamino dihidrocloridrico (Merck)
- Ácido fosfórico concentrado (J. T. Baker)
-Nitrato de sódio (Carlo Erba)
-Nitrito de sódio (Carlo Erba)

41
3.5 EQUIPAMENTOS
Os equipamentos utilizados nos procedimentos microbiológicos foram:
- Autoclave Vertical mod 415- FANEN
- Estufa BOD Q – 315D (QUIMIS ®)
- Centrifuga mod C.T. A 2815 . Para a determinação de nitrito foi empregado espectrofotômetro Agilent 8453.

42
4 RESULTADOS E DISCUSSÕES
A biossíntese de nitrato redutase e a otimização de sua atividade na redução do nitrato
foi estudada.
4.1 Fusarium como fonte de nitrato redutase
4.1.1 Extrato como fonte enzimática
Na Figura 8, através do método de Griess, apresenta-se a variação da concentração de
nitrito formado durante a incubação do extrato de duas espécies de fungos, F. oxysporum 55.1
e F.cactus, a partir de 6,0 mgN/L de nitrato. Observa-se um aumento contínuo da
concentração de nitrito no meio contendo extrato de F. oxysporum, atingindo-se um patamar a
partir das 72 horas de incubação, com uma eficiência de conversão de nitrato à nitrito de 30 %
e não foi observado um período de indução. Para a espécie F.cactus, foi observado um
máximo de concentração de nitrito atingindo um máximo de 24 horas. A eficiência observada
foi de 8,3 %. A eficiência foi calculada comparando a concentração de nitrito medida no meio
reacional (em mgN/L) com a concentração de nitrato inicialmente no meio (também em
mgN/L).

43
0
0 , 2
0 , 4
0 , 6
0 , 8
1
1 , 2
1 , 4
1 , 6
1 , 8
2
0 2 0 4 0 6 0 8 0 1 0 0 1 2 0T e m p o /h o r a s
Con
cent
raçã
o ni
trito
( m
gN/L
)
Figura 8 - Variação da concentração de nitrito formado em função do tempo. Meio formado por extrato filtrado em membrana 0.45 µm das espécies: F. cactus ( ) e F. oxysporum n° 55.1 ( ) . Concentração inicial de nitrato foi de 6,0 mgN/L.
4.1.2 Biomassa como fonte enzimática
4.1.2.1 Avaliação da atividade da nitrato redutase com meio reacional com água ou
tampão.
Na figura 9 é apresenta-se a variação da concentração de nitrito formado durante a
incubação da biomassa da espécie cactus a partir de 15 mgN/L de nitrato. Observa-se um
aumento contínuo da concentração de nitrito no meio contendo a biomassa da espécie cactus
atingindo-se um máximo em 38 horas. Observou-se uma eficiência de 33,3 %.

44
0
0 ,5
1
1 ,5
2
2 ,5
3
3 ,5
4
4 ,5
5
5 ,5
0 5 1 0 1 5 2 0 2 5 3 0 3 5 4 0
T e m p o /ho ras
Con
cent
raçã
o ni
trito
(mgN
/L)
Figura 9 - Variação da concentração de nitrito formado em função do tempo. Meio
formado pela biomassa da espécie cactus diretamente em contato com a solução de nitrato. Concentração inicial de nitrato 15 mgN/L.
Na avaliação da atividade da nitrato redutase com meio reacional com água ou tampão a
figura 10 mostra a variação da concentração de nitrito formado durante a incubação da
biomassa da espécie cactus a partir de 15 mgN/L de nitrato nos diferentes meios tamponados.
Observa-se um aumento contínuo da concentração de nitrito no meio contendo a biomassa da
espécie cactus atingindo-se um máximo em 27 horas. Observou-se uma eficiência de 77,1 %
para o meio não tamponado (água), 61,4 % para pH 6.0, 0,09 % para pH 5.0, e 10,86 % para
pH 7.2.
O ponto ótimo para a redução do nitrato parece estar próximo ao valor 6,0. Na solução
não tamponada (água) esse valor de pH pode ter sido atingido, obtendo-se então, uma maior
eficiência. A ausência de tamponamento é uma condição experimental não adequada, pois não
há controle.

45
0
2
4
6
8
1 0
1 2
1 4
0 1 0 2 0 3 0 4T em po / h o ras
Con
cent
raçã
o ni
trito
(mgN
/L)
0
Figura 10 - Variação da concentração de nitrito formado em função do tempo. Meio
formado pela biomassa da espécie cactus diretamente em contato com a solução de nitrato nos diferentes meios em : água ( ), pH 5,0 ( ♦ ), pH 6,0 ( ), pH 7,2 ( ). Concentração inicial 15 mgN/L.
4.1.2.2 Avaliação da atividade da nitrato redutase com diferentes proporções de
biomassa
A atividade da nitrato redutase proveniente da espécie cactus com diferentes proporções
de biomassa foi avaliada como mostra a figura 11 através da variação da concentração de
nitrito formado a partir de 15 mgN/L. Observa-se um aumento contínuo da concentração de
nitrito no meio contendo a biomassa da espécie cactus atingindo um máximo em 15 horas. A
eficiência observada foi de 50,5 % para 1g de biomassa (suspensa em 10 mL de tampão),
8,7% para 2 g de biomassa (suspensa em 10 mL de tampão) e 6% para 3 g de biomassa (
suspensa em 10 mL de tampão). A quantidade elevada de material orgânico em suspensão
pode ter apresentado problemas de transporte de massa na solução, determinando as
eficiências baixas com o aumento da massa de fungos no meio.

46
0,0
1,0
2,0
3,0
4,0
5,0
6,0
7,0
8,0
0 5 10 15 20 25 30
Tempo/ horas
Con
cent
raçã
o ni
trito
(mgN
/L)
Figura 11 - Variação da concentração de nitrito formado em função do tempo. Meio formado por diferentes proporções biomassa da espécie cactus diretamente em contato com a solução de nitrato. Concentração de nitrato inicial 15 mgN/L. Proporções: 1g ( ), 2g ( ) e 3g ( ).
4.1.2.3 Avaliação da atividade da nitrato redutase na presença de ácido ascórbico e
glicose
Na avaliação da influência de ácido ascórbico e glicose a figura 12 mostra a variação da
concentração de nitrito formado durante a incubação de biomassa da espécie cactus a partir de
15 mgN/L de nitrato na presença de 0,03% de glicose e 0,1% de vitamina C, e outro meio
contendo apenas 0,1% de vitamina C na solução de nitrato . Observa-se um aumento contínuo
da concentração de nitrito no meio contendo a biomassa da espécie cactus atingindo-se um
máximo em 20 horas. A eficiência observada foi de 63,4 % para o meio com glicose e
vitamina C, 53,4% e de 33,4% para o meio com vitamina C. Observa-se que a vitamina C
pode estar atuando como anti-oxidante auxiliando na estabilidade da enzima, já que neste
experimento a queda brusca do nitrito após atingir eficiência máxima diminuiu.

47
0 ,0
1 ,0
2 ,0
3 ,0
4 ,0
5 ,0
6 ,0
7 ,0
8 ,0
9 ,0
1 0 ,0
0 5 1 0 1 5 2 0 2 5 3 0T e m p o / h o ra s
Con
cent
raçã
o ni
trito
(m
gN/L
)
Figura 12 - Variação da concentração de nitrito formado em função do tempo. Meio
formado por biomassa da espécie cactus diretamente em contato com a solução tampão com nitrato e vitamina C, e em contato com solução tampão com nitrato, vitamina C e glicose nos meios centrifugados e ressuspedidos. Concentração inicial 15 mgN/L de nitrato. Sobrenadante-vitamina C ( ), ressuspendido-vitamina C ( ), sobrenadante-vitamina C e glicose ( ♦ ), ressuspendido-vitamina C e glicose ( ).
No caso da glicose, parece estar envolvida na redução do cofator NAD+(forma oxidada)
para NADH (forma reduzida necessária para que ocorra a atividade enzimática), conforme a
reação:
H2O + NAD+ + GLICOSE-6-P → 6-P-GLUCONATO + NADH + H+
Observa-se a relação entre a glicose e a regeneração parcial do cofator, pois NAD+
apresenta uma absorbância direta no UV de 260 nm, como pode ser verificado no
sobrenadante antes de iniciar a cinética, na Fig.13.

48
Figura 13 - Espectro do sobrenadante, antes de adicionar a solução de nitrato, glicose e ácido ascórbico. O espectro evidencia uma banda a 260 nm característica do NAD+.
Foi avaliada a influência da concentração de ácido ascórbico juntamente com a glicose
na atividade da nitrato redutase presente no sobrenadante e no ressuspendido da biomassa de
F.cactus.
Neste estudo, empregou-se diferentes concentrações de ácido ascórbico (0,1, 0,5 e 1%),
mantendo-se a concentração da glicose em 0,03%.
Para o sobrenadante, observa-se um aumento contínuo da concentração de nitrito para
todos os meios citados (fig. 14), com exceção do meio contendo 1% de ácido ascórbico. A
eficiência observada no sobrenadante foi de 47,7% com 0,1% de ácido ascórbico e 40% com
0,5% de ácido ascórbico.

49
0,0
1,0
2,0
3,0
4,0
5,0
6,0
7,0
8,0
0 5 10 15 20 25 30
Tempo/h
Con
cent
raçã
o ni
trito
(mgN
/L)
Figura 14 - Variação da concentração de nitrito formado em função do tempo. Meio
formado por biomassa da espécie cactus diretamente em contato com a solução tampão com nitrato e diversas concentrações de vitamina C no sobrenadante. Concentração inicial 15 mgN/L de nitrato.Concentrações de vitamina C: 0,1% ( ), 0,5% ( ), 1% ( ).
Para o ressuspendido (fig.15), a eficiência observada foi de 80% para 0,1% de ácido
ascórbico, 6,7% com 0,5% ácido ascórbico e 33,4% com 1% de ácido ascórbico.
Nos meios contendo 1% de ácido ascórbico pode ser que o mesmo esteja agindo como
inibidor, principalmente no caso do sobrenadante. Tanto no sobrenadante quanto no
ressuspendido, o máximo de formação de nitrito ocorreu em 25 horas.

50
0,0
2,0
4,0
6,0
8,0
10,0
12,0
14,0
0 5 10 15 20 25 30
Tempo /horas
Con
cent
raçã
o ni
trito
(mgN
/L)
Figura 15 - Variação da concentração de nitrito formado em função do tempo. Meio
formado por biomassa da espécie cactus diretamente em contato com a solução tampão com nitrato e diversas concentrações de vitamina C no ressuspendido. Concentração inicial 15 mgN/L de nitrato. Concentrações de vitamina C: 0,1% ( ), 0,5% ( ), 1% ( ).
4.1.2.4 Avaliação da atividade da nitrato redutase na presença de sulfito de sódio
Outra espécie redutora cuja influência na atividade da nitrato redutase foi avaliada
consistiu em sulfito de sódio. Foi realizada a incubação de biomassa da espécie cactus com 15
mgN/L de nitrato na presença e na ausência de sulfito de sódio. Observou-se um aumento
contínuo da concentração de nitrito apenas na ausência do sulfito atingindo um máximo em
24 horas (Figura 16), com uma eficiência de 60 % enquanto que no meio contendo sulfito
houve uma total inibição na redução do nitrato. A solução tampão fosfato 100 mmol/L pH 6,0
com biomassa sem que a solução de nitrato fosse adicionada é para garantir de que não existe
a presença de nitrito no meio, ou seja, o nitrito estará presente se houver redução do nitrato
adicionado ao meio pela ação enzimática da nitrato redutase.

51
0,0
1,0
2,0
3,0
4,0
5,0
6,0
7,0
8,0
9,0
10,0
0 5 10 15 20 25 30
Tempo /h
Con
cent
raçã
o ni
trito
(mgN
/L)
Figura 16 - Variação da concentração de nitrito formado em função do tempo. Meio formado por biomassa da espécie cactus diretamente em contato com a solução tampão com nitrato ( ), solução tampão com nitrato e sulfito ( ), e sol tampão apenas ( ♦ ). Concentração inicial 15 mgN/L de nitrato.
4.1.2.5 Avaliação da atividade da nitrato redutase na ausência de oxigênio
Na Figura 17 apresenta-se a variação da concentração de nitrito formado durante a
incubação de biomassa da espécie cactus a partir de 15 mgN/L de nitrato em meio reacional
saturado de nitrogênio para expulsão de oxigênio (possível agente oxidante). Observa-se um
aumento contínuo da concentração de nitrito no meio contendo a biomassa da espécie cactus.
Então, no meio saturado com nitrogênio atingiu-se um máximo em 30 horas e a eficiência
observada foi de 80 % .

52
0
2
4
6
8
10
12
14
0 10 20 30 4Tem po/ horas
Con
cent
raçã
o ni
trito
(mgN
/L)
0
Figura 17 - Variação da concentração de nitrito formado em função do tempo. Meio formado por biomassa da espécie cactus diretamente em contato com a solução tampão com nitrato em meio em anaerobiose (passagem de nitrogênio). Concentração inicial 15 mgN/L de nitrato.
Na Figura 18 é apresentada a variação da concentração de nitrito formado durante a
incubação de biomassa da espécie cactus a partir de 15 mgN/L , 5mN/L e 7,5 mgN/L de
nitrato, e a cepa 07 a partir de 15 mgN/L de nitrato, sendo que todos os meios estão saturados
de nitrogênio (pois o estudo anterior indicou ser melhor na ausência do oxigênio). Observa-se
um aumento contínuo da concentração de nitrito no meio contendo a biomassa da espécie
cactus atingindo um máximo em 24 horas. A eficiência observada foi de 80 % para o meio da
espécie cactus com 15 mgN/L, de 100% para a concentração 7,5 mgN/L e 90 % para a
concentração 5 mgN/L . Enquanto que a cepa 07 com 15 mgN/L apresentou uma eficiência
máxima de 42,7 %.

53
0
2
4
6
8
1 0
1 2
0 5 1 0 1 5 2 0 2 5 3 0 3 5te m p o /h
conc
entra
ção
nitri
to (m
gN/L
)
Figura 18 - Variação da concentração de nitrito formado em função do tempo. Meio formado por biomassa da espécie cactus diretamente em contato com a solução tampão com nitrato e diversas concentrações de nitrato em meio em anaerobiose. F.cactus: 15mgN/L ( ), 7,5 mgN/L ( ), 5 mgN/L ( ) e cepa 07 ( ).
Na Figura 19 apresenta-se a variação da concentração de nitrito formado durante a
incubação de biomassa da cepa n°69 a partir de 5 mgN/L de nitrato em meio reacional
saturado de nitrogênio para expulsão de oxigênio. Observa-se um aumento contínuo da
concentração de nitrito no meio contendo a biomassa da cepa n° 69 atingindo um máximo em
30 horas. A eficiência observada foi de 4 % para o meio em anaerobiose.

54
0,00
0,05
0,10
0,15
0,20
0,25
0 5 10 15 20 25 30Tempo/ horas
Con
cent
raçã
o ni
trito
(mgN
/L)
Figura 19 - Variação da concentração de nitrito formado em função do tempo. Meio formado por biomassa da cepa n° 69 diretamente em contato com a solução tampão com nitrato em meio em anaerobiose (passagem de nitrogênio). Concentração inicial 5 mgN/L de nitrato.
4.2. Escherichia coli como fonte de nitrato redutase
4.2.1 Produção de nitrato redutase em condições aeróbicas de crescimento microbiano e
cinética enzimática
Na Figura 20 apresenta-se a variação da concentração de nitrito formado durante a
incubação de células de E.coli a partir de 15 mgN/L , 5 mN/L , 3 mgN/L e 1 mgN/L de nitrato
em condições reacionais de aerobiose. Pode-se identificar a formação de um patamar de
concentração de nitrito no período analisado de 20 a 27 horas.

55
0 , 0
0 , 5
1 , 0
1 , 5
2 , 0
2 , 5
3 , 0
3 , 5
4 , 0
4 , 5
0 1 0 2 0T e m p o / h o r a s
Con
cent
raçã
o ni
trito
(mgN
/L)
3 0
Figura 20 - Variação da concentração de nitrito formado em função do tempo. Meio formado por células de E.coli suspensas em solução tampão com diversas concentrações de nitrato de em meio aerobiótico. Concentrações: 15 mgN/L ( ), 5 mgN/L ( ), 3 mgN/L( ) e 1 mgN/L ( ).
Uma curva analítica foi obtida lançando-se em gráfico a concentração de nitrito
determinada em função da concentração de nitrato adicionado ao meio. Observa-se, através da
inclinação da curva obtida, uma eficiência de 27% na conversão do nitrato a nitrito (Figura
21).

56
y = 0,2714x - 0,2223R2 = 0,9895
0
0,5
1
1,5
2
2,5
3
3,5
4
4,5
0 5 10 15 20
Concentração nitrato ( mg N/L)
Con
cent
raçã
o ni
trito
( m
g N
/L)
Figura 21 - Curva analítica da concentração de nitrito formado e a concentração de nitrato adicionado.
4.2.2 Produção de nitrato redutase em condições anaeróbicas de crescimento microbiana
e cinética enzimática
Na Figura 22 é apresentada a variação da concentração de nitrito formado durante a
incubação de células de E.coli da a partir de 15 mgN/L , 5 mN/L ,3 mgN/L e 1 mgN/L de
nitrato em condições reacionais de anaerobiose, onde pode-se observar uma eficiência muito
baixa.

57
0 ,0
0 ,1
0 ,2
0 ,3
0 ,4
0 ,5
0 ,6
0 ,7
0 ,8
0 ,9
0 5 1 0 1 5 2 0 2 5 3 0T e m p o / h o ra s
Con
cent
raçã
o ni
trito
(mgN
/L)
Figura 22 - Variação da concentração de nitrito formado em função do tempo. Meio
formado por células de E.coli suspensas em solução tampão com diversas concentrações de nitrato de em meio anaerobio. Concentrações: 15 mgN/L ( ), 5 mgN/L ( ), 3 mgN/L( ) e 1 mgN/L ( ).
4.3 Testes utilizando fontes vegetais de nitrato redutase
As fontes vegetais como alface e gengibre embora tenham apresentado atividade
foram descartadas devido à dificuldades na obtenção de extratos limpos, o que foi possível
utilizando broto de feijão.
4.3.1 Teste utilizando broto de feijão como fonte de nitrato redutase
4.3.2 Avaliação da atividade da nitrato redutase no meio reacional com tampão apenas.

58
Na Figura 23 é apresentada a variação da concentração de nitrito formado durante a
incubação de tecido vegetal a partir de 15 mgN/L de nitrato e tampão. Observa-se um
aumento contínuo da concentração de nitrito formado no meio contendo tecido vegetal
atingindo um máximo em apenas 4 horas. A eficiência observada foi de 28,07 %.
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
4,0
4,5
0 50 100 150 200 250 300 350Tempo/ minutos
Con
cent
raçã
o ni
trito
(mgN
/L)
Figura 23 - Variação da concentração de nitrito formado em função do tempo. Meio formado por tecido vegetal triturado suspenso em solução tampão sem nitrato () e com nitrato ().
4.3.3 Avaliação da atividade da nitrato redutase utilizando o protetor PVP-VA.
Na Figura 24 apresenta-se a variação da concentração de nitrito formado durante a
incubação de tecido vegetal a partir de 15 mgN/L de nitrato, tampão e polivinilpirrolidona
(PVP). Observa-se um aumento contínuo da concentração de nitrito no meio contendo tecido
vegetal atingindo um máximo em 3 horas. Já houve um aumento pois a eficiência observada
foi de 37,8 % .

59
0
1
2
3
4
5
6
0 2 4 6T e m p o / h o ra s
Con
cent
raçã
o ni
trito
(mgN
/L)
8
Figura 24 - Variação da concentração de nitrito formado em função do tempo. Meio
formado por tecido vegetal triturado suspenso em solução tampão apenas () ou contendo: PVP () e PVP com nitrato ().
4.3.4 Avaliação da atividade da nitrato redutase utilizando o protetor PVP-VA com
diferentes concentrações de nitrato.
Na Figura 25 é apresentada a variação da concentração de nitrito formado durante a
incubação de tecido vegetal a partir de 15 mgN/L ,10 mgN/L e 5 mgN/L de nitrato, tampão e
0,01g/mL polivinilpirrolidona (PVP). Observa-se um aumento contínuo da concentração de
nitrito no meio contendo tecido vegetal atingindo um máximo em 3,5 horas. A eficiência
observada foi de 34,7 % para 15 mg N/L.

60
0
1
2
3
4
5
6
0 5 0 1 0 0 1 5 0 2 0 0 2 5 0 3 0 0T e m p o / m in
Con
cent
raçã
o ni
trito
(mgN
/L)
Figura 25 - Variação da concentração de nitrito formado em função do tempo. Meio formado por tecido vegetal triturado suspenso em solução tampão com diversas concentrações de nitrato: 15 mgN/L ( ), 10 mgN/L ( ), 5 mgN/L ( )
4.3.5 Avaliação da atividade da nitrato redutase utilizando diferentes concentrações do
PVP-VA uma concentração fixa de 15 mgN/L de nitrato.
Na Figura 26 apresenta-se a variação da concentração de nitrito formado durante a
incubação de tecido vegetal a partir de 15 mgN/L de nitrato, tampão fosfato 100 mmol/l pH
6,0 e diversas concentrações de polivinilpirrolidona (PVP). Observa-se um aumento contínuo
da concentração de nitrito no meio contendo tecido vegetal atingindo um máximo em 4 horas.
A eficiência observada foi de 46 %,49% e 45% para 0,04 g/mL, 0,08 g/mL e 0,16 g/mL de
PVP, respectivamente. Esse comportamento pode ser explicado, pois a faixa de eficiência
máxima para essas concentração de PVP neste experimento estão muito próximas.
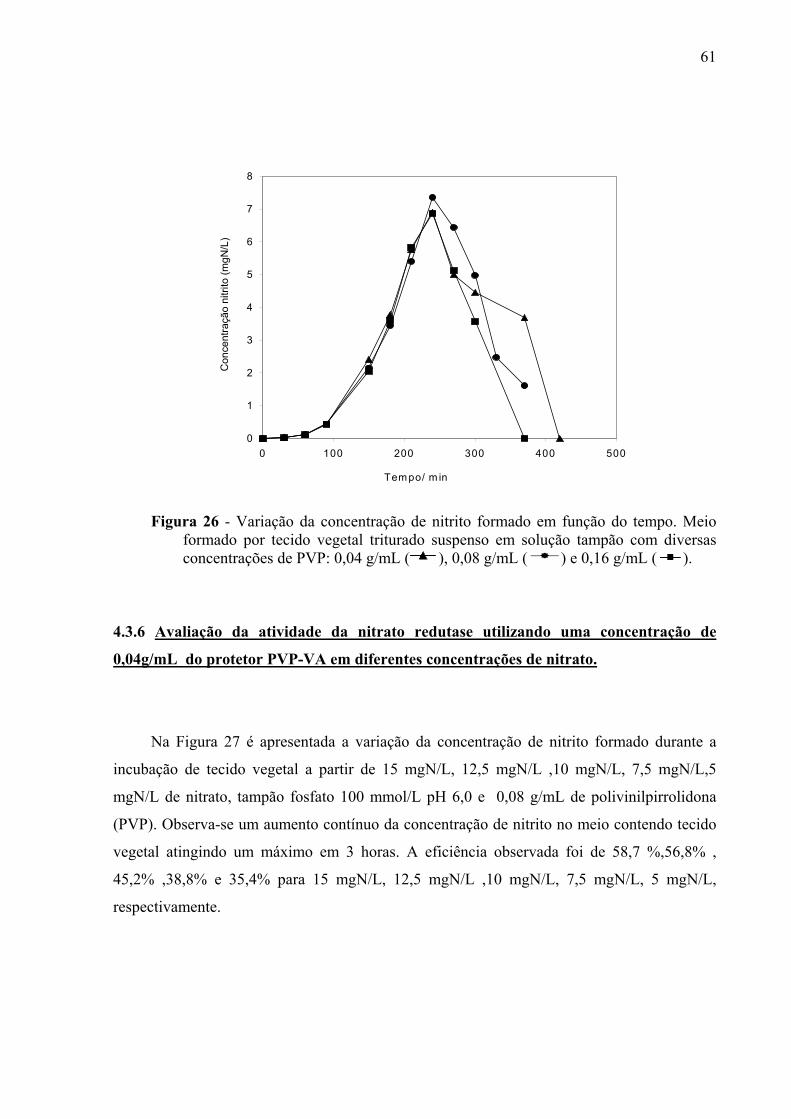
61
0
1
2
3
4
5
6
7
8
0 100 200 300 400 500
Tem po/ m in
Con
cent
raçã
o ni
trito
(mgN
/L)
Figura 26 - Variação da concentração de nitrito formado em função do tempo. Meio formado por tecido vegetal triturado suspenso em solução tampão com diversas concentrações de PVP: 0,04 g/mL ( ), 0,08 g/mL ( ) e 0,16 g/mL ( ).
4.3.6 Avaliação da atividade da nitrato redutase utilizando uma concentração de
0,04g/mL do protetor PVP-VA em diferentes concentrações de nitrato.
Na Figura 27 é apresentada a variação da concentração de nitrito formado durante a
incubação de tecido vegetal a partir de 15 mgN/L, 12,5 mgN/L ,10 mgN/L, 7,5 mgN/L,5
mgN/L de nitrato, tampão fosfato 100 mmol/L pH 6,0 e 0,08 g/mL de polivinilpirrolidona
(PVP). Observa-se um aumento contínuo da concentração de nitrito no meio contendo tecido
vegetal atingindo um máximo em 3 horas. A eficiência observada foi de 58,7 %,56,8% ,
45,2% ,38,8% e 35,4% para 15 mgN/L, 12,5 mgN/L ,10 mgN/L, 7,5 mgN/L, 5 mgN/L,
respectivamente.

62
0
1
2
3
4
5
6
7
8
9
1 0
0 5 0 1 0 0 1 5 0 2 0 0 2 5 0 3 0 0 3 5 0
T e m p o / m in
Con
cent
raçã
o ni
trito
(mgN
/L)
Figura 27 - Variação da concentração de nitrito formado em função do tempo. Meio formado por tecido vegetal triturado suspenso em solução tampão com 0,08 g/mL de PVP e diversas concentrações de nitrato: 15 mgN/L ( ), 12,5 mgN/L ( ), 10 mgN/L ( ), 7,5 mgN/L ( ), 5 mgN/L ( ).
A partir dos dados da figura 27, uma curva analítica foi traçada que poderá ser utilizada
na determinação da concentração de nitrato numa possível amostra a ser analisada.

63
y = 0 ,7307x - 2 ,2844R 2 = 0 ,983
0
1
2
3
4
5
6
7
8
9
1 0
0 5 10 15 20
C oncentração ( m gN /L)
Conc
entr
ação
( m
gN/L
)
Figura 28 - Curva analítica da concentração de nitrito formado em função da concentração de nitrato adicionado.
4.3.7 Avaliação da atividade da nitrato redutase utilizando uma concentração de
0,04g/mL do protetor PVP-VA em diferentes concentrações de nitrato e pHs.
Na figura 29 é apresentada a variação de nitrito formado durante a incubação de tecido
vegetal a partir de 15 mgN/L, 10 mgN/L e 5 mgN/L de nitrato, 0,04 g/mL de
polivinilpirrolidona (PVP) em diferentes pHs. Observa-se a melhor eficiência para a maioria
dos pHs em 6,0.

64
0
1
2
3
4
5
6
7
8
9
1 0
4 4 , 5 5 5 , 5 6 6 , 5 7 7 , 5 8p H
Con
cent
raçã
o ni
trito
( m
gN/L
)1 5 m g N / L1 0 m g N / L5 m g N / L
Figura 29 - Variação da concentração de nitrito em função do pH do meio reacional, considerando os valores de eficiência máxima para as de nitrato adicionadas
4.3.8 Avaliação da atividade da nitrato redutase utilizando uma concentração de
0,04 g/mL do protetor PVP-VA em diferentes pHs e concentrações de fosfato.
Para verificar a influência do poder tamponante apresenta-se na, Figura 30, a variação
de nitrito formado durante a incubação de tecido vegetal a partir de 15 mgN/L de nitrato; 0,04
g/mL de polivinilpirrolidona (PVP) em diferentes pHs e concentrações de fosfato. Observa-se
a melhor eficiência (49,6%) para pH 5,5 em fosfato 50 mmol/L, sendo a segunda melhor
eficiência (46,6%) para pH 6,2 em fosfato 100 mmol/L.

65
0
1
2
3
4
5
6
7
8
0 50 100 150 200 250
Concentração de fosfato (mmol/L)
Con
cent
raçã
o de
nitr
ito (m
gN/L
)
Figura 30 - Variação da concentração de nitrito em função da concentração de fosfato,
considerando os valores de eficiência máxima para os diferentes pHs a uma concentração fixa de nitrato adicionada. Legenda dos pHs 5,5 ( ), 6,0 ( ), 6,2 ( ) e 7,0 ( ).
Esses dados também foram representados pela eficiência em função do pH (Figura 31).
Observa-se uma diminuição da eficiência em tampão fosfato 200 mmol/L e 50 mmol/L
quando o pH aumenta, enquanto em tampão fosfato 100 mmol/L ao aumentar o pH até 6,2
ocorre aumento da eficiência. Esse comportamento pode ser explicado devido a variação da
força iônica.

66
0
1
2
3
4
5
6
7
8
5 5,5 6 6,5 7 7,5
pH
Con
cent
raçã
onitr
ito (m
gN/L
)
Figura 31 - Variação da concentração de nitrito em função do pH, considerando os valores de eficiência máxima para as diferentes concentrações de fosfato no tampão a uma concentração fixa de nitrato adicionada.Legenda das concentrações de fosfato 50 mmol/L (♦), 100 mmol/L (), 200 mmol/L ().
5 CONCLUSÕES E ETAPAS FUTURAS
A partir dos estudos realizados com três tipos distintos de extratos brutos pode-se
resumir suas principais características.
5.1 Fusarium
Na avaliação da nitrato redutase de Fusarium, na redução enzimática do nitrato para fins
analíticos, embora a conversão tenha chegado até 100% o mesmo apresentou baixa
linearidade e diversas desvantagens como:

67
- Tempo de 10 dias necessário para o crescimento da biomassa;
- O período de incubação de no mínimo 20 hs;
- Necessidade de equipamentos microbiológicos (muitas vezes ausentes nos laboratórios
de química analítica);
- O preparo e o custo dos meios de cultura;
- Está suscetível a contaminações;
- O descarte precisa ser auto-clavado;.
5.2 E.coli
Quando se utilizou nitrato redutase de E.coli o método apresentou boa linearidade, baixa
turbidez, mas apresentou eficiência reduzida de 27% e diversas desvantagens microbiológicas
assim como Fusarium. O cuidado deve ser ainda maior, pois E.coli é uma enterobacteria
capaz de contaminar os analistas.
5.3 Vigna radiata L.
Quando se utilizou nitrato redutase de broto de feijão o método apresentou boa
linearidade, baixa turbidez, boa eficiência 73% e muitas vantagens quando comparados com
as fontes de nitrato redutases microbiológicas:
- Preparo rápido e muito fácil;
- Obtido facilmente no comércio local;
- O tempo de reação é pequeno, de no máximo 6 hs;
- Os equipamentos utilizados são simples, facilitando o uso dessa fonte em laboratório
de análises clínicas;
- É inócuo para os analistas;
- O descarte é simples.
Desta forma observa-se que o método de análise utilizando nitrato redutase de broto de
feijão para a determinação de nitrato mostrou-se promissor para o monitoramento desta

68
espécie em amostras de água, com as seguintes vantagens: emprego de reagentes não
carcinogênicos, não adiciona cofatores (uma vez que os extratos brutos fornece os mesmos),
baixo consumo de reagentes, extremamente econômico, não gera resíduos tóxicos.
Etapas Futuras
- Padronizar protocolos de análise visando aumentar a repetibilidade do método;
-Validação do método (comparar com outros métodos utilizando águas de efluentes de
estação de tratamento de água);
-Estudar fontes similares;
-Uso de extrato bruto aplicado em análise por injeção em fluxo, substituindo a coluna de
cádmio.

69
Referências
ANDERSON,L. Simultaneous spectrofotometric determination of nitrite and nitrate by flow injection analysis. Analytica Chimica Acta, v. 110, p. 123-128, 1979. ANDRADE, R. Desenvolvimento de métodos analíticos para determinação de nitrato, nitrito e n-nitrosaminas em produtos cárneos. Campinas, p.11, 2004. Tese de doutorado. BARBIER, G.C.; CAMPBELL, W.H.Viscosity Effects on Eukaryotic Nitrate Reductase Activity. The Journal of Biological Chemistry, v. 280, n. 28, p. 26049–26054, 2005. BORCHERDING, H.; LEIKEFILD, S.; FREY, C.; DIEKMANN, S.; STEINRIICKE, P.Enzymatic microtiter plate-based nitrate detection in environmental and medical analysis. Analytical Biochemistry, v. 282, p. 1-2, 2000. BOYER, P.D.; LARDY, H.; MYRBACK. The enzymes, v.7, 2 .ed. New York and London:Academic Press, cap.25, p. 591-592,1963. CAMPBELL , E. R; CAMPBELL, W. H. Nitrate measurement whit biosensor tecnology. Appalachian rivers II, v. 28-29, p.1-5, 1999. CAMPBELL, E.R.; PATTON, C.J.; CAMPBELL, W.H.; KRYSKALLA, J.R. Nitrate analysis using environmentally benign nitrate reductase methods. NWQMC, p. 1-9, 2002. CAVICCHIOLI, A.; GUTZ, I.G.R. Uso da radiação ultravioleta para o pré-tratamento de amostras em análises inorgânicas. Quimica Nova, vol. 26, n. 6, p. 919-921, 2003. CHOW, F.; OLIVEIRA, M.C.; PEDERSEN, M. In vitro assay and light regulation of nitrate reductase in red alga Gracilaria chilensis. Journal of Plant Physiology, v. 161, p. 769–776, 2004. CLESCERI, L.S.; GREENBERG, A.E.; EABN, A.D., APHA – AMERICAN PUBLIC HEALTH ASSOCIATION In: Standard methods for the Examination of Water and Waste Water, Washington : APWA,AWWA,WEF, 20 . ed., cap. 4, p.112-116, 1998. COSS, A.; CANTOR, K. P; REIF, J. S.; LYNCH, C. F.; WARD, M. H. Pancreatic Cancer and Drinking Water and Dietary Sources of Nitrate and Nitrite. American Journal of Epidemiology, v.159, n.7, p.693-701, 2004. COTRAN, R.S.; KUMAR, V.; COLLINS, T., Robbins Patologia estrutural e funcional, 6 .ed., Rio de Janeiro: Guanabara Koogan, cap.10, p. 376-378, 2000. DEMBNY,K.D.; ROSA, A.M.; JOHNSON, C.; ADAMS, M.B.; PIEPER, G.M. Heparin interferes with the determination of plasma nitric oxide by inhibition of enzymatic conversion of nitrate to nitrite by nitrate reductase.Clinica Chimica Acta, v. 275, p.107-114, 1998.

70
DOMBROWSKI, L. J.; PRATT, E. J. Fluorometric method determining nanogram quantities of nitrite ion. Analytical Chemistry, v. 44, n. 14, p. 2268-2272, 1972. FANQUIN,V; ANDRADE, A.T. Nutrição mineral e diagnose do estado nutricional de hortaliças. Lavras: UFLA/FAEPE, p. 88, 2004. Disponível em:< File;//A:\ EFEITOS DO NITRITO.htm>. Acesso em: 05 de março de 2005. FERREIRA, F.N.; MONTEIRO, M.I.C.; OLIVEIRA, N.M.M. Determinação de nitrato em águas pelo método simplificado do salicilato, p.76-83. Disponível em: < http://www.cetem.gov.br/publicacao/serie_anais_X_jic_2002/Fernanda.pdf > Acesso em: 10 de setembro de 2006. FERREYRA, N.F; SOÝ´S, V.M. An amperometric nitrate reductase –phenosafranin electrode:kinect aspects and analytical applications. Bioelectrochemistry, v. 64, p. 61-70, 2004. FINAN, A.; KEENAN,P.; O´ DONOVAN, F.O.; MAYNE, P.; MURPHY, J.. Methaemoglobinaemia associated with sodium nitrite in three siblings. BMJ.Clinical Review, v. 317, p. 1138-1139, 1998. FOX, J.B. The determination of nitrite: a critical review. CRC Critical Reviews in Analytical Chemistry, v.15, cap.3, p. 283-313, 1983. FRIEDBERG, M.A; HINSDALE, M.E.; SHIHABI, K. Analysis of nitrate in biological fluids by capillary electrophoresis. Journal of Chromatography A, n. 781, p. 491-496, 1997 FURLANI, P. R.; COMETTI, N. N. Segurança alimentar e produtos hidropônicos. Disponível em:< http://www.labhidro.cca.ufsc.br/seguranca_alimentar_NITRATO.htm> Acesso em: 10 de setembro de 2006. GEFFNER, M.E.; POWARS, D.R.; CHOCTAW, W.T.,Acquired Methemoglobinemia.The Western Journal of Medicine, v. 134,n. 1, p.7-10, 1981. GEWITZ, H.S.; PIEFKE, J.; VENNESLAND, B. Purification and characterization of demolybdo nitrate reductase ( NADH-cytochrome c oxidoredutase) of Chlorella vulgaris. The Journal of Biological Chemistry, v. 256, n. 22, p.11527-11531, 1981. GRANGER, D. L.; TAINTOR, R.R.; BOOCKVAR, K.S.; HIBBIS, J.B. Measurement of nitrate and nitrite in biological samples using nitrate reductase and Griess reaction. Methods in Enzymology , v. 268, n. 13, p. 142-151, 1996. GRISHAM, M.B; JOHNSON, G. G.; LANCASTER, J.R.Quantitation of nitrate and nitrite in extracellular fluids. Methods in enzymology, v. 268, n. 23, p.237-246,1996. HOFF, T.; STUMMAN, B.M.; WENNINGSEN, K.W. Struture, formation and regulation of nitrate reductase in higher plants. Physiology Plantarum,v. 84, p.616-624,1992. JEPSON, B.J.N.J; ANDERSON, L.J.; RUBIO, L.M.; TAYLOR, C.J.; BUTLER, C.S.; FLORES, E.; HERRERO, A., BUTT, J.N.; RICHARDSON, D.J.. Tuning a nitrate

71
reductase for function. The Journal the Biological Chemystry, v. 279, n. 31, p. 32212-32218, 2004. KHAN, A.; SARKAR, D. Identi.cation of a respiratory-type nitrate reductase and its role for survival of Mycobacterium smegmatis in Wayne model. Microbial Pathogenesis, v. 41, p. 90–95, 2006. KIANG,C.H; KUAN,S.S; GUIBAULT,G.G. Enzymatic determination of nitrate: Eletrochemical detection after reduction with nitrate reductase and nitrite reductase. Analytical Chemistry, v. 50, n. 9, p.1319-1322, 1978. LI, H.; MEININGER, C. J.; WU, G. Rapid determination of nitrite by reversed-phase high-performance liquid chromatography with fluorescence detection. Journal of Chromatography B, n.746 , p.199-207, 2000. LILLO, C.; MEYERS, C.; LEA, U. S.; PROVAN, S.; OLTE dal, S. Mechanism and importance of post-translational regulation of nitrate reductase. Journal of Experimental Botany, v. 55, n. 401, p. 1275-1282, 2004. LIMA, V. L. A.G. ; MELO, E. A.; MACIEL, M.I.S; SILVA, G.S.B.; LIMA, D.E.S., Fenólicos totais e atividade antioxidante do extrato aquoso de broto de feijão-mungo (Vigna radiata L.), Revista de Nutrição, n. 17, p. 53, 2004.
LORENZI, T. F., Manual de Hematologia, 2. ed. São Paulo: Medsi, cap. 3, p. 294-295, 1999.
LUNDBERG, J. O.; GOVONI, M. Inorganic nitrate is a possible source for systemic generation of nitric oxide. Free Radical Biology & Medicine, v. 37, n. 3, p. 395-400, 2004
MACGREGOR, C.H.; SCHAITMAIN,C..A. ;.NORMANSELL, D.E. Purification and properties of nitrate reductase from Escherichia coli. The Jounal of Biological Chemistry, v. 249, n.16, p. 5321-5327, 1974. MANTOVANI, J.; CRUZ, M.C.P.; FERREIRA, M.E.; BARBOSA, J.C. Comparação de procedimentos de quantificação de nitrato em tecido vegetal. Pesquisa Agropecuária Brasileira, Brasília, v. 40, n.1, p.54-59, 2005. MARTINEZ-ESPINOSA, R.M; MARHUENDA-EGEA, F.C, BONETE, M.J. Assimilatory nitrate reductase from the haloarchaeon Haloferax mediterranei: purification and characterisation.FEMS Microbiology Letters, v. 204, p.381-385, 2001. MIKAMI, B.; IDA, S. Purification and properties of ferredoxin-nitrate reductase from the Cyanobacterium Plectonema boryanum. Biochimica et Biophysica Acta, v. 791, p. 294-304,1984. MORPETH, F.F.; BOXER, D.H. Kinetic analysis of respiratory nitrate reductase from Escherichia coli K 12. Biochemistry, v. 24 , p.40-46, 1985.

72
MOUREAUX, T.; LEYDECKER, M.T.; MEYER, C. Purification of nitrate reductase from Nicotiana plumbaginifolia by affinity chromatography using 5’ AMP-sepharose and monolonal antibodies. European Journal of Biochemistry, v. 179 , p.617-620, 1989. MURRAY, K.D.; GRANNER, D.K.; MAYES, P.A.; RODWELL,V.W. Haper: Bioquímica, 8.ed. São Paulo: Atheneu , cap. 60, p. 738-739, 1998. NOLLET, L. M. L. Handbook of Water Analysis, 2. ed., New York: Marcel Dekker, cap. 12, p. 2001-217, 2000. OLIVEIRA, M. A. J.; BOVI, M. L. A.; MACHADO, E. C. ; RODRIGUEZ, J. D. Atividade da redutase de nitrato em mudas de pupunheira (Bactris gasipaes). Ciência Rural, v.35, n.3, p.515-522, 2005. PATTON, C. P.; FISHER, A.; CAMPBELL, W.; CAMPBELL, A.R., Corn leaf nitrate reductase – a nontoxic alternative to cadmium for photometric nitrate determinations in water samples by-air segmented continuous flow analysis. Environmental Science & Technology, v.36, n. 4, p. 729-735, 2002. PEREIRA, J.F. LIMA, J.O.; ROCHA, R.B.; MEDINA, E.F.; QUEIROZ, M.V. Nitrato redutase em fungos filamentosos. Revista de Biotecnologia Ciência e desenvolvimento. 31. ed.,p.74-85, 2003. Disponível em:<http://www.biotecnologia.com.br/revista/bio31/nitrato_red.pdf> Acessso em: 10 de setembro de 2006. POONNACHIT,U.; DARNELL, R. Effect of and nitrate on ferric chelate reductase and nitrate reductase in Vaccinium species. Annals of Botany ,v. 93 , p. 399-405, 2004 RIDLEY, H.; WATTS, C.A.; RICHARDSON, D.J.; BUTLER, C.S. Resolution of Distinct Membrane-Bound Enzymes from Enterobacter cloacae SLD1a-1 That Are Responsible for Selective Reduction of Nitrate and Selenate Oxyanions. Applied and Environmental Microbiology, v. 72, n. 8, p. 5173–5180, 2006. SANCHES, P.J.S.T.;ZANIN, K.D; CAMARÃO, E.B; GARCIA, R.C; RIOS, A;VALCARCEL,M. Pré concentração de nitrosaminas a partir de amostras aquosas por extração em fase sólida e cromatografia capilar eletrocinética micelar. Química Nova, v. 26, n. 2, p.193-196, 2003. SANTOS, J.; BECK, L.; WALTER, M.; SOBCZAK, M.; OLIVO, C.J.; COSTABEBER, I.; EMANUELLI, T. Nitrato e nitrito em leite produzido em sistema convencional e orgânico. Ciência e Tecnologia de Alimentos, v. 25, p. 304-309, 2005. SCHIMIDT, H. H.H.W. Determination of nitric oxide via measurement of nitrite and nitrate in culture media. Biochemica , n .02, p. 22, 1995. SILVA, P. Farmacologia, 5. ed., Rio de Janeiro : Guanabara Koogan, cap. 62, p. 606-607, 1998.

73
SOLOMONSON, L.P. ; BARBER, M.J. Assimilatory nitrate reductase: functional properties and regulation. Annual Review of Plant Physiology. Plant Molecular Biology, v. 41, p. 225-253, 1990. SOLOMONSON, L. P. ; LORIMER, G.H.; HALL, R.L.; BORCHERS, R.; BAILEY, J.L. Reduced nicotinamide adenine dinucleotide-nitrate reductase of Chlorella vulagaris. The Journal of Biological Chemistry, v. 250 , n.11, p. 4120-4127, 1975. TURRA, M.; AYUB, M.A.Z.. Estudo da variação do teor de nitritos e nitratos em embutidos coloniais: possíveis implicações para a saúde pública. Revista do Instituto Adolfo Lutz, v. 58, n. 2 , p.113-120, 1999. UNKLES, S.E.; WANG, R.; WANG, Y.; GLASS, A.D.M.; CRAWFORD, N.M.; KINGHORN, J.R. Nitrate Reductase Activity Is Required for Nitrate Uptake into Fungal but Not Plant Cells. The Journal of Biological Chemistry, v. 279, n. 27, p. 28182-28186, 2004. VAN´T RIET, J., PLANTA, R.J. Purifiaction, struture and properties of respiratory nitrate reductase of Klebsiella aerogenes. Biochimica et Biophysica Acta, v. 379 , p. 81-94, 1975. XUEIJUANG, W.; DZYADEVYCH, V.; CHOVELON, J.M.; RENAULT, N.J.; LING, C.; SIQING, X.; JIANFU, Z., Conductometric nitrate biosensor based on methyl viologen / Nafton ®/ nitrate reductase interdigitated electrodes. Talanta , v. 69, p. 450–455, 2006. YANG, G. C. C.; LEE, H. L. Chemical reduction of nitrate by nanosized iron:kinetics and pathways. Water Research, v. 39, p. 884–894, 2005. WARD, M.H.; DEKOK, T. M.; LEVALLOIS, P.; BRENDER, J.; GULIS, G.; NOLAN, B.T.; VANDERSLICE, J. Drinking-water nitrate and health - recent findings and research needs. Environmental Health Perspectives, v. 113 , n.11, p.1608-1614, 2005. ZEMAN, C.L.; KROSS, B.; VLAD, M., A nested case-control study of methemoglobinemia risk factors in children of Transylvania, Romania. Environmental Health Perspectives, v.110, n. 8, p.817-822, 2002. ZHU, X.; HO, CHOO, K.;PARK, J. M. Nitrate removal from contaminated water using polyelectrolyte-enhanced ultrafiltration. Desalination, v. 193, p. 350–360, 2006.

74

Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo