unit 3. basic of biopolymers (3) control of protein function spectroscopy of biopolymers
TRANSCRIPT

Unit 3. Basic of Biopolymers (3) Control of Protein
Function
Spectroscopy of Biopolymers

Major MechanismMajor Mechanismss of Protein Regulation of Protein Regulation
• Controlled by localization localization of the gene product or the species it interacts.
• Controlled by the covalent covalent or noncovalentnoncovalent binding of effector molecules.
• Controlled by the amount amount and lifetimelifetime of the active protein.

Proteins can be targeted to specific compartments
and complexesProtein is only present in its active form in the specific compartment where it is needed, or when bound in a complex with other macromolecules that participate in its function.
Localization Specification
targeted to cellular compartments by signal sequences or by attachment of a lipid tail that inserts into membranes.
directed to a complex of interacting proteins by a structural interaction domain
Localization is a dynamic process and a given protein may be targeted to different compartments at different stages of the cell cycle

Interaction domainA protein domain that recognizes another protein, usually via a specific recognition motif.
Interaction domains The name of the particular example shown for each family is given below each structure, along with the function and specificity of the domain.

• Protein function is modulated by the environment in which the protein operates
• Changes in redox environment can greatly affect protein structure and function
• Changes in pH can drastically alter protein structure and function
Control by pH & Redox environment
Cathepsin D conformational switching by pH
At neutral pH
Avtive siteis blocked by the N-terminal segment.
At low pH
The active site is opened by reorientation of the N-terminal segment.

Protein activity can be regulated by binding of an effectoreffector and by covalent modification
• Protein activity can also be controlled by the binding of effector molecules, which often work by inducing conformational changes that produce inactive or active forms of the protein.

Effector LigandEffector Ligand
• Effector ligand: a ligand that induces a change in the properties of a protein.
• Effectors may be as small as a proton or as large as another macromolecule.
• Effectors may bind noncovalently or may modify the covalent structure of the protein, reversibly or irreversibly.
• Effectors that regulate activity by binding to the active site usually take the form of inhibitors that compete with the substrate for binding.

EffectorCompetitive Binding and Cooperativity
• cooperative binding: interaction between two sites on a protein such that the binding of a ligand to the first one affects the properties of the second one.
•positive cooperativity the first ligand molecule to bind is bound weakly, but its binding alters the conformation of the protein in such a way that binding of the second and subsequent ligand molecules is promoted. •negative cooperativity the first ligand binding weakens and thereby effectively inhibits subsequent binding to the other sites.

Cooperative Ligand Binding
1. Protein function can be controlled by effector ligands that bind competitively to ligand-binding or active sites
2. Cooperative binding by effector ligands amplifies their effects

Effector Binding and AllosteryEffector Binding and Allostery
Effector molecules can cause conformational changes at distant sites
allostery: the property of being able to exist in two structural states of differing activity. The equilibrium between these states is modulated by ligand binding.
•allosteric activator: a ligand that binds to a protein and induces a conformational change that increases the protein’s activity.•allosteric inhibitor: a ligand that binds to a protein and induces a conformational change that decreases the protein’s activity.

Ligand-induced conformational change activates aspartate transcarbamoylase

Iron binding regulates the repressor of the diphtheria toxin geneComparison of the structures of the aporepressor DtxR (red, left, PDB 1dpr) and the ternary complex (right) of repressor (green), metal ion (Fe2+, orange) and DNA (grey) (PDB 1fst). Iron binding induces a conformational change that moves the recognition helices (X) in the DtxR dimer closer together, providing an optimal fit between these helices and the major groove of DNA. In addition, metal-ion binding changes the conformation of the amino terminus of the first turn of the amino-terminal helix (N) of each monomer. Without this conformational change, leucine 4 in this helix would clash with a phosphate group of the DNA backbone. Thus, DtxR only binds to DNA when metal ion is bound to the repressor.

Effector Binding• Binding of effector molecules can be covalent or c
an lead to covalent changes in a protein.
Examples :•Phosphorylation on the hydroxyl group of the side
chains of serine, threonine or tyrosine residues•side-chain methylation, •covalent attachment of carbohydrates and lipids, •amino-terminal acetylation and •limited proteolytic cleavage, in which proteases c
ut the polypeptide chain in one or more places.

Protein activity may be regulated by protein quantity and lifetime
• The activity of a protein can also be regulated by controlling its amount and lifetime in the cell.
• The amount of protein can be set by the level of transcription
• At the level of the protein, quantities are controlled by the lifetime of the molecule, which is determined by its rate of degradation.
• there are several specific mechanisms for targeting protein molecules to degradative machinery in the cell, including covalent attachment of the small protein ubiquitin.

Nobel Prize in Chemistry 2004
• Aaron Ciechanover
Technion – Israel Institute of Technology
• Avram Hershko Technion – Israel Institute of Technology
• Irwin RoseUniversity of California Irvine
"for the discovery of ubiquitin-mediated protein degradation"
http://nobelprize.org/chemistry/laureates/2004/
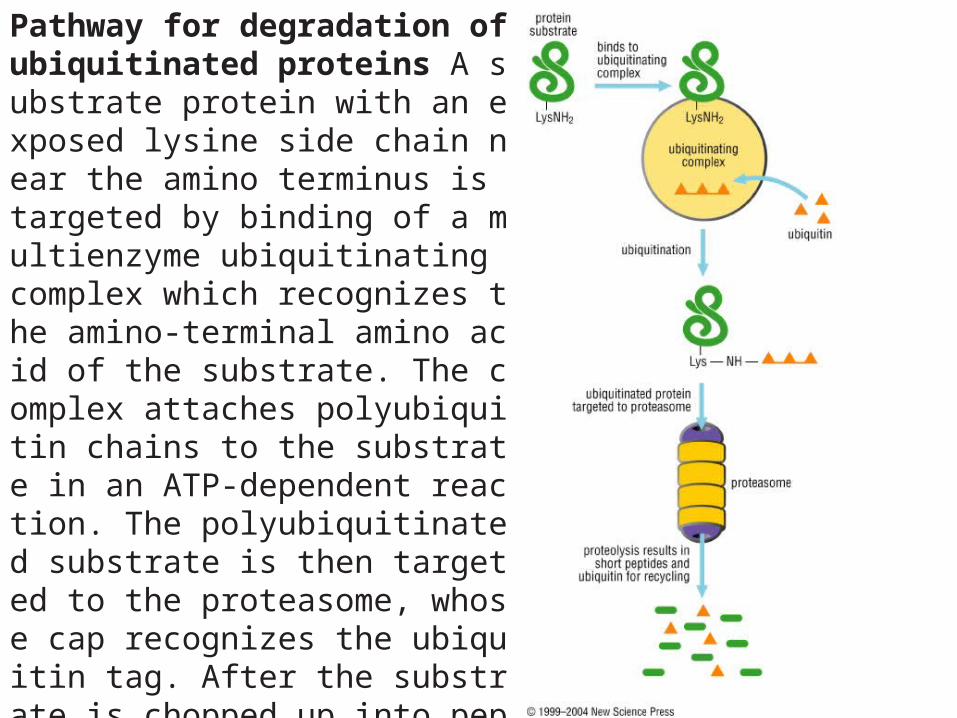
Pathway for degradation of ubiquitinated proteins A substrate protein with an exposed lysine side chain near the amino terminus is targeted by binding of a multienzyme ubiquitinating complex which recognizes the amino-terminal amino acid of the substrate. The complex attaches polyubiquitin chains to the substrate in an ATP-dependent reaction. The polyubiquitinated substrate is then targeted to the proteasome, whose cap recognizes the ubiquitin tag. After the substrate is chopped up into peptide fragments (which may then be degraded further by other proteases), the ubiquitin is recycled.

The eukaryotic proteasome Proteins targeted for destruction (green) are fed into the multiprotein complex called the proteasome. In prokaryotes, these machines of destruction consist simply of a tunnel-like enzymatic core; in eukaryotes they have an additional cap (here shown in purple) at either or both ends. The core is formed by four stacked rings surrounding a central channel that acts as a degradation chamber. The caps recognize and bind to proteins targeted by the cell for destruction. On entry into the proteasome, proteins are unfolded in a process that uses the energy released by ATP hydrolysis and injected into the central core, where they are enzymatically degraded into small fragments. http://doegenomestolife.org.

Ubiquitin and protein degradation

Ubiquitin Structure

Di-ubiquitin
http://www.ebi.ac.uk/thornton-srv/databases/cgi-bin/pdbsum/GetPage.pl?pdbcode=1aar

complex of the vps23 uev with ubiquitin
http://www.ebi.ac.uk/thornton-srv/databases/cgi-bin/pdbsum/GetPage.pl?pdbcode=1uzx

A single protein may be subject to many regulatory influences
Coordination and integration of regulatory signals is achieved largely through signal transduction networks that set the balance of activities and thereby the balance of metabolism and cell growth and division pathways.
The cyclin-dependent protein kinases that control progression through the cell cycle are regulated by a number of different mechanisms