ultrastructural changes in rat livers induced by repeated … · liver changes induced by trypan...
TRANSCRIPT

[CANCER RESEARCH 32, 2393-2399, November 1972]
Ultrastructural Changes in Rat Livers Induced by RepeatedInjections of Trypan Blue
T. Gillman and R. C. Hallowes
Institute of Animal Physiology, Agricultural Research Council, Brabraham, Cambridge, England ¡T.G.J, and Department of Pathology, ImperialCancer Research Fund, Lincoln 's Inn Fields, London, WC2A 3PX, Engfand /R. C. H.]
SUMMARY
Repeated injections of trypan blue into the s.c. tissue ofinbred Wistar rats induced tumors in the liver. A sequence ofmorphological changes has been observed during induction ofthe tumors, and these were studied ultrastructurally. Theearliest recognized change was the appearance in the portaltracts of focal collections of monocytic cells that resembledKupffer cells. They appeared either to collect in the periportalconnective tissue or to migrate through the endothelial walland collect within the portal venules or lymphatics. Thiscaused periportal cyst formation and pressure atrophy of thesurrounding hepatocytes. No mitotic activity was seen in themonocytic cells, but their numbers increased and occludedlymph flow. The more numerous these cells became the lessdid they resemble Kupffer cells and the more they came toresemble cells of the established tumors. Collagen fibrils andbundles as well as fibrin appeared among the cells.Subsequently, nonperiodic fibrillar material was also seenaround the cells. The cellular areas were then indistinguishablefrom established primary tumors. The morphological changesseen in the liver resemble those reported in histopathologicaldescriptions of Kupffer cell sarcoma in humans.
INTRODUCTION
In 1949 Gillman et al. (4) described the induction of tumorsin the livers of rats by repeated injections of trypan blue. Thetumors were classified as reticulum cell tumors. They weretransplantable (11), and the cells of transplanted s.c. tumorsand metastatic renal tumors were similar morphologically andin enzyme content and were associated with extracellularmaterial similar to those of the primary tumors (7).
A sequence of morphological changes has been shown tooccur in the liver during tumor induction. The early changeswere present after a total of 50 mg of trypan blue had beenadministered by s.c. injection and late changes after 150 mg oftrypan blue had been administered (4, 5). This paper describesthe ultrastructure of these changes and discusses theirrelevance to other reticuloendothelial tumors.
MATERIALS AND METHODS
Twenty-five male Wistar rats were given repeated injectionsof trypan blue, and 12 of these rats were used in this study.
Received March 3, 1972; accepted July 28, 1972.
The rats were from an inbred strain that had been maintainedat the Brabraham Laboratories of the Agricultural ResearchCouncil for 6 years and had originally been purchased fromthe Medical Research Council Animal Laboratories,Carshalton, Surrey, England. The trypan blue was from thesame batch of Grubler's dye as used in previous studies (4-6,
8). Each rat received s.c. injections of 10 mg of dye freshlydissolved in 1 ml of glass-distilled water. The injections weregiven once every other week and commenced when the ratswere 9 to 10 weeks old and weighed approximately 220 g.Eight rats, weighing approximately 330 g each at the time ofbiopsy, were subjected to liver biopsies when they hadreceived a total of 50 mg of trypan blue over 10 weeks. Fourrats, weighing approximately 360 g each at the time of biopsy,were subjected to liver biopsies when they had received a totalof 150 mg of trypan blue over 30 weeks. At biopsy, pieces ofliver tissue were removed from each of these rats underhalothane anesthesia. Liver tissue was also removed from two19-week-old and 40-week-old untreated rats of the same strain.One-half of the tissue was placed in 3% glutaraldehyde in 0.1M phosphate buffer at pH 7.3 for 5 min and reduced inthickness to a maximum of 1.5 mm; the fixation continued at4°for a total of 3 hr. The tissues were transferred to the
phosphate buffer and washed with continuous agitationovernight. They then were embedded in agar, chopped into50-jum slices, and postfixed in 1% buffered osmic acid for 1 hr.The slices were dehydrated through graded alcohols, infiltratedwith Araldite, and embedded (9). After polymerization wascomplete, the slices were examined by phase-contrastmicroscopy and portal tracts of pathologically altered livertissue were marked for examination. Sections 0.5 to 1 urnwere cut from these areas with an LKB Ultratome and stainedwith 0.1% toluidine blue in 0.1% sodium tetraborate at 60°for
2 min; suitable areas then were chosen for ultrathin sectioning.These sections were stained with a solution of 20% uranylacetate (analytical grade) in absolute methanol for 10 min,washed in deionized water, restained in Reynolds lead citrate(19) for 5 min, and examined with an Hitachi 7S electronmicroscope. The other one-half of the tissue was fixed in 4%buffered formaldehyde, dehydrated, and embedded in paraffinwax. Sections from these blocks were stained withhematoxylin and eosin to ensure that representative lesionswere being studied.
RESULTS
The macroscopic and microscopic changes that occur duringthe induction period have been described (4, 5). These changes
NOVEMBER 1972 2393
on March 22, 2020. © 1972 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

T. Gillman and R. C. Hallowes
were not uniformly distributed throughout the liver. Theearliest changes could be seen in one area of a liver thatshowed more advanced changes in adjacent areas. Thissuggested that a progressive lesion had been induced by trypanblue, and the following morphological descriptions are basedon this conclusion.
Apart from some congestion, the external surface of theliver appeared macroscopically normal even after 5 injectionsof 10 mg of dye had been given. On section, however, blueareas 0.5 to 1 mm in diameter were seen on the cut surface.Microscopically, the earliest detectable lesion in the liversections consisted of collections of a few monocytic cells in 1corner of several portal tracts (Fig. 1). As the lesionsprogressed, the monocytic cells became more numerous andlay in cystic spaces round the portal tracts (Fig. 2). The cystsbecame larger and sometimes more cellular and replaced partsof the surrounding hepatic lobules (Fig. 3). An eosinophilicmaterial appeared within the cysts which then became morecellular. Elongated cells with large, pale nuclei appeared amongthe monocytic cells, filled up the cysts, and extended into thehepatic lobules. Mitotic activity was not seen in any of thesecellular areas until they were densely cellular and severalhepatic lobules had been destroyed. Liver biopsies from ratsthat had received 15 injections of 10 mg of dye showed thatthe liver was partly replaced by a variable mixture of small andlarge cysts and tumors. These occurred throughout the liverand most appeared to have originated in a portal tract ratherthan in a hepatic lobule. The tumors consisted of pleomorphiccells loosely arranged in fibrillar connective tissue (Fig. 4).Numerous small cysts and tumors were present alsoimmediately under the serosal surface.
Electron Microscopy
Portal Tract of Untreated Rats. The ultrastructure andarrangement of structures in the portal tracts from the 2untreated rats were similar to those described by Rouiller andJezequel (20). In the periportal connective tissue 2 types ofcells were recognized. One type (Fig. 5, A) had a denselyheterochromatic irregular nucleus and a small amount ofcytoplasm that contained a few mitochondria and somedilated endoplasmic reticulum. These were similar to"adventitious connective tissue cells" described by Schnack et
al. (23). The other cell type (Fig. 5, R) was larger, with an ovalor elongated, less heterochromatic nucleus and with abundantcytoplasm. The perinuclear cytoplasm contained a fewmitochondria, a little rough endoplasmic reticulum, andnumerous ribosomes. The peripheral cytoplasm containedmany filaments but few organdÃes and extended into short orlong pseudopodial processes between collagen bundles andother portal structures. These cells were most numerous roundbile ducts. They were similar to the reticuloendothelial cellsdescribed by Nicolson and Rouiller (16), and Rouiller et al.(21).
Portal Tracts of Treated Rats. The earliest abnormality tobe detected was the presence of a few monocytic cells in aspace lined by endothelial cells in the portal tracts. Thepredominant monocytic cell had an irregular nucleus,approximately 5 ^m in diameter, with abundant marginated
heterochromatin and very rarely a nucleolus. The cytoplasmwas extensive and contained numerous vacuoles withelectron-dense contents, free ribosomes, rough and smoothendoplasmic reticulum, and a few mitochondria. Thecytoplasm that extended into long or short pseudopodial orfilamentous processes protruded in many directions from thecell surface and entwined with similar processes of adjoiningcells. Occasional lymphocytes were present also; but red cells,polymorphonuclear leukocytes, and platelets were not seen.The spaces around the cells were free of electron-densematerial and appeared empty (Fig. 6). The space was limitedby flattened cell processes of endothelial cells devoid ofbasement membrane and by the monocytic cells or theirprocesses which were in direct contact with underlyingcollagen bundles. Similar monocytic cells were present in thesurrounding connective tissue. The other structures in theportal tracts were normal. The hepatocytes and the sinusoidalendothelial cells adjacent to the portal tracts were normal.Kupffer cells formed part of the sinusoid wall. The cells hadsimilar nuclei and their cytoplasm contained similar organdÃesto those in the monocytic cells. The Kupffer cells, however,did not have the numerous pseudopodial processes of themonocytic cells.
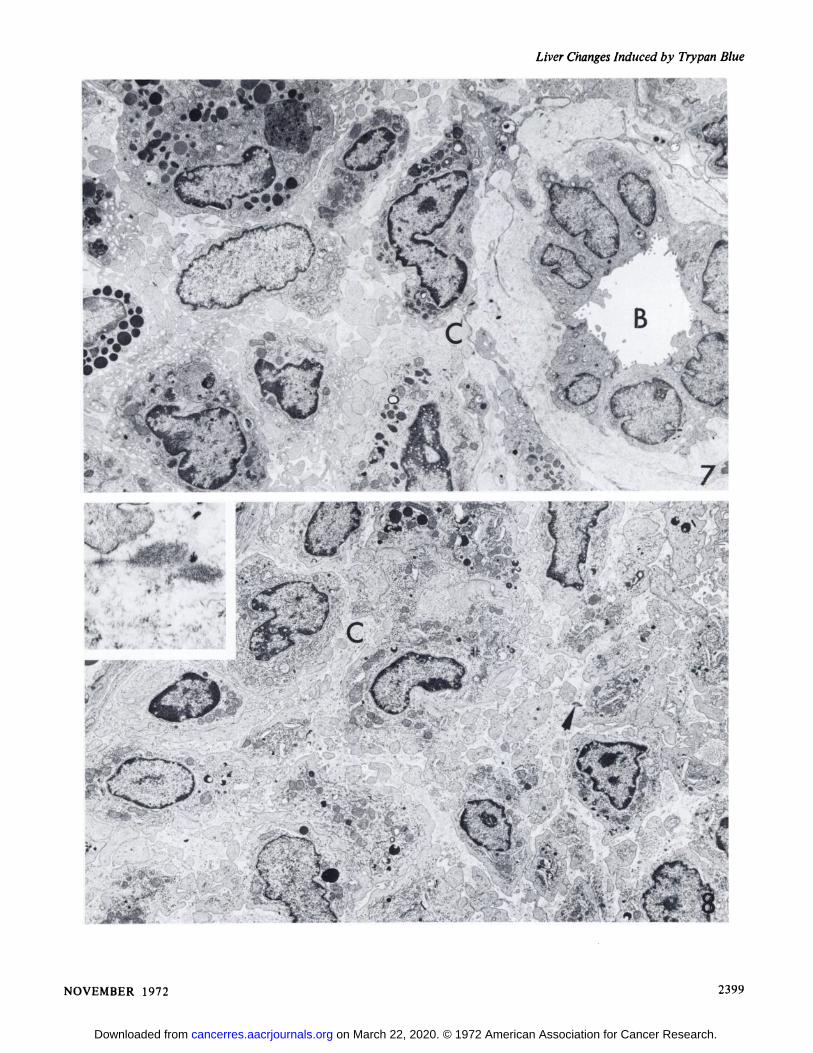
The monocytic cells became more numerous in theperiportal connective tissue and migrated into portal venulesbut not into the spaces of Disse or along the sinusoids. Thecells became more pleomorphic; some cells containednumerous phagosomes, whereas others contained none. Thespaces surrounding them contained finely granular materialand fibrin. Collagen fibers and their bundles appeared in theextracellular spaces (Fig. 7). Hepatocytes adjacent to theseareas underwent both atropic and hypertropic changes butfinally became necrotic. The pleomorphic monocytic cellsextended along the sinusoids as the hepatocytes disappeared.From this stage onwards one cell type became predominant.The cells had elongated, irregular nuclei with some marginatedheterochromatin and nucleoli. The cytoplasm was extensiveand contained numerous mitochondria, scattered profiles ofsmooth and rough endoplasmic reticulum, occasional vacuoles,and few phagosomes. Pseudopodial processes extended fromthe cell surfaces into the amorphous surrounding groundsubstance (Fig. 8). The cells, some of which were binucleate,still varied considerably in size and shape. They were eitherpacked closely together or arranged loosely in a granularground substance in which electron-dense, nonperiodicfibrillary material was seen (Fig. 8, inset). As the tumorsenlarged, the tumor cell nuclei became larger and thecytoplasm contained fewer organelles.
DISCUSSION
The lesions produced in the liver of inbred Wistar rats bytrypan blue are usually present before histologicalabnormalities are seen in lymph nodes, bone marrow, orperipheral blood (4-6); but local infiltration by small numbersof mononuclear cells would be easier to detect in portal tractsof the liver, which are almost devoid of such cells, than inlymph nodes or bone marrow, which already contain largenumbers of mononuclear cells. The 1st change detected in the
2394 CANCER RESEARCH VOL. 32
on March 22, 2020. © 1972 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

Liver Changes Induced by Trypan Blue
tumor process was the focal collection of cells that resembledKupffer cells (15). The subsequent tumor was derived fromthese cells and so could be classified as a Kupffer-cell sarcoma(3). This tumor has been described in Moselle wine growers(22), patients receiving Fowler's solution (18) (both due to
long exposure to arsenic), patients exposed to Thorotrast (13,17), and patients exposed to urethan (24). In all of theseconditions ingestion of foreign material by Kupffer cells couldoccur. A standard procedure for demonstrating these cells insitu is by their intravital uptake of trypan blue. Thehistopathology of the liver in Kupffer-cell sarcoma and theinvolvement of portal lymph node, spleen, and lung has beenreviewed recently by Blackwell and Joske (1). There was noevidence of leukemia in any of these cases, but the histológica!appearance of the liver tumors resembled those seen in thetrypan blue-induced tumors in the livers of rats.
The origin of Kupffer cells is not known. Evidence suggeststhat they arise from either thoracic duct cells (10) or fromprecursors in the bone marrow (12). If trypan blue initiatedneoplastic transformation of these precursors, it would bedifficult to detect in situ because of the problem of locatingsuch cells in diffuse cellular tissues. Early in the inductionperiod abnormal Kupffer cells collected in the portal areas,while surrounding adjacent hepatocytes appearedmorphologically normal. The abnormal cells were not,therefore, part of the response of the rat to hepatocellulardamage, but their presence led to local alteration and possiblyblocking of blood flow in terminal portal tracts. This in turnled to hepatocellular damage. Some abnormal cells containednumerous phagosomes in their cytoplasm, others contained afew, and some cells were without phagosomes. The longer therats were exposed to repeated doses of trypan blue the lesswell differentiated did the abnormal cells become. No mitoticactivity was seen in the abnormal cells in the portal tracts (4).Trypan blue appeared to stimulate Kupffer cell precursors,either directly or indirectly, to produce cell lines withdifferent cytoplasmic capabilities. Cells of one line that hadretained a phagocytic capability may have been stimulated tobecome hypertropic and actively phagocytic when surroundedby necrotic and fragmenting hepatocytes. This proliferativestimulus would be lost once the cellular debris had beenremoved. Then the proliferation may have been stimulated ofanother cell line that had not retained a phagocytic capabilityand that preferred to grow in semisolid rather than fluidconditions that followed the formation of fibrin and collagenin the cysts. The appearance of fibrin and collagen in sinusoidshas been reported in toxic conditions (2) and appears to be afeature of hepatocellular disorders that affect the endotheliumof the liver sinusoids (23). Reticuloendothelial cells in theportal tract connective tissue the morphology of which wasintermediate between that of endothelial cells and Kupffercells have been reported (21). Such cells become hyperplasticafter injections of zymosan, a yeast hydrolysate, and resemblemorphologically the abnormal monocytic cells seen in trypanblue-treated rats (4).
In previous papers, Gillman et al. (5, 6) have presentedevidence suggesting that the neoplastic cells of tumors inducedin rats by trypan blue are abnormal histiocytes that arise fromendothelium. The present paper does not demonstrate anendothelial origin for the tumor, but it does show
morphological similarities between this tumor and thoseinduced by arsenic or Thorotrast. Trypan blue is not a purechemical compound (14). The isolation of a carcinogenicfraction or fractions from this dye might simplify furtherstudies on a dye that is both a teratogen and an inducer ofproliferative changes in reticuloendothelial tissues of rats.
ACKNOWLEDGMENTS
Dr. Hallowes gratefully acknowledges the help of members of thelate Dr. Gillman's department in the preparation of this manuscript. Dr.
Hallowes would also like to thank Mr. D. G. West for technicalassistance, Mrs. S. Whittles for typing the manuscript, and Mr. J. Leachfor photographic assistance.
REFERENCES
1. Blackwell, J. B., and Joske, R. A. Kupffer-cell Sarcoma. Am. J.Digest. Diseases, /5: 133-138, 1970.
2. Boler, R. R., and Bibighaus, A. J. Ultrastructural Alterations ofDog Livers during Endotoxin Shock, Lab. Invest., 17: 537-561,
1967.3. Edmundson, H. A. Tumours of the Liver and Intrahepatic Bile
Ducts. Atlas of Tumour Pathology, Section 7, Fascicle 25, p. 139.Washington: Air Force Institute of Pathology.
4. Gillman, J., Gillman, T., and Gilbert, C. Reticulosis andReticulum-cell Tumours of the Liver Produced in Rats by TrypanBlue with Reference to Hepatic Necrosis and Fibrosis. SouthAfrican J. Med. Sci., 14: 21-84, 1949.
5. Gillman, J., Gillman, T., and Gilbert, C. The Pathogenesis ofExperimentally Produced Lymphomata in Rats (IncludingHodgkin's-like Sarcoma). Cancer, 5. 792-846, 1952.
6. Gillman, T. II. Abnormal Monocyte and PolymorphonuclearLeucocyte Formation in Trypan Blue Treated Rats. PossibleSignificance for Understanding the Pathogenesis of Monocytic andRelated Leukaemias in Man. Medicine in South Africa, 50-67,
1957.7. Gillman, T., and Hallowes, R. C. Ultrastructural and Histochemical
Observations on a Transplantable Reticuloendothelial Tumor inRats. Cancer Res., 32: 2383-2392, 1972.
8. Gillman, T., Kinns, M., and Cross, R. M. Hodgkin's Disease: A
Possible Experimental Model in Rats. Lancet, 2: 1421-1422,
1969.9. Hallowes, R. C., and Streck, P. W. The Use of the Tissue Sectioner
in Selecting Histological Features for Electron MicroscopicalStudy. J. Microscop., 92: 217-222, 1970.
10. Howard, J. G., Boad, J. L., and Christie, G. H. Further Studies onthe Transformation of Thoracic Duct Cells into Liver Macrophages.Ann. N. Y. Acad. Sci., 129: 327-339, 1967.
11. Kinns, A. M. Growth in the Rats Brown Fat of a Hodgkin's-like
Transplantable Reticulo-Endothelial Tumour. Brit. J. Exptl.Pathol.,5/: 411-414, 1970.
12. Kinsky, R. G. Extra-hapatic Derivation of Kupffer Cells duringOestrogenic Stimulation of Parabiotic Mice. Brit. J. Exptl. Pathol.,50: 438-447, 1969.
13. Kluge, T., and Hovig, T. Ultrastructural Localization of Thorotrastin the Reticuloendothelial System. Am. J. Pathol., 54: 355-372,1969.
14. Lloyd, J. B., Beck, F., Griffiths, A., and Parry, L. M. TheMechanism of Action of Acid Bisazo Dyes. In: P. N. Campbell(ed.), The Interaction of Drugs and Subcellular Components onAnimal Cells, pp. 171-202. London: J. & A. Churchill Ltd.
15. Mills, D. M., and Zucker-Franklin, D. Electron Microscope Studyof Isolated Kupffer Cells. Am. J. Pathol., 54: 147-166, 1969.
NOVEMBER 1972 2395
on March 22, 2020. © 1972 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

T. Gillman and R. C. Hallowes
16. Nicolson, P., and Rouiller, C. Beziehungen Zwischen denEndothelzellen der Lebersinusoide und den von KupfferschenSternzellen. Z. Zellforsch. Mikroscop. Anat. Abt. Histochemie, 76:313-338, 1967.
17. Rakov, H. L., Smalldon, T. R., and Dermon, H. HepaticHemangioendothelio-Sarcoma. Arch. Intern. Med., 112: 173-178,1963.
18. Regelson, W., Kim, U., Ospina, J., and Holland, J. F.Hemangioendothelial Sarcoma of Liver from Chronic ArsenicIntoxication by Fowler's Solution. Cancer, 21: 514-522, 1968.
19. Reynolds, E. S. Use of Lead Citrate at High pH as an ElectronOpaque Stain in Electron Microscopy. J. Cell Biol., 17: 208-212,1963.
20. Rouiller, C., and Jezequel, A. M. Electron Microscopy of the Liver.
In: C. Rouiller (ed.), The Liver, Vol. 1, pp. 195-264. NewYork:Academic Press, Inc., 1963.
21. Rouiller, C., Pictet, R., Nicoleson, P., and Orci, L. Les CellulesReticulo-Endotheliales du Foie et l'Espace de Disse. Rev. Intern.
Hepatol., l 7: 827-854, 1967.22. Roth, F. Ãœberdie chronische Arsenvergiftung der Moselwinzer
unter besonderer Berücksichtigung des Arsenkrebes. Z.Krebsforsch., 61: 287-319, 1956.
23. Schnack, H., Stockinger, L., and Wewalka, F. AdventitiousConnective Tissue Cells in the Space of Disse and Their Relation toFibre Formation. Rev. Intern. Hepatol., 17: 855-860, 1967.
24. Trainin, N. Neoplastic Nature of Liver "Blood Cyst" Induced by
Urethane in Mice. J. Nati. Cancer Imi.,31: 1489-1499, 1963.
Fig. 1. Part of a portal tract showing a portal vein (v) surrounded by numbers of monocytic cells (m) situated in periportal connective tissue (50mg trypan blue). Toluidine blue, X 500.
Fig. 2. Cyst in liver containing large monocytic cells (m). Three hepatocytes (//) in various stages of atrophy are present (50 mg trypan blue).Toluidine blue, X 500.
Fig. 3. Three portal veins (v) and a bile duct (d) completely surrounded by mononuclear cells. The remains of at least 1 hepatocyte (h) is present(50 mg trypan blue). Toluidine blue, X 500.
Fig. 4. Pleumorphic cellular and fibrillar tumor replacing liver (150 mg trypan blue). Toluidine blue, X 500.Fig. 5. Normal rat hepatic portal tract. In the portal connective tissue between bile ducts (B), portal vein (K), and hepatocytes (H) are 3
reticuloendothelial cells (R) and an adventitial connective tissue cell (A). X 4,000.Fig. 6. Trypan blue-treated rat (50 mg). The portal connective tissue contains a focal collection of monocytic cells (M) resembling Kupffer cells
within an endothelial-lined (£')space adjacent to a bile duct (B). X 4,000.
Fig. 7. Trypan blue-treated rat (50 mg). Several pleomorphic monocytic cells are present among collagen fibers (Q adjacent to a bile duct(B). X 4,000.
Fig. 8. Trypan blue-treated rat (150 mg). The periportal tissues are replaced by pleomorphic cells. Collagen fibers (O are present in theintercellular spaces as well as a single group of nonperiodic fibrils (arrow) characteristic of trypan blue-induced tumors. X 4,000. Inset, detail ofthese nonperiodic fibrils. X 20,000.
2396 CANCER RESEARCH VOL. 32
on March 22, 2020. © 1972 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

Liver Changes Induced by Trypan Blue
fy'ìf ' '* *4r *' * ^ "
•¿�"V~ C\ CO «1 %'< JiJ: À '
•¿�-^>. li - M^wf
ÃŒ5r- •¿�•¿�''^^^.-..L,,' «»%-^|w*-.'"^y
.. •¿�-v;*^; v ^*'tff^T*"-'Ä«^;w^ ^»«sÃP̂fer ¿^i'̂ ^^?^C?f2
NOVEMBER 1972 2397
on March 22, 2020. © 1972 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

T. Gillman and R. C. Hallowes
6. .v"--
2398 CANCER RESEARCH VOL. 32
on March 22, 2020. © 1972 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from
Liver Changes Induced by Trypan Blue
¿;;
NOVEMBER 1972 2399
on March 22, 2020. © 1972 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

1972;32:2393-2399. Cancer Res T. Gillman and R. C. Hallowes Injections of Trypan BlueUltrastructural Changes in Rat Livers Induced by Repeated
Updated version
http://cancerres.aacrjournals.org/content/32/11/2393
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/32/11/2393To request permission to re-use all or part of this article, use this link
on March 22, 2020. © 1972 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from