ufrrj instituto de florestas programa de pÓs...
TRANSCRIPT

UFRRJ
INSTITUTO DE FLORESTAS
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS
AMBIENTAIS E FLORESTAIS
TESE
Caracterização estrutural do complexo lignina-carboidrato-ácidos fenólicos da
parede celular do caule de Euterpe oleracea Mart. (Arecaceae)
Gisely de Lima Oliveira
2017

ii
UNIVERSIDADE FEDERAL RURAL DO RIO DE JANEIRO
INSTITUTO DE FLORESTAS
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS AMBIENTAIS E
FLORESTAIS
CARACTERIZAÇÃO ESTRUTURAL DO COMPLEXO
LIGNINA-CARBOIDRATO-ÁCIDOS FENÓLICOS DA
PAREDE CELULAR DO CAULE DE Euterpe oleracea Mart.
(Arecaceae)
Gisely de Lima Oliveira
Sob a orientação do Professor
Dr. Alexandre Monteiro de Carvalho
Dr. Heber dos Santos Abreu (in memoriam)
e Coorientação da Professora
Dra. Helena Regina Pinto Lima
Tese submetida como requisito parcial para
obtenção do grau de Doutor em Ciências, no
Programa de Pós-Graduação em Ciências
Ambientais e Florestais, Área de
concentração em Tecnologia e Utilização de
Produtos Florestais.
Seropédica, RJ
Maio de 2017

iii
Ficha catalográfica a ser elaborada pela Biblioteca Central

iv
UNIVERSIDADE FEDERAL RURAL DO RIO DE JANEIRO
INSTITUTO DE FLORESTAS
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS AMBIENTAIS E
FLORESTAIS
GISELY DE LIMA OLIVEIRA
Tese submetida como requisito parcial para obtenção do grau de Doutor em Ciências, no
Programa de Pós-Graduação em Ciências Ambientais e Florestais, Área de Concentração em
Tecnologia e Utilização de Produtos Florestais.
TESE APROVADA EM: 26/05/2017
____________________________________________________
Alexandre Monteiro de Carvallho. Prof. Dr. UFRRJ
(Orientador)
____________________________________________________
Fábio Akira Mori. Prof. Dr. UFLA
____________________________________________________
Juarez Benigno Paes. Prof. Dr.UFES
____________________________________________________
Fernando José Borges Gomes. Prof. Dr. UFRRJ
____________________________________________________
Gilmara Pires de Moura Palermo. Prof. Dr. UFRRJ

v
A Deus.
Ao meu marido Leandro Oliveira de Brito.
A minha mãe Maristela de Lima Oliveira.
Aos meus irmãos Vitor e Wagner de Lima Oliveira.
A minha avó Maria José de Cavalcante de Lima.
A todos os meus familiares.
DEDICO

vi
AGRADECIMENTOS
Nesta etapa tão importante da minha vida, não poderia deixar de agradecer primeiramente a
Deus por mais uma vez mostrar sua fidelidade em minha vida me ajudando a vencer com
sabedoria todos os obstáculos. Sinto-me verdadeiramente abençoada por Deus ter-me
concedido a oportunidade de concluir o Doutorado em Ciências Ambientais e Florestais na
Universidade Federal Rural do Rio de Janeiro.
A Universidade Federal Rural do Rio de Janeiro, ao Programa de Pós-graduação em Ciências
Ambientais e Florestais (PPGCAF), Instituto de Florestas pela oportunidade que me
concederam em participar no doutorado bem como a disponibilidade das instalações.
A Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) pela concessão
da bolsa durante todo o período de realização deste doutorado.
A Fundação de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ) pela infraestrutura
e aquisição de equipamentos do Laboratório de Química da Madeira do Departamento de
Produtos Florestais do Instituto de Florestas/ Universidade Federal Rural do Rio de Janeiro.
Ao Prof. Dr. Heber dos Santos Abreu, que como orientador, colaborou em todos os aspectos
para a realização deste trabalho, demonstrando notável atenção perante minhas dúvidas
(mesmo diante de tantas obrigações e responsabilidade), paciência, interesse e amizade na
tarefa de orientação. Infelizmente, não foi possível que o senhor visse a conclusão deste
trabalho. É uma lástima que não esteja mais em nosso convívio, mas gostaria de deixar
registrado aqui meu agradecimento póstumo. Somos assim, despreparados para perda e
impacientes com o tempo, o qual deixamos responsável por apagar o nosso sofrimento. Eu
estou triste pela falta que o senhor me faz, mas ao mesmo tempo, grata a Deus por ter
conhecido o senhor um dia. Tê-lo como orientador (mais uma vez) foi um privilégio. Muito
obrigada por tudo!
Ao professor Alexandre Monteiro de Carvalho pela orientação, apoio e que tanta ajuda
forneceu para que este trabalho fosse concluído com o êxito esperado.
À professora Helena Regina Pinto Lima, do Departamento de Botânica, do Instituto de
Ciências Biológicas e de Saúde pela orientação, sobretudo pelo apoio, companheirismo e
amizade durante a realização deste trabalho. Obrigada por sua atenção e paciência em mostrar
quais os caminhos que deveria seguir, na hora em que ficava sem rumo e incentivando na
conclusão dessa tese.
Aos professores Mário Geraldo de Carvalho e Rosane Nora Castro pelo apoio, incentivo,
ensinamentos, colaboração e parceria, por todos os momentos em que necessitei da ajuda de
vocês. Mais uma vez minha sincera gratidão.
Ao Prof. Roberto Carlos Costa Lelis pela concessão do Laboratório de Tecnologia da Madeira
do Instituto de Florestas/ Universidade Federal Rural do Rio de Janeiro.

vii
Aos técnicos Vitor dos Santos Almeida e Maurício Lemos Mattos do Laboratório de Química
Orgânica do Departamento de Química do Instituto de Ciências Exatas da Universidade
Federal Rural do Rio de Janeiro pelos registros dos espectros de RMN de 13
C.
Ao amigo José Carlos Ferreira Batista pelo carinho e pela valiosa colaboração que foi de
fundamental importância para o processo do desenvolvimento deste trabalho.
Ao meu marido Leandro, meu amigo e companheiro de todas as horas, meu sincero
agradecimento pelo apoio incondicional nos momentos difíceis, pela disponibilidade e
paciência ao longo deste período. Sou grata por cada gesto carinhoso e amoroso, sorriso e
lágrimas em que você esteve ao meu lado. Obrigada meu AMOR.
À minha amada avó Maria José Cavalcante de Lima, por quem serei eternamente grata, pelo
carinho, dedicação e ensinamentos; aos meus estimados tios pelo carinho, amizade e
colaboração que me apoiaram ao longo da minha jornada universitária, mas também ao longo
de toda minha vida.
À minha mãe Maristela de Lima Oliveira, aos meus queridos irmãos Vitor e Wagner de Lima
Oliveira, a minha cunhada Cíntia Martins de Lima pelo carinho, amor e dedicação e também
pela compreensão pelos momentos em que estive ausente.
Aos meus amados sobrinhos Laís e Davi Martins de Lima pelas brincadeiras, pelas risadas e
principalmente pelo apoio e por entender que muitas vezes foi necessário estar ausente. Vocês
sempre poderão contar com a tia!
A todos os colegas e amigos do Laboratório de Química da Madeira do Instituto de
Florestas/Universidade Federal Rural do Rio de Janeiro.
A todos que direta ou indiretamente contribuíram para a realização desse sonho. Obrigada a
todos pelo carinho, amizade, companheirismo e principalmente pela paciência ao longo desta
jornada que ao mesmo tempo foi extremamente difícil e gratificante.

viii
RESUMO
OLIVEIRA, Gisely de Lima. Caracterização estrutural do complexo lignina-carboidrato-
ácidos fenólicos da parede celular no caule de Euterpe oleracea Mart. (Arecaceae). 2017.
84 f. Tese (Doutorado em Ciências Ambientais e Florestais). Instituto de Florestas,
Universidade Federal Rural do Rio de Janeiro, Seropédica, RJ, 2017.
Euterpe oleracea Mart. caracteriza-se como uma espécie importante pelo seu potencial
econômico e pela utilização, principalmente, de seus frutos. Esse trabalho teve como objetivo
caracterizar a lignina e ácidos fenólicos constituintes, bem como as ligações entre esses nas
paredes celulares do caule de Euterpe oleracea Mart. As análises foram realizadas com a base
do caule de palmeiras amostradas nas regiões externa (RE) e interna (RI) da secção tranversal,
sendo estas extraídas com solventes orgânicos e tratadas com NaOH 1%. Foram quantificados
os teores de holocelulose, α-celulose e lignina. Também foram realizadas análises
espectroscópicas (Espectroscopia Infravermelho com Transformada de Fourier e de
Ressonância Magnética Nuclear de 13
C no estado sólido) e cromatográficas com as amostras
do caule, com e sem tratamento com NaOH 1%. As ligninas das amostras RE e RI foram
isoladas pelo método de Björkman para a caracterização estrutural desta, pela da técnica de
RMN 1H e
13C no estado líquido. Para o estudo anatômico e avaliação do processo de
lignificação, as amostras RE e RI foram processadas de acordo com técnicas usuais de
microscopia óptica e microscopia de fluorescência. O caule extraído apresentou um teor de
74,40% (RE) e 74,42% (RI) de holocelulose e um teor 10,33% (RE) e 15,66% (RI) de lignina.
Após o tratamento com NaOH 1%, houve um aumento do teor de holocelulose de 6,08% e
4,73% (RE e RI, respectivamente) e uma redução do teor de lignina, resultando na dissolução
de 1,83% e 4,33% da lignina original das amostras RE e RI. Os resultados da espectroscopia
no infravermelho (FTIR) e RMN 13
C no estado sólido permitiram observar, que após o
tratamento com NaOH 1%, ocorreu uma redução de grupos funcionais ligados a lignina e as
hemiceluloses, indicando que o processo se mostrou favorável à hidrólise. Verificou-se que a
lignina das amostras RE e RI de E. oleracea é do tipo Guaiacila Siringila (GS) e incorporada
com ácido p-hidroxibenzoico e protocatecuico, os quais foram confirmados pela
cromatografia líquida de alta performance. Também foram observadas as principais unidades
estruturais presentes na lignina como ligação β-O-4, β-O-4’ ϒ-acetiladas, spiro-dienona,
unidade guaiacila, unidade siringila e p-hidroxibenzoato. As análises das dimensões das fibras
apresentaram valores médios de comprimento de 2570,82 e 2460,04 µm, diâmetro de 60,67 e
62,89 µm, diâmetro do lume de 43,28 e 35,93 µm, espessura da parede celular de 10,39 e
13,48 µm, respectivamente para as amostras RE e RI. Os testes histoquímicos e a microscopia
de fluorescência foram considerados eficientes e mostraram a localização e a composição da
lignina, revelando que as ligninas são do tipo GS e as unidades de guaiacila e surge
principalmente na lamela média e nos ângulos comuns as celulas.
Palavras chave: açaí, derivados do ácido benzóico, análises espectroscópicas.

ix
ABSTRACT
OLIVEIRA, Gisely de Lima. Structural characterization of the lignin-carbohydrate-
phenolic acids complex of the stem cell wall of Euterpe oleracea Mart. (Arecaceae). 2017.
84 f. Thesis (Doctor Science in Environmental and Forestry Sciences). Instituto de Florestas,
Universidade Federal Rural do Rio de Janeiro, Seropédica, RJ, 2017.
Euterpe oleracea Mart. is characterized as an important species due to its economic potential
and the use, mainly, of its fruits. This work aimed to characterize the lignin and phenolic acids
constituents, as well as the connections between them in the cell walls of the stem of Euterpe
oleracea Mart. The analyzes were carried out with the base of the palm stem sampled in two
regions: external region (ER) and internal region (IR) of the cross section, which were
extracted with organic solvents and treated with 1% NaOH. The levels of holocellulose, α-
cellulose and lignin were quantified. We also performed spectroscopic (FTIR and 13
C NMR in
solid state) and chromatographic analyzes with stem samples, with and without 1% NaOH
treatment. The lignins from ER and IR samples were isolated by Björkman method for the
structural characterization of this by the 1H and
13C NMR in the liquid state. For anatomical
study and evaluation of the lignification process, ER and IR samples were processed
according to usual techniques under light microscopy and fluorescence microscopy. The
extracted stem had a content of 74.40% (ER) and 74.42% (IR) of holocellulose and a content
of 10.33% (ER) and 15.66% (IR) of lignin. After treatment with 1% NaOH, there was an
increase in the holocellulose content of 6.08% and 4.73% (ER and IR, respectively) and a
reduction of the lignin content of these, resulting in the dissolution of 1.83% and 4.33% of the
original lignin from samples ER and IR. The results of infrared spectroscopy (FT-IR) and 13
C
NMR in the solid state allowed to observe, after treatment with 1% NaOH, a reduction of
functional groups bound to lignin and hemicellulose, indicating which process was favorable
to the hydrolysis. The lignin of the E. oleracea ER and IR samples was found to be GS-type
and incorporated with benzoic derivatives, p-hydroxybenzoic and protocatecuic acid, in which
they were confirmed by high performance liquid chromatography. The main structural units
present in lignin as β-O-4, β-O-4'-acetylated, spiro-dienone, guaiacyl, syringyl and p-
hydroxybenzoate units were also observed. The analysis of the fiber dimensions presented
mean fiber length values of 2570.82 and 2460.04 μm, fiber diameter of 60.67 and 62.89 μm,
flame diameter of 43.28 and 35.93 μm, thickness of the cell wall of 10.39 and 13.48 μm,
respectively for the ER and IR samples. Histochemical tests and fluorescence microscopy
were considered efficient and showed the location and composition of the lignin corroborating
with the chemical analyzes, revealing that the lignins are of the GS type and the units of
guaiacil and arise mainly in the middle lamella and in the cellular corner.
Keywords: açaí, benzoic acid derivatives, spectroscopic analysis.

x
LISTA DE TABELAS
Tabela 1. Teor de celulose, hemicelulose e lignina de diferentes materiais lignocelulósicos e
de alguns resíduos agrícolas (%). ............................................................................................... 6
Tabela 2. Relação de indivíduos, número de registros e parâmentros dendrométricos das
amostras de Euterpe oleracea estudados. ................................................................................. 11
Tabela 3. Teor de holocelulose e lignina das diferentes amostras do caule de Euterpe
oleracea. ................................................................................................................................... 18
Tabela 4. Valores de referência dos constituintes macromoleculares de diversas espécies
(adaptado de FONSECA, 2012). .............................................................................................. 19
Tabela 5. Atribuições dos sinais dos espectros das amostras de Euterpe oleracea (região
externa e interna). ..................................................................................................................... 24
Tabela 6. Sinais FTIR normalizados pela razão da intensidade em 1505 (RE) e 1503 cm-1
(RI) do caule de Euterpe oleracea. ........................................................................................... 25
Tabela 7. Atribuições aos deslocamentos químicos observados nos espectros de RMN 13
C
(estado sólido) das amostras do caule de Euterpe oleracea (RE e RI). ................................... 26
Tabela 8. Atribuição dos sinais de hidrogênio em regiões do espectro de RMN 1H das
ligninas. .................................................................................................................................... 29
Tabela 9. Atribuição dos sinais dos espectros de RMN 2D-HSQC da lignina acetilada de
Euterpe oleracea (RE e RI). ..................................................................................................... 38
Tabela 10. Dimensões das fibras da região externa e interna do caule de Euterpe oleracea. . 45

xi
LISTA DE FIGURAS
Figura 1. Parede celular contendo microfibrilas de celulose, hemicelulose, pectina e proteínas
solúveis (STICKLEN, 2008). ..................................................................................................... 3
Figura 2. Camadas S1, S2 e S3 da parede celular (adaptado de STICKLEN, 2008). ............... 4
Figura 3. Álcoois precursores da lignina p-hidroxifenila (H), guaiacila (G) e siringila (S). ..... 4
Figura 4. Ligações inter-monoméricas comuns em uma molécula de lignina (adapatado de
LATIF et al., 2015). .................................................................................................................... 5
Figura 5. Principais ligações de CLC e subestruturas da lignina (adaptado de YOU et al.,
2015). .......................................................................................................................................... 7
Figura 6. Esquema representativo de uma ligação α éter (a) e β éter (b) formadas entre lignina
e hemicelulose por meio de um hidroxicinamato. (LAM et al., 2001)....................................... 8
Figura 7. Esquema do caule de Euterpe oleracea. (A) Região de coleta das amostras. (B)
Seção transversal do caule de Euterpe oleracea com a identificação das regiões estudadas. .. 12
Figura 8. Fluxograma de preparo das amostras da região externa (RE) e interna (RI) do caule
de Euterpe oleracea. ................................................................................................................. 13
Figura 9. Estrutura dos derivados benzóicos. (A) Ácido p-hidroxibenzóico e (B) ácido
protocatecuico. .......................................................................................................................... 20
Figura 10. Espectros de infravermelho das amostras extraídas, extraídas e tratadas com
NaOH 1%, lignina de Björkman e da lignina acetilada da região externa (RE) do caule de
Euterpe oleracea....................................................................................................................... 21
Figura 11. Espectros de infravermelho das amostras extraídas, extraídas e tratadas com
NaOH 1%, lignina de Björkman e da lignina acetilada da região interna (RI) do caule de
Euterpe oleracea....................................................................................................................... 22
Figura 12. Esquema do complexo lignina-carboidrato (CLC) reagindo com NaOH (HE et al.,
2008). ........................................................................................................................................ 25
Figura 13. Espectros de RMN 13
C (estado sólido) das amostras extraídas e amostras extraídas
e tratadas com NaOH 1% do caule de Euterpe oleracea (região externa e interna). ............... 28
Figura 14. Espectros de RMN 1H das ligninas acetiladas de Euterpe oleracea das regiões
(RE) e (RI). ............................................................................................................................... 30
Figura 15. Espectros de RMN 13
C DEPTQ (400 MHz, CDCl3) da lignina das amostras RE e
RI de Euterpe oleracea. ............................................................................................................ 32

xii
Figura 16. Espectro HSQC (400/100 MHz, CDCl3) da fração da lignina acetilada da região
externa de Euterpe oleracea (RE). ........................................................................................... 34
Figura 17. Espectro HSQC (400/100 MHz, CDCl3) da fração da lignina acetilada da região
externa de Euterpe oleracea (RI). ............................................................................................ 35
Figura 18. Espectro HMBC (400/100 MHz, CDCl3) da fração da lignina acetilada da região
externa de Euterpe oleracea (RE). ........................................................................................... 36
Figura 19. Espectro HMBC (400/100 MHz, CDCl3) da fração da lignina acetilada da região
externa de Euterpe oleracea (RI). ............................................................................................ 37
Figura 20. Principais unidades estruturais presentes na lignina das amostras RE e RI de
Euterpe oleracea....................................................................................................................... 39
Figura 21. Estrutura da lignina da palmeira Euterpe oleracea, obtida pelas informações (A)
do espectro de RMN 1H e (B) RMN
13C. ................................................................................. 39
Figura 22. Curvas de UV dos padrões contidos nas amostras RE e RI de E. oleracea
adquiridas pela análise por CLAE-DAD. ................................................................................. 41
Figura 23. Cromatogramas dos ácidos fenólicos encontrados nos extratos obtidos com acetato
de etila e metanol de Euterpe oleracea (RE e RI). ................................................................... 42
Figura 24. Cromatogramas dos ácidos fenólicos encontrados no caule de Euterpe oleracea
(RE e RI). .................................................................................................................................. 44
Figura 25. Seções transversais da região externa do caule de Euterpe oleracea. ................... 48
Figura 26. Seções transversais da região interna do caule de Euterpe oleracea.. ................... 49

xiii
LISTA DE ABREVIAÇÕES, SIGLAS E SÍMBOLOS
H p-Hidroxifenila
G Guaiacila
S Siringilaa
UDP Uridina Difosfato
β-O-4 Ligação Beta-O-4 formada por unidades fenilpropanóides
α-O-4 Ligação Alfa-O-4 formada por unidades fenilpropanóides
β-5 Ligação Beta-5 formada por unidades fenilpropanóides
5-5 Ligação 5-5 formada por unidades fenilpropanóides
4-O-5 Ligação 4-O-5 formada por unidades fenilpropanóides
β-1 Ligação Beta-1 formada por unidades fenilpropanóides
β-β Ligação Beta-Beta formada por unidades fenilpropanóides
CLC Complexo Lignina-Carboidrato
AF Ácido Ferúlico
APC Ácido p-cumárico
RE Região externa do caule de Euterpe oleracea
RI Região interna do caule de Euterpe oleracea
Km Quilômetro
cm Centímetro
ASTM American Society for Testing and Materials
NaOH Hidróxido de sódio
RMN Ressonância Magnética Nuclear
ppm Partes por milhão
mg/L Miligrama por litro
CLAE Cromatografia Líquida de Alta Eficiência
TAPPI Technical Association of the Pulp and Paper Industry
mL Mililitro
ºC Grau Celsius
mg Miligrama
rpm Rotação por minuto
mbar Milibar
L Litro
KOH Hidróxido de potássio
KBr Brometo de potássio
CP/MAS Cross Polarization Magic Angle Spinning
CDCl3 Clorofórmio deuterado
δ Deslocamento químico
HCl Ácido clorídrico
H2O Água
AcOH Ácido acético
MeOH Metanol
MHz Megahertz
Hz Hertz
HPLC High performance liquid chromatography
µL Microlitro

xiv
nm Nanômetro
IAWA International Association of Wood Anatomists
H2SO4 Ácido sulfúrico
FTIR Infravermelho por transformada de Fourier
IV Infravermelho
HSQC Heteronuclear single quantum correlation
HMQC Heteronuclear multiple quantum correlation
HMBC Heteronuclear multiple bond correlation
TR Tempo de retenção

xv
SUMÁRIO
1 INTRODUÇÃO ..................................................................................................................... 1
2 REVISÃO DE LITERATURA ............................................................................................. 2
2.1 Parede Celular................................................................................................................... 2
2.2 Complexo Lignina-Carboidrato-Ácidos fenólicos ........................................................... 6
2.3 Família Arecaceae ............................................................................................................ 9
2.4 Euterpe oleracea Mart. ................................................................................................... 10
3 MATERIAL E MÉTODOS ................................................................................................ 11
3.1 Coleta do material ........................................................................................................... 11
3.2 Preparo das Amostras ..................................................................................................... 12
3.3 Análises Químicas .......................................................................................................... 13
3.4 Determinação do Teor de Holocelulose ......................................................................... 13
3.5 Determinação do Teor de α-Celulose ............................................................................. 14
3.6 Lignina de Klason ........................................................................................................... 14
3.7 Lignina de Björkman ...................................................................................................... 15
3.8 Acetilação ....................................................................................................................... 15
3.9 Espectroscopia no Infravermelho ................................................................................... 15
3.10 Ressonância Magnética Nuclear (RMN 13
C) no estado sólido do caule de Euterpe
oleracea ................................................................................................................................ 16
3.11 Ressonância Magnética Nuclear (RMN 1H e
13C) em solução da lignina de Euterpe
oleracea ................................................................................................................................ 16
3.12 Determinação de Ácidos Fenólicos da parede celular por Cromatografia Líquida de
Alta Eficiência (CLAE) ........................................................................................................ 16
3.13 Microscopia Óptica de Campo Claro ........................................................................... 17
3.14 Testes Histoquímicos .................................................................................................... 18
3.15 Microscopia de Fluorescência ...................................................................................... 18

xvi
4 RESULTADOS E DISCUSSÃO ........................................................................................ 18
4.1 Determinação do teor dos constituintes macromoleculares ........................................... 18
4.2 Análise por espectroscopia de infravermelho (FTIR) do caule de Euterpe oleracea .... 20
4.3 Análises por espectrometria de Ressonância magnética nuclear de 13
C no estado sólido
do caule de Euterpe oleracea ............................................................................................... 26
4.4 Análises por espectrometria de Ressonância magnética nuclear de 1H
13C em solução da
lignina de Euterpe oleracea .................................................................................................. 29
4.5 Análises cromatográficas de ácidos fenólicos dos extratos e do caule de Eutepe
oleracea ................................................................................................................................ 40
4.6 Anatomia quantitativa das fibras caule de Euterpe oleracea ......................................... 44
4.7 Análises histológicas do caule de Eutepe oleracea ........................................................ 46
5 CONCLUSÕES .................................................................................................................... 50
6 REFERÊNCIAS BIBLIOGRÁFICAS .............................................................................. 51
ANEXO ......................................................................................... Erro! Indicador não definido.

1
1 INTRODUÇÃO
A parede celular é um sistema multimolecular constituído por celulose, hemicelulose,
lignina e pelos ácidos p-hidroxicinâmicos que funcionam como âncora nesse sistema. A
lignina tem sido definida como macromolécula de origem fenilpropanoídica constituída
majoritariamente pelas subunidades de álcoois p-cumarílico, coniferílico e sinapílico que
incrustam a parede celular dos vegetais, que pelo acoplamento combinatório destes, formam
as ligninas do tipo p-hidroxifenila (H), guaiacila (G) e siringila (S).
As quantidades relativas das unidades de monolignóis diferem consideravelmente
entre as plantas. Em geral, a lignina das eudicotiledôneas é composta de unidades G e S e
vestígios de unidades H, enquanto a lignina de gimnospermas é principalmente composta por
unidades G, com quantidades menores de H. Em monocotiledôneas, a lignina contém níveis
semelhantes de ambas as unidades S e G e a quantidade de H relativamente mais elevada
(VANHOLME et al. 2010; CESARINO et al., 2012). Além disso, a lignina de gramíneas
contém quantidades significativas de ácido ferúlico e ácido p-cumárico (VOGEL, 2008;
CESARINO et al., 2012).
Em monocotiledôneas os ácidos p-hidroxicinâmicos são envolvidos no processo de
lignificação. Ésteres ferulatos-polissacarídeos são comuns nesse táxon e estão incorporados à
lignina via processos de acoplamento de radicais que caracterizam a lignificação, produzindo
fortes ligações entrecruzadas entre lignina e os polissacarídeos da parede celular (RALPH et
al., 1995, 1998a). Esse autor em 2010 tem divulgado diferentes ligações em ligninas de
espécies de monocotiledôneas, diferindo de outros grupos de plantas.
A composição química da parede celular de monocotiledôneas, nas últimas décadas,
tem recebido atenção considerável de pesquisadores que têm visado não somente a melhoria
dos processos de extração da lignina, mas também na elucidação da estrutura de ligninas, da
caracterização da reatividade química, das propriedades funcionais e do desenvolvimento de
novas aplicações.
A busca de tecnologia para à utilização de matéria-prima proveniente de fontes
renováveis e preservação do ambiente têm motivado centros de pesquisas a estudar a
utilização de recursos naturais e subprodutos agroindustriais (WAMBUA, IVENS e
VERPOEST, 2003; FROLLINI et al., 2004; KEENER, STUART e BROWN, 2004). O
interesse na utilização de fibras vegetais por diferentes segmentos da indústria é crescente,
assim como a demanda por novas fontes de fibras naturais. Neste contexto, o estipe de
palmeiras pode se tornar importante matéria-prima para diversas finalidades e um fator
importante é que esses são muitas vezes descartados em favor de outros produtos de interesse
(ISMAIL e MAMAT, 2002).
Dentre as palmeiras, a Euterpe oleracea Mart., conhecida como açaí, é considerada
uma matéria-prima importante, pelo aproveitamento de seus frutos e extensa comercialização.
Esta espécie tem um valor social e cultural na Região Norte do Brasil e isso pode ser
observada por sua aplicação na economia local, tais como, alimentação, produção de celulose,
fabricação de casas, ração animal, arborização, medicina caseira e corante natural (QUEIROZ
e MELÉM JUNIOR, 2001; GOUVÊA, 2007).
De acordo com dados estatísticos, o Brasil, em 2013, produziu 202.216 toneladas de
açaí, sendo a região Norte a maior produtora com 186.379 toneladas dessa produção,
respondendo por 94% do que é extraído no país, e o Pará é o estado que mais contribuiu para
a região Norte sendo responsável por 59 % do que é produzido na região (IBGE, 2015).

2
A cultura ao longo dos anos vem despertando interesse dos agroextrativistas,
produtores e pesquisadores, os quais têm procurado praticar diferentes formas de manejo
objetivando preservar a espécie e aumentar a produtividade. No entanto, faltam informações
sobre a composição química, principalmente sobre a lignina de palmeiras. A ênfase neste
componente da parede celular ocorre, pois esta é o fator limitante do valor nutritivo dos
materiais fibrosos como fonte de energia para os ruminantes, é um determinante importante
das propriedades mecânicas dos materiais e também por impedir a separação das células de
fibra que é o objetivo principal de polpação.
O conhecimento acerca da lignificação em palmeiras pode expandir os caminhos
futuros para uma melhor utilização do estipe, além da obtenção de produtos de maior valor
agregado como, por exemplo, biocombustíveis lignocelulósicos. Diante do exposto o presente
trabalho teve como objetivo geral verificar as seguintes hipóteses, a saber: i) como se
caracteriza a estrutura da lignina da parede celular das fibras de Euterpe oleracea? ii) Quais
são os ácidos fenólicos constituintes presentes na parede celular desta espécie? iii) Quais os
tipos de ligações ocorrem entre os monolignóis e os demais componentes da parede celular?
iv) Quais alternativas de utilização do estipe da Euterpe oleracea poderão ser sugeridas a
partir das avaliações realizadas?
O trabalho teve como objetivos complementares caracterizar a lignina e ácidos
fenólicos, constituintes da parede celular, a fim de se estudar as ligações cruzadas entre
lignina e o complexo carboidratos-ácidos fenólicos no caule de Euterpe oleracea Mart.;
avaliar os teores dos constituintes da parede celular em duas condições de pré-tratamento para
analisar possíveis alterações do gradiente de concentração de lignina, entre a região externa
(região periférica) e região interna (região central) do caule; caracterizar a lignina e derivados
cinâmicos e benzoicos das paredes celulares, estabelecer as ligações cruzadas entre estes por
de técnicas espectroscópicas e cromatográficas; analisar as características anatômicas e
histoquímicas por microscopia de campo claro e de fluorescência.
2 REVISÃO DE LITERATURA
2.1 Parede celular
A parede celular é rígida e, portanto, limita o tamanho do protoplasto, evitando a
ruptura da membrana plasmática, quando o protoplasto amplia seu tamanho, em decorrência
da entrada de água e também determina o tamanho e a forma da célula, a textura do tecido e a
forma final de um órgão vegetal (EVERT e ESAU, 2013).
Todas as paredes das células provêm da placa equatorial formada durante citocinese,
para separar as células filhas. Cada protoplasto forma sua parede de fora para dentro, de modo
que a camada mais recente de uma determinada parede está em posição mais interna, próxima
do protoplasto (EVERT e ESAU, 2013). A região de união das paredes primárias de células
adjacentes é chamada lamela média ou substância intercelular (GUILLEMIN et al, 2005;
KNOX, 2008).
As primeiras camadas formadas constituem a parede primária, que são finas e
extensíveis (ALBERTS et al., 2002), na qual ocorre a deposição das microfibrilas por
intussuscepção, ou seja, por arranjo entrelaçado (GLÓRIA e GUERREIRO, 2003). Estas são
implantadas em uma matriz hidratada, e tal estrutura fornece resistência e flexibilidade à
célula (TAIZ e ZEIGER, 2004).

3
As paredes primárias contêm três classes principais de polissacarídeos. A primeira é a
celulose, um polímero não ramificado de subunidades de glicose que é sintetizada e
depositadas na parede por um complexo enzimático (celulose sintase) na membrana
plasmática. As microfibrilas de celulose da parede estão embebidas em uma matriz de
moléculas não celulósicas. Essas moléculas são os polissacarídeos conhecidos como
hemicelulose e pectinas, bem como as proteínas estruturais chamadas glicoproteínas (Figura
1) (SMITH, 2001; EVERT e ESAU, 2013).
Figura 1. Parede celular contendo microfibrilas de celulose, hemicelulose, pectina e proteínas
solúveis (STICKLEN, 2008).
As hemiceluloses são formadas por unidades básicas, chamadas de monossacarídeos
(arabinose, ramnose, galactose, xilose e manose). A biossíntese desses monômeros e a
incorporação dos mesmos na parede celular envolvem a atividade de várias enzimas e
substratos de nucleotídeo-açúcares, como, UDP-glicose, UDP-glucuronato, UDP-galactose,
UDP-xilose e UDP-arabinose (REITER e VANZIN, 2001).
As hemiceluloses constituem um grupo heterogêneo de polissacarídeos que se ligam
com firmeza à parede, apresentam conformação ramificada e não linear, constituindo uma
estrutura amorfa (TAIZ e ZEIGER, 2004).
Segundo Sticklen (2008), existem dois tipos de parede celular primária, que são
classificadas de acordo com o tipo de ligações cruzadas. Paredes Tipo I estão presentes nas
eudicotiledôneas e consistem em quantidades iguais de glucana e xiloglucana embebidas
numa matriz de pectina, enquanto as paredes do tipo II são encontradas em cereais e outras
gramíneas, tem glucuronoarabinoxilanas com suas glicanas de ligações cruzadas e carecem de
pectina e proteínas estruturais (CARPITA e McCANN, 2002).
Quando o crescimento celular é finalizado, em algumas células, internamente à parede
primária ocorre à deposição de camadas adicionais (GLÓRIA e GUERREIRO, 2003),
iniciando-se a formação da parede secundária (RAVEN, EVERT e EICHHORN, 2001), onde
as microfibrilas são depositadas por aposição, ou seja, por um arranjo ordenado. Estas
camadas são designadas S1, S2 e S3, sendo delimitadas pela mudança de deposição, que varia
nas diferentes camadas (Figura 2) (GLÓRIA e GUERREIRO, 2003).

4
Figura 2. Camadas S1, S2 e S3 da parede celular (Adaptado de STICKLEN, 2008).
Timell et al. (1967) afirmam que o número de camadas nas paredes secundárias pode
mudar em resposta a estímulos ambientais, e as mesmas em algumas espécies de plantas
podem ter mais do que três camadas. Por exemplo, GRITSCH e MURPHY (2005)
observaram até seis camadas distintas nas paredes secundárias de fibras de bambu.
Durante a deposição da parede secundária inicia-se a lignificação, que consiste em um
processo bioquímico que abrange desde a formação dos monolignóis até a polimerização da
lignina na parede celular (GLÓRIA e GUERREIRO, 2003; MONTEIRO, PEREIRA e
ABREU, 2004). Quando depositada, a lignina estabelece ligações covalentes e não covalentes
com a matriz de carboidratos, formando o chamado complexo lignina-carboidratos (CLC)
(SARKANEN, 1998; LAWOKO, HENRIKSSON e GELLERSTEDT, 2005).
A lignina é um complexo composto por unidades de fenilpropanos não linearmente e
ligados aleatoriamente, formados por três monômeros principais que são os álcoois p-
cumarílico, coniferílico e sinapílico (Figura 3).
Figura 3. Álcoois precursores da lignina p-hidroxifenila (H), guaiacila (G) e siringila (S).
Esses monolignois constituintes das paredes secundárias variam consideravelmente em
estrutura nos diferentes grupos vegetais. Em eudicotiledôneas lenhosas e nas
monocotiledôneas contêm, predominantemente, unidades guaiacila (G) e siringila (S) com

5
vestígios de unidades p-hidroxifenila (H); as gramíneas, geralmente, contêm níveis mais
elevados de unidades H; e em gimnospermas as ligninas consistem principalmente em
unidades de G com menores quantidades de unidades H (PATTATHIL et al., 2015).
A abundância relativa dessas unidades principais varia entre as espécies de plantas,
tecidos, tipos celulares e estágios de desenvolvimento (STEWART et al., 2009) e pode
ocorrer ligações em qualquer um dos diferentes locais em cada unidade fenólica, fazendo com
que diferentes tipos de ligações possam ocorrer em uma molécula de lignina como, ligações
β-O-4, α-O-4, β-5, 5-5, 4-O-5, β-1, e β-β (Figura 4). Embora estas sejam as dominantes, pelo
menos 20 diferentes tipos de ligações foram identificados (WHETTEN, MACKAY e
SEDEROFF, 1998).
Del Río et al. (2007a) afirmam que ligninas de gramíneas incluem as três unidades
ligadas a ferulatos e p-cumaratos, no entanto, Dixit e Nair (2014), citam que outros
monômeros típicos também podem ser encontrados em ligninas naturais.
Figura 4. Ligações inter-monoméricas comuns em uma molécula de lignina (Adapatado de
LATIF et al., 2015).
Essas formas complexas de paredes representam a maior parte da biomassa vegetal e
um imenso reservatório de carbono sob a forma de lignoceluloses. Vários dados estão
disponíveis para demonstrar que a composição química de lignocelulose é variável,
dependendo da espécie, do estádio de desenvolvimento e as condições ambientais (BOUDET,
2003), conforme a Tabela 1.

6
Tabela 1. Teor de celulose, hemicelulose e lignina de diferentes materiais lignocelulósicos e
de alguns resíduos agrícolas (%).
Material lignocelulósico Celulose Hemicelulose Lignina
Caule de Eudicotiledôneas 40-55 24-40 18-25
Caule de Gimnospermas 45-50 25-35 25-35
Gramíneas 25-40 35-50 10-30
Casca de nozes 25-30 25-30 30-40
Espiga de milho 45 35 15
Palha de trigo 50 30 15
Fonte: Adaptado de SUN e CHENG (2002).
As paredes celulares são resistentes à desconstrução por catalisadores biológicos ou
químicos, e esta propriedade é, muitas vezes, referida como “recalcitrância” (HIMMEL et al.,
2007) e é considerada como a principal barreira para a produção de biocombustíveis
celulósicos de baixo custo (HIMMEL et al., 2007; CHUNDAWAT et al., 2011). Desta forma,
é indiscutível obter uma melhor compreensão da estrutura e da arquitetura da parede celular, a
fim de desenvolver abordagens racionais para superar tal problema.
Pattathil et al. (2015) citam que as informações sobre as características estruturais e a
composição das paredes celulares têm sido obtidas a partir de análises de todos os
componentes ou de polímeros isoladamente. Esses autores consideram desafiador o estudo da
parede celular, em função do grande número de espécies vegetais ocorrentes na natureza,
aliado à complexidade estrutural, heterogeneidade e diversidade dessas estruturas.
2.2 Complexo lignina-carboidrato-ácidos fenólicos
O conhecimento do monolignois formadores da lignina e o sistema complexo formado
a partir de outras unidades químicas constituem uma importante linha de pesquisa, para
compreensão do funcionamento e desenvolvimento de diferentes grupos vegetais ao longo da
evolução. Esta organização ocorre quando ligninas se tornam ancoradas à parede primária
(YAMAMOTO, BOKELMAN e LEWIS, 1989; TERASHIMA, 1993), ocorrendo uma
adcrustação sobre a parede primária (JUNG e DEETZ, 1993), e continua durante o
amadurecimento da parede celular.
Uma parte da estrutura da lignina está ligada quimicamente às hemiceluloses, ou seja,
existe um complexo lignina-carboidrato (CLC) na estrutura química da parede celular. A
formação desse complexo também pode ocorrer entre os polissacarídeos, componentes
fenólicos e a própria lignina (GRABBER, RALPH e HATFIELD, 2000; BARAKAT et al.,
2007).
Os ácidos fenólicos são incorporados à parede celular em resposta ao estresse biótico,
pela via dos fenilpropanóides, que resulta na síntese de ácido cinâmico e derivados de ácido
benzoico. Esses últimos são esterificados e incorporados na fração da parede celular (DE
ASCENSAO e DUBERY, 2003).
Diversos estudos demonstram que existem interações físicas e químicas como ligações
de hidrogênio, forças de Van der Walls, e ligações do tipo éster ou éter, que formam uma
espécie de “rede” entre os componentes da parede celular. Estas ligações covalentes formadas
entre a lignina e os polissacarídeos, contribuem ainda mais para “recalcitrância” da parede
celular (SCALBERT et al., 1985; ALÉN, 2000; EBRINGEROVÁ, 2005). Algumas dessas

7
ligações foram descritas por Balakshin, Capanema e Chang (2007), que descreveu ligações
fenol glicosídeo (FGlc), ésteres (Est) e éteres benzílicos (Eb) (Figura 5).
Em que: FGlc: fenol glicosídeo; Est: ϒ-éster; Eb: éter benzílico; (A) ligação β-O-4; (A’) ligação β-O-4 com γ-
carbono acetilado; (A’’) ligação β-O-4 com carbono γ ligado a p-cumaril; (B) Estrutura resinol formada por
ligações β-β’, α-O- γ’ e γ-O- α’; (C) Estruturas fenilcumaranas formada por ligações β-5’ e α-O-4’; (D)
Estruturas spiro-dienona formada por ligações β-1’ e α-O- α; (E) Subestruturas α, β-diaril-éter; (AF) Ácido
ferúlico; (APC) Ácido p-cúmarico; Glc: Ácido glucorônico; Man: Manose; Gal: Galactose; Ara:Arabinose; Xil:
Xilose.
Figura 5. Principais ligações de CLC e subestruturas da lignina (Adaptado de YOU et al.,
2013).
Vários monolignóis estão conjugados à lignina (LU e RALPH, 2008; RALPH e
LANDUCCI, 2010; RALPH, 2010). As Angiospermas lenhosas apresentam baixo nível de
lignina aciladas (SARKANEN, CHANG e ALLAN, 1967), enquanto ligninas das fibras de
Hibiscus cannabinus e lignina em várias outras plantas são amplamente aciladas (RALPH,
1996; RALPH e LU, 1998; LU e RALPH, 2004; DEL RÍO et al., 2007b; DEL RÍO et al.,
2008; MARTINEZ et al., 2008).
Essa complexidade da estrutura e das ligações da lignina com os outros componentes
da parede celular pode ser explicada por dois fatores. O primeiro é causado pela diversidade
de grupos funcionais associados aos polissacarídeos e à lignina, como álcoois primários e
secundários, carboxilas e carbonilas. No caso de gramíneas, as hidroxilas fenólicas de ácidos
fenólicos esterificados constituem um tipo de grupo funcional adicional. O segundo ocorre
pela ação de diferentes enzimas, como glicosil-transferases e peroxidases, que participam na
construção da arquitetura da parede celular (CORNU et al., 1994).

8
As Poaceae produzem a enzima p-cumaril-CoA: monolignol transferase (PMT)
(WITHERS et al., 2012; PETRIK et al., 2014), formando monolignóis com carbono γ ligado a
uma estrutura p-cumaril que é incorporada no polímero de lignina crescente, em geral nos
indivíduos adultos (RALPH et al., 1994a; CRESTINI e ARGYROPOULOS, 1997).
Algumas madeiras têm ligninas parcialmente p-hidroxibenzoiladas, como por
exemplo, salgueiros (Salix spp.- Salicaceae) (LANDUCCI, DEKA e ROY, 1992), choupos e
álamos (Populus spp. - Salicaceae) (VENVERLOO, 1971; TOMIMURA, 1992a; MORREEL
et al., 2004; LU et al., 2004) e Aralia cordata (Araliaceae) (HIBINO et al., 1994). As
palmeiras também apresentam ligninas com alguns grupos p-hidroxibenzoila (TOMIMURA,
1992b; SUN, FANG e TOMKINSON, 1999; KURODA, OZAMA e UENO, 2001;
RENCORET et al., 2013).
Em plantas herbáceas, o CLC é estruturalmente diferente das lenhosas, por causa da
incorporação de hidroxicinamatos na parede da célula. A formação da "lignina-ferulato-
polissacarídeo" foi bem estabelecida pela da ligação do ácido ferúlico esterificado aos
carboidratos e éter à lignina (IIYAMA, LAM e STONE, 1990; RALPH et al., 1998;
BURANOV e MAZZA, 2008).
Os ácidos p-cumárico e ferúlico são bifuncionais e capazes de formar ligações éster ou
éter por reação dos seus grupos carboxílicos ou fenólicos, respectivamente. O ácido p-
cumárico está associado, principalmente, à lignina; ácido ferúlico, por outro lado, é
principalmente esterificado com a hemicelulose (ATUSHI, AZUMA e KOSHIJIMA, 1984;
SCALBERT et al, 1985). Esta ligação pode ocorrer pela ligação do ácido à lignina na posição
α ou β (Figura 6) (LAM, KADOYA e IIYAMA, 2001).
(a) Ligação α-éter entre lignina e hidroxicinamatos
(b) Ligação β-éter entre lignina e hidroxicinamatos
R=H: p-cumarato, R=OCH3: ferulato
Figura 6. Esquema representativo de uma ligação α éter (a) e β éter (b) formadas entre lignina
e hemicelulose por meio de um hidroxicinamato (LAM, KADOYA e IIYAMA, 2001).

9
Os resultados de vários estudos de composição da parede celular e lignificação em
monocotiledôneas (HARTLEY et al., 1990; HATFIELD, 1993; RALPH et al., 1994a,b)
mostraram que o conteúdo e a composição dos ácidos hidroxicinâmicos são dependentes da
localização morfológica e a fase de diferenciação. Tem sido relatado que o ácido ferúlico
rapidamente se deposita nas paredes das células na sua fase inicial de lignificação,
subsequentemente o ácido p-cumárico, sendo o majoritário entre os ácidos hidroxicinâmicos
(MORRISON et al., 1998).
No campo teórico, tem havido debate sobre a estrutura do complexo lignina-
carboidrato (CLC), quanto o processamento de biomassa, a presença do CLC ou a sua
formação durante o processamento. Esse complexo parece dificultar o processamento químico
ou bioquímico de lignocelulose, como deslignificação durante a polpação química (GIERER e
WANNSTRÖM, 1986; IVERSON e WANNSTRÖM, 1986; CHOI, CHOI e FAIX, 2007) e
na fermentação de materiais lignocelulósicos para a produção de etanol (KIM et al., 2003;
AITA, SALVI e WALKER, 2011).
2.3 Família Arecaceae
Arecaceae é um grupo monofilético que inclui 183 gêneros e 2.364 espécies
(GOVAERTS e DRANSFIELD, 2005; DRANSFIELD et al., 2008). Grande número de
espécies de palmeiras se distribui nos trópicos e poucas nas regiões subtropicais
(HENDERSON, GALEANO e BERNAL, 1995).
McCann e Roberts (1991), Carpita e Gibeaut (1993) e Carpita (1996) realizaram uma
extensa revisão sobre as diferenças entre as espécies de monocotiledôneas e outras
angiospermas quanto à estrutura do polímero, arquitetura e biossíntese dos polissacárideos
não celulósicos e suas associações/ligações. Também foram analisados os tipos de substâncias
aromáticas e proteínas estruturais por meio de ligações covalentes à parede primária e
secundária.
Horn et al. (2009) estudaram os principais padrões evolutivos para Arecaceae com
base no estudo anatômico da lâmina foliar dos gêneros de palmeiras. Nos caules das
monocotiledôneas, o sistema vascular primário é formado por feixes dispersos no tecido
parenquimático formando um atactostelo (GLÓRIA e GUERREIRO, 2003).
As espécies de monocotiledôneas geralmente não possuem crescimento secundário a
partir de um câmbio, mas podem apresentar um aumento em diâmetro, como por exemplo,
nas palmeiras, por meio de um espessamento resultante da divisão e aumento das células do
parênquima fundamental sem que haja estabelecimento de uma faixa meristemática contínua
(GLÓRIA e GUERREIRO, 2003). Esse tipo de crescimento é chamado de secundário difuso,
porque a atividade meristemática não está restrita a determinada região do órgão.
As camadas secundárias consecutivas da parede celular com diferentes ângulos de
microfibrila (hélice Z) são depositadas durante o envelhecimento, resultando em uma
estrutura multilamelar da parede celular, o que permite uma adaptação contínua em nível
celular ao longo da vida útil da planta (PARAMESWARAN e LIESE, 1976; LIESE, 1987,
BHAT, LIESE e SCHMITT, 1990).
Recentemente, foi mostrado para Washingtonia robusta, que as palmeiras podem
seguir uma estratégia diferente de ajustar o desempenho mecânico nas calotas de fibras
vasculares (RÜGGEBERG et al., 2008), uma vez que as fibras da palmeira permanecem vivas
no corpo da planta, elas podem adicionar continuamente novas camadas de parede celular até

10
que o lume da célula inteira esteja cheio. Além disso, as fibras podem aumentar o grau de
lignificação da parede celular (RÜGGEBERG et al., 2008).
As palmeiras pertencem a uma família importante dentre as monocotiledôneas que
afetam significativamente a economia e o cotidiano de milhões de pessoas no mundo. A sua
contribuição para a economia mundial, as economias locais e os estilos de vida de pessoas são
enormes, por causa do surpreendente número de produtos que podem ser obtidos (RIVAS,
BARBIERI e MAIA, 2012). Estas contribuem para a alimentação, habitação, móveis, energia,
vestuário e jardins.
Entre os produtos alimentares podem ser destacados os frutos, sementes, o "palmito",
méis, 'sagu' (material com amido extraído a partir do centro dos troncos), diferentes bebidas
obtidas a partir da seiva ou os frutos e açúcar cristalizada a partir da seiva. Outros tipos de
produtos são as fibras, óleos e ceras (JONES, 1995).
Em decorrência dos usos citados, os caules muitas vezes são cortados e não são
utilizados (ISMAIL e MAMAT, 2002). A fim de utilizar estes resíduos eficientemente,
muitos estudos têm sido realizados sobre usá-los como matéria-prima para produtos à base de
resíduos ligno-celulósicos, incluindo painel de média densidade, celulose e papel (WANG et
al., 2014; TLIJANI et al., 2014; MURATA et al., 2015; DERROUICHE et al., 2015; LU et
al., 2015).
2.4 Euterpe oleracea Mart.
Euterpe oleracea Mart. (açaizeiro) é uma palmeira delgada e alta que pode atingir uma
altura de 20 a 25 m e apresenta farta perfilhação e alcança, no estado nativo, cerca de 20
palmeiras por “touceira” (das quais pelo menos três em produção). Cada touceira produz entre
6 a 8 cachos com 2,5 kg cada um, representando de 15 a 20 kg de frutos por palmeira (em
duas safras). Os troncos são lisos, roliços, longos, de cor clara, sem espinhos (MATTOS,
1993). É uma espécie florestal típica da região que compõe o estuário do Amazonas, com
características de cultura permanente, o que a torna indicada para as condições tropicais de
grande precipitação pluviométrica (superior a 2.300 mm anuais), elevada temperatura e
umidade relativa do ar e lençol freático superficial (MIRANDA et al., 2001).
É uma das espécies mais abundantes e frequentes nas áreas de várzea, constituindo-se
na espécie nativa de maior importância econômica para a região do estuário amazônico.
Estudos realizados na região têm demonstrado que a concentração de açaizeiros pode atingir
até 25% da população botânica das áreas de várzea (ANDERSON et al., 1985). As maiores
áreas ocupadas com essa espécie, encontram-se na Amazônia Oriental Brasileira, mais
precisamente na região do estuário do rio Amazonas, considerada como seu centro de origem
e onde existem densas e diversificadas populações, que segundo Calzavara (1972) e
Cavalcante (1991), ocupavam em torno de 1 milhão de hectares.
Em 2002, observou-se o aumento da área colhida com açaí de 16.115 ha (2002) para
26.671 ha (2004), ou seja, uma taxa de crescimento de 65,50% num período de três anos. Essa
significativa taxa média de crescimento da área colhida do açaí é uma resposta do crescimento
da demanda regional, nacional e mesmo internacional do açaí, que pode estar induzindo os
produtores rurais a aumentarem área plantada do açaí. A expansão da área colhida indica o
grande interesse dos produtores rurais quanto à necessidade da formação de uma base
produtiva de cultivo do açaí como condição básica para a atração de agroindústrias de
processamento (SANTANA, CARVALHO e MENDES, 2006).

11
O açaizeiro inicia seu ciclo de produção de frutos 4 anos após o plantio, podendo
eventualmente, algumas plantas, entrarem em fase de produção aos 3 anos de idade. A
produção máxima ocorre entre 5 e 6 anos de idade, por terem atingido a estabilidade
produtiva e, consequentemente, um percentual de frutificação mais elevado (NOGUEIRA et
al., 1998).
Essa é uma espécie que tem multiplicidade de usos, dentre os quais se destacam as
folhas para cobertura de casas, fibras, celulose, ração animal, adubo e proteção de plantações;
os frutos para bebida, alimento, adubo, curtimento de couro, álcool, remédio anti-diarréico e
ração animal; o palmito para alimento; as inflorescências para adubo, vassouras e proteção de
plantações; os estipes para construções, celulose, lenha e isolamento elétrico e as raízes são
usados como vermífugo (JARDIM, 2005).
A exploração e o aproveitamento dos frutos e palmito do açaizeiro são as atividades
mais importantes e lucrativas praticadas nas várzeas do estuário amazônico. A produção de
frutos e de palmito depende da relação entre o número de touceiras de açaizeiros por hectare,
demais palmeiras e espécies lenhosas e os produtores ribeirinhos já perceberam que a
exploração madeireira contribui para o surgimento e ampliação dos açaizais (QUEIROZ e
MOCHIUTTI, 2002).
3 MATERIAL E MÉTODOS
3.1 Coleta do material
Para as análises foram utilizados três indivíduos de Euterpe oleracea Mart. com 15
anos, coletados no Bairro de Patrimônio, próximo à divisa com o estado de São Paulo,
localizado no Km 586 da Rodovia Rio-Santos, munícipio de Paraty (23°11'41" latitude Sul e
44°43'41" longitude oeste), estado do Rio de Janeiro. Os indivíduos foram coletados,
processados e registrados na xiloteca do Departamento de Produtos Florestais, Instituto de
Florestas, Universidade Federal Rural do Rio de Janeiro, Seropédica (Tabela 2).
Tabela 2. Relação de indivíduos, número de registros e parâmentros dendrométricos das
amostras de Euterpe oleracea estudados.
Amostra Número do Registro Diâmetro da base
(cm)
Altura (m)
1 7639 16 12
2 7640 18 14
3 7641 16,5 12
Para o estudo da química lignoídica e da anatomia de E. oleracea, foram retiradas
amostras da base da palmeira (Figura 7A), por ser a parte mais desenvolvida do caule, de duas
regiões: externa (RE - mais próxima à casca) e interna (RI - mais central do disco) (Figura
7B).

12
Figura 7. Esquema do caule de Euterpe oleracea. (A) Região de coleta das amostras. (B)
Seção transversal do caule de Euterpe oleracea com a identificação das regiões estudadas (RE
– Região externa; RI – Região interna).
3.2 Preparo das amostras
Fragmentos do caule de E.oleracea das regiões externa e interna, foram secos ao ar e
moídos em moinho de facas do tipo Wiley, classificadas e utilizadas a fração que ficou retida
na peneira número 24 internacional (malha 60 ASTM). Após o preparo das amostras, as
partículas foram submetidas à extração com cicloexano, acetato de etila e metanol em um
extrator do tipo soxhlet, por um período de 24 horas para cada solvente (ABREU et al., 2006).
Com o material extraído foi realizado também um tratamento alcalino ao utilizar
NaOH 1% (TAPPI, 1979), totalizando quatro tratamentos:
1) Amostra RE extraída
2) Amostra RE extraída e tratada com NaOH 1%
3) Amostra RI extraída
4) Amostra RI extraída e tratada com NaOH 1%

13
3.3 Análises químicas
Com o material obtido das amostras RE e RI extraídas com cicloexano, acetato de etila
e metanol, e amostras RE e RI extraídas e tratadas com NaOH 1%, foram obtidos a lignina
Klason, teor de holocelulose e teor de α-celulose (ABREU et al., 2006). Também foi realizada
a determinação de ácidos fenólicos por meio de cromatografia líquida de alta eficiência
(CLAE) das amostras proveniente s da região externa e interna do caule e dos extratos obtidos
com o solvente acetato de etila e metanol (GEETHA et al., 2011).
Para a caracterização da estrutura da lignina das amostras RE e RI, esta foi isolada
pelo método de Björkman, nos quais foram realizadas as análises no infravermelho, RMN 1H
e 13
C no estado líquido e sólido.
Um fluxograma da parte experimental, consta na Figura 8.
Figura 8. Fluxograma de preparo das amostras da região externa (RE) e interna (RI) do caule
de Euterpe oleracea.
3.4 Determinação do teor de holocelulose
Para determinação do teor de holocelulose foi utilizado o método de cloração. Foram
utilizadas amostras extraídas e extraídas e tratadas com NaOH 1% da região externa (RE) e
interna (RI) de E. oleracea. Para tanto, foram utilizadas 2,5 g de amostra seca e adicionaram-

14
se 8 mL de água destilada quente, 0,5 mL de ácido acético e 1g de clorito de sódio. Depois,
essas amostras foram mantidas em banho-maria a 70°C, sendo a cada 1 h adicionado mais 0,5
mL de ácido acético e 1g de clorito de sódio até as fibras ficarem completamente separadas. A
amostra foi mantida em repouso sem adição de qualquer reagente, durante 24 horas.
A amostra foi filtrada sob vácuo, e lavada com água destilada até a cor amarela e o
odor de cloro desaparecer completamente. O material foi seco em uma estufa a 103 ± 2ºC
durante 24 horas e depois foi pesado (ABREU et al., 2006). A porcentagem de holocelulose
foi calculada pela Equação 1.
olocelulose eso seco do resíduo (mg)
eso da amostra de madeira 1 (1)
3.5 Determinação do teor de α-celulose
Foram utilizadas amostras extraídas e extraídas e tratadas com NaOH 1% das regiões
externa (RE) e interna (RI) de E. oleracea para determinação do teor de α-celulose. Para isto,
dois gramas de holocelulose seca foram transferidos para um recipiente fechado e adicionados
10 mL da solução de NaOH 17,5% mantendo a temperatura a 20 ºC, em um banho-maria.
Depois foram adicionados, em intervalos de 5 minutos, mais 5 mL da solução de NaOH
17,5%, agitando-se cuidadosamente. A amostra foi mantida a 20 ºC por 30 minutos,
perfazendo um total de tempo de tratamento de 45 minutos. Foram adicionados 33 mL de
água destilada a 20ºC, permanecendo em repouso em repouso durante 1 hora, antes de
filtragem.
O material foi filtrado sob vácuo, em um filtro de porosidade média, previamente
tarado e lavado com 100 mL de NaOH 8,3%. Posteriormente foi lavado duas vezes com água
destilada. No próprio filtro foram adicionados 15 mL de ácido acético 10%, aguardando 3
minutos e foi retirado todo ácido acético sob vácuo. O resíduo celulósico foi lavado
novamente com 250 mL de água destilada e seco em uma estufa a 103 ± 2ºC, durante 24
horas. O material foi resfriado em um dessecador por 1 hora. A porcentagem de α-celulose foi
calculada pela Equação 2.
α celulose eso seco do resíduo celulósico
eso da amostra holocelulósica 1 (2)
3.6 Lignina Klason
Para determinação do teor da lignina Klason foi utilizado amostras extraídas e
extraídas e tratadas com NaOH 1% da região externa (RE) e interna (RI) de E. oleracea.
Foram utilizadas aproximadamente 300 mg de amostra que foram digeridas com ácido
sulfúrico 72%, sendo homogeneizada por agitação contínua durante 1 minuto e conservada
por 1 hora entre 25 e 30°C em banho-maria. O material obtido foi diluído a 84 mL de água
destilada, permanecendo em refluxo por 4 horas.
O produto obtido foi lavado com 500 mL de água destilada quente em um funil de
placa sinterizada e seco em estufa a 103 ± 2ºC durante 24 horas (EFFLAND, 1977). A
porcentagem de lignina foi calculada pela Equação 3.
Lignina eso seco do resíduo (mg)
eso da amostra de madeira 1 (3)

15
3.7 Lignina de Björkman
As amostras extraídas e tratadas com NaOH 1% de E. oleracea (RE e RI) foram
trituradas em um moinho de bolas de aço inoxidável para a redução da granulometria até
apresentar nível coloidal ao utilizar a velocidade de 310 rpm durante 36 horas. O material foi
transferido para um erlenmeyer e extraído com dioxano (96:4) por 4 dias sob agitação
constante. A solução de dioxano obtida foi filtrada por duas vezes em um funil de Büchner ao
utilizar papel de filtro. A solução de dioxano:água foi evaporada em um evaporador rotatório
a 40°C em 55 mbar de pressão negativa.
O material concentrado foi dissolvido com uma solução de ácido acético:água (9:1) e
filtrado com um funil de placa sinterizada n° 3 e com placa sinterizada n° 2. O material
filtrado foi transferido para um funil de separação, no qual foi gotejado em 1 L de água
bidesionizada sob agitação. Foi realizada uma nova filtragem da solução aquosa em um funil
de Büchner com papel de filtro e ao término desse procedimento o produto permaneceu em
um dessecador contendo anidrido fosfórico.
Após a secagem, o material foi dissolvido com uma solução de 1,2-dicloroetano e
etanol absoluto (2:1), e o material dissolvido foi gotejado em éter etílico por duas vezes,
alternada por duas operações de centrifugação. A lavagem constituiu a última etapa e foi
realizada com éter de petróleo.
3.8 Acetilação
Após a obtenção da lignina de Björkman, 50 mg desse material foram dissolvidos em
1 mL de piridina e adicionado 1 mL de anidrido acético na proporção 1:1 e permaneceu na
solução reacional sob agitação a frio durante 72 horas. Depois, a lignina foi recuperada pela
precipitação da solução reacional em éter etílico e lavada por 3 vezes com este mesmo
solvente sob centrifugação. Posteriormente, o material foi seco em dessecador de vidro sob
pressão reduzida, na presença de KOH durante 3 dias.
3.9 Espectroscopia no infravermelho
Foi realizada análises de espectroscopia no infravermelho do caule (região interna e
externa) de E. oleracea nas seguintes condições de tratamento:
1) amostras extraídas conforme metodologia descrita no item 3.2;
2) amostras extraídas e tratadas com NaOH 1%;
3) amostras obtidas da lignina de Björkman;
Os espectros de infravermelho foram registrados em um espectrômetro (VARIAN
640-IR FT-IR spectrometer) ao utilizar 1mg de amostra e 100mg de KBr para confecção das
pastilhas para análise e realizadas no modo experimental de transmitância com 4 cm-1
de
resolução, 32 varreduras, amplitude espectral entre 4000-400cm-1
.
Com a intenção de comparar as alterações dos grupos funcionais com as mudanças dos
espectros, após o tratamento alcalino, foram calculadas as absorbâncias relativas (AR) para os
principais sinais de absorção de acordo com a Equação 4.
A Aab
A15 5-15 3
1 (4)

16
Em que: AR é a absorbância relativa da banda de absorção; Aab é a absorbância da
banda de absorção AB; A1505-1503 é a absorbância do sinal de referência a 1505-1503 cm-1
.
As absorbâncias relativas foram calculadas relativamente à absorbância do sinal de
absorção em 1505 e 1503 cm-1
para as amostras RE e RI, respectivamente. Este sinal foi
escolhido como referência, pois se admite que o número de anéis aromáticos seja constante
durante o tratamento e, portanto, a intensidade do sinal em 1505 e 1503 cm-1
causado pelas
vibrações C=C do anel aromático, deve ser constante. Foi realizada a correção de linha de
base para calcular a absorbância de determinado sinal de absorção, e esta foi determinada
como a altura do pico (SMITH, 1982; BYKOV, 2008).
3.10 Ressonância magnética nuclear (RMN 13
C) no estado sólido do caule de Euterpe
oleracea
As amostras RE e RI extraídas e extraídas e tratadas com NaOH 1% foram
processadas em um moinho de bolas para redução da granulometria. Após moagem o material
foi seco e em um dessecador contendo anidrido fosfórico. Foram utilizados 500 mg para
análise de RMN 13
C no estado sólido (ABREU et al., 2006). Os espectros foram obtidos em
um aparelho Bruke Avance II 400 MHz, com rotor de 4 mm, com a técnica CP/MAS com
giro de 8000 Hz com polarização cruzada.
3.11 Ressonância magnética nuclear (RMN 1H e
13C) em solução da lignina de Euterpe
oleracea
A elucidação estrutural da lignina das amostras RE e RI de E. oleracea foi realizada
por técnica espectroscópica de ressonância magnética nuclear (RMN) para os átomos de
carbono (RMN de 13
C) e prótons (RMN de 1H) uni e bidimensionais.
Os espectros foram obtidos na Central Analítica da Universidade Federal Rural do Rio
de Janeiro (Instituto de Ciências Exatas/Departamento de Química) e registrados em um
aparelho Bruker Advance II 400 MHz. As amostras das ligninas acetiladas RE e RI (item 3.8)
foram dissolvidas em CDCl3 (clorofórmio deuterado) e os deslocamentos químicos (δ) foram
expressos em partes por milhão (ppm).
3.12 Determinação de Ácidos Fenólicos da parede celular por Cromatografia Líquida de
Alta Eficiência (CLAE)
Para a determinação do teor de ácido fenólicos na parede celular foi utilizada a
metodologia descrita por Geetha et al. (2011). Esta análise foi realizada com as amostras
extraídas e extraídas e tratadas com NaOH 1% de E.oleracea (RE e RI). Além da análise dos
ácidos fenólicos presentes na parede celular também foi realizada a análise cromatográfica
destes no estado livre, nos quais foram utilizados os extratos obtidos com acetato de etila e
metanol conforme metodologia descrita no item 3.2.
As amostras (100 g) do caule foram extraídas num aparelho de soxhlet com metanol
(300 mL x 3), até o material vegetal ficar incolor. O extrato metanólico combinado foi
concentrado à vácuo em um evaporador rotativo sob vácuo. O resíduo obtido foi dissolvido
em 50 mL de água e submetido a hidrólise ácida com HCl a 5% (10 mL). A solução
hidrolisada foi extraída com acetato de etila (50 mL x 3) e a camada orgânica combinada foi

17
seca sobre sulfato de sódio anidro. A solução foi concentrada e o resíduo resultante dissolvido
em metanol para análise cromatográfica (GEETHA et al., 2011).
A determinação do perfil químico nas amostras foi realizada por um cromatógrafo
líquido Prominence (Shimadzu) equipado com duas bombas (LC-20AT), detector de
fotodiodos (SPD-M20A), injetor automático (SIL-10A) e forno para coluna (CTO-20A) foi
utilizado para determinação das substâncias.
A separação cromatográfica foi realizada em uma coluna analítica de fase reversa C-
18 (25cm 4,6 mm, 5 μm de partícula, Luna-Phenomenex), tendo como fase móvel água
Mill-Q com 1% de ácido acético (solvente A) e metanol (solvente B), fluxo constante de 1,0
mL/min e temperatura de 28 ºC. O volume de injeção foi de 2 μL e a detecção foi realizada a
280nm. O gradiente de eluição iniciou-se com 35% do solvente B até 80% em 2 minutos, 80 a
92% de B em 4 min, 92 a 35% B em 2 min, em que foi mantido a 35% de B por 3 minutos.
Os cromatogramas foram adquiridos e processados pelo “software” LcSolution (Shimadzu).
3.13 Microscopia óptica de campo claro
Foram coletados fragmentos dos caules dos três indivíduos e fixados em FAA 70% -
formaldeído 40%, ácido acético glacial e álcool 70%, na proporção 1:1:18 (JOHANSEN,
1940), por 3 dias e conservados em álcool 70%.
As amostras foram incluídas em polietileno glicol (PEG) 1500 e seções transversais
obtidas em micrótomo de deslize, na espessura média de 20 μm. Essas seções foram
clarificadas com hipoclorito de sódio a 50%, neutralizadas em água acética (1:500), coradas
com uma mistura de azul de astra 2% e safranina 0,5% em solução aquosa e montadas em
lâminas semipermanentes (BUKATSCH, 1972).
Para a dissociação dos elementos traqueais das amostras das regiões interna e externa do
caule foi utilizada o método modificado de Franklin (1945), que consiste na mistura de
peróxido de hidrogênio 30% e ácido acético glacial, na proporção 1:1. Os fragmentos obtidos
foram corados com safranina aquosa 1% e depois montados em lâminas semipermanentes.
Foram realizadas 25 mensurações das fibras de cada indivíduo segundo as
recomendações das normas IAWA Committee (1989). A partir dos valores dos diâmetros total
e do lume das fibras, foram obtidos os parâmetros anatômicos espessura da parede (μm) e
fração parede (%), conforme Equações 5 e 6.
E (μm) Dt-Dl
2 (5)
F ( ) 2 E
Dt 1 (6)
Em que: EP é espessura da parede (μm); Dt é Diâmetro total (μm); Dl é Diâmetro do
lúmen (μm); FP é a fração parede (%).
Na análise dos dados, foi utilizada estatística descritiva, em que os resultados obtidos
para as propriedades anatômicas que foram tabulados e interpretados por meio dos valores
mínimos, máximos, média e desvio padrão.

18
3.14 Testes histoquímicos
Para a realização dos testes histoquímicos foram obtidas seções transversais do
material fresco de E. oleracea (RE e RI). O teste de Wiesner confirma a presença de lignina
com grupos cinamaldeído, com a cor variando de rosa a vermelho (VALLET et al., 1996); e
Maüle que consiste em três tratamentos sequenciais do tecido lignificado com permanganato
de potássio 1%, ácido clorídrico 3% e hidróxido de amônio concentrado, para identificação da
lignina do tipo guaiacílica (G) pela cor marrom e siringílica (S) pela cor vermelha (LIN e
DENCE, 1992).
3.15 Microscopia de fluorescência
Para a microscopia de fluorescência, seções transversais do caule (regiões interna e
externa) foram transferidas para uma solução de auramina 0,01% por 15 segundos, no qual
esse fluorocromo fluoresce amarelo quando ligado a lignina (HALL et al., 1989), e em
seguida lavadas com água bidesionizada. As mesmas foram observadas em microscópio de
fluorescência utilizando cubo de excitação azul em 450-480 nm e filtro FITC (U-MWB2)
(BARROS e MIGUENS, 1998) e as fotomicrografias foram obtidas com um microscópio
óptico (Olympus BX 51) com um sistema digital de processamento de imagens (cellf imaging
software).
4 RESULTADOS E DISCUSSÃO
4.1 Determinação do teor dos constituintes macromoleculares
Os resultados das análises da composição química das amostras do caule de E.
oleracea e das amostras tratadas com NaOH 1% estão apresentados na Tabela 3. O teor de
celulose, hemicelulose e lignina correspondem a 90% da massa total do caule. Os 10%
restante são ceras, taninos, goma e algumas substâncias inorgânicas, conhecidas como
extrativos.
O teor de α-celulose e de holocelulose das amostras RE e RI do caule apresentou
valores médios bastante similares entre estas. Estes valores foram semelhantes aos
encontrados em fibras do estipe dos palmiteiros branco e macho (IPT, 1988). No entanto,
quando comparado, esses resultados foram superiores aos encontrados em diversas espécies
de madeiras e também de várias monocotiledôneas como consta na Tabela 4.
Tabela 3. Teor de holocelulose e lignina das diferentes amostras do caule de Euterpe
oleracea.
Amostras Holocelulose (%) α-Celulose (%) Lignina (%)
RE extraída 74,40 64,08 10,33
RE extraída + NaOH 1% 80,48 64,92 8,50 RI extraída 74,42 63,08 15,66
RI extraída + NaOH 1% 79,15 68,17 11,33 RE – Região externa; RI – Região interna do caule de E. oleracea.

19
Tabela 4. Valores de referência dos constituintes macromoleculares de diversas espécies
Espécie Teor dos constituintes (%)
Referência Celulose Hemicelulose Lignina
Calamus manan 39,05 - 22,22 Kalima e Jasni (2004)
Calamus occidentalis 49,95 22,39
Bambu 57-66* 21-31 García Hortal (2007)
Saccharum sp.
- - 20,88 Paula et al. (2011)
54,3-55,2 16,8 25,3-24,3 Satyanarayana, Guimarães e
Wypych (2007)
49-62 - 19-24 García Hortal (2007)
Agave sisalana 59,18 24,58 6,97 Salazar e Leão (2006)
Cocos nucifera 45,47 20,36 31,83
Agave sisalana 74-75,2 10-13,9 7,6-8
Satyanarayana, Guimarães e
Wypych (2007)
Juta 60 22,1 15,9
Curauá 70,7 21,1 7,5-11,1
Piaçava 31,6 - 48,4
Açai 57,3* - 19,8
IPT (1988) Palmiteiro branco 61,2* - 15,7
Palmiteiro macho 65,9* - 18,9
Fonte: Adaptado de FONSECA (2012).
Foi obtido um teor de lignina de 10,33% e 15,66% para amostras RE e RI extraídas,
respectivamente. Essas variações observadas podem ser atribuídas ao estágio de maturação.
Os valores observados foram inferiores a outras biomassas, como o endocarpo de babaçu
(27,9%), bambu gigante (22,7%) e madeiras de Eucalyptus grandis (23,9%) e Pinus oocarpa
(25,1%), obtidos por Silva, Barrichelo e Brito (1986), Marinho et al. (2012), Pereira et al.
(2000) e Moraes, Nascimento e Melo (2005), respectivamente.
O baixo teor de lignina de um material lignocelulósico reduz o tempo de produção de
polpa e a carga química em comparação com as de outras matérias-primas não madeireiras
(DIAZ et al., 2007; LOPEZ et al., 2008).
Após o tratamento com NaOH 1%, ocorreu um aumento do teor de holocelulose de
6,08% e 4,73% (RE e RI, respectivamente), porém houve uma redução do teor de lignina,
resultando na dissolução de 1,83% e 4,33% da lignina original das amostras RE e RI,
respectivamente, permitindo propor que parte do material incrustante foi solubilizada. Tal fato
se relaciona à ligação da lignina da amostra RI e polissacarídeos, que são relativamente mais
lábeis entre os monômeros da lignina, ou ainda entre lignina e polissacarídeos.
Esses resultados podem ser explicados pelo tratamento alcalino que pode promover a
solubilização de hemiceluloses por ruptura e quebra de ligações de hidrogênio. Essas
substâncias, quando ligadas por meio de grupamento do tipo éster, podem ser quebradas por
uma base (XIAO, SUN e SUN, 2001).
O pré-tratamento alcalino é uma das várias tecnologias que têm sido investigadas, no
qual se utilizam reagentes como hidróxido de sódio (CARRILLO et al., 2005; SILVERSTEIN
et al., 2007) e o álcali cliva ligações hidrolisáveis em ligações de lignina e de polissacarídeos,
o que provoca uma redução no grau de polimerização e cristalinidade, inchaço das fibras, bem
como a ruptura da estrutura da lignina (HSU, 1996).
Gierer (1985) e Scalbert e Monties (1986) afirmam que as partes ácidas, como grupos
carboxílicos ionizados ou fenólicos, em solução alcalina, podem promover a solubilização da
lignina. Xiao, Sun e Sun (2001) afirmam que essa elevada solubilidade da lignina e

20
hemiceluloses também pode ser atribuída à quebra de ligações éster entre os ácidos
hidroxicinâmicos, como os ácidos p-cumárico e ferúlico.
No entanto, o caule de E. oleracea, ao ser analisado por técnica espectroscópica
(infravermelho e RMN 1H e
13C) e por CLAE-DAD permitiu observar a presença do ácido p-
hidroxibenzóico e ácido protocatecuico (Figura 9), formando ligações entre esses e a lignina.
Além disso, as palmeiras também têm parcialmente p-hidroxibenzoil conectado à lignina
(TOMIMURA, 1992a, b; SUN et al., 1999b; KURODA, OZAMA e UENO, 2001;
RENCORET et al., 2013).
Figura 9. Estrutura dos derivados benzóicos. (A) Ácido p-hidroxibenzóico e (B) ácido
protocatecuico.
4.2 Análise por espectroscopia de infravermelho (FTIR) do caule de Euterpe oleracea
A espectroscopia de infravermelho (IV) com transformada de Fourier é comumente
utilizada para identificar os grupos funcionais de biomassa lignocelulósica e as alterações
causadas por diferentes tratamentos.
Nesse estudo, as amostras extraídas e extraídas e tratadas com NaOH 1% de E.
oleracea (RE e RI) foram submetidas à análise por FTIR e apresentaram espectros similiares,
apresentando importates regiões de absorção das amostras RE e RI (Figuras 10 e 11).
Também foram submetidas à análise por FTIR as amostras RE e RI das ligninas de Björkman
para uma melhor análise da lignina dessa espécie.
A caracterização dos sinais de absorção no IV das amostras foi realizada
principalmente de modo qualitativo por comparação com dados da literatura, conforme
descrito por Lin e Dence (1992), Boeriu et al. (2004), Schwanninger et al. (2004), Kubo e
Kadla (2005), Yan et al. (2009), Gwon et al. (2010), Xu et al. (2013); Latif et al (2015).

21
Figura 10. Espectros de infravermelho das amostras extraídas, extraídas e tratadas com
NaOH 1% e lignina de Björkman e da região externa (RE) do caule de Euterpe oleracea.
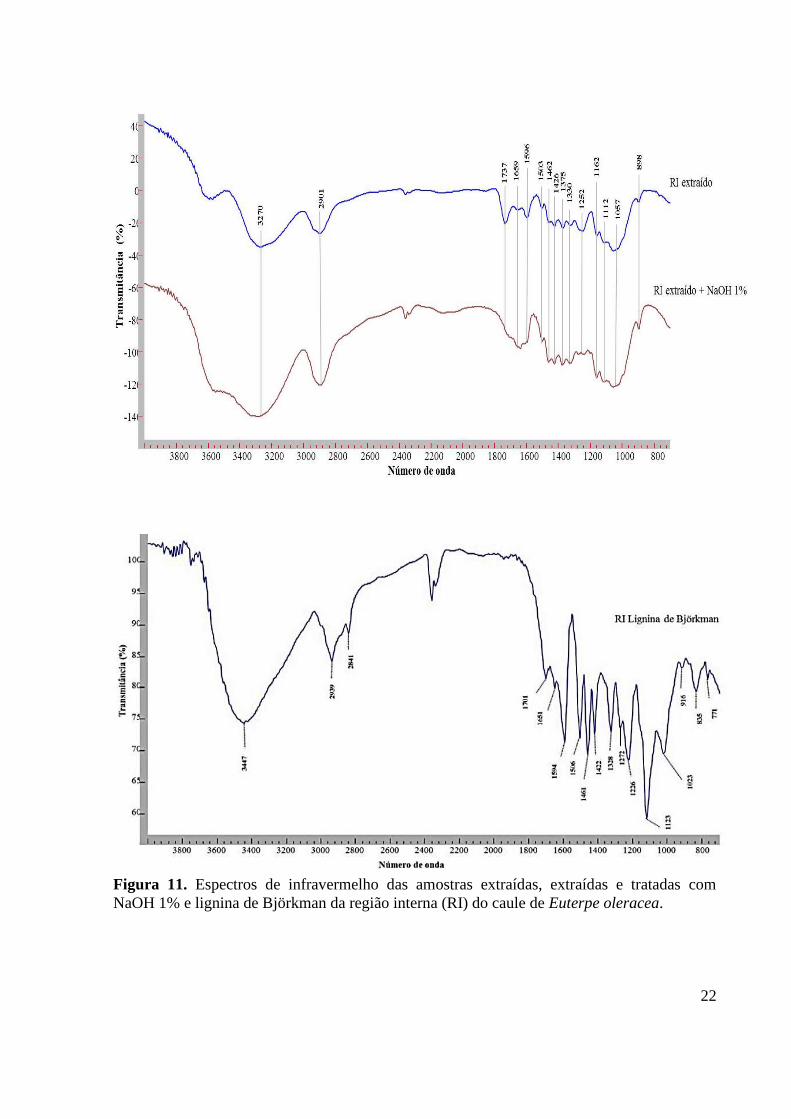
22
Figura 11. Espectros de infravermelho das amostras extraídas, extraídas e tratadas com
NaOH 1% e lignina de Björkman da região interna (RI) do caule de Euterpe oleracea.

23
Os espectros de ambas as amostras (RE e RI) extraídas foram similares, e
apresentaram um sinal forte de estiramento de O-H em 3271-3270 cm-1
. A absorbância em
2904-2850 cm-1
foi decorrente do estiramento de C-H em grupos metoxílicos aromáticos e em
grupos metílicos e metilênicos das cadeias laterais da lignina.
Também foi observado o sinal em 1737-1735 cm-1
nos espectros e foi atribuída ao
estiramento de C=O de éster certamente derivados dos grupos acetilicos ligados às
hemiceluloses.
Os sinais a partir de 1600 a 1450 cm-1
foram devido à vibração do esqueleto aromático
da lignina. Os sinais em 1596, 1505-1503, 1427-1426 cm-1
surgem a partir das vibrações do
esqueleto aromático da lignina (-C=C-C- e -C=C-C=C- ou ligação dos monômeros da lignina
β-1, β-β, 5-5, e β-5).
A absorção em 1462-1456 cm-1
foi atribuído à deformação angular assimétricas de C-
H de grupamento metila e metilenos. Estes sinais podem ser observados com mais clareza
após o isolamento da lignina pelo método de Björkman das amostras de E.oleracea (RE e RI).
O sinal em 1382-1375 cm-1
foi atribuído ao dobramento de C-H de grupos CH3 da
celulose e hemicelulose e em 1225 cm-1
ao estiramento C-O do grupo hidroxílico fenólico na
lignina. Além destes sinais, a lignina é composta por unidades guaiacila propano (G) e
unidades de siringila propano (S) que contêm um e dois grupos metoxílicos, respectivamente.
Os espectros, Figuras 10 e 11, mostram sinais característicos de lignina que são
observados em 1272 cm-1
para as unidades de guaiacila propano (G) ligado a C=O e 1329-
1328 cm-1
para unidades de siringila propano (S) acrescido do anel G condensado (isto é, anel
G substituído na posição 5), e em torno de 1225 cm-1
atribuído ao estiramento de C-C mais C-
O mais C-C(=O)-C; G condensado > G eterificado (FAIX, 1991).
De acordo com o sistema de classificação de lignina com espectros infravermelhos por
Faix (1991), ao se analisar os espectros das ligninas de Björkman das amostras RE e RI de E.
oleracea, pode-se concluir que as mesmas pertencem aos tipos típicos de lignina guaiacila
siringila (GS) pelas seguintes características: a absorbância em 1462 cm-1
superior a
absorbância do sinal de referência a 1505-1503 cm-1
; o sinal em 1329-1328 cm-1
superior ao
sinal em 1272 cm-1
; o sinal em 1272 cm-1
baixo (algumas vezes aparece apenas um ombro); o
sinal em 1125 cm-1
muito alto e o sinal em 835 cm-1
pronunciado.
Os sinais em 1160 cm-1
e 1049 cm-1
foi atribuído, respectivamente, ao estiramento de
C-O-C da celulose ou hemicelulose e estiramento C-O da celulose, hemicelulose ou lignina.
Sun, Lawther e Bank (1996) e Sun et al. (2002) afirmam que o sinal em 1044 cm-1
que é
atribuído ao C-O e C-C é característico para xilanas e com contribuição da ligação glicosídica
(C-O-C). O sinal em 898 cm-1
corresponde ao C1 do anel e é atribuída às ligações glicosídicas
(1→4) entre unidades de xilose em hemicelulose. As atribuições típicas dos sinais de
infravermelho FT-IR que representam os principais componentes da parede celular, constam
na Tabela 5.

24
Tabela 5. Atribuições dos sinais dos espectros das amostras de Euterpe oleracea (região
externa e interna).
Para analisar as diferenças dos espectros das amostras do caule de E.oleracea após
tratamento com NaOH 1%, os sinais foram normalizados pela razão da intensidade da
absorção de cada sinal pela da intensidade da absorção do sinal em 1505-1503 cm-1
para
amostras RE e RI, respectivamente. Este sinal corresponde ao estiramento de anéis
aromáticos, que foi selecionado como uma referência interna, uma vez que tem uma
Número
de ondas
(cm-1
)
Referências Atribuição Polímero
3417-3445
FAIX (1991); SCHWANNINGER et al.
(2004); KUBO e KADLA (2005); YAN et al.
(2009); XU et al. (2013)
Estiramento de O-H Celulose,
Lignina
2903-2904
FAIX (1991); SCHWANNINGER et al.
(2004); KUBO e KADLA (2005); YAN et al.
(2009); XU et al. (2013)
Estiramento de C-H Celulose,
Lignina
1736
FAIX (1991); SCHWANNINGER et al.
(2004); YAN et al. (2009); SILLS e
GOSSETT (2012); XU et al. (2013)
Estiramento de C=O de éster Hemicelulose
1595
FAIX (1991); SCHWANNINGER et al.
(2004); YAN et al. (2009); SILLS e
GOSSETT (2012)
Vibrações do esqueleto
aromático; S>G;
G Condensado > G eterificado
Lignina
1500-1510
FAIX (1991); SCHWANNINGER et al.
(2004); YAN et al. (2009); SILLS e
GOSSETT (2012); XU et al. (2013)
Vibração do esqueleto aromático Lignina
1457-1459 FAIX (1991); YAN et al. (2009); SILLS e
GOSSETT (2012); XU et al. (2013) Deformações angular de C-H Lignina
1425-1427 KUBO e KADLA (2005) Deformação de C-H no plano Lignina
1376-1377 SCHWANNINGER et al. (2004); SILLS e
GOSSETT (2012); XU et al. (2013)
Deformação angular de C-H de
CH3
Celulose,
hemicelulose,
lignina
1322-1329
FAIX (1991); SCHWANNINGER et al.
(2004) ; KUBO e KADLA (2005);YAN et al.
(2009); XU et al. (2013)
C-O em anel siringílico Lignina
1280 SILLS e GOSSETT (2012); XU et al. (2013) Deformação angular de C-H Celulose
1272
FAIX (1991); SCHWANNINGER et al.
(2004); SILLS e GOSSETT (2012); XU et al.
(2013)
Vibração do anel guaiacil,
influenciada por estiramento
C=O
Lignina
1226-1227
FAIX (1991); SCHWANNINGER et al.
(2004); YAN et al. (2009); SILLS e
GOSSETT (2012); XU et al. (2013)
Estiramento de C-C e C-O Lignina
1160-1161 SCHWANNINGER et al. (2004); SILLS e
GOSSETT (2012); XU et al. (2013) Estiramento assimétrico C-O-C
Celulose,
hemicelulose
1049-1051 BOERIU et al. (2004) Estiramento C-O, C-C, C-OH
Celulose,
hemicelulose,
lignina
1024-1026 SILLS e GOSSETT (2012) Estiramento C-O e C-C-O
Celulose,
hemicelulose,
lignina
898 SCHWANNINGER et al. (2004)
Deformação de C-H de anel
aromático; Deformação de
grupos de C1-H
Celulose,
hemicelulose,
lignina

25
intensidade relativamente constante. Os resultados dessas diferenças podem ser observados na
Tabela 6.
A maioria dos sinais característicos de absorção da lignina e hemicelulose nas
amostras tratadas com NaOH 1% foi significativamente enfraquecido quando comparado aos
sinais das amostras extraídas do caule de E. oleracea (Tabela 6) sugerindo que a lignina e
hemicelulose foram parcialmente degradadas durante o pré-tratamento.
Tabela 6. Sinais FTIR normalizados pela razão da intensidade em 1505 (RE) e 1503 cm-1
(RI) do caule de Euterpe oleracea.
Número de onda
(cm-1
)
Razão das intensidades dos
sinais
da amostra RE Número de onda
(cm-1
)
Razão das intensidades dos
sinais
da amostra RI
Extraídas
Extraídas e
tratadas com
NaOH 1%
Extraídas
Extraídas e
tratadas com
NaOH 1%
3272-3273 2,67 2,22 3270-3285 1,45 1,73
2900-2911 1,72 1,87 2900-2901 1,29 1,31
1736 1,64 - 1737 1,18 -
1649-1650 1,43 1,41 1657-1659 1,03 0,85
1457-1461 1,74 1,50 1462-1463 1,17 1,04
1426-1428 1,88 1,75 1426-1427 1,20 1,06
1374-1383 1,88 1,74 1375-1380 1,24 1,06
1320-1332 1,58 1,64 1330-1331 1,19 0,91
1242-1253 1,89 1,29 1243-1252 1,27 0,91
1161-1162 2,12 1,93 1162 1,31 1,21
1112-1114 2,44 2,15 1112-1115 1,40 1,26
1053-1059 2,78 2,30 1057-1059 1,49 1,35
898-899 0,53 0,61 897-898 0,92 0,58
O símbolo (-) refere-se à ausência do sinal após o tratamento com NaOH 1%.
As alterações mais significativas após tratamento com NaOH 1% foram observadas
em 1737-1736 cm-1
no qual o sinal desapareceu tanto nas amostras RE e RI; em 1162-1161
cm-1
(estiramento assimétrico de C-O de grupos ésteres) e em 1059-1053 cm-1
(estiramento C-
O, C-C, C-OH) que apresentaram uma redução das intensidades nas amostras RE e RI. Este
resultado permite entender que ocorreu uma reação de saponificação durante o pré-tratamento
alcalino (Figura 12). Tal reação quebra a ligação éster entre lignina e carboidratos e libera a
celulose a partir do encapsulamento de lignina, tornando mais exposta a celulose (HE et al.,
2008).
Figura 12. Esquema do complexo lignina-carboidrato (CLC) reagindo com NaOH (HE et al.,
2008).

26
Koullas et al. (1992) afirmam que o mecanismo de hidrólise alcalina ocorre pela
saponificação de ligações de ésteres intermoleculares, que reticulam hemiceluloses de xilanas
e outros componentes, por exemplo, lignina e outra hemicelulose. Durot, Gaudard e Kurek
(2003) destacam que o tratamento alcalino pode liberar um pouco de celulose e hemicelulose
por hidrólise das ligações éster entre lignina e celulose ou hemicelulose.
4.3 Análises por espectrometria de ressonância magnética nuclear de 13
C no estado sólido
do caule de Euterpe oleracea
A anisotropia do material lignocelulósico conduz a espectros com sinais muitos largos,
visto que os sinais estão concentrados na região entre 50 e 110 ppm, no qual a grande parte é
atribuído a carbonos da estrutura da celulose, com contribuição de sinais de hemicelulose e
lignina (REZENDE et al., 2011). Nesse trabalho, essa técnica foi utilizada para avaliar do
efeito do tratamento alcalino com NaOH 1% com as amostras do caule (RE e RI) de
E.oleracea e conhecer quais as estruturas dos grupos aromáticos no material.
Os espectros de RMN de 13
C e suas atribuições estão apresentados na Tabela 7. As
amostras de E. oleracea (RE e RI) extraídas apresentaram um perfil similar. No entanto, após
o tratamento alcalino com NaOH 1%, ocorre o desaparecimento dos sinais de carbono do
esqueleto lignoídico e diferenciando dos sinais δH 173 e 22 causado pela a hidrólise dos
ésteres e acetatos.
Tabela 7. Atribuições aos deslocamentos químicos observados nos espectros de RMN 13
C
(estado sólido) das amostras do caule de Euterpe oleracea (RE e RI).
Deslocamento químico (ppm) Atribuições
RE
extraído
RE
extraído +
NaOH 1%
RI extraído
RI extraído
+ NaOH
1%
173,45 - 173,81 - COOR, ésteres de glicosídeos e de acetato
154,24 153,51 153,87 154,24 Siringil éter ligada, C-3/C-5
133,80 133, 80 C-4 de Siringil
121,59 122,34 C-1 de Guaiacil, C-4 de Siringil
106,50 106,64 106,64 106,57 C-5 de Guaiacil
105,92 105,92 105,99 105,70 C-1 Arabinofuranosil
89,74 89,81 89,74 89,71 C-1 de celulose, C-1 de xilanas
85,19 85,05 85,05 85,12 C-4 de celulose cristalina
75,80 75,80 75,80 75,80 C-4 de celulose amorfa, C-4 de xilanas
73,35 73,28 73,20 73,20 Celulose e xilanas, C-2/C-3/C-5
65,62 65,76 65,84 65,76 C-6 de celulose cristalina, C-5 de xilanas
63,24 63,09 63,53 63,09 C-6 de celulose amorfa
56,66 57,03 56,81 56,74 Metoxilas aromáticas
21,78 - 22,07 - Grupos metileno alifáticos; acetatos

27
Nas amostras RE e RI extraídas pode-se observar que os sinais mais intensos foram de
carbonos de celulose e da cadeia C3 da lignina que aparecem entre 60 a 110 ppm. Os sinais
em torno de 60 ppm foram atribuídos a C6, de 70 a 80 ppm para C2-C5 (Figura 13).
Os sinais de carbonos de metilas e carboxilas de grupos acetílicos ligados a
hemiceluloses estão indicadas em 21,78 – 22,07 e 173,45 – 173,81 ppm, para amostra RE e
RI, respectivamente (Figura 13).
Os sinais menos intensos em 56,66 – 56,38 ppm foram atribuídos respectivamente a
grupos metoxílicos ligados a sistemas aromáticos de lignina. O sinal intenso em δH 154,23 –
153,87 foi atribuído a carbonos originados em sistema aromático, podendo estar sustentando
grupos -OCH3, –OH ou OR. Segundo Lin e Dence (1992), os sinais em torno de 180 ppm
correspondem a carboxilas conjugadas de ácidos p-hidroxicinâmicos e de segmentos
moleculares da lignina.
Para as amostras RE e RI extraídas e tratadas com NaOH 1%, pode-se obsevar que os
sinais a 21,78 – 22,07 e 173,45 – 173,81 ppm, atribuídos a grupos acetil de hemiceluloses,
desapareceram, confirmando a remoção de hemiceluloses em ambas as amostras (Figura 13).
Os sinais em 130-140 ppm e 154-158 ppm podem estar representando os carbonos do
anel siringil sendo 136-138 ppm do C-4 e 152-158 ppm dos carbonos C-3 e C-5. Apesar de
serem carbonos originados sp2, o carbono C-4 está vizinho a dois grupos originados em orto
que, desta forma, exerce efeito mesomérico de proteção. Na faixa de 130-140 ppm estão
também representando os carbonos C-1 do anel guaiacila com o deslocamento do carbono em
aproximadamente 130 ppm.
Da mesma forma, os sinais em δH 130-136 e de 154-158 foram reduzidos
gradualmente, o que indica a remoção parcial do teor de lignina das amostras RE e RI (Figura
13). Esses resultados também foram encontrados por Karnaouri, Rova e Christakopoulos
(2016) que estudaram diferentes formas de pré-tratamento em cascas de bétula visando
fracionamento de seus componentes. Essas observações também estão de acordo com os
trabalhos de Stewart et al. (1997), Wikberg e Mannu (2004) e Martins et al. (2006).
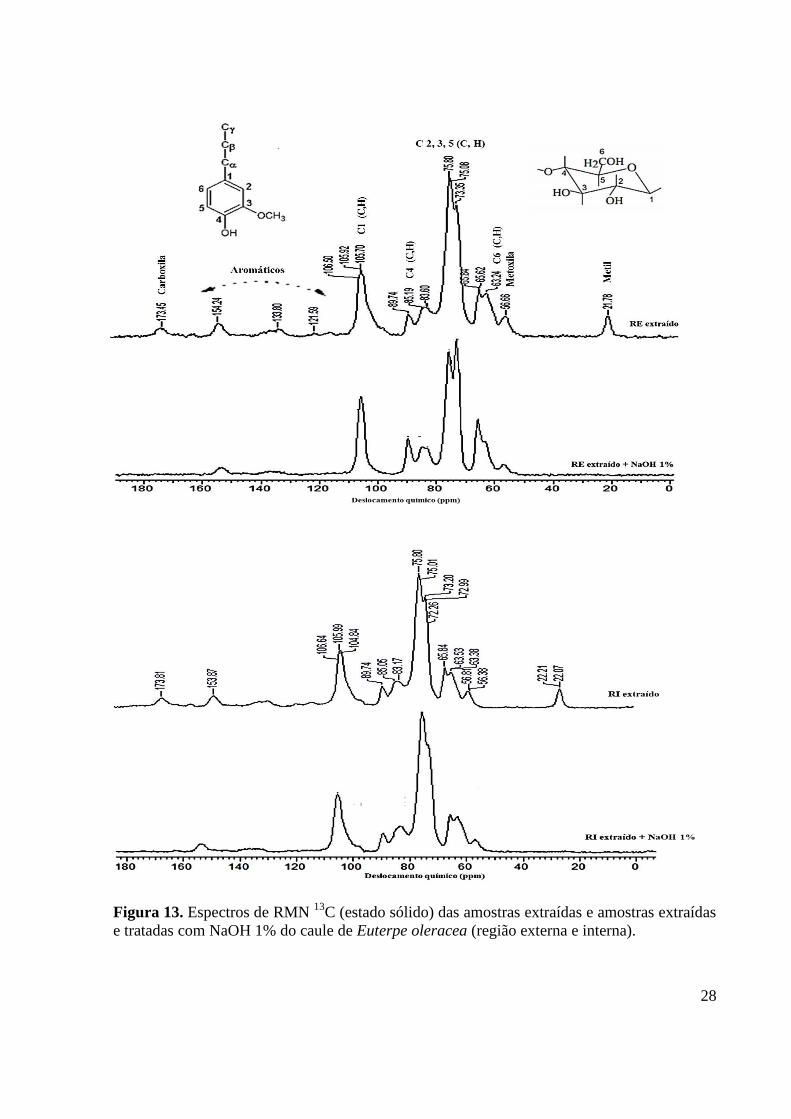
28
Figura 13. Espectros de RMN 13
C (estado sólido) das amostras extraídas e amostras extraídas
e tratadas com NaOH 1% do caule de Euterpe oleracea (região externa e interna).

29
4.4 Análises por espectrometria de ressonância magnética nuclear de 1H
13C em solução da
lignina de Euterpe oleracea
As técnicas analíticas de RMN 1H e
13C têm sido amplamente aplicadas para a
identificação da estrutura da lignina e podem ser utilizadas para atribuir estruturas diferentes a
várias fontes de lignina. Nimz et al. (1981) compararam as diferenças estruturais entre as
ligninas de folhosas, coníferas e gramíneas com análise de espectros RMN 13
C de lignina
acetilada, revelando as características estruturais das ligninas e justificando a sua classificação
como ligninas G (coníferas), ligninas GS (folhosas), ligninas HGS (gramíneas) ou ligninas
HG (madeira de compressão).
Os espectros de RMN 1H das amostras de lignina acetilada de E. oleracea (RE e RI)
foram similares e os resultados das análises foram realizados por comparação com os dados
da literatura e estão apresentados na Tabela 8, cujas frequências estão agrupadas em regiões
principais do espectro das ligninas.
Tabela 8. Atribuição dos sinais de hidrogênio em regiões do espectro de RMN 1H das
ligninas.
Região Deslocamento químico
( Grupos funcionais contendo hidrogênios responsáveis pelo
sinal
1 7,00-8,00 Hidroxila fenólica
2 6,00-7,00 Ar-H; Olefínico
3 4,00-5,00 H-C=O; Ar-CH-O
4 3,7 OCH3; OCH2
5 1,7-2,5 Metoxílico e H-C-Ar; H3-C-C
Referências: (MORAIS, 1992; ABREU e OERTEL, 1999; GONÇALVES et al., 2000; SALIBA et al., 2001;
YAN, XU e YU, 2009).
Os sinais entre 6,0 e 8,0 ppm são atribuídos aos hidrogênios de anéis aromáticos em
unidades G e S, nos quais os sinais em 6,7 e 6,8 ppm representam os hidrogênios aromáticos
em unidades G, enquanto 6,6 ppm originam-se das unidades S (FAIX, GRUNWALD e
BEINHOFF, 1992).
A maior intensidade do sinal em H 6,60 do que em H 6,90 (Figura 14), permite
sugerir que a lignina foi composta por uma proporção relativamente elevada de núcleos
siringílicos do que de guaiacilicos, tanto das amostras RE e RI de E. oleracea. Esses
resultados também foram encontrados por Jin, Jin e Shao (2012) ao estudarem a lignina
isolada pela metodologia de Björkman das fibras de Hibiscus cannabinus (Kenaf).
Os sinais em H 2,0, 2,1 e 2,2 correspondem a absorção de metilas adjacentes a
ligações duplas ou a grupos carboxílicos (MARCHESSAULT et al., 1982) e os sinais fortes
em H 3,4 e 3,7 podem ser atribuídos às metoxilas (-OCH3). Os sinais de H 4,1-4,5 são
compatíveis aos Hα, Hβ e Hγ da unidade C3 da lignina (Figura 14).
Lapierre, Lallemand e Monties (1982) afirmam que o sinal em torno de 4,9 ppm
representa Hα em ligações β-O-4 e os sinais na região de H 4,6-4,7 e 4,0-4,5 são originados
de Hβ e Hγ de estruturas β-O-4', respectivamente.
Um sinal acentuado em H 7,29 é dos hidrogênios do clorofórmio não deuterado que
aparece como impureza do solvente deuterado utilizado ou como resíduo de solvente

30
proveniente da extração do produto na acetilação. O pequeno sinal em H 7,9 foi atribuído aos
hidrogênios de aldeídos como nas estruturas de cinamaldeído e benzaldeído (Figura 14).
Figura 14. Espectros de RMN 1H das ligninas acetiladas de Euterpe oleracea das regiões
(RE) e (RI).

31
As amostras de lignina de E. oleracea (RE e RI) também foram analisadas com
espectros de RMN 13
C em solução com experimentos de sequência de pulso DEPTQ. Esses
espectros representam sinais em duas fases, diferenciando de acordo com a multiplicidade. No
caso da Figura 15, os sinais que aparecem na fase superior são C e CH2 e os sinais na fase
inferior representam os carbonos e metílicos (CH3) e metinicos (CH).
Os espectros das amostras apresentaram sinais em 20-30 ppm que foram atribuídos a
átomos de carbono saturados em cadeias alifáticas. Sun et al. (1997) afirmam que os sinais
que apareceram no espectro entre 22,2-33,8 ppm para –CH2- foram atribuídos aos grupos α e
β- ou γ-metilenos não oxigenados das cadeias laterais n-propil ou, mais certamente, de
hidrocarbonetos como impureza no processamento de acetilação. Xiao, Sun e Sun (2001)
afirmam que os sinais entre 13 e 33 ppm podem ser atribuídos a grupos metil na cadeia
propílica ou a algumas impurezas alifáticas.
Os sinais dos Cγ, Cα e Cβ das ligações éter β-O-4 aparecem em 62, 74 e 80 ppm,
respectivamente. Estes resultados também foram encontrados por Sun et al. (1997) ao estudar
lignina das fibras do fruto de palmeira.
Os átomos de carbono de sistemas aromáticos em ligninas apresentam sinais na faixa
entre 104 a 160 ppm (NIMZ et al., 1981; SUZUKI et al., 1998). O espectro de RMN 13
C das
ligninas de E. oleracea (RE e RI) foi relativamente simples na região de sistema aromático
mostrando sinais de CH em 104 e 153, que podem ser atribuídos ao C2/6 e C3/5 em unidades
de siringila (Figura 15), respectivamente. Esta análise confirma as observações da
espectroscopia no IV de que a lignina do caule de E. oleracea é do tipo guaiacila-siringila
(GS).
As unidades siringila (S) têm simetria, seus carbonos 2/6 dos anéis aromáticos
apresentaram deslocamento químico em torno de 104 ppm e os seus carbonos quaternários
oxigenados 3/5 estão a 153 ppm (Figura 15). Esta simetria separa-os dos seus homólogos
guaiacila (G), nos quais os carbonos protonados 2, 5 e 6 variam entre 110 a 125 ppm, embora
se verifique alguma sobreposição com os carbonos quaternários 3 e 4, que variam de 145 a
147 ppm (NIMZ et al., 1981; SUZUKI et al., 1998; JIN, JIN e SHAO, 2012).
Com base em estudos de modelo de compostos de lignina, o sinal do C5 em unidades
guaiacila substituídas (condensadas) é protegido (menor deslocamento químico) e os valores
do deslocamento químico (δ) nas estruturas em que C5-5 e C5-O-4 variam em grande medida
dependendo dos seus substituintes no C1 sendo eterificados em seus grupos fenólicos (NIMZ
et al., 1984; MIYAKOSHI e CHEN, 1991).
Os sinais amplos de 169-170 ppm nos espectros de 13
C das ligninas de E.oleracea (RE
e RI) sugerem a presença de carbonos de carboxilas de éster alifáticos ou de sistema
aromático (Figura 15). Esses resultados foram semelhantes aos encontrados por Lüdemann e
Nimz (1973) ao estudarem a madeira de Fagus silvatica e Picea excelsa e por Jin, Jin e Shao
(2012) para a lignina das fibras de Hibiscus cannabinus (Kenaf).
A maioria dos sinais observados para a lignina de E. oleracea (RE e RI) são
semelhantes aos previamente observados em espectro de lignina de bagaço de cana-de-açúcar
(RAMIRES et al., 2010; MANCERA et al., 2010), de bambu (SUN et al., 2013), de palha de
centeio (SUN et al., 2000), de cevada (SUN et al., 2002), de trigo (SUN, FANG e
TOMKISON, 2000) e da palmeira Elaeis guineensis (LU et al., 2015).
Os espectros de RMN 13
C das amostras RE e RI também apresentaram sinais em 111,
113 e 123 ppm (Figura 15), nos quais podem ser atribuídos a presença da estrutura de
derivados de ácidos benzoicos na parede celular de E. oleracea. Lu et al. (2015) afirmam que
estas estruturas são utilizadas como "monômeros" no processo de lignificação nas palmeiras

32
(bem como em salgueiros, choupos e álamos) e que este processo ocorre pelas transferases
que são responsáveis pela produção de conjugados de monolignois p-hidroxibenzoatos.
Figura 15. Espectros de RMN 13
C DEPTQ (400 MHz, CDCl3) da lignina das amostras RE e
RI de Euterpe oleracea.

33
Também foram realizados experimentos de RMN 2D (1Hx
13C-COSY) que foi bastante
útil para elucidar a estrutura química da lignina analisada. Os espectros de RMN 2D (1Hx
13C-
COSY) podem ser obtidos com diferentes sequencias de pulsos conduzindo a obtenção de
sinais de interação de acoplamento a uma ligação [JH-C (HSQC)] e a duas outras ligações
[2,3
JH-C (HMBC)]. Esta técnica tem sido utilizada, por vezes em conjunção com a RMN 13
C
quantitativa para identificar novas estruturas da lignina e/ou reticulação de interpolímeros,
como por exemplo, no caso dos chamados complexos lignina-carboidrato que resultam do
acoplamento da lignina-polissacárideos (BALAKSHIN, CAPANEMA e CHANG, 2007,
BALAKSHIN et al., 2011, CHEN et al., 2012, KIM e RALPH, 2010, MANSFIELD et al.,
2012).
Os espectros HSQC de RE e RI mostraram sinais de interação a uma ligação H2,6 e
C2,6 (δH 6,8 com δc 104) e os espectros MBC dos hidrogênios 2,6 (δH 6,8) com C-3,5 (δH
153); C-1 (δC 134) e C-α (δCH 74). A intensidade dos sinais de δH 2,6 e as interações
facilmente observados mesmo com pouca acumulação, realizado nos experimentos dos
espectros 2D, que o material analisado tem grande quantidade de siringila na estrutura
(Figuras 16, 17, 18 e 19). Os espectros HSQC mostraram mais correlações 1H-
13C (
1JHC) entre
os respectivos núcleos de cadeias alifáticas laterais, oxigenadas e não oxigenadas, e de anéis
aromáticos (Figuras 16 e 17).
Os sinais de interação 1JH-C, observados na frequência de núcleos de sistema alifático
oxigenado (δH/δC 3,9/6,2/62-80 ppm), e nas regiões de núcleos de sistemas aromáticos (δH/δC
6,6-7,8/104-132 ppm) são compatíveis com frequências de núcleos de ligninas nas amostras
analisadas (Figuras 16 e 17). O sinal de interação 1JH-C em δH/δC 3,7/56 ppm observado nos
espectros das amostras de lignina de E. oleracea (respectivamente RE e RI) corresponde à
correlação dos sinais de grupos metoxilas nas estruturas (DEL RÍO et al., 2009).
Também foram observados sinais das correlações 1JH-C entre os hidrogênios de CH2-ϒ
livre (δH/δC 3,9/61) e de CH2 sustentando grupo aceto ila (δH 4,1 e δH/δC 4,4/62-64), cujos
valores são compatíveis com a presença desses grupos nesta posição. Os sinais de 1JHC em
δH 4,4-4,8 com δC 74 e 8 podem ser atribuídos aos carbonos Cα e Cβ da unidade C3 da
lignina.
Os principais sinais de interação 1JH-C na região de absorção de núcleos de sistemas
aromáticos dos espectros HSQC correspondem aos anéis benzênicos das diferentes unidades
de lignina. As unidades S apresentaram um sinal para correlação H2,6/C2,6 em δH/δC 6,6/104
ppm e as unidades G em 7,2/120 para a correlação H6–C6, 6,9/113 de H-2/C-2 e 6,8/112 de H-
5/C-5. A intensidade do sinal 7,6/104 revela a grande quantidade de unidades S na estrutura
da lignina.
Outro sinal significativo na região de absorção de núcleos de sistema aromático foi
observado nas correlações H2,6-C2,6 em 7,9/131 que foi atribuído às subestrutura de p-
hidroxibenzoato de sinapila (PB).

34
Figura 16. Espectro HSQC (400/100 MHz, CDCl3) da fração da lignina acetilada da região externa de Euterpe oleracea (RE).

35
Figura 17. Espectro HSQC (400/100 MHz, CDCl3) da fração da lignina acetilada da região externa de Euterpe oleracea (RI).

36
Figura 18. Espectro HMBC (400/100 MHz, CDCl3) da fração da lignina acetilada da região externa de Euterpe oleracea (RE).

37
Figura 19. Espectro HMBC (400/100 MHz, CDCl3) da fração da lignina acetilada da região externa de Euterpe oleracea (RI).

38
As atribuições das principais correlações das amostras das ligninas acetiladas RE e RI
estão listadas na Tabela 9. Estes resultados foram comparados com os resultados encontrados
na literatura obtidos na análise de RMN com valores tabelados para lignina.
Tabela 9. Atribuição dos sinais dos espectros de RMN 2D-HSQC da lignina acetilada de
Euterpe oleracea (RE e RI).
Sigla δH/δC (ppm) Atribuições Referências
-OMe 3,7/56 H-C em metoxilas
SUN et al., (2010); SHE et al.,
(2012); WEN et al., (2014);
OLIVEIRA (2015)
Aϒ 3,9/61 e 3,7/59 Hϒ-Cϒ em subestruturas β-O-4
SUN et al., (2010); WEN et al.,
(2014)
A’ϒ 4,1 e 4,4/64 Hϒ-Cϒ em subestruturas β-O-4’
ϒ-acetiladas
SUN et al., (2010); SHE et al.,
(2012)
Dβ 5,9/75 Hα-Cα em subestruturas spiro-
dienona OLIVEIRA (2015)
Dα 4,5/80 Hβ-Cβ em spiro-dienona OLIVEIRA (2015)
S2,6 6,6/104 H2,6 -C2,6 em unidades siringila SUN et al., (2010); SHE et al.,
(2012); WEN et al., (2014)
G6 7,2/120 H6–C6 em unidades guaiacila SUN et al., (2010); SHE et al.,
(2012); WEN et al., (2014)
PB2,6 7,9/131 H2,6 -C2,6 em ácido p-
hidroxibenzoico WEN et al., (2014)
Esses resultados também foram obtidos por Yuan et al. (2011) ao analisarem a lignina
da madeira de Populus tomentosa Carr. (choupo) de crescimento rápido que é parcialmente
acilada no carbono γ nas ligações aril éter de β-O-4’ das cadeias laterais. No entanto, alguns
grupos esterificados naturais presentes na lignina podem ter sido hidrolisados e removidos
quando se utilizou o presente método de isolamento.
Esse resultado é compatível com as análises estruturais da lignina de Populus tremula
(SMITH, 1955), clones híbridos de Salix (LANDUCCI, DEKA e ROY, 1992), Elaeis
guineensis (SUN et al., 1999), Populus tremula x Populus alba (MEYERMANS et al. 2000),
Populus tremula x Populus alba (MORREEL et al., 2004) que mostraram que os ésteres de γ-
p-hidroxibenzoato estão presentes.
As análises dos sinais na região de núcleos de anéis aromáticos dos espectros
revelaram que a fração da lignina das amostras RE e RI de E. oleracea pode ser considerada
como uma lignina do tipo GS contendo unidades de ácidos p-hidroxibenzoico e
protocatecuico, nos quais a presença destes foram confirmados por meio da análise
cromatográfica. As principais subestruturas da lignina estão apresentadas na Figura 20 e a
proposta para lignina de E.oleracea (RE e RI) está apresentada na Figura 21.

39
Figura 20. Principais unidades estruturais presentes na lignina das amostras RE e RI de
Euterpe oleracea. A – Ligação β-O-4; A’ – Subestrutura β-O-4’ ϒ-acetiladas; D –
Subestrutura spiro-dienona; G- Unidade guaiacil; S- Unidade siringil; PB – Subestrutura p-
hidroxibenzoato.
Figura 21. Estrutura da lignina da palmeira Euterpe oleracea, obtida pelas informações (A)
do espectro de RMN 1H e (B) RMN
13C.

40
A lignina siringila é mais reativa em sistemas alcalinos que a guaiacila (DEL RIO et
al., 2005) e pode ser justificado pelas unidades guaiacila terem um carbono na posição C-5
disponível para realizar ligação carbono-carbono muito forte, contribuindo para um grau de
condensação mais alto da lignina e consequentemente dificultando a remoção desta
macromolécula.
Chiang (2005) relata que a reatividade da lignina depende da frequência de suas
unidades monoméricas guaiacila (G) e siringila (S). Mais unidades dos monômeros siringila,
ou elevadas relações S/G proporcionam elevada reatividade da lignina, facilitando a sua
retirada em processos que possuem este objetivo, como na produção industrial de polpa de
celulose. O índice de deslignificação é diretamente proporcional à relação S/G e não depende
apenas da acessibilidade da lignina, mas de sua estrutura química (RODRIGUES et al., 1999).
Desta forma, a compreensão da estrutura da lignina pode fornecer informações importantes
quanto à recalcitrância e desta maneira auxiliar na estratégia para um melhor aproveitamento
da matéria prima (PATTATHIL et al., 2015).
4.5 Análises cromatográficas de ácidos fenólicos dos extratos e do caule de Eutepe oleracea
A determinação de ácidos fenólicos na parede celular pode oferecer informações
importantes para a compreensão de como estes estão envolvidos na parede celular e na
conexão entre a lignina e os carboidratos, bem como a influência destes na recalcitrância da
lignina.
Os ácidos fenólicos ocorrem nas plantas nas formas livres que são extraídos da matriz
utilizando solventes orgânicos e ligados que se encontram conjugados a parede celular e estão
associados por ligação éster ou éter. A hidrólise ácida ou básica é necessária para liberar estes
compostos ligados a matriz celular. A identificação dos ácidos foi realizada pela comparação
dos tempos de retenção (TR) em minutos, juntamente com as análises dos respectivos
espectros de absorção no UV em comparação com padrões autênticos (Figura 22).

41
Figura 22. Curvas de UV dos padrões contidos nas amostras RE e RI de E. oleracea
adquiridas pela análise por CLAE-DAD.
As análises cromatográficas dos extratos de E.oleracea das amostras RE e RI obtidos
com acetato de etila mostraram dois picos (Figura 23A e B), e com metanol mostraram dois
picos para a amostra RE (Figura 21C), porém a amostra RI apresentou cinco picos (Figura
21D), dos quais estes foram identificados como ácido protocatecuico (TR 4,060 min) e ácido
p-hidroxibenzóico (TR 5,543 min).
Ao observar a área dos picos é possível visualizar as diferentes concentrações desses
ácidos fenólicos presentes em cada amostra (Figura 23). Foi constatada a presença dos ácidos
protocatecuico e p-hidroxibenzoico em todas as amostras, sendo o ácido protocatecuico
predominante nos extratos obtidos com acetato de etila e o ácido p-hidroxibenzoico
predominante nos extratos metanólicos.
Shein et al. (2001) afirmam que a acúmulo de ácido p-hidroxibenzoico desempenha
um papel importante na proteção de células de coníferas agindo como um agente fungicida no
estágio de penetração fúngica do citosol e o ácido protocatecuico representa um dos principais
compostos antioxidantes, que foram extraídos da casca de pinheiro (SALLEEM, KIVELA e
PIHLAJA, 2003).

42
Vários compostos fenólicos simples ocorrem em plantas. Os derivados do ácido
cinâmico são os mais abundantes, enquanto os de ácido benzóico e aldeídos estão restritos a
alguns grupos (JUNG, DEETZ e GARST, 1983; HARTLEY e KEENE, 1984). Foi sugerido
que as substâncias fenólicas limitam a digestão dos carboidratos; a digestibilidade dos
carboidratos das paredes celulares é aumentada quando as substâncias fenólicas são liberadas
das plantas forrageiras por tratamento com álcali (HARTLEY e JONES, 1978).
Muitos ácidos benzóicos e cinâmicos servem como precursores de compostos mais
complexos, enquanto alguns deles desempenham papéis reguladores e metabólicos. Apenas
uma fração menor existe como ácidos livres. Ao invés disso, a maioria está ligada por
ligações glicosídicas, éster, éter ou acetal a componentes estruturais de plantas, como a
lignina e celulose, ou a polifenóis maiores, como flavonoides e taninos (WÓJCIAK-KOSIOR
e ONISZCZUK, 2008).
Figura 23. Cromatogramas dos ácidos fenólicos encontrados nos extratos obtidos com acetato
de etila e metanol de Euterpe oleracea (RE e RI); A- amostra RE acetato de etila; B- Amostra
RI acetato de etila; C- Amostra RE metanol; D- Amostra RI metanol.
Identificação dos picos pelo tempo de resposta: 1- ácido protocatecuico; 2- ácido p-
hidroxibenzoico.

43
As análises cromatográficas das amostras do caule de E. oleracea das regiões externa
e interna mostraram cinco picos para a amostra RE extraída e sete picos para a amostra RI
extraída (Figura 24A e B), sendo identificados o ácido protocatecuico em ambas as amotras,
no entanto, apenas a amostra RE extraída foi verificada a presença de ácido p-
hidroxibenzóico. A presença desses ácidos na parede celular corrobora com a análise
espectrométrica de RMN.
O envolvimento de ácidos fenólicos ligados à parede celular no mecanismo de
resistência foi relatado em várias interações hospedeiro-patógeno (BOLWELL, MATERN e
KNEUSEL, 1985; MATERN e KNEUSEL, 1988). O acúmulo desses nas paredes celulares
tem sido frequentemente discutido, não somente como uma parte importante de resposta de
defesa, mas também em relação à lignificação e suberização (BOUDET et al., 1995;
ASCENSAO e DUBERY, 2003).
Lam, Iiyama e Stone (1992) citam que os ácidos fenólicos podem funcionar como
pontos de “ancoragem” para deposição de polímeros de lignina. Ascensao e Dubery (2003)
afirmam que o reforço das paredes celulares com ésteres hidroxicinamoil pode representar
proteção imediata contra fungos e estes podem se conectar a parede celular pela ação de
peroxidases e fornecer uma matriz para posterior lignificação.
Sun, Fang e Bolton (1999) apontaram que 58% de ácido p-hidroxibenzóico foram
esterificados na lignina das paredes celulares do estipe e dos frutos da palmeira Elaeis
guineensis. Kim et al. (1995) relataram uma abundância significativa de ácido p-
hidroxibenzóico éster e éter ligado na lignina de Björkman de Populus maximowiczii Henry
usando a diferença entre os rendimentos de ácido em hidrolisados alcalinos à temperatura
ambiente e a 170ºC.
O aumento na quantidade de ácidos fenólicos ligados às paredes celulares
correlacionou-se com a deposição de lignina em Picea abies. O envolvimento destes ligados à
parede celular no mecanismo de resistência foi relatado para várias outras interações
hospedeiro-patógeno (BOLWELL, ROBINS e DIXON, 1985; MATERN e KNEUSEL,
1988).
Após o tratamento alcalino foi possível observar que os picos dos ácidos
protocatecuico e p-hidroxibenzoico da amostra RE desapareceram (Figura 24C), porém não
foi observada esta perda na amostra RI (Figura 24D). Possivelmente esta perda pode ser
explicada por uma maior quantidade de ligações do tipo éster na região externa do que na
interna, que podem ser confirmados pela razão do sinal de estiramento de C=O nos espectros
de infravermelho, nos quais a região externa apresentou uma maior razão do que a interna.
Os tratamentos alcalinos dissolvem a lignina e liberam ácidos fenólicos das paredes
celulares por clivagem de ligações éster no complexo de lignina. Sabe-se que a lignina torna
os polissacarídeos resistentes à hidrólise ácida ou por enzima. A solubilização da lignina por
meio de produtos alcalinos aumenta a biodegradabilidade dos polissacarídeos da parede
celular (PAN, BOLTON e LEARY, 1998).

44
Figura 24. Cromatogramas dos ácidos fenólicos encontrados no caule de Euterpe oleracea
(RE e RI); A- amostra RE extraído; B - Amostra RI extraído; C - Amostra RE extraído +
NaOH 1%; D - Amostra RI extraído + NaOH 1%.
Identificação dos picos pelo tempo de resposta: 1- ácido protocatecuico; 2- ácido p-
hidroxibenzoico.
4.6 Anatomia quantitativa das fibras caule de Euterpe oleracea
Os resultados das análises das dimensões das fibras de E. oleracea das regiões externa
e interna, como o comprimento, espessura da parede e fração da parede celular estão
apresentadas na Tabela 10.
No estudo do estipe da palmeira, sendo este considerado um material heterogêneo, é
necessário o conhecimento detalhado de sua constituição química e anatômica para subsidiar
sua melhor aplicação. Desta forma, características como o teor de lignina, morfologia das
fibras, a proporção em que os diversos tecidos ocorrem, sua organização e distribuição
exercem papel fundamental na definição de suas propriedades.

45
Tabela 10. Dimensões das fibras da região externa e interna do caule de Euterpe oleracea.
Amostras Variáveis
Comprimento das
fibras (µm)
Diâmetro
total (µm)
Diâmetro
do lúmen
(µm)
Espessura
da parede
(µm)
Fração
parede
(%)
Região externa
Indivíduo
1
Min. 1507,63 35,17 14,67 6,69 15,21
Max. 3394,62 87,88 74,51 18,69 67,11
Média 2616,16 58,18 33,03 12,57 44,33
Desvio
Padrão 489,09 10,62 12,51 3,64 13,16
Indivíduo
2
Min. 1261,84 9,89 8,13 0,47 7,85
Max. 3718,57 122,26 99,93 15,94 39,91
Média 2211,19 55,39 43,18 6,10 19,71
Desvio
Padrão 727,95 41,96 33,52 5,14 8,81
Indivíduo
3
Min. 1778,34 12,33 10,00 0,71 10,08
Max. 4291,92 108,32 94,57 33,08 77,74
Média 2912,43 68,44 43,46 12,49 36,60
Desvio
Padrão 717,79 29,48 26,83 8,20 20,90
Região interna
Indivíduo
1
Min. 959,13 16,02 15,9 1,92 11,12
Max. 3222,38 96,11 77,21 41,96 96,39
Média 2244,98 67,10 35,20 15,95 46,72
Desvio
Padrão 526,56 23,16 22,91 11,37 26,49
Indivíduo
2
Min. 1154,06
10,55 4,25 0,79 15,04
Max. 4007,03 102,45 76,90 27,52 92,83
Média 2289,63 55,04 27,96 13,54 50,53
Desvio
Padrão 704,74 24,37 22,29 7,171 24,90
Indivíduo
3
Min. 1335,22 38,72 8,11 6,18 16,07
Max. 4075,26 99,47 83,49 16,87 80,62
Média 2845,51 66,52 44,63 10,94 36,10
Desvio
Padrão 789,50 18,87 20,73 3,29 16,98
O comprimento, as fibras da região externa e interna apresentaram valores médios de
2570,82 µm e 2460,04 µm, respectivamente e foram classificadas como muito longas de
acordo com as normas da IAWA (1989). O comprimento médio acima de 2 mm da região
externa e interna do caule de E. oleracea, classifica as fibras dessa espécie em uma posição
intermediária entre as fibras curtas de Eucalyptus (média de 1mm) e as longas de Pinus (3,5
mm).
Para Manimekalai, Pavichandran e Balasubramanian (2002) as fibras de maior
comprimento são melhores aproveitadas para a fabricação de papel, pois estão diretamente
relacionadas à resistência tensora e elasticidade. Em todas as variedades de Sorghum bicolor
estudadas por esses pesquisadores, as fibras periféricas apresentaram maior comprimento em
relação à central.

46
Em relação ao diâmetro da fibra foi encontrado um valor médio de 60,67 e 62,89 µm e
do lume de 43,28 e 35,93 µm, para RE e RI, respectivamente. O parâmetro diâmetro do lume
apresenta significância como o comprimento, estando estritamente correlacionado com as
propriedades mecânicas, especialmente tensão, elasticidade e ruptura.
Manimekalai, Pavichandran e Balasubramanian (2002) constataram para cinco
variedades de Sorghum bicolor L., constataram que as fibras periféricas, que possuem lume
menor, apresentaram propriedades mecânicas favoráveis à utilização como suprimento de
matéria-prima em indústrias de celulose.
Os valores médios de espessura de parede das fibras para RE e RI foram de 10,39 e
13,48 µm, respectivamente. Este resultado é interessante, pois é um dos fatores que mais se
relaciona com a resistência da celulose, e conforme Foelkel (1977), fibras com paredes mais
espessas possuem maior teor relativo de celulose que as de paredes mais delgadas.
O espessamento da parede das fibras está relacionado à resistência mecânica, quanto
menor a espessura da parede da fibra maior será o coeficiente de flexibilidade,
consequentemente será menor o grau de colapso em função da maior flacidez
(MANIMEKALAI, PAVICHANDRAN e BALASUBRAMANIAN 2002).
Quando se realizam estudos visando a produção de celulose as características
anatômicas como, comprimento, largura, diâmetro de lume e espessura da parede celular dos
traqueídeos ou fibras são importantes, pois, conhecendo essas dimensões pode-se
correlacionar com as propriedades físico-mecânicas da polpa celulósica e papel (COSTA,
2011).
As amostras RE e RI apresentam s valores médios da fração parede das fibras de 33,55
e 44,45 µm. Estes resultados indicam, percentualmente, a quantidade de celulose,
hemiceluloses e lignina existentes nas paredes das fibras, demonstrando que, quanto mais
espessa é a parede, menos espaços vazios e mais componentes químicos estruturais terão as
fibras (PAULA e SILVA JÚNIOR, 1994).
4.7 Análises histológicas do caule de Eutepe oleracea
Nas seções transversais do caule de E. oleracea foi observado uma estrutura
anatômica padrão das monocotiledôneas, exibindo feixes vasculares distribuídos de maneira
aleatória no tecido fundamental e permitindo verificar uma maior quantidade de feixes
vasculares na região externa (RE) do que na interna (RI), circundados por calotas de fibras
conspícuas.
Tomlinson (1990) relatou que esses feixes fibrovasculares constituídos de grande
proporção de fibras, localizados próximo à epiderme, são os responsáveis pelo aumento da
resistência mecânica do caule. Já que o centro do estipe é altamente lacunoso, apresentando
muitos feixes vasculares separados pelo tecido fundamental.
Foram observados, nas amostras RE e RI, dois tipos de fibras associados aos
elementos condutores do floema, sendo o primeiro mais interno com células com paredes
espessas e lume reduzido e o segundo formado por células com lume mais amplo e paredes
menos espessas, que apresentam uma lignificação gradual.
Tomlinson e Huggett (2012) descreveram esse processo de espessamento e
lignificação das fibras para espécies da família Arecaceae, no qual ocorre a maturação gradual
dessas células, a partir dos elementos condutores do floema, com uma progressão radial, que
finaliza na porção mais externa da calota.

47
Para obter uma indicação na localização e na composição da lignina, as seções
transversais foram submetidas aos testes de Wiesner e de Maüle, respectivamente. O teste de
Wiesner revelou unidades de cinamaldeídos por meio de uma cor avermelhada observada nas
paredes das fibras (Figuras 25A e B; 26A e B), indicando uma maior intensidade de cor na
lamela média das células e ângulos comuns as células. Também foi observada uma variação
de intensidade de cor nas calotas de fibras floemáticas, sendo mais intensa nas paredes das
fibras próximas ao floema, tanto nas amostras RE, quanto nas amostras RI (Figuras 25A e B;
26A e B).
De acordo com Wu, Fukazama e Ohtani (1990) a intensidade da cor pela reação de
Wiesner é causada pela presença de grupos coniferaldeído na lignina guaiacila. Pomar,
Merino e Barceló (2002) afirmam que este teste também pode revelar os grupos terminais p-
hidroxicinamaldeído e p-hidroxi-benzilaldeído 4-O ligado. A presença dessas substâncias na
parede celular corrobora com as análises espectroscópicas (IV e RMN).
Segundo Fukushima e Terashima (1991) a lignina é formada na lamela média e nos
ângulos comuns as demais células, nos estágios iniciais da lignificação. A lignina G é
primeiramente depositada na região da lamela média e depois na parede secundária, enquanto
lignina S é depositada em quantidades menores durante o estágio final da lignificação da
parede secundária.
A formação e a distribuição da lignina, na espécie estudada, ocorreram de forma
semelhante ao descrito pelos autores, reveladas respectivamente, pelo teste de Wiesner e
microscopia de fluorescência. Estes também permitiram observar nas amostras RE e RI fibras
com paredes polilameladas (Figuras 25 e 26).
De acordo com Itoh (1990) e Weiner, Liese e Schmitt (1996), as paredes das fibras de
palmeiras e bambus podem mostrar uma polilamelação resultante de variações no depósito da
lignina em cada camada da parede celular. Segundo estes autores tais fibras permanecem
vivas por um longo período e por isso continua a desenvolver novas lamelas. Donaldson e
Lausberg (1998) afirmam que as diferenças na intensidade da fluorescência também podem
fornecer indícios quanto à distribuição de lignina na fibra, ou seja, a concentração de lignina
reflete no aumento ou na diminuição da intensidade da fluorescência. Em Euterpe oleracea
foram observadas também pequenas variações de intensidade nas fibras.
A localização e a composição da lignina são propriedades importantes dos materiais
lignocelulósicos, porque as ligninas do tipo guaiacila (G) são mais fortemente reticuladas e,
portanto, mais resistentes a degradação química do que as ligninas com alto teor de unidades
siringila (S) (NUOPPONEN et al., 2006). Higuchi (1990) relatou que as características da
macromolécula de lignina podem prevenir a hidrólise de celulose "in situ" por vários
organismos.

48
Figura 25. Seções transversais da região externa do caule de Euterpe oleracea. A, B, C e D –
Testes histoquímicos aplicados em seção transversal das amostras RE; E e F – Microscopia de
fluorescência aplicada em seção transversal das amostras RE. A e B- Detalhe da lignificação
das fibras do floema evidenciado pelo teste de Wiesner. B- Lignina evidenciada pelo teste de
Wiesner na lamela média e nos ângulos das células. C e D - Lignina evidenciada pelo teste de
Wiesner na lamela média e nos ângulos das células. E e F - Fluorescência da lignina,
mostrando processo de lignificação nos cantos das células e na lamela média.
Barras: A, C e E= 200 µm; F= 100 µm; B e D= 50 µm.

49
Figura 26. Seções transversais da região interna do caule de Euterpe oleracea. A, B, C e D –
Testes histoquímicos aplicados em seção transversal das amostras RI; E e F – Microscopia de
fluorescência aplicada em seção transversal das amostras RI. A e B- Detalhe da lignificação
das fibras do floema evidenciado pelo teste de Wiesner. B- Lignina evidenciada pelo teste de
Wiesner na lamela média e nos ângulos das células. C e D - Lignina evidenciada pelo teste de
Wiesner na lamela média e nos ângulos das células. E e F – Fluorescência da lignina,
mostrando processo de lignificação nos cantos das células e na lamela média.
Ac= Ângulo comum às células; Lm= Lamela média.
Barras: A, C e E= 200 µm; F= 100 µm; B e D= 50 µm.

50
5 CONCLUSÕES
A caracterização química da parede celular de amostras do caule de Euterpe oleracea
mostrou que o pré-tratamento com NaOH 1% possibilitou aumento no teor de holocelulose
das regiões externa e interna, e redução no teor de lignina.
O pré-tratamento com NaOH 1% mostrou efeito favorável à hidrólise, concluindo-se
que parte do material incrustante foi solubilizada.
Nas análises realizadas, as amostras RE e RI tratadas com NaOH 1% apresentaram
uma redução de grupos funcionais ligados a lignina e a hemicelulose, indicando que o
processo se mostrou favorável à hidrólise.
Verificou-se que a lignina das amostras RE e RI de E. oleracea é de tipo GS e
incorporada com os ácidos p-hidroxibenzoico e protocatecuico. As análises também
indicaram as principais unidades estruturais presentes, ligação β-O-4, β-O-4’ ϒ-acetiladas,
spiro-dienona, unidade guaiacila, unidade siringila e p-hidroxibenzoato.
Os resultados das análises anatômicas do estipe de Euterpe oleracea, juntamente com
os resultados da caracterização química, indicaram que este material, geralmente
caracterizado com um resíduo após a utilização dos frutos e folhas dessas palmeiras, pode ser
convertido em produtos de maior valor agregado e químicos.

51
6 REFERÊNCIAS BIBLIOGRÁFICAS
ABREU, H. S.; CARVALHO, A. M.; MONTEIRO, M. B. O.; PEREIRA, R. P. W.; SILVA,
H. R.; SOUZA, K. C. A.; AMPARADO, K. F.; CHALITA, D. B. Métodos de análise em
química da madeira, Floresta e Ambiente, Série técnica. 2006, 20p.
ABREU, H. S.; OERTEL, A. C. Estudo químico da lignina de Paullinia rubiginosa. Cerne, v.
5, n. 1, p. 52-60, 1999.
AITA, G. A.; SALVI, D. A.; WALKER, M. S. Enzyme hydrolysis and ethanol fermentation
of dilute ammonia pretreated energy cane. Bioresource Technology, v. 102, n. 6, p. 4444–
4448, 2011.
ALBERTS, B.; JOHNSON, A.; LEWIS, RAFF, M.; ROBERTS, K.; WALTER, P. Molecular
Biology of the Cell, 4th edition, New York: Garland Science; 2002.
ALÉN, R. Structure and chemical composition of wood. In: STENIUS, P. (Editor). Forest
Products Chemistry. Finlândia: Gummerus Printing, 2000. p. 12 – 55.
ANDERSON, A.B.; GELY, A.; STRUDWICK, J.; SOBEL, G.L.; PINTO, M.C. Um sistema
agroflorestal na várzea do estuário amazônico (Ilha das Onças, município de Barcarena,
Estado do Pará). Acta Amazônica. v. 15, n. 1-2, p. 195-224, 1985.
ATUSHI, K.; AZUMA, J-I.; KOSHIJIMA, T. Lignin-carbohydrate complexes and phenolic
acids in bagasse. Holrfornchung, v. 38, n.3, p. 141-149, 1984.
BALAKSHIN, M.; CAPANEMA, E.; GRACZ, H.; CHANG, H.; JAMEEL, H. Quantification
of lignin-carbohydrate linkages with high-resolution NMR spectroscopy, Planta, v. 233, n. 6,
p. 1097-1110, 2011.
BALAKSHIN, M. Y.; CAPANEMA, E. A.; CHANG, H. M. MWL fraction with a high
concentration of lignin-carbohydrate linkages: Isolation and 2D NMR spectroscopic analysis.
Holzforschung, v. 61, n. 1, p. 1–7, 2007.
BARAKAT, A.; WINTER, H.; RONDEAU-MOURO, C.; SAAKE, B.; CHABBERT, B.;
CATHALA, B. Studies of xylan interactions and cross-linking to synthetic lignins formed by
bulk and end-wise polymerization: a model study of lignin carbohydrate complex formation.
Planta, v. 226, n.1, p. 267–281. 2007.
BARROS, C. F.; MIGUENS, F. C. Ultrastructure of the epidermal cells of Beilshmiedia
rigida (Mez) Kosterm. (Lauraceae). Acta Microscopica, v. 6, n. 3, p. 451-461, 1998.
BHAT, K. M.; LIESE, W.; SCHMITT, U. Structural variability of vascular bundles and cell
wall in rattan stem. Wood Science and Technology, v.24, n.3, p. 24, p. 211–224, 1990.

52
BOERIU, C.; BRAVO, D.; GOSSELINK, R. J. A., DAM, G. J. E .G. V. Characterisation of
structure-dependent functional properties of lignin with infrared spectroscopy. Industrial
Crops and Products, v. 20, n.2, p. 205-218, 2004.
BOLWELL, G. P.; ROBINS, M. P.; DIXON, R. A. Metabolic changes in elicitor treated bean
cells. Enzymic responses associated with rapid changes in cell wall components. European
Journal of Biochemistry, v. 148, n.3, p. 571–578, 1985.
BOUDET, A-M. Towards an understanding of the supramolecular organization of the
lignified wall. In: The Plant Cell Wall. Oxford: Blackwell Publishing, 2003. p. 155 – 182.
BUKATSCH, F. Bemerkungen zur Doppelfarbung: Astrablau-Safranin. Mikrokosmos, v. 61,
p. 255, 1972.
BURANOV, A. U.; MAZZA, G. Lignin in straw of herbaceous crops. Industrial Crops and
Products, v. 28, n. 3, p. 237–259, 2008.
BYKOV, I. Characterization of Natural and Technical Lignins using FTIR
Spectroscopy. 2008. 43 f. Tese (Doutorado) - Curso de Chemical Technology, Department
Of Chemical Engineering And Geosciences, Luleå University Of Technology, Luleå, 2008.
CALZAVARA, B. B. G. As possibilidades do açaizeiro no estuário amazônico. Boletim da
Faculdade de Ciências Agrárias do Pará - FCAP. Belém, (5): 103p. 1972.
CARPITA, N.; McCANN, M. In Biochemistry & Molecular Biology of Plants Ch. 2 (eds
Buchanan, B., Gruissem, W. & Jones, R. L.) John Wiley & Sons, New Jersey, 2002.
CARPITA, N. C. Structure and biogenesis of the cell walls of grasses. Annual review of
plant physiology and plant molecular biology, v. 47, n.1, p. 445-476, 1996.
CARPITA, N. C.; GIBEAUT, D. M. Structural models of primary cell walls in flowering
plants: consistency of molecular structure with the physical properties of the walls during
growth. The Plant Journal, v. 3, n. 1, p. 1-30, 1993.
CARRILLO, F.; LIS, M. J.; COLOM, X.; LÓPEZ-MESASA, M.; VALLDEPERAS, J. Effect
of alkali pretreatment on cellulase hydrolyiss of wheat straw: Kinetic study. Process
Biochemistry, v. 40, n 10, p. 3360–3364, 2005.
CAVALCANTE, P. Frutas comestíveis da Amazônia. Belém: CEJUP, 1991. 271p. 1991.
CESARINO, I.; ARAÚJO, P.; DOMINGUES JÚNIOR, A. P.; MAZZAFERA, P. An
overview of lignin metabolism and its effect on biomass recalcitrance. Brazilian Journal of
Botany, v. 35, n. 4, p. 303-311, 2012.
CHEN, F.; TOBIMATSU, Y.; HAVKIN-FRENKEL, D.; DIXON, R. A.; RALPH, J. A
polymer of caffeyl alcohol in plant seeds. Proceedings of the National Academy of Sciences
of the United States of America, v. 109, n. 5, p. 1772-1777, 2012.

53
CHIANG, V. L. Understanding gene function and control in lignin formation in wood.
Agricultural Biotechnology, v. 17, p. 139-144, 2005.
CHOI, J. W.; CHOI, D. H.; FAIX, O. Characterization of lignin–carbohydrate linkages in the
residual lignins isolated from chemical pulps of spruce (Picea abies) and beech wood (Fagus
sylvatica). Journal of Wood Science, v. 53, n. 4, p. 309-313, 2007.
CHUNDAWAT, S. P. S.; BECKHAM, G. T.; HIMMEL, M. E.; DALE, B. E. Deconstruction
of lignocellulosic biomass to fuels and chemicals. Annual Review of Chemical and
Biomolecular Engineering, v. 2, n.1, p. 121–145, 2011.
CORNU, A.; BESLE, J. M.; MOSONI, P.; GRENET, E. Lignin-carbohydrate complexes in
forages: structure and consequences in the ruminal degradation of cell-wall carbohydrates.
Reproduction Nutrition Development Journal, v. 34, n.5, p. 385-398, 1994.
COSTA, J de A. Qualidade da madeira de Eucalyptus urograndis, plantado no Distrito
Federal, para produção de celulose Kraft. 2011. 86 f. Dissertação (Mestrado) - Curso de
Engenharia Florestal, Universidade de Brasília, Brasília, 2011.
CRESTINI, C.; ARGYROPOULOS, D. S. Structural analysis of wheat straw lignin by
quantitative 31
P and 2D NMR spectroscopy. The occurrence of ester bonds and α-O-4
substructures. Journal of Agricultural and Food Chemistry, v. 45, n. 4, p. 1212–1219,
1997.
DE ASCENSAO, A. R. F. D. C.; DUBERY, I. A. Soluble and wall-bound phenolics and
phenolic polymers in Musa acuminata roots exposed to elicitors from Fusarium oxysporum
f.sp. cubense. Phytochemistry, v.63, n.6, p.679–686, 2003.
DEL RÍO, J. C.; RENCORET, J.; MARQUES, G.; LI, J.; GELLERSTEDT, G.; JIMÉNEZ-
BARBERO, J.; MARTÍNEZ, A. T.; GUTIÉRREZ, A. Structural Characterization of the
Lignin from Jute (Corchorus capsularis) Fibers. Journal of Agricultural and Food
Chemistry, v. 57, n. 21, p. 10271-10281, 2009.
DEL RIO, J. C.; RENCORET, J.; MARQUES, G.; GUTIERREZ, A.; IBARRA, D.;
SANTOS, J. I.; JIMENEZ-BARBERO, J.; ZHANG, L. M.; MARTINEZ, A. T. Highly
acylated (acetylated and/or p-coumaroylated) native lignins from diverse herbaceous plants.
Journal of Agricultural and Food Chemistry, v. 56, n. 20, p. 9525–9534, 2008.
DEL RIO, J. C.; GUTIÉRREZ, A.; RODRÍGUEZ, I. M.; IBARRA, D.; MARTÍNEZ, A. T.
Composition of non-woody plant lignins and cinnamic acids by Py-GC/MS, Py/TMAH and
FT-IR. Journal Analytical Applied Pyrolysis, v. 79, p. 39-46, 2007a.
DEL RÍO, J. C.; MARQUES, G.; RENCORET, J.; MARTÍNEZ, A. T.; GUTIÉRREZ, A.
Occurrence of naturally acetylated lignin units. Journal of Agricultural and Food
Chemistry, v. 55, n. 14, p. 5461-5468, 2007b.

54
DERROUICHE, I.; MARZOUG, I. B.; SAKLI, F.; ROUDESLI, S. Study of Extraction and
Characterization of Ultimate Date Palm Fibers. Advances in Materials, v. 4 n. (5-1), p. 7-14,
2015.
DIAZ, M. J.; GARCIA, M. M.; EUGENIO, M. M.; TAPIAS, R.; FERNANDEZ, M.; LOPEZ,
F. Variations in fiber length and some pulp chemical properties of leucaena varieties.
Industrial Crops and Products, v. 26, n. 2, p. 142-150, 2007.
DIXIT, S.; NAIR, S. A Review on Delineating the Distribution of p-Coumarate vs. p-
Hydroxybenzoate as Acylating Groups in Lignin across Monocot Families. Lignocellulose, v.
3, n. 2, p. 131-144, 2014.
DONALDSON, L. A.; LAUSBERG, M. J. F. Comparison of conventional transmitted light
and confocal microscopy for measuring wood cell dimensions by image analysis. IAWA
Journal, v. 19, n 3, p. 321-336, 1998.
DRANSFIELD, J.; UHL, N. W.; ASMUSSEN, C. B.; BAKER, W. J.; HARLEY, M . M.;
AND LEWIS C. E. Genera palmarum: The evolution and classification of palms. Royal
Botanic Gardens, Kew, UK. 2008.
DUROT, N.; GAUDARD, F.; KUREK, B. The unmasking of lignin structures in wheat straw
by alkali. Phytochemistry, v. 63, p. 617–623, 2003.
EBRINGEROVÁ, A. Structural diversity and application potential of hemicelluloses.
Macromolecular Symposia, v. 232, n. 1, p. 1–12, 2005.
EFFLAND, M. J. Modified procedure to determine acid-insoluble lignin in wood and pulp.
Tappi, v. 60, n. 10, 143-144, 1977.
EVERT, R. F.; ESAU, K. Anatomia das plantas de Esau, meristemas, células e tecidos do
corpo da planta: sua estrutura, função e desenvolvimento. Tradução da Terceira Edição
americana, Editora Edgard Blücher. 2013, 728 p.
FAIX, O.; GRUNWALD, C.; BEINHOFF, O. Determination of phenolic hydroxyl group
content of milled wood lignins (MWL’s) from different botanical origins using selective
aminolysis, FTIR, 1H NMR and UV spectroscopy. Holzforschung, v. 46, n. 5, p. 425-532,
1992.
FAIX, O. Classification of Lignins from Different Botanical Origins by FT-IR Spectroscopy.
Holzforschung,v. 45, p.21-27, 1991.
FOELKEL, C. E. B. Estrutura da madeira. 85p. 1977.
FRANKLIN, G. L. Preparation of thin sections of synthetic resin and wood-resin composites,
and a new macerating method for wood. Nature, v. 155, p. 51, 1945.

55
FROLLINI, E.; PAIVA, J. M. F.; TRINDADE, W. G.; RAZERA, A. T.; TITA, S. P.;
Lignophenolic and phenolic resins as matrix in vegetal fibers reinforced composites. In:
Wallenberger FT, Weston N, editors. Natural fibers, plastics and composites. New York:
Kluwer Academic Publishers; p. 193–219, 2004.
FUKUSHIMA, K.; TERASHIMA, N. Heterogeneity in formation of lignin XIV. Formation
and structure of lignin in differentiating xylem of Ginkgo biloba. Holzforschung, v. 45, p.
87-94, 1991.
GARCÍA HORTAL, J. A. Fibras papeleras. Barcelona: UPC, 243 p. 2007.
GEETHA, N. P.; MAHESH, M.; BETTADAIAH, B. K.; KINI, R. K.; PRAKASH, H. S.
HPLC Method for Determination of p-coumaric acid from the Medicinal Herb Leptadinia
reticulate. International Journal of Phytomedicine, v. 3, p. 319-324, 2011.
GIERER, J.; WANNSTRÖM, S. The formation of ether bonds between lignins and
carbohydrates during alkaline pulping processes. Holzforschung, v. 40, p. 347–352, 1986.
GIERER, J. Chemistry of delignification, Part 1: General concept and reactions during
pulping.Wood Science and Technology, v. 19, n. 4, p. 289–312, 1985.
GOUVÊA, D. D. S. Caracterização anatômica e ultraestrutural de sementes de Euterpe
oleracea Mart. 2007, 64p. Dissertação (Mestrado em Agronomia), Universidade Federal
Rural da Amazônia – UFRA, Belém – AM, 2007.
GLÓRIA, B. A.; GUERREIRO, S. M. C. Anatomia Vegetal. Viçosa: UFV, 2003. 438 p.
GOVAERTS, R.; DRANSFIELD, J. World checklist of palms. Royal Botanic Gardens, Kew.
UK. 2005.
GRABBER, J. H.; RALPH, J.; HATFIELD, R. D. Cross-linking of maize walls by ferulate
dimerization and incorporation into lignin. Journal of Agricultural and Food Chemistry, v.
48, p. 6106 – 6113, 2000.
GRITSCH, C. S.; MURPHY, R. J. Ultrastructure of fiber and parenchyma cell walls during
early stages of culm development in Dendrocalamus asper. Annals of Botany, v. 95, n. 4, p.
619–629, 2005.
GUILLEMIN, F.; GUILLON, F.; BONNIN, E.; DEVAUX, M. F.; CHEVALIER, T.; KNOX,
J. P.; LINERS, F.; THIBAULT, J. F. Distribution of pectic epitopes in cell walls of the sugar
beet root. Planta, v. 222, p. 355–371, 2005.
GWON, J. G.; LEE, S. Y.; DOH, G. H.; KIM, J. H. Characterization of Chemically Modified
Wood Fibers Using FTIR Spectroscopy for Biocomposites. Journal of Applied Polymer
Science, v. 116, p. 3212–3219, 2010.

56
HALL, D. O.; SCURLOCK, J. M. O.; Bolhàr-Nordenkampf, H. R.; Leegood, R. C. in
“ hotosynthesis and production in a changing environment”. A field and laboratory manual.
London: Chapman & Hall, 1989,110.
HARTLEY, R. D.; MORRISON, W. H. III; HIMMELSBACH, D. S.; BORNEMAN, W. S.;
Cross-linking of cell wall phenolic arabinoxylans in graminaceous plants. Phytochemistry, v.
29, p. 3705 – 3709, 1990.
HATFIELD, R. D. Cell-wall polysaccharide interactions and degradability. In: Jung, H.G.,
Buxton, D.R., Hatfield, R.D., Ralph, J. (Eds.), Forage cell-wall structure and digestibility.
ASA, CSSA, and SSSA, Madison, WI, pp. 285 – 313. 1993.
HE, Y.; PANG, Y.; LIU, Y.; LI, X.; WANG, K. Physicochemical characterization of rice
straw pretreated with sodium hydroxide in the solid state for enhancing biogas production.
Energy Fuels, v. 22, n. 4, p. 2775–2781, 2008.
HENDERSON, A.; GALEANO, G.; BERNAL, R. Field guide to the palms of the Americas.
Princeton University Press, Princeton, New Jersey, USA. 1995.
HIBINO, T.; SHIBATA, D.; ITO, T.; TSUCHIYA, D.; HIGUCHI, T.; POLLET, B.;
LAPIERRE, C. Chemical properties of lignin from Aralia cordata. Phytochemistry, v. 37, n.
2, p. 445–448, 1994.
HIGUCHI, T.; ITO, T.; UMEZAWA, T.; HIBINO, T.; SHIBATA, D. Red-brown color of
lignified tissues of transgenic plants with antisense CAD gene: Wine-red lignin from coniferyl
aldehyde. Journal of Biotechnology, v. 37, p. 151-158, 1994.
HIMMEL, M. E.; DING, S. Y.; JOHNSON, D. K.; ADNEY, W. S.; NIMLOS, M. R.;
BRADY, J. W.; FOUST, T. D. Biomass recalcitrance: engineering plants and enzymes for
biofuels production. Science, v. 315, p. 804–807, 2007.
HORN, J. W.; FISHER, J. B.; TOMLINSON, P. B.; LEWIS, C. E.; LAUBENGAYER, K.
Evolution of lamina anatomy in the palm family (Arecaceae). American Journal of Botany,
v. 96, n. 8, p. 1462–1486, 2009.
HSU, T. A. In Handbook on bioethanol, production and utilization: Pretreatment of biomass.
Washington DC: Taylor and Francis: Edited by Wyman, CE; 1996, p. 179-212.
IAWA COMMITTEE. List of microscopic features for hardwood identification. Iawa
Bulletin, Leiden, v.10, p.219–332, 1989.
IIYAMA, K.; LAM, T. B. T.; STONE, B. A. Phenolic acid bridges between polysaccharides
and lignin in wheat internodes. Phytochemistry, v. 29, p. 733–737, 1990.
ISMAIL, A.; MAMAT, M. N. The optimal age of oil palm replanting. Oil Palm Industry
Economic Journal, v. 2, n. 1, p. 11-18, 2002.

57
ITOH, T. Lignification of bamboo (Phyllostachys heterocycla Mitf.) during its growth.
Holzforschung, v. 44, p. 191–200, 1990.
IVERSON, T.; WANNSTRÖM, S. Lignin–carbohydrate bonds in a residual lignin isolated
from pine kraft pulp. Holzforschung, v. 40, p. 19–22, 1986.
JARDIM, M. A. G. Possibilidade do cultivo do açaizeiro (Euterpe oleracea Mart.) em
áreas de capoeira como alternativa para agricultores do nordeste do Nordeste Paraense.
Museu Paraense Emílio Goeldi/Conselho Nacional de Desenvolvimento Científico e
Tecnológico. Relatório Técnico-Científico, 35 p. 2005.
JIN, Z.; JIN, G.; SHAO, S. Lignin characteristics of bast fiber and core in kenaf, bark and
wood of paper mulberry and mulberry. Journal of Wood Science, v. 58, p. 144-152, 2012.
JOHANSEN, D. A. Plant microtechnique. Mc Graw Hill, New York. 1940.
JONES, D. L. Palms throughout the world. Washington: Smithsonian Institution, 1995. 410p.
JUNG, H. G.; DEETZ, D. A. Cell-wall lignification and degradability. In: Forage cell-wall
structure and digestibility (HJ Jung et al, eds) Am Soc Agron, Madison, WI, USA, p. 315-
346, 1993.
JUNG, H. G.; FAHEY Jr, G. C.; GARST J. E. Simple phenolic monomers of forages and
effects of in vitro fermentation on cell wall phenolics. Journal of animal Science, v. 57, p.
1294-1305, 1983.
KALIMA, T.; JASNI. Study of Calamus occidentalis J. R. Witono; J. Dransf. species
commercial values and possible utilization. Biodiversitas, v. 5, n. 2, p. 61-65, 2004.
KARNAOURI, A.; ROVA, U. CHRISTAKOPOULOS, P. Effect of Different Pretreatment
Methods on Birch Outer Bark: New Biorefinery Routes. Molecules, v., 21, p. 427-443; 2016.
KEENER, T. J.; STUART, R. K.; BROWN, T. K. Maleated couping agents for natural fibre
composites. Composites Part A: Applied Science and Manufacturing, Kidlington, v. 35, n. 3;
p. 357-362, 2004.
KIM, H.; RALPH, J. Solution-state 2D NMR of ball-milled plant cell wall gels in DMSO-
d(6)/pyridine-d(5), Organic & Biomolecular Chemistry, v. 8, n. 3, 576-591, 2010.
KIM, T. H.; KIM, J. S.; SUNWOO, C.; LEE, Y. Y. Pretreatment of corn stover by aqueous
ammonia. Bioresource Technology, v. 90, n. 1, p. 39-47, 2003.
KIM, Y.-S.; IIYAMA, K.; KURAHASHI, A.; MESHITSUKA, G. Structural feature of lignin
in cell walls of normal and fast-growing poplar (Populus maximowiczii Henry) linkage types
of wall-bound hydroxybenzoic and hydroxy-cinnamic acids. Mokuzai Gakkaishi, v. 41, p.
837-843, 1995.

58
KNOX, J. P. Revealing the structural and functional diversity of plant cell walls. Current
Opinion in Plant Biology, v. 11, p. 308–313, 2008.
KOULLAS, D. P.; CHRISTAKOPOULOS, P.; KEKOS, D.; MACRIS, B. J.; KOUKIOS,
E.G. Correlating the effect of pretreatment on the enzymatic hydrolysis of straw.
Biotechnology and Bioengineering, v. 39, p. 113–116, 1992.
KUBO, J. H.; KADLA, J. F. Hydrogen bonding in lignin: a Fourier transform infrared model
compound study. Biomacromolecules, v. 6, p. 2815-2821, 2005.
KURODA, K-I.; OZAWA, T.; UENO, T. Characterization of sago palm (Metroxylon sagu
Rottb.) lignin by analytical pyrolysis. Journal of Agricultural and Food Chemistry, v. 49,
p. 1840–1847, 2001.
LAM, T. B. T.; KADOYA, K.; IIYAMA, K. Bonding of hydroxycinnamic acids to lignin:
ferulic and p-coumaric acids are predominantly linked at the benzyl position of lignin, not the
β-position, in grass cell walls. Phytochemistry, v. 57, p. 987–992, 2001.
LAM, T. B. T.; IIYAMA, K.; STONE, B. A. Cinnamic acid bridges between cell wall
polymers in wheat and phalaris internodes. Phytochemistry, 31, p. 1179-1183, 1992.
LANDUCCI, L. L.; DEKA, G. C.; ROY, D. N. A 13
C NMR study of milled wood lignins
from hybrid Salix clones. Holzforschung, v. 46, n. 6, p. 505–511, 1992.
LAPIERRE, C.; LALLEMAND, J. Y.; MONTIES, B. Evidence of poplar lignin
heterogeneity by combination of 13
C and 1H NMR spectroscopy. Holzforschung, v. 36, n. 6,
p. 275-282, 1982.
LATIF, M. H. A.; ATTIYA, H. G.; AL-ABAYAJI, M. A.; ALSOUZ, M. A. K. Isolation and
Characterization of Lignin from Iraqi Date Palm Phoenix Dactylifera L. Frond Bases as a
Good and Cheap Source of Lignin. Journal of Natural Sciences Research, v.5, n.4, p. 83-
90, 2015.
LAWOKO, M.; HENRIKSSON, G.; GELLERSTEDT, G. Structural differences between the
lignin-carbohydrate complexes presents in wood and chemical pulps. Biomacromolecules,
Washington, DC, v. 6, p. 3467-3473, 2005.
LIESE, W. Research on bamboo. Wood Science and Technology, v. 21, p. 189–209, 1987.
LIN, S. Y.; DENCE, C. W. Methods in lignin chemistry. Berlim: Spring-Verlag, 1992, 568
p.
LOPEZ, F.; GARCIA, M. M.; YANEZ, R.; TAPIAS, R.; FERNANDEZ, M.; DIAZ, M. J.
Leucaena species valoration for biomass and paper production in 1 and 2 year harvest,
Bioresource Technology, v. 99, p. 4846-4853, 2008.

59
LU, F.; KARLEN, S. D.; REGNER, M.; KIM, H.; RALPH, S. A.; SUN, R-C.; KURODA, K-
I.; AUGUSTIN, M. A.; MAWSON, R.; SABAREZ, H.; SINGH, T.; JIMENEZ-MONTEON,
G.; ZAKARIA, S.; HILL, S.; HARRIS, P. J.; BOERJAN, W.; WILKERSON, C. G.;
MANSFIELD, S. D.; RALPH, J. Naturally p-Hydroxybenzoylated Lignins in Palms.
BioEnergy Research, v. 8, p. 934–952, 2015.
LU, F.; RALPH, J. Novel tetrahydrofuran structures derived from β–β-coupling reactions
involving sinapyl acetate in Kenaf lignins. Organic and Biomolecular Chemistry, v. 6, n.
20, p. 3681–3694, 2008.
LU, F.; RALPH, J.; MORREEL, K.; MESSENS, E.; BOERJAN, W. Preparation and
relevance of a cross-coupling product between sinapyl alcohol and sinapyl p-hydroxy
benzoate. Organic & biomolecular chemistry, v. 2, p. 2888 – 2890, 2004.
LÜDEMANN, H. D.; NIMZ, H. H. Carbon-13 nuclear magnetic resonance spectra of lignins.
Biochemical and Biophysical Research Communications, v. 52, n. 4, p. 1162–1973, 1973.
MANCERA, A.; FIERRO, A.; DUMARÇAY, S.; GÉRARDIN, P.; VELÁSQUEZ, J.;
QUINTANA, G.; CELZARD, A. Physicochemical characterisation of sugar cane bagasse
lignin oxidized by hydrogen peroxide. Polymer degradation and stability, v. 35, p. 470-476,
2010.
MANIMEKALAI, V.; PAVICHANDRAN, P.; BALASUBRAMANIAN, A. Fibres of
Sorghum bicolor (L.) Moench and their potential use in paper and board making.
Phitomorphology, v. 52, n. 1, p. 55-59, 2002.
MANSFIELD, S. D.; KIM, H.; LU, F.; RALPH, J. Whole plant cell wall characterization
using solution-state 2D NMR. Nature Protocols, v. 7, n. 9, p. 1579-1589, 2012.
MARCHESSAULT, R. H.; COULOMBE, S.; MORIKAWA, H.; ROBERT, D.
Characterization of aspen exploded lignin. Canadian Journal of Chemistry, v. 60, n. 18, p.
2372-2382, 1982.
MARINHO, N. P.; NISGOSKI, S.; KLOCK, U.; ANDRADE, A. S.; MUNIZ, G. I. B. Análise
química do bambu-gigante (Dendrocalamus giganteus Wall. Ex Munro) em diferentes idades.
Revista Ciência Florestal, v. 22, n. 2, p. 417-422, 2012.
MARTINEZ, A. T.; RENCORET, J.; MARQUES, G.; GUTIERREZ, A.; IBARRA, D.;
JIMENEZ-BARBERO, J.; DEL RIO, J. C. Monolignol acylation and lignin structure in some
nonwoody plants: a 2D NMR study. Phytochemistry, v. 69, n. 16, p. 2831–2843, 2008.
MARTINS, M. A.; FORATO, L. A.; MATTOSO, L. H. C.; COLNAGO, L. A. A solid state 13
C high resolution NMR study of raw and chemically treated sisal fibers, Carbohydrate
Polymers, v. 64, p. 127-133, 2006.
MATERN, U.; KNEUSEL,R. E. Phenolic compounds in plant disease resistance,
Phytoparasitica, v. 16, p. 153–170, 1988.

60
McCANN, M. C.; ROBERTS, K. Architecture of the primary cell wall. In: The Cytoskeletal
Basis of Plant Growth and Form (C. W. Lloyd, ed.). London: Academic Press, p. 109-129.
1991.
MEYERMANS, H.; MORREEL, K.; LAPIERRE, C.; POLLET, B.; DE BRUYN, A.;
BUSSON, R.; HERDEWIJN, P.; DEVREESE, B.; VAN BEEUMEN, J.; MARITA, J. M.;
RALPH, J.; CHEN, C.; BURGGRAEVE, B.; VAN MONTAGU, M.; MESSENS, E.;
BOERJAN, W. Modification in lignin and accumulation of phenolic glucosides in poplar
xylem upon downregulation of caffeoyl-coenzyme A O-methyltransferase, an enzyme
involved in lignin biosynthesis. Journal Of Biological Chemistry, v. 275, n. 47, p. 36899-
36909, 2000.
MIRANDA, I.P.A.; RABELO, A.; BUENO, C.R.; BARBOSA, E.M.; RIBEIRO, M.N.S.
Frutos de Palmeiras da Amazônia. Manaus: MCT/INPA, 2001. 119p.
MIYAKOSHI, T.; CHEN, C. L. 13
C NMR spectroscopic studies of phenylcoumaran and 1,2-
diarylpropane type lignin model compounds. Part 1. Synthesis of model compounds.
Holforschung, v. 45, Suppl. 1, p. 41–48, 1991.
MONTEIRO, M. B. O.; PEREIRA, R. P. W.; ABREU, H. S. Bioquímica da lignificação de
células xilemáticas. Floresta e Ambiente, v. 11, p. 48 - 57, 2004.
MORAES, S. A. L.; NASCIMENTO, E. A.; MELO, D. C. Análise da madeira de Pinus
oocarpa Parte I: estudo dos constituintes macromoleculares e extrativos voláteis. Revista
Árvore, v. 29, n. 3, p. 461-470, 2005.
MORAIS, S. A. L. Contribuição ao estudo químico e espectroscópico da lignina de
madeira moída do Eucalyptus grandis: Isolamento, quantificação e análise estrutural. Belo Horizonte, 1992. 260p. Tese (Doutorado em Química) - UFMG/ICEx, 1992.
MORREEL, K.; RALPH, J.; KIM, H.; LU, F.; GOEMINNE, G.; RALPH, S. A.; MESSENS,
E.; BOERJAN, W. Profiling of oligolignols reveals monolignol coupling conditions in
lignifying poplar xylem. Plant Physiology, v. 136, n. 3, p. 3537–3549, 2004.
MORRISON, T. A.; JUNG, H. G.; BUXTON, D. R.; HATFIELD, R. D. Cell wall
composition of maize internodes of vary maturity. Crop Science, v. 38, n. 2, p. 455 – 460,
1998.
MURATA, Y.; KUBO, S.; TOGAWA, E.; RAMLE, S. F. B. M.; SULAIMAN, O.; HASHIM,
R.; IBRAHIM, W. A.; KOSUGI, A.; HIROOKA, A.; ABE, H. Detection of vascular bundles
using cell wall birefringence on exposure to polarized light. Industrial Crops and Products,
v. 65, p. 190–197, 2015.
NIMZ, H. H.; ROBERT, D.; FAIX, O.; NEMR, M. Carbon-13 NMR spectra of lignins. 8:
Structural differences between lignins of hardwood, softwoods, grasses and compression
wood. Holzforschung, v. 35, p. 16–26, 1981.

61
NIMZ, H. H.; TSCHIRNER, U.; STÄHLE, M.; LEHMANN, R.; SCHLOSSER, M. Carbon-
13 NMR spectra of lignins, 10. Comparison of structural units in spruce and beech lignin.
Journal of Wood Chemistry and Technology, v. 4, n. 3, p. 265–284, 1984.
NUOPPONEN, M. H.; WIKBERG, H. I.; BIRCH, G. M.; JÄÄSKELÄINEN, A. S.;
MAUNU, S. L.; VUORINEN, T.; STEWART, D. Characterization of 25 tropical hardwood
with Fourier transform infrared, ultraviolet resonance Raman, and 13C-NMR cross-
polarization/magic-angle spinning spectroscopy. Journal of Applied Polymer Science, v.
102, p. 810–819, 2006.
OLIVEIRA, F. C. Oxidação de lignina proveniente de resíduos lignocelulósicos
agroindustriais para obtenção de compostos químicos aromáticos de maior valor
agregado. 2015. 199 f. Tese (Doutorado)- Faculdade de Engenharia Química de Lorena,
Lorena, São Paulo, 2015.
PAN, X. G. L.; BOLTON, J. J.; LEARY, G. Determination of ferulic and p-coumaric acids in
wheat straw and the amounts released by mild acid and alkaline peroxide treatment. Journal
of Agricultural and Food Chemistry, v. 46, p. 5283–5288, 1998.
PARAMESWARAN, N.; LIESE, W. On the fine structure of bamboo. Wood Science and
Technology, v. 10, p. 231–246, 1976.
PATTATHIL, S.; HAHN, M. G.; DALE, B. E.;CHUNDAWAT, S. P. S. Insights into plant
cell wall structure, architecture, and integrity using glycome profiling of native and AFEXTM
-
pre-treated biomass. Journal of Experimental Botany, v. 66, n. 14, p. 4279-4294, 2015.
PAULA, L. E. R.; TRUGILHO, P. F.; NAPOLI, A.; BIANCHI, M. L. Characterization of
residues from plant biomass for use in energy generation. Cerne, v. 17, n. 2, p. 237-246,
2011.
PAULA, J. E.; SILVA JÚNIOR, F. G. Anatomia de madeiras indígenas com vistas à
produção de energia e papel. Pesquisa Agropecuária Brasileira, v. 19, n. 12, p. 1807-1821,
1994.
PEREIRA, J. C. D.; STURION, J. A.; HIGA, A. R.; HIGA, R. C. V.; SHIMIZU, J. Y.
Características da madeira de algumas espécies de eucalipto plantadas no Brasil. Colombo:
EMBRAPA Florestas, (Comunicado Técnico, n. 38). p. 55, 2000.
PETRIK, D.; KARLEN, S. D.; CASS, C.; PADMAKSHAN, D.; LU, F.; LIU, S.; LE BRIS,
P.; ANTELME, S.; SANTORO, N.; WILKERSON, C. G.; SIBOUT, R.; LAPIERRE, C.;
RALPH, J.; SEDBROOK, J. C. p-Coumaroyl-CoA:monolignol transferase (PMT) acts
specifically in the lignin biosynthetic pathway in Brachypodium distachyon. The Plant
Journal, v. 77, n. 5, p. 713–726, 2014.
POMAR F.; MERINO F.; ROS BARCELÓ, A. O-4-Linked coniferyl and sinapyl aldehydes
in lignifying cell walls are the main targets of the Wiesner (phloroglucinol-HCl) reaction.
Protoplasma, v. 220, p. 17–28, 2002.

62
QUEIROZ, J. A. L. de.; MOCHIUTTI, S. Tipos de manejo de açaizais e seu efeito sobre a
diversidade florestal no Estuário Amazônico. Anais... Belém: SIMPÓSIO
INTERNACIONAL DA IUFRO, 2000. Livro do Simpósio. IUFRO, p.344-350, 2002.
QUEIROZ, J. A. L.; MELÉM JUNIOR, N. J. Efeito do tamanho do recipiente sobre o
desenvolvimento de mudas do açaí (Euterpe oleracea Mart.). Revista Brasileira de
Fruticultura, v. 23, p. 460–462, 2001.
RALPH, J. Hydro ycinnamates in lignification. Phytochemistry Reviews, v. 9, p. 65–83,
2010.
RALPH, J.; LANDUCCI, L. L. NMR of Lignins. In: Heitner C, Di mmel DR, Schmidt JA
(eds) Lignin and lignans: advances in chemistry. CRC Press (Taylor & Francis Group), Boca
Raton, p. 137–234, 2010.
RALPH, J.; HATFIELD, R. D.; GRABBER, J. H.; JUNG, H. J. G. Cell wall cross-linking in
grasses by ferulates and diferulates. Journal of Agricultural and Food Chemistry, v. 79, p.
403–407, 1998.
RALPH, J.; LU, F. The DFRC method for lignin analysis. Part 6. A modified method to
determine acetate regiochemistry on native and isolated lignins. Journal of Agricultural and
Food Chemistry, v. 46, 11, p. 4616–4619, 1998.
RALPH, J. An unusual lignin from kenaf. Journal of Natural Products, v. 59, n. 4, p. 341–
342, 1996.
RALPH, J.; GRABBER, J. H.; HATFIELD, R. D. Lignin-ferulate crosslinks in grasses: active
incorporation of ferulate polysaccharide esters into ryegrass lignins. Carbohydrate
Research. v. 275, p. 167-178, 1995.
RALPH, J.; HATFIELD, R. D.; QUIDEAU, S.; HELM, R. F.; GRABBER, J. H.; JUNG, H. J.
Pathway of p-coumaric acid incorporation into maize lignin as revealed by NMR. Journal of
the American Chemical Society, v. 116, p. 9448 – 9456, 1994a.
RALPH, J.; QUIDEAU, S.; GRABBER, J. H.; HATFIELD, R. D. Identification and synthesis
of new ferulic acid dehydrodimers present in grass cell walls. Journal of the Chemical
Society, Perkin Transactions 1, p. 3485 – 3498, 1994b.
RAMIRES, E. C.; MEGIATTO, J. D.; GARDRAT, C.; CASTELLAN, A.; FROLLINI, E.
Valorization of an industrial organosolv-sugarcane bagasse lignin: characterization and use as
a matrix in biobased composites reinforced with sisal fibers. Biotechnology and
Bioengineering, v. 107, n. 4, p. 612-621, 2010.
RAVEN, P. H.; EVERT, R. F.; EICHHORN, S. E. Biology of plants. New York: Worth
Publ., 2001. 791 p.

63
REITER, W.D.; VANZIN, G.F. Molecular genetics of nucleotide sugar interconversion
pathways in plants. Plant Molecular Biology, New York, v. 47, n.1/2, p. 95-113, 2001.
RENCORET, J.; RALPH, J.; MARQUES, G.; GUTIÉRREZ, A.; MARTÍNEZ, Á. T.; DEL
RIO, J. C. Structural characte rization of the lignin from coconut (Cocos nucifera) coir fibers.
Journal of Agricultural and Food Chemistry, v. 61, n. 10, p. 2434 –2445, 2013.
REZENDE, C. A.; LIMA, M. A.; MAZIERO, P.; AZEVEDO, E. R.; GARCIA, W.;
POLIKARPOV, I. Chemical and morphological characterization of sugarcane bagasse
submitted to a delignification process for enhanced enzymatic digestibility. Biotechnology
for Biofuels, v. 4, p. 54, 2011.
RIVAS, M.; BARBIERI, R. L.; MAIA, L. C. Plant breeding and in situ utilization of palm
trees. Ciência Rural, v. 42, n. 2, p. 261-269, 2012.
RODRIGUES, J. et al. Determination of tree to tree variation in syringyl:guaiacyl ratio of
Eucalyptus globulus wood lignin by analytical pyrolysis. Journal of Analytical and Applied
Pyrolysis, v. 48, n. 2, p. 121–128, Jan. 1999.
RÜGGEBERG, M.; SPECK, T.; PARIS, O.; LAPIERRE, C.; POLLET, B.; KOCH, G.;
BURGERT, I. Stiffness gradients in vascular bundles of the palm Washingtonia robusta.
Proceedings of the royal society B, v. 275, p. 2221–2229, 2008.
SALAZAR, V. L. P.; LEÃO, A. L. Biodegradação das fibras de coco e de sisal aplicadas na
indústria automotiva. Revista Energia na Agricultura, v. 21, n. 2, p. 99-133, 2006.
SALLEEM, A.; KIVELA, H.; PIHLAJA, K. Antioxidant activity of pine bark constituents,
Zeitschrift für Naturforschung, v. 58, p. 351–354, 2003.
SALIBA, E. O. S.; PILÓ-VELOSO, N. M. R. D.; MORAIS, S. A. L.; PILÓ-VELOSO, D.
Lignins – Isolation methods and chemical characterization. Ciência Rural, v. 31, n. 5, p.917-
928, 2001.
SANTANA, A. C.; CARVALHO, D. F.; MENDES, F.A. T. Organização e competitividade
das empresas de polpas de frutas no Estado do Pará : 1995 a 2004. Unama, 2006.
SARKANEN, S. Template polymerization in lignin biosynthesis. In: LEWIS, N.G.;
SARKANEN, S. (Ed.). Lignin and lignan biosynthesis. Washington, DC: American Chemical
Society Symposium, 1998. p. 194-208.
SARKANEN, K. V.; CHANG, H-M.; ALLAN, G. G. Species variation in lignins. III.
Hardwood lignins. Tappi, v. 50, n. 12, p. 587–590, 1967.
SATYANARAYANA, K. G.; GUIMARÃES, J. L.; WYPYCH F. Studies on lignocellulosic
fibers of Brazil. Composites, n. 38, p. 1694–1709, 2007.

64
SCALBERT, A.; MONTIES, B. Comparison of Wheat Straw Lignin Preparations. II. Straw
Lignin Solubilisation in Alkali. Holzforschung, v. 40, n. 4, p. 249-254, 1986.
SCALBERT, A.; MONTIES, B.; LALLEMAND, J. Y.; GUITTET, E.; ROLANDO, C. Ether
linkage between phenolic acids and lignin fractions from wheat straw. Phytochemistry, v. 24,
n. 6. P. 1359 – 1362, 1985.
SCHWANNINGER, M.; RODRIGUES, J. C.; PEREIRA, H.; HINTERSTOISSER, B. Effects
of short-time vibratory ball milling on the shape of FT-IR spectra of wood and cellulose.
Vibrational Spectroscopy, v. 36, n. 1, p. 23-40, 2004.
SHE, D.; NIE, X. N.; XU, F.; GENG, Z. C.; JIA, H. T.; JONES, G. L.; BAIRD, M. S.
Physico-chemical characterization of different alcohol-soluble lignins from rice straw,
Cellulose chemistry and technology, v. 46, n. (3-4), p. 207-219, 2012.
SHEIN, I. V.; POLYAKOVA, G. G.; ZRAZHEVSKAYA, G. K.; PASHENOVA, N.V.;
VETROVA, V. P. Accumulation of phenolic compounds in conifer callus cultures in response
to wood blue-stain fungi, Russ. Journal of Plant Physiology, v. 48, p. 251–256, 2001.
SILLS, D. L.; GOSSETT, J. M. Using FTI to predict saccharification from enzymatic
hydrolysis of alkali pretreated biomasses. Biotechnology Bioengineering, v. 109, p. 353–
362, 2012.
SILVA, J. C.; BARRICHELO L. E. G.; BRITO, J. O. Endocarpos de babaçu e de macaúba
comparados a madeira de Eucalyptus grandis para a produção de carvão vegetal. Revista
IPEF, Piracicaba, n. 34, p. 31-34, 1986.
SILVERSTEIN, R. A.; CHEN, Y.; SHARMA-SHIVAPPA, R. R.; BOYETTE, M. D.;
OSBORNE, J. A comparison of chemical pretreatment methods for improving
saccharification of cotton stalks. Bioresource Technology, v. 98, n. 16, p. 3000-3011, 2007.
SMITH, L. G. Plant cell division: building walls in the right places. Nature Reviews
Molecular Cell Biology, vol 2, p. 33-39, 2001.
SMITH, A. Applied spectroscopy. Moscow: Mir. 1982.
SMITH, D. C. C. p-Hydroxybenzoate groups in the lignin of aspen (Populus tremula). Journal
of the Chemical Society, p. 2347-2351, 1955.
STICKLEN, M. B. Plant genetic engineering for biofuel production: Towards affordable
cellulosic ethanol. Nature Reviews. Genetics, v. 9, p. 433-443, 2008.
STEWART, J. J.; AKIYAMA, T.; CHAPPLE, C.; RALPH, J.; MANSFIELD, S. D. The
effects on lignin structure of overexpression of Ferulate 5-hydroxylase in hybrid poplar. Plant
Physiology, v. 150, p. 621–635, 2009.

65
STEWART, D.; AZZINI, A.; HALL, A. T.; MORRISON, I. M. Sisal fibers and their
constituent non-cellulosic polymers. Industrial Crops and Products, v. 6, n. 1, p. 27-26,
1997.
SUN, Z.Y., TANG, Y. Q.; MORIMURA, S.; KIDA, K. Reduction in environmental impact of
sulfuric acid hydrolysis of bamboo for production of fuel ethanol. Bioresource technology,
v.128, p.87-93, 2013.
SUN, Y. C.; WEN, J. L.; XU, F.; SUN, R. C. Fractional and structural characterization of
organosolv and alkaline lignins from Tamarix austromogoliac. Scientific Research and
Essays, v. 5, n. 24, p. 3850-3864, 2010.
SUN, Y.; CHENG, J. Hydrolysis of lignocellulosic materials for ethanol production: A
review. Bioresource Technology, v. 83, p. 1-11, 2002.
SUN, R. C.; SUN, X. F.; FOWLER, P.; TOMKINSON, J. Strucutural and physicochemical
characterization of lignins solubilized during alkaline peroxide treatment of barley straw.
European Polymer Journal, v. 38, n. 7, p. 1399-1407, 2002.
SUN, R. C.; TOMKINSON, J.; WANG, S. Q.; ZHU, W. Characterization of lignins from
wheat straw by alkaline peroxide treatment. Polymer Degradation and Stability, v. 67, n. 1,
p. 101-109, 2000a.
SUN, R. C.; FANG, J. M.; TOMKISON, J. Delignification of rye straw using hydrogen
peroxide. Industrial Crops and Products, v. 12, n. 2, p. 71-83, 2000.
SUN, R. C.; FANG, J. M.; TOMKINSON, J. Fractional isolation and structural
characterization of lignins from oil palm trunk and empty fruit bunch fibres. Journal of
Wood Chemistry and Technology, v. 19, n. 4, p. 335–356, 1999.
SUN, R. C.; FANG, J.; TOMKINSON, J.; BOLTON, J. Physico-chemical and structural
characterization of alkali soluble lignins from oil palm trunk and empty fruit-bunch fibers.
Journal of Agricultural and Food Chemistry, v. 47, p. 2930-2936, 1999.
SUN, R. C.; LAWTHER, J. M.; BANK, W. B. Effects of pre-treatment time and different
alkalis on the composition of alkali-soluble wheat straw lignin. Journal of Agricultural and
Food Chemistry, v. 44, p. 3965-3970, 1996.
SUZUKI, S.; SHINTANI, H.; PARK, S. Y.; SAITO, K.; LAEMSAK, N.; OKUMA, M.;
IIYAMA, K. Preparation of binderless boards from steam exploded pulps of oil palm (Elaeis
guneensis Jaxq.) fronds and structural characteristics of lignin and wall polysaccharides in
steam exploded pulps to be discussed for self-bindings. Holzforschung, v. 52, n. 4, :417–426,
1998.
TAIZ, L.; ZEIGER, E. Paredes celulares: estruturas, biogênese e expansão. In: Fisiologia
vegetal. 3.ed. Porto Alegre: Artmed, 2004. cap. 15, p. 339-364.

66
TAPPI - Technical Association of the Pulp and Paper Industry. TAPPI test methods T 212 os-
76: one per cent sodium hydroxide solubility of wood and pulp. Atlanta: Tappi Technology
Park, 1979.
TERASHIMA, N. Formation and ultrastructure of lignified plant cell walls. ISINPC 1, p. 24-
33, 1993.
TIMELL, T. E. Recent progress in the chemistry of wood hemicelluloses. Wood Science and
Technology, v. 1, n. 1, p. 45–70, 1967.
TLIJANI, M.; GOUADRIA, A.; BENYOUNES, R.; DURASTANTI, J. F.; MAZIOUD, A.
Alkaline Treatment Effect on Mechanical Properties of Date palm Wood Fiber. International
Journal of Engineering Sciences & Research Technology, v. 3, n. 2, p. 956-961, 2014.
TOMIMURA, Y. Chemical characteristics and utilization of oil palm trunks. Japan
Agricultural Research Quarterly, v. 25, n. 4, p. 283–288, 1992a.
TOMIMURA, Y. Chemical characteristics of oil palm trunk. Bulletin of the Forestry and
Forest Products Research Institute, n. 362, p. 133–142, 1992b.
TOMLINSON, P. B.; HUGGETT, B. Cell longevity and sustained primary growth in palm
stems. American Journal of Botany, v. 99, n. 12, p. 1891-1902, 2012.
TOMLINSON, P. B. The structural biology of palms. Oxford University Press, Oxford, UK.
492 p. 1990.
TOMLINSON, P. B. The structural biology of palms. Oxford University Press, Oxford,
UK. 1990, 492 p.
VALLET, C.; CHABBERT, B.; CZANINSKI, Y.; MONTIES, B. Histochemistry of lignin
deposition during sclerenchyma differentiation in Alfalfa stems. Annals of Botany, v. 78, n.
5, p. 625–632, 1996.
VANHOLME, R.; DEMEDTS, B.; MORREEL, K.; RALPH, J.; BOERJAN, W. Lignin
biosynthesis and structure. Plant Physiology, v. 153, p. 895-905, 2010.
VENVERLOO, C. J. Lignin of Populus nigra L. cv. ‘Italica’ and some other Salicaceae.
Holzforschung, v. 25, n. 1, p. 18–24, 1971.
VOGEL, J. Unique aspects of the grass cell wall. Current Opinion of Plant Biology, v. 11,
p. 301-307, 2008.
WAMBUA P, IVENS J, VERPOEST I. Natural fibers: can they replace glass in fiber-
reinforced plastics. Composites Science and Technology, v. 63, p. 1259–1264, 2003.

67
WANG, N.; LIU, W.; HUANG, J.; MA, K. The structure– mechanical relationship of palm
vascular tissue. Journal of the mechanical behavior of biomedical materials, v. 36, p. 1–
11, 2014.
WEINER, G.; LIESE, W.; SCHMITT, U. Cell wall thickening in fibers of the palms Rhapis
excelsa (Thunb.) Henry and Calamus axillaris Becc. In: DONALDSON, L. A., SINGH, A.
P., BUTTEFIELD, B. G., WHITE-HOUSE, L. Recent Advances in Wood Anatomy. NZFRI
Ltd, Rotorua, New Zealand, 1996, p. 191–197.
WEN, J. L.; YUANI, T. Q.; SUN, S. L.; XU, F.; SUN, R. C. Understanding the Chemical
Transformations of Lignin during Ionic Liquid Pretreatment. Green Chemistry, v. 16, p.
181-190, 2014.
WHETTEN, R. W.; MACKAY, J. J.; SEDEROFF, R. R. Recent advances in understanding
lignin biosynthesis. Annual Review of Plant Physiology and Plant Molecular Biology, v.
49, n. 1, p. 585-609, 1998.
WIKBERG, H.; MANNU, S. L. Characterization of thermally modified hard and soft woods
by 13
C CP MAS NMR. Carbohydrate Polymers, v.58, p. 461-466, 2004.
WITHERS, S.; LU, F.; KIM, H.; ZHU, Y.; RALPH, J.; WILKERSON, C. G. Identification of
a grass-specific enzyme that acylates monolignols with p-coumarate. The Journal of
Biological Chemistry, v. 287, n. 11, p. 8347–8355, 2012.
WÓJCIAK-KOSIOR, M,; ONISZCZUK, A. Sample Preparation and TLC Analysis of
Phenolic Acids. In: M. WAKSMUNZKA-HAJNOS; J. SHERMA; T. KOWALSKA. Thin
Layer Chromatography in Phytochemistry. Boca Raton: Taylor & Francis., 2008, p. 332–
359.
WU, J.; FUKAZAMA, K.; OHTANI, J. Lignin Analysis in Some Tropical Hardwoods U sing
Ultraviolet Microscopy. Research Bulletins of the College Experiment Forests. v. 47, n. 2,
p. 353-366, 1990.
XIAO, B.; SUN, X. F.; SUN, R. Chemical, structural, and thermal characterizations of alkali-
soluble lignins and hemicelluloses, and cellulose form maize stem, rye stram, and rice straw.
Polymer Degradation and Stability, v. 74, n. 2, p. 307-319, 2001.
XU, F.; YU, J.; TESSO, T.; DOWELL, F.; WANG, D. Qualitative and quantitative analysis
of lignocellulosic biomass using infrared techniques: A mini-review. Applied Energy, v. 104,
p. 801–809, 2013.
YAMAMOTO, E.; BOKELMAN, G. H.; LEWIS, N. G. Phenylpropanoid metabolism in cell
walls: an overview. In: Plant Cell Wall Polymers: Biogenesis and Biodegradation (GN Lewis,
MG Paice, eds) ACS Symposium Series, 676 p, Am Chem Soc, Washington, DC, USA, 399,
68-88, 1989.

68
YAMAMOTO, E.; BOKELMAN, G. H.; LEWIS, N. G. Phenylpropanoid metabolism in cell
walls: an overview. In: LEWIS, N.; PAICE, M. Plant cell wall polymers. Washington, DC:
American Chemical Society, 1989.p. 68-88, 1989.
YAN, T.; XU, Y.; YU, C. The Isolation and Characterization of Lignin of Kenaf Fiber.
Journal of Applied Polymer Science, v. 114,n 3, p. 1896–1901, 2009.
YOU, T-T.; MAO, J-Z.; YUAN, T-Q.; WEN, J-L.; XU, F. Structural Elucidation of the
Lignins from Stems and Foliage of Arundo donax Linn. Journal of agricultural and food
chemistry, v. 61, n 22, p. 5361–5370, 2013.
YUAN, T. Q.; SUN, S. N.; XU, F.; SUN, R. C. Characterization of ligninstructures and LCC
linkages by quantitative 13C and 2D HSQC NMR spectroscopy. Journal of Agricultural
and Food Chemistry, v. 59, n.19, 10604 –10614, 2011.
YUAN, T.; SUN, S.; XU, F.; SUN, R. Characterization of Lignin Structures and Lignin–
Carbohydrate Complex (LCC) Linkages by Quantitative13C and 2D HSQC NMR
Spectroscopy. Journal of Agricultural and Food Chemistry, v. 59, n. 19, p. 10604-10614,
2011.