triple expression of b7-1, b7-2 and 4-1bbl enhanced antitumor immune response against mouse h22...
TRANSCRIPT

J Cancer Res Clin Oncol (2011) 137:695–703
DOI 10.1007/s00432-010-0905-9ORIGINAL PAPER
Triple expression of B7-1, B7-2 and 4-1BBL enhanced antitumor immune response against mouse H22 hepatocellular carcinoma
Guoqiang Li · Xiaofeng Wu · Feng Zhang · Xiangcheng Li · Beicheng Sun · Yue Yu · Aihong Yin · Lei Deng · Jie Yin · Xuehao Wang
Received: 26 January 2010 / Accepted: 26 April 2010 / Published online: 19 June 2010© Springer-Verlag 2010
AbstractObjectives Costimulatory signals are essential for T-cellactivation and hence play a very important role in antitumorimmunity. B7 and 4-1BBL which belongs to tumor necro-sis factor (TNF) family provide costimulatory interactionfor T-cell activation and function. This study investigatedthe role of B7 and 4-1BBL in the ampliWcation of tumorimmunity by transduction of the B7-1, B7-2 and 4-1BBLinto mouse hepatocellular carcinoma cell line H22.Methods The tumorigenicity of H22 variants expressingeither B7-1, B7-2 (H22/B7-1/B7-2) or 4-1BBL wascompared with an H22 variant expressing B7-1, B7-2 and4-1BBL (H22/B7-1/B7-2/4-1BBL). The study next investi-gated whether the combination of B7-1/B7-2 and 4-1BBLcell injection induced cytotoxic T lymphocyte (CTL)response and IL-2/IFN-� secretion. The immune mecha-nisms underlying this combination treatment were thenanalyzed.Results Syngeneic BALB/c mice injected with H22/B7-1/B7-2/4-1BBL cells that expressed elevated levels of B7-1,B7-2 and 4-1BBL showed a tumor development frequency
of 50% compared with 100% in mice injected with the H22parental line, H22/neo, H22/B7-1/B7-2 and H22/4-1BBL.Mice inoculated with H22 tumor cells expressing B7-1, B7-2 and 4-1BBL developed a strong cytotoxic T lymphocyteresponse and long-term immunity against wild-type tumor,suggesting a synergistic eVect between the B7 and 4-1BBLcostimulatory pathways. Results showed that H22/B7-1/B7-2/4-1BBL tumor vaccines probably protect the inWltrat-ing lymphocytes from apoptosis and induce NF-�B activa-tion to improve T-cell-mediated antitumor response.Conclusions In this study, the antitumor consequences ofusing B7-1, B7-2 and 4-1BBL gene transfer have demon-strated the therapeutic potential of gene therapy approachfor hepatocellular carcinoma.
Keywords Carcinoma · Hepatocellular · Tumor immunity · B7-1 · B7-2 · 4-1BBL · NF-�B
Introduction
The activation of T cells plays a central role in antitumorimmunity. In order to fully activate a naive T cell, two keysignals are thought to be required to destroy tumor cells.The Wrst signal is provided by the interaction of the T-cellreceptor (TCR) with major histocompatibility class (MHC)antigens on antigen-presenting cells (APCs). The second,or costimulatory, signal is antigen independent and is pro-vided by members of the B7 and/or tumor necrosis factor(TNF) family. The B7-CD28 is one of the best character-ized costimulatory pathways, which are generated by bind-ing of B7-1 (CD80) and B7-2 (CD86) expressed on APCsto the T-cell surface molecule CD28. This provides the pri-mary costimulatory interaction for initial T-cell activationand induction of eVector function (Greenwald et al. 2005).
Guoqiang Li and Xiaofeng Wu contributed equally to this work.
G. Li · X. Wu · F. Zhang · X. Li · B. Sun · Y. Yu · A. Yin · L. Deng · X. Wang (&)Liver Transplantation Center, First AYliated Hospital of Nanjing Medical University, 300 Guangzhou Road, Nanjing 210029, Chinae-mail: [email protected]
J. Yin (&)Department of Respiratory Medicine, Jinling Hospital, 305 Zhongshan East Road, Nanjing 210002, Chinae-mail: [email protected]
123

696 J Cancer Res Clin Oncol (2011) 137:695–703
However, another class of molecules, which belongs to theTNF family, includes the 4-1BB ligand (4-1BBL,CD137L). It can also provide costimulation to T cells andmay have important functions in modulating the immuneresponse. This ligand expressed on APCs interacts with itsreceptor (4-1BB) expressed on the surface of T cells. Inaddition to TCR engagement, it induces T-cell proliferationand cytokine secretion (Croft 2003). The 4-1BBL signalingpathway appears to modulate the magnitude and duration ofthe immune response and the size of subsequent immunememory compartments (Bertram et al. 2002; Takahashiet al. 1999).
Antigen encounter with either inadequate costimulationor a lack of costimulation, however, results in T-cell desen-sitization or anergy. There is now increasing evidence toshow that tumor cells are generally poor APCs, due to anabsence or downregulation in costimulatory molecules and/or peptide presentation through MHC molecules, allowingthem to escape protective immune responses (Pardoll 2003;Schultze et al. 1996). Hepatocellular carcinoma (HCC) is aleading cause of cancer-related death worldwide, especiallyin Asia and South Africa (Parkin et al. 2005). Surgicalresection is curative but is restricted to patients with earlystaging and compensated liver function. Although themulti-kinase inhibitor sorafenib is currently the onlyapproved agent for advanced HCC (Llovet et al. 2008;Melero et al. 2007), HCC is clearly a disease for whichnovel therapeutic strategies must be developed to improvethe prognosis of aVected patients. The development ofcostimulatory strategies with the common goal of elicitingtumor cell destructive immune responses is currently thefocus of cancer immunotherapeutic approaches (Meleroet al. 2007; Tamada and Chen 2006). Accumulating datahave revealed that costimulatory molecules are often down-regulated or lost in HCC cells (Fujiwara et al. 2004; Tats-umi et al. 1997). Previous investigators have shown that thegenetic modiWcation of HCC cells to express elevated lev-els of costimulatory molecules can elicit protective andsometimes curative immunity against unmodiWed HCCtumor cells (Li et al. 2004). In addition, the expression ofmany costimulatory molecules in a tumor cell seems to bemore eYcient than the expression of single molecule ineliciting tumor-speciWc T-cell immunity (Disis et al. 2009).
In this study, a gene therapy approach was taken totransfect HCC cells with the immunostimulatory antigensB7-1, B7-2 and/or 4-1BBL onto surfaces of tumor cells.This study demonstrated that B7 synergizes with the4-1BBL costimulatory pathway to amplify systemic T-cell-mediated antitumor immunity, as demonstrated by the initi-ation of an eVective CTL response to tumor antigens andthe rejection of the parental tumor in vivo. Further analysiswas done on the immune mechanisms underlying this com-bination treatment.
Materials and methods
Cell lines, transfection and cloning
H22, a mouse HCC cell line (BALB/c background) wasobtained from the China Center for Type Culture Collec-tion (Wuhan, China) and cultured in DMEM with 10% FCSin a humidiWed 5% CO2 atmosphere. The murine full-length B7-1, B7-2 and 4-1BBL cDNA were obtained andsubcloned into a PCI-neo mammalian expression vector(Promega, Madison, WI). The recombinants were namedPCI-neo-B7-1, PCI-neo-B7-2 and PCI-neo-4-1BBL, anddigested with restriction enzyme and sequenced. Therecombinant named pcDNA3.1-B7.1-IRES-B7.2 was con-structed and conWrmed with restriction enzyme digestion.Subsequently, four recombinants were transfected into H22with Lipofectamine Reagent (Invitrogen, Carlsbad, CA).G418-resistance colonies were then acquired and namedH22/B7-1, H22/B7-2, H22/4-1BBL and H22/B7-1/B7-2,respectively. B7-1, B7-2 and 4-1BBL triple transfectantswere generated by retransfection of H22/B7-1/B7-2 cellswith PCI-neo-4-1BBL. Flow cytometry were applied todetermine whether there was stable expression of targetgenes in variants.
Cell counts and phenotyping
Cell counts were performed using 0.04% trypan blue/PBS(Sigma–Aldrich, Co. Ltd., Dorset, UK). Expression of cellsurface molecules on parental H22 cells and transducedH22 cells was determined using immunoXuorescence stain-ing according to standard procedures. For phenotypicanalysis of transfectants, FITC-conjugated anti-B7-1 mAb,PE-conjugated anti-B7-2 mAb and FITC-conjugated mAbto murine 4-1BBL and appropriate isotype controls(Diaclone, Besançon, France) were used. Analysis wasperformed using a FACSCalibur Xow cytometer (Becton–Dickinson, San Jose, CA).
In vivo tumor vaccination
BALB/c mice, 6–8 weeks of age, obtained from the Shang-hai Laboratory Animal Center of Chinese Academy of Sci-ences, were used for inoculation of hepatoma H22 cells. Allanimal procedures were conducted in compliance with theguidelines approved by the China Association of Labora-tory Animal Care and the Institutional Animal Care Com-mittee.
For primary tumor inoculation, parental H22 (H22/wt),H22/neo, H22/B7-1/B7-2, H22/4-1BBL and H22/B7-1/B7-2/4-1BBL at 1 £ 106 cells/mouse were injected subcutane-ously into the left Xank of BALB/c mice using 26-gaugeneedles. For challenge experiments, mice that had achieved
123

J Cancer Res Clin Oncol (2011) 137:695–703 697
long-term survival after the initial inoculation were chal-lenged with 1 £ 106 wild-type H22 cells injected into theright Xank. The mice were scored for tumor growth twice aweek. Tumor volume was estimated using the followingequation: tumor volume (mm3) = [long length (mm) £(short length (mm))2]/2 (Mogi et al. 2000). The survivaltime of the mice was also monitored.
Cytotoxic T lymphocyte (CTL) assays
Mice that survived primary injection with the H22 variants,as well as those challenged with parental H22 at 56 days,were killed for CTL assay. The cells were analyzed forcytotoxicity by quantitative measurements of lactate dehy-drogenase (LDH) using CytoTox96 nonradioactive cyto-toxicity assay kits (Promega, Madison, WI) according tothe manufacturer’s protocol. This system quantiWes therelease of LDH from target cells. The assays were per-formed with eVector cells and target cells (1 £ 104 perwell) mixed together at various ratios in a Wnal volume of200 �l. After incubation at 37°C and 5% CO2 for 6–9 h, a50-�L sample of supernatant was collected from each wellto assess the amount of LDH. The percentage of speciWcLDH release was calculated as follows: percentagecytotoxicity = (experimental release ¡ spontaneous release)/(maximum release ¡ spontaneous release) £ 100.
Cytokine measurements
At diVerent times (14, 35, 56 and 84 days) post Wrst inocula-tion of tumor cells, mice were killed, and the mixed T-lym-phocyte culture (MLC) was prepared according to standardtechniques. Killed mice included those that survived primaryinjection and those that were subsequently challenged withparental tumors. Spleens were collected, and single-cell sus-pensions were prepared. Splenocytes were mixed with H22stimulator cells at a 1:1 cell ratio in 96-well plates (Corning,Rochester, NY). Three days after initiation of the culture, ali-quots of the culture supernatant were collected to measureIL-2 and IFN-� production by ELISA (BD Pharmingen, CA).A total of 5 £ 106 mouse spleen cells were incubated with105 H22 cells in 96-well plates. Controls included spleencells from irrelevantly immunized mice. Culture supernatantswere collected after 48 h for measuring the mean levels (pg/ml) of IL-2 and IFN-� using commercial ELISA kits (BDPharmingen, San Diego, CA). ELISA was performed accord-ing to the manufacturer’s instructions. All ELISA determina-tions were carried out in triplicate.
Detection of apoptosis
To measure T-cell activation-induced cell death (AICD),the quantiWcation of DNA fragmentation was determined
by Xow cytometry. Spleens were removed on day 21 postinoculation. Spleen T cells were pelleted, washed with ice-cold PBS in 96-well plates, and gently resuspended in250 �L buVer (propidium iodide 50 �g/mL, 0.1 mol/Lsodium citrate, 0.1% Triton £100). After incubation over-night in the dark at 4°C, genomic DNA content was deter-mined using a FACSCalibur Xow cytometer (Becton–Dickinson, CA, USA) and CellQuest analysis software. Theexperiment was performed in quadruplicate. Nuclei dis-playing a hypodiploid, or sub-G1, population were identi-Wed as apoptotic.
Electrophoretic mobility-shift assay (EMSA)
EMSA was performed using nuclear extracts prepared fromspleen, using the Promega gel shift assay system and 32P-labeled NF-�B consensus oligonucleotide. For EMSAexperiments, the following double-stranded oligonucleo-tides were used: NF-�B, 5�-AGT GAG GGG ACT TTCCCA GGC-3�, 3�-TCA ACT CCC CTG AAA GGG TCCG-5�. Oligonucleotides were annealed with the use ofKlenow DNA polymerase and then 5�-end labeled with[�-32P]ATP with the use of T4 polynucleotide kinase, usingstandard procedures (Curry et al. 1999). For competitionexperiments, 50-fold molar excess of unlabeled oligonu-cleotide was added to the binding reaction. Samples wereloaded on a 4.8% polyacrylamide gel and detected by auto-radiography. Supershift studies were carried out by prein-cubation with anti-p65 and anti-p50 (Santa CruzBiotechnology, Santa Cruz, CA) for 30 min.
Statistical analysis
The data are expressed as the means § SEM. Analysis wasperformed using SPSS 12.0 software (SPSS, Chicago, IL).Statistical signiWcance was determined by one-wayANOVA test and P < 0.05 was considered to be statisti-cally signiWcant. The Kaplan–Meier method was used toestimate the probability of survival, and statistical signiW-cance was tested using log-rank statistics.
Results
Generation and phenotyping of variants
H22 cells were transfected with PCI-neo-B7-1, PCI-neo-B7-2, PCI-neo-4-1BBL, pcDNA3.1-B7-1-IRES-B7-2, andthe G418-resistant (PCI-neo-B7-1, PCI-neo-B7-2, and PCI-neo-4-1BBL) or hygromycin-resistant (pcDNA3.1-B7-1-IRES-B7-2) cells were selected. The wild-type tumor,the vector-only transfected H22-PCI-neo-B7-1, H22-PCI-neo-B7-2, H22-PCI-neo-4-1BBL, the double transfectant
123
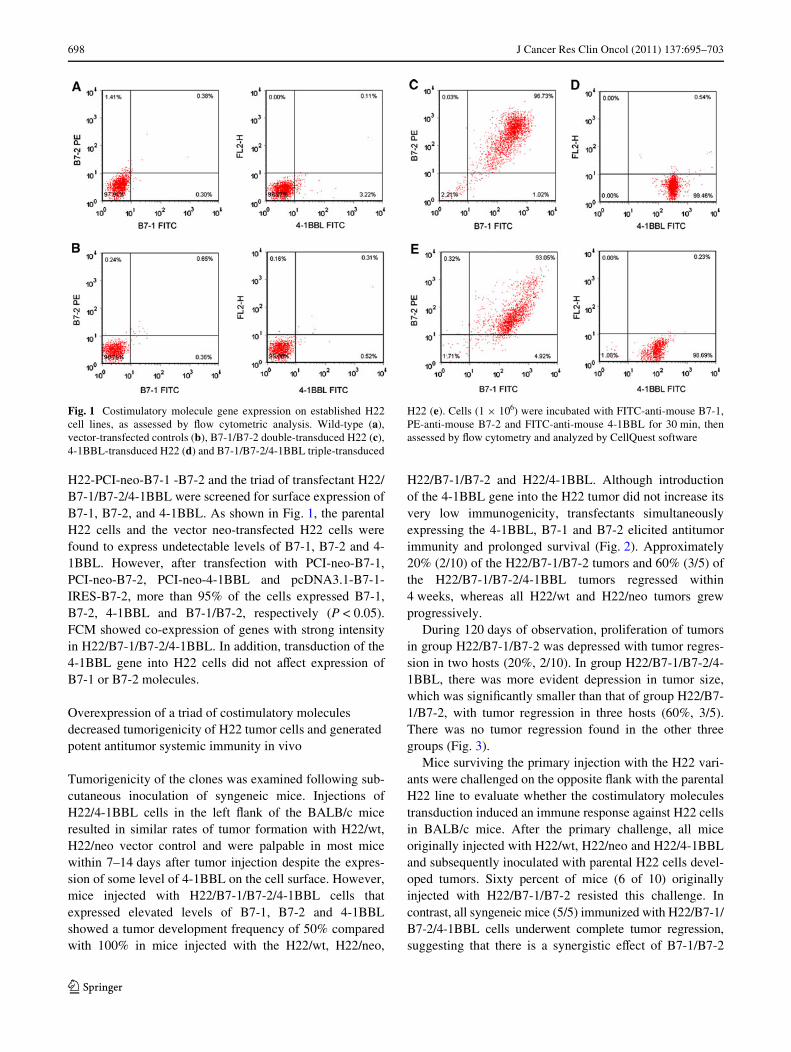
698 J Cancer Res Clin Oncol (2011) 137:695–703
H22-PCI-neo-B7-1 -B7-2 and the triad of transfectant H22/B7-1/B7-2/4-1BBL were screened for surface expression ofB7-1, B7-2, and 4-1BBL. As shown in Fig. 1, the parentalH22 cells and the vector neo-transfected H22 cells werefound to express undetectable levels of B7-1, B7-2 and 4-1BBL. However, after transfection with PCI-neo-B7-1,PCI-neo-B7-2, PCI-neo-4-1BBL and pcDNA3.1-B7-1-IRES-B7-2, more than 95% of the cells expressed B7-1,B7-2, 4-1BBL and B7-1/B7-2, respectively (P < 0.05).FCM showed co-expression of genes with strong intensityin H22/B7-1/B7-2/4-1BBL. In addition, transduction of the4-1BBL gene into H22 cells did not aVect expression ofB7-1 or B7-2 molecules.
Overexpression of a triad of costimulatory molecules decreased tumorigenicity of H22 tumor cells and generated potent antitumor systemic immunity in vivo
Tumorigenicity of the clones was examined following sub-cutaneous inoculation of syngeneic mice. Injections ofH22/4-1BBL cells in the left Xank of the BALB/c miceresulted in similar rates of tumor formation with H22/wt,H22/neo vector control and were palpable in most micewithin 7–14 days after tumor injection despite the expres-sion of some level of 4-1BBL on the cell surface. However,mice injected with H22/B7-1/B7-2/4-1BBL cells thatexpressed elevated levels of B7-1, B7-2 and 4-1BBLshowed a tumor development frequency of 50% comparedwith 100% in mice injected with the H22/wt, H22/neo,
H22/B7-1/B7-2 and H22/4-1BBL. Although introductionof the 4-1BBL gene into the H22 tumor did not increase itsvery low immunogenicity, transfectants simultaneouslyexpressing the 4-1BBL, B7-1 and B7-2 elicited antitumorimmunity and prolonged survival (Fig. 2). Approximately20% (2/10) of the H22/B7-1/B7-2 tumors and 60% (3/5) ofthe H22/B7-1/B7-2/4-1BBL tumors regressed within4 weeks, whereas all H22/wt and H22/neo tumors grewprogressively.
During 120 days of observation, proliferation of tumorsin group H22/B7-1/B7-2 was depressed with tumor regres-sion in two hosts (20%, 2/10). In group H22/B7-1/B7-2/4-1BBL, there was more evident depression in tumor size,which was signiWcantly smaller than that of group H22/B7-1/B7-2, with tumor regression in three hosts (60%, 3/5).There was no tumor regression found in the other threegroups (Fig. 3).
Mice surviving the primary injection with the H22 vari-ants were challenged on the opposite Xank with the parentalH22 line to evaluate whether the costimulatory moleculestransduction induced an immune response against H22 cellsin BALB/c mice. After the primary challenge, all miceoriginally injected with H22/wt, H22/neo and H22/4-1BBLand subsequently inoculated with parental H22 cells devel-oped tumors. Sixty percent of mice (6 of 10) originallyinjected with H22/B7-1/B7-2 resisted this challenge. Incontrast, all syngeneic mice (5/5) immunized with H22/B7-1/B7-2/4-1BBL cells underwent complete tumor regression,suggesting that there is a synergistic eVect of B7-1/B7-2
Fig. 1 Costimulatory molecule gene expression on established H22cell lines, as assessed by Xow cytometric analysis. Wild-type (a),vector-transfected controls (b), B7-1/B7-2 double-transduced H22 (c),4-1BBL-transduced H22 (d) and B7-1/B7-2/4-1BBL triple-transduced
H22 (e). Cells (1 £ 106) were incubated with FITC-anti-mouse B7-1,PE-anti-mouse B7-2 and FITC-anti-mouse 4-1BBL for 30 min, thenassessed by Xow cytometry and analyzed by CellQuest software
123

J Cancer Res Clin Oncol (2011) 137:695–703 699
and 4-1BBL. These results indicate that tumor variantsexpressing B7-1/B7-2 and B7-1/B7-2/4-1BBL are immu-nogenic and protect against further systemic challenge withunmodiWed parental H22 cells. Furthermore, long-termtumor immunity is induced by the modiWed tumors bysimultaneously expressing B7-1, B7-2 and 4-1BBL. Theseexperiments also demonstrated that 4-1BBL is not requiredfor the eVector phase of the immune response. The systemicimmunity induced by H22/B7-1/B7-2 and H22/B7-1/B7-2/4-1BBL protected the hosts against a subcutaneous chal-lenge with parental wild H22 cells. The length of immuneprotection from H22/B7-1/B7-2 was shorter than that fromH22/B7-1/B7-2/4-1BBL, which converted the H22 cell lineinto a long-term antitumor vaccine.
Co-expression of B7-1, B7-2 and 4-1BBL genes induced cytolytic activity against H22 cells and IL-2/IFN-� production
The study next investigated whether the combination ofB7-1/B7-2 and 4-1BBL cell injection induced CTLs thatcan lyse tumor cell targets. As shown in Fig. 4, splenocytes
from mice treated with B7-1/B7-2 or B7-1/B7-2/4-1BBLtransduced cells displayed high-level cytolytic activityagainst H22 cells, whereas CTLs from mice treated withnon- or neo-transduced cells displayed low reactivity.Importantly, splenocytes from mice treated with B7-1/B7-2/4-1BBL cells displayed signiWcantly stronger cytolyticactivity versus mice treated with B7-1/B7-2 cells. Theseresults demonstrate that the combined B7-1/B7-2 and 4-1BBL therapy was superior in its ability to stimulate CTLactivity.
The study then evaluated whether H22 cells expressingB7-1/B7-2 and/or 4-1BBL could stimulate BALB/c spleno-cytes to produce IL-2/IFN-�. After a 3-day MLC, the con-centration of IL-2/IFN-� in the culture supernatants wasassessed by ELISA. The MLC with B7-1/B7-2/4-1BBL-transfected H22 cells generated signiWcantly elevatedamounts of IL-2 (276.5 § 31.4 pg/ml) and IFN-�(305.6 § 21.3 pg/ml) when compared those with H22/neocells (IL-2: 12.4 § 4.1 pg/ml; IFN-�: 7.4 § 2.3 pg/ml) orH22 cells alone (IL-2: 9.2 § 2.4 pg/ml; IFN-�: 11.2 §3.6 pg/ml; P < 0.05 Figs. 5a, b). Also, H22 cells transfectedwith B7-1/B7-2/4-1BBL stimulated syngeneic BALB/c
Fig. 2 Growth kinetics of wild-type H22 tumors or transfected with mutant variants. Syngeneic mice were challenged with ei-ther wild-type, vector-transfec-ted controls, B7-1/B7-2 double-transduced H22, 4-1BBL-trans-duced H22 or B7-1/B7-2/4-1BBL triple-transduced H22 cells. The mice were inoculated subcutaneously with 1 £ 106 tumor cells, and the volume of tumors that subsequently developed was estimated twice weekly
Fig. 3 Survival of syngeneic mice vaccinated with H22 cell variants.Survival of the mice (y-axis) over a 120-day follow-up period (x-axis)indicated that there was signiWcant diVerence in the tumorigenicity ofwild-type, vector-transfected controls, B7-1/B7-2 double-transduced
H22, 4-1BBL-transduced H22 or B7-1/B7-2/4-1BBL triple-trans-duced H22 cells. The B7-1/B7-2/4-1BBL triple-transduced variantsformed tumors with low frequency in the BALB/c mice
123

700 J Cancer Res Clin Oncol (2011) 137:695–703
splenocytes to produce signiWcantly more IL-2 or IFN-�than cells transfected only with B7-1/B7-2 (IL-2:74.9 § 14.7 pg/ml; IFN-�: 184.4 § 23.3 pg/ml, P < 0.01)or 4-1BBL (IL-2: 97.2 § 11.2 pg/ml; IFN-�: 97.6 § 7.8 pg/ml).
4-1BBL together with B7-1 and B7-2 protect T lymphocytes from apoptosis
Recent reports showed a correlation between the capacityof CD28 itself to induce the expression of Bcl-xL and toprotect T cells from apoptosis. This notion was conWrmedin the current study by FACS of T cells 21 days after H22cells inoculation. H22/wt, H22/neo immunotherapyresulted in more T-cell apoptosis than B7-1/B7-2 and/or 4-1BBL immunotherapy at the same time points (P < 0.05,Fig. 6). In addition, at the same time points, T cells fromthe group treated with combination of B7-1/B7-2 and 4-1BBL tumor vaccine had lower apoptotic percentages thanthose from the group treated only with tumor vaccine(P < 0.003). The results suggest that the H22/B7-1/B7-2/4-1BBL tumor vaccines probably protect the inWltrating lym-phocytes from apoptosis, thus increasing the T-cell numberand function.
B7-1, B7-2 and 4-1BBL mediated NF-�B activation
Previous study conWrmed the capability of 4-1BB to induceNF-�B-mediated gene expression (Arch and Thompson1998). In this study, NF-�B activation was apparent in B7-1/B7-2 and/or 4-1BBL tumor vaccines measured by EMSA(Fig. 7). NF-�B activity of T cells in both H22/wt andH22/neo groups was very low. However, its activity in
B7-1/B7-2/4-1BBL group was stable and remarkablyhigher than those in B7-1/B7-2 and 4-1BBL groups at thesame time point post Wrst inoculation.
Fig. 4 CTL responses of splenocytes in mice rejecting tumor chal-lenge with H22 cell variants. Splenocytes from mice that survived pri-mary injection and systemic challenge with the parental tumor cellswere restimulated in vitro for 5 days and used in a CTL assay. Cyto-toxic activity against parental H22 cells was measured in a LDH-re-lease assay and speciWc lysis was measured at four ratios of eVector/target cells (E:T ratio). The data are representative of three experiments
Fig. 5 IL-2 and IFN-� secretion by H22 cell variants as measured byELISA. Splenocytes from BALB/c mice that survived primary injec-tion with wild-type, vector transfected controls, B7-1/B7-2 double-transduced H22, 4-1BBL-transduced H22 and B7-1/B7-2/4-1BBLtriple-transduced H22 cells were restimulated in vitro for 48 h withH22 cells. Supernatants were analyzed by ELISA for the secretion ofIL-2 (a) and IFN-� (b), respectively. The assay was performed in trip-licate. Each bar represents the level of cytokine in the supernatant
Fig. 6 Decreased apoptosis of spleen T cells of mice treated with B7-1/B7-2 and/or 4-1BBL tumor vaccine. Mice were inoculated subcuta-neously with B7-1/B7-2 and/or 4-1BBL cells, and spleens wereremoved on day 21 post inoculation. Splenocytes were isolated andthen analyzed for apoptosis using a FACSCalibur Xow cytometer. Theexperiment was performed in quadruplicate
123

J Cancer Res Clin Oncol (2011) 137:695–703 701
Discussion
Tumor cells display poor costimulatory molecule expres-sion which leads them to escape immune surveillance.Costimulatory molecules play an important role in initiat-ing and maintaining antitumor immune responses. For thisreason, genetically modiWed tumor cells expressingcostimulatory molecules have been used as cancer vaccinesin both experimental tumor models and in clinical trials. Inthis study, it was shown that neither B7-1/B7-2 on H22/B7-1/B7-2 variants nor 4-1BBL on H22/4-1BBL variants wereable to mediate long-term antitumor immunity. However,expression of transfected 4-1BBL in conjunction with B7-1and B7-2 levels on the H22 variants resulted in long-termimmunity. Half of the mice injected with H22 variantsexpressing transfected B7-1/B7-2/4-1BBL developedtumors in a 56-day follow-up period (n = 5), while all miceinjected with the H22 parental (n = 10), vector control(n = 10) or H22/4-1BBL variants cells (n = 10) formedtumors within 21 days. Mice that resisted tumor formationfollowing injection with H22/B7-1/B7-2/4-1BBL demon-strated long-term immunity (at least 56 days) when laterchallenged with the H22 parental cell line. In contrast, 60%of mice (6/10) immunized with H22/B7-1/B7-2 were pro-tected during the 21 days following challenge with theparental H22, but 100% of them formed tumors within56 days following challenge. All unimmunized controlmice (n = 10) developed progressive tumors. This study
thus indicates the potential utility of engineered tumor cellsexpressing costimulatory molecules B7-1 and B7-2, espe-cially in combination with costimulatory molecule 4-1BBL,in converting this tumorigenic cell line into an eVectivelong-term antitumor vaccine.
Gene transfer of costimulatory ligands into the tumorcells elicited an immunologic response against the tumor.Although B7-1 and B7-2 have been shown to have diVeren-tial antitumor eVects (Foreman et al. 1999; Martin-Fonte-cha et al. 2000), the double-positive (B7-1 and B7-2)vaccine showed a superior eVect due to the fact that bothB7-1 and B7-2 bind to CD28 and thus produce an additivesignaling eVect. The ligand 4-1BBL (also called CD137L)is a type 2 transmembrane protein and a TNF superfamilymember. The interaction of 4-1BBL and its receptor 4-1BB(CD137) provides a costimulatory signal leading to T lym-phocyte proliferation and survival (Locksley et al. 2001).Tumor cells engineered to overexpress 4-1BBL have beenshown to inhibit tumor outgrowth associated with a strongcytotoxic T-cell response and long-term immunity indiverse mouse models (Muller et al. 2008). However, in thepresent study it was found that 4-1BBL is not required forthe eVector phase of the immune response. This conclusionis supported by a previous study which showed that expres-sion of 4-1BBL on tumors did not increase their sensitivityto lysis by CTL, since 4-1BBL+ and 4-1BBL¡ tumors had acomparable susceptibility to being killed by activated CTLin vitro (Melero et al. 1998). The failure of the 4-1BBL+
tumors to elicit a therapeutic eVect on established wild-typetumors may be caused by a weaker ligand-receptor interac-tion, since higher association/lower dissociation ratesbetween the mAb and 4-1BB have been documented(Shuford et al. 1997). Of note, synergy of 4-1BB and CD28costimulation ampliWed an antitumor immune response andconferred long-lasting protection against challenge withparental tumor (Guinn et al. 1999; Melero et al. 1998). Inline with the data presented here, CD28 appears to play akey role during early primary T-cell activation, while 4-1BB-mediated stimuli seem to be more important duringlate primary and secondary T-cell responses (Bertram et al.2004; Dawicki and Watts 2004). Inclusion of diVerent typesof costimulatory signals, and hence the use of multiple sig-naling pathways will also diVerentially aVect properties thatare not measurable in vitro (Finney et al. 2004). Similarly,preliminary study on another mouse HCC cell line Hepa1-6revealed the same protective immune response (data notshown).
Costimulation is balanced between induction of T lym-phocyte proliferation and inhibition of activation-inducedcell death (AICD). Elimination of T cells by AICD servesas an important mechanism to terminate immune responses,and mainly occurs through the extrinsic pathway of apopto-sis (Plas et al. 2002). Re-exposure to antigen and CD28
Fig. 7 B7-1/B7-2 and/or 4-1BBL-induced activation of NF-�B DNA-binding activity by EMSA. Nuclear extracts from splenocytes of miceinoculated with B7-1/B7-2 transfected H22 cells (Lane 3), 4-1BBLtransfected H22 cells (Lane 4), B7-1/B7-2/4-1BBL triple-transducedH22 cells (Lane 5), and wild type (Lane 1). Transfection with emptyvector (Lane 2) is shown as negative control. Excess of nonlabeledNF-�B probes (£ 100) were used as competitors
123

702 J Cancer Res Clin Oncol (2011) 137:695–703
costimulation reduces AICD and increases the number of Tcells performing eVector functions (KirchhoV et al. 2000).The interaction of CD28 and B7-2 not only enhances IL-2production, which can act as an extrinsic regulator of cellsurvival, but also stimulates T cells actively secrete IL-8and Bcl-xL upregulation protects T cells from apoptosis(Marinari et al. 2004). Okkenhaug et al. employed a trans-genic approach to induce Y170F mutation in CD28, whichprevented the induction of anergy and promoted T-cellproliferation, interleukin 2 secretion and B cell help(Okkenhaug et al. 2001). Furthermore, Starck et al. demon-strated that CD137/4–1BB and CD28-mediated costimula-tory signals share downstream components such asmembers of the PI3 kinase and AKT/PKB signaling path-way, which induced inhibition of AICD (Starck et al.2005). In this HCC model, 4-1BBL cooperated with B7-1and B7-2 to protect T cells from apoptosis.
T-cell costimulation to NF-�B serves as a paradigm for asignalling system that requires input from the TCR as wellas the CD28 system to be fully activated (Kane et al. 2002).Marinari et al. have recently demonstrated that CD28engagement by B7 can generate TCR-independent signalsleading to I�B kinase (IKK) and NF-�B activation(Marinari et al. 2004). Most T-cell autonomous eVects ofCD28 signalling are under tight regulation of NF-�B as, forexample, the prosurvival function of CD28 involves theNF-�B-dependent upregulation of Bcl-xL22 and the inhibi-tion of p73 expression (Wan and DeGregori 2003). Simi-larly, NF-�B proteins are important for the CD28-mediatedinduction of many chemokines and lymphokines such asIL-2. The interaction of 4-1BBL with 4-1BB providescostimulatory signals to both CD4+ and CD8+ T cellsthrough the activation of NF-�B, c-Jun and p38downstream pathways (Cannons et al. 2000). Data from thepresent study suggest (1) a close association between B7-1,B7-2, and 4-1BBL expression and a strong link between theexpression of B7-2 and 4-1BBL and NF-�B activity in thismodel system, and (2) 4-1BBL synergy with the CD28costimulatory in promoting NF-�B activation.
Conclusions
In the present study, tumor cells were transduced to expressB7-1/B7-2 and/or 4-1BBL. It was demonstrated that B7synergizes with the 4-1BBL costimulatory pathway,thereby inducing an eVective CTL response to tumor anti-gens and amplifying systemic T-cell-mediated antitumorimmunity. Furthermore, results showed that H22/B7-1/B7-2/4-1BBL tumor vaccines probably protect the inWltratinglymphocytes from apoptosis and induce NF-�B activationto improve T-cell-mediated antitumor response. The genetherapy approach taken in this study to test the antitumor
consequences of using B7-1, B7-2 and 4-1BBL gene trans-fer has been shown to have therapeutic potential for HCCtreatment.
Acknowledgments This work was funded in part by National Natu-ral Science Foundation of China (No. 30901442); Natural ScienceFoundation of Jiansu Province (No. BK2009439); and Open Founda-tion from Health Department, Jiangsu Province (No. ZX05 200901).
References
Arch RH, Thompson CB (1998) 4–1BB and Ox40 are members of atumor necrosis factor (TNF)-nerve growth factor receptor sub-family that bind TNF receptor-associated factors and activate nu-clear factor kappaB. Mol Cell Biol 18(1):558–565
Bertram EM, Lau P, Watts TH (2002) Temporal segregation of 4–1BBversus CD28-mediated costimulation: 4–1BB ligand inXuences Tcell numbers late in the primary response and regulates the size ofthe T cell memory response following inXuenza infection. JImmunol 168(8):3777–3785
Bertram EM, Dawicki W, Sedgmen B, Bramson JL, Lynch DH, WattsTH (2004) A switch in costimulation from CD28 to 4–1BB duringprimary versus secondary CD8 T cell response to inXuenza in vi-vo. J Immunol 172(2):981–988
Cannons JL, Choi Y, Watts TH (2000) Role of TNF receptor-associ-ated factor 2 and p38 mitogen-activated protein kinase activationduring 4–1BB-dependent immune response. J Immunol165(11):6193–6204
Croft M (2003) Co-stimulatory members of the TNFR family: keys toeVective T-cell immunity? Nat Rev Immunol 3(8):609–620
Curry HA, Clemens RA, Shah S, Bradbury CM, Botero A, GoswamiP, Gius D (1999) Heat shock inhibits radiation-induced activationof NF-kappaB via inhibition of I-kappaB kinase. J Biol Chem274(33):23061–23067
Dawicki W, Watts TH (2004) Expression and function of 4–1BB dur-ing CD4 versus CD8 T cell responses in vivo. Eur J Immunol34(3):743–751
Disis ML, Bernhard H, JaVee EM (2009) Use of tumour-responsive Tcells as cancer treatment. Lancet 373(9664):673–683
Finney HM, Akbar AN, Lawson AD (2004) Activation of resting hu-man primary T cells with chimeric receptors: costimulation fromCD28, inducible costimulator, CD134, and CD137 in series withsignals from the TCR zeta chain. J Immunol 172(1):104–113
Foreman KE, Wrone-Smith T, Krueger AE, NickoloV BJ (1999)Expression of costimulatory molecules CD80 and/or CD86 by aKaposi’s sarcoma tumor cell line induces diVerential T-cell acti-vation and proliferation. Clin Immunol 91(3):345–353
Fujiwara K, Higashi T, Nouso K, Nakatsukasa H, Kobayashi Y, UemuraM, Nakamura S, Sato S, Hanafusa T, Yumoto Y, Naito I, ShiratoriY (2004) Decreased expression of B7 costimulatory molecules andmajor histocompatibility complex class-I in human hepatocellularcarcinoma. J Gastroenterol Hepatol 19(10):1121–1127
Greenwald RJ, Freeman GJ, Sharpe AH (2005) The B7 family revis-ited. Annu Rev Immunol 23(1):515–548
Guinn BA, DeBenedette MA, Watts TH, Berinstein NL (1999) 4–1BBL cooperates with B7–1 and B7–2 in converting a B cell lym-phoma cell line into a long-lasting antitumor vaccine. J Immunol162(8):5003–5010
Kane LP, Lin J, Weiss A (2002) It’s all Rel-ative: NF-kappaB andCD28 costimulation of T-cell activation. Trends Immunol23(8):413–420
KirchhoV S, Muller WW, Li-Weber M, Krammer PH (2000) Up-regu-lation of c-FLIPshort and reduction of activation-induced cell
123

J Cancer Res Clin Oncol (2011) 137:695–703 703
death in CD28-costimulated human T cells. Eur J Immunol30(10):2765–2774
Li ZS, Yang XW, Chen Z, Dong HL, Ye J, Qu P, Lu SY, Zhang XM,Sui YF (2004) In vivo tumor co-transfection with superantigenand CD80 induces systemic immunity without tolerance and pro-longs survival in mice with hepatocellular carcinoma. CancerBiol Ther 3(7):660–666
Llovet JM, Ricci S, Mazzaferro V, Hilgard P, Gane E, Blanc JF, de Oli-veira AC, Santoro A, Raoul JL, Forner A, Schwartz M, Porta C,Zeuzem S, Bolondi L, Greten TF, Galle PR, Seitz JF, Borbath I,Haussinger D, Giannaris T, Shan M, Moscovici M, Voliotis D,Bruix J (2008) Sorafenib in advanced hepatocellular carcinoma.N Engl J Med 359(4):378–390
Locksley RM, Killeen N, Lenardo MJ (2001) The TNF and TNF recep-tor superfamilies: integrating mammalian biology. Cell104(4):487–501
Marinari B, Costanzo A, Marzano V, Piccolella E, Tuosto L (2004)CD28 delivers a unique signal leading to the selective recruitmentof RelA and p52 NF-kappaB subunits on IL-8 and Bcl-xL genepromoters. Proc Natl Acad Sci U S A 101(16):6098–6103
Martin-Fontecha A, Moro M, Crosti MC, Veglia F, Casorati G, Della-bona P (2000) Vaccination with mouse mammary adenocarci-noma cells coexpressing B7–1 (CD80) and B7–2 (CD86)discloses the dominant eVect of B7–1 in the induction of antitu-mor immunity. J Immunol 164(2):698–704
Melero I, Bach N, Hellstrom KE, AruVo A, Mittler RS, Chen L (1998)AmpliWcation of tumor immunity by gene transfer of the co-stim-ulatory 4–1BB ligand: synergy with the CD28 co-stimulatorypathway. Eur J Immunol 28(3):1116–1121
Melero I, Hervas-Stubbs S, Glennie M, Pardoll DM, Chen L (2007)Immunostimulatory monoclonal antibodies for cancer therapy.Nat Rev Cancer 7(2):95–106
Mogi S, Sakurai J, Kohsaka T, Enomoto S, Yagita H, Okumura K,Azuma M (2000) Tumour rejection by gene transfer of 4–1BBligand into a CD80(+) murine squamous cell carcinoma and therequirements of co-stimulatory molecules on tumour and hostcells. Immunology 101(4):541–547
Muller D, Frey K, Kontermann RE (2008) A novel antibody-4–1BBLfusion protein for targeted costimulation in cancer immunother-apy. J Immunother 31(8):714–722
Okkenhaug K, Wu L, Garza KM, La Rose J, Khoo W, Odermatt B,Mak TW, Ohashi PS, Rottapel R (2001) A point mutation inCD28 distinguishes proliferative signals from survival signals.Nat Immunol 2(4):325–332
Pardoll D (2003) Does the immune system see tumors as foreign orself? Annu Rev Immunol 21(1):807–839
Parkin DM, Bray F, Ferlay J, Pisani P (2005) Global cancer statistics.CA Cancer J Clin 55(2):74–108
Plas DR, Rathmell JC, Thompson CB (2002) Homeostatic control oflymphocyte survival: potential origins and implications. NatImmunol 3(6):515–521
Schultze J, Nadler LM, Gribben JG (1996) B7-mediated costimulationand the immune response. Blood Rev 10(2):111–127
Shuford WW, Klussman K, Tritchler DD, Loo DT, Chalupny J, SiadakAW, Brown TJ, Emswiler J, Raecho H, Larsen CP, Pearson TC,Ledbetter JA, AruVo A, Mittler RS (1997) 4–1BB costimulatorysignals preferentially induce CD8 + T cell proliferation and leadto the ampliWcation in vivo of cytotoxic T cell responses. J ExpMed 186(1):47–55
Starck L, Scholz C, Dorken B, Daniel PT (2005) Costimulation byCD137/4–1BB inhibits T cell apoptosis and induces Bcl-xL andc-FLIP(short) via phosphatidylinositol 3-kinase and AKT/proteinkinase B. Eur J Immunol 35(4):1257–1266
Takahashi C, Mittler RS, Vella AT (1999) Cutting edge: 4–1BB is abona Wde CD8 T cell survival signal. J Immunol 162(9):5037–5040
Tamada K, Chen L (2006) Renewed interest in cancer immunotherapywith the tumor necrosis factor superfamily molecules. CancerImmunol Immunother 55(4):355–362
Tatsumi T, Takehara T, Katayama K, Mochizuki K, Yamamoto M,Kanto T, Sasaki Y, Kasahara A, Hayashi N (1997) Expression ofcostimulatory molecules B7–1 (CD80) and B7–2 (CD86) on hu-man hepatocellular carcinoma. Hepatology 25(5):1108–1114
Wan YY, DeGregori J (2003) The survival of antigen-stimulated Tcells requires NFkappaB-mediated inhibition of p73 expression.Immunity 18(3):331–342
123