transepithelial of system - proceedings of the national ... · proc. natl. acad.sci. usa77(1980)...
TRANSCRIPT

Proc. Natl. Acad. Sci. USA 77 (1980) 6941
Correction. In the article "A theoretical study of the structureof parathyroid hormone" by James E. Zull and Naomi B. Lev,which appeared in the July 1980 issue of Proc. Natl. Acad. Sci.USA (77, 3791-3795), the authors request that the followingcorrections be noted. On p. 3791, a citation of ref. 12 should beinserted at the end of line 8 in the right column. On p. 3792, thecitation in the last line of the left column should be ref. 10. Onp. 3794, the citation in line 12 of the left column should be onlyref. 17, and the beginning of the next line should be Met-8. Inthe reference list on p. 3795, the pages for ref. 8 should be5723-5729.
Correction. In the article "Cytokinin inhibition of respirationin mitochondria from six plant species" by Carlos 0. Miller,which appeared in the August 1980 issue of Proc. Nati. Acad.Sci. USA (77, 4731-4735), there was a printer's error in Table2 on p. 4733. The inhibitions of NADH oxidation by bush beanmitochondria (line 2) have been misplaced: the numbers 4, 13,and 82 should appear under the column headings BAP, SHAM,and Antimycin A, respectively.
Correction. In the article "Interferon inhibits the conversionof 3T3-L1 mouse fibroblasts into adipocytes" by Susan Keayand Sidney E. Grossberg, which appeared in the July 1980 issueof Proc. Nati. Acad. Sci. USA (77,4099-4103), ref. 15, cited inline 32 of Materials and Methods, should be: De Maeyer-Guignard, J., Tovey, M. G., Gresser, I. & De Maeyer, E. (1978)Nature (London) 271, 622-625.
Correction. In the article "Presumptive common precursor forneuronal and glial cell lineages in mouse hypothalamus" by F.De Vitry, R. Picart, C. Jacque, L. Legault, P. Dupouey, and A.Tixier-Vidal, which appeared in the July 1980 issue of Proc.Natl. Acad. Sci. USA (77, 4165-4169), an undetected printer'serror occurred in the legend to Fig. 4 (p. 4168). In the third line,the second 0 (for double-stained cell) should have been *.
Correction. In the article "Transepithelial migration of humanneutrophils: An in vitro model system" by Eva B. Cramer,Linda C. Milks, and George K. Ojakian, which appeared in theJuly 1980 issue of Proc. Natl. Acad. Sci. USA (77, 4069-4073),an error by the Proceedings editorial office appeared on p.4070. The words "scanning electron microscopy" should bedeleted from the first two footnotes (* and t) to Table 1.
Corrections

Proc. Nat!. Acad. Sci. US!AVol. 77, No. 7, pp. 4069-4073, July 1980Cell Biology
Transepithelial migration of human neutrophils: An in vitromodel system
(leukocytes/chemotaxis/cell-cell interaction/intercellular junctions)
EVA B. CRAMER, LINDA C. MILKS, AND GEORGE K. OJAKIANDepartment of Anatomy and Cell Biology, State University of New York, Downstate Medical Center, Brooklyn, New York 11203
Communicated by Aron A. Moscona, February 19, 1980
ABSTRACT An in vitro model system for studying trans-epithelial migration of human neutrophils has been developed.Canine kidney epithelial cells grown on micropore filters forma confluent, polarized monolayer with an average transepi-thelial electrical resistance of 181 ohms-cm2. Neutrophils in achemotactic chamber are stimulated to undergo random mi-gration, chemokinesis, or chemotaxis through the epithelium.When stimulated by a gradient of the synthetic chemoattractantfMet-Leu-Phe, significantly more neutrophils traverse thelow-resistance epithelium than do under conditions of randommigration or chemokinesis. Transmission and scanning electronmicroscopy of this process reveal that neutrophils traverse theepithelium through the intercellular space. After leukocyteemigration, lateral epithelial cell membranes reapproximate.Neutrophils undergoing chemotaxis can also traverse the pola-rized epithelium from the basal epithelial surface, whichsuggests that the chemotactic gradient and not the apical-basalpolarity of the epithelial cells determines the direction oftransepithelial migration. The data further suggest that (i) thein vitro model of leukocyte transepithelial migration morpho-logically simulates the in vivo process, (ii) neutrophils morereadily penetrate the epithelium when attracted by a chemo-tactic factor, and (iii) neutrophils can traverse a low-resistanceepithelium in the absence of serum and connective tissue fac-tors.
Prompt accumulation of neutrophils in tissues invaded bybacteria is essential for the suppression of bacterial growth andinfection. To accomplish this, neutrophils leave the vascularsystem by passing through the endothelium of postcapillaryvenules (1) and, frequently, through a second epithelium (2-4)toward the inflammatory stimuli. However, because this processis difficult to study in vvo, little is known about the restrictionsthat the permeability of the endothelium or other epithelia mayimpose on (i) the directionality or rate of leukocyte migration,(ii) the requirements for serum and connective tissue factors,(iii) the method by which neutrophils traverse occludingjunctions, or (iv) the effects the emigrating neutrophils mayhave on epithelial permeability and intercellular junction dis-assembly and reformation.An in vitro model of transepithelial migration was designed
to answer such questions. This system combines the chemotacticmethod of Boyden (5) with a technique that assesses confluencyand tight junctional permeability of cell monolayers by trans-epithelial electrical resistance measurements (6, 7). At present,only kidney (6, 7) and mammary gland (8) epithelia formzonulae occludentes and produce a measurable transepithelialelectrical resistance when grown in vitro. Since neutrophilstraverse kidney epithelium in response to infection (4) we grewa monolayer of canine kidney epithelial cells (MDCK) on mi-cropore filters (6) and used this epithelium to separate humanneutrophils placed in the upper compartment of a modified
The publication costs of this article were defrayed in part by pagecharge payment. This article must therefore be hereby marked "ad-vertisement" in accordance with 18 U. S. C. §1734 solely to indicatethis fact.
4069
Boyden chemotactic chamber from a synthetic chemoattractantin the lower compartment. Under these conditions, a chemo-tactic gradient is established across the filter and neutrophilsare stimulated to migrate through the confluent, polarized,low-resistance, epithelial monolayer (9, 10).
MATERIALS AND METHODSPreparation of MDCK Monolayer. MDCK originally iso-
lated from a whole adult dog kidney (provided by J. Leighton,Medical College of Pennsylvania, Philadelphia) were main-tained in culture by serial passage in Eagle's medium withEarle's salts, penicillin (100 units/ml), streptomycin (100jg/ml), and 10% (vol/vol) fetal calf serum or were stored inliquid nitrogen after the addition of 10% (vol/vol) dimethylsulfoxide to the culture medium. The cells were harvested with0.25% trypsin/2 mM EDTA in Dulbecco's phosphate-bufferedsaline (Ca2+- and Mg2+-free), plated (1-2 X 105 cells per ml)on 13-mm-diameter micropore filters (6), and grown in a 95%air/5% CO2 atmosphere for 2-3 days at 370C. All tissue culturematerials were from GIBCO.
Transepithelial Electrical Resistance. Transepithelialelectrical resistance of the monolayers grown on the microporefilters was measured prior to all experiments. The filters weremounted between the two halves of a Ussing chamber (exposing0.32 cm2 of the monolayer), and the transepithelial electricalresistance was measured at room temperature in the modifiedEagle's medium containing penicillin and streptomycin (6,7).
Preparation of Neutrophils. Citrated venous blood fromnormal human volunteers was separated into a granulocyte-richfraction by Hypaque/Ficoll (11) and dextran-sedimentationtechniques (12). Residual erythrocytes were eliminated byhypotonic lysis in 0.2% sodium chloride. This resulted in a cellfraction containing approximately 98% neutrophils with 98-99% viability as determined from trypan blue dye exclusion.Prior to use, neutrophils were suspended (5 X 106 cells per ml)in Gey's balanced salt solution (Microbiological Associates,Bethesda, MD) containing 2% (wt/vol) bovine serum albumin,penicillin (63 units per ml), streptomycin (138 ,ug/ml). Al-though Gey's solution with 2% albumin was used in most ex-periments, studies also were performed in Hanks' balanced saltsolution without phenol red (GIBCO) or in Gey's solutionwithout albumin.
Transepithelial Neutrophil Migration. To study transepi-thelial migration from the apical epithelial surface, MDCKmonolayers were grown on Millipore filters (cellulose acetateand nitrate; 0.45-,um pore) or Sartorius filters (cellulose nitrate;3.0-Am pore). The filters were washed in Gey's solution with2% albumin and then were positioned to face the neutrophilsin the upper compartment of a blind well or Boyden chemo-
Abbreviation: MDCK, canine kidney epithelial cells.
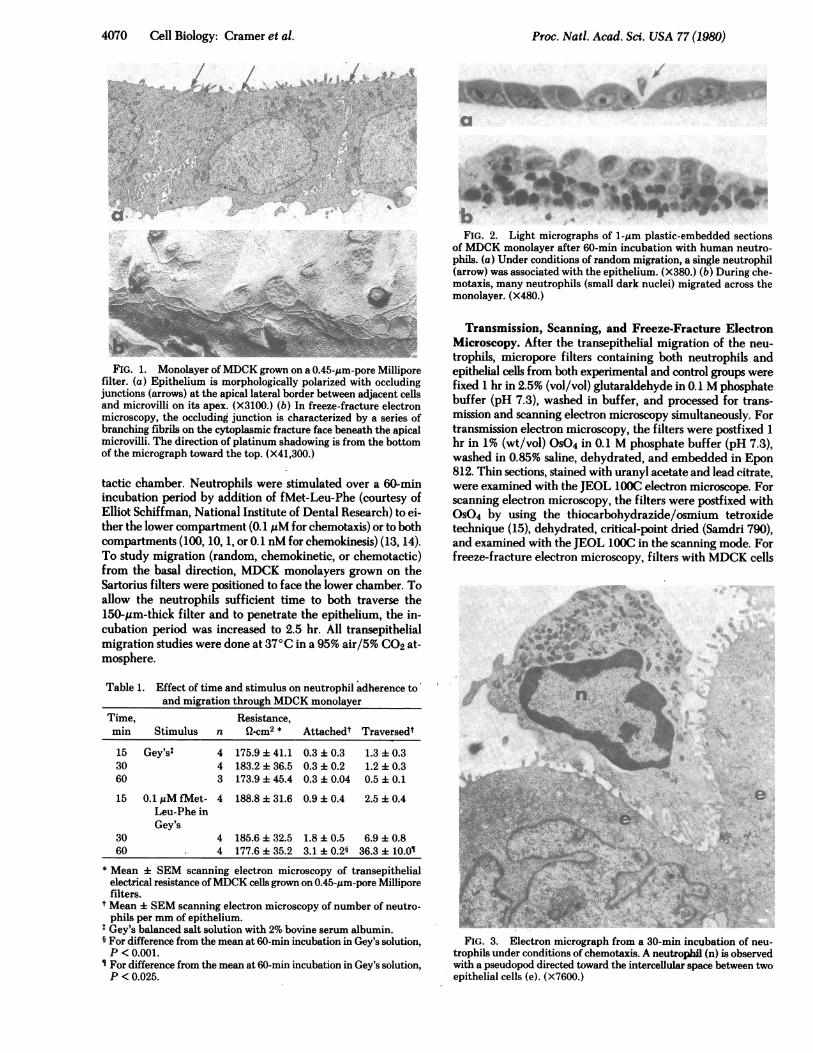
4070 Cell Biology: Cramer et al.
a
FIG. 1. Monolayer ofMDCK grown on a 0.45-,um-pore Milliporefilter. (a) Epithelium is morphologically polarized with occludingjunctions (arrows) at the apical lateral border between adjacent cellsand microvilli on its apex. (X3100.) (b) In freeze-fracture electronmicroscopy, the occluding junction is characterized by a series ofbranching fibrils on the cytoplasmic fracture face beneath the apicalmicrovilli. The direction of platinum shadowing is from the bottomof the micrograph toward the top. (X41,300.)
tactic chamber. Neutrophils were stimulated over a 60-minincubation period by addition of fMet-Leu-Phe (courtesy ofElliot Schiffman, National Institute of Dental Research) to ei-ther the lower compartment (0.1 ,uM for chemotaxis) or to bothcompartments (100, 10, 1, or 0.1 nM for chemokinesis) (13, 14).To study migration (random, chemokinetic, or chemotactic)from the basal direction, MDCK monolayers grown on theSartorius filters were positioned to face the lower chamber. Toallow the neutrophils sufficient time to both traverse the150-tsm-thick filter and to penetrate the epithelium, the in-cubation period was increased to 2.5 hr. All transepithelialmigration studies were done at 370C in a 95% air/5% CO2 at-mosphere.
a
FIG. 2. Light micrographs of 1-um plastic-embedded sections
of MDCK monolayer after 60-min incubation with human neutro-phils. (a) Under conditions of random migration, a single neutrophil(arrow) was associated with the epithelium. (X380.) (b) During che-motaxis, many neutrophils (small dark nuclei) migrated across themonolayer. (X480.)
Transmission, Scanning, and Freeze-Fracture ElectronMicroscopy. After the transepithelial migration of the neu-
trophils, micropore filters containing both neutrophils andepithelial cells from both experimental and control groups werefixed 1 hr in 2.5% (vol/vol) glutaraldehyde in 0.1 M phosphatebuffer (pH 7.3), washed in buffer, and processed for trans-mission and scanning electron microscopy simultaneously. Fortransmission electron microscopy, the filters were postfixed 1hr in 1% (wt/vol) OS04 in 0.1 M phosphate buffer (pH 7.3),washed in 0.85% saline, dehydrated, and embedded in Epon812. Thin sections, stained with uranyl acetate and lead citrate,were examined with the JEOL 10OC electron microscope. Forscanning electron microscopy, the filters were postfixed withOS04 by using the thiocarbohydrazide/osmium tetroxidetechnique (15), dehydrated, critical-point dried (Samdri 790),and examined with the JEOL 100C in the scanning mode. Forfreeze-fracture electron microscopy, filters with MDCK cells
i* sil
i,
.0
W
Table 1. Effect of time and stimulus on neutrophil adherence toand migration through MDCK monolayer
Time, Resistance,min Stimulus n Q-cm2 * Attachedt Traversedt
15 Gey'st 4 175.9 + 41.1 0.3 + 0.3 1.3 + 0.330 4 183.2 + 36.5 0.3 + 0.2 1.2 ± 0.360 3 173.9 + 45.4 0.3 i 0.04 0.5 ± 0.1
15 0.1 ,tM fMet- 4 188.8 + 31.6 0.9 + 0.4 2.5 ± 0.4Leu-Phe inGey's
30 4 185.6 : 32.5 1.8 + 0.5 6.9 d 0.860 4 177.6 : 35.2 3.1 0.2§ 36.3 : 10.01
* Mean + SEM scanning electron microscopy of transepithelialelectrical resistance ofMDCK cells grown on 0.45-,um-pore Milliporefilters.
t Mean + SEM scanning electron microscopy of number of neutro-phils per mm of epithelium.Gey's balanced salt solution with 2% bovine serum albumin.
§ For difference from the mean at 60-min incubation in Gey's solution,P<0.001.
1 For difference from the mean at 60-min incubation in Gey's solution,P < 0.025.
e
KS ..
..
FIG. 3. Electron micrograph from a 30-min incubation of neu-trophils under conditions of chemotaxis. A neutrophil (n) is observedwith a pseudopod directed toward the intercellular space between twoepithelial cells (e). (X7600.)
Proc. Natl. Acad. Sci. USA 77 (1980)
1,704., W
14
Ir",
'.. ...e_:W
...i..r,
.00, ..
,ip ;., ."St .'T,- -:,q.
0%, ,
ON
ow
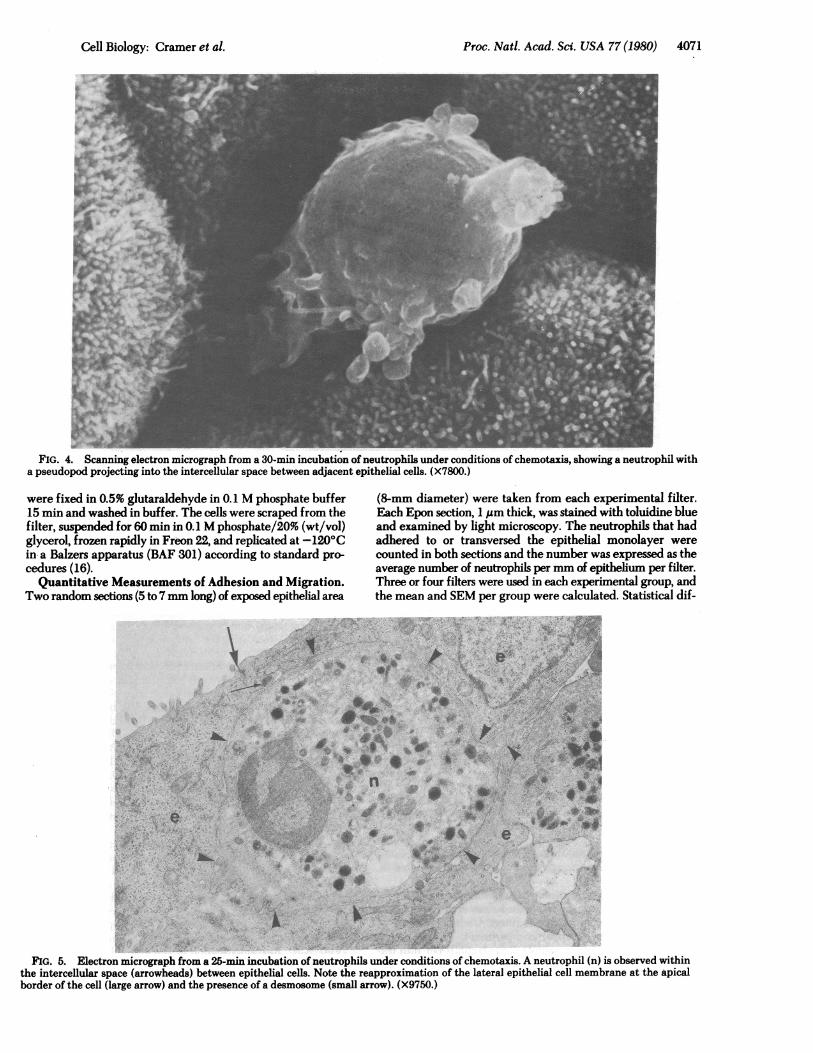
Proc. Natl. Acad. Sci. USA 77 (1980)
FIG. 4. Scanning electron micrograph from a 30-min incubation of neutrophils under conditions of chemotaxis, showing a neutrophil witha pseudopod projecting into the intercellular space between adjacent epithelial cells. (X7800.)
were fixed in 0.5% glutaraldehyde in 0.1 M phosphate buffer15 min and washed in buffer. The cells were scraped from thefilter, suspended for 60 min in 0.1 M phosphate/20% (wt/vol)glycerol, frozen rapidly in Freon 22, and replicated at -1200Cin. a Balzers apparatus (BAF 301) according to standard pro-cedures (16).
Quantitative Measurements of Adhesion and Migration.Two random sections (5 to 7 mm long) of exposed epithelial area
'44Aa
4bi.. *
'A Se, II
e.4
4k*
(8-mm diameter) were taken from each experimental filter.Each Epon section, 1 um thick, was stained with toluidine blueand examined by light microscopy. The neutrophils that hadadhered to or transversed the epithelial monolayer werecounted in both sections and the number was expressed as theaverage number of neutrophils permm of epithelium per filter.Three or four filters were used in each experimental group, andthe mean and SEM per group were calculated. Statistical dif-
Zksr-ffkft; Y
#1a*
s 's d'9s_
Xk~~~~~~~~~~~~A'i :j 9,2*;.-)its'IA
aO ~ **9 e./ *
I;I
"
FIG. 5. Electron micrograph from a 25-min incubation of neutrophils under conditions of chemotaxis. A neutrophil (n) is observed withinthe intercellular space (arrowheads) between epithelial cells. Note the reapproximation of the lateral epithelial cell membrane at the apicalborder of the cell (large arrow) and the presence of a desmosome (small arrow). (X9750.)
Cell Biology: Cramer et al. 4071
.Oi# .r

4072 Cell Biology: Cramer et al.
.,
q., * ,J
a
bya
ibid
b40".'4
P60 AI~MO ~ ~ ~ _Ii
/
"t~ n
C..Aea
e
- I&,
n
ferences between groups were determined by Student's ttest.
RESULTSEpithelial Monolayer. Two to 3 days after plating, the
MDCK had formed a confluent monolayer on the upper surfaceof both the 0.45-Aim-pore Millipore and 3.0-,um-pore Sartoriusfilters. The epithelium was morphologically polarized withnumerous microvilli and belt-like occluding intercellularjunctions at the apical border of the cells (Fig. la). The baso-lateral regions of adjacent cells had extensive interdigitationsheld together by occasional desmosomes. Freeze-fractureelectron microscopy confirmed the presence of zonulae oc-cludentes at the apical border of the intercellular space (Fig.lb). The transepithelial electrical resistance of the epithelialmonolayers used in this study ranged from 64 to 262 ohms-cm2,with an average of 181 ohms-cm2. Epithelia with transepithelialelectrical resistances >300 ohms cm2 were not utilized becauseneutrophil migration through these monolayers is impeded (9,10).
Transepithelial Migration. Neutrophils, when stimulated,were quite capable of penetrating the kidney epithelium grownon the Millipore or the Sartorius filters. Under conditions ofchemotaxis (buffer in upper compartment, 100 nM fMet-Leu-Phe in lower compartment), 7 neutrophils per mm tra-versed the low-resistance epithelium in 30 min and 36 neu-trophils permm in 60 min (Fig. 2b; Table 1). However, under
FIG. 6. Neutrophils stimu-lated to migrate through a 3.0-,um-pore Sartorius filter towardthe basal surface of the epithelialmonolayer. (a) Under conditions ofchemotaxis, many neutrophilshave traversed the filter and someare penetrating the epithelium(arrow). (X200.) (b) Higher mag-nification of the area in the rec-tangle in a (X1000.) (c) Neutro-phils (n) migrating between twoepithelial cells (e). (X5400.)
conditions of random migration (buffer in both compartments),one neutrophil per mm migrated through the epithelium in 30min (Table 1). Increasing the incubation time to 60 min did notincrease the number of neutrophils traversing the epithelium.After 60 min under conditions of chemokinesis (100, 10, 1, or0.1 nM fMet-Leu-Phe in both compartments), there was nosignificant increase in neutrophil penetration of the monolayer.Neither the type of buffer. (Gey's or Hanks' balanced salt so-lutions) nor the presence of albumin affected neutrophil ad-herence or ability to penetrate and traverse the epithelium.Under conditions of random migration, neutrophils adhered
to the surface of plain Millipore filters (98 cells per mm), butrelatively few neutrophils (0.3 cells per mm) attached to thesurface of the epithelium after 60 min of incubation. Duringchemokinesis, the number of cells adhering to the epithelialsurface did not change, whereas during chemotaxis it signifi-cantly increased to three cells permm (Table 1). This remainslow in comparison to adherence to plain filters.
Transmission and scanning electron microscopy of the mo-nolayer revealed that the neutrophils attached primarily to thetips of the epithelial microvilli. However, in areas where mi-crovilli were scarce or at times when neutrophils were passingbetween epithelial cells, neutrophils made contact with theepithelial surface. During transepithelial' migration the neu-trophils traversed the epithelium through the intercellular space(Figs. 3-5). Reapproximation of the apical lateral epithelialmembranes was observed following neutrophil migration (Fig.5).
Proc. Natl. Acad. Sci. USA 77 (1980)
_XI it"
"
.AA,-1;,
IN O.- I;.,%
c Ad6,&-

Proc. Nati. Acad. Scz. USA 77 (1980) 4073
The ability of the neutrophils to migrate across the polarizedepithelium from the basal direction also was examined underconditions of random migration, chemokinesis, and chemotaxis.After 2.5 hr of incubation, less than one neutrophil per mm wasobserved traversing the epithelium under conditions of randommigration or chemokinesis. In contrast, when attracted by achemotactic gradient significantly, more neutrophils (23 cellsper mm) were observed penetrating the epithelium (Fig. 6).
DISCUSSIONThe use of an in vitro model system of transepithelial migrationhas several advantages over in vivo studies in that (i) humanneutrophils can be studied, (ii) the composition of the fluid onboth sides of the epithelium can be controlled to permit a sys-tematic analysis of the effects of serum and connective tissuefactors, (iii) the type of chemoattractant can be varied and theconcentration can be controlled, and (iv) various aspects of themigration process can be examined that are not possible to studyin vivo, such as the effect of epithelial permeability on neu-trophil emigration.The in vitro migration of neutrophils across an epithelium
simulates the in vivo phenomenon. In both situations, leukocytesinitially adhere to the epithelial cell surface (1, 17-19) and passbetween epithelial cells (1, 20-22). After leukocyte emigration,the epithelial tight junctions appear to re-form (21).
It seems likely that, in this in vitro system, neutrophils tra-verse intercellular tight junctions. Our results, as well as thoseof others (6, 7), demonstrate that MDCK form a confluentmonolayer with apical occluding junctions. Observations ofneutrophils either projecting pseudopods into the apical regionof the intercellular space or enclosed within the intercellularspace support the premise that neutrophils migrate betweenepithelial cells. Therefore, it is reasonable to assume that whenneutrophils traverse the monolayer they penetrate occludingjunctions.The results of this study indicate that the invasive ability of
neutrophils is stimulated by a chemotactic gradient. Althoughserum and connective tissue components such as histamine (23,24), kinins (25), 3a and 5a components of complement (26, 27),and prostaglandins E1 and E2 (28) may influence the perme-ability of the epithelium, neutrophils are capable of penetratingthe epithelium in the absence of these factors. In fact, duringthe inflammatory process, vascular permeability and leukocyteemigration appear to be separate phenomena (1, 25, 29, 30).
Emigration of leukocytes from venules during inflammationseems to be primarily unidirectional, because reverse emigra-tion in vivo is only occasionally observed for macrophages andrarely for neutrophils (17). Despite the structural and functionalpolarity of epithelia in general (31), and dog kidney tubules (32)and MDCK (6, 7, 33) in particular, it appears that migrationof human neutrophils across an MDCK monolayer from bothapical and basal directions is determined by the orientation ofthe chemotactic gradient, not by the polarity of the epithelialcell.
Although leukocyte emigration is of primary importance inthe inflammatory process, a clear understanding of the struc-tural, biochemical, and physiological aspects of this phenom-enon has not been attained. The in vitro model we have de-veloped provides a simple, precisely controlled system to in-vestigate the normal dynamics of this process and a means todetect and study impaired neutrophil emigration. In addition,this model is sufficiently flexible to enable the study of varioustypes of leukocytes and epithelia, including endothelium. Inthe latter case, the system also could be modified to study the
We thank Dr. Alice Deutsch, Dr. Thomas Easton, and Dr. DonaldFischman for their critical reading of the manuscript. We also ac-knowledge the expert technical assistance of Ms. Claire Lipp, thephotographic assistance of Mr. Jack Illari and Mr. Louis Dienes, andthe secretarial assistance of Mrs. Lynn Abott, Mrs. Ruth Cintron, andMrs. Ruth Ginsberg. We are grateful to Dr. Saiyid Shafiq for the useof his Balzars apparatus. These studies were supported in part by theNational Institutes of Health Contract 263-79-G-002206 and GrantGM25125.
1. Grant, L. (1973) in The Inflammatory Process, eds. Zeifach, B.,Grant, L. & McCluskey, R. (Academic, New. York), Vol. 2, pp.205-249.
2. Phillips, D. & Mahler, S. (1977) Anat. Rec. 189,45-0.3. Seelig, L. & Beer, A. (1978) Biol. Reprod. 18, 736-744.4. Zollinger, H. & Mihatsch, M. (1978) Renal Pathology in Biopsy,
(Springer, New York), pp. 424-437.5. Boyden, S. (1962) J. Exp. Med. 115,453-466.6. Misfeldt, D., Hamamoto, S. & Pitelka, D. (1976) Proc. Natl. Acad.
Sci. USA 73, 1212-1216.7. Cereijido, M., Robbins, E., Dolan, W., Rotunno, C. & Sabatini,
D. (1978) J. Cell Biol. 77, 853-880.8. Bisbee, C., Machen, T. & Bern, H. (1979) Proc. Natl. Acad. Sci.
USA 76, 536-540.9. Cramer, E., Milks, L. & Ojakian, G. (1979) Fed. Proc. Fed. Am.
Soc. Exp. Biol. 38,1085 (abstr.).10. Cramer, E., Milks, L. & Ojakian, G. (1979) in The Cytoskeleton:
Membranes and Movement, eds. Condeelis, J., Satir, P. & Bur-ridge, K. (Cold Spring Harbor Laboratory, Cold Spring Harbor,NY), p. 142 (abstr.).
11. Boyum, A. (1968) Scand. J. Clin. Lab. Invest. (Suppl.) 21, 77-89.
12. Clark, R. & Kimball, H. (1971) J. Clin. Invest. 50,2645-2652.13. Schiffman, E., Corcoran, B. & Wahl, S. (1975) Proc. Natl. Acad.
Sci. USA 72,1059-1062.14. Cramer, E. & Gallin, J. (1979) J. Cell Biol. 82,369-379.15. Malick, L. & Wilson, R. (1975) in Scanning Electron Micros-
copy/1975, eds. Johari, 0. & Corvin, I. (IIT Research Institute,Chicago), pp. 259-266.
16. Moor, H. & Muhlethaler, K. (1963) J. Cell Biol. 17, 609-628.17. Clark, E., Clark, E. & Rex, R. (1936) Am. J. Anat. 59, 123-
173.18. Atherton, A. & Born, G. (1972) J. Phystol. (London) 222, 447-
474.19. Anderson, A. & Anderson, N. (1976) Immunology 31, 731-
748.20. Schoefl, G. I. (1972) J. Exp. Med. 136,568-588.21. Hayashi, H., Yoshinaga, M. & Yamamoto, S. (1974) Antibiot. &
Chemother. (Basel) 19,296-32.22. Wenk, E. J., Orlic, D., Reith, E. J. & Rhodin, J. A. G. (1974) J.
Ultrastruct. Res. 46,214-241.23. Riley, J. & West, G. (1953) J. Physiol. (London) 120,528-537.24. Graham, H., Lowry, O., Wheelwright, F. & Lenz, M. (1955)
Blood 10, 467-481.25. Wilhelm, D. (1973) in The Inflammatory Process, eds. Zeifach,
B., Grant, L. & McCluskey, R. (Academic, New York), Vol. 2, pp.
251-301.26. Dias DaSilva, W. & Lepow, I. (1967) J. Exp. Med. 125, 921-
946.27. Cochrane, C. & Muller-Eberhard, H. (1968) J. Exp. Med. 127,
371-386.28. Crunckhorn, P. & Willis, A. (1971) Br. J. Pharmacol. 41, 49-
56.29. Hurley, J. (1963) Aust. J. Exp. Biol. Med. Sci. 41, 171-186.30. Logan, G. & Wilhelm, D. (1963) Nature (London) 198, 968-
969.31. Ernst, S. A. & Mills, J. W. (1977) J. Cell Biol. 75, 74-94.32. Kyte, J. (1976) J. Cell Biol. 68, 287-303.33. Rodriguez-Boulan, E. & Sabatini, D. D. (1978) Proc. Natl. Acad.
mechanisms of invasion and metastasis of malignant cells.
Cell Biology: Cramer et al.
Sci. USA 75,5071-5075.