to elucidate pharmacological profile of bioactive...
TRANSCRIPT

To Elucidate Pharmacological Profile of Bioactive Principles of some Himalayan
Medicinal and Aromatic Plants
A Thesis Submitted for the Degree of DOCTOR OF PHILOSOPHY
in
CHEMISTRY KUMAUN UNIVERSITY
2009
by
SANGEETA PILKHWAL M. Pharm
DEPARTMENT OF PHARMACY
KUMAUN UNIVERSITY, NAINITAL UTTARAKHAND

CC EE RR TT II FF II CC AA TT EE
This is to certify that the work incorporated in this thesis
entitled “To Elucidate Pharmacological Profile of Bioactive Principles
of some Himalayan Medicinal and Aromatic Plants” has been carried
out by Mrs. Sangeeta Pilkhwal, under our supervision. She has fulfilled
the requirement of prescribed period for the Degree of Doctor of
Philosophy in Chemistry of Kumaun University, Nainital.
Unless stated otherwise, the work included in this thesis is
original and has not been submitted for any other degree.
DR. KANWALJIT CHOPRA PROF. C. S. MATHELA Co-supervisor Supervisor Head, Pharmacology Division Professor & Head UIPS, Panjab University Department of Chemistry Chandigarh. Kumaun University, Nainital.

DDeeddiiccaatteedd TToo
MMyy FFaammiillyy MMeemmbbeerrss

AACCKKNNOOWWLLEEDDGGEEMMEENNTT
As I sit back to write this little piece, I think of the innumerable hands that
have steered me through this arduous journey and put me on the right path of
learning and enlightment.
At the outset, I deem it my prime duty and obligation to express my
sagacious sense of gratitude and indebtedness to my esteemed teacher and guide
PPrrooffeessssoorr CC.. SS.. MMaatthheellaa,, HHeeaadd,, DDeeppaarrttmmeenntt ooff CChheemmiissttrryy,, for gracious initiation,
valuable guidance, congenial discussion and constant encouragement throughout the
course of this study. My inner conviction does not only allow me to express heartfelt
gratitude but something else which is inexpressible for my co guide Dr.((MMrrss..))
KKaannwwaalljjiitt CChhoopprraa, Head, Pharmacology Division, University Institute of
Pharmaceutical Sciences, Panjab University, Chandigarh. Her inspiration, moral
support and valuable suggestions goes a long way in the successful execution of the
research protocols.
I wish to express my sincerest thanks to PPrrooff.. KKaattaarree,, Chairman, University
Institute of Pharmaceutical Sciences, Panjab University, Chandigarh, for providing
the necessary facilities for pharmacological studies. I am also thankful to PPrrooff.. SS.. KK..
KKuullkkaarrnnii for his generous help and constant encouragement.
Words fail to express my feelings towards my B. Pharm teachers MMrr.. SS..CC..
SShhaarrmmaa who has always been a great source of inspiration for me. I wish to express
my sincere thanks to Prof. Vijay JJuuyyaall aanndd Mr. B.K. Singh, Reader in Department
of Pharmacy for tthheeiirr ccoonnssttaanntt hheellpp aanndd ssuuppppoorrtt dduurriinngg mmyy wwoorrkk
I am elated with delight to express my deep sense of gratitude to all my
seniors and colleagues especially DDrr.. KKuummuudd UUppaaddhhaayyaayy,, DDrr.. AAnniittaa SSiinngghh,, MMrrss..
DDiivvyyaa JJuuyyaall,, MMrrss.. AArrcchhaannaa NNeeggii SSaahh,, MMrr.. MMaahheennddrraa SSiinngghh RRaannaa,, MMrr.. TTiirraatthh
KKuummaarr,, MMrr.. RRaajjeesshhwwaarr KKaammaallkkaanntt who have always lend a helping hand during the
hours of need.

Batch mates and friends are always the pacemakers of life as they polish you
in proper shape of personality. It is my pleasure to acknowledge my special thanks to
my friends and classmates AAnnuurraagg,, MMaahheennddrraa aanndd AAsshhiisshh for their constant
cooperation, day to day help, love, affection and care.
I would like to extend my heartiest gratitude to DDrr.. RRaajjeennddrraa PPaaddaalliiyyaa and
other co-workers for GC-MS screening of the samples and help in chemical
investigations and spectral interpretation.
I am also thankful to Dr. L.S. Rautela, Lab technician, Department of
Pharmacy and others in nonteaching staff MMrr.. GG..CC.. BBhhaatttt, MMrr.. UUmmeesshh,, MMrr..
JJaaggddiisshh,, MMrr.. MMaanniisshh aanndd MMrr.. BBhhuuppeennddrraa for their help and support throughout the
project duration. I also thank, Mr. S.S. Mehra and Mr. Mukesh Chandra in the
library staff for their help and support.
I also express my special thanks to Ms. Maninder Kaur of CPEBS and Mr.
Kiran of Pharmacology division, UIPS, Panjab University, Chandigarh for their
help and cooperation. I would also like to acknowledge the laboratory staff of
Pharmacology division of UIPS, Panjab University for their help and support.
It is beyond my capacity to express my gratitude to my people and my family
who are pillars of my strength. My vocabulary fails in expressing my indebtness to
all my family members for their unending love, inspiration and moral support. I wish
to reciprocate the feeling of thanks to my husband MMrr.. MMuukkeesshh LLaall ssaahh who has
always encouraged me with constant help and valuable suggestions throughout the
study.
I find myself devoid of words in my lexicon to acknowledge my indebtness to
MMuummmmyy,, PPaappaa aanndd mmyy mmootthheerr--iinn--llaaww for their love, affection, inspiration,
unquestionable patience, encouragement and unparallel sacrifice.
I thank all beloved and respected people around me who directly and
indirectly helped me during my degree program whose names could not be mentioned
here.
AApprriill 22000099 SSAANNGGEEEETTAA PPIILLKKHHWWAALL

ABBREVIATIONS
A1, A2A Adenosine receptors
ATP Adenosine triphosphate
BDZs Benzodiazepines
BHA Butylated hydroxyanisole
BHT Butylated hydroxytoluene
CBR Conditioned behavior response
cGMP Cyclic guanosine monophosphate
COX Cyclooxygenase
DCM Dichloromethane
DNA Deoxyribose nucleic acid
DPPH 1,1 diphenyl 2 picryl hydrazyl
ECG Electrocardiogarm
ED50 Median lethal concentration
EEG Electroencephalogram
EPM Elevated plus maze
FST Forced swim test
GABA Gamma amino butyric acid
GC-MS Gas chromatography-Mass spectroscopy
5HT 5 Hydroxytryptamine
GSH Reduced glutathione
H-NMR Proton nuclear magnetic resonance
HPLC High performance liquid chromatography
HPTLC High performance thin layer chromatography
IL Interleukin
iNOS Inducible nitric oxide synthase
IR Infrared
IVHD Isovaleroxy hydroxyl didrovaltrate
LC50 Median lethal concentration
L-NAME N-omega-nitro-l-arginine methyl ester

LPS Lipopolysaccharide
LSD Lysergic acid diethylamine
MDA Malondialdehyde
MES Maximal electro shock seizures
ML Melatonin
MPO Myeloperoxidase
NBT Nitrobluetetrazolium
NF-κB Nuclear factor kappa B
NMDA N-methyl-D –aspartate
NMR Nuclear magnetic resonance
NO Nitric oxide
NOS Nitric oxide synthase
NSAIDs Non steroidal anti-inflammatory drugs
OD Optical density
PMS Post mitochondrial supernatant
PTZ Pentylenetetrazol
Rf Retention factor
ROS Reactive oxygen species
SOD Superoxide dismutase
TLC Thin layer chromatography

CONTENTS Introduction................................................................................... Aims and objectives.................................................................................... CHAPTER 1
Review of literature.................................................................................... CHAPTER 2 Chemical screening of Valeriana wallichii collected from different regions of Kumaun Himalaya
Section A: Isolation and identification of constituents from Valeriana wallichii essential oils 2.1.a. Introduction …………….......................................................... 2.2.a. Material and methods............................................................... 2.3.a. Results..................................................................................... Section B: Isolation and analysis of valepotriates of Valeriana wallichii chemotypes 2.1.b. Introduction………………………………………………………………… 2.2.b Material and methods……………………………………………………… 2.3.b Results…………………………………………………………………………
CHAPTER 3
Psychopharmacological profile of Valeriana wallichii chemotypes
3.1. Introduction …………………………………………………………. 3.2. Material and methods ………………………………………………. 3.3. Results ………………………………………………………………………. 3.4. Discussion ...…………………………………………………………
CHAPTER 4
Screening of Valeriana wallichii chemotypes for antidepressant effect
4.1. Introduction ………………………………………………………… 4.2. Material and methods ……………………………………………… 4.3. Results ……………………………………………………………… 4.4. Discussion …………………………………………………………..
CHAPTER 5

Pharmacological and neurobiochemical evidence for antidepressant like effect of Valeriana wallichii chemotypes
5.1. Introduction ………………………………………………………… 5.2. Material and methods ……………………………………………… 5.3. Results ……………………………………………………………… 5.4. Discussion …………………………………………………………..
CHAPTER 6
Studies on analgesic activity of Valeriana wallichii chemotypes
6.1. Introduction ……………………………………………………… 6.2. Material and methods …………………………………………… 6.3. Procedure ………………………………………………………… 6.4. Results …………………………………………………………… 6.5 Discussion………………………………………………………………..
CHAPTER 7
Evaluation of in vitro antioxidant profile of Valeriana wallichii chemotypes
7.1. Introduction ……………………………………………………… 7.2. Material and methods …………………………………………… 7.3. In vitro antioxidant assays ………………………………………. 7.4. Results …………………………………………………………… 7.5. Discussion ………………………………………………………..
CHAPTER 8
Modulation of antioxidant defense system in mice brain by Valeriana wallichii chemotypes
8.1. Introduction ………………………………………………………. 8.2. Material and methods ……………………………………………. 8.3. Results ……………………………………………………………. 8.4. Discussion …………………………………………………………
Conclusion ……………………………………………………………………. Bibliography ………………………………………………………………….

INTRODUCTION
Medicinal plants are the nature’s gift to human being to make disease free
healthy life. India is one of the most medico culturally diverse country in the world
where the medicinal plant sector is part of a time-honored tradition that is respected
even today. Medicinal plants are believed to be much safer and in our country more
than two thousand medicinal plants are recognized. Chemical and biological diversity
of aromatic and medicinal plants depending on factors, such as cultivation area,
climatic conditions, vegetation phase, genetic modifications and others is an important
impetus to study flora present in different growing sites, countries and geographical
zones. A great number of aromatic, spicy and other plants contain chemical
compounds exhibiting antioxidant and psychopharmacological properties. Numerous
studies were carried out on some of these plants, e.g. rosemary, sage, oregano, which
resulted in a development of natural antioxidant formulations for food, cosmetic and
other applications. However, scientific information on biological properties of various
plants, particularly those that are less widely used in culinary and medicine, is still
rather scarce. Therefore, the assessment of such properties remains an interesting and
useful task, particularly for finding new sources for natural antioxidants, functional
foods and neutraceuticals. Besides this, efficacy of medicinal plants is more evident in
problems related to the nervous system; as stress, anxiety, tension and depression are
intimately connected with most illnesses.
Valerianaceae is a family of about 13 genera and more or less 400 species,
chiefly confined to temperate regions with exception of Andes of South America.
Plants under this genus are mostly herbs or shrubs, annual to perennial; with or
without basal aggregation of leaves. Many members of the Valerianaceae have a
distinctive odor due to the presence of valerianic acid and its derivatives. Several
species have medicinal properties, and root and leaf extracts are used in treating nerve
complaints. A few species are used for perfumes and dyes. The major constituent
present in Valeriana is iridoids. Iridoids are found in many medicinal plants and may
be responsible for some of their pharmaceutical activities. Isolated and purified,
iridoids exhibit a wide range of bioactivities including cardiovascular,

antiphepatotoxic, chlorectic, hypoglycemic, anti-inflammatory, antispasmodic,
antitumor, antiviral, immunomodulator and purgative (Didna et al., 2007). A
considerable number of investigations on Valeriana species have yielded other
compounds like sesquiterpenoids, lignans and alkaloids with pharmacological
properties, including sedative, cytotoxic, antitumor, antioxidant, and vasorelaxant
(Piccinelli et al., 2004; Thies et al., 1981; Bounthanh et al., 1981).
This work, therefore is aimed at evaluating psychopharmacological profile and
preliminary screening of free radical scavenging activities of the extracts and oils
isolated from Valeraina wallichii chemotypes found in Himalayan region.

AIMS AND OBJECTIVES
Herbal medicine is the oldest and most natural form of medicine whose history
of efficacy and safety spans centuries and covers every country on the planet. Because
herbal medicine is holistic medicine, it is, in fact, able to look beyond the symptoms
to the underlying systemic imbalance when skillfully applied by the trained
practitioner. Herbal medicine offers very real and permanent solution to problems,
many of them seemingly intractable to pharmaceutical intervention. The efficacy of
herbal medicine is more evident in problems related to the nervous system as stress,
anxiety, tension and depression are intimately connected with most illness. So there is
great scope for the development of herbal medicine in the area of nervous diseases
and of its application in mental illness. Another area where medicines from herb can
be used is in the treatment of disorders caused by oxidative stress. The majority of
disease conditions like atherosclerosis, hypertension, ischemic diseases, Alzheimer’s
disease, parkinsonism, cancer, diabetes mellitus and inflammatory conditions are
considered to be primarily due to the imbalance between prooxidant and antioxidant
homeostasis. Antioxidant principles from natural resources possess multifacetedness
in their magnitude of activities and provide enormous scope in correcting the
imbalance. Hence, there is no doubt that phytochemicals deserve a proper position in
the therapeutic armamentarium.
Herbal medicine can justifiably boast of Valeriana, the ideal "tranquillizer".
Valeriana is an important medicinal genus in many Pharmacopoeial systems. The
Valeriana wallichii DC. has long been used in Ayurveda (Charak Samhita, Susruta
Samhita) and Unani systems of medicine. The rhizomes of this plant contain volatile
oil (which includes valerianic acid), volatile alkaloids (including chatinine), and
iridoids (valepotriates) which have been shown to reduce anxiety and aggression and
even to counteract the effects of ethanol. Valeriana is effective in treating anxiety
while maintaining normal mental awareness as it enables the patient to continue the
most complicated mental exercise without drowsiness, loss of consciousness or
depression.

Thus the aim of the present study was to compare the
psychoneuropharmacological and antioxidant effects of Valeriana wallichii
chemotypes growing in different areas of Kumaun hills and to correlate it with
interspecific variation in chemical constituents under the following experimental
protocols:
A) Chemical profiling of Valeriana wallichii collected from different regions of
Kumaon:
• Chemical screening for terpenoids.
• Chemical screening for valepotriates.
B) Bioactivity evaluation of Valeriana wallichii extracts and oils:
• Evaluation of psychopharmacological profile.
• Screening of antidepressant effect.
• Pharmacological and neurobiochemical evidence for antidepressant like effect.
• Preliminary studies on the analgesic activity.
• Evaluation of in vitro antioxidant profile.
• Modulation of antioxidant defense system in mice brain.

CHAPTER-1
Review of Literature

1.1. INTRODUCTION
India is one of the 12 mega biodiversity centre having 45,000 plant species
and diversity is unmatched due to 16 different agroclimatic zones, 10 vegetative
zones, and 15 biotic provinces. The country has a rich floral diversity, about 500
plants with medicinal use are mentioned in ancient texts and around 800 plants have
been used in indigenous systems of medicine. Indian subcontinent is a vast repository
of medicinal plants that are used in traditional medical treatments (Chopra et al.,
1956), which also forms a rich source of knowledge. The various indigenous systems
such as Siddha, Ayurveda, Unani and Allopathy use several plant species to treat
different ailments (Rabe and Staden, 1997). In India around 20,000 medicinal plant
species have been recorded recently (Dev, 1997), and more than 500 traditional
communities use about 800 plant species for curing different diseases (Kamboj,
2000). Currently 80% of the world population depends on plant-derived medicine for
the first line of primary health care for human alleviation because it has no side
effects.
In the last century, roughly 121 pharmaceutical products were formulated
based on the traditional knowledge obtained from various sources. The estimation of
total phytomedicine sale reported in country wise European Union was about US $ 6
billion in 1991 and $ 4 billion in 1996, of which almost half were sold in Germany $ 3
billion, in France $ 1.6 billion, in Italy $ 0.6 billion and in Japan $ 1.5 billion. The
present global market is said to be US $ 250 billion. In India the sale of total herbal
products is estimated at $ 1 billion and the export of herbal crude extract is about $ 80
million, of which 50% is contributed by Ayurvedic classical preparations.
A number of scientific investigations have highlighted the importance and the
contribution of many plant families used as medicinal plants and they have played a
vital role for the development of new drugs. Plant derived drugs are used to cure
mental illness, skin diseases, tuberculosis, diabetes, jaundice, hypertension and
cancer. Medicinal plants play an important role in the development of potent
therapeutic agents. Plant derived drugs came into use in the modern medicine through
the uses of plant material as indigenous cure in folklore or traditional systems of
medicine. Determining the biological (activity) properties of plants used in traditional

medicine is helpful to the rural communities and informal settlements. Several
attempts are currently being undertaken to isolate the active compound(s) by
bioassay-guided fractionation from the species that showed high biological activity
during screening. Therefore, these scientific investigations may be utilized to develop
drugs for these diseases. Further research is deserved to isolate the compounds
responsible for the observed biological activity. So the present study also aims at
determining the biological activity of certain plant species found in Himalayan region
with special emphasis on evaluation of psychopharmacological and antioxidant
activity.
1.2. VALERIANA
The genus Valeriana constitutes a group of perennial herbs or undershrubs,
with a short, often strong-smelling rootstock, distributed chiefly in the temperate and
cold region of the northern hemisphere. Several species are however also found in
Andean Chile, and some in Brazil, South Africa, and South and South-east subtropical
Asia. The genus is scarce in the mountain of the tropics and about 12 species occur in
India.
The dried rhizomes and roots of V. jatamansi as well as those of V. officinalis
were official in the earlier editions (pre 1966) of the Pharmacopoeia of India (The
Wealth of India, 2005). In view of pharmaceutical and perfumery value, the Valeriana
species of various global regions have been investigated in the recent years. Three
distinct classes of compounds have been associated with the medicinal properties of
valerian: a) mono- and sesquiterpenes, b) iridoid triesters (valepotriates), and c)
pyridine alkaloids. The composition of the volatile oil varies markedly between
cultivars and species, as does the amount and relative proportion of valepotriates,
making chemical standardization difficult but highly desirable (Moore et al., 2003;
The Wealth of India, 2005).
1.2.1. Historical uses
The valerian oil in large amounts is said to produce dullness of intellect,
drowsiness ending in deep sleep, reduced frequency of pulse and increased urine flow.

It is useful in cases of irregular nervous action, when not connected with
inflammation, or an excited condition of the system. It has also been used in
intermittents, combined with Peruvain bark and in acute rheumatism. It is said that the
virtues of valerian reside chiefly in the volatile oil. By 1923, there was the first
indication that the action of valerian could also act through an odourous pathway
(BPC, 1923). The overall action of valerian rhizome is virtually because of its volatile
oil and valepotriates. It has been used as carminative and antispasmodic in hysteria
and similar nervous manifestations and as the perfect herbal tranquillizer. It was used
for this purpose in the First World War to treat soldiers suffering from shell shock
(Howard, 1987).
1.2.2. Chemical constituents of Valeriana officinalis
The roots of V. officinalis contain several compounds with demonstrable
pharmacological activity. These include the essential oil and its sesquiterpenoids
(valerenic acid), epoxy iridoid esters (valepotriates) and their decomposition products
such as baldrinal and homobaldrinal, amino acids (arginine, GABA, glutamine,
tyrosine) and alkaloids. Valerian also possesses small amounts of phenolic acids,
flavonoids, valerosidatum, chlorogenic acid, caffeic acid, choline, β-sitosterol, fatty
acids, and various minerals (Herbalist, 1999).
1.2.3. Modern uses
Smooth muscle relaxant
Valeriana compounds probably relax stimulated smooth muscle cells by
acting as musculotropic agents and not by interacting with receptors of the autonomic
nervous system (Hazelhoff, 1982). Valeriana officinalis var. latifolia has property to
relieve smooth muscle spasms and is a powerful vasodilator (Yang, 1994). A
preparation of a volatile oil fractionated from the root was used to treat patients with
angina pectoris, among whom ST-T ischemic changes appeared on ECG in 50 cases
before treatment. Its effective rate in simple angina was found to be 87.8% while in
angina with ischemic findings the rate was 88%. The mild myorelaxant action of
valerian is attributed to the valepotriate component of the herb (Dunaev, 1987). The
valepotriates, isovaltrate and valtrate and the essential oil compound valeranone were

observed to suppress the rhythmic contraction in a closed part of the guinea-pig ileum
in vivo (Hazelhoff, 1982). The same compounds and didrovaltrate relaxed potassium
stimulated contractions and inhibited BaCl2 contractions in guinea pig ileum
preparations in vitro. Potassium stimulated smooth muscle cells were also relaxed by
the Valeriana compounds even when autonomic receptors were blocked by
appropriate antagonists thus suggesting that the effects are not mediated through
receptors of the cholinergic or adrenergic nervous system, but rather demonstrated a
direct effect on the muscle tissue. The inhibition of muscle contractions by the valium
chemicals valeranone and didrovalrate were as potent as papaverine (Hazelhoff,
1982).
Neuroprotective effect
V. officinalis was studied against the toxicity induced by amyloid beta peptide
25-35 Abeta. Cultured rat hippocampal neurons were exposed to Abeta (25-35) (25
microM) for 24-48 h. Valerian extract prevented Abeta evoked neuronal injury which
is characterized by a decrease in cell reducing capacity and associated neuronal
degradation. It also partially inhibited ascorbate /iron-induced peroxidation and
inhibited excess influx of Ca2+ following neuronal injury. Neuroprotective properties
of Valerian against Abeta toxicity at long term can contribute to introduction of
Valerian extract to prevent degeneration in aging or neurodegenerative disorders
(Malva et al., 2004).
Sedative effect
Sedative and sleep enhancing effects were found with Valeriana officinalis
and Valeriana wallichii. Studies from around the world have demonstrated that
Tagara taken before bedtime can improve the quality of sleep, reducing the time to
fall asleep, number of awakenings during the night producing minimal or no
drowsiness in the morning and did not affect anterograde memory (Leathwood and
Chauffard, 1982; Leathwood and Chauffard, 1985). Bicuculline antagonized the
inhibitory effects of both the valerian extract and valerenic acid depicting GABAergic
mechanism in sedative effect. Thus valerian may potentiate the effects of anesthetics

that act on GABA receptors and presurgical valerian use may cause a valerian-
anesthetic interaction (Yuan et al., 2004). Recently, it has been suggested that
valepotriates could be useful in improving the condition of animals (Andreatini and
Leite, 1994) and humans (Poyares et al., 2002) during benzodiazepine withdrawal.
The sedative action could be attributed to the essential oil and valepotriate fractions
(Hendricks et al., 1981). Valerenic acid also produced a dose-related increase in
pentobarbital-induced sleep with 50 and 100 mg/kg i.p. (Hendricks et al., 1985).
Hesperidin present in valerian has sedative and sleep enhancing properties but is not a
ligand for BDZ’s (Fernandaz et al., 2004).
Attempts have been made using in vitro assay techniques to delineate the
mechanism of action for the sedative effects of valerian. Several researchers have
linked the effects of valerian extracts and/or its components with an effect on the
inhibitory neurotransmitter GABA (Mennini et al., 1993; Santos et al., 1994a, 1994b).
GABA mediates sedation in the central nervous system and benzodiazepines exert
their actions via this system. Valerenic acid and acetylvalerenic acids have been
reported to inhibit GABA transaminase, thereby prolonging the inhibitory effect of
GABA (Riedel et al., 1982). However, the effect was small and required mM
concentrations. An aqueous extract of valerian, containing 55 mg valerenic acids/100
g extract, was recently shown to displace radiolabeled GABA from its binding sites
on synaptosomes isolated from rat brains (Santos et al., 1994a). Analysis of the
content of the extract for amino acids revealed that the extract itself contained GABA
in sufficient quantity (4.6 mM) to account for the displacement activity (Santos et al.,
1994b) however, it is unlikely to account for the activity of valerian in vivo because
GABA does not cross the intact blood-brain barrier. Analysis also showed that the
extract contained high amounts of glutamine (13 mM) which is able to cross the
blood-brain barrier. The usual concentration of glutamine in the brain extracellular
fluid is in the range of 0.2 to 0.5 mM. Glutamine has been shown in vitro to stimulate
GABA synthesis in synaptosomes and brain slices, and the authors are investigating
whether valerian extracts have any effect on rat brain amino acid levels in vivo.

Anxiolytic effect
Clinical research demonstrates that standardized valerian extract from
Valeriana officinalis effectively relieves anxiety-related insomnia and suggests that
valerian extract may be comparable to some prescription anti-anxiety drugs for
relieving anxiety. Unlike many drugs, however, valerian is not addictive or habit-
forming when taken in recommended doses. SEREDYN a formulation contains the
optimal dose of standardized Valerian extract (minimum 0.8% valerenic acid) to
provide maximum relief without side effects or drowsiness and only SEREDYN
combines valerian with Passion Flower extract and L-theanine to provide a complete
anti-anxiety supplement. Valeriana produces anxiolytic effects via gamma-
aminobutyric acidergic mechanism i.e inhibition of GABA-transaminase, interaction
with GABA/benzodiazepine receptors and interference in uptake and release of
GABA in synaptosomes (Morazzoni and Bombardelli, 1995; Houghton, 1999).
Apigenin derivatives (6 methylapigenin) isolated from Valeriana officinalis
also exhibited medium-high affinity for the benzodiazepine-binding site and has
anxiolytic effects in mice (Fernandaz et al., 2004). In one study the effect of a mixture
of valepotriates was evaluated on the elevated plus maze performance of diazepam
withdrawn rats and both the diazepam and valerian 12 mg/kg reversed the anxiogenic
effect (Andreatini & Leite, 1994).
Antioxidant effect
In one study efficiency of valerian as antioxidant was tested by the influence
of its extract on the yield of photochemiluminescence of Gly-Trp solutions.
Antioxidant property was examined under conditions when its own absorption was
minimized. Riboflavin as additional sensitizer was used for superoxide generation
(Bol’shakova et al., 1997). The aglycones 8 hydroxypinoresinol and prinsepiol were
isolated from valerian and were found to display powerful antioxidant activity.

Anti-inflammatory effect
In traditional European medicine valerian has also been reported as an
antiinflammatory remedy. A study reports that the ethanolic (EtOAc) extract of the
underground parts of V. officinalis showed inhibitory activity against NF-kappaB at
100 µg/mL in the IL-6/Luc assay on HeLa cells and provided protection against
excitotoxicity in primary brain cell cultures at micromolar concentrations. Bioassay-
guided fractionation of the EtOAc extract led to the isolation of three known
sesquiterpenes: acetylvalerenolic acid, valerenic acid and valerenal. The first two
were active as inhibitors of NF-kappaB at a concentration of 100 µg/mL.
Acetylvalerenolic acid reduced NF-kappaB activity to 4%, whereas valerenic acid
reduced NF-kappaB activity to 25% (Jacob- Herrera et al., 2006).
Muscle relaxant action
The essential oil of valerian and the isolated components valerenal, valerenic
acid, valeranone, and isoeugenyl-isovalerate were screened for central nervous system
effects on mice upon intraperitoneal administration (Hendricks et al., 1985). The
essential oil showed muscle relaxant activity with the oxygenated components
exhibiting more activity than the hydrocarbon fraction. Valerenal and valerenic acid
were more active than valeranone, producing ataxia at 50 mg/kg. In a subsequent
study on valerenic acid, the authors reported a decrease in rotorod and traction
performance in mice given 100 mg/kg ip.
Other actions
Fixed valerian-hops extract combination Ze91019 which is used as a sleep aid
was studied for in vitro binding study at 14 subtypes of five classes of system
receptors (dopamine, serotonin, melatonin, MCH and neuropeptide Y). Binding
affinities were demonstrated at some of the screened melatonin (ML1 and ML2) and
serotonin (5-HT4, 5-HT6 and 5-HT7) receptor subtypes. However the nature of affinity
(agonist/antagonist) of Ze91019 to the respective receptors is yet to be determined.

Serotonin (5-HT4, 5-HT6, 5-HT7) receptor has its role in cognitive performance,
depression and sleep disorders. It was found that valerian extract (V. officinalis) has
affinity to ML1 and ML2 receptors (Abourashed et al., 2004; Fauteck et al., 1996). In
other study polar extracts of valerian roots (V. officinalis) activated A1 receptors
(partial agonistic activity) while nonpolar extracts showed antagonistic or inverse
agonistic activity at A1 receptors as demonstrated by GTPγs binding assays at human
recombinant A1 receptors stably expressed in Chinese hamster ovary cells. Isovaltrate
was characterized as a potent, highly efficacious inverse agonist at adenosine A1
receptors (Lacher et al., 2007). Adenosine appears to be one of the main sleep
inducing substances in the brain, which accumulates during wake time, and its
sedative effects may be mediated by both adenosine A1 and A2A receptors. Activation
of adenosine A1 receptors results in sedative, anticonvulsive, analgesic, antidiuretic,
negative inotropic and antiarrhythmic effects (Yan et al., 2003). Selective antagonists
for A2A receptors are promising novel therapeutics for Parkinson’s disease and may
also exhibit neuroprotective and antidepressive activities (Chen, 2003). In one
radioligand binding study it was found that valerian extracts (DCM and petroleum
ether) were found to have strong affinity to 5HT (5a) receptor, and only weak binding
affinity to the 5HT (2b) and the serotonin transporter. 5HT (5a) receptor is located in
suprachiasmatic nucleus of the brain, which is implicated in the sleep-wake cycle.
Petroleum ether extract inhibited [(3) H] lysergic acid diethylamide (LSD) binding to
the human 5-HT (5a) receptor (86% at 50 µg/ml) and DCM extract inhibited LSD
binding by 51%. These results indicate that valerian and valerenic acid are new partial
agonists of the 5HT (5a) receptor (Dietz et al., 2005).
1.3 VALERIANA WALLICHII Synonym: Valeraina jatamansi (Indian valerian)
Hindi- Muskhbala, Tagar Bengal- Mushkbala, Tagar
Kashmir- Mushkbala Punjab- Balamushk, Bala, Mushkwali
Garhwal- Sumaiya Bombay- Tagarganthoda
Kumaon- Samyo

Valeriana wallichii commonly known as Indian valerian is one of the
important plant species of commerce, which belongs to the family Valerianaceae. It is
native to Indian Himalayas.
1.3.1. Distribution
It is found in the temperate zones of Himalayas, from Kashmir to Bhutan at an
altitude of 1300-3300 m, in the Khasia Hills (India) at an altitude of 1300-2000 m,
and in Afghanistan and Pakistan (Bos et al., 1997).
1.3.2. Characteristics
V. wallichii is a perennial, erect herb; 60-100 cm high. The roots are yellowish
brown, 1.5-7 cm long and 1-2 mm thick. The rhizome is yellowish to brownish, 4-7
cm long and 1 cm thick, sub-cylindrical. Leaves are radical, persistent, stalked,
cordate-ovate, acute, toothed. The flowers are white or tinged with pink in a terminal
corymb, 2.5-8 cm across, often unisexual, and the male and female on different plants.
Fruits are small, smooth, without hairs (Kapoor, 1990).
V. jatamansi is abundant in the Western Himalayas whereas V. hardwickii and
V. officinalis are rather scarce. It prefers deep rich soil and flourishes in shady and
moist localities. The plant flowers during April-June and the rhizomes and roots are

dug out in the autumn. They are cleaned with water and dried in the sun on mats, or
sometimes by spreading them in trays, supported over wood-fire. Considerable
quantities of the drug are transported from the hills, especially from Kashmir and
other places in North-West Himalayas, to the plains. Some quantities of this drug are
also imported from Afghanistan. A survey in Pakistan showed that drug is sometimes
adulterated with other miscellaneous rhizomes upto extent of 27%. The dried
rhizomes and roots of V. jatamansi have been recognized as Indian valerian in the
IPC, distinct from those of V. officinalis recognized as valerian.
Dried Indian Valerian
Indian valerian occurs in the market in dull yellowish brown pieces of
rhizome, 4-8cm long × 5-12 mm thick, sub cylindrical, somewhat flattened usually
slightly curved and unbranched, upper surface bearing numerous raised leaf scars and
the under having prominent circular root scars, with a few roots attached; fracture and
horny; taste bitter. The drug according to official specification shall contain ash not
more than 12%, foreign organic matter not more than 12%; and alcohol (60%),
solvent extractive not more than 30%. On steam distillation, the dried rhizomes and
roots of both the forest and cultivated types yield a sweet smelling essential oil. The
yield of the oil from the fresh rhizomes and roots of cultivated types was found to be
higher (1.8%) whereas the dried ones of the forest origin have been found to yield
only 0.5-0.7 %. The physicochemical characteristics of the oils obtained from the
cultivated and wild rhizomes and roots are significantly different (Sood, 1965). The

oil is a pale-brown or amber-yellow coloured liquid with root-like colour with a
distinct note of valeric acid, more or less pronounced according to the age of the oil.
Musk-like and patchouli-like camphoraceous notes are quite characteristic. The oil
distilled from shade-dried roots, not too old, has a finer odour reminiscent of violet
leaf oil. The odor and flavor of Indian valerian oils are considered poor as compared
to those of V. officinalis oils. Solvent extraction of Indian valerian roots by benzene,
petroleum ether or alcohol yields semisolid resinoids which also have a fair demand
because of the relative low price and the ease with which it can blend with other
perfumes.
1.3.3. Chemical constituents
Valepotriates
These compounds are chemically unstable iridoid triesters in which the
various hydroxy groups are esterified with acetic, isovaleric, hydroxyisovaleric, and
α-methylvaleric acids. The valepotriates are divided into the monoene valepotriate
didrovaltrate and several diene valepotriates (valtrate, isovaltrate, homovaltrate, and
acevaltrate). Valtrate is the most abundantly present diene valepotriate alongwith 1-
acevaltrate and dihydrovaltrate in V. wallichii (Bounthanh et al., 1981). The
valepotriates content varies greatly among the species with higher percentage in
underground parts but lower quantities are reported from aerial parts. Because of their
lipophilicity and instability in aqueous solutions, these compounds are present in only
small amounts in commercially available root extracts. Several active degradation
products of valepotriates (baldrinal, 11-ethoxyviburtinal, and homobaldrinal) and an
iridoid ester glycoside designated as valersidatum (isovaleryl glucoside, m.p. 78-
80°C) has been identified. V. wallichii contains IVHD-valtrate, valerosidate (Yu et al.,
2006) and five new iridoids, 1-homoacevaltrate, 1-homoisoacevaltrate, 11-
homohydroxyldihydrovaltrate, 10-acetoxy-1-homovaltrate hydrin, and 10-acetoxy-1-
acevaltrate hydrin, were isolated from the rhizomes and roots of V. jatamansi (Tang et
al., 2002). A new iridoid, 11-methoxyviburtinal (Chen et al., 2005), 8-
methylvalepotriate and 1,5-dihydroxy-3,8-epoxyvalechlorine were also isolated from
the roots of V. wallichii.

O
O
O
H
O
O
CH3CH3
O
O
O
O
O
O
O
O H3C
CH3
H
O
O
CH3
CH2OCOCH2CH(CH3)2 Acevaltrate Didrovaltrate
O
O
O
H
O
CH3
O
CH3
O
O
CH3
O
O
O
O H3C
CH3
HO
O
O
CH3
O
CH3CH3
O
O
O
1-Homoisovaltrate Isovaleroxyhydroxy didrovaltrate (IVHD)
O
O
O
HO
O
CH3
O
CH3
O
O
CH3
O
O
O
O H3C
CH3
H
O
O
CH3
CH2OCOCH2CH(CH3)2
11-Homohydroxyldidrovaltrate Didrovaltrate
O
O
O
O
O
O
O
OH
H3CO
OH
H
O
OH
O
O
H
OH
Cl
O
8-methylvalepotriate 1,5-dihydroxy-3,8-epoxyvalechlorine A

O
CH2O CH3
O
CHO
O
CH2O
O
CHO
CH3
CH3
Baldrinal Homobaldrinal
Volatile oil
The essential oil from root contains calarene, β-bargamotene, α-santalene, α-
curumene, xanthorrhizol, valeranone, curcumene, α, β and γ-patchoulenes, α-
fenchene, patchouli alcohol, maaliol, β-sitosterol, valeranone, maali-oxide, valerenic
acid, isovaleric and β-methylvaleric acids (chief constituents) (Khare, 2007), formic,
propionic, butyric, palmitic acid and stearic acids, and isovaleryl ester of D(-)α-
hydroxyisovaleric acid. The oil also contains valerianian, α-pinene, camphene and
terpineol (Arora & Arora, 1963). From the rhizome, citric acid, malic acid, maliol,
succinic acid and tartaric acid have been isolated (Kapoor, 1990).
A study revealed that the chemical compositions of the oils show two
chemotypes within V. wallichii. The type-I was characterized by presence of maaliol
(64.3%), viridiflorol (7.2%) and sesquiterpene hydrocarbons (19.2%). The type-II
contained patchouli alcohol (40.2%), viridiflorol (5.2%), 8-acetoxy-patchouli alcohol
(4.5%) and sesquiterpene hydrocarbons (34.5%). Viridiflorol and 8-acetoxy-patchouli
alcohol have been isolated from V. wallichii for the first time. Didrovaltrate was
major compound in the dichloromethane extract in both the chemotypes. 11-α-
acevaltrate was present in both varieties but was in greater percentage in V. wallichii
(maaliol type). Isovaleroxyhydroxy didrovaltrate (IVHD) was common in both
varieties. Homoisovaltrate is present in V. wallichii (patchouli alcohol type), and is
absent in V. wallichii (maaliol type) (Sammal, 2005; Mathela et al., 2005a; Mathela et
al., 2005b).

H
OH
H
H
OH
H
H Maaliol Patchouli alcohol
H
COOH
H
O
Valerenic acid Valeranone
H
OHH
OH
Viridiflorol Terpineol
Flavonoids
Two new flavone glycosides, acacetin 7-O- -sophoroside and acacetin 7-O-
(6”-O-α-l-rhamnopyranosyl)-β-sophoroside were isolated from the rhizomes and roots
of V. jatamansi Jones (Tang et al., 2003). Plant contains 6-methylapigenin and
hesperidin (Marder et al., 2003).
OOH
HO O
OH
6-methylapigenin

Other constituents
Rhizomes and roots contain cyclopentapyrans, acacetin-7-O-rutinosides,
linarin iso-valerinate (Thies, 1968), 4-methoxy-8-pentyl-1-naphthoic acid (Pandey
and Shukla, 1993), lignan prinsepiol-4-omicron-beta-D-glucoside, coniferin,
hexacosanic acid, limonene, choline, chatinine, valerianine, actinidine, tannins and
resins. 4-methoxy 8-pentyl-1-napthoic acid and methyl eicosanoate (Pande et al.,
1994), cubenol, caryophyllene oxide, cadinol and aristolene (Dua et al., 2008) are
other constituents isolated from this plant.
N
CH3
H3C
N
CH2CH
H3C
O Actinidine Limonene Valerianine
1.3.4 Characterization of V. wallichii by TLC
a) Under 254 nm UV V. wallichii is characterized by two dominating
bands at Rf 0.37 and 0.53, and the bands at the higher Rf are missing
(Fig. 1).
b) Under visible light, the band at Rf 0.25 in V. wallichii is blue. The
most intense band of V. wallichii is a brown band at Rf 0.53. There is
another intense brown band at Rf 0.37. No bands appear at Rf 0.55 and
0.8 and V. wallichii shows a sharp violet band at Rf 0.48 between two
broad brown bands (Fig. 2), (Herbalist, 1999).

Fig. 1. HPTLC plate viewed under 254 nm UV light
Fig. 2. HPTLC plate viewedafter application of the HCl-acetic acid reagent in visiblelight
1.3.5. Ethnomedical uses
Locally it is being used for medicinal purpose especially for headache and eye
trouble. In Ayurvedic medicine, it is used as aromatic, stimulant, diuretic (Said,
1970), carminative, and antispasmodic. It is also used for the treatment of epilepsy,
hysteria, chorea, shell shock and neurosis (Sharma, 2003). The crushed leaves are
rubbed on the forehead in extreme headache (Bhattacharjee, 1998). Powdered drug,
mixed with sugar is used in urinary troubles. It is useful in jaundice (Awan, 1990),
ulcers, wounds, cardiac debility, dry cough, asthma, chronic and intermittent fever. A
decoction of the drug is reported to be given in Nepal to mother after parturition
probably as sedative. The extract showed antimicrobial effect against Micrococcus
pyogens var. aureus and Entamoeba histolytica. It is also used as an insect repellent.
1.3.6. Other uses
V. wallichii is mainly used in different pharmaceutical or medical
manufacturing for the proper cure of migraine. In India, the dried rhizomes are used in
perfumes, hair preparations and as incense (Bhattacharjee, 1998). The oil is used as
adjunct of certain flavours for tobacco, honey, root-beer types, etc. In perfumery, it
can be employed for blending in high-grade perfumes and as a fixative. The oil finds
use as a tonic and stimulant in certain medicinal preparations.

1.3.7. Actions of chemical constituents
Valerenic acid
• It reduces reactive oxygen species production and enhances the cell viability.
• Valerenic acid inhibits the enzyme responsible for the central catabolism of
GABA (Riedel, 1982).
• Valerenic acid has also been shown to depress CNS activity (Hendricks et al.,
1985).
• It causes activation of K+ ATP channel, thus act as hypotensive.
• It enhances sedative and sleeping properties.
• It causes increase in short-term memory.
• Valerenic acid (VA) antagonized picrotoxin induced convulsions in mice at
12.5 and 25 mg/kg i.p. (Hiller and Zetler, 1996).
• Valerenic acid makes substantial contribution to the sedative and spasmolytic
activity of the essential oil and extract of V. officinalis (Singh et al., 2006).
• The valerenic acids (val. acid, AcO-val. acid) inhibit the enzymatic
inactivation of GABA in the brain by 20-38% (accumulation of GABA may
result in a decreased central activity and increased sedation).
Valepotriates
• The valtrates (especially dihydrovaltrate) have specific affinity for barbiturate
and peripheral benzodiazepine receptors.
• In one study valtrate was found to be a new Rev-transport inhibitor (a HIV-1
viral regulatory protein) with anti-HIV activity. Valtrate also inhibited the p-
24 production of HIV-1 virus without any cytotoxicity against the host MT-4
cells (Murakami et al., 2002).

Glutamime
• It may also contribute to increasing the GABA concentrations, because in the
brain glutamine (which easily passes the blood-brain-barrier) can be converted
into GABA.
Actinidine
• It appears to affect the GABA neurosystem.
Cyclopentapyrans
• It exhibit sedative, tranquillising and bacteriocidal properties.
1.3.8. Pharmacological actions of V. wallichii
Anxiolytic effect
V. wallichii extract is useful in the treatment of human stress related disorders.
It not only significantly reduces stress and anxiety, but also significantly improved
depression and enhanced the willingness to adjustment (Bhattacharyya et al., 2007).
Sedative effect
It has been observed that defatted extract of V. wallichii is much more
effective than aqueous one. The former (100 mg/kg) reduces the motility to about
30% and the animal remains drowsy. 50 and 100 mg/kg of the defatted extract
showed a marked potentiation on thiopental sleep. EEG shows a reduction in voltage
and frequency of the fast waves. The animal is less alert and responsive to pricking.
While neither of the extract was effective against electro- and chemo-convulsion.
Conditioned behavior response (CBR) remains unaffected. Valeriana wallichii +
Humulus lupulus + Passiflora incarnate +Trifolium pretense + Momordica charantia
is a combination used for treating insomnia.

Neuroprotective effect
Bilateral carotid artery occlusion followed by reperfusion produce significant
cerebral infarction and impair short-term memory, motor co-ordination and lateral
push response. In a study done on ischemia-reperfusion induced cerebral injury,
extracts of V. wallichii markedly attenuated cerebral injury in terms of decreased
infarct size, increase in short-term memory, motor incoordination and lateral push
response (Rehni et al., 2007).
Antiprotozoal activity
A root extract of V. jatamansi (code BAL-O) exhibited larvicidal and
adulticidal activity against different mosquito species. The median lethal
concentration (LC50) of BAL-O against larvae of Anopheles stephensi, Anopheles
culicifacies, Aedes aegypti, Aedes albopictus, and Culex quinquefasciatus were 68.1,
42.8, 51.2, 53.8, and 80.6 mg/liter, respectively. The LC50 and the 90% lethal
concentration against adult Anopheles stephensi, Anopheles culicifacies, Anopheles
aegypti, Anopheles albopictus, and Culex quinquefasciatus were 0.14, 0.16, 0.09,
0.08, and 0.17 and 0.24, 0.34, 0.25, 0.21, and 0.28 mg/cm2, respectively (Dua et al.,
2008).
Nematicidal action
In a study, essential oil of V. wallichii exhibited good nematicidal activity
against the pine wood nematode, Bursaphelenchus xylophilus (Kim et al., 2008).
Anxiolytic action
A study was done on V. wallichii, for its role in stress disorders in hospital
based clinical set up. The observations exhibited that, V. wallichii not only
significantly (p < 0.001) attenuated stress and anxiety, but also significantly (p <
0.001) improved depression and also enhanced the willingness to adjustment.
Nevertheless it did not alter memory, concentration or attention of the volunteers. The

results suggest that V. wallichii may be useful in the treatment of stress related
disorders in human and may be a promising anti-stress agent in near future
(Bhattacharyya et al., 2007). Apigenin derivatives (6 methylapigenin) isolated from V.
wallichii and V. officinalis also exhibited medium-high affinity for the
benzodiazepine-binding site and has anxiolytic effects in mice (Fernandaz et al.,
2004). In one study the effect of a mixture of valepotriates was evaluated on the
elevated plus maze performance of diazepam withdrawn rats. Both the diazepam and
valerian 12 mg/kg reversed the anxiogenic effect (Andreatini and Leite, 1994).
Anticonvulsant effect
The water extract (i.p or i.g) of V. jatamansi together with pentobarbital
sodium can antagonize convulsive action induced by thiosemicarbazide. Although it
is ineffective on the convulsion induced by picrotoxin, it can rather prolong the latent
period of convulsion induced by picrotoxin in mice (Cao and Hong, 1994). It was
shown that pure valerenic acid (VA) antagonized picrotoxin induced convulsions in
mice at 12.5 and 25 mg/kg ip (Hiller and Zetler, 1996). VA was assumed to be the
most important active component in Valeriana.
Analgesic effect
There’s a study which depicts that a substance isolated from the rhizomes and
roots of Indian valerian has marked effect in the Haffner test (Schultz and Eckstein,
1962). Dried leaves of V. jatamansi at a dose of 2 mg acts as central analgesic
(Shrivastava and Sisodia, 1970).
Antispasmodic effect
The valepotriates and sesquiterpenes are reported earlier for their
antispasmodic activity (Hazelhoff et al., 1982) and they might have a role in the
relaxant effect of the extract. Spasmolytic effect of the V. wallichii has been used in
different gastrointestinal disorders such as diarrhea and abdominal spasm. Crude

extract of V. wallichii rhizomes (0.1-3 mg/ml) and its fractions caused relaxation of
spontaneous contractions in rabbit jejunum preparations. When tested against high K+
(80 mM)-induced contractions it produced weak inhibitory effect, while caused
complete relaxation of the contractions induced by low K+ (20mM). These results
indicate that the antispasmodic and hypotensive effects of V. wallichii are mediated
possibly through KATP channel activation, which justify its use in gastrointestinal and
cardiovascular disorders (Anwar et al., 2005). 8 hydroxypinoresinol also showed a
higher vasorelaxant activity.
1.4. VALERIANA HIMALAYANA
Synonyms: Valeriana dioica C.B. Clarke, Valeriana pusilla Royle
1.4.1. Distribution
It is distributed in N.W. Asia, Kashmir and the Karakorums and is fairly
common in marshy or moist places, also in shade, among rocks or meadows from
3500-5000 m.
1.4.2. Characteristics
A dioecious or polygamo-monoecious plant, 11-38 cm tall. Rootstock with
slender roots. Scape solitary or 2-3, glabrous, striate. Nodes minutely pilose. Radical
leaves entire, crenulate or subserrate, oblong to suborbiculate, 1-2.5 cm x 0.8-1.6 cm;
nerves prominent; petiole up to 6.5 cm long. Lower cauline leaves pinnati-partite,
with 1-3 lateral lobes, the terminal one largest, ovate-oblong to elliptic-ovate, entire or
slightly toothed; the uppermost cauline leaves with narrower segments. Flowers in
dichotomous corymbs or compact heads, pink or white. Upper bracts lanceolate, 4
mm long, acute or obtuse. Ovary glabrous; stigma 2-3-fid. Achene 2.5-3 mm long,
glabrous, shorter than the upper bracts; pappus segments 10-13. Flowering period is
from June-August. The female flower can be recognized by its smaller size and 2-3-
fid stigma; the male flowers are broader with more or less included anthers.

1.4.3. Uses
Roots of this plant are used for psychiatric problems, for increasing memory,
and in cough, asthma, paralysis and nerve problems (Khan and Khatoon, 2008).
1.4.4. Chemical constituents
It contains maaliol, valeranone, kessane and α-kessyl acetate as major
compounds (Mathela et al., 2005b).
1.5. VALERIANA PYROLAEFOLIA. DECNE
1.5.1. Distribution
It is found in the temperate Himalayas from Kashmir to Bhutan.
1.5.2. Characteristics
Valeriana pyrolaefolia Decne is closely related to V. jatamansi in morphology
and both these species occur more or less interspersed. A preliminary chemical
investigation of the former species has indicated that it may form a substitute for the
valerian. It is therefore not unlikely that the rhizomes and roots of both the species
may be mixed at the time of collection of the drug. Pharmacognostic study of V.
pyrolaefolia has, however, shown that it has smaller rhizome than that of V.
jatamansi. V. pyrolaefolia can also be differentiated from the latter species by
differences in the dimensions of cells in various tissues (The Wealth of India, 2005).
V. pyrolaefolia is a less known Himalayan species within the genus Valeriana. The V.
pyrolaefolia is a 5-25 cm sub-succulent herb that grows along waysides, hilly slopes
and shrubberies at an altitudinal range of 2,100-3,200 m. The distinguishing
morphological feature of the herb V. pyrolaefolia from V. wallichii is in having 1-3
pairs of stalkless broadly ovate entire stem leaves, white flushed pink flowers, hairless
fruits and other characteristics like the dimension of the cells in various tissues. The
roots are highly aromatic and the fresh leaves have a strong odor though weaker than
that of the roots. No popular uses are referred to in the literature examined, but it is

mentioned that based on closely related morphology with V. wallichii, V.
pyrolaefolia is also collected along with V. wallichii for commercial purpose and has
been reported as a substitute for V. wallichii (Prakash, 1999).
1.5.2. Chemical constituents
The chemical composition of root and leaf oils of V. pyrolaefolia Decne. was
determined by GC and GC/MS, and 45 components were identified. Both oils were
rich in sesquiterpenoids and characterized by high contents of valeranone (31.8-
37.9%) and patchouli alcohol (8.6-12.9%). The leaf oil composition showed
remarkable presence of a non-terpenoid, 2,6-dimethoxyphenol (19.8%), among the
major compounds. The presence of valeranone and patchouli alcohol suggests that it
has commercial potential (Mathela et al, 2005b).
O
O
OH
2, 6 dimethoxyphenol
1.6. VALERIANA HARDWICKII
Hindi - Tagger, Shumeo Bengali- Tagger, Bal, Chur, Ushusr, Saru tagar
Punjab-Taggar, Balachar Kumaun- Asarun, Shumeo
Lepcha- Chammaha Bombay- Tagger-genthoda
1.6.1. Distribution
It is found from Easren Asia - China to the Himalayas. Usually found amongst
herbaceous vegetation on humus-rich soils, 1900 - 3100 metres in Kashmir, Bhutan
(Singh and Kachroo, 1976). Also found in the Khasi and Jaintia hills between 1500
and 1800 m.

1.6.2. Characteristics
The herb is common along road sides, in shady areas of forests, hilly slopes
and flowers during rainy season. An erect herb, 0.3-1.6 m high. Rootstocks are
descending. Radical leaves long petiolate, ovate, soon disappearing, cauline leaves
opposite, odd-pinnate and very deeply pinntified; leaflets 3-9, ovate to oblong-
lnaceolate usually entire, terminal segments largest, petiolate, lateral ones subsessile.
Flowers in compound corymbs forming a terminal lax panicle corolla white about 1.5-
2mm long. The flowers are dioecious (individual flowers are either male or female,
but only one sex is to be found on any one plant so both male and female plants must
be grown if seed is required) and are pollinated by insects. Fruit ovate-oblong,
compressed with 3-dorsal, 1 ventral and 2 marginal ribs, hairy on one side or both
crowned with long, plumose calycinal bristles (Pandey, 1995). The plant prefers light
(sandy), medium (loamy) and heavy (clay) soils and requires moist soil. Taxonomists
have classified two distinct varieties of Valeriana hardwickii viz. V. hardwickii var.
hardwickii and Valeriana hardwickii var. arnottiana. Both these subspecies occur in
the Kumaun (Uttarakhand) in higher altitude region (8,000- 10,000 ft).
The rhizomes and roots of V. hardwickii possess more or less the same
properties as those of V. jatamansi and V. officinalis and are, therefore a good
substitute for the drug valerian. Their tincture also has shown to be equivalent in
pharmacological activity to that of valerian. The rhizomes and roots of V. hardwickii
are transported to the plains and not infrequently they are mixed with the official drug
valerian. Besides the use in medicine they are also employed as an incense and to
scent the hair.
1.6.3. Medicinal Uses
Used as V. jatamansi and V. officinalis. The root is bitter, carminative,
diuretic, expectorant, nervine and stimulant (Usher, 1974) It is used as a nerve tonic
and in the treatment of conditions such as epilepsy and hysteria. It is also used in the

treatment of rheumatism and low blood pressure. The pounded root or leaves are used
as a poultice to treat boils. The plant is antispasmodic, aphrodisiac, emmenagogue,
diaphoretic and stimulant (Duke and Ayensu, 1985). This plant is an effective
substitute for V. officinalis (Chopra et al., 1986).
1.6.4 Chemical constituents
Roots contain a crystalline acid, α-eudesmol, edemo, angelicin etc (Sharma,
2003). Epoxysesquithujene, a new sesquiterpene epoxide has been characterized in
the essential oil of Valeriana hardwickii var. hardwickii on the basis of chemical
reactions and extensive NMR data (Mathela et al., 2007). In case of V. hardwickii var
hardwickii total analysis revealed that the sesquiterpenoids constitute 75.1% out of
which 18.19 % are the sesquiterpene hydrocarbons and 56.91% as oxygenated
sesquiterpenes while monoterpenoids are 19.77 %, out of which monoterpene
hydrocarbons constitute 1.9% and oxygenated are 17.87%. Alpha-agarofuran
(46.52%), bornyacetate (14.46%), 1,7-di-epi alpha-cedrene (8.62%) and alpha-
gurjunene (2.45%) are among the major compounds identified. While in case of V.
hardwickii var arnotianna the analysis of oil revealed dominance in oxygenated
sesquiterpenoids (50.95%) followed by sesquiterpene hydrocarbons (22.43%),
oxygenated monoterpenes (16.35%) and monoterpene hydrocarbons (2.43%). The
chief components of the oil are valeracetate (17.26%), bornyl acetate (15.25%) methyl
linoleate (11.73%), cuprarene (10.37 %) and alpha-cedrene (6.22%) (Sati, 2002; Sati
and Mathela, 2004). Most interesting feature in the composition of V. hardwickii var
arnotianna is the presence of sesquiterpene ester, valeracetate which is used for
sedation and antispasmodic purposes.
V. hardwickii var. arnottiana was found to exist as two independent
chemotype i.e. type I contains α-kessyl acetate, valeracetate and 8-epi-kessylglycol
diacetate whereas chemotype-II contains maaliol and kessyl acetate as major
constituents.

O
H
H
AcO
O
H
H
OAc
α-kessyl acetate Valeracetate
O
H
H
AcO
OAc
O 8-epi-kessyl glycol diacetate Epoxysesquithujene
OO
H
OH
H
H Bornyl acetate Maaliol
1.6.5. Uses
It is used as insecticide for scenting hair and is useful in mental disorder,
scenting hair (Sharma, 2003).
Chemosystematic markers within the genus Valeriana
S. No. Name of the species Major/characterstic compounds
1. Valeriana wallichii chemotype I Maaliol, virdifloral
2. Valeriana wallichii chemotype II Patchouli alcohol, 8-acetoxy-patchouli
alcohol, virdifloral
3. Valeriana pyrolaefolia Patchouli alcohol, valeranone

S. No. Name of the species Major/characterstic compounds
4. Valeriana himalayana Valeranone , maaliol, α-kessyl acetate
5. Valeriana hardwickii var.
arnottiana chemotype I
Valeracetate, 8-epikessyl glycol diacetate,
α-kessyl acetate
6. Valeriana hardwickii var.
arnottiana chemotype II
Maaliol, kessanyl acetate
7. Valeriana hardwickii var.
hardwickii
Bornyl acetate
1.7. BIOACTIVITY EVALUATION
The extracts (steam distilled oils and solvent extracts) from above plant
species have been studied for biological activity with major emphasis on following
activities.
1.7.1. Analgesic activity
Analgesics are the drugs which increase the threshold for pain either by acting
on opioid receptors (e.g morphine) or by inhibiting the synthesis of prostaglandins
(aspirin) which are responsible for pain production. Painful reactions in animals can
be produced by intraperitoneal injection of chemicals like phenylquinone, bradykinin
or acetic acid and also by thermal method (application of heat).
1.7.2 Antiepileptic activity
Epilepsy is a heterogeneous symptom complex- a chronic disorder
characterized by recurrent seizures. Seizures are finite episodes of brain dysfunction
resulting from abnormal discharge of cerebral neurons. The causes of seizures are
many and include full range of neurologic diseases, from infection to neoplasm and

head injury. Experimentally epilepsy can be produced in mice by electroshock, by
administration of chemicals like pentylenetetrazole, strychinine and kainic acid.
1.7.3. Anxiolytic activity
Anxiety is a physiological and psychological state characterized by cognitive,
somatic, emotional, and behavioral components (Seligman et al., 2001). These
components combine to create an uncomfortable feeling that is typically associated
with uneasiness, apprehension, or worry. The elevated plus-maze is commonly used
to assess anxiety-like behaviour in laboratory animals (rats/mice). The maze is usually
a cross shaped maze with two open arms and two closed arms, which is elevated
above the floor.
This task exploits the conflict between the innate fear that rodents have of
open areas versus their desire to explore novel environments. Security is provided by
the closed arms whereas the open arms offer exploratory value. When anxious, the
natural tendency of rodents is to prefer enclosed dark spaces to opened brightly lit
spaces. In this context, anxiety-related behaviour is measured by the degree to which
the rodent avoids the unenclosed arms of the maze.
1.7.4. Antidepressant activity
Major depression is one of the most common psychiatric disorders. Despite
intensive research, the mechanisms of action of various pharmacologic treatments are
still not understood, though most are believed to involve effects on two monoamine
neurotransmitters: serotonin and norepinephrine. Antidepressant activity of a drug can
be evaluated using forced swim test in mice.
1.7.5. Antioxidant activity
Living systems have evolved to survive in the presence of molecular oxygen
and for most biological systems; life depends upon its presence. However, oxygen has

double-edged properties i.e. it is essential for life, but it can also provoke damaging
oxidative events within cells. Free radicals are highly reactive substances with one or
more unpaired electrons in its outer orbital and are formed in the body's cells as a
result of metabolic processes (Niki, 1992; Niki, 2001). Free radicals react rapidly with
adjacent molecules via a variety of reactions including: hydrogen abstraction
(capturing), electron donation and electron sharing (McCord, 2000). Reactive oxygen
species (ROS) are found intracellularly and extracellularly and may be produced
endogenously or arise from exogenous sources, i.e. taken in from the environment
(Sies and Cadenas, 1985; Sies, 1997). Important sources of endogenous free radicals
include prooxidative enzyme systems (e.g. lipoxygenase), drugs and their metabolites,
pollutants, and other chemicals and toxins (Halliwell, 1996; Spiteller, 2001). External
sources such as sunlight and other forms of radiation can generate endogenous ROS,
which can lead to a number of diseases (Halliwell, 1996; Stief, 2003). ROS can also
be formed in food through lipid oxidation and photosensitizers exposed to light (Fang
et al., 2002). ROS can be classified into oxygen-centered radicals such as superoxide
anion (O2•−), hydroxyl radical (OH•), alkoxyl radical (RO•), peroxyl radical (ROO•)
and oxygen-centered nonradical derivatives such as hydrogen peroxide (H2O2) and
singlet oxygen (1O2). Other common reactive species are nitrogen species such as
nitric oxide (NO•), nitric dioxide (NO2•), and peroxynitrite (OONO –).
ROS cause lipid oxidation, protein oxidation, DNA strand breaks, and
modulation of gene expression (Evans and Halliwell, 1999; Halliwell, 2000). ROS are
involved in many diseases such as atherosclerosis, cancer, stroke, asthma, arthritis and
other age related diseases. (Halliwell et al., 1992). Because antioxidant defense in the
human body is not completely efficient, increased free radical formation may produce
a continuous level of oxidative damage. So there is a continuous need of antioxidants
from outside source to prevent oxidative stress which refers to a severe disturbance in
the prooxidant-antioxidant balance in favor of the prooxidant, leading to potential
damage. Primary antioxidants most often act by donating a hydrogen atom, while
secondary antioxidants may act by binding metal ions able to catalyse oxidative

processes, by scavenging oxygen, by absorbing UV radiation, by inhibiting enzymes
or by decomposing hydroperoxides (Schwarz et al., 2001). It is known that different
natural phenolic compounds function as both primary and secondary antioxidants by
different mechanisms. Monitoring of either the decrease of the radical or the
antioxidant, or the formation of products can be used for assessing the antioxidant
activity (Decker et al., 2005).

CHAPTER-2
Chemical Screening of Valeriana wallichii Collected from Different Regions of Kumaun
Himalaya

Section A
Isolation and Identification of Constituents from Valeriana wallichii
Essential oils

2.1. a. INTRODUCTION
Valeriana wallichii has long been used in Ayurveda and Unani systems of
medicine (Gupta and Shah, 1981). Roots of V. wallichii are aromatic, possess sedative
activity and are useful in hysteria, epilepsy and neurosis (Kapoor, 1990). The roots of
V. wallichii have been investigated for terpenoids and valepotriates with α-pinene, β-
pinene, camphene and β-cymene as the prinicipal monoterpene hydrocarbons in the
essential oil (Bos et al., 1997). Major sesquiterpenoids reported in the essential oil
V.wallichii are α-curcumene, β-farnesene, α patchoulene, β-patchoulene,
cryptomeridiol, konokonol, maaliol, xanthrorizol and patchouli alcohol (Naryanan et
al., 1964). Present study deals with chemical screening of V. wallichii natural samples
for comparison of their constituents.
2.2. a. MATERIAL AND METHODS
2.2.1. a. Plant material and its identification
V. wallichii (Voucher specimen No. Chem/DST/V.II) and (Voucher specimen
No. Chem/DST/V.01) were collected from Kumaun region during August to October.
The plant materials were identified from Botanical Survey of India, Dehradun and
were coded as VW-I (Voucher specimen No. Chem/DST/V.01) and VW-II (Voucher
specimen No. Chem/DST/V.II) .
2.2.2. a. Extraction of essential oil
The collected plant material was first freed from other mixings and washed
with a stream of water. The fresh plant material (roots 2 kg) each time was subjected
to steam distillation in a copper electric still fitted with spiral glass condensers. The
distillate was saturated with NaCl and the oil was extracted with n-hexane and
dichloromethane. The organic phase was dried over anhydrous Na2SO4 and the
solvent was evaporated under reduced pressure in a thin film rotatory evaporator at
30°C. The oil yield was 0.60% (v/w) for plant VW-II and 0.16% for VW-I. The oil
obtained from VW-II was coded as V-PA and oil isolated from VW-I was coded as
V-MA.

Valeriana wallichii essential oil
(4.0 g)
Column chromatography (Silica gel)
n-hexane 5% Et2O in 10% Et2O in 20% Et2O in 30% Et2O in EtOAC n-hexane n-hexane n-hexane n-hexane
A B C D E Hydrocarbon mixture (1.0 g)
Scheme 2.1.a: Isolation of components from V. wallichii (VW-II) root oil
Valeriana wallichii essential oil
(4.0 g)
Et2O: n-hexane (5% to 10%)
Et2O: n-hexane (10% to 20%)
VW-IIC VW-IID 210 mg 180 mg
n-hexane 5% Et2O in 10 n-hexane
A B C (1 Hydrocarbon Et2O: n-hexane Emixture (1.0 g) (5% to 10%) (
VW-IB VW-IC
(50 mg) (80 m
Scheme 2.2.a: Isolation of compon
Column chromatography (Silica gel)
% Et2O in 30% Et2O in EtOAC n-hexane n-hexane
.0 g) D (0.2 g) E
t2O: n-hexane 8% to 20%)
g)
ents from V. wallichii (VW-I) root oil

2.2.3.a. GC-MS analysis
GC-MS was done using fused silica capillary column (30m x 0.25mm) liquid
phase DB-5 with helium as a carrier gas in Thermoquest Trace GC 2000 interfaced
with Finnigan MAT Polaris Q mass spectrometer. The column temperature was
programmed at 3°/min from 60° to 210 °C. The mass spectra corresponding to GC
peaks were scanned at 70 eV under EI conditions.
2.2.4.a. Isolation of compounds
The oil isolated from the roots of plant VW-II was chromatographed over
silica gel (75 g, 230-400 mesh, Merck). The column was eluted with n-hexane
followed by a mixture of n-hexane: ether (5% to 30% ether in n-hexane) and finally
washed with ethyl acetate. Similar fractions were mixed to give a total of five
workable fractions among several others. The fractions were concentrated and
examined by TLC, IR, GC and GC-MS under isothermal and column temperature
programmed conditions. They were subjected to repeated column chromatography to
isolate pure compounds.
VW-IIC was isolated from fraction C (5-10 % Et2O in hexane) by repeated
column chromatography and VW-IID from Fraction D with 10-20 % Et20 in hexane
(Scheme 2.1.a). Similar procedure was repeated for sample VW-I. Repeated column
chromatography of the fractions gave compound VW-IB and VW-IC (Scheme 2.2.a)
2.3.a. RESULTS
2.3.1.a. GC- MS screening of the essential oil of Valeriana wallichii (VW-II)
The essential oil of the Valeriana wallichii (VW-II) was analysed using GC-
MS. The aim was to identify minor constituents which could not be isolated in the
pure form. The constituents were identified using NIST Library Search Programme
and compared with literature reports. The gas chromatogram showed the presence of
nearly 28 peaks of which 20 have been identified (Fig. 2.1.a). The results of the
analysis are summarized in Table 2.1.a. The total identified constituents comprise
approximately 95% of the total, which is dominated by sesquiterpenoids followed by
monoterpenes. V. wallichii possesses patchouli alcohol (30.72%) as the major
constituent followed by the presence of δ-guaiene (10.69%), seychellene (10.29%),

acetoxyl patchouli alcohol (10.45%), α-guaiene (4.87%), α-humulene (4.22%) and α-
patchoulene (4.28%). Among minor constituents are kessane, δ-selinene,
caryophyllene, bornyl acetate and valencene. The major oxygenated sesquiterpenes in
the previous reports are cryptomendiol, kanokonol, maaliol, xanthrorhizol and
patchouli alcohol but except for patchouli alcohol, other oxygenated sesquiterpenoids
were found to be totally absent in V. wallichii material under investigation.
Fig. 2.1.a. Gas Chromatogram of V. wallichii (VW-II)

Table 2.1.a. Chemical composition of the essential oil V. wallichii (VW-II)
S.No. Content
(% oil)
M+ Major Fragments Compound identified
1. 1.44 136 93,91,77, 121,136, 105 α-pinene
2. 2.87 136 93,121,107,80,79,41 β –pinene
3. 1.75 136 93,91,121,79,77,94,107 Camphene
4. 1.83 164 149,163,134,105,119,150 thymol methyl ether
5. 2.50 164 149,164,134,105,119,150,135 carvacrol methyl ether
6. 1.92 196 95,121,136,154,108,80 bornyl acetate
7. 4.87 204 105,91,147,133,161,189,204 α-guaiene
8. 1.17 204 122,105,79,133,147,161,189,204 γ- patchoulene
9. 4.22 204 93,41,107,121,133,147,204 α-humulene
10. 4.28 204 107,93,119,135,189,147,175 α- patchoulene
11. 10.29 204 91,105,119,133,147,161,175,189 Seychellene
12. 1.09 204 105,91,161,79,119,133,189,204 Valencene
13. 0.92 204 204,105,91,161,189,119,133,175 δ-selinene
14. 10.69 204 187,93,135,119,161,204 δ- guaiene
15. 1.60 204 108, 93,81, 149,189,204 Unidentified
16. 1.73 204 105,161,119,133,147,189,204 α-murrolene
17. 1.62 204 93,107,91,108,189,204 Caryophyllene
18 0.41 204 161,107,105,91,147,189,204 β-gurjunene
19. 1.19 222 108,55,67,126,135,149,189,205 Kessane
20. 2.35 222 107,91,176,187,205,220 Unidentified
21. 30.72 222 138,205,81,95,109,125,161,189,222 patchouli alcohol
22. 10.45 280 125,81,107,177,159,79,205,220,282 8-acetoxyl patchouli alcohol

2.3.2.a. GC-MS screening of essential oil of Valeriana wallichii (VW-I)
The essential oil of VW-I was analyzed using GC-MS with the primary aim of
identification of the major and minor constituents of the essential oil. The gas
chromatogram (Fig. 2.2.a) showed the presence of 30 compounds, out of which 26
compounds have been identified constituting about 92.41% of the total oil. The
separation of solid compound of the root oil was done through the process of
decantation i.e. separation of crystalline part from that of the mother liquor. The
patchouli alcohol which is considered as the characteristic of the V. wallichii was
separated in the form of crystalline part and traces of it were found present in the
mother liquor part giving minor peaks in the GC-MS analysis which was also
confirmed through Co-TLC. Maaliol (36.82%) was found to be the major component
followed by β-gurjunene (21.28%), guaiol (8.60%) and α-santalene (5.42%). The
detailed analysis is given in the Table 2.2.a. The present results show maaliol as the
chief component of the oil (36.79 %) while patchouli alcohol being present as trace
constituent. The present chemical investigation has also shown the presence of
bicyclogermacrene (2.87%), α-longipinene and α-trans-bergomotene (trace) in the
root oil. The results are shown in Table 2.2.a.
Fig. 2.2.a. Gas Chromatogram of VW-I

Table 2.2.a. GC-MS data of essential oil of Valeriana wallichii (VW-I) t= trace percentage
S.No. Content(% oil)
Major Fragments Compound identified
1. 0.65 93,91,79,94,67,107,92,60,105,41,43, 65,121,122,136 α-pinene
2. t 119,91,117,134,120,115,77,41,105,55 p-cymene
3. t 67,93,91,94,79,77,107,121,65,105,53, 51,136 limonene
4. 0.42 91,93, 77,92,121,136,79,105,119,107,65,51,41 γ-terpinene
5. t 93,91,121,79,136,105,77,107,119,92, 67,51,41 α-terpinolene
6. 0.41 95,67,93,91,121,41,77,136,65,105,51, 121 borneol
7. 0.11 149,164,91,134,117,119,150,77,93, 105,51,121 thymol methyl ether
8. 0.68 149,164,91,117,119,150,134,115,105,77,135,65,79 carvacrol methyl ether
9. 0.79 93,95,121,67,91,81,79,136,108,107,43,41,154 bornyl acetate
10. 1.12 105,119,161,204,189,133,91,147,107,81,93 α-longipinene
11. 5.42 91,93,94,79,105,95,189,119,161,77,133,67,148,41,65,204,51 α-santalene
12. 21.28 161,105,119,133,91,189,147,162,93,79,77,204,175,55,41 β-gurjunene
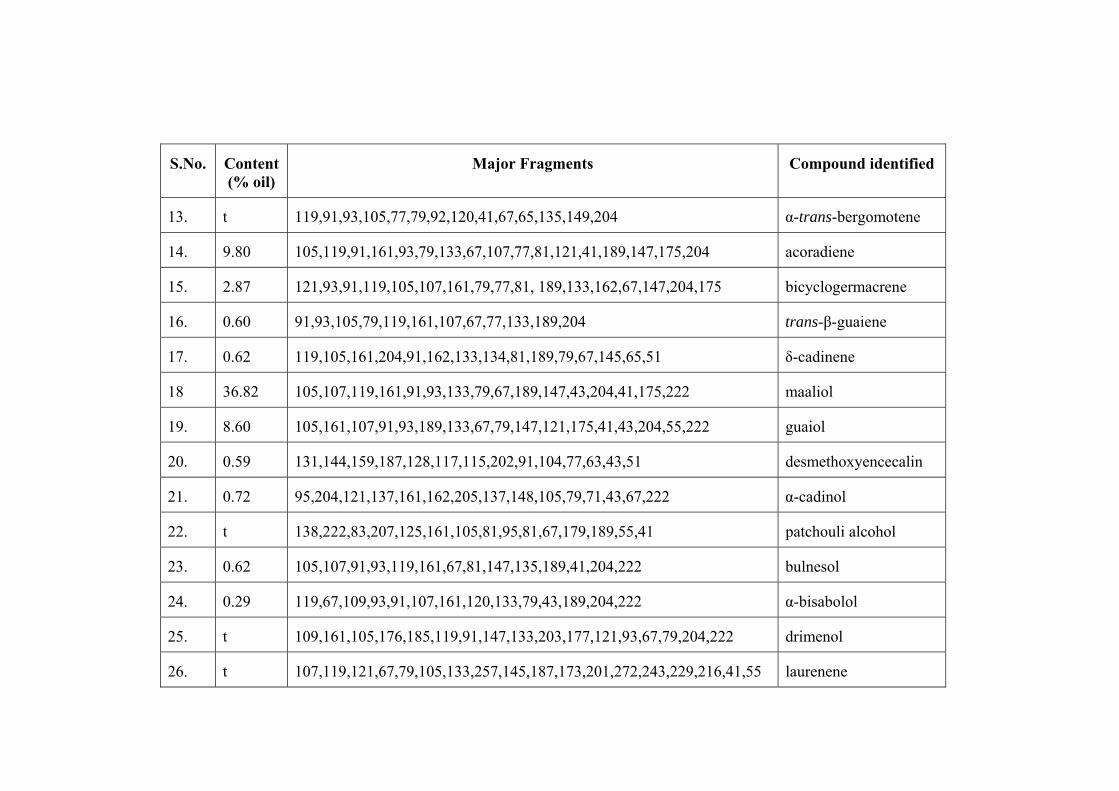
S.No. Content (% oil)
Major Fragments Compound identified
13. t 119,91,93,105,77,79,92,120,41,67,65,135,149,204 α-trans-bergomotene
14. 9.80 105,119,91,161,93,79,133,67,107,77,81,121,41,189,147,175,204 acoradiene
15. 2.87 121,93,91,119,105,107,161,79,77,81, 189,133,162,67,147,204,175 bicyclogermacrene
16. 0.60 91,93,105,79,119,161,107,67,77,133,189,204 trans-β-guaiene
17. 0.62 119,105,161,204,91,162,133,134,81,189,79,67,145,65,51 δ-cadinene
18 36.82 105,107,119,161,91,93,133,79,67,189,147,43,204,41,175,222 maaliol
19. 8.60 105,161,107,91,93,189,133,67,79,147,121,175,41,43,204,55,222 guaiol
20. 0.59 131,144,159,187,128,117,115,202,91,104,77,63,43,51 desmethoxyencecalin
21. 0.72 95,204,121,137,161,162,205,137,148,105,79,71,43,67,222 α-cadinol
22. t 138,222,83,207,125,161,105,81,95,81,67,179,189,55,41 patchouli alcohol
23. 0.62 105,107,91,93,119,161,67,81,147,135,189,41,204,222 bulnesol
24. 0.29 119,67,109,93,91,107,161,120,133,79,43,189,204,222 α-bisabolol
25. t 109,161,105,176,185,119,91,147,133,203,177,121,93,67,79,204,222 drimenol
26. t 107,119,121,67,79,105,133,257,145,187,173,201,272,243,229,216,41,55 laurenene

2.3.3.a. Isolation of compounds from Valeriana wallichii (VW-II)
Two compounds VW-IIC and VW-IID were isolated from the plant.
Characterization of compound VW-IIC
IR film (cm-1): 3500, 2925, 1450 and 1370
MS m/z (%): 222 (M+, 32), 207 (24), 204 (20), 189 (21), 161(33), 138 (100), 121
(29), 98 (107), 83 (38), 81(25)
1H-NMR (δ ppm): δ 0.76 (d, 3H), 0.83 (s, 3H), 1.03 (s, 3H), 1.06 (s, 3H), 1.09(t,
2H), 1.27 (q, 2H), 1.40 (q, 2H), 1.92 (t, 2H), 1.70 (t, 2H), 1.20 (m, 1H), 1.50 (m, 1H),
1.80 (t, 1H).
13C-NMR (δ ppm): δ 18.4, 20.5, 24.2, 24.2, 24.4, 26.7, 28.0, 28.5, 28.7, 32.6, 37.5,
39.0, 40.0, 43.7 and 75.6.
The IR spectrum (Fig. 2.3.a.) and mass spectral data with M+ at 222
corresponding to C15H26O and fragment ion (M-18) suggest the compound to be a
sesquiterpene alcohol. The 13C-NMR spectrum with 15 carbon signals and 1H-NMR
spectral signals at δ 0.76, 0.83, 1.03 and 1.06 showed the presence of 4 methyls. The
signals at δ 1.09, 1.27, 1.40, 1.92 and 1.70 showed the presence of five –CH2- and
signals at δ 1.21, 1.50 and 1.85 showed the presence of three –CH-. The spectral data
suggest the compound as patchouli alcohol and the NMR values of compound VW-
IIC match well with those given in literature for the proposed structure (Nishiya et al.,
1995; Sati, 2002; Tewari, 2002; Sammal, 2005).
OH
H
H
VW-IIC (Patchouli alcohol)

Fig. 2.3.a. IR spectrum of compound VW-IIC
Fig. 2.4.a. MS spectrum of compound VW-IIC

Fig. 2.5.a. 13C-NMR spectrum of compound VW-IIC
Fig. 2.6.a. 1H NMR spectrum of compound VW-IIC

Charcterization of compound VW-IID
IR film (cm-1): 1275, 1735, 3500
MS m/z (%): 280 (M+, 2), 222 (2), 220 (19), 205 (22), 177 (54), 159 (39), 125 (100),
107 (71), 81(76), 79 (21), 67(16), 55(8).
1H-NMR (δ ppm): δ 0.8 (d, 3H), 0.9 (s, 3H), 1.10 (s, 3H), 1.15 (s, 3H), 2.02 (s, 3H),
148 (t, 2H), 1.52 (t, 2H), 1.64 (q, 2H), 1.76 (t, 2H), 1.34 (m, 1H), 1.43 (q, 1H), 1.96
(m,1H), 4.8 (t, 1H).
13C-NMR (δ ppm): δ 18.6, 19.8, 21.5, 23.3, 24.9, 27.7, 27.7, 28.4, 32.5, 35.8,
38.9, 39.9, 42.3, 43.1, 74.5, 75.0, 170.9
The mass spectrum showed, M+ peak at 280 corresponding to C17H28O3 (Fig
2.7.a) while the IR spectrum showed a hydroxyl (3500 cm-1) and acetoxyl group
(1735 and 1275 cm-1). The 1H-NMR spectrum showed methyl signals at (δ 0.80, 0.90,
1.10, 1.15 and an acetoxyl methyl group (δ 4.80) (Fig. 2.8.a.). The 13C-NMR
spectrum indicated the presence of 17 carbons (Fig. 2.9.a.). The compound with 4
degree of unsaturation is a tricyclic sesquiterpene with one hydroxyl and the other as
acetoxyl group. The positions of –OH and –COCH3 groups at C1 and C8 is supported
by comparison of 1H and 13C-NMR data of closely related structures given in
literature (Nishiya et al., 1995; Arantes et al., 1999). On the basis of above spectral
data the compound was identified as 8-acetoxyl patchouli alcohol (Tewari, 2002,
Sammal, 2005).
OH
H
H
OAc
W-IID (8-acetoxyl patchouli alcohol)

Fig. 2.7.a. MS spectrum of compound VW-IID
Fig. 2.8.a. 1H-NMR spectrum of compound VW-IID

Fig. 2.9.a. 13C-NMR spectrum of compound VW-IID
2.3.4.a. Isolation of compounds from Valeriana wallichii (VW-I)
Characterization of compound VW-IB
White needle shaped crystals
IR (Nujol) (cm-1): 3507, 3007, 2917, 1457, 1387, 1373, 1325, 1174, 1103.
MS (m/z) (%): 222 (M+) (3), 207 (2), 204 (16), 189 (36), 175 (13), 161(74), 62(18),
147(35), 133(48), 119(75), 107(80), 105(100), 93 (68), 91(69), 79(45), 67(45),
55(10), 43 (17), 41(14).
1H-NMR δ (ppm): 0.48 (1H, dd, J=5.1, 7.2 Hz), 0.59 (1H, dd, J=7.2, 7.2 Hz),0.76
(1H, m), 0.88 (3H, s), 1.08 (3H, s), 1.27 (3 H, s)1.47 (2H, s), 1.49 (2H, s), 1.74 (2H,
s),1.79 (2H, m),1.80 (2H, s) (Table 2.9.a).

13C-NMR δ (ppm): 72.84, 49.63, 42.70, 41.25, 39.61, 33.02, 29.15, 23.02, 20.14,
19.78, 19.17, 18.76, 17.54, 15.99 and 15.49.
Compound VW-IB was the major component (36.82%) of the oil. The IR
absorption band at 3507 cm-1, m/z at 204 through loss of H2O from M+ confirms the
hydroxyl (-OH) in the molecule [222 (M+)- H2O].
The 1H-NMR exhibits two singlets at δ 0.88 and δ 0.93 ascribed to -CH3 and
CH3-C(OH)- respectively. Two singlets at δ 0.59 (1H, dd, J=7.2, 7.2 Hz) and δ 0.76
(1H, m) must be due to two methine protons fused on cyclopropane ring. Two singlets
at δ 1.08 (3H) and δ 1.27 (3H) are showing a gemdimethyl group. The mass spectrum
with molecular ion peak (m/z) at 222 (M+) corresponds to C15H26O. The carbon
attached to –OH showed up at δ 72.8 ppm. The spectral data show the compound to
be maaliol which is also supported by the literature reports (Nishiya et al., 1992, Sati,
2002, Sammal, 2005).
H
OH
H
H VW-IB (Maaliol)
Fig. 2.10.a. Infra Red spectrum of compound VW-IB

Fig. 2.11.a. Mass spectrum of compound VW-IB
Fig. 2.12.a. 13C-NMR spectrum of compound VW-IB

Fig. 2.13.a. 1H-NMR spectrum of compound VW-IB
Characterization of compound VW-IC
White needle crystals with pleasant odour.
IR (Nujol) (cm-1): 3507, 3007, 2980, 2917, 2856, 1457, 1387, 1373, 1325, 1174,
1141, 1103, 953.
MS (m/z, %): 222 (M+) (65), 207 (32), 205 (5), 204 (15), 189 (17), 179 (20), 161
(30), 138 (100), 125 (32), 105 (24), 95 (22), 91 (20), 83 (34), 81 (22), 67 (18), 55 (7),
41 (12).
1H-NMR δ (ppm): 0.79 (3H, d, J=6.7 Hz), 0.82 (3H, s), 1.02 (3H, s), 1.05 (3H, s),
1.16 (2H, t), 1.22 (2H, q), 1.25 (1H, q), 1.40 (2H, t), 1.45 (1H, m), 1.48 (2H, q), 1.73
(1H, m), 1.83 (2H, t).
13C-NMR δ (ppm):72.87, 49.57, 42.66, 41.22, 39.57, 32.98, 29.12, 22.97, 20.10,
19.76, 19.12, 18.73, 17.49, 15.46 and15.49

Compound VW-IC was another major component. The IR absorption band at
3507 indicates the compound VW-IC possesses an alcoholic group (Fig. 2.14.a). The 1H-NMR spectral data δ 0.82 (3H, s), δ 1.02 (3H, s) and δ 0.79 (3H, d) showed –CH3,
(CH3)2C-OH and >C-CH3, respectively (Fig. 2.15.a). The mass spectrum showed
molecular ion peak (m/z) 222 corresponding to C15H26O which is for a tricyclic
sesquiterpene alcohol (Fig 2.16.a). The carbon attached to –OH showed up at δ 72.8
ppm. Comparison of the data with literature report ((Nishiya et al., 1995; Sati, 2002;
Tewari, 2002; Sammal, 2005) confirms the compound VW-IC as patchouli alcohol.
OH
H
H VW-IC (Patchouli alcohol)
Fig. 2.14.a. Infra Red spectrum of compound VW-IC

Fig. 2.15.a. 1H-NMR spectrum of compound of compound VW-IC
Fig. 2.16.a. Mass spectrum of compound VW-IC

2.4.a. DISCUSSION
The percentage yield of the essential oil was found to be 0.60% (v/w) in case
of plant VW-II. Gas chromatogram revealed 28 peaks out of which 20 compounds
have been identified. The major constituents were separated by column
chromatography followed by their purification. The minor constituents were identified
with the help of GC-MS. Patchouli alcohol (VW-IIC), an isolated compound was
found to be major constituent. 8-Acetoxyl patchouli alcohol (VW-IID) constituted
10.45 % of the oil. The percentage yield of the essential oil of plant VW-I was found
to be 0.16% (v/w). The GC-MS analysis showed the presence of 30 compounds, out
of which 26 compounds constituting 92.41% of the total oil have been identified. An
interesting feature in the mother liquor was the presence of oxygenated
sesquiterpenoid like maaliol (36.82%) followed by β-gurjunene (21.28%), guaiol
(8.60%), α-santalene (5.42%), bicyclogermacrene (2.87%). The major component was
found to be maaliol. The results showed that the composition of the essential oil
isolated from roots and rhizomes of V. wallichii varied depending on the origin. The
plant VW-I was found to be chemotype maaliol of V. wallichii and VW-II was found
to be patchouli alcohol chemotype.

Section B
Isolation and Analysis of Valepotriates of Valeriana wallichii Chemotypes

2.1. b. INTRODUCTION
Same as in section 2.1.a. of chapter 2.
2.2.b. MATERIAL AND METHODS
2.2.1.b Plant material
Roots and rhizomes of Valeriana wallichii (Maaliol type), Valeriana wallichii
(Patchouli alcohol type) were collected from regions as mentioned in section 2.2.1.a.
2.2.2.b. Extraction
After collection, the plant materials were immediately stored at cool and dry
place. The plant material (1kg) were dried in shady place and subjected to
dichloromethane (CH2Cl2) extraction. The extract was concentrated to dryness with
rotary evaporator at 35°C to yield a brown dry mass (2 g). So the yield being 20% for
both the chemotypes. Extract from plant VW-II (Patchouli alcohol type) was coded as
VVDR03 and that from VW-I (Maaliol type) was coded as VVW02. Extraction of the
plant material for HPLC screening was carried out by homogenizing 10 g of dried
material of each Valeriana plant with 50 ml of dichloromethane for 3 days. The
homogenate was filtered and the residue was washed with 5 ml portion of
dichloromethane. Evaporation of the solvent was done below 35°C by rotary
evaporator, and the residue was again dissolved in dichloromethne (3 ml).
2.2.3.b HPLC Analysis
Thermoelectron Quaternary HPLC system with an injection valve system and
a 20µl sample loop and stainless steel column (Thermoelectron C-18) 25 cm x 4.6 mm
i.d was used. The column outlet was connected to UV detector (UV-1000) and RI-
detector (RI-150). The detection wavelengths were 254 nm for diene valepotriates and
210 nm for monene valepotriates.
2.2.4.b. Solvents
Dichloromethane and acetonitrile used were of HPLC grade (Qualigens). The
water used for solvent system was deionised and the HPLC system was isocratic with
binary solvent system, acetonitrile: water (70:30).

2.2.5.b. Analytical separation of valepotriates
Diene valepotriates show absorption maximum at 254 nm, due to their two
conjugated double bonds, and can be detected with high sensitivity at this wavelength.
Monene valepotriates were detected at 210 nm. Acetonitrile: water (70:30) was used
as eluent with 1ml/ min flow rate (Fig. 2.1.b, 2.2.b, 2.3.b & 2.4.b).
Fractionation
The residue (oily mass) was fractionated on column chromatography silica gel
(60-120 mesh) in Et2O: n-hexane (8% to 30%). Repeated column chromatography of
the column fractions followed by purification afforded three compounds (VW-I≠01,
VW-I≠02 and VW-I≠03) from Valeriana wallichii (Maaliol type) and three
compounds (VW-II≠04, VW-II≠05 and VW-II≠06) from Valeriana wallichii
(Patchouli alcohol type).
Valeriana wallichii (Maaliol type) Extract (0.5 g)
Column Chromatography
(n-hexane: Et2O) Silica gel (50 g)
n-hexane 8% Et2O 15% Et2O 20% Et2O 30% Et2O
(A) (B) (C) (D) (E)
Recolumn (n-hexane:Et2O) (5-10%) Si gel (10g)
VW-I≠01 VW-
10 mg 8 m
Scheme 2.1.b: Isolation of components fro
Recolumn (n-hexane: Et2O) (15-20%) Si gel (10g)
I≠02 VW-I≠
g 10mg
m V. wallichii (VW-I
Recolumn (n -exane:Et2O) (25-30%) Si gel (10g)
Hydrocarbonmixture
03
)

Valeriana wallichii (patchouli alcohol type) Extract (0.5 g)
Column chromatography (n-hexane: Et2O) Silica gel (80.0g)
n-hexane 8% Et2O 10% Et2O 15% Et2O 30% Et2O
Hydrocarbon mixture
VW-II≠04 VW-II≠05 VW-II≠06 F
8 mg 10 mg 12mg
(A) (B) (C) (D) (E)
Scheme 2.2
Fig. 2.1.b. HPLC of C
Recolumn (n-hexane:Et2O) (0-5%) Si gel (10g)
.b: Isolation of components from V.
H2Cl2 extract of V. wallichii (Maali
Recolumn (n-hexane:Et2O) (5-10%) Si gel (15g)
wallichii (VW-II)
ol type) detected at 210 nm.

Fig. 2.2.b. HPLC of CH2Cl2 extract of V. wallichii (Maaliol type) detected at RI detector
Fig. 2.3.b. HPLC of CH2Cl2 extract of V. wallichii (Patchouli alcohol type) at 210 nm

Fig. 2.4.b. HPLC of CH2Cl2 extract of V. wallichii (Patchouli alcohol type) detected at RI detector
2.3.b. RESULTS
2.2.1.b. Compounds isolated from Valeriana wallichii (Maaliol type)
Three compounds were isolated from the plant viz. VW-I≠01, VW-I≠02 and VW-
I≠03.
Characterization of compounds
VW-I≠01
IRKBr υ max cm-1: 2925, 1766, 1733, 1250
1H-NMR (CDCl3, 300 MHz) δ ppm: 6.52 (1H, s), 5.84 (1H, d, J=0.5), 4.95 (1H, t,
J=5.5), 4.69 (2H, d, J=12.5), 4.69 (2H, d, J=12.5), 2.97-2.99 (1H, m), 2.72 (1H, d,
J=8.0), 2.05 (3H, s), 1.0 (3H, s).
13C-NMR (CDCl3, 75 MHz) δ ppm: C-1 (88.3), C-3 (145.4), C-4 (11.2), C-5 (60.8),
C-6 (83.0), C-7 (73.2), C-8 (62.2), C-9 (48.8), C-10 (48.2), C-11 (61.9), R1’-C-1’
(170.9), C-2’ (43.3), C-3’ (25.7), C-4’ (17.2), C-5’ (18.8), R2-OAc-(169.8), (20.8),
R3”-C”-1 (169.6), C”-2 (43.3), C”-3 (25.6), C”-4 (22.3), C”-5 (22.4).

The compound VW-I≠01 was obtained as a light brown viscous liquid. 13C-
NMR showed the presence of twenty-three carbons. Valtrate type skeleton was
confirmed by the presence of one epoxide group at the C-8 carbon, one methyl
protons at δ 4.74 adjacent to an oxygen function and one methine carbon δ 83.0
having an oxygen function with an acetate residue at C-7. 13C-NMR (Fig. 2.5.b) and 1H-NMR (Fig. 2.6.b) signals confirmed two isovaltrate groups at C-1 and C-11. The
structure of the compound VW-I≠01 was therefore characterized as didrovaltrate.
Finally, identity was confirmed by comparison of its spectral data with those reported
in literature (Tang et al., 2002; Thies et al., 1981, Sammal, 2005).
O
O
O
O H3C
CH3
H
O
O
CH3
CH2OCOCH2CH(CH3)2 VW-I≠01 (Didrovaltrate)
Fig. 2.5.b. 13C-NMR spectrum of VW-I≠01

Fig. 2.6.b. 1H-NMR spectrum of VW-I≠01
VW-I≠02
IRKBr υ max cm-1: 3500, 1766, 1633, 1255.
1H-NMR (CDCl3, 300 MHz) δ ppm: 6.65 (1H, s), 6.07 (1H, d, J=2.0), 4.92 (2H, d,
J=12.5), 4.93 (1H, d, J=10.0), 4.73 (1H, d, J=12.5), 3.11(2H, d, J=5.0), 2.91 (2H, d,
J=2.0), 2.83 (2H, d, J=5.0), 2.06 (3H, s), 1.99 (2H, t, J=13.0).
13C-NMR (CDCl3, 75 MHz) δ ppm: C-1 (88.1), C-3 (145.2), C-4 (111.2), C-5
(69.5), C-6 (40.25), C-7 (73.1), C-8 (62.1), C-9 (48.02), C-10 (48.2), C-11 (61.1), R1-
C’-1 (169.5), C’-2 (76.6), C’-3 (172.9), C’-4 (42.93), C’-5 (25.51), C’-6 (22.1), C’-7
(22.1), C’-8 (29.81), C’-9 (17.03), C’-10 (18.6), R2-(C”-1) 169.7, (C”-2) 20.64 (R3-
OAc, C”-1), (170.8), C”-2 (42.83), C”-3 (25.55), C”-4 (22.2), C”-5 (22.2).

Compound VW-I≠02 is a viscous light brown liquid. The 13C-NMR showed
27 signals, indicating 7 methyls, 5 methylene, 8 methine and 7 quaternary carbons
(Fig. 2.7.b). The chemical shift of two olefinic methine carbons at δ 145.2 and 111.2
assigned for C-3 and C-4 respectively. The quaternary carbon at δ 62.1 (C-8 and the
methylene carbon at δ 48.2 C-10) are characteristic for an epoxide ring at C-8 and C-
10 and corresponds to a valtrate-type iridoid. A singlet, at δ 2.06 in the 1H-NMR
spectrum was assigned to the methyl protons of an acetate residue (Fig. 2.8.b). 13C-
NMR signal at δ 169.7 was assigned to the carbonyl carbon of the acetate residue.
13C-NMR showed 10 carbon signals due to 4 methyl carbons (δ 22.2, 22.2, 18.6 and
17.03), methylene carbons (δ 42.93) linked to the carbonyl carbon (δ 172.9) three
methine carbons (δ 25.5, 29.8 & 76.6) linked to a carbonyl residue. All these data
were used to assign compound VW-I≠02 as isovaleroxyhydroxy didrovaltrate.
Finally, identity was confirmed by comparison of its spectral data with those reported
in literature (Tang et al., 2002; Becker et al., 1984, Sammal, 2005).
O
O
O
O H3C
CH3
HO
O
O
CH3
O
CH3CH3
O
O
O
VW-I≠02 (Isovaleroxyhydroxy didrovaltrate)

Fig. 2.7.b. 13C-NMR spectrum of VW-I≠02
Fig. 2.8.b. 1H-NMR spectrum of VW-I≠02

VW-I≠03
IRKBr υ max cm-1: 2925, 1766, 1733, 1640.
1H-NMR (CDCl3, 300 MHz) δ ppm: 6.72 (1H, s), 5.98 (1H, d, J=10), 5.89 (2H, d, J=
3.6), 5.39 (1H, d, J=2.5), 2.36-2.21 (2H, m), 2.08 (3H, s), 1.0 (3H, s).
13C-NMR (CDCl3, 75 MHz) δ ppm: C-1 (92.4), C-3 (148.6), C-4 (108.21), C-5
(141.1), C-6 (118.3), C-7 (83.3), C-8 (64.0), C-9 (42.9), C-10 (47.7), C-11 (60.7), R1-
C’-1 (169.5), C’-2 (79.2), C’-3 (170.8), C’-4 (22.1), C’-5 (26.6), C’-6 (26.5), C’-7
(26.5), R2-OAc- (170.3), (20.84), R3-C”-1 (170.4), C”-2 (43.9), C”-3 (25.5), C”-4
(22.1), C”-5 (22.2).
The compound VW-I≠03 was obtained as yellow viscous liquid. 13C-NMR
spectrum of the compound showed the presence of twenty four carbons. 1H-NMR
spectrum revealed signals for methyl groups at δ 0.99, 1.31, 1.52 and 2.06, one methyl
proton at δ 4.69 adjacent to an oxygen function and one methyl carbon δ 79.2 having
oxygen function. Furthermore , the 13C-NMR spectrum showed 7 carbon signal due to
2 methine carbons (δ 25.5, 79.2), 3 methyl carbons (δ 22.1, 26.5 and 26.5) and 2 ester
carbonyl carbons (δ 170.8 and 170.3). These spectral data suggested the presence of
acetoxylisovaleryl group and isovaleryl group present at C-11 and C-1, respectively.
The structure of the compound VW-I≠03 was therefore characterized as 1-α
acevaltrate. Finally, identity was confirmed by comparison of its spectral data with
those reported in literature (Thies et al., 1981; Sammal, 2005).
O
O
O
H
O
O
CH3CH3
O
O
O
O
VW-I≠03 (Acevaltrate)

2.2.2.b Isolation of compounds from Valeriana wallichii (Patchouli alcohol type)
Three compounds were isolated from the plant viz. VW-II≠04, VW-II≠05 and VW-
II≠06.
Characterisation of compounds
VW-II≠04
IRKBr υ max cm-1: 3500, 1770, 1640, 1250.
1H-NMR (CDCl3, 300 MHz) δ ppm: 6.65 (1H, s), 6.07 (1H, d, J=2.0), 4.92 (2H, d,
J=12.5), 4.93 (1H, d, J=10.0), 4.73 (1H, d, J=12.5), 3.11 (2H, d, J=5.0), 2.91 (2H, d,
J=2.0), 2.83 (2H, d, J=5.0), 1.99 (2H, t, J=13.0).
13C-NMR (CDCl3, 75 MHz) δ ppm: C-1 (88.2), C-3 (145.4), C-4 (111.2), C-5
(69.7), C-6 (40.3), C-7 (73.2), C-8 (88.2), C-9 (48.1), C-10 (48.8), C-11 (61.9), R1-C’-
1 (169.6), C’-2 (76.8), C’-3 (173.1), C’-4 (43.0), C’-5 (25.5), C’-6 (22.3), C’-7 (22.3),
C’-8 (29.6), C’-9 (17.1), C’-10 (18.7), R2-OAc- (169.8), (20.84), R3-C”-1 (170.9), C”-
2 (43.0), C”-3 (25.7), C”-4 (22.3), C”-5 (22.3).
Compound VW-II≠04 was obtained as a viscous light brown liquid. It’s 13C-
NMR showed 27 resonance lines. Analysis of the overall NMR spectral data revealed
the presence of an iridoid skeleton with ester units. The quaternary carbons at δ 88.2
(C-8) and the methylene carbon at δ 48.8 (C-10) were characterstic for an epoxide
ring at C-8 and C-10 and corresponds to a valtrate type iridoid. A methyl singlet,
which appeared at δ 2.06 in the 1H-NMR spectrum was assigned to the methyl protons
of an acetate residue. The 13C-NMR spectrum showed 10-carbon signals due to 4-
methyl carbons (δ 22.3, 22.3, 18.7 and 17.1), methylene carbon (δ 42.93) linked to the
carbonyl carbon (δ 173.1), three methine carbons (δ 25.5, 29.9 and 76.6) linked to a
carbonyl residue. All the data show the compound VW-II≠04 as isovaleroxy hydroxy
didrovaltrate. Finally its identity was confirmed by comparison of its spectral data
with literature reports (Thies et al., 1981; Sammal, 2005).

O
O
O
O H3C
CH3
HO
O
O
CH3
O
CH3CH3
O
O
O
VW-II≠04 (Isovaleroxyhydroxy didrovaltrate)
VW-II≠05
IRKBr υ max cm-1: 2925, 1766, 1740, 1640, 1610.
1H-NMR (CDCl3, 300 MHz ) δ ppm: 6.71 (1H, s), 5.98 (1H, d, J=10.0), 5.86 (2H, d,
J=2.5), 5.38 (1H, d, J=2.5), 2.06 (3H, s), 1.10 (3H, s)
13C-NMR (CDCl3, 75 MHz ) δ ppm: C-1 (92.4), C-3 (148.6), C-4 (108.2), C-5
(141.1), C-6 (118.3), C-7 (83.3), C-8 (64.0), C-9 (42.9), C-10 (47.7), C-11(60.7), R1-
C’-1 (169.5), C’-2 (79.2), C’-3 (170.8), C’-4 (22.1), C’-5 (26.6), C’-6 (26.5), C’-7
(26.5), R2-OAc-(170.3), (20.84), R3-C”-1 (170.4), C”-2 (43.9), C”-3 (25.5), C”-4
(22.1), C”-5 (22.2).
The compound VW-II≠05 was obtained as a pale yellow viscous liquid. 13C-
NMR spectrum of the compound showed the presence of twenty four carbons. 1H-
NMR spectrum revealed signals for methyl group at δ 0.99, 1.31, 1.52 and 2.06, one
methyl proton at δ 4.69 adjacent to an oxygen function and one methyl carbon δ 79.2
having an oxygen function. Furthermore, the 13C-NMR spectrum showed seven
carbon signal due to two methine carbons (δ 25.5, 79.2), three methyl carbons (δ 22.1,
26.5 and 26.5) and two ester carbonyl carbons (δ 170.8 and 170.3). These spectral
data suggested the presence of acetoxylisovaleryl group and isovaleryl group present
at C-11 and C-1, respectively. The structure of compound VW-II#05 was therefore
characterized as 1-α acevaltrate. Finally, identity was confirmed by comparison of its
spectral data with those reported in literature (Becker et al., 1984; Thies et al., 1981;
Sammal, 2005).

O
O
O
H
O
O
CH3CH3
O
O
O
O
VW-II≠05 (Acevaltrate)
VW-II≠06
IRKBr υ max cm-1: 1770, 1740, 1640, 1610, 1250.
1H-NMR (CDCl3, 300 MHz ) δ ppm: 6.69 (1H, s), 5.86 (1H, d, J=3.0), 5.96 (1H, d,
J= 10.0), 4.76 (2H, d, J= 12.0), 4.66 (2H, d, J= 12.0), 3.03 (1H, d, J=5.0), 2.91 (1H, d,
J=5.0), 2.06 (3H, s).
13C-NMR (CDCl3, 75 MHz ) δ ppm: C-1 (92.5), C-3 (148.5), C-4 (108.3), C-5
(140.9), C-6 (118.6), C-7 (83.0), C-8 (64.1), C-9 (43.0), C-10 (47.8), C-11(60.8), R1-
C’-1 (170.8), C’-2 (42.9), C’-3 (26.1), C’-4 (22.2), C’-5 (22.3), R2-C”-1(172.4), C”-2
(42.9), C”-3 (25.6), C”-4 (29.6), C”-5 (11.2), C”-6 (19.9).
The compound VW-II#06 was obtained as light brown viscous liquid. 13C-
NMR showed 23 carbons, indicating 5 methyls, 5 methylene carbons, 7 methine
carbons and 6 quaternary carbons (Fig 2.9.b). The signals at δ 6.69 and δ 92.5
respectively were assigned to H-1 and C-1. The chemical shifts of two olefinic
methine carbons at δ 108.3 and 140.9 respectively were assigned to C-3, C-6, C-4 and
C-5. The chemical shift of one methine carbon (δ 83.0), one quaternary carbon (δ
64.1) and one methylene carbon (δ 43.0) were assigned C-7, C-8 and C-9
respectively. The quaternary carbon at δ 64.1 (C-8) and the methine carbon at δ 47.8
(C-10) were characteristic for an epoxide ring at C-8 and C-10. 1H and 13C-NMR data
of compound showed the presence of acetate residue at C-7 and a β-me-isovaltrate
group at C-11. Therefore, compound VW-II#06 was identified as 1-homoisovaltrate.
Finally, the identity was confirmed by comparison of spectral data (Fig. 2.9.b) with
that reported in literature (Tang et al., 2002; Thies et al., 1981; Sammal, 2005).

O
O
O
H
O
CH3
O
CH3
O
O
CH3
VW-II≠06 (1-Homoisovaltrate)
Fig. 2.9.b 13C-NMR spectrum of VW-II≠06

CHAPTER-3
Psychopharmacological Profile of Valeriana wallichii chemotypes

3.1. INTRODUCTION
Although a multitude of pharmaceutical agents are available for the treatment
of mood disorders, anxiety, insomnia and epilepsy; many patients have difficulty
tolerating the side effects, do not respond adequately, or eventually lose their
response. Many therapeutic herbs and nutrients have far fewer side effects and may
provide an alternative treatment or can be used to enhance the effect of prescription
medications.
Epilepsy is one of the most common disorders of the central nervous system,
worldwide. Though, the advent of newer techniques in neurobiology has provided
some insight into the pathophysiology of epilepsies, many aspects of this phenomenon
still remain unknown. It is, therefore not surprising that the currently used
antiepileptic drugs fail to provide satisfactory seizure control for nearly 15 % to 20 %
per cent of patients with epilepsy. For such patients, the combinations of antiepileptic
drugs are often prescribed in attempts to improve seizure control. However, toxicities
associated with these drugs further compromise the quality of life, while drug-drug
interactions may complicate management. Thus there is an ever lasting need for
search of newer molecules for treating seizures.
Valeriana wallichii DC. (Syn: V. jatamansi Jones) is a plant that belongs to
family “Valerianaceae” commomly called as Indian valerian. It is a small perennial
herb of 15-45 cm height with rootstock, thick branching stem, sharply pointed leaves,
white or pink flowers in clusters and hairy fruit. It is indigenous to the temperate
Himalayas and found in Kashmir, Nepal, Bhutan, Burma and Afghanistan. The plant
is widely known for its use in anxiety, insomnia, epilepsy, failing reflexes, hysteria,
neurosis and sciatica (Nadkarni, 1976). It is also considered useful as a potent
tranquilizer, emmenagogue (Nadkarni, 1976), diuretic (Said, 1970) and
hepatoprotective (Awan, 1990). Its active principles are valepotriates (Becker and
Chavadej, 1985) (like dihydrovaltrate) (Bounthanh et al., 1981), linarin-isovalerianate
(Thies, 1968), sesquiterpenoids (Ron et al., 2000), 6- methylapigenin and hesperidin
(Marder et al., 2003). Valerian preparations yield isovaleric acid, a substance
analogous to valproic acid and likely to possess anticonvulsant properties (Eadie,
2004). It is well known that the content and composition of active compounds in

herbal medicines are strongly influenced by many factors, such as genotype, climate,
harvest and preparation process. For example, V. wallichii roots from different
sources were found to contain between 0.09 and 1.30% v/w of essential oil (Kapoor,
1990). In the present work the effect of variation of active compounds between V.
wallichii growing under different environmental conditions has been correlated with
their psychopharmacological profile
3.2. MATERIAL AND METHODS
3.2.1. Plant material and its extraction
Plants were collected and identified as mentioned in section 2.2.1.a. of chapter
2 (section A). The oil was extracted by steam distillation as described in section
2.2.2.a. of chapter 2 (section B). The oil obtained from VW-II (Patchouli chemotype)
was coded as V-PA and that obtained from VW-I (Maaliol chemotype) was coded as
V-MA. Similarly the extracts were obtained by the method as mentioned in section
2.2.2.b. and were coded as VVW02 (maaliol chemotype) and VVDR03 (patchouli
alcohol chemotype).
3.2.2. Animals
Animals used in the study were laca mice of either sex (20-40g) bred in
Central Animal House facility of Panjab University, Chandigarh housed in cages with
food and water ad libitum and maintained on a natural light and dark cycle. The
experimental protocols were approved by the Panjab University Animal Ethical
committee.
3.2.3. Drugs
The drugs used in the present study were Pentylenetetrazole (PTZ) (Sigma,
USA), Diazepam (Sigma, USA). PTZ was dissolved in normal saline. Diazepam was
suspended in a drop of Tween 80 and volume was made up with sterile water. The test
drugs VVW02, VVDR03, V-PA and V-MA were also suspended in Tween-80.
Vehicle treated group received a drop of Tween-80 dissolved in sterile water and
acted as control group.

3.2.4. Experimental protocol and procedure
3.2.4.1. Anticonvulsant activity
Pentylenetetrazol (PTZ, Metrazol) is a central nervous system stimulant and
was synthesized in 1924. In 1926, Hildebrant showed its convulsant action. It
produces jerky type of clonic convulsions in mice. Pentylenetetrazole has been used
experimentally to study seizure phenomenon and to identify pharmaceuticals that may
control seizure susceptibility. The convulsive effect of this drug is considered to be
analogous to petit mal type of convulsions in man. It is considered as a GABA
antagonist but the mechanism of the epileptogenic action of PTZ at the cellular
neuronal level is still unclear.
For evaluating the effect of V. wallichii chemotypes on PTZ-induced
convulsions, animals were divided into following groups, each group consisting of six
animals.
Group 1: Vehicle treated
Group 2-4: VVDR03 (20, 40 and 80 mg/kg p.o.)
Group 5-7: VVW02 (20, 40 and 80 mg/kg p.o.)
Group 8-10: V-PA (20, 40 and 80 mg/kg p.o.)
Group 11-13: V-MA (20, 40 and 80 mg/kg p.o)
Group 14: Diazepam (1 mg/kg, i.p.)
Animals were injected pentylenetetrazol 80mg/kg intraperitoneally (i.p.) one
hour after the drug treatment. Onset of seizures, delay in death and the mortality were
noted after 1 hr in all the groups (Kulkarni, 1999).
3.2.4.2 . Measurement of anxiety levels
Elevated plus maze, is a novel test to study the anxiogenic and anxiolytic drug
effects in rodents (Kulkarni, 1999). Exposure of the animals to novel maze alley
evokes an approach-avoidance conflict which is stronger in open arm as compared to
enclosed arm. Rodents (rats and mice) have aversion for high and open arm, they
freeze, become immobile, defecate and show fear like movements. The plasma

cortisol level is also reported to be increased, as a true reflection of anxiety. Major
advantages of this test procedure are a) it is simple, fast and less time consuming b)
no prior training or noxious stimuli is required and c) it is predictable and reliable
procedure for studying anxiety response as well as antianxiety action of drugs.
The plus maze apparatus for mice consist of two open (16x5 cm) and two
closed arms (16x5x12 cm) and is placed at a height of 25 cm. Anxiety reduction in the
plus-maze is indicated by an increase in the proportion of time spent in the open arms
(time in open arms/total time in open or closed arms), and an increase in the
proportion of entries into the open arms (entries into open arms/total entries into open
or closed arms). Total number of arm entries and number of closed-arm entries are
usually employed as measures of general anxiety in rodents (Hogg, 1996).
Animals were divided into following thirteen groups, each group consisting of
six animals.
Group 1: Vehicle treated
Group 2-4: VVDR03 (10, 20 and 40 mg/kg p.o.)
Group 5-7: VVW02 (10, 20 and 40 mg/kg p.o.)
Group 8-10: V-PA (10, 20 and 40 mg/kg p.o.)
Group 11-13: V-MA (10, 20 and 40 mg/kg p.o)
The animals were then placed individually at the center of the maze with head
facing the open arm one hour after the treatment. During the 5 min test, the number of
entries in closed arms and the time spent in closed arm were recorded (Kulkarni,
1999). Similarly in a separate protocol the effect of above treatments were noted in
elevated plus maze after 14 days of daily dosing.
3.2.4.3. Rota rod test
The test was carried out using an apparatus consisting of a horizontal rod of 3
cm diameter which was made to rotate at a speed of 20 rpm. A pretest was carried out
and only those animals who demonstrated the ability to remain on the revolving rod
for at least 1 min were selected. Animals were divided into following groups, each
group consisting of six animals.

Group 1: Vehicle treated
Group 2-4: VVDR03 (10, 20 and 40 mg/kg p.o.)
Group 5-7: VVW02 (10, 20 and 40 mg/kg p.o.)
Group 8-10: V-PA (10, 20 and 40 mg/kg p.o.)
Group 11-13: V-MA (10, 20 and 40 mg/kg p.o.)
Group 14: Diazepam (1 m/kg, i.p.)
The drugs were given 1 h before the test and the time for which mice remained
on the revolving rod was noted and compared with the control group. Mouse unable
to remain on the rod at least for three min was considered as a positive test and the
time of its fall was recorded (Viswanatha et al., 2006).
3.2.4.4. Statistical analysis
Results were expressed as mean±SEM. The intergroup variation was measured
by one way analysis of variance (ANOVA) followed by Tukey’s test. Statistical
significance was considered at P<0.05. The statistical analysis was done using the
Jandel Sigma Statistical Software version 2.0.
3.3. RESULTS
3.3.1. Acute toxicity
The oral administration of V-PA, V-MA, VVDR03 and VVW02 at doses
ranging from 10-80mg/kg did not produce any lethal effect. No adverse effect or
mortality was detected in albino mice up to 2000mg/kg, p.o dose of VVDR03 and
VVW02 during 24 h observation period. Similarly no adverse effect or mortality was
detected in albino mice up to 1000mg/kg dose of V-PA and V-MA.3.3.2. Effect on
PTZ induced convulsions
In PTZ induced convulsions both the extracts and oils were found to have no
effect on onset of convulsions when PTZ was injected to the mice (data not shown).
VVW02 and VVDR03 were found to produce dose dependent effect in delaying onset

of death but the effect was significant at 80 mg/kg in comparison to control and other
groups (Table 3.1.). VVDR03 delayed the time by 40.8 % at 80 mg/kg while VVW02
increased the time for onset of death by 30.4 %.
In case of valerian oils a dose dependent response was obtained with V-PA
and an inverted U shaped curve was obtained with V-MA but only 40mg/kg dose
significantly delayed the onset of death with respect to control group. When V-PA
was evaluated for effect on PTZ-induced convulsions then only 80 mg/kg dose of V-
PA produced significant delay in onset of death (73.9 %) (Table 3.2.).
The delay in onset of death is similar for VVDR03 and VVW02 as there is no
statisitical significant difference between them. Similarly V-PA was found to be more
effective in delaying the onset of death than V-MA. And out of all four V-PA was
most effective in delaying onset of death after PTZ injection.
3.3.3. Effect on elevated plus maze
Oral administration of both the extracts and oils at doses 10, 20 & 40 mg/kg
were found to have no anxiolytic effect in plus maze after 1 hr of administration
(Table 3.3. & 3.4.). Similarly no anxiolytic effect was found when the above drugs
were administered daily for 14 days and then tested in elevated plus maze (Table 3.5.
& 3.6.).
3.3.4. Effect on rota rod apparatus
All the doses (10, 20 and 40 mg/kg p.o) of VVDR03, VVW02, V-PA and V-
MA were found to have no effect on the time of fall in rota rod test suggesting that at
these doses valerian extracts and oils do not have muscle relaxant effect (Fig. 3.1.,
3.2., 3.3. & 3.4.). Diazepam the standard drug produced a significant decrease in time
of fall (49 sec) after intraperitoneal injection, depicting skeletal muscle relaxant
effect.

Table 3.1. Effect of different doses of VVDR03 and VW02 on convulsions induced by pentylenetetrazole. * denotes significance at p <0.05 in comparison to vehicle treated group
Treatment Dose (mg/kg) Onset of death (secs)
Vehicle 743 ±80
VVDR03 20 437±35
VVDR03 40 581 ±54
VVDR03 80 1257 ±140*
VVW02 20 566±50
VVW02 40 740.3±30
VVW02 80 1068±90*
Diazepam 1 No death
Table 3.2. Effect of different doses of V-PA and V-MA on convulsions induced by pentylenetetrazole. * denotes significance at p <0.05 in comparison to vehicle treated group
Treatment Dose (mg/kg) Onset of death (secs)
Vehicle 513±69
V-PA 20 323±67.9
V-PA 40 592.2±23
V-PA 80 1972±118*
V-MA 20 538±28
V-MA 40 735.7±39.6 *
V-MA 80 360±50
Diazepam 1 No death

Table 3.3. Effect of single administration different doses of VVDR03 and VVW02 on elevated plus maze
Treatment Dose Number of entries Time duration (secs) (mg/kg) (Closed arm) (Closed arm)
Vehicle 8±2 237.3±19.7
VVDR03 10 5±1.2 257±10.0
VVDR03 20 6±0.47 230±20.0
VVDR03 40 4±1 260±11.0
VVW02 10 3.8±1.39 265.2±14.7
VVW02 20 4±1.67 276±6.84
VVW02 40 4.4 ±1.83 248.4±42.4
Table 3.4. Effect of single administration of different doses of V-PA and V-MA on
elevated plus maze
Treatment Dose Number of entries Time duration (secs) (mg/kg) (Closed arm) (Closed arm)
Vehicle 5.3±0.3 211±42.8
V-PA 10 6.0±0.9 260±13
V-PA 20 5.3±1.7 262±23
V-PA 40 4.3± 1.3 251.3±31.4
V-MA 10 5.0±0.9 242±28
V-MA 20 4.6 ±1 237±48.5
V-MA 40 4± 1 271±3

Table 3.5. Effect of different doses of VVDR03 and VVW02 on elevated plus maze after 14 days of dosing.
Treatment Dose Number of entries Time duration (secs) (mg/kg) (Closed arm) (Closed arm)
Vehicle 8±2 237.3±19.7
VVDR03 10 4.2±1.7 222.6±13
VVDR03 20 6±2.1 211±9.8
VVDR03 40 9± 3.8 234±15.6
VVW02 10 5.2±0.47 209±14.3
VVW02 20 9.5±2.17 236±17.7
VVW02 40 6.6 ±1.8 242±16.1
Table 3.6. Effect of different doses of V-PA and V-MA in plus maze after 14 days of dosing Treatment Dose Number of entries Time duration (secs) (mg/kg) (Closed arm) (Closed arm)
Vehicle 3.6±1.2 230.7±19.7
V-PA 10 5±1.9 260±13.0
V-PA 20 3.4±0.7 278.2±9.8
V-PA 40 4.6±1.9 228.2±25.3
V-MA 10 6±1.2 222±28.0
V-MA 20 5±1.3 209.2±39.6
V-MA 40 7.2±2 257.8±10.6

0
50
100
150
200
250
300
350
Vehicle VVDR03(40)
VVDR03(20)
VVDR03(10)
Diazepam(1)
Tim
e of
fall
(sec
s)
0 min60 min
*
Fig. 3.1. Effect of single oral administration of VVDR03 (10, 20 & 40 mg/kg) on rota rod apparatus. * denotes significance at p <0.05 in comparison to 0 min reading
0
50
100
150
200
250
300
350
Vehicle VVW02(40)
VVW02(20)
VVW02(10)
Diazepam(1)
Tim
e of
fall
(sec
s)
0 min60 min
*
Fig. 3.2. Effect of single oral administration of VVW02 (10, 20 & 40 mg/kg) on rota
rod apparatus. * denotes significance at p <0.05 in comparison to 0 min reading

0
50
100
150
200
250
300
350
Vehicle V-PA(40) V-PA(20) V-PA (10) Diazepam(1)
Tim
e of
fall
(sec
s)
0 min60 min
*
Fig. 3.3. Effect of single oral administration of V-PA (10, 20 & 40 mg/kg) on rota rod apparatus. * denotes significance at p <0.05 in comparison to 0 min reading
0
50
100
150
200
250
300
350
Vehicle V-MA(40) V-MA(20) V-MA (10) Diazepam(1)
Tim
e of
fall
(sec
s)
0 min60 min
*
Fig. 3.4. Effect of single oral administration of V-MA (10, 20 & 40 mg/kg) on rota rod apparatus. * denotes significance at p <0.05 in comparison to 0 min reading

3.4. DISCUSSION
The present study reports some neuropharmacological activities of valerian
extracts and oils in mice. Epilepsy is a common neurological disorder affecting about
0.5-1 % of the world’s population (Hackinski, 1998). The most popular and widely
used animal seizure models are the traditional Maximal Electroshock Seizures and
PTZ tests. Prevention of seizures induced by PTZ in laboratory animals is the most
commonly used preliminary screening test for characterizing potential anticonvulsant
drugs. Though the PTZ test predicts activity against absence seizures, the underlying
neuronal abnormality is poorly understand. Diminution of brain GABA level has been
reported after sub convulsive dose of PTZ (Ha et al., 2000). The MES test is
considered to be a predictor of likely therapeutic efficacy against generalized tonic-
clonic seizures. By contrast, the PTZ assay has been used preliminarily to evaluate
antiepileptic drugs. A large body of experimental evidences supports the involvement
of γ-Amino butyric acid (GABA) in seizures. GABA is the principal inhibitory
neurotransmitter in the cerebral cortex maintaining the inhibitory tone that
counterbalances neurons excitation. When this balance is disturbed, seizures ensue.
Reduction in GABA-mediated inhibitory activity of glutamate decarboxylase has
been reported in studies of human epileptic brain tissues (David, 2001). Glutamate
concentration increases before seizure onset and is found to be highest in the epileptic
hippocampus than non-epileptic hippocampus while GABA levels increased during
seizures was greater in non-epileptic hippocampus than in epileptic hippocampus
showing decreased levels of GABA in epilepsy (During and Spancer, 1993). The
GABAA receptor is responsible for most fast inhibitory neurotransmission in the
central nervous system. Consequently, this receptor has been targeted for the
pharmacological control of anxiety, sleep, and epilepsy. Numerous natural and
synthetic compounds interact with the GABAA receptor at distinct, yet incompletely
defined, sites. These compounds include barbiturates, benzodiazepines, neurosteroids
and picrotoxin (Sieghart, 1992; Smith and Olsen, 1995). GABAA receptor agonists as
well as drugs, which allosterically modulate the GABA receptor channel complex, are
therapeutically active against convulsive seizures (David, 2001).

In our study valerian extracts and oils were found to have delayed onset of
death in comparison to vehicle treated group while no effect was produced on onset of
convulsions and it failed to produce protection against mortality. There is an assertion
also that most drugs with anticonvulsant activity do not counteract pentylenetetrazole
seizures but only retard them (Loscher et al., 1991). We found that both VVDR03 and
VVW02 produced a dose dependent effect in delaying onset of death and the effect
being significant at 80 mg/kg dose in comparison to control group. But none of the
dose prolonged the time of onset of convulsions as compared to control group. The
delay in onset of death produced by valerian extracts may result from the effect of its
components on inhibitory neurotransmitter GABA (Menini et al., 1993; Santos et
al.,1994a, 1994 b). The effect produced by VVDR03 and VVW02 is comparable. It is
suggested that different constituents of valerian extract interact with the GABA
system in the brain like inhibition of GABA transaminase, interaction with
GABA/benzodiazepine receptors and interference in uptake and release of GABA in
synaptosomes (Morazzoni and Bombardell, 1995, Houghton, 1999). GABA is the
major inhibitory neurotransmitter in the brain and the inhibition of its
neurotransmission has thought to be an underlying factor in epilepsy (Olsen, 1981).
Standard antiepileptic drugs, phenobarbitone and diazepam are thought to produce
their antiepileptic effects by enhancing GABA neurotransmission. This also may be
responsible for anticonvulsant and sedative action produced by V. wallichii.
Overproduction of NO has been linked to neurotoxicity during ischemia, some
forms of neurodegenerative brain diseases, and induction of seizures (Garthwaite,
1991). Nitric oxide (NO) has been linked to epileptic activity through the formation of
cyclic GMP (cGMP). Stimulation of the brain N-methyl-D-aspartate (NMDA)
receptors with glutamate or excitatory amino acids increases calcium influx, which
results in activation of cNOS and formation of NO. NO in turn activates guanylate
cyclase to synthesize cGMP, which is assumed to initiate seizures (Garthwaite, 1991).
NO modulates experimentally induced seizures in a complex manner. In addition to
stimulation of cGMP, NO has several other actions: it blocks NMDA receptors in a
negative feedback manner, thereby attenuating excitable activity (Manzoni et al.,

1992); promotes and suppresses glutamate release (Pelligrino et al., 1996) and reduces
the receptor activity of the inhibitory γ-aminobutyric acid (GABA) neurotransmitter
(Robello et al., 1996). PTZ induction of seizures may be related to the antagonistic
activity of the compound at the GABA-A receptor and to its activation of the NMDA
receptor (Kaputlu and Uzbay, 1997). Thus, the induced NO may augment the
capability of PTZ to induce seizures at both pathways. In case of valerian oils V-PA
produced a dose dependent effect and the results being significant at 80 mg/kg in
comparison to control group. V-MA produced an inverted U shaped curve in which
the effect produced at 40 mg/kg is significant in comparison to control group.
Valerian oil V-PA was found to be most effective in delaying onset of death in PTZ-
induced convulsions. Valerian oils may be effective because of terpenes present in
them as some researchers have reported anticonvulsant activity of monoterpenes
(Librowski et al., 2000). Therefore it seems that antiseizure profile of V. wallichii oils
may be related in part to terpenoids present in them like α-pinene, camphene and
terpineol (Arora and Arora, 1963) and the terpenes might be mediating anticonvulsant
effect by affecting nitric oxide pathway (Ahn et al., 2003). In another study done by
us we found that valerian oils produced an antidepressant effect mediated by NO
pathway and a similar inverted U-shaped activity curve or biphasic dose response
curve was observed with V-MA in producing antidepressant effect. Such biphasic
curve has been reported in other studies and it may be due to facilitatory and
inhibitory effect of NO on the NMDA receptor. In addition such mechanisms may
help explain why so many contradictory results have been found suggesting that NO
either facilitates or inhibits NMDA receptor mediated events such as seizures
(Rundfeldt et al., 1995), neurotoxicity (Weissman et al., 1992), anxiolysis and
nociception (Harkin et al., 1999).
The elevated plus maze (EPM) is a rodent model of anxiety that is used as a
screening test for putative anxiolytic compounds and as a general research tool in
neurobiological anxiety research. The test setting consists of a plus-shaped apparatus
with two open and two enclosed arms, each with an open roof, elevated 15 cm from
the floor. The model is based on rodents aversion for open spaces. This aversion leads

to the behavior termed thigmotaxis, which involves avoidance of open areas by
confining movements to enclosed spaces or to the edges of a bounded space (Pellow
et al., 1985; Treit et al., 1993; Rodgers, 1997; Carobrez and Bertoglio, 2005). So the
total time duration spent in closed arm was noted and compared with that of control
and standard drug treated group. Valerian extracts and oils were not found to have
neither anxiolytic nor muscle relaxant effect at the tested doses (10, 20 and 40 mg/kg
p.o).

CHAPTER-4
Screening of Valeriana wallichii chemotypes for Antidepressant effect

4.1. INTRODUCTION
Depression has attracted strong attention as a major public health problem
worldwide that’s why prevention and treatment of depression are urgent issues need
to be resolved. Exposure to chronic stress is one of the major factor that provoke
depressive illness (Sakakibara et al., 2008). The abnormalities of noradrenergic and
serotonergic transmission are known to cause mental depression. Subsequently a
dopamine hypothesis of depression was also put forward (Dhir and Kulkarni, 2007).
Biochemical theories of mood disorders have focused on the biogenic amines but also
the importance of gamma butyric acid (GABA) in the neurochemical pathophysiology
of depression has been demonstrated (Petty, 1995). Clinical data also indicate that a
decreased GABA function accompanies depression and there is accumulating
evidence which implicates a GABAergic dysfunction in depression (Kubacka et al.,
2006). On the other hand there are several evidences which suggest that deficiency of
behaviors in depressive illness is associated with an increase in presynaptic inhibitory
α2 receptors leading to a secondary decrease in norepinephrine (NE) release (David,
2006). Drugs that block α2 receptors and increase NE release have been shown to have
antidepressant effect in various animal models of depression.
Previous studies have demonstrated that antagonists at the NMDA receptor are
as efficacious as tricyclic antidepressants in pre-clinical antidepressant screening
procedures and in blocking or reversing the behavioral deficits associated with animal
analogs of major depressive symptomatology. The NMDA receptor complex gates
Ca2+ (Southam and Garthwaite, 1993), which interacts with calmodulin to
subsequently activate nitric oxide (NO) synthase. Since NMDA receptor antagonists
possess significant antidepressant-like properties (Trullas, 1997), it is hypothesized
that NO synthase inhibitors would possess antidepressant properties and there are
some reports supporting this hypothesis (Harkin et al., 1999). So L-arginine-nitric
oxide (NO)-cyclic guanosine monophosphate (cGMP) is an important signaling
pathway involved in depression (Dhir and Kulkarni, 2008).

So whatever is the cause there is a wide variety of antidepressant drugs
available to treat depression but most of the synthetic drugs are not without side
effects. On the other hand, drugs obtained from natural sources are perceived to have
the least risk and low side-effect profiles, while having the ability to cure psychiatric
disorders in much the same way as their synthetic counterparts. Ayurveda, the ancient
traditional system of medicine, mentions a number of single and compound drug
formulations of plant origin that are used for the treatment of psychiatric disorders.
Recently several clinical studies have presented some biological activities of valerian
root extracts and introduced the plant as anti-HIV (Murukami et al., 2002), sleep aid
(Francis and Dempster, 2002; Fernandez et al., 2004), tranquilizer (Houghton, 1999),
antidepressant (Andreatini et al., 2002; Miyasaka et al., 2006; Hattesohl et al., 2008)
agent. So the aim of the present study is to evaluate the effect of Valeriana wallichii
chemotypes on forced swim test and to explore the mechanism responsible for this
effect as there are few studies done on antidepressant-like effect of herbal
formulations like Trans-a1 (Shalam et al., 2007) and Sumind (Nanjappa et al., 2007)
containing Valeriana wallichii as one of the ingredient.
4.2. MATERIALS AND METHODS
4.2.1. Plant material and extraction
Same as in section 3.2.1. of chapter 3.
4.2.2. Animals
Same as in section 3.2.2. of chapter 3.
4.2.3. Drugs and treatment
Imipramine, L-NAME (N-omega-nitro-l-arginine-methyl-ester) a nitric oxide
synthase inhibitor were used. Extracts VVDR03, VVW02 and their oils V-PA and V-
MA were used at doses of 10, 20 and 40 mg/kg. Both the extracts and oils were
administered orally as an emulsion in 2% Tween-80 to the animals one hour prior to

forced swim test (FST). And then effect of extracts and oils in combination with L-
NAME (10 mg/kg, intraperitoneally) was studied in FST for mechanistic study.
4.2.4. Procedure
Locomotor activity
Six groups of animals were fed orally with VWDRO3 and VVWO2 at a dose
of 10, 20 and 40 mg/kg and another six groups were given 10, 20 and 40 mg/kg dose
of V-PA and V-MA. One group was given vehicle and acted as control and another
group received imipramine (10mg/kg i.p). The locomotor activity was monitored after
one hour using actophotometer (IMCORP, India). The animals were individually
placed in activity meter for three minutes before counting of actual locomotor activity
for the next 5 minutes. The locomotor activity was expressed in terms of total
photobeams counts, 5 min per animal (Reddy, 1998). Percentage increase or decrease
in locomotion was calculated.
Forced swim test
Briefly, a group of 64 mice was randomly divided into 14 groups and treated
as follows:
Group 1: Vehicle treated (control group)
Group 2-4: VVDR03 (10, 40 and 40 mg/kg p.o.)
Group 5-7: VVW02 (10, 20 and 40 mg/kg p.o.)
Group 8-10: V-PA (10, 20 and 40 mg/kg p.o.)
Group 11-13: V-MA (10, 20 and 40 mg/kg p.o)
Group 14: Impiramine (10 m/kg, p.o.)
Then one hour after the treatment mice were individually forced to swim
inside a rectangular glass jar (25x12x25 cm3) containing 15 cm of water maintained at
23–25°C. After the initial 2–3 min of vigorous activity the animals showed period of
immobility by floating with minimum movements. An animal is considered to be
immobile whenever it remained floating passively in the water in a slightly hunched
but upright position, its nose above the water surface (Fig. 4.1.). The total immobility

period for the period of 6 minutes was recorded with the help of stop-watch (Kulkarni
and Mehta, 1985).
Fig.4.1. Rectangular jar depicting the mobile (left) and immobile (right) postures of the animals
Extracts and oils in combination with L-NAME in forced swim test
In order to explore mechanism of action of valerian in decreasing immobility
period in FST, mice received following treatments:
Group 1: Vehicle treated (control group)
Group 2: L-NAME (10 mg/kg i.p.)
Group 3: VVDR03 (10 mg/kg, p.o.) + L-NAME (10 mg/kg i.p.)
Group 4: VVW02 (10 mg/kg, p.o.) + L-NAME (10 mg/kg i.p.)
Group 5: V-PA (10 mg/kg, p.o.) + L-NAME (10 mg/kg i.p.)
Group 6: V-MA (10 mg/kg, p.o.) + L-NAME (10 mg/kg i.p.)
All the test drugs were administered orally. L-NAME was given half an hour
after administration of test drugs and then mice were allowed to swim in Porsolt
apparatus 30 minutes after intraperitoneal injection of L-NAME.

4.2.5. Statistical analysis
The data were expressed as mean± SEM of 6 animals. Results were analysed
statistically by One-way ANOVA followed by Tukey’s multiple comparison using
sigma stat software. The difference was considered statistically significant if p<0.05.
4.3. RESULTS
4.3.1. Acute toxicity
Same as in section 3.3.1. of chapter 3.
4.3.2. Effect on locomotor activity
None of the dose of VVDR03 significantly decreased the locomotor activity
while VVW02 at 40 mg/kg produced significant decrease (45.8%) in locomotor
activity (Table 4.1.). V-MA did not alter the spontaneous locomotor activity except at
dose 40 mg/kg which significantly produced 44.9 % decrease in this parameter (Table
4.2.). While 40 mg/kg dose of V-PA produced a decrease in locomotor activity but the
effect was not significant. So the results depict that none of the doses of VVDR03 and
V-PA altered locomotor activity in mice. Imipramine the standard drug was also
devoid of any effect on locomotor activity.
4.3.3. Effect of acute dosing of extracts in FST
When single oral administration of different doses of VVDR03 and VVW02
were studied for effect in FST, 40 mg/kg dose of both the extracts (Fig. 4.2. & Fig.
4.3.) significantly decreased immobility period as compared to vehicle treated group.
The effect was found to be more pronounced with VVW02 (57% decrease in
immobility) than VVDR03 (32.7 % decrease in immobility) (statistical significant)
and group treated with standard drug imipramine (36.2 % decrease).

4.3.4. Effect of acute dosing of oils in FST
Different doses of V-PA produced a dose response curve i.e on increasing the
dose response was increased (Fig. 4.4.). The effect being significant at 40 mg/kg
(57.6% decrease) and 20 mg/kg (46.9% decrease). Whereas different doses of V-MA
produced an inverted U shaped curve, the effect being significant at 40 mg/kg and 10
mg/kg dose (Fig. 4.5.). 10 mg/kg produced 39.7% decrease while 40 mg/kg dose
produced 58% decrease in immobility as compared to control group. When compared
statistically both the oils were found equally effective in producing antidepressant
effect.
4.3.5. Effect of NOS inhibitor L-NAME on the action of VVDR03 and VVW02
(10 mg/kg) in FST
L-NAME was combined with 10 mg/kg dose of VVW02 and VVDR03 and
the modulation by L-NAME was then studied after half and hour of L-NAME
injection. L-NAME per se did not have any effect on the immobility period in FST
neither it potentiated the effect of VVDR03 and VVW02 (Fig 4.6.). The combination
did not change the locomotor activity (results not shown).
4.3.6. Effect of NOS inhibitor L-NAME on the action of V-PA and V-MA (10
mg/kg) in FST
To assess the modulatory effect of NOS inhibitor (L-NAME) on FST, V-PA,
V-MA (10 mg/kg p.o) was combined with L-NAME (10 mg/kg i.p). L-NAME per se
did not have any effect on the immobility period in FST. However, when L-NAME
was given 30 min after V-PA and V-MA it significantly potentiated the effect of V-
PA and V-MA (Fig. 4.7.). The combination did not increase the locomotor activity
(results not shown).

Table 4.1. Effect of different doses of extracts (VVDR03 & VVW02) of Valeriana wallichii in actophotometer apparatus in mice. (*) show significance at P<0.05 vs. vehicle group. Value in parenthesis is % decrease in locomotor counts. Treatment Dose (mg/kg. p.o) Mean ambulatory scores
Vehicle 240 ±32.4
VVDR03 10 235±64
VVDR03 20 139.6±65.4
VVDR03 40 210 ±58.5
VVW02 10 188.75±10
VVW02 20 105.3±62.7
VVW02 40 139±21 (45.8) *
Imipramine 10 200 ±6
Table 4.2. Effect of different doses of oils (V-MA & V-PA) of Valeriana wallichii in actophotometer apparatus in mice. (*) show significance at P<0.05 vs. vehicle group. Value in parenthesis is % decrease in locomotor counts.
Treatment Dose (mg/kg. p.o) Mean ambulatory scores
Vehicle 414 ±13.6
V-PA 10 383 ±45.4
V-PA 20 415 ±62.9
V-PA 40 319 ±36
V-MA 10 391±40
V-MA 20 324± 20.3
V-MA 40 228 ±48.7*(44.9)
Imipramine 10 300 ±40

Fig. 4.2. Effect of valerian extract VVW02 on the immobility period induced by FST. VVW02 at different doses were administered orally 60 min before FST challenge. *P<0.05 vs. vehicle group.
Fig. 4.3. Effect of valerian extract VVDR03 on the immobility period induced by FST. VVDR03 at different doses were administered orally 60 min before FST challenge. *P<0.05 vs. vehicle group.

Fig. 4.4. Effect of V-PA on the immobility period induced by FST. V-PA at different doses was administered orally 60 min before FST challenge. *P<0.05 vs. vehicle group.
Fig. 4.5. Effect of V-MA on the immobility period induced by FST. V-MA at different doses was administered orally 60 min before FST challenge. *P<0.05 vs. vehicle group.

Fig. 4.6. Effect of 10 mg/kg dose of VVDR03 & VVW02 in combination with L-NAME (10mg/kg i.p.).
Fig. 4.7. Effect of 10 mg/kg dose of V-PA & V-MA in combination with L-NAME (10mg/kg i.p.). a denotes significance at P < 0.05 in comparison to L-NAME per se, b denotes significance at P < 0.05 in comparison to V-PA per se and c denotes significance at P < 0.05 in comparison to V-MA per se. # denotes significance at P < 0.05 in comparison to vehicle treated group.

4.4. DISCUSSION
One of the purposes of this study was to evaluate the antidepressant-like effect
of valeriana by an animal model for depression. We employed the forced swimming
test (FST) on mice. The immobility displayed by rodents when subjected to an
unavoidable stress such as forced swimming is thought to reflect state of despair or
lowered mood, which reflects depressive illness in humans. Additionally, immobility
time has been shown to be reduced by treatment with antidepressant drugs (Porsolt,
1981). The FST was designed to detect potential antidepressant compounds based on
the abilities of clinically effective antidepressants to reduce the immobility in FST
(Porsolt et al., 1977). Though the FST does not adequately reflect the symptomalogy
of human depression, it appears to have a higher predictive validity compared to other
animal models Additionally, it is sufficiently specific, since it discriminates
antidepressants from neuroleptics and anxiolytics (Borsini and Meli, 1988; Willner,
1984). The present study has shown that valerian oils and extracts posses
antidepressant effect in FST.
In our study DCM extracts of both the chemotypes (VVW02 and VVDR03) at
40mg/kg dose significantly reduced the immobility time in FST in acute study.
VVW02 was found to produce 57 % decrease in immobility at 40 mg/kg as compared
to vehicle treated control group (Fig. 4.2.) while 40 mg/kg dose of VVDR03 produced
32.7% decrease in immobility which is comparable to the effect produced by standard
drug imipramine (Fig. 4.3.). In case of valerian oils V-MA produced significant
decrease in immobility at dosage 10 and 40 mg/kg while 20 mg/kg did not affect the
immobility period (Fig.4.5.) and produced an inverted U shaped curve. V-PA in single
dose study produced a dose dependent effect i.e. 20 and 40 mg/kg dose of V-PA
produced a significant decrease in immobility as compared to group receiving 10
mg/kg and control group (Fig.4.4.). Psychostimulants, which exert an indiscriminate
motor stimulating activity, have previously been shown to have efficacy at decreasing
immobility in FST, but no antidepressant activity (Porsolt, 1981). So in order to

exclude a false positive result, we employed an additional test to check motor
stimulating activity of extracts and oils. In the present study VVW02 and V-MA
lowered the locomotor activity at 40 mg/kg dose only whereas no alteration was seen
with VVDR03 and V-PA. Therefore the reduction in immobility by valerian extract
time can be attributed to an inherent antidepressant like effect.
A study depicted antidepressant effect of kessyl glycol diacetate, a valepotriate
possibly due to blockade of monoamine uptake (Miller and Murray, 1998). Methanol
extracts of Japanese valerian (Valeriana fauriei) also exhibited strong antidepressant
activity in the forced swimming test in mice (Oshima and Matsuoka, 1995). The
active fraction was identified as a sesquiterpenoid called a kessyl alcohol and its
activity compared favorably to that of imipramine. These studies support earlier
findings of a Japanese valerian root extract which significantly inhibited immobility
induced by a forced swimming test in rats (Sakomoto et al., 1992). Another recent
study showed anti-anxiety and anti-depressant benefits of valerian but failed to show
sedative or muscle relaxant benefits (Hattesohl et al., 2008). A study done on
Japanese valerian confirms that valerian extract and imipramine (a tricyclic anti-
depressant) significantly inhibited immobility in FST and reversed reserpine-induced
in mice, a test which suggests antidepressant actions (Sakamoto et al., 1992).
During the past few years, nitric oxide (NO), an inorganic free radical has
emerged as an important signal and effector molecule in several biological processes
including vasodilatation, inflammation and neurotransmission (Garthwaite, 1991;
Lowenstein, 1994; Moncada, 1991). NO is produced in neurons from the L-arginine
by the calmodium-dependent enzyme. NO synthase and NO fulfill some of the most
important criteria for a neurotransmitter (Knowles, 1994; Dawson, 1994). An
important target of NO is soluble guanlyl cyclase which is activated by NO binding to
heme part of the enzyme, causing elevation in cGMP. It appears that in different
systems, NO may be able to modulate vesicular release of neurotransmitter in either
direction or not all depending on the coincident level of presynaptic activity and NO
concentration (Garthwaite, 1995). In order to explore the involvement of nitric oxide

(NO) signaling pathway in the antidepressant-like effect of valerian extracts and oils,
10 mg/kg dose of VVDR03, VVW02, V-PA and V-MA were combined with L-
NAME (10 mg/kg) (a selective nitric oxide synthase inhibitor) (Babbedge et al., 1993;
Re es et al., 1990) and then tested for alteration in immobility in FST. No potentiation
was found in case of valerian extracts. It was found that L-NAME per se did not alter
the immobility period compared to control whereas it potentiated the action of V-PA
and V-MA significantly. The potentiation was even significant to group receiving L-
NAME only. The L-NAME was devoid of locomotor stimulatory property at 10
mg/kg dose which is in accordance to earlier study (Harkin et al., 1999). Moreover, a
study depicts that stimulation of NO synthase not only antagonized the behavioral
effect of NO synthase inhibitors but also of the prototypic antidepressant, imipramine.
So it may be possible that one or more component of essential oil (sesquiterpenes) of
Valeriana might be acting as NOS inhibitor as various studies report inhibition of NO
synthesis in lipopolysac-charide (LPS)-activated RAW 264.7 cells by sesquiterpenes
(Ahn et al., 2003; Motai and Kitanaka, 2005). In other studies, also sesquiterpenoids
have been found to have suppressive effect on NOS and COX-2 activity (Lee et al.,
2002; Yoon et al., 2008).
An inverted U-shaped activity curve or biphasic dose response curve observed
with V-MA has been reported in previous studies and it may be due to facilitatory and
inhibitory effect of NO on the NMDA receptor. NO regulates NMDA receptor
activity in a biphasic manner playing both a positive (via activation of guanylate
cyclase) and negative (via feedback effects on the NMDA receptor resulting in
decreased NMDA receptor and NO synthase activity) modulatory role in NMDA
receptor mediated events (Lei et al., 1992; Manzoni and Bockaert., 1993). In addition
such mechanisms may help explain why so many contradictory results have been
found suggesting that NO either facilitates or inhibits NMDA receptor mediated
events such as seizures (Rundfeldt et al., 1995), nociception, neurotoxicity (Weissman
et al., 1992) and anxiolysis (Harkin et al., 1999). Moreover, a study depicts that NO
has a dual role in the modulation of depression (Da Silva et al., 2000).

From our results it can be speculated that the level of NOS inhibition elicited
by valerian oils leads to a critical NO concentration which in turn, alters the vesicular
release of neurotransmitter like norepinephrine involved in depression. There are
other studies which also show that chronic blockade of nitric oxide synthesis elevates
plasma levels of catecholamines and their metabolites at rest and during stress in rats
(Richard et al., 1997). So we can say that the nitric oxide signaling pathway is
involved in the antidepressant-like effect of Valerian oils while valerian extracts
might be producing antidepressant by potentiating the action of GABA. As it has been
discussed that GABAergic dysfunction is also involved in depression (Kubacka et al.,
2006) and valepotriates and other constituents present in the extracts tends to enhance
the action of GABA (Riedel, 1982).

CHAPTER-5
Pharmacological and Neurobiochemical Evidence for Antidepressant like Effect of
Valeriana wallichii chemotypes

5.1. INTRODUCTION
Major depression has a lifetime prevalence rate of 17% (Andrews and
Nemeroff, 1994). Depression is more common in women and is seen throughout the
life. A major depressive disorder may occur as either a single or recurrent event.
Despite the advent of new molecules in the pharmacotherapy of depression, it is
unfortunate that this disorder goes undiagnosed and untreated. Although the currently
prescribed molecules provide some improvement in the clinical condition of the
patient, it is at the cost of having to bear the burden of their adverse effects (Stahl,
1998; Tripathi, 2005; Hardman et al., 2001). On the other hand, drugs obtained from
natural sources are perceived to have the least risk and low side-effect profiles, while
having the ability to cure psychiatric disorders in much the same way as their
synthetic counterparts. Ayurveda, the ancient traditional system of medicine,
mentions a number of single and compound drug formulations of plant origin that are
used for the treatment of psychiatric disorders. Patients with depression have
symptoms that generally reflect changes in the brain neurotransmitters serotonin,
norepinephrine, and dopamine. The "dysregulation hypothesis" proposes that
depression may be due to abnormalities in the pattern and responsiveness of
neurotransmitter systems that result from modifications in the receptor sensitivity and
function. Antidepressants are thought to increase levels of norepinephrine, serotonin,
or dopamine at the synapse to restore equilibrium to neurotransmitter activity.
The previous study revealed antidepressant effect of Valerian oils and extracts.
So this study aims at evaluating the alterations in neurotransmitter levels after 14 days
dosing of Valeriana wallichii extracts and oils.
5.2. MATERIALS AND METHODS
5.2.1. Plant material and extraction
Same as in section 3.2.1. of chapter 3.
5.2.2. Animals
Same as in section 3.2.2. of chapter 3.

5.2.3. Drugs and treatment
Extracts VVDR03, VVWO2 and their oils V-PA and V-MA were used at
doses of 10, 20 and 40 mg/kg. Both the extracts and oils were administered orally as
suspension in Tween-80 to the animals for 14 days and then the effect in forced swim
test (FST) was seen one hour after the drug administration on 14th day.
5.2.4. Procedure
Forced swim test
Briefly, a group of 64 mice was randomly divided into 14 groups, each group
containing six animals and treated as follows: one group was administered
imipramine (10mg/kg), three groups were administered VVDR03 (10, 20, 40 mg/kg,
p.o), three groups received VVW02 (10, 20, 40 mg/kg, p.o), three groups received V-
PA (10, 20, 40 mg/kg, p.o) and other three groups received V-MA (10, 20, 40 mg/kg.
p.o). One group receiving vehicle served as control. All the groups received the
treatment continuously for two week. Then one hour after the last day treatment mice
were individually forced to swim inside a rectangular glass jar (25×12×25 cm3)
containing 15 cm of water maintained at 23–25°C. After the initial 2–3 min of
vigorous activity the animals showed period of immobility by floating with minimum
movements. Depression produced in animal is characterized by increased immobility
period. An animal is considered to be immobile whenever it remained floating
passively in the water in a slightly hunched but upright position, its nose above the
water surface. The total immobility period for the period of 6 minutes was recorded
with the help of stop-watch (Kulkarni and Mehta, 1985).
Measurement of biogenic amines
Biogenic amines (dopamine and norepinephrine) were estimated by high
performance liquid chromatography (HPLC) with electrochemical detector (ECD)
(Beyer et al., 2002). Waters® standard system consisting of a high pressure isocratic
pump, a 20µl sample injector valve, C18 reverse phase column and electrochemical
detector were used. Data were recorded and analyzed with the help of Empower
software. Mobile phase consisting of 0.15M NaH2PO4, 0.25mM EDTA, 1.75mM 1-

octane sulfonic acid, 2% isopropanol and 4% methanol (pH 4.8). Electrochemical
conditions for the experiment were +0.800 V, sensitivity ranges from 1 to 100 nA.
Separation was carried out at a flow rate of 1 ml/min. Samples (20µl) were injected
manually. Brain samples were homogenized in homogenizing solution containing
0.1M perchloric acid. After that samples were centrifuged at 24,000×g for 15 min.
The supernatant was further filtered through 0.25µm nylon filters before injecting in
the HPLC injection pump. Data were recorded and analyzed with the help of
Empower® software provided by Waters® (Beyer et al., 2002).
5.2.5. Statistical analysis
The data were expressed as mean± SEM of 6 animals. Results were analysed
statistically by One-way ANOVA followed by Tukey’s multiple comparison using
Jandel Sigma Stat Software. The difference was considered statistically significant at
p<0.05.
5.3. RESULTS
5.3.1. Acute toxicity
Same as in section 3.3.1 of chapter 3.
5.3.2. Effect of extracts in FST after 14 days of dosing
VVDR03 and VVW02 were found to produce a dose dependent response
curve in chronic study. 20 and 40 mg/kg dose of both the extracts decreased
immobility period significantly as compared to control group. The decrease being
30.8% and 55.9 % with VVW02 (Fig. 5.1) while VVDR03 (Fig. 5.2) produced 17.2
% and 45 % decrease in immobility in FST.
5.3.3. Effect of oils in FST after 14 days of dosing
Only 20 mg/kg dose of V-PA produced a significant decrease in immobility in
FST (Fig. 5.3.) as compared to vehicle treated group. V-PA (20 mg/kg) produced 69.6
% decrease which is much more marked than the effect produced by 40 mg/kg dose of

V-PA. 40 mg/kg dose of V-MA produced a significant decrease (70.9%) in
immobility while 20 mg/kg V-MA has no significant effect in FST (Fig. 5.4.).
5.3.4. Effect of extracts and oils on the neurotransmitter levels in mice brain
When checked for the alterations in the neurotransmitter levels after 14 days
of dosing 20 and 40 mg/kg dose of VVW02 increased norepinephrine levels dose
dependently and significantly. No alteration in dopamine levels is seen with any dose
of VVW02 (Fig. 5.5.). In case of VVDR03 both 20 and 40 mg/kg increased the
norepinephrine levels in whole brain significantly. The alterations being dose
dependent. None of the dose altered dopamine levels as compared to control except
20 mg/kg (Fig. 5.6.). 40 mg/kg and 20 mg/kg of V-PA increased norepinehrine levels
by 48.5 % and 84.96% respectively but the effect is significant at 20mg/kg dose (Fig.
5.7.). Similarly V-MA at 40 mg/kg produced a marked increase in norepinehrine
levels (Fig. 5.8.) while 20 mg/kg did not affect norepinehrine levels. None of the dose
of V-PA and V-MA altered dopamine levels in brain.
0
50
10 0
150
2 0 0
2 50
3 0 0
V ehicle V V W 0 2( 10 )
V V W 0 2( 2 0 )
V V W 0 2( 4 0 )
Mea
n im
mob
ility
per
iod
(sec
)
*
*
Fig. 5.1. Effect of VVW02 on the immobility period induced by FST. VVW02 at different doses was administered orally repeatedly for 14 days before FST challenge. *P<0.05 vs. vehicle group.

0
50
10 0
150
2 0 0
2 50
3 0 0
V ehicle V V D R 0 3( 10 )
V V D R 0 3( 2 0 )
V V D R 0 3( 4 0 )
Mea
n im
mob
ility
per
iod
(sec
)
*
*
Fig. 5.2. Effect of VVDR03 on the immobility period induced by FST. VVDR03 at different doses was administered orally repeatedly for 14 days before FST challenge. *P<0.05 vs. vehicle group.
0
50
10 0
150
2 0 0
2 50
V ehicle V - PA ( 10 ) V - PA ( 2 0 ) V - PA ( 4 0 )
Mea
n im
mob
ility
per
iod
(sec
)
* *
Fig. 5.3. Effect of V-PA on the immobility period induced by FST. V-PA at different doses was administered orally repeatedly for 14 days before FST challenge. *P<0.05 vs. vehicle group.
0
50
10 0
150
2 0 0
2 50
V ehicle V - M A ( 10 ) V - M A ( 2 0 ) V - M A ( 4 0 )Mea
n im
mob
ility
per
iod
(sec
)
*
Fig.5.4. Effect of V-MA on the immobility period induced by FST. V-MA at different doses was administered orally repeatedly for 14 days before FST challenge. *P<0.05 vs. vehicle group.

Fig. 5.5. Effect of different doses of VVW02 on the alteration in neurotransmitter levels in the mouse whole brain. VVW02 was administered for 14 days before sacrificing the animals. (*) P<0.05 as compared to vehicle treated group.
Fig. 5.6. Effect of different doses of VVDR03 on the alteration in neurotransmitter levels in the mouse whole brain. VVDR03 was administered for 14 days before sacrificing the animals. (*) P<0.05 as compared to vehicle treated group.

Fig. 5.7. Effect of different doses of V-PA on the alteration in neurotransmitter levels in the mouse whole brain. V-PA was administered for 14 days before sacrificing the animals. (*) P<0.05 as compared to vehicle treated group.
Fig. 5.8. Effect of different doses of V-MA on the alteration in neurotransmitter levels in the mouse whole brain. V-MA was administered for 14 days before sacrificing the animals. (*) P<0.05 as compared to vehicle treated group
5.4. DISCUSSION
In this chronic study repeated administration of VVW02 and VVDR03 for 14
days produced a dose dependent response curve (Fig. 5.1 & 5.2). The effect being
significant at 20 and 40 mg/kg for both the extracts and out of them VVW02 (Maaliol

chemotype) was found to more effective than VVDR03 (Patchouli alcohol
chemotype).
In case of oils, V-PA produced a U shaped curve and the effect was found to
be significant at 20mg/kg in comparison to control (Fig. 5.3). Such U-shaped activity
curve may be due to multiple receptor action and has also been reported for some
herbal medicines (Butterweck et al., 2000, Butterweck et al., 2001 & Sakakibara et
al., 2006). V-MA produced a dose dependent response curve and the effect being
significant at 40 mg/kg (Fig. 5.4).
When checked for the alterations in the neurotransmitter levels after 14 days
of dosing, 20 and 40 mg/kg dose of VVW02 increased norepinephrine levels dose
dependently and significantly. No alteration in dopamine levels was seen with any
dose of VVW02 (Fig. 5.5). Similarly 20 and 40 mg/kg dose of VVDR03 increased the
norepinephrine levels in whole brain significantly and dose dependently (Fig. 5.6).
None of the dose altered dopamine levels as compared to control except 20 mg/kg of
VVDR03. 20 mg/kg of V-PA increased norepinehrine levels significantly and V-MA
at 40 mg/kg produced a marked increase in norepinehrine levels (Fig. 5.7 & 5.8).
From the previous study done by us we found that a NOS inhibitor (L-NAME)
potentiated the antidepressant effect of valerian oils in FST depicting the alteration of
NO level in mice brain by valerian oils leading to antidepressant effect. So it appears
that in different systems, NO may be able to modulate vesicular release of
neurotransmitter in either direction or not all depending on the coincident level of
presynaptic activity and NO concentration (Garthwaite, 1995). There are other studies
which also show that blockade of nitric oxide synthesis elevates plasma levels of
catecholamines and their metabolites at rest and during stress in rats (Richard et al,
1997). So from our results it can be speculated that the level of nitric oxide synthase
(NOS) inhibition elicited by valerian oil leads to a critical NO concentration which in
turn, alters the vesicular release of neurotransmitter like norepinephrine involved in
depression and we can say that the nitric oxide signaling pathway is involved in the
antidepressant-like effect of V. wallichii oils. While in case of valerian extracts no

potentiation was seen with L-NAME which suggests that some other pathway is
involved in antidepressant effect of extracts (VVDR03 and VVW02).
Biochemical theories of mood disorders have focused on the biogenic amines
but the importance of gamma butyric acid (GABA) in the neurochemical
pathophysiology of depression has also been demonstrated (Petty, 1995). Clinical data
also indicate that a decreased GABA function accompanies depression and there are
accumulating evidences which implicates a GABAergic dysfunction in depression
(Kubacka et al., 2006). There are various studies showing that the constituents present
in valeriana enhance GABA levels (Morazzoni and Bombardell, 1995, Houghton,
1999). So we can hypothesize that valerian extracts might be altering norpeinephrine
levels in brain by enhancing GABA neurotransmission.

CHAPTER-6
Studies on Analgesic Activity of Valeriana wallichii chemotypes

6.1. INTRODUCTION
Pain is a sensorial modality which is universally understood as a signal of
disease and it is the most common symptom requiring treatment with analgesic
agents. So pain often has a protective function. Throughout history, man has used
many different forms of therapy for the relief of pain, among them; medicinal herbs
are highlighted due to their wide popular use. In the relief of pain, opiates are
generally considered to act on the central nervous system exercising their effects
through three opioid receptors (µ, κ and δ) and such drugs are especially important for
the treatment of chronic pain. Although morphine has reigned for centuries as the king
of painkillers, its rule hasn’t been totally benign. There are concerns about its
addictive properties and side effects, which include respiratory depression,
drowsiness, decreased gastrointestinal motility, nausea and several alterations of the
endocrine and autonomic nervous system. Natural products in general and medicinal
plants in particular, are believed to be an important source of new chemical
substances with potential therapeutic efficacy. Taking into account the most important
analgesic, prototypes (e.g. salicylic acid and morphine) were originally derived from
the plant sources. The study of plant species traditionally used as pain killers should
still be seen as a fruitful research strategy in the search of new analgesic and anti-
inflammatory drugs.
Valeriana wallichii or Indian Valerian is a plant that steeped in history. Its
original use in herbal medicine today is as sedative and calming agent. Valepotriates
are responsible for the sedative action of the plant. It is used as a carminative and
antispasmodic in hysteria and similar nervous manifestations. Spasmolytic effect of
the V. wallichii has been used in different gastrointestinal disorders such as diarrhea
and abdominal spasm (Holmes, 1989). The studies depict that mild myorelaxant
action of Valeriana is attributed to the valepotriates component of the herb (Dunaev et
al., 1987). The valepotriates isovaltrate and valtrate, and the essential oil compound
valeranone were observed to suppress the rhythmic contractions in a closed part of the
guinea-pig ileum in vivo (Hazelhoff et al., 1982). The same compounds and
didrovaltrate relaxed potassium stimulated contractures and inhibited BaCl2
contractions in guinea-pig ileum preparations in vitro. The inhibition of muscle

contractions by the valium chemicals valeranone and didrovaltrate were as potent as
papaverine. There is a report showing analgesic effect of dried leaves of V. jatamansi
at a dose of 2 mg (Shrivastava and Sisodia, 1970). It has potential ethnomedicinal
uses like V. officinalis but till date there is no evidence-based clinical report. In this
context, we aimed to investigate the medicinal plant, V. wallichii for its analgesic
effect.
6.2. MATERIAL AND METHODS
6.2.1. Plant material and its extraction
Same as in section 3.2.1. of chapter 3.
6.2.2. Animals
Same as in section 3.2.2. of chapter 3.
6.2.3. Drugs
Aspirin was procured from Panacea Biotech, Lalru, India. To the test drugs
(VVW02, VVDR03, V-PA and V-MA) one drop of Tween-80 was added and then
volume was made up with distilled water. Solution of aspirin was also prepared using
Tween-80.
6.2.4. Procedure
Effect of valerian extracts and oils in Acetic acid-induced writhing
Acetic acid (1%) was injected i.p. in mice and the writhing response
characterized by abdominal constriction and hind limb stretching was counted for 10
min (Kulkarni, 1999). V-PA, V-MA, VVDR03 and VVW02 at doses of 20, 40 and 80
mg/kg were administered orally to mice 1 hr before the test. Aspirin 100mg/kg, p.o.
was used as a standard for comparison while vehicle treated group was kept as
control. The number of writhings in these groups were recorded and compared with
control and standard groups.

Effect of valerian extracts and oils on tail-flick latency
Analgesia was assessed with analgesiometer (Techno Electronics, India).
Basal reaction time of animals to radiant heat was recorded by placing the tip (last 1-2
cm) of the tail on the radiant heat source. The tail withdrawal from the heat (flicking
response) was taken as the end point. The animals, which showed flicking response
within 3-5 secs, were selected for the study. A cut off period of 12 secs was observed
to avoid damage to the tail (Kulkarni, 1999). The measurement of tail flick latency
was recorded at 15, 30, 60, 120 and 180 min after administration of V-PA, V-MA,
VVDR03 and VVW02 (20, 40 and 80 mg/kg).
Effect of valerian extracts and oils in combination with aspirin in acetic acid
writhing
For mechanistic study sub-effective dose i.e 20 mg/kg dose of VVDR03,
VVW02, V-PA and V-MA was combined with 5 mg/kg dose of aspirin and was then
studied in acetic acid induced writhing. Aspirin was given intraperitoneally 30
minutes after the administration of test drugs and acetic acid was injected 30 minutes
after aspirin. Per se effect of 5 mg/kg dose of aspirin was also studied in acetic acid
writhing.
6.2.5. Statistical analysis
The data were expressed as mean± SEM of 6 animals. Results were analysed
statistically by One-way ANOVA followed by Tukey’s multiple comparison using
sigma stat software. The difference was considered significant at p<0.05.
6.3. RESULTS
Acute toxicity
Same as in section 3.3.1 of chapter 3.
Effect of valerian extracts and oils in Acetic acid-induced writhing
Different doses of VVDR03 and VVW02 dose dependently inhibited the
number of writhings. VVDR03 produced significant inhibition of writhing at 80mg/kg

(54.3%) and 40 mg/kg (29.8%) (Fig. 6.1.). While VVW02 produced significant effect
only at 80 mg/kg and the percentage inhibition with it was found to be 29.8% (Fig
6.2.). The standard drug aspirin at 100mg/kg dose significantly inhibited the writhing
movements (64.9%) and the results were found to be highly significant (p<0.05) in
comparison to control group.
Similarly V-MA and V-PA at doses 20, 40 and 80 mg/kg, p.o. produced dose
dependent inhibition. V-PA at doses 80 and 40 mg/kg produced significant inhibition
of writhings i.e. 50% and 31.1% respectively (Fig. 6.3.). V-MA produced significant
inhibition in number of writhings at 80 mg/kg (63.6 %) and 40 mg/kg (27.2 %) as
compared to control group (Fig. 6.4.).
Effect of valerian extracts and oils on tail flick latency
In tail flick model different doses of VVDR03 (Table 6.1.) and V-PA (Table
6.3.) showed no significant increase in tail flick latency at different time interval when
compared to the predrug reaction time. But both VVW02 (Table 6.2.) and V-MA
(Table 6.4.) at 80 mg/kg dose produced significant increase in tail flick latency after
two hours of drug administration in comparison to predrug reaction. Similarly the
standard drug aspirin produced significant increase in tail flick latency after one and
two hours of drug administration.
Effect of valerian extracts in combination with aspirin in acetic acid writhing
Aspirin (5 mg/kg i.p) produced significant inhibition of writhings as compared
to vehicle treated group. But when 20 mg/kg dose of VVDR03 and VVW02 was
given in combination with aspirin (5 mg/kg) no potentiation was seen.
Effect of valerian oils in combination with aspirin in acetic acid writhing
V-PA (20 mg/kg) per se was found to have no effect on acetic acid writhing.
Aspirin per se produced 35% inhibition of writhing as compared to vehicle treated

group while when aspirin was given in combination with V-PA (20 mg/kg)
percentage inhibition was found to be 61.7% (Fig. 6.7.). Similarly V-MA in
combination with aspirin produced 52.4 % decrease in number of writhings (Fig.
6.8.). Thus the results show that aspirin potentiated the action of V-MA and V-PA in
acetic acid induced writhing.
0
5
10
15
2 0
2 5
3 0
3 5
4 0
4 5
V V D R 0 3( 2 0 )
V V D R 0 3( 4 0 )
V V D R 0 3( 8 0 )
V ehicle A sp ir in
Num
ber o
f writ
hing
s
*
*
*
Fig. 6.1. Effect of different doses of VVDR03 in acetic acid induced writhing in mice. Aspirin (100 mg/kg p.o) was used as standard. (*) denotes significance at P<0.05 versus vehicle treated group
0
5
1015
2 0
2 5
3 0
3 54 0
4 5
50
V V W 0 2( 2 0 )
V V W 0 2( 4 0 )
V V W 0 2( 8 0 )
V ehicle A sp ir in
Num
ber o
f writ
hing
s
*
*
Fig. 6.2. Effect of different doses of VVW02 in acetic acid induced writhing in mice. Aspirin (100 mg/kg p.o) was used as standard. (*) denotes significance at P<0.05 versus vehicle treated group

0
5
10
15
2 0
2 5
3 0
3 5
4 0
4 5
V - PA( 2 0 )
V - PA( 4 0 )
V - PA( 8 0 )
V ehicle A sp ir in
Num
ber o
f writ
hing
s
* *
*
Fig. 6.3. Effect of different doses of V-PA in acetic acid induced writhing in mice. Aspirin (100 mg/kg p.o) was used as standard. (*) denotes significance at P<0.05 versus vehicle treated group.
0
5
10
15
2 0
2 5
3 0
3 5
4 0
4 5
V - M A( 2 0 )
V - M A( 4 0 )
V - M A( 8 0 )
V ehicle A sp ir in
Num
ber o
f writ
hing
s
*
*
*
Fig. 6.4. Effect of different doses of V-MA in acetic acid induced writhing in mice. Aspirin (100 mg/kg p.o) was used as standard. (*) denotes significance at P<0.05 versus vehicle treated group. Table 6.1. Effect of different doses of VVDR03 in tail flick latency in mice. (*) denotes significance at P<0.05 versus predrug reaction Tail flick latency in seconds
Group (mg/kg) Pre-drug reaction
0.25 h 0.5 h 1 h 2 h 3 h
VVDR03 (20) 2.04±0.17 1.75±0.6 2.11±0.2 2.4±0.01 2.5±0.4 2.15±0.4
VVDR03 (40) 2.08±0.4 2.44±0.3 2.46±0.27 3.48±0.6 3.88±0.8 3.57±0.7
VVDR03 (80) 2.22±0.3 1.89±0.18 2.18±0.5 3±0.5 2.7±0.3 2.28±0.6
Aspirin (100) 3.79±0.2 3.94±0.8 4.5±0.5 7.17±0.07* 5.77±0.8* 3.88±0.19

Table 6.2. Effect of different doses of VVW02 in tail flick latency in mice. (*) denotes significance at P<0.05 versus predrug reaction
Tail flick latency in seconds
Group (mg/kg)
Pre-drug reaction
0.25 h 0.5 h 1 h 2 h 3 h
VVW02 (20) 2.00±0.17 2.75±0.6 2.13±0.2 2.6±0.01 2.5±0.4 2.15±0.4
VVW02 (40) 2.32±0.3 2.47±0.3 2.6±0.5 2.9±0.3 3.48±0.6 3.57±0.7
VVW02 (80) 1.18±.07 1.37±0.01 1.69±0.2 2±0.4 3.14±0.8* 2.3±0.8
Aspirin (100) 3.79±0.2 3.94±0.8 4.5±0.5 7.17±0.07* 5.77±0.8* 3.88±0.19
Table 6.3. Effect of different doses of V-PA in tail flick latency in mice. (*) denotes significance at P<0.05 versus predrug reaction.
Tail flick latency in seconds
Group (mg/kg)
Pre-drug reaction
0.25 h 0.5 h 1 h 2 h 3 h
V-PA (20) 2.89±0.13 3.31±0.15 2.79±0.41 2.99±0.36 3.32±0.3 3.0±0.31
V-PA (40) 2.23±0.27 2.49±0.18 3.15±0.3 3.29±0.4 3.2±0.4 2.3±0.4
V-PA (80) 2.74±0.19 2.8±0.07 3.24±0.3 2.9±0.29 3.6±0.4 3.3±0.195
Aspirin (100) 3.79±0.2 3.94±0.8 4.5±0.5 7.17±0.07* 5.77±0.8* 3.88±0.19
Table 6.4. Effect of different doses of V-MA in tail flick latency in mice. (*) denotes significance at P<0.05 versus predrug reaction
Tail flick latency in seconds
Group (mg/kg)
Pre-drug reaction
0.25 h 0.5 h 1 h 2 h 3 h
V-MA (20) 3.5±0.17 2.14±0.19 2.88±0.17 2.5±0.15 3.29±0.5 2.97±0.2
V-MA (40) 2.88±0.3 3.3±0.12 6.17±1.9 3.8±0.9 3.97±0.6 3.14±0.7
V-MA (80) 2.57±0.2 2.9±0.3 3.32±0.52 3.4±0.95 4±0.4* 3.4±0.175
Aspirin (100) 3.79±0.2 3.94±0.8 4.5±0.5 7.17±0.07* 5.77±0.8* 3.88±0.19

0
5
10
15
2 02 5
3 0
3 5
4 0
4 5
V V D R 0 3( 2 0 )
V V D R 0 3( 2 0 ) +A sp
( 5)
A sp ( 5) V ehicle
Num
ber o
f writ
hing
s
*, a *
Fig. 6.5. Effect of aspirin (5 mg/kg, i.p) in combination with VVDR03 (20 mg/kg, p.o) in acetic acid induced writhing in mice. (*) denotes significance at P<0.05 as compared to vehicle treated group. (a) denotes significance at P<0.05 as compared to VVDR03 per se.
05
1015
2 02 53 03 54 04 550
V V W 0 2( 2 0 )
V V W 0 2( 2 0 ) +A sp
( 5)
A sp ( 5) V ehicle
Num
ber o
f writ
hing
s
*, a *
Fig. 6.6. Effect of aspirin (5 mg/kg, i.p) in combination with VVW02 (20 mg/kg, p.o) in acetic acid induced writhing in mice. (*) denotes significance at P<0.05 as compared to vehicle treated group. (a) denotes significance at P<0.05 as compared to VVW02 per se.
0
5
10
15
2 0
2 5
3 0
3 5
4 0
4 5
V - PA ( 2 0 ) V - PA( 2 0 ) +A sp
( 5)
A sp ( 5) V ehicle
Num
ber o
f writ
hing
s
*, a, b
*, a
Fig. 6.7. Effect of aspirin (5 mg/kg, i.p) in combination with V-PA (20 mg/kg, p.o) in acetic acid induced writhing in mice. (*) denotes significance at P<0.05 as compared to vehicle treated group. (a) denotes significance at P<0.05 as compared to V-PA per se and (b) denotes significance at P<0.05 as compared to Aspirin per se.

0
5
10
15
2 0
2 5
3 0
3 5
4 0
4 5
V - M A( 2 0 ) )
V - M A( 2 0 ) +A sp
( 5)
A sp ( 5) V ehicle
Num
ber o
f writ
hing
s
*, a, b *, a
.Fig. 6.8. Effect of aspirin (5 mg/kg, i.p) in combination with V-MA (20 mg/kg, p.o) on acetic acid induced writhing in mice. (*) denotes significance at P<0.05 as compared to vehicle treated group. (a) denotes significance at P<0.05 as compared to V-MA per se and (b) denotes significance at P<0.05 as compared to Aspirin per se.
6.4. DISCUSSION
In this study, analgesic activity of valerian extract and oils were investigated
in both acetic acid-induced writhing and tail flick models in mice. The acetic acid-
induced writhing reaction in mice, described as a typical model for inflammatory pain,
has long been used as a screening tool for the assessment of analgesic or anti-
inflammatory properties of new agents (Collier et al., 1968; Khandelwal, 2007). This
method presents a good sensitivity; however, it shows poor specificity, leaving scope
for the misinterpretation of results. This can be avoided by complementing the test
with other models of nociception and by a performance motor test. Acetic acid causes
inflammatory pain by inducing capillary permeability (Amico-Roxas et al, 1984) and
liberating endogenous substances that excite pain nerve endings (Raj, 1996). The
abdominal constriction is related to sensitization of nociceptive receptors to
prostaglandins, bradykinin and substance-P (Huo et al, 2007, Singh and Majumdar,
1995). The nociceptive response caused by acetic acid is also dependent on the release
of TNF-α, interleukin-8 via modulation of macrophages and mast cells localized in
the peritoneal cavity (Ikeda et al., 2001). Because the writhing test usually lasts for
less than an hour, it is too short to cover central prostaglandin-dependent sensitization,

which requires spinal COX-2 induction (Hanns, 2007) however it primarily involves
COX-1 derived prostanoids and thus non-selective COX inhibitor offers a better
antinociceptive effect as compared to selective COX-2 inhibitors (Kulkarni and Jain,
2005).
The results suggest that in acetic acid writhing model VVDR03 produced
significant effect at 40 and 80 mg/kg while VVW02 was found to produce significant
effect at only 80 mg/kg. Both V-MA and V-PA produced significant effect in acetic
acid induced writhing at 40 and 80 mg/kg dose. The highest dose of V-MA tested (80
mg/kg) produced 63.6 % inhibition whereas that of V-PA produced 50 % inhibition.
Statistical comparison shows that both the oils are equally effective in producing
antinociceptive in acetic acid writhing. Aspirin also inhibited the number of writhings
significantly and the effect produced by 80 mg/kg V-MA is comparable to that of 100
mg/kg aspirin (64.9%).
Tail flick is an acute spinally mediated reflex to noxious thermal stimuli. The
fast rising pain in the tail flick gives rise to rapid tail withdrawal at the lowest possible
threshold for pain before it reaches to higher level. In the tail flick model, none of the
doses of V-PA and VVDR03 were found to have effect on tail flick latency when
compared to the predrug reaction time while 80 mg/kg dose of V-MA and VVW02
produced significant increase in tail flick latency after two hours. This indicates that
higher center is also involved in antinociceptive effect of the maaliol chemotype but
at higher doses.
The present study demonstrated that both valerian extracts and oils were found
to be more effective as peripheral analgesics. When studied for mechanism of action
in acetic acid writhing model aspirin potentiated the action of valerian oils while no
potentiation was seen with extracts. NSAIDs can inhibit COX in peripheral tissues
and, therefore, interfere with the mechanism of transduction of primary afferent
nociceptors (Fields, 1987). So the mechanism of analgesic effect of valerian oil could
probably be due to blockade of the effect or the release of endogenous substances that
excite pain nerve endings similar to that of indomethacin and other NSAIDs.

There are few reports on analgesic activity of Valeriana wallichii (Cao and
Hong, 1994; Schultz and Eckstein, 1962; Shrivastava and Sisodia, 1970). There is
another report which demonstrates analgesic activity of the volatile oil of Valeriana
amurensis in acetic acid writhing (Wu et al., 2007). There are many studies reporting
analgesic activity of essential oils and sesquiterpenes (Golshani et al, 2004; Santos et
al, 1997; Sayyah et al, 2002; Sayyah et al, 2003; Koudou et al, 2005; Ahmed et al,
1997). A recent report presents natural terpene 1,8-cineole for its analgesic effect
(Santos and Rao, 2000). In a study β-caryophyllene inhibited in vitro formation of
PGE1 (Burstein et al, 1975) and sesquiterpenoids have been found to have suppressive
effect on iNOS and COX-2 activity (Lee et al., 2002; Yoon et al., 2008). Anti-
inflammatory effect has been reported for limonene and terpineol (Duke and
Beckstrom, 1996). The limonene has been tested in the lipopolysaccharide (LPS)-
induced pleurisy model and when administered orally was able to inhibit the LPS-
induced inflammation including cell migration and inhibiting the production of nitric
oxide, gamma-interferon and IL-4 (Souza, 2003). The inhibitory effect of terpineol, a
volatile terpenoid alcohol on the compound action potential of rat sciatic nerve has
been reported (Moreira, 2001).
Few studies on Indian valerian have demonstrated that it contain flavonoid
compounds (Marder et al., 2003; Tang and Liu, 2003). Flavonoids are known to target
prostaglandins which are involved in the late phase of acute inflammation and pain
perception (Rajnarayana et al., 2001). Iridoids found in many medicinal plants exhibit
a wide range of bioactivities including cardiovascular, antiphepatotoxic, chlorectic,
hypoglycemic, anti-inflammatory, antispasmodic, antitumor, antiviral, immuno-
modulator and purgative activities (Didna et al., 2007). In the present study, GC-MS
screening of both the plant showed that their essential oil is dominated by
sesquiterpenes so we can speculate that the inhibitory effect of extracts and essential
oils in acetic acid writhing and hot plate test is due to the presence of terpenes,
sesquiterpenes (Cabo et al, 1986; Ozbek et al., 2006) and iridoids. Natural terpenoids
are reported to be natural inhibitors of NF-κB signaling (Salminen et al., 2008) just
like aspirin and other NSAIDs (Muller et al., 2001; Tegeder et al., 2001). Valerian

oils inhibited number of writhings by acting in a way similar to aspirin while effect of
extracts in acetic acid writhing may be attributed to some other mechanism possibly
to KATP channel activation which can then led to antispasmodic effect (Gilani et al,
2005). Analgesic effect of DCM extract of maaliol chemotype on tail flick model may
be attributed to mechanism other than that of COX-inhibiton. Hence the presence of
flavonoids, terpenes and iridoids may be contributory to the analgesic activity of
Valeriana in acetic acid induced writhing and tail flick model.

CHAPTER-7
Evaluation of in vitro Antioxidant Profile of Valeriana wallichii chemotypes

7.1. INTRODUCTION
Free radicals have aroused significant interest among scientists in the past
decade. Their broad ranges of effects in the biological systems have drawn on the
attention of many experimental works. Highly reactive free radicals, especially
oxygen-derived radicals which are formed by exogenous chemicals or endogenous
metabolic processes in the human body, are capable of oxidizing biomolecules,
resulting in cell death and tissue damage. Oxidative stress, defined as an imbalance
between oxidants and antioxidant in favour of the former, leads to many biochemical
changes due to overproduction of free radicals. DNA is probably the most
biologically significant target of oxidative attack, and it is widely thought that
continuous oxidative damage to DNA is a significant contributor to the age-related
development of the major cancers (Ames et al., 1993). Not only this, oxidative
damage has been implicated in the pathology of many other diseases and conditions
including diabetes, cardiovascular disease, inflammatory conditions and aging.
Almost all organisms possess antioxidant defenses and numerous damage removal
and repair enzymes to remove or repair damaged molecules. However, the natural
antioxidant mechanisms can be inefficient, hence dietary intake of antioxidant
compounds become important. Several authors demonstrated that antioxidant intake is
inversely related to mortality from coronary heart disease and to the incidence of heart
attacks (Hertog et al., 1993; Anderson et al., 1996). Antioxidants like vitamin C,
vitamin E (tocopherol), carotenoids and several polyphenolic compounds including
flavonoids that are mainly supplied as dietary consumptions can also impede
carcinogenesis by scavenging oxygen radical or interfering with the binding of
carcinogens to DNA (Stoner and Mukhtar, 1995). Synthetic antioxidants are widely
used because they are effective and cheaper than natural types. However, the safety
and toxicity of synthetic antioxidants have been important concerns (Imaida et al.,
1983).
Many herbal plants contain antioxidant compounds and these compounds
protect cells against the damaging effects of reactive oxygen species, such as singlet
oxygen, superoxide, peroxyl radicals, hydroxyl radicals and peroxynitrite. The food
industries also use natural antioxidants as a replacement of conventional synthetic

antioxidants in food by natural products that are considered to be promising and a safe
source. As a result of which, much attention has been directed towards the
characterization of antioxidant properties of plant extracts/their fractions and
identification of the constituents responsible for those activities. Considering the
pathological implications of reactive oxygen species (ROS), it is important to identify
antioxidants, which can scavenge several free radicals and prevent multiple diseases.
This can be achieved by simple in vitro test systems. Subsequent to this preliminary
screening, the results can be confirmed with ex vivo and in vivo systems.
V. wallichii is used as carminative and forms an ingredient of a number of
ayurvedic recepies. The drug contains a group of iridoid or monoterpene derivatives
known as valepotriates, which are useful as tranquilizers. It is also used for its
antispasmodic properties. V. officinalis a related species is reported to posses
neuroprotective activity and in another study its efficiency as antioxidant was tested
by the influence of its extract on the yield of chemiluminesence of Gly-Trp solution
(Bol’shakova et al., 1997). V. wallichii and V. officinalis share a group of active
constituents and both of them are characterized by an unusual fragnance, derived from
the essential oil component. There are reports depicting the presence of lignans,
terepenes, iridoids and flavonoids in Indian valerian. Besides this V. wallichii is being
used as a substitute for V. officinalis. We thus evaluated the antioxidant properties of
V. wallichii in different in vitro models. The antioxidant activity of the tested
compounds was compared with the standard antioxidant vitamin C.
7.2. MATERIAL AND METHODS
7.2.1. Chemicals
Potassium ferricyanide, sodium nitroprusside, ferric chloride and
trichloroacetic acid. Sulfanilamide, napthylethylene diamine hydrochloride,
orthophosphoric acid and 1, 1,-diphenyl 2-picryl hydrazyl (DPPH) were obtained
from Sigma Chemicals, USA. Ascorbic acid was procured from Ranbaxy, Gurgaon.
All other unlabeled chemicals and reagents were of analytical grade and were used
without further purification.

7.2.2. Plant material and its extraction
Same as in section 3.2.1. of chapter 3.
7.3. IN VITRO ANTIOXIDANT ASSAYS
In vitro antioxidant activity was determined by using DPPH radical
scavenging, nitric oxide scavenging and reductive ability. Different concentrations of
both the extracts (VVDR03 and VVW02) and oils (V-PA and V-MA) were prepared
in dimethyl sulfoxide (DMSO) ranging from 1-10mg/ml.
7.3.1. DPPH Radical Scavenging assay
DPPH (1,1,-diphenyl 2-picryl hydrazyl) scavenging activity was measured by
the spectrophotometric method (Sreejayan and Rao, 1996). DPPH, is a stable free
radical having a purple colour. When free radical scavengers are added, DPPH is
reduced and its colour is changed to yellow, based on the efficacy of antioxidants
(Fig. 1.). To a methanolic solution of DPPH (100 µM) 2.95 ml, 0.05 ml of test
compounds (extracts and oils of both the chemotypes) dissolved in DMSO were
added at different concentrations (1-10 mg/ml). Equal amount of DMSO was added to
the control. Absorbance was recorded at 517 nm after 20 min. Ascorbic acid was used
as a standard. Radical scavenging activity was expressed as percent inhibition and
was calculated using the following formula.
% DPPH radical scavenging activity= (1- Sample OD) x100 Control OD
NO2
NO2
NN NO2[ H ]
NO2N N
NO2
NO2
H
DPPH Reduced form of DPPH
Fig. 1. Reduction of 1,1, Diphenyl -2-Picryl hydrazyl (DPPH) free radical.

7.3.2. Nitric Oxide Scavenging Effect
Nitric oxide is generated from sodium nitroprusside, which at physiological
pH (7.4) liberates nitric acid. This nitric acid gets converted to nitrous acid and further
forms nitrite ions on contact with air. The nitrite ions diazotise with sulphanilic acid
and couple with naphthylethylenediamine (Griess reagent), producing pink colour
which can be measured spectrophotometrically (Sreejayan and Rao, 1997) (Fig. 2.).
Sodium nitroprusside (5 mM) in phosphate buffer saline was mixed with different
concentrations of extracts and oils (1-10mg/ml) dissolved in DMSO and incubated at
25º C for 30 min. After 30 min 1.5 ml of incubated solution were removed and diluted
with 1.5 ml of Griess reagent (1% sulphanilamide, 2% phosphoric acid and 0.1%
napthylethylene diamine dihydrochloride). The absorbance of the chromophore
formed during diazotisation of the nitrite with sulphanilamide and subsequent
coupling with napthylethylene diamine was measured at 546 nm along with a control.
Ascorbic acid was used as a standard. The percentage inhibition of nitric oxide
generated was measured by comparing the absorbance values of control and test.
% nitric oxide scavenging activity= (1- Sample OD) X 100 Control OD
H 2 NO 2 S N = N NH NH 2
+
NH NH 2
+N
N
SO 2 NH 2
H 2+
SO 2 NH 2
N
NO
O 2
+NO
NH 2
SO 2 NH 2
NO
NO + O 2
SO 2 NH 2
NH 2
Fig. 2. Generation of nitric oxide and its reaction with Griess reagent.

7.3.3. Reducing Power
Reductive ability of the extracts and oils was measured according to the
method of Oyaizu (Oyaizu, 1986). Different concentrations (1-10mg/ml) of extract
were mixed with 2.5 ml of sodium phosphate buffer (pH 6.6) and 2.5 ml of potassium
ferricyanide (1%). The mixture was incubated at 50ºC for 20 min. Trichloroacetic
acid (2.5 ml of 10%) was added to it, the mixture was mixed and centrifuged at 650
rpm for 10 min. The upper layer (5 ml) was mixed with 5 ml of deionised water and 1
ml of ferric chloride (1%) and absorbance was measured at 700 nm. Higher
absorbance indicated higher reducing power.
7.3.4. Statistical Analysis
For all the drugs and in all the test systems, percentage inhibition curve was
plotted by taking concentration on x-axis and percentage scavenging on the y-axis and
IC50 values (amount of drug required to scavenge 50% of free radicals) were obtained
from the linear plots (r value ranging from 0.92-0.98 for all the test systems). The data
was statistically analyzed by one-way ANOVA followed by Tukey’s test for multiple
pair wise comparison in all the test systems.
7.4. RESULTS
7.4.1. DPPH Radical Scavenging Activity
This activity has been used by various researchers as quick and reliable
parameters to assess the in vitro antioxidant activity of crude plant extracts (Navarro
et al., 1992; Thabrew et al., 1998). In this test DPPH radicals react with suitable
reducing agents and then electrons become paired off and the solution loses its colour
and becomes yellow. The ability of a compound to act as donor for hydrogen atoms or
electrons is measured spectrophotometrically. The DPPH scavenging activity exerted
by VVDR03 and VVW02 as well as ascorbic acid is summarized in Fig. 7.1. and
exhibited a concentration dependent inhibition. VVDR03 at 10 mg/kg exhibited
63.8% inhibition, VVW02 and ascorbic acid at the same concentration exhibited 34.5
% and 96% inhibition. In this study VVDR03 at different concentration’s

demonstrated significant DPPH scavenging activity as compared to VVW02. Free
radical scavenging activity of both the extracts is moderate as compared to ascorbic
acid. IC50 values were found to be 7.028 mg/ml for VVDR03 and 13.653 mg/ml for
VVW02 (Fig. 7.7.). While the IC50 of standard drug Ascorbic acid is 0.022 mg/ml. So
the study demonstrated that both have abilities to act as radical scavengers. Valerian
oils also produced concentration dependent inhibition but were found to have weak
scavenging action on DPPH as compared to extracts. Both V-PA and V-MA exhibited
almost similar scavenging effect in DPPH assay i.e 23.2 and 26.2 % at 10mg/ml (Fig.
7.4.).The IC50 was found to be 26.707 mg/ml for V-PA and 21.06 for V-MA which is
much greater than that of ascorbic acid (Fig. 7.7.).
7.4.2. Nitric Oxide Scavenging Effect
Nitric oxide (NO) is an important chemical mediator generated by endothelial
cells, macrophages, neurons etc. and is involved in the regulation of various
physiological processes. Excessive nitric oxide is associated with several diseases.
Nitric oxide reacts with superoxide and forms peroxynitrite radicals and is responsible
for the inflammatory response by the release of prostaglandin. Some scientists believe
that repeated infections throughout life, cause an excessive production of NO, which,
over time, causes toxic damage to the body resulting in age-related diseases such as
heart disease, Alzheimer’s disease and Diabetes. Peroxynitrite, produced in vivo from
nitric oxide and superoxide, leads to oxidation of low-density lipoprotein.
Peroxynitrite is a strong nitrating species with oxidizing effect. It may oxidize all
major types of biomolecules like DNA, proteins and lipids. Peroxynitrite is thought to
be involved in neurodegenerative diseases like Alzheimer’s disease, Parkinson’s
disease etc. Thus nitric oxide scavenging may prevent the diseases caused by it.
Fig. 7.2. illustrates the percentage inhibition of nitric oxide generation by
VVDR03 and VVW02. Ascorbic acid was used as a standard compound and its IC50
is 0.02243 mg/ml. IC50 of VVW02 was found to be 8.3 mg/ml whereas that of
VVDR03 is 0.6 mg/ml (Fig. 7.8.). This shows that nitric oxide scavenging activity of
VVDR03 is more than VVW02.

Similarly valerian oils were tested for nitric oxide scavenging effect and
showed a concentration dependent scavenging effect. The IC50 of V-MA was found to
be 12.7 mg/ml while that of V-PA is 7.09 mg/ml (Fig. 7.8.).
7.4.3. Reductive Ability
The antioxidant activity has been reported to be concomitant with the
development of reducing power. The reductive ability of VVDR03 and VVW02 has
been shown in Fig. 7.3. The reducing power increased as the extract concentration
increased, indicating that some compound in extract is both electron donor and could
react with free radicals to convert them in to more stable products and to terminate
radical chain reaction. All the concentrations of both the extracts showed higher
activity than the control and out of them VVDR03 was found to be having more
reducing power than VVW02. Both the oils when tested for reducing power were
found to have almost similar reducing power at tested concentrations (Fig. 7.6.). The
reducing power was found to be low for oils than the extracts. Ascorbic acid at all the
concentrations gave absorbance as 3.9 nm while control gave an absorbance of 0.054
nm. This shows that ascorbic acid has very high reducing property.
0
20
40
60
80
100
120
0 1 2 4 6 8 10
Conc (mg/ml)
% in
hibi
tion
V V D R 0 3 V V W 0 2 A sco rb ic acid
Fig.7.1. Percentage inhibition of DPPH radical by different concentrations of VVDR03, VVW02 and ascorbic acid

0
20
40
60
80
100
120
0 1 2 4 6 8 10
Conc (mg/ml)
% in
hibi
tion
V V D R 0 3 V V W 0 2 A sco rb ic acid
Fig. 7.2. Percentage inhibition of nitric oxide by different concentrations of VVDR03, VVW02 and Ascorbic acid
0
1
2
3
4
5
0 1 2 4 6 8 10
Conc (mg/ml)
Abs
orba
nce
(nm
)
V V D R 0 3 V V W 0 2 A sco rb ic acid
Fig. 7.3. Reductive ability of different concentrations of VVDR03, VVW02 and Ascorbic acid

0
20
40
60
80
100
120
0 1 2 4 6 8 10
Conc (mg/ml)
% in
hibi
tion
V - PA V =M A A sco rb ic acid
Fig. 7.4. Percentage inhibition of DPPH radical by different concentrations of V-PA, V-MA and ascorbic acid
0
20
40
60
80
100
120
0 1 2 4 6 8 10
Conc (mg/ml)
% in
hibi
tion
V - PA V =M A A sco rb ic acid
Fig. 7.5. Percentage inhibition of nitric oxide by different concentrations of V-PA, V-MA and Ascorbic acid

0
1
2
3
4
5
0 1 2 4 6 8 10
Conc (mg/ml)
Abs
orba
nce
(nm
)
V - PA V =M A A scorb ic acid
Fig. 7.6. Reductive ability of different concentrations of V-PA, V-MA and Ascorbic acid
7.0 2 8
13 .6 53
2 6 .70 7
2 1.0 6
0 .0 2 2 20
5
10
15
2 0
2 5
3 0
Conc
entra
tion
(mg/
ml)
V V D R 0 3 V V W 0 2 V - PA V - M A A scorb icacid
Fig. 7.7. Histogram showing IC50 values of V. wallichii extracts and oils in DPPH
assay

0 .6
8 .3
7.0 9
12 .7
0 .0 2 2 4 30
2
4
6
8
10
12
14
Conc
entra
tion
(mg/
ml)
V V D R 0 3 V V W 0 2 V - PA V - M A A scorb icacid
Fig. 7.8. Histogram showing IC50 values of V. wallichii extracts and oils in nitric oxide scavenging assay 7.4. DISCUSSION
Free radical oxidative stress has been implicated in the pathogenesis of a
variety of clinical disorders, resulting usually from deficient natural antioxidant
defenses. Potential antioxidant therapy therefore should include either natural free
radical scavenging enzymes or agents which are capable of augmenting the activity of
these enzymes. If human disease is believed to be due to the imbalance between
oxidative stress and antioxidative defense, it is possible to limit oxidative tissue
damage and hence prevent disease progression by antioxidant defense supplements.
Medicinal herbs are known to contain a variety of antioxidants. It has been revealed
that various phenolic antioxidants such as flavonoids, tannins, coumarins, xanthones
and more recently procyanidins present in plants scavenge radicals dose dependently,
thus they are viewed as promising therapeutic drugs for free radical pathologies. The
antioxidant activity of putative antioxidants have been attributed to various
mechanisms, among which are prevention of chain initiation, binding of transition
metal ion catalysts, decomposition of peroxides, prevention of continued hydrogen
abstraction, reductive capacity and radical scavenging (Diplock, 1997; Oktay et al.,
2003). Numerous antioxidant methods and modifications have been proposed to

evaluate antioxidant activity and to explain how antioxidants function. Of these, total
antioxidant activity, reducing power, DPPH assay, metal chelating, active oxygen
species such as H2O2, O2•− and OH• quenching assays are most commonly used for the
evaluation of antioxidant activities of extracts (Duh et al., 1999; Amarowicz et al.,
2000; Chang et al., 2002).
DPPH radicals react with suitable reducing agents losing colour
stoichometrically with the number of electrons consumed which is measured
spectrophotometrically at 517 nm. The DPPH method is widely used to determine
antiradical/antioxidant activity of purified phenolic compounds as well as natural
plant extracts (Brand-Williams et al., 1995; Sripriya et al., 1996; Bondet et al., 1997;
Mahinda and Shahidi, 2000; Peyrat-Mallard et al., 2000). The method also has good
repeatability and is used frequently however color interference of DPPH with samples
that contain anthocyanins leads to underestimation of antioxidant activity (Arnao,
2000). Used as a reagent, DPPH evidently offers a convenient and accurate method
for titrating the oxidizable groups of natural or synthetic antioxidants. Lower
absorbance of the reaction mixture indicated higher free radical scavenging activity
(Cao et al., 1997). All the tests were performed in triplicate and the results averaged.
From the present results it may be postulated that DCM extracts, VVDR03 and
VWR02 reduces the radical to the corresponding hydrazine when it reacts with the
hydrogen donors in the antioxidant principles. The free radical scavenging effect was
found to be dose dependent. Standard ascorbic produced 96% scavenging of DPPH at
10mg/ml and 72.3% at 1 mg/ml. The IC50 of VVDR03 (7.028 mg/ml) and VVW02
(13.653 mg/ml) demonstrated that VVDR03 is more potent than VVW02 in
scavenging free radicals, however the scavenging potential is moderate in comparison
to ascorbic acid.
Nitric oxide is generated from the amino acid L-arginine by vascular
endothelial cells, phagocytes and certain cells in the brain. Some scientists believe
that repeated infections throughout life cause an excessive production of NO, which
over time, causes toxic damage to the body resulting in age related diseases such as

heart disease, Alzheimer’s disease and diabetes. NO reacts with superoxide and forms
peroxynitrite radicals and is responsible for the inflammatory response by the release
of prostaglandin. The extracts moderately inhibited nitric oxide in dose dependent
manner, the effect being more prominent in case of VVDR03 than VVW02. Whereas
the oils V-PA and V-MA showed a week nitric oxide scavenging activity. Thus the
present study offers scientific evidence for the use of the plant in the indigenous
system in inflammatory conditions.
Reducing power of extracts and oils was good and increased with increasing
concentration. On a comparative basis, the extracts were found to have more
antioxidant property than the oils, which posses only weak antioxidant activity. Out of
the two extracts VVDR03 was found to have more antioxidant potential than the
VVW02. The antioxidant property of extracts can be attributed to the presence of
iridoids (Gamboa and Castro, 2004; Raju et al., 2004), lignans (Yamauchi et al., 2007;
Piccinelli et al., 2004) and flavonoids. There are studies showing that extract of
valerian root are endowed with flavonoids (Wasowski et al., 2002; Marder et al.,
2003) making this popular and very well tolerated plant attractive for the search of a
daily-used neuroprotective product for human consumption (Tang et al., 2003). The
weak antioxidant effect of valerian oils could be due to their chemical composition, as
the essential oil contains mainly monoterpene hydrocarbons such as sabinene,
terpinene-4-ol, α-pinene, p-cymene, terpinene, limonene, β-pinene and α-terpineol.
These compounds are known to possess a weak antioxidant activity (Tepe et al.,
2005).
In vitro scavenging is only a preliminary test for selecting potent antioxidants.
These compounds should be tested in vivo for their activity in different disease states
so that their potential uses will be explored. Their bioavailability and interaction with
biomembranes can also be determined. If these parameters are established properly,
the scope for developing phytochemicals into pharmaceuticals will be practical.

CHAPTER-8
Modulation of Antioxidant Defense System in Mice Brain by Valeriana wallichii
chemotypes

8.1. INTRODUCTION
Oxygen is essential for the survival of all on this earth. During the process of
oxygen utilization in normal physiological and metabolic processes approximately 5%
of oxygen gets univalently reduced to oxygen-derived free radicals. Free radicals are
partially reduced metabolites of oxygen and are highly toxic, mutagenic and reactive.
A free radical has one or more unpaired electrons in its outer orbital and they react
rapidly with adjacent molecules, donating, abstracting and even sharing its outer
orbital electron(s). Sometimes free radicals generate a second free radical or other
reactive oxygen species (ROS). Oxidative damage by oxygen free radicals are known
to be one of the mechanisms of chronic disorders such as atherosclerosis or cancer
(Halliwell and Gutteridge, 1989). Oxygen free radicals are created through aerobic
metabolism and are mostly removed by antioxidants in vivo (Diplock, 1994). An
antioxidant is a substance, which at low concentration relative to oxidisable substrate
significantly delays or reduces oxidation of the substrate by itself becoming oxidized.
The antioxidant defense systems in the body include superoxide dismutase (in
mitochondria and cytosol), catalase (in peroxisomes), glutathione peroxidase, α-
tocopherol (in membranes and lipoproteins). As with the chemical antioxidants, cells
are protected against oxidative stress by an interacting network of antioxidant
enzymes (Davies, 1995; Sies, 1997). Here, the superoxide released by processes such
as oxidative phosphorylation is first converted to hydrogen peroxide and then further
reduced to give water. This detoxification pathway is the result of multiple enzymes,
with superoxide dismutases catalysing the first step and then catalases and various
peroxidases removing hydrogen peroxide. The glutathione system includes
glutathione, glutathione reductase, glutathione peroxidases and glutathione S-
transferases (Meister and Anderson,1983). This system is found in animals, plants and
microorganisms (Meister and Anderson, 1983; Creissen et al., 1996). Glutathione
peroxidase is an enzyme containing four selenium-cofactors that catalyzes the
breakdown of hydrogen peroxide and organic hydroperoxides. But when generation
of ROS overtakes the antioxidant defense of the cells, the free radicals start attacking
the cell proteins, lipids and carbohydrates leading to a number of physiological
disorders. They have also been implicated in pathogenesis of a number of disorders

like diabetes, liver damage, nephrotoxicity, inflammation, cancer, cardiovascular
disorders, and neurological disorders and in the process of aging. Antioxidants can
cancel out the cell-damaging effects of free radicals (Sies, 1997). Furthermore, people
who eat fruits and vegetables, which are good sources of antioxidants, have a lower
risk of heart disease and some neurological diseases (Stanner, 2004) and there is
evidence that some types of vegetables, and fruits in general, probably protect against
a number of cancers. People taking vitamin E supplements had a lower risk of
developing heart disease (Rimm et al., 1993). There is also some evidence that
antioxidants might help prevent diseases such as macular degeneration (Bartlett and
Eperjesi, 2003), suppressed immunity due to poor nutrition (Wintergerst et al., 2006),
and neurodegeneration (Wang, 2006). These observations suggested that antioxidants
might help prevent these conditions.
Therefore there is a constant need to replenish antioxidant resources, either
endogenously or through supplementation. For this, there is a need to identify
antioxidants, which can scavenge several free radicals and prevent multiple diseases.
Although synthetic antioxidants such as butylated hydroxytoluene (BHT), butylated
hydroxyanisole (BHA) and tertiary butylhydroquinone are widely used as
antioxidants in various industries, they are suspected of being responsible for liver
damage and carcinogenesis in laboratory animals (Grice, 1986). Therefore, the
development and utilization of more effective antioxidants of natural origin is desired
(Gulcin et al., 2002). Therefore search into the determination of natural antioxidant
sources is much important. The present study has been directed to investigate the
modulatory effect of Valeriana wallichii on antioxidant defense system in mice.
8.2. MATERIAL AND METHODS
8.2.1. Plant material and its extraction
Same as in section 3.2.1. of chapter 3.
8.2.2. Animals
Same as in section 3.2.2. of chapter 3.

8.2.3. Experimental protocol and procedure
Both the extracts VVW02 and VVDR03 and oils V-PA and V-MA were
studied for modulatory effect on antioxidant defense system of mice brain. Mice were
divided into thirteen groups, each group consisting of six animals. Group I, II and III
received 10, 20 and 40 mg/kg of VVDR03. Group IV, V and VI received 10, 20 and
40 mg/kg dose of VVW02. Group VII, VIII and IX received 10, 20 and 40 mg/kg
dose of V-PA. Group X, XI and XII received 10, 20 and 40 mg/kg dose of V-MA.
Group XIII received vehicle only i.e a drop of Tween-80 dissolved in distilled water
and served as control. The drugs were dissolved in one drop of Tween 80 and solution
was made up with sterile water. All the drugs were given orally to mice continuously
for 14 days. On 14th day all the animals were sacrificed, blood was collected and
whole brains were removed and weighed. Brains were processed to get 10%
homogenate in cold buffer using glass Teflon homogenizer. The homogenates were
used to estimate brain malondialdehyde (MDA), catalase and reduced glutathione
(GSH). Brain homogenates were centrifuged at 800g for 5 min at 4ºC to get the post
mitochondrial supernatant (PMS) which was used to assay superoxide dismutase
(SOD) activity.
Estimation of lipid peroxidation
The quantitative measurement of lipid peroxidation in terms of
malondialdehyde levels in the whole brain was performed according to the method of
Wills (Wills, 1966). The malondialdehyde (MDA) content, a measure of lipid
peroxiadtion, was assayed in the form of thiobarbituric acid reacting substances
(TBARS). Briefly, the reaction mixture consisted of 0.5 ml homogenate and 0.5 ml
Tris HCl pH 7.4 incubated at 37ºC for 2 hrs. Then 1ml of 10% Trichloroacetic acid
was added to the above mixture and centrifuged at 1000 rpm for 10 min. 1ml of
supernatant was removed, to which is added 1 ml of 0.067% thiobarbitric acid. The
tubes containing reaction mixture were then kept in boiling water for 10 min. They
were then cooled 1ml of distilled water was added. The absorbance of the colour
developed in the tubes was measured at 532 nm.

Estimation of reduced glutathione
Content of reduced glutathione in the brain was assayed by the method of
Jollow (Jollow et al., 1974). 1ml of tissue homogenate (10%) was precipitated with
1ml of sulfosalicylic acid (4%). The samples were kept at 4ºC for at least 1 hr and
then subjected to centrifugation at 1200g for 15 min at 4ºC. The assay mixture
contained 0.1 ml aliquot and 2.7 ml phosphate buffer (0.1 M pH 7.4) in a total volume
of 3 ml. The yellow colour developed was read immediately at 412 nm on a
spectrophotometer.
Estimation of superoxide dismutase (SOD)
Assay of SOD was done in post mitochondrial supernatant (Kono, 1978). The
assay system consisted of EDTA 0.1mM, sodium carbonate 50mM and 96mM of
nitroblue tetrazolium (NBT). In the cuvette, 2 ml of above mixture, 0.05 ml
hydroxylamine and 0.05 ml of PMS were taken and the auto-oxidation of
hydroxylamine was observed by measuring the absorbance at 560 nm.
Estimation of catalase
Catalase activity was assayed by the method of Luck (Luck, 1963). Briefly the
assay mixture consisted of 3 ml of H2O2 phosphate buffer (1.25x10–2 M H2O2) and
0.05 ml of supernatant of the brain homogenate (10%) and the change in absorbance
was read at 240 nm using the spectrophotometer. Enzyme activity was calculated
using the millimolar extinction coefficient of H2O2 (0.07). The results were expressed
as micromoles of H2O2 decomposed per min, per mg of protein.
8.2.4 Statistical analysis
All the values are expressed as mean±SEM. The data were analysed by
analysis of variance (ANOVA) followed by Tukey test. The criterion for statistical
significance was at p<0.05.

8.3. RESULTS
Effect on TBARS levels
Peroxidation of lipids causes severe damage to cell membrane because lipids
are the main component of the cell membrane. Its initiation occurs when a radical
species attacks and removes allylic hydrogen from an unsaturated fatty acid, resulting
in a radical chain reaction. Once lipid peroxidation begins, iron may participate in
driving the process. The possible sources of iron in body are iron-containing proteins.
There are a number of methods available to measure the markers of lipid
peroxidation, either by measuring the formation of products, or the degradation of
reactants. The most commonly used method is the thiobarbituric acid reactive
substances (TBARS) test. The results of the effect of VVW02 and VVDR03 on the
lipid peroxidation of mice brain homogenates are shown in Fig. 8.1 & 8.3. VVW02
did not alter the lipid peroxiadtion in brain as compared to control group. Similarly
none of the dose of VVDR03 except 40 mg/kg attenuated the level of lipid
peroxidation significantly. In case of oils none of the dose of V-PA and V-MA altered
the TBARS levels in mice brain as compared to control group (Fig. 8.5 & 8.7).
Effect on glutathione levels
Glutathione is a cysteine-containing peptide found in most forms of aerobic
life (Meister and Anderson, 1983). It is not required in the diet and is instead
synthesized in cells from its constituent amino acids (Meister, 1988). Glutathione has
antioxidant properties since the thiol group in its cysteine moiety is a reducing agent
and can be reversibly oxidized and reduced. In cells, glutathione is maintained in the
reduced form by the enzyme glutathione reductase and in turn reduces other
metabolites and enzyme systems as well as react directly with oxidants (Linster and
Van, 2007). Due to its high concentration and its central role in maintaining the cell's
redox state, glutathione is one of the most important cellular antioxidants.
In the present study control group was found to have 0.0121micromoles/mg pr
as level of reduced glutathione. VVW02 (10, 20 & 40 mg/kg) did not result in a
significant alteration in reduced glutathione levels (Fig. 8.1.) While 40 mg/kg of
VVDR03 significantly increased the GSH levels (Fig. 8.3.) as compared to control

group. No effect was found on brain glutathione levels with V-PA (Fig. 8.5.) and V-
MA (Fig. 8.7.) (10, 20 and 40 mg/kg).
Effect on catalase
Catalases are enzymes that catalyse the conversion of hydrogen peroxide to
water and oxygen, using either an iron or manganese cofactor (Chelikani, 2004;
Zámocký and Koller, 1999). This protein is localized to peroxisomes in most
eukaryotic cells (Del Río, 1992). Hydrogen peroxide is generated by a number of
intracellular flavin oxidases like xanthine oxidase and also from the autooxidation of
numerous xenobiotics (Hochstein and Atallah, 1988). Hydrogen peroxide can directly
affect the DNA and sulfhydryl groups apart from its participation in hydroxyl radical
generation. H2O2 is reduced by three general mechanisms. 1) It is the substrate for two
enzymes, catalase and glutathione (GSH) peroxidase that catalyze the conversion of
H2O2 to H2O + O2; this presumably is a detoxification mechanism (Maddipati and
Marnett, 1987). 2) H2O2 is converted by myeloperoxidase (MPO) in neutrophils to
hypochlorous acid (HOCl). This appears to be a mechanism for a physiological toxic
agent, since HOCl is a strong oxidant that acts as a bactericidal agent in phagocytic
cells. Reaction of HOCl with H2O2 yields singlet oxygen (1O2) and water. 3) H2O2 is
converted in a spontaneous reaction catalyzed by Fe2+ (Fenton reaction) to the highly
reactive hydroxyl radical (OH.). The hydroxyl radical reacts instantaneously with any
biological molecule (RH) from which it can abstract a hydrogen atom. The resulting
free radical is more stable and hence longer-lived than the hydroxyl radical Thus
removal of hydrogen peroxide significantly lowers the toxic effects of hydroxyl
radicals. VVW02 at 10, 20 and 40 mg/kg failed to improve the levels of catalase (Fig.
8.2.) in mice brain but VVDR03 significantly increased the level of catalase at all the
doses (Fig. 8.4.). No alteration was seen on catalase level with different doses of V-
PA and V-MA (Fig. 8.6. & 8.8.).
Effect on SOD
Superoxide anion is generated from mitochondria during respiratory chain,
enzymatic glycosylation of proteins and also from the activation of phagocytes (Beyer

et al., 1991). Superoxide anions undergo dismutation which causes the production of
hydrogen peroxide (Hochstein and Atallah, 1988; Riley and Behrman, 1991).
Superoxide anion is normally involved in inflammatory conditions. Formation of
superoxide anion radical leads to a cascade of other ROS (Grisham, 1992).
Superoxide dismutates to hydrogen peroxide (H2O2) and oxygen. It is less reactive
when compared to the other free radicals. But it leads to the production of hydroxyl
and peroxynitrite radicals by combination with hydrogen peroxide and nitric oxide
radical respectively. The pathological effects of superoxide anion are indirect in the
sense that it is the subsequently formed hydroxyl and peroxynitrite radicals which are
involved in different pathological conditions like cancer, cardiac, renal ischemia,
atherosclerosis, diabetes etc. Thus if, superoxide anion production is controlled or
scavenged, the formation of hydroxyl and peroxynitrite radicals can be limited, such
that, the diseases caused by them can sufficiently be reduced.
In our study none of the dose of VVW02 (Fig. 8.2.) altered brain SOD levels.
While VVDR03 at 40 mg/kg produced significant increase in SOD levels as
compared to control group (Fig. 8.4.). Similarly when valerian oils V-PA and V-MA
were studied for modulation of SOD levels in brain no effect was found when
compared with control group (Fig. 8.6. & 8.8.).
00 .1
0 .20 .3
0 .40 .50 .6
0 .70 .8
0 .91
V V W 0 2( 10 )
V V W 0 2( 2 0 )
V V W 0 2( 4 0 )
V ehicle
nmol
es/m
g pr
0
0 .0 0 2
0 .0 0 4
0 .0 0 6
0 .0 0 8
0 .0 1
0 .0 12
0 .0 14
0 .0 16
0 .0 18
V V W 0 2( 10 )
V V W 0 2( 2 0 )
V V W 0 2( 4 0 )
V ehicle
mic
rom
oles
/mg
pr
(a) (b)
Fig 8.1. Effect of different doses of VVW02 on a) Lipid peroxidation in mice brain b) Glutathione content in mice brain

0
2
4
6
8
10
12
14
16
18
V V W 0 2( 10 )
V V W 0 2( 2 0 )
V V W 0 2( 4 0 )
V ehicle
mic
rom
oles
of
H2O
2 de
com
pose
d/m
in/m
g p
r
0
0 .5
1
1.5
2
2 .5
3
V V W 0 2( 10 )
V V W 0 2( 2 0 )
V V W 0 2( 4 0 )
V ehicle
un
its/
mg
pr
(a) (b)
Fig. 8.2. Effect of different doses of VVW02 on a) Catalase level in mice brain b) Superoxide dismutase level in mice brain
0
0 .1
0 .2
0 .3
0 .4
0 .5
0 .6
0 .7
0 .8
0 .9
V V D R 0 3( 10 )
V V D R 0 3( 2 0 )
V V D R 0 3( 4 0 )
V ehicle
nmol
es/m
g pr * a
0
0 .0 0 5
0 .0 1
0 .0 15
0 .0 2
0 .0 2 5
0 .0 3
V V D R 0 3( 10 )
V V D R 0 3( 2 0 )
V V D R 0 3( 4 0 )
V ehicle
mic
rom
oles
/mg
pr * a b
(a) (b)
Fig. 8.3. Effect of different doses of VVDR03 on a) Lipid peroxidation in mice brain b) Glutathione content in mice brain. Results are significant at P<0.05. (*) versus vehicle group, a versus VVDR03 (20 mg/kg), b versus VVDR03 (10 mg/kg)
02
46
810
1214
1618
2 0
V V D R 0 3( 10 )
V V D R 0 3( 2 0 )
V V D R 0 3( 4 0 )
V ehicle
mic
rom
oles
of H
2O2
deco
mpo
sed/
mim
/mg
pr
**
*
00 .5
11.5
22 .5
3
3 .54
4 .55
V V D R 0 3( 10 )
V V D R 0 3( 2 0 )
V V D R 0 3( 4 0 )
V ehicle
units
/mg
pr
* a b
(a) (b)
Fig. 8.4. Effect of different doses of VVDR03 on a) Catalase level in mice brain b) Superoxide dismutase level in mice brain. Results are significant at P<0.05. * versus vehicle group, a versus VVDR03 (20 mg/kg) b versus VVDR03 (10 mg/kg)

2 . 8
2 . 9
3
3 . 1
3 . 2
3 . 3
3 . 4
3 . 5
3 . 6
V- P A ( 10 ) V- P A ( 2 0 ) V- P A ( 4 0 ) Ve hi c l e
nmol
es/m
g pr
0 .0 12
0 .0 12 5
0 .0 13
0 .0 13 5
0 .0 14
0 .0 14 5
0 .0 15
V- P A ( 10 ) V- P A ( 2 0 ) V- P A ( 4 0 ) Ve hi c l e
mic
rom
oles
/mg
pr
(a) (b)
Fig. 8.5. Effect of different doses of V-PA on a) Lipid peroxidation in mice brain b) Glutathione content in mice brain
2 .6
2 .7
2 .8
2 .9
3
3 .1
3 .2
3 .3
3 .4
3 .5
V- P A ( 10 ) V- P A ( 2 0 ) V- P A ( 4 0 ) Ve hi c l e
mic
rom
oles
of H
2O2
deco
mpo
sed/
min
/mg
pr
0
0 .5
1
1.5
2
2 .5
3
3 .5
4
4 .5
5
V- P A ( 10 ) V- P A ( 2 0 ) V- P A ( 4 0 ) Ve hi c l e
units
/mg
pr
(a) (b)
Fig. 8.6. Effect of different doses of V-PA on a) Catalase level in mice brain b) Superoxide dismutase level in mice
0
0 .5
1
1.5
2
2 .5
V- M A ( 10 ) V- M A ( 2 0 ) V- M A ( 4 0 ) Ve hi c l e
nm
ole
s/m
g p
r
0
0 .0 0 1
0 .0 0 2
0 .0 0 3
0 .0 0 4
0 .0 0 5
0 .0 0 6
0 .0 0 7
0 .0 0 8
V- M A ( 10 ) V- M A ( 2 0 ) V- M A ( 4 0 ) Ve hi c l e
mic
rom
oles
/mg
pr
(a) (b)
Fig. 8.7. Effect of different doses of V-MA on a) Lipid peroxidation in mice brain b) Glutathione content in mice brain

0
0 . 5
1
1. 5
2
2 . 5
V- M A ( 10 ) V- M A ( 2 0 ) V- M A ( 4 0 ) Ve hi c l e
mic
rom
oles
of H
2O2
deco
mpo
sed/
min
/mg
pr
0
0 .5
1
1.5
2
2 .5
3
3 .5
4
4 .5
V- M A ( 10 ) V- M A ( 2 0 ) V- M A ( 4 0 ) Ve hi c l e
units
/mg
pr
(a) (b)
Fig. 8.8. Effect of different doses of V-MA on a) Catalase level in mice brain b) Superoxide dismutase level in mice
8.4. DISCUSSION
Antioxidants are radical scavengers which protect the human body against free
radicals that may cause pathological conditions such as ischemia, anaemia, asthma,
arthritis, inflammation, neuro-degenertion, Parkinson's diseases, mongolism, ageing
process and perhaps dementias (Polterait, 1997). Plants are potent biochemical
factories and have been components of phytomedicine since times immemorial and
man is able to obtain from them a wondrous assortment of industrial chemicals. Plant
based natural constituents can be derived from any part of plant like bark, leaves,
flowers, roots, fruits, seeds. The beneficial medicinal effects of plant materials
typically result from the combinations of secondary products present in the plant.
Flavonoids and flavones are widely distributed secondary metabolites with
antioxidant and antiradical properties (Nakayoma and Yamada, 1995). Antioxidant-
based drugs/formulations for the prevention and treatment of complex diseases like
atherosclerosis, stroke, diabetes, Alzheimer's disease, and cancer have appeared
during the last 3 decades (Devasagayam et al., 2004). This has attracted a great deal of
research interest in natural antioxidants. Subsequently, a worldwide trend towards the
use of natural phytochemicals present in berry crops, tea, herbs, oilseeds, beans, fruits,
and vegetables has increased.
In the present study different doses (10, 20 and 40 mg/kg) of valerian extracts
VVDR03, VVW02 and valerian oils V-PA and V-MA were administered orally to

mice daily for 14 days. And then on 14th day mice were sacrificed and brain
homogenate was prepared. Brain homogenates were then evaluated for modulation of
antioxidant defense system. It was found that none of the dose of VVW02 produced
any alteration in TBARS levels, glutathione content, SOD and catalase levels in brain
as compared to control group. VVDR03 however exerted antioxidant effect at 40
mg/kg and significantly increased the glutathione levels and SOD levels. All the doses
(10, 20 and 40 mg/kg) of VVDR03 increased catalase levels significantly. Similarly
40 mg/kg dose of VVDR03 significantly decreased the TBARS levels as compared to
control group. When the effect of valerian oils on antioxidant system was evaluated
then none of the dose of V-PA and V-MA altered TBARS levels, glutathione content,
catalase and SOD levels in mice brain as compared to control group.
From the in vitro study it was demonstrated that valerian extracts VVDR03
and VVW02 exhibited antioxidant activity out of which VVDR03 was more effective.
In the present study however only VVDR03 at 40 mg/kg was found to have
modulated antioxidant defense system in mice brain after two weeks dosing. VVW02,
V-PA and V-MA were not found to be effective in this study and the results correlate
well with the results of in vitro study. Iridoids, present in roots of Valeriana wallichii
might be responsible for modulatory effect on antioxidant defense system of mice
brain (Gamboa and Castro, 2004; Raju et al., 2004; Ahmad et al., 2008).

CONCLUSION
Chemical Screening
• The major constituents of the oils from roots and rhizomes of Valeriana
wallichii collected from different geographical region were separated and
characterized by GC, GC/MS and NMR. The chemical compositions of the
oils show two chemotypes within Valeriana wallichii. Patchouli alcohol and
8-Acetoxyl patchouli alcohol were isolated from the plant VW-II and since
Patchouli alcohol was the major compound it was characterised as V. wallichii
patchouli alcohol chemotype. From the plant VW-I, Maaliol and Patchouli
alcohol were isolated. Since Maaliol was one the major component of the
essential oil it was characterised as V. wallichii maaliol chemotype.
OH
H
H
OH
H
H
OAc
H
OH
H
H Patchouli alcohol 8-Acetoxyl patchouli alcohol Maaliol
• The extracts from the roots and rhizomes of both the chemotypes were
subjected to HPLC screening. The residue obtained after extraction was
fractionated on column chromatography to yield valepotriates. Didrovaltrate,
Isovaleroxyhydroxy didrovaltrate and 1-α Acevaltrate were isolated from V.
wallichii maaliol chemotype while Isovaleroxy didrovaltrate, 1-α Acevaltrate
and 1-Homoisovaltrate from patchouli alcohol chemotype.
O
O
O
O H3C
CH3
H
O
O
CH3
CH2OCOCH2CH(CH3)2
O
O
O
H
O
O
CH3CH3
O
O
O
O
Didrovaltrate Acevaltrate

O
O
O
H
O
CH3
O
CH3
O
O
CH3
O
O
O
O H3C
CH3
HO
O
O
CH3
O
CH3CH3
O
O
O
1-Homoisovaltrate Isovaleroxyhydroxy didrovaltrate (IVHD)
Biological screening
• When evaluated for psychopharmacological effect, none of the chemotypes
was found to have anxiolytic and muscle relaxing effect at tested doses (10, 20
& 40 mg/kg p.o).
• Extracts and oils of both the chemotypes were found effective in delaying
onset of death when tested in PTZ-induced convulsions. However they didn’t
afford any effect on percentage mortality as compared to control group. The
standard drug diazepam produced 100 % protection from mortality. Essential
oil of patchouli alcohol chemotype (V-PA) was more effective than maaliol
chemotype (V-MA) while both the extracts were equally effective in delaying
onset of death. V-PA was found to be most effective. Thus we can say that
both chemotypes possess very weak anticonvulsant effect which can be
attributed to the presence of valepotriates and sesquiterpenes.
• In acute study DCM extracts and essential oils of both the chemotypes
produced antiimmobilty effect without producing any motor stimulating
effect, however extract and oil of maaliol chemotype significantly decreased
locomotor activity as compared to vehicle treated group. Essential oils of both
the chemotypes were equally effective as antidepressant while extract of
maaliol type was found to be more effective than patchouli alcohol type. Out
of all, extract of patchouli alcohol type was found to be least effective in
producing antimmobilty effect. Since nitric oxide synthase inhibitor, L-

NAME, potentiated the effect of essential oils they are postulated to produce
antidepressant effect by modulating nitric-oxide signaling pathway while
GABA potentiating effect of extracts might be responsible for antidepressant
effect.
• In chronic study, both the oils were found to be equally effective as
antidepressants as in acute study. Extract of maaliol type was more effective
than patchouli alcohol type and this correlates well with acute study. The
results are further supported by neurobiochemical studies used to check
alterations in the neurotransmitter levels and both oils and extracts of both the
chemotypes significantly increased norepinephrine levels in mice brain after
14 days of treatment. Thus we can conclude that DCM extract of maaliol type
was more effective than patchouli alcohol type. Both the oils and DCM extract
of maaliol type were equally effective and the effect produced by them is
slightly more than the standard antidepressant, imipramine.
• While evaluating for analgesic effect, extract of chemotype patchouli alcohol
was found to be more effective than maaliol type in acetic acid writhing test
while both the oils produced comparative results in inhibiting acetic acid
writhings. From the present results we can conclude that analgesic effect
produced by both the oils in acetic acid writhing is equal to standard drug,
Aspirin. In tail flick test, only maaliol chemotype (both extract and oil) was
found to be effective but only at higher dose. Essential oils exerted action in
acetic acid writhing through COX pathway as they potentiated the effect of
aspirin while DCM extracts acted through pathway other than COX pathway.
• From the present study we found that DCM extracts of both the chemotypes
possess moderate in vitro antioxidant activity. Out of which, patchouli alcohol
type was more effective than maaliol type and this correlates well with in vivo
study also, done on mice. While in case of oils, both possess very weak in
vitro antioxidant activity while none was effective in modulating antioxidant
defense system in mice brain at the tested doses. Thus we can say that the
mixture of iridoids and essential oil present in DCM extracts of both the
chemotypes may be responsible for antioxidant effect.

BIBLIOGRAPHY
Abourashed EA, Koetter U, and Brattstrom A (2004) In vitro binding experiments
with a valerian, Hops and their fixed combination extract (Ze91019) to selected CNS
receptors. Phytomedicine 11: 633-638.
Ahmed M, Sadhu SK, Datta BK, Kunu JK, and Bachar SC (1997) Preliminary studies
on the antiinflammatory, analgesic and diuretic activity of stagninol, a sesquiterpene
isolated from Persicaria stagnina. Pharmazie 52(6): 472-475
Ahmad I, Chen S, Peng Y, Chen S, and Xu L (2008) Lipoxygenase inhibiting and
antioxidant iridoids from Buddleja crispa. J Enzyme Inhib Med Chem 23(1): 140-143.
Ahn H, Kim JY, Lee HJ, Kim YK, and Ryu J (2003) Inhibitors of inducible nitric
oxide synthase expression from Artemisia iwayomogi. Archives of Pharmacal
Research 26: 301-305.
Amarowicz R, Naczk M, and Shahidi F (2000) Antioxidant activity of crude tannins
of canola and rapeseed hulls. J Am Oil Chem Soc 77: 957-961.
Ames BN, Shigenaga MK, and Hagen TM (1993) Oxidants, antioxidants, and the
degenerative diseases of aging. Proc Natl Acad Sci U. S. A. 90: 7915-7922.
Amico-Roxas M, Caruso A, Trombadore S, Scifo R, and Scapagini U (1984)
Gangliosides antinociceptive effects in rodents. Arch Int Pharmacodyn Ther 272:
103-117.
Anderson D (1996) Antioxidant defences against reactive oxygen species causing
genetic and other damage. Mutat Res 350:103-108.
Andreatini R and Leite JR (1994) Effect of valepotriates on the behaviour of rats in
the elevated plus-maze during diazepam withdrawal. Eur J Pharmacol 260 (2-3):
233-235.

Andreatini R, Sartori ML, Seabra ML, and Leite JR (2002) effect of valepotriates
(valerian extract) in generalized anxiety disorder: A randomized placebo-controlled
pilot study. Phytother Res 16: 650-654.
Andrews JM and Nemeroff CB (1994) Contemporary management of depression. Am
J Med 97: 24S-30S.
Anwar HG, Arif-ullah K, Qaiser J, Fazal S, and Rukhsana G (2005) Antispasmodic
and blood pressure lowering effects of Valeriana wallichii are mediated through
K+channel activation. J Ethnopharmacol 100: 347-352.
Arnao MB (2000) Some methodological problems in the determination of antioxidant
activity using chromogen radicals: a practical case. Trends Food Sci Tech 11: 419-
421.
Arantes SF, Hanson JR, and Hitchcock PB (1999) The microbiological hydroxylation
of the sesquiterpenoid patchoulol by Mucor plumbeus. Phytochemistry 52: 635-638.
Arora RB and Arora C (1963) Hypotensive and tranquillizing activity of jatamansone
(valeranone) a sesquiterpene from Nardostachys jatamansi DC. in Pharmacology of
Oriental Plants (Chen KK and Mukerji B eds) pp 51-60, Pergamon Press, Oxford.
Awan HMH (1990) Kitabul Mufradat, GA Printers, Lahore.
Babbedge RC, Hart SL, and Moore PK (1993) Anti-nociceptive activity of nitric
oxide synthase inhibitors in the mouse: dissociation between the effect of L-NAME
and L-NMMA. J Pharm Pharmacol 45: 77-79.
Bartlett H and Eperjesi F (2003) Age-related macular degeneration and nutritional
supplementation: A review of randomised controlled trials. Ophthalmic Physiol Opt
23 (5): 383-99.

Becker H and Chavadej S (1985) Valepotriate production of normal and colchicine-
treated cell suspension cultures of Valeriana wallichii. J Nat Prod 48: 17-21.
Becker H, Chavadej S, Thies PW, Finner E (1984) Die Struktar neuer valepotriate aus
colchicin-behandelten Zelkulturen von Valeriana wallichii. Planta Med 50: 245-248.
Beyer CE, Boikess S, Luo B, and Dawson LA (2002) Comparison of the effects of
antidepressants on norepinephrine and serotonin concentrations in the rat frontal
cortex: an in-vivo microdialysis study. J Psychopharmacol 16: 297-304.
Beyer W, Imlay J, and Fridovich I (1991) Superoxide dismutases. Prog Nucl Acids
Res Mol Biol 40: 221-253.
Bhattacharjee (1998) Handbook of Medicinal Plants, Pointer Publisher, Jaipur.
Bhattacharyya D, Jana U, Debnath PK, and Sur TK (2007) Initial exploratory
observational pharmacology of Valeriana wallichii on stress management: a clinical
report. Nepal Med Coll J 9(1): 36-39.
Bol’shakova IV, Lozovskaia EL, and Sapezhinskii II (1997) Antioxidant properties of
a series of extracts from medicinal plants. Biofizika 42 (2): 480-483.
Bondet V, Brand-Williams W, and Berset C (1997) Kinetics and mechanism of
antioxidant activity using the DPPH free radical method. Lebensm Wiss Technol 30:
609-615.
Borsini F and Meli A (1988) Is the forced swimmining test a suitable model for
revealing antidepressant activity? Psychopharmacol 94:147-161.
Bos R, Woerdenbag HJ, Hendriks H, Smit HF, Wilkstrom HV, and Scheffer JJC
(1997) Composition of the essential oil from roots and rhizomes of Valeriana
wallichii DC. Flav Frag J 12: 123-131.

Bounthanh C, Bergmann C, Beck JP, Hagg-Berrurier M, and Anton R (1981)
Valepotriates, a new class of cytotoxic and antitumor agents. Planta Med 41: 21-28.
Brand-Williams W, Cuvelier ME, and Berset C (1995) Use of a free radical method to
evaluate antioxidant activity. Lebensm Wiss Technol 28: 25-30.
British Pharmaceutical Codex (1923) An imperial Dispensary for the use of Medical
Practitioners and Pharmacists, Pharmaceutical Press, London.
Burstein S, Varanelli C, and Slade LT (1975) Prostaglandins and cannabis. Inhibition
of biosynthesis by essential oil components of marihuana. Biochem Pharmacol 24(9):
1053-1054.
Butterweck V, Jurgenliemk G, Nahrstedt A, and Winterhoff H (2000) Flavonoids
from Hypericum perforatum show antidepressant activity in the forced swimming
test. Planta Med 66: 3-6.
Butterweck V, Nishibe S, Sasaki T, and Uchida M (2001) Antidepressant effects of
Apocynum venetum leaves in a forced swimming test. Biol Pharm Bull 24: 848-851.
Cabo J, Crespo ME, Jimenez J, and Zarzuelo A (1986) The spasmolytic activity of
various aromatic plants from the province of Granada. I. The activity of the major
components of their essential oils. Plantes Medicinales et Phytotherapie. 20: 213-218.
Cao G, Sofic E, and Prior RL (1997) Antioxidant and pro-oxidant behavior of
flavonoids; structure activity relationships. Free Rad Biol Med 22: 749-760.
Cao B and Hong GX (1994) Central inhibition action of Valeriana jatamansi Jones.
Zhongguo Zhong Yao Za Zhi 19(1): 40-42.
Carobrez AP and Bertoglio LJ (2005) Ethological and temporal analyses of anxiety-
like behavior: the elevated plus-maze model 20 years on. Neurosci Biobehav Rev
29:1193-1205.

Chang LW, Yen WJ, Huang SC, and Duh PD (2002) Antioxidant activity of sesame
coat. Food Chem 78: 347-354.
Chelikani P, Fita I, and Loewen P (2004) Diversity of structures and properties among
catalases. Cell Mol Life Sci 61 (2): 192–208.
Chen JF (2003) The adenosine A2A receptors as an attractive target for Parkinson’s
disease treatment. Drugs News Perspect 16: 597-604.
Chen YG, Yu LL, Huang R, Lv YP, and Gui SH (2005) 11-Methoxyviburtinal, a new
iridoid from Valeriana jatamansi. Arch Pharm Res 28 (10):1161-1163.
Chopra RN, Nayar SL, and Chopra IC (1956) Glossary of Indian Medicinal Plants,
Council of Scientific & Industrial Research, New Delhi,
Chopra RN, Nayar SL, and Chopra IC (1986) Glossary of Indian Medicinal Plants
(Including the Supplement), Council of Scientific & Industrial Research, New Delhi.
Collier HOJ, Dinneen JC, Johnson CA, and Schneider C (1968) The abdominal
constriction response and its suppression by analgesic drugs in the mouse. Br J
Pharmacol Chemother 32: 295-310.
Creissen G, Broadbent P, Stevens R, Wellburn A, and Mullineaux P (1996)
Manipulation of glutathione metabolism in transgenic plants. Biochem Soc Trans 24
(2): 465-469.
Da Silva GL, Matteussi AS, dos Santos ARS, Calixto JB, and Rodrigues ALS (2000)
Evidence for dual effects of nitric oxide in the forced swimming test and in the tail
suspension test in mice. Neuroreport 11(17): 3699-3702.
David JN (2006) The role of dopamine and norepinephrine in depression and
antidepressant treatment. J Clin Psychiatry 67[suppl 6]: 3-8.

David M (2001) Gabaergic mechanism in epilepsy. Epilepsia 42(3): 8-12.
Davies K (1995) Oxidative stress: the paradox of aerobic life. Biochem Soc Symp 61:
1-31.
Dawson TM and Synder SH (1994) Gases as biological messengers; nitric oxide and
carbon monoxide in the brain. J Neurosci 14: 5147-5159.
Decker EA, Warner K, Richards MP, and Shahidi F (2005) Measuring antioxidant
effectiveness in food. J Agric Food Chem 53: 4303-4310.
Del Río L, Sandalio L, Palma J, Bueno P, and Corpas F (1992) Metabolism of oxygen
radicals in peroxisomes and cellular implications. Free Radic Biol Med 13 (5): 557-
580.
Dev S (1997) Ethnotherapeutic and modern drug development: The potential of
Ayurveda. Cur Sci 73 (11): 909-928.
Devasagayam TPA, Tilak JC, and Boloor KK (2004) Free radicals and antioxidants in
human health. Curr Stat Fut Prosp JAPI 52: 794-804.
Dhir A and Kulkarni SK (2008) Possible involvement of nitric oxide (NO) signaling pathway
in the antidepressant-like effect of MK-801(dizocilpine), a NMDA receptor antagonist in mouse
forced swim test. Indian J Exp Biol 46 (3): 164-170.
Dhir A and Kulkarni SK (2007) Involvement of dopamine (DA)/serotonin (5-
HT)/sigma (σ) receptor modulation in mediating the antidepressant action of
ropinirole hydrochloride, a D2/D3 dopamine receptor agonist. Brain Res Bull 74: 58-
65.
Didna B, Debnath S, and Harigaya Y (2007) Naturally occurring iridoids: A Review.
Chem Pharm Bull 55 (2):159-222.

Dietz BM, Mahady GB, Pauli GF, and Farnsworth NR (2005) Valerian extract and
valerenic acid are partial agonists of the 5-HT(5a) receptor in vitro.
Mol Brain Res 138: 191-197.
Diplock AT (1994) Antioxidant and free radical scavengers, in Free Radical Damage
and its Control (Rice-Evans CA and Burdon RH eds) pp 113-130, Elsevier,
Amsterdam.
Diplock AT (1997) Will the ‘good fairies’ please proves to us that vitamin E lessens
human degenerative of disease? Free Rad Res 27: 511-532.
Dua VK, Alam MF, Pandey AC, Rai S, Chopra AK, Kaul VK, and Dash AP (2008)
Insecticidal activity of Valeriana jatamansi (Valerianaceae) against mosquitoes. J Am
Mosq Control Assoc 24(2): 315-318.
Duh PD, Tu YY, and Yen GC (1999) Antioxidant activity of water extract of Harug
Jyur (Chrysanthemum morifolium Ramat). Lebensm Wiss Technol 32: 269-277.
Duke JA and Ayensu ES (1985) Medicinal Plants of China, Reference Publications,
Washington.
Duke JA and Beckstrom SM (1996) Handbook of Medicinal Mints, Phytochemicals
and Biological Activities, CRC Press, Florida.
Dunaev VV, Trzhetsinskii SD, Tishkin VS, Fursa NS, and Linenko VI (1987)
Biological activity of the some of the valepotrites isolated from Valeriana
alliariifolia. Farmakol Toksikol 50(6): 33-37.
During MJ and Spancer DD (1993) Extracellular hippocampal glutamate and
spontaneous seizures in the conscious human brain. Lancet 341: 1607-1610.
Eadie MJ (2004) Could Valerian Have Been the First Anticonvulsant? Epilepsia 45:
1338 -1343.

Evans P and Halliwell B (1999) Free radicals and hearing. Cause, consequence, and
criteria. Ann N Y Acad Sci 884: 19-40.
Fang YZ, Yang S, and Wu G (2002) Free radicals, antioxidants, and nutrition.
Nutrition 18: 872-879.
Fauteck JD, Pietz B, Winterhoff H, and Wittkowski W (1996) Interaction of
Valeriana officinalis with melatonin receptors: a possible explanation of its biological
action. Proceedings of the 2nd International Conference on Phytomedicines, Munich:
ESCOP, Munich.
Fernandez S, Wasowski C, Paladini AC, and Marder M (2004) Sedative and sleep-
enhancing properties of linarin, a flavonoid isolated from Valeriana officinalis.
Pharmacol Biochem Behav 77(2): 399-404.
Fields HL (1987) Analgesic drugs, in Pain (Day W, ed) pp 272, Mac-Graw-Hill,
USA.
Foster S and Duke JA (1990) A Field Guide to Medicinal Plants. Houghton Mifflin
Co, US.
Francis AJP and Dempster RJW (2002) Effect of valerian, Valeriana edulis, on sleep
difficulties in children with intellectual deficits: randomized trial. Phytomedicine 9:
273-279.
Gamboa IC and Castro O (2004) Iridoids from the aerial parts of Verbena littoralis
(Verbenaceae) J Phytochem 65: 2369-2372.
Garthwaite J (1991) Glutamate, nitric oxide and cell-cell signaling in the nervous
system. Trends Neurosci 14: 60-67.
Garthwaite J and Boulton CL (1995) Nitric oxide signaling in the central nervous
system. Annu Rev Physiol 57: 683-706.

Gilani AH, Khan A, Jabeen Q, Subhan F, and Ghafar R (2005) Antispasmodic and
blood pressure lowering effects of Valeriana wallichii are mediated through K+
channel activation. J Ethnopharmacol 100: 347-352.
Golshani S, Karamkhani F, Monsef-esfehani HR, and Abdollahi M (2004)
Antinociceptive effects of the essential oil of Dracocephalum kotschyi in the mouse
writhing test. J Pharm Pharmaceut Sci 7(1): 76-79.
Grice HC (1986) Safety evaluation of butylated hydroxytoluene (BHT) in the liver,
lung and gastrointestinal tract. Food Chem Toxicol 24: 1127-1130.
Grisham MB (1992) Reactive metabolites of oxygen and nitrogen in biology and
medicine, RG Landes Co., Austin.
Gülçin I, Büyükokuro¡glu MF, Oktay M, and Küfrevio¡glu ÖI (2002) On the in vitro
antioxidant properties of melatonin. J Pineal Res 33: 167-171.
Gupta LK and Shah SC (1981) Cultivation and importance of Valeriana wallichii in
the hills of Uttar Pradesh. Ind Drugs 393-395.
Ha JH, Lee DU, Lee JT, Kim, JS, Yong CS, and Kin JA (2000) 4
Hydroxybenzaldehyde from Gastrodia elata as active in the antioxidation and
GABAergic neuromodulation of the rat brain. J Ethnopharmacol 73: 329-333.
Hackinski V (1998) New antiepileptic drugs, the cost of innovation. Arch Neurol 55:
1142-1145.
Halliwell B (1996) Antioxidants in human health and disease. Annu Rev Nutr 16: 33-
50.
Halliwell B (2000) Why and how should we measure oxidative DNA damage in
nutritional studies? How far have we come? Am J Clin Nutr 72: 1082-1087.

Halliwell B and Gutteridge JMC (1989) Free radicals, ageing and disease, in Free
radicals biology and medicine (Halliwell B and JMC Gutteridge JMC eds) pp 416,
Clarendon Press, Oxford.
Halliwell B, Gutteridge JM, and Cross CE (1992) Free radicals, antioxidants, and
human disease: where are we now? J Lab Clin Med 119: 598-620.
Hanns UZ (2007) Prostanoids in nociception and pain. Biochem Pharmacol 73: 165–
174.
Hardman JG, Limbird LE, and Goodman GA (2001) Goodman and Gilman's: The
Pharmacological Basis of Therapeutics, 10th ed. The McGraw Hill Co., New York.
Harkin AJ, Karen HB, Brandy C, and Paul IA (1999) Nitric oxide synthase inhibitors
have antidepressant-like properties in mice 1. Acute treatments are active in the
forced swim test. Eur J Pharmacol 372: 207-213.
Hattesohl M, Feistel B, Sievers H, Lehnfeld R, Hegger M, and Winterhoff H (2008)
Extracts of Valeriana officinalis show anxiolytic and antidepressant effects but neither
sedative nor myorelaxant properties. Phytomedicine 15: 2-15.
Hazelhoff B, Malingre TM, and Meijer DK (1982) Antispasmodic effects of
Valeriana compounds: an in-vivo and in-vitro study on the guinea pig ileum. Arch Int
Pharmacodyn Ther 257 (2): 274-287
Hendricks H, Bos R, Allersma DP, Malingré TM, and Koster AS (1981)
Pharmacological screening of valerenal and some other components of essential oil of
Valeriana officinalis. Planta Med 42: 62-68.
Hendriks H, Bos R, Woerdenbag HJ, and Koster AS (1985) Central nervous
depressant activity of valerenic acid in the mouse. Planta Med 51: 28-31.

Herbalist RU (1999) Valerian Root: Valeriana officinalis Analytical, Quality control
and Therapeutic Monograph. American Herbal Pharmacopoeia, Scotts Valley, CA.
Hertog MG, Feskens EJ, Hollman PC, Katan MB, and Kromhout D (1993) Dietary
antioxidant flavonoids and risk of coronary heart disease: the Zutphen Elderly Study.
Lancet 342: 1007-1011.
Hiller KO and Zetler G (1996) Neuropharmacological studies on ethanol extracts of
Valeriana officinalis: Behavioural and anticonvulsant properties. Phytother Res 10:
145-151.
Hochstein P and Atallah AS (1988) The nature of oxidant and antioxidant systems in
the inhibition of mutation and cancer. Mut Res 202: 363-375.
Hogg S (1996) A review of the validity and variability of the elevated plus-maze as an
animal model of anxiety. Pharmacol Biochem Behav 54: 21-30.
Holmes P (1989) The Energetics of Western Herbs, Artemis Press, Boulder, CO.
Houghton PJ (1999) The scientific basis for the reputed activity of valerian. J Pharm
Pharmacol 51: 505-512.
Howard M (1987) Traditional Folk Remedies, A comprehensive Herbal. Ebury Press,
London.
Huo Y, Guo C, Didna B, Debnath S, and Harigaya Y (2007) Naturally Occurring
Iridoids: A Review, Part 1 Chem Pharm Bull 55(2): 159-222.
Ikeda Y, Ueno A, Naraba H, and Oh-ishi S (2001) Involvement of vanilloid receptor
VR1 and prostanoids in acetic acid induced writhing responses of mice. Life Sci 69:
2911-2919.

Imaida K, Fukushima S, Shivai T, Ohtani M, Nakanishi K, and Ito N (1983)
Promoting activities of butylated hydroxyl anisole and butylated hydroxytoulene on 2-
stage urinary bladder carcinogenesis and inhibition of γ-gluatamyl transpeptidase-
positive foci development in the liver of rats. Carcinogen 4: 885-889.
Jacobo-Herrera NJ , Vartiainen N, Bremner P, Simon Gibbons S, Jari Koistinaho, and
Heinrich M (2006) NF-κB modulators from Valeriana officinalis. Phytother Res 20:
917-919.
Jollow D, Mitchell L, Zampaglione N, and Gillete J (1974) Bromobenze induced liver
necrosis: protective role of glutathione and evidence for 3, 4-bromobenzenoxide as
the hepatotoxic intermediate. Pharmacol 111: 151-159.
Kamboj VP (2000) Herbal Medicine. Cur Sc 78(1): 35-39.
Kapoor LD (1990) CRC Handbook of Ayurvedic Medicinal Plants, CRC Press, Boca
Raton.
Kaputlu I and Uzbay T (1997) L-NAME inhibits pentylenetetrazol and strychnine-
induced seizures in mice. Brain Res 753: 98-101.
Khan SA and Khatoon S (2008) Ethnobotanical studies on some useful herbs of
Haramosh and Bugrote valleys in Gilgit, northern areas of Pakistan. Pak J Bot 40(1):
43-58.
Khandelwal KR (2007) Practical Pharmaocology,Techniques and Experiments,
Nirali Prakashan, Pune, India.
Khare CP (2007) Indian Medicinal Plants: An illustrated Dictionary, Springer
Publisher, US.

Kim J, Seo SM, Lee SG, Shin SC, and Park IK (2008) Nematicidal activity of plant
essential oils and components from coriander (Coriandrum sativum), Oriental
sweetgum (Liquidambar orientalis), and valerian (Valeriana wallichii) essential oils
against pine wood nematode (Bursaphelenchus xylophilus). J Agric Food Chem
56(16): 7316-7320.
Knowles RG and Moncada S (1994) Nitric oxide synthases in mammals. Biochem J
298: 249-258.
Kono Y (1978) Generation of superoxide radical during autooxidation of
hydroxylamine and an assy for superoxide dismutase. Arch Biochem Biophys 186:
189-195.
Koudou J, Abena AA, Ngaissona P and Bessière JM (2005) Chemical composition
and pharmacological activity of essential oil of Canarium schweinfurthii. Fitoterapia
76: 700-703
Kubacka M, Tadeusz L, Ryszard C, Bozena F, and Stanislaw L (2006) Anticonvulsant
and antidepressant activity of the selected terpene GABA derivatives in experimental
tests in mice. Pharmacol Rep 58: 936-943.
Kulkarni SK (1999) Handbook of Experimental Pharmacology, 3rd ed, Vallabh
Prakashan, New Delhi.
Kulkarni SK and Jain NK (2005) Coxibs: The new super aspirins or unsafe pain
killers? 37: 86-89.
Kulkarni SK and Mehta AK (1985) Purine nucleoside-mediated immobility in mice:
reversal by antidepressants. Psychopharmacol 85: 460-463.
Lacher SK, Mayer R, Sischardt K, Nieber K, and Muller CE (2007) Isolation of valerian
extracts of different polarity with adenosine receptors: Identification of isovaltrate as an
inverse agonist at A1 receptors. Biochem Pharmacol 73: 248-258.

Leathwood PD and Chauffard F (1985) Aqueous extract of valerian reduces latency to
fall asleep in man. Planta Med 51: 144-148.
Leathwood PD, Chauffard F, Heck E, and Munoz-Box R (1982) Aqueous extract of
valerian root (Valeriana officinalis L.) improves sleep quality in man. Pharmacol
Biochem Behav 17: 65-71.
Lei SZ, Pan ZH, Aggarwal SK, Chen HS, Hartman J, Sucher NJ, and Lipton SA
(1992) Effect of nitric oxide production on the redox modulatory site of the NMDA
recptor-channel complex. Neuron 8: 1087-1099.
Lee SK, Hong CH, Huh SK, Kirn SS, Oh OJ, Min HY, Park KK, Chung WY and
Wang JK (2002) Suppressive effect of natural sesquiterpenoids on inducible
cycloxygenase (COX-2) and nitric oxide synthase (iNOS) activity in mouse
macrophase cells. J Env Path Toxicol Oncology 21:141-148.
Librowski T, Czarnecki R, Mendyk A, and Jastrzebska M (2000) Influence of new
monoterpene homologues of GABA on the central nervous system activity in mice.
Pol J Pharmacol 52: 317-321.
Linster CL and Van SE (2007) Vitamin C. Biosynthesis, recycling and degradation in
mammals. FEBS J 274 (1): 1-22.
Loscher W, Fassbender CP, and Nolthing B (1991) The role of technical, biological
and pharmacological factors in the laboratory evaluation of anticonvulsant drugs.
Maximal electroshock seizure models. Epilepsy Res 8: 79-84.
Lowenstein CJ, Dinerman JL, and Synder SH (1994) Nitric oxide-a physiological
messenger. Ann Intern Med 120: 227-230.
Luck H (1963) Catalase, in Methods of Enzymatic Analysis. (Bergmeyer HU, ed) pp
885-888, Academic Press, New York.

Maddipati KR and Marnett LJ (1987) Characterisation of the major hydroperoxide-
reducing activity of human plasma. Purification and properties of a selenium-
dependent glutathione peroxidase. J Biol Chem 262: 17398-17403.
Mahinda W and Shahidi F (2000) Scavenging of reactive-oxygen species and DPPH
free radicals by extracts of borage and evening primrose meals. Food Chem 70:17-26.
Malva JO, Santos S, and Macedo T (2004) Neuroprotective properties of Valeriana
officinalis extracts. Neurotox Res 6(2): 131-140.
Manzoni O and Bockaert J (1993) Nitric oxide synthase activity endogenously
modulates NMDA receptors. J Neurochem 61: 368-370.
Manzoni O, Prezeau L, Marin P, Deshager S, Bockaert J, and Fagni L (1992) Nitric
oxide-induced blockade of NMDA receptors. Neuron 8: 653-662.
Marder M, Viola H, Wasowski C, Fernandez S, Medina JH, and Paladini AC (2003)
6-Methylapigenin and hesperidin: new Valeriana flavonoids with activity on the
CNS. Pharmacol Biochem Behav 75: 537–545.
Mathela CS, Tiwari M, Sammal SS, and Chanotiya CS (2005a) Valeriana wallichii
DC, a new chemotype from Northwestern Himalaya. JEOR 17: 672-675.
Mathela CS, Chanotiya CS, Sati S, Sammal SS, and Wray V (2007)
Epoxysesquithujene, a novel sesquiterpenoid from Valeriana hardwickii var.
hardwickii. Fitoterpia 78: 279-282.
Mathela CS, Chanotiya CS, Sammal SS, Pant AK, and Pandey S (2005b)
Compositional diversity of terpenoids in the himalayan Valeriana genera. Chem
Biodiver 2: 1174-1182.
McCord JM (2000) The evolution of free radicals and oxidative stress. Am J Med Sci
108: 652-659.

Meister A (1988) Glutathione metabolism and its selective modification. J Biol Chem
263 (33): 17205-17208.
Meister A and Anderson M (1983) Glutathione. Annu Rev Biochem 52: 711-760.
Mennini T, Bernasconi P, Bombardelli E, and Morazzoni P (1993) In vitro study on
the interaction of extracts and pure compounds from Valeriana officinalis roots with
GABA, benzodiazepine and barbiturate receptors in rat brain. Fitoterapia 44(4): 291-
300.
Miller LG and Murray WJ (1998) Herbal Medicinals: A Clinicians Guide, Haworth
Press, Philadelphia.
Miyasaka LS, Atallah AN, and Soares BG (2006) Valerian for anxiety disorders.
Cochrane Database Syst Rev Issue 4. doi:10.1002/14651858.CD004515.
Moncada S, Palmer RMJ, and Higgs EA (1991) Nitric oxide: physiology,
pathophysiology and pharmacology. Pharmacol Rev 43: 109-142.
Moore LL, Minne K, and Moore MB (2003) AltCareDex®
System. MICROMEDEX,
Inc., Englewood, Colorado.
Morazzoni P and Bombardelli E (1995) Valeriana officinalis: Traditional use and
recent evaluation of activity. Fitoterapia 66: 99-112.
Moreira MR, Cruz GM, Lopes MS, Albuquerque AA, and Leal-Cardoso JH (2001)
Effects of terpineol on the compound action potential of the rat sciatic nerve. Braz J
Med Biol Res 34: 1337-1340.
Motai H and Kitanaka S (2005) Sesquiterpene phenylpropanoids from Ferula
fukanensis and their nitric oxide production inhibitory effects. J Nat Prod 68(3): 365-8.

Muller DN, Heissmeyer V, Dechend R, Hamich F, Park JK, Fiebeler A, Shagdarsuren
E, Theuer J, Elger M, and Pilz B (2001) Aspirin inhibits NF-κB and protects from
angiotensin II-induced organ damage. FASEB J 15: 1822-1824.
Murakami N, Ye Y, Kawanishi M, Aoki, S, Kudo N, Yoshid M, Nakayama EE,
Shioda T, and Kobayashi M (2002) New Rev-transport inhibitor with anti-HIV
activity from Valerianae radix. Bioorg Med Chem Lett 12(20): 2807-2810.
Nadkarni KM (1976) Indian Materia Medica, 3rd ed, Popular Prakashan, Bombay.
Nakayoma J and Yamada M (1995) Suppression of active oxygen-induced
cytotoxicity by flavonoids. Biochem Pharmcol 45: 265-267.
Nanjappa KN, Shalam M, Harish MS, Rao PS, and Kutty BM (2007)
Pharmacological and neurobiochemical evidence for antidepressant-like effect of
Sumind, a herbal product in animals. The Internet Journal of Nutrition and Wellness
4:Number 1.
Naryanan CS, Kulkarni KS, Vaidya AS, Kanthamani S, Lakshmi KG, Bapat BV,
Patnikar SK, Kulkarni SN, Kelkar GR, and Bhattacharya SC (1964) Terpenoids-
XLVI:Components of Indian valerian root oil. Tetrahadron 20: 963-968.
Navarro MC, Montilla MP, Martin A, Jimenez J and Utrilla MP (1992) Free radicals
and antihepatotoxic activity of Rosmarinus tomentosus. Planta Med 59: 312-314.
Niki E (1992) Free radical pathology and antioxidants: overview. J Nutr Sci Vitaminol
12: 538-540.
Niki E (2001) Free Radicals in the 1900's: from in vitro to in vivo. Free Rad Res 33:
693-704.
Nishiya K, Kimura T, Takeya K, and Itokawa H (1992) Sesquiterpenoids and iridoid
glycosides from Valeriana fauriei. Phytochemistry 31: 3511-3514.

Nishiya K, Tsyjiyama T, Kimura T, Takeya K, Itokawa H, and Iitaka Y (1995)
Sesquiterpenoids from Valeriana fauriei Phytochemistry 39: 713-714.
Oktay M, Gülçin I, and Küfrevioˇglu ÖI (2003) Determination of in vitro antioxidant
activity of fennel (Foeniculum vulgare) seed extracts. Lebensm Wiss Technol 36: 263-
271.
Olsen RWJ (1981) GABA-benzodiazepine-barbiturate receptor interactions. Journal
of Neurochem 37: 1-3.
Oshima Y and Matsuoka S (1995) Antidepressant principles of Valeriana fauriei
roots. Chem Pharm Bull 43(1):169-170.
Oyaizu M (1986) Studies on product of browning reaction prepared from glucose
amine. Jap J Nutr 44: 307-315.
Ozbek H, Tas A, Ozgokce F, Selcuk N, Alp S, and Karagoz S (2006) Evaluation of
median lethal dose and analgesic activity of Foeniculum vulgare Miller essential oil.
Int J Pharmacol 2: 181-183.
Pande A, Uniyal GC, and Shukla YN (1994) Determination of chemical constituents
of Valeriana wallichii by reverse phase HPLC plus. Ind J Pharm Sci 56: 56-58.
Pandey A and Shukla YN (1993) Naphthoic acid derivative from Valeriana wallichii.
Phytochem 32: 1350-1359.
Pandey G (1995) Medicinal Plants of Himalaya, Sri Satguru Publications, New Delhi.
Pelligrino DA, Baugham VL, and Koenig HM (1996) Nitric oxide and the brain, in
International Anesthesiology Clinics. Topics in Neuroanesthesia (Jaffe RA and
Gifford RG eds) pp. 113-132, Mass: Little Brown and Co, Boston.

Pellow S, Chopin P, File SE, and Briley M (1985) Validation of open:closed arm
entries in an elevated plus-maze as a measure of anxiety in the rat. J Neurosci
Methods 14: 149-167.
Petty F (1995) GABA and mood disorders: a brief review and hypothesis. J Affect
Disord 34: 275-281.
Peyrat-Maillard MN, Bonnely S, and Berset C (2000) Determination of the
antioxidant activity of phenolic compounds by coulometric detection. Talanta 51:
709-716.
Piccinelli AL, Arana S, Caceres A, Bianca REV, Sorrentino R, and Rastrelli L (2004)
New Lignans from the roots of Valeriana prionophylla with antioxidative and
vasorelaxant activities. J Nat Prod 67: 1135-1140.
Polterait O (1997) Antioxidants and free-radical scavengers of natural origin. Current
Org Chem 1: 415-440.
Porsolt RD (1981) Behavioural despair, in Antidepressants: Neurochemical,
Behavioural and Clinical Perspectives (Enna SJ, Malick JB, and Richelson E eds) pp
121-139, Raven Press, New York.
Porsolt RD, Bertin A, and Jalfre M (1977) Behaviour despair models in mice: a
primary screening test for antidepressants, Arch Int Pharmacodyn 229: 327-336.
Poyares DR, Guilleminault C, Ohayon MM, and Tufik S (2002) Can valerian improve
the sleep of insomniacs after benzodiazepine withdrawal? Prog
Neuropsychopharmacol Biol Psychiatry 26: 539-545.
Prakash V (1999) Indian Valerianaceae: A Monograph on a Medicinally Important
Family. Scientific Publisher, Jodhpur.

Rabe T and Staden JV (1997) Antibacterial activity of South African plants used for
medicinal purposes. J Ethnopharmacol 56: 81-87.
Raj PP (1996) Pain mechanism, in Pain medicine: A Comprehensive Review. (Raj PP
ed) pp 12-23, Mosby-Year Book, Missouri.
Rajnarayana K, Reddy MS, Chaluvadi MR, and Krishna DR (2001) Bioflavonoids
classification, pharmacological, biochemical effects and therapeutic potential. Indian
J Pharmacol 33: 2-16.
Raju BL, Lin SJ, Hou WC, Lai ZY, Liu PC, and Hsu FL (2004) Antioxidant iridoid
glucosides from Wendlandia formosana. Natl Prod Res 18(4): 357-364.
Re es DD, Palmer RM, Schulz R, Hodson HF, and Moncada S (1990)
Characterization of three inhibitors of endothelial nitric oxide synthase in vitro and in
vivo. Br J Pharmacol 101: 746-752.
Reddy DS, Kaur G, and Kulkarni SK (1998) Sigma (sigma1) receptor mediated anti-
depressant-like effects of neurosteroids in the porsolt swim test. Neuroreport 9: 3069-
3073.
Rehni AK, Pantlya HS, Shri R, and Singh M (2007) Effect of chlorophyll and
aqueous extracts of Bacopa monniera and Valeriana wallichii on ischemia and
reperfusion-induced cerebral injury in mice. Indian J Exp Biol 45(9): 764-769.
Richard K, Karel P, Dmitrij T, Jana J, Daniela J, and Milan R (1997) Chronic
blockade of nitric oxide synthesis elevates plasma levels of catecholamines and their
metabolites at rest and during stress in rats. Neurochem Res 22: 995-100.
Riedel E, Hänsel R, and Ehrke G (1982) Inhibition of gamma-aminobutyric acid
catabolism by valerenic acid derivatives. Planta Med 46: 219-220.

Riley JCM and Behrman HR (1991) Oxygen radicals and reactive oxygen species in
reproduction. Proc Soc Exp Biol Med 198: 781-791.
Rimm EB, Stampfer MJ, Ascherio A, Giovannucci E, Colditz GA, and Willett WC
(1993) Vitamin E consumption and the risk of coronary heart disease in men. N Engl
J Med 328(20): 1450-1456.
Robello M, Amico C, Bucossi G, Cupello A, Rapallino MV, and Thellung S (1996)
Nitric oxide and GABAA receptor function in the rat cerebral cortex and cerebellar
granule cells. Neuroscience 74: 99-105.
Rodgers RJ (1997) Animal models of anxiety: where next? Behav Pharmacol 8:
477-496.
Ron BH, Willis CA, Bone K, and Morgan M (2000) Herbal products: active
constituents, mode of action and quality control. Nutrition Research Reviews 13:
47-77.
Rundfeldt C, Koch R, Richter A, Mevissen M, Gerecke U, and Loscher W (1995)
Dose dependent anticonvulsant and proconvulsant effects of nitric oxide synthase
inhibitors on seizure threshold in a cortical stimulation model in rats. Eur J
Pharmacol 274: 73-81.
Said HM (1970) Hamdard Pharmacopoeia of Eastern Medicine, Times Press,
Karachi.
Sakakibara H, Ishida K, Grundmann O, Nakajima J, Seo S, Butterweck V, Minami Y,
Saito S, Kawai Y, Nakaya Y, and Terao J (2006) Antidepressant effect of extracts
from Ginkgo biloba leaves in behavioural models. Biol Pharm Bull 29: 1767-1770.
Sakakibara H, Yoshino S, Yoschchika K, and Terao J (2008) Antidepressant-like
effect of Onion (Alliun cepa) powder in a rat behavioural model of depression. Biosci
Biotechnol Biochem 72(1): 94-100.

Sakamoto T, Mitani Y, and Nakajima K (1992) Psychotropic effects of Japanese
valerian root extract. Chem Pharm Bull 40: 758-761.
Salminen A, Lehtonen M, Suuronen T, Kaarniranta K, and Huuskonen J (2008)
Terpenoids: natural inhibitors of NF-κB signaling with anti-inflammatory and
anticancer potential. CMLS 65: 2979-2999.
Sammal SS (2005) PhD Thesis, Kumaun University, Nainital.
Santos FA and Rao VSN (2000) Antiinflammatory and antinociceptive effect of 1,8-
cineole, a terpenoid oxide present in many plant essential oil. Phytother Res 14:
240-244.
Santos FA, Rao VSN, Silveira ER (1997) Anti-inflammatory and analgesic activities
of the essential oil of Psidium guianense. Fitoterapia 68: 65-68.
Santos MS, Ferreira F, Cunha AP, Carvalho AP, and Macedo T (1994a) An aqueous
extract of valerian influences the transport of GABA in synaptosomes. Planta Med
60: 278-279.
Santos MS, Ferreira F, Faro C, Pires E, Carvalho AP, Cunha AP, and Macedo T
(1994b) The amount of GABA present in aqueous extracts of valerian is sufficient to
account for (3H)GABA release in synaptosomes. Planta Med 60: 475-476.
Sati S (2002) PhD Thesis, Kumaun University, Nainital.
Sati S and Mathela CS (2004) Essential oil composition of Valeriana hardwickii var.
arnottiana from the Himalayas. Flav Frag J 20: 299-301.
Sayyah M, Peirovi A, and Kamalinejad M (2002) Anti-nociceptive effect of the fruit
essential oil of Cuminum cyminum L. in rat. Iran Biomed J 6(4): 141-145.

Sayyah M, Saroukhani G, Peirovi A, and Kamalinejad M (2003) Analgesic and anti-
inflammatory activity of the leaf essential oil of Laurus nobilis Linn. Phytother Res
17: 733-736.
Schultz OE and Eckstein K (1962) Isolation of a substance from the rhizomes and
roots of Indian valerian (Valeriana wallichii D.C.) with marked effect in the Haffner
test. Arzneimittelforschung 12: 1005-1012.
Schwarz K, Bertelsen G, Nissen LR, Gardner PT, Heinonen MI, Hopia A, Tuong HB,
Lambelet P, McPhail D, Skibsted LH, and Tijburg L (2001) Investigation of plant
extracts for the protection of processed foods against lipid oxidation. Comparison of
antioxidant assays based on radical scavenging, lipid oxidation and analysis of the
principal antioxidant compounds. Eur Food Res Technol 212: 319-328.
Seligman MEP, Walker EF, and Rosenhan DL (2001) Abnormal Psychology, 4th ed.
W.W. Norton & Company, New York.
Shalam M, Shantakumar SM, and Narasu ML (2007) Pharmacological and
biochemical evidence for the antidepressant effect of the herbal preparation Trans-01.
Indian J Pharmacol 39: 231-234.
Sharma R (2003) Medicinal Plants of India: An encyclopedia. Daya Publishing
House, New Delhi.
Shrivastava SC and Sisodia CS (1970) Analgesic studies on Vitex negundo and
Valeriana wallichii. Indian Vet J 47: 170-175.
Sieghart W (1992) GABAA receptors: ligand-gated Cl− ion channels modulated by
multiple drug-binding sites. Trends Pharmacol Sci 13: 446-450.
Sies H (1997) Oxidative stress: oxidants and antioxidants. Exp Physiol 82 (2): 291-295.

Sies H and Cadenas E (1985) Oxidative stress: damage to intact cells and organs.
Philos Trans R Soc Lond B Biol Sci 311: 617-631.
Singh G and Kachroo P (1976) Forest Flora of Srinagar. Bishen Singh Mahendra Pal
Singh, Dehradun.
Singh N, Gupta AP, Singh B, and Kaul VK (2006) Quantification of Valerenic Acid
in Valeriana jatamansi and Valeriana officinalis by HPTLC. Chromatographia 63:
209-213.
Singh S and Majumdar DK (1995) Analgesic activity of Ocimum sanctum and its
possible mechanism of action. Int J Pharmacol 33(3):188-192.
Smith GB and Olsen CE (1995) Functional domains of GABAA receptors. Trends
Pharmacol Sci 16: 162-168.
Sood VK (1965) Isolation of Indian Valerian oil from V. wallichii roots and rhizomes.
Perfum Essent oil Rec 56: 656-657.
Southam E and Garthwaite J (1993) The nitric oxide-cyclic GMP signaling pathway
in rat brain. Neuropharmacol 32: 1267-1277
Souza MC, Siani AC, Ramos MF, Menezes-de-Lima OJ, and Henriques MG (2003)
Evaluation of anti-inflammatory activity of essential oils from two Asteraceae
species. Pharmazie 58: 582-586.
Spiteller G (2001) Lipid peroxidation in aging and age-dependent diseases. Exp
Geront 36: 1425-1457.
Sreejayan N and Rao MNA (1997) Nitric oxide scavenging by curcuminoids. J
Pharm Pharmacol 49: 105-107.

Sreejayan N and Rao MNA (1996) Free radical scavenging by curcuminoids. Drug
Research 46: 169-171.
Sripriya G, Chandrasekharan K, Murty VS, and Chandra TS (1996) ESR
spectroscopic studies on free radical quenching action of finger millet (Eleusine
coracana). Food Chem 47: 537-540.
Stahl SM (1998) Essential Psychopharmacology: Neuroscientific Basis and Practical
Applications. Cambridge University Press, Cambridge.
Stanner SA, Hughes J, Kelly CN, and Buttriss J (2004) A review of the
epidemiological evidence for the ‘antioxidant hypothesis’. Public Health Nutr 7 (3):
407-422.
Stief TW (2003) The physiology and pharmacology of singlet oxygen. Med
Hypotheses 60: 567-572.
Stoner GD and Mukhtar H (1995) Polyphenols as cancer chemopreventive agent. J
Cell Biochem 22:169-180.
Tang YP, Liu X, and Biao Yu (2003) Two New Flavone Glycosides from Valeriana
jatamansi. J Asian Nat Prod Res 5: 257-261.
Tang YP, Liu X, and Yu B (2002) Iridoids from the rhizomes and roots of Valeriana
jatamansi. J Nat Prod 65: 1949-1952.
Tegeder I, Niederberger E, Israr E, Guhring H, Brune K, Euchenhofer C, Grosch S,
and Geisslinger G (2001) Inhibition of NF- B and AP-1 activation by R- and S-
flurbiprofen. FASEB J 15: 2-4.
Tepe B, Sokmen M, Akpulat HA, Daferera D, Polissiou M, and Sokmen A (2005)
Antioxidative activity of the essential oils of Thymus sipyleus subsp. Sipyleus var.
rosulans. J Food Eng 66: 447-454.

Tewari M (2002) PhD Thesis, Kumaun University, Nainital.
Thabrew MI, Hughes RD, and McFarlane IG (1998) Antioxidant activity of Osbeckia
aspera. Phytother Res 12: 288-290.
The Wealth of India. Raw Materials Vol. X (2005) CSIR, New Delhi, 424-427.
Thies PW (1968) Linarin-isovalerianate, a currently unknown flavonoid from
Valeriana wallichii D.C. Report on the active substances of Valeriana. Planta Med
16(4): 363-371.
Thies PW, Finner E, and David S (1981) Über die wirkstoffe des baldrians. Planta
Med 41: 15-20.
Treit D, Menard J, and Royan C (1993) Anxiogenic stimuli in the elevated plus-
maze. Pharmacol Biochem Behav 44: 463-469.
Tripathi KD (2005) Essentials of Medical Pharmacology, 5th ed. Medical Publishers
Ltd, New Delhi.
Trullas R (1997) Functional NMDA antagonists: a new class of antidepressant agents,
in Antidepressants: New pharmacological Strategies (Skolnick P ed) pp 103–124,
Humana Press, Totowa, NJ.
Usher G (1974) A Dictionary of Plants Used by Man, Hafner press, New York.
Viswanatha SAHM, Thippeswamy AHM, Manjula DV, and Mahendra KCB (2006)
Some neruopharmacological effects of the methanolic root extract of Cissus
quadrangularis in mice. African Journal of Biomedical Research 9: 69-75.
Wang J, Wen L, Huang Y, Chen Y, and Ku M (2006) Dual effects of antioxidants in
neurodegeneration: direct neuroprotection against oxidative stress and indirect
protection via suppression of glia-mediated inflammation. Curr Pharm Des 12(27):
3521-3533.

Wasowski C, Marder M, Viola H, Medina JH, and Paladini AC (2002) Isolation and
identification of 6-methylapigenin, a competitive ligand for the brain GABA(A)
receptors, from Valeriana wallichii. Planta Med 68: 934-936.
Weissman BA, Kadar T, Brandeis R, and Shapira S (1992) NG-nitro-L-arginine
enhances neuronal death following transient forebrain ischemia in gerbils. Neurosci
Lett 146: 139-142.
Willner P (1984) The validity of animal models of depression. Psychopharmacol 83:
1-16.
Wills ED (1966) Mechanisms of lipid peroxide formation in animal tissues. Biochem
J 99: 667-676.
Wintergerst E, Maggini S, and Hornig D (2006) Immune-enhancing role of vitamin C
and zinc and effect on clinical conditions. Ann Nutr Metab 50 (2): 85-94.
Wu JK, Huo JH, and DU XW (2007) Pharmacological effects of volatile of Valeriana
amurensis on CNS. Zhong Yao Cai 30: 977-980.
Yamauchi S, Sugahara T, Matsugi J, Someya T, Masuda T, Kishida T, Akiyama K,
and Maruyama M (2007) Effect of the benzylic structure of lignan on antioxidant
activity. Biosci Biotechnol Biochem 71(9): 2283-2290.
Yan L, Burbiel JC, Maa BA, and Muller CE (2003) Adenosine receptor agonists from
basic medicinal chemistry to clinical development. Expert Opin Emerg Drugs 8: 537-
576.
Yang GY and Wang W (1994) Clinical studies on the treatment of coronary heart
disease with Valeriana officinalis var latifolia. Chung Kuo Chung Hsi I Chieh Ho Tsa
Chih 14(9): 540-542.

Yoon JH, Lim HJ, Lee HJ, Hee-doo K, Raok J, and Jae-ha R (2008) Inhibition of
lipopolysaccharide-induced inducible nitric oxide synthase and cyclooxygenase-2
expression by xanthanolides isolated from Xanthium strumarium. Bioorg Med Chem
Letters 18: 2179-2182.
Yu LL, Han CR, Huang R, Lv YP, Gui SH, and Chen YG (2006) A new iridoid
tetraester from Valeriana jatamansi. Pharmazie 61(5): 486-488.
Yuan CS, Mehendale S, Xiao Y, Aung HH, Xie JT, and Ang-Lee MK (2004) The
gamma-aminobutyric acidergic effects of valerian and valerenic acid on rat brain stem
neuronal activity. Anesth Analg 98(2): 353-358.
Zámocký M and Koller F (1999) Understanding the structure and function of
catalases: clues from molecular evolution and in vitro mutagenesis. Prog Biophys Mol
Biol 72(1): 19-66.