the yunnan biodiversity hotspot project - startseite · miehe: the yunnan biodiversity hotspot...
TRANSCRIPT

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
1
Impressum
Unpublished Symposium Volume
International Symposium
“The Yunnan Biodiversity Hotspot Project – Program, Objectives, and Application”
April 02 – 06 2016, Dresden, Germany
Copyright: Senckenberg Natural History Collections Dresden & Technical University Dresden, Germany
Edited by: Lutz Kunzmann (Senckenberg)
Cover and project logo: Markward Fischer (Senckenberg)
Maps: Susann Stiller (Senckenberg)
Layout: Felix P. Herrmann (TU Dresden)

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
2
Content
Program 3
List of participants 7
Scientific committee / Organizing team 11
Preliminary Project Outline and Drafts of Group Proposals 12
A. Brief Project Outline 12
B. Drafts of Superordinated Project Proposals, Group Proposals and Abstracts of
Subprojects 13
Project summary
0. Synthesis and integration 15
0.1. Additional overview presentations 15
1. The physical basis 16
1.1. Current state of geo- and climate diversity 16
1.2. Uplift history and climate reconstruction 17
2. Evolution of Biodiversity – the fossil record 18
2.1. Standard section/drill core to study vegetation (pollen), climate (incl. glacial
history) using a multiproxy approach (incl. isotopes, biomarkers, etc.) 18
2.2. Neogene biota: paleo-biodiversity, paleoecology, biomarkers, paleoclimate,
stratigraphy, biogeography, altitudinal zonation 21
2.3. The hominoids 30
3. Evolution of Biodiversity – the genomic record 32
3.1. Plant groups 32
3.2. Invertebrate groups 42
3.3. Vertebrate groups 48
4. Present-day diversity distribution in Yunnan – a macroecology and
modelling analysis of present-day patterns 49
5. Future threats 53
Field trips 54
Map with field trip destinations 55
Practical paleontological field work part 1 56
Practical paleontological field work part 2 72
Useful information 81

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
3
Program
Friday & Saturday – Arrival and Welcome
Date Time Program item (early arrivals) Program item (late
arrivals)
April 1
afternoon Arrival at airport Dresden,
transport to hotel, check-in
19:00 – 22:00 Dinner
April 2
8:30 – 13:00 Visit Senckenberg Natural History
Collections Dresden (collections,
labs)
Arrival at airport or main
station Dresden, hotel
check-in
13:00 – 14:00 Lunch
14:00 – 18:00 Transport to hotel, afternoon:
sightseeing and shopping
possibility
Afternoon: sightseeing and
shopping possibility
18:00 – 22:00 Welcome Party in the exhibition building Japanese Palace with
visit to the Senckenberg special exhibition 'Tibet'
Sunday - Symposium I
April 3 Workshop: oral presentations and discussion (day 1)
8:00 – 9:00 Registration, upload presentations
9:00 – 9:20 Welcome talks (Lutz Kunzmann, Volker Mosbrugger, Zhou Zhe-
Kun, Technical University Dresden representative)
9:20 – 9:40 Zhe-Kun Zhou: Evolution and extinction of some plants and
their response to Asia Monsoon climate.
9:40 – 10:00 Sun Hang: xxx
10:00 – 10:30 Volker Mosbrugger/Zhe-Kun Zhou/Lars Opgenoorth/Georg
Miehe: The Yunnan Biodiversity Hotspot Project: state-of-the-art
and application procedure
ongoing 1.2. Uplift history and climate reconstruction, geo- and climate
diversity
- Andreas Mulch

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
4
- Bob and Teresa Spicer / Torsten Utescher
- Cheng-Long Deng
11:00 – 11:20 Tea and coffee break
ongoing 2.1. Standard section/drill core to study vegetation (pollen),
climate (incl. glacial history) using a multiproxy approach (incl.
isotopes, biomarkers, etc.)
- Torsten Utescher
- Wei-Ming Wang
- Martina Stebich
- Christian Rolf
ongoing 2.2. Neogene leaf floras: stratigraphy, taxonomy, biogeography,
climate and altitudinal zonation
- Lutz Kunzmann
- Tao Su
- Torsten Wappler
- Karolin Moraweck
- Ulf Linnemann
ongoing The hominoids
- Christine Hertler
- Tina Lüdecke
ongoing Present-day diversity distribution in Yunnan – a macroecology
and modelling analysis of present-day patterns
- Alice Hughes
- Matthew Forrest
13:00 – 14:00 Group photo and Lunch
14:00 – 15:30 Evolution of Biodiversity / the genomic record
3.1. Plant groups
- Georg Miehe
- Anita Roth-Nebelsick/Wilfried Konrad/Stefan Wanke
- Jian-Quan Liu/Lars Opgenoorth
3.2. Invertebrates
- Joachim Schmidt/Lars Opgenoorth
- Sonja Wedmann
- Martin Brändle
3.3 Vertebrates
- Martin Päckert

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
5
15:30 – 16:00 Tea and coffee break
16:00 – 18:00 "World Café" (round-table discussion):
- Discussion within the project groups (organized by PI)
- Networking between project groups
18:00 – 18:30 Summary first day: Volker Mosbrugger & Zhe-Kun Zhou
18:30 – 19:00 Break (hotel)
19:30 – 22:00 Dinner (Italian restaurant)
Monday - Symposium II
April 4 Workshop: discussion and synthesis
(day 2)
8:30 – 10:00 Presentation of results of “World Café” (PIs) and general
discussion
10:00 – 10:20 Tea and coffee break
10:20 – 12:00 Synthesis and coordination of project groups (Volker
Mosbrugger, Zhe-Kun Zhou, Torsten Utescher, Georg
Miehe)
12:00 – 13:00 Lunch
13:00 – 14:30 Final discussion and outlook (Volker Mosbrugger, Zhe-Kun
Zhou)
14:30 – 15:00 Tea and coffee break
15:00 – 18:00 Visit botanical garden of the Institute of Botany, Technical
University Dresden (guide: Barbara Ditsch, curator)
18:30 – 19:00 Break (hotel)
19:30 – 22:00 Symposiums dinner (Spanish restaurant) with Closing remarks
(Volker Mosbrugger)
International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
6
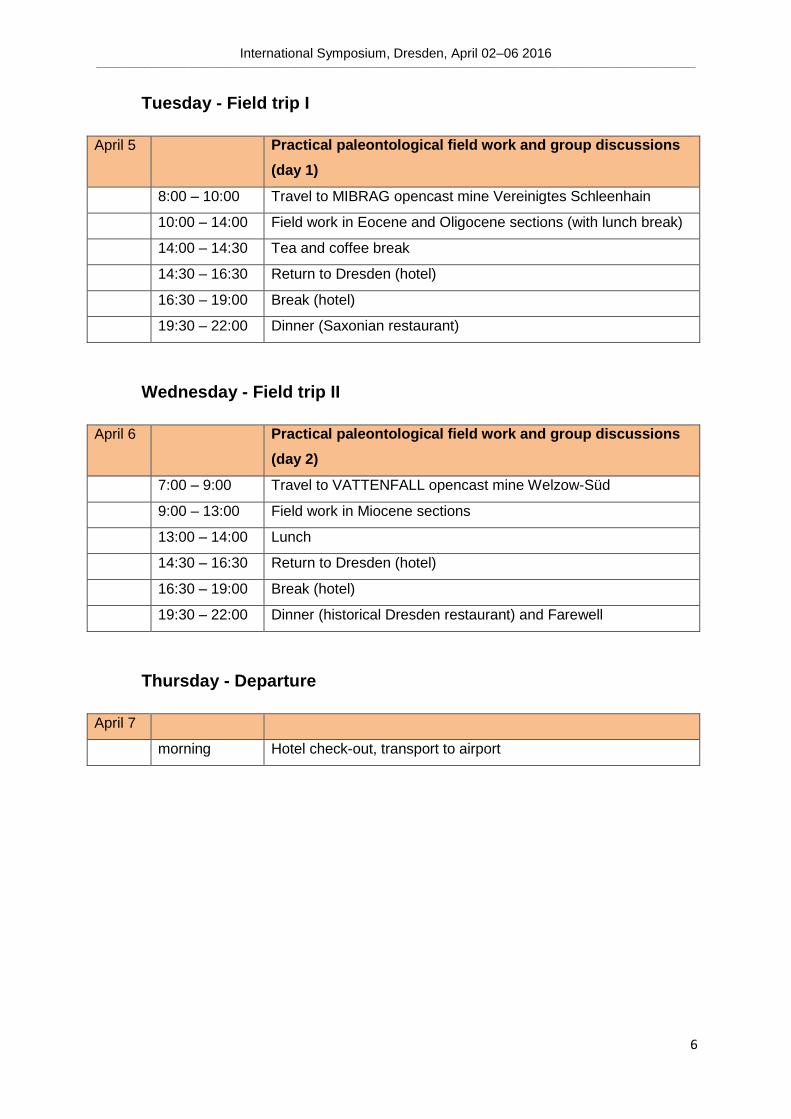
Tuesday - Field trip I
April 5 Practical paleontological field work and group discussions
(day 1)
8:00 – 10:00 Travel to MIBRAG opencast mine Vereinigtes Schleenhain
10:00 – 14:00 Field work in Eocene and Oligocene sections (with lunch break)
14:00 – 14:30 Tea and coffee break
14:30 – 16:30 Return to Dresden (hotel)
16:30 – 19:00 Break (hotel)
19:30 – 22:00 Dinner (Saxonian restaurant)
Wednesday - Field trip II
April 6 Practical paleontological field work and group discussions
(day 2)
7:00 – 9:00 Travel to VATTENFALL opencast mine Welzow-Süd
9:00 – 13:00 Field work in Miocene sections
13:00 – 14:00 Lunch
14:30 – 16:30 Return to Dresden (hotel)
16:30 – 19:00 Break (hotel)
19:30 – 22:00 Dinner (historical Dresden restaurant) and Farewell
Thursday - Departure
April 7
morning Hotel check-out, transport to airport

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
7
List of participants
Martin BRÄNDLE [email protected]
Philipp University of Marburg, Marburg, Germany
Cheng-Long DENG [email protected]
Institute of Geology and Geophysics, CAS, Beijing, PR China
Matthew FORREST [email protected]
Senckenberg, Frankfurt, Germany
Christine HERTLER [email protected]
Heidelberg Academy of Sciences and Humanities, Heidelberg, Germany
Sylvia HOFMANN [email protected]
Natural History Museum Erfurt, Erfurt, Germany
Yong-Jiang HUANG [email protected]
Kunming Institute of Botany, CAS, Kunming, PR China
Alice HUGHES [email protected]
Xishuangbanna Tropical Botanical Garden, Yunnan, PR China
Phyo Kay KHINE
Philipp University of Marburg, Germany
Annette KLUSSMANN-KOLB [email protected]
Senckenberg, Frankfurt, Germany
Wilfried KONRAD [email protected]
Technical University of Dresden
Lutz KUNZMANN [email protected]
Senckenberg, Dresden, Germany
Zhimin LI [email protected]
Kunming Institute of Botany, CAS, Kunming, PR China

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
8
Ulf LINNEMANN [email protected]
Senckenberg, Dresden, Germany
Jian-Quan LIU [email protected]
School of Life Science, Lanzhou University, Lanzhou, PR China
Tina LÜDECKE [email protected]
Senckenberg, Frankfurt, Germany
Georg MIEHE [email protected]
Philipp University of Marburg, Marburg, Germany
Karolin MORAWECK [email protected]
Senckenberg, Dresden, Germany
Volker MOSBRUGGER [email protected]
Senckenberg, Frankfurt, Germany
Andreas MULCH [email protected]
Senckenberg, Frankfurt, Germany
Christoph NEINHUIS [email protected]
Technical University of Dresden, Germany
Lars OPGENOORTH [email protected]
Philipp University of Marburg, Marburg, Germany
Martin PÄCKERT [email protected]
Senckenberg, Dresden, Germany
Christiane RITZ [email protected]
Senckenberg, Görlitz, Germany
Christian ROLF [email protected]
Leipniz Institute for Applied Geophysics, Hannover, Germany

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
9
Anita ROTH-NEBELSICK [email protected]
State Museum of Natural History Stuttgart, Stuttgart, Germany
Joachim SCHMIDT [email protected]
University of Rostock, Rostock, Germany
Teresa SPICER [email protected]
Institute of Botany, CAS, Beijing, PR China
Martina STEBICH [email protected]
Senckenberg, Weimar, Germany
Tao SU [email protected]
Xishuangbanna Tropical Botanical Garden, Yunnan, PR China
Hang SUN [email protected]
Kunming Institute of Botany, CAS, Kunming, PR China
Torsten UTESCHER [email protected]
Senckenberg, Frankfurt; University of Bonn, Germany
Truong Van DO [email protected]
Vietnam National Museum of Nature, Ha Noi, Vietnam
Wei-Ming WANG [email protected]
Institut of Geology and Palaeontology, Nanjing, PR China
Yun WANG [email protected]
Senckenberg, Görlitz, Germany
Stefan WANKE [email protected]
Technical University of Dresden, Germany
Torsten WAPPLER [email protected]
University of Bonn, Bonn, Germany

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
10
Sonja WEDMANN [email protected]
Senckenberg, Frankfurt, Germany
Karsten WESCHE [email protected]
Senckenberg, Görlitz, Germany
Zhe-Kun ZHOU [email protected]
Xishuangbanna Tropical Botanical Garden, Yunnan, PR China

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
11
Scientific committee
Volker Mosbrugger Senckenberg Society for Nature Research, Frankfurt/M., Germany
Georg Miehe Philipp University of Marburg, Marburg, Germany
Torsten Utescher Senckenberg Society for Nature Research Frankfurt/M., / Universität
Bonn, Germany
Lutz Kunzmann Senckenberg Natural History Collections Dresden, Germany
Christoph Neinhuis Technical University Dresden, Germany
Karolin Moraweck Technical University Dresden / Senckenberg Natrual History
Collections Dresden, Germany
Zhe-Kun Zhou Xishuangbanna Tropical Botanical Garden (CAS), Yunnan, PR China
Tao Su Xishuangbanna Tropical Botanical Garden (CAS), Yunnan, PR China
Sun Hang Kunming Institute of Botany (CAS), Yunnan, VR China
Organizing team
Sigrid Beutner Senckenberg, Dresden, Germany
Mina Breuer Technical University of Dresden, Germany
Franziska Ferdani Senckenberg, Dresden, Germany
Markward Fischer Senckenberg, Dresden, Germany
Denise Hennig Senckenberg, Dresden, Germany
Felix P. Herrmann Technical University of Dresden, Germany
Carola Kunzmann Senckenberg, Dresden, Germany
Lutz Kunzmann Senckenberg, Dresden, Germany
Karolin Moraweck Senckenberg, Dresden, Germany
Madeleine Streubig Senckenberg, Dresden, Germany
Susann Stiller Senckenberg, Dresden, Germany
Birgit Walker Senckenberg, Dresden, Germany

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
12
Preliminary Project Outline and Drafts of Group Proposals
How orogenesis drives evolution: Processes, status & trends of geo-biodiversity in
the Himalayan-Yunnan Hotspot.
DFG Trans-Regio: Dresden, Frankfurt, Marburg
A. Brief Project Outline
0 Synthesis and integration
1. The physical basis
1.1 Current state of geo- and climate diversity
1.2 Uplift history and climate reconstruction
2. Evolution of Biodiversity – the fossil record
2.1 Standard section/drill core to study vegetation (pollen), climate (incl. glacial history)
using a multiproxy approach (incl. isotopes, biomarkers, etc.)
2.2 Neogene biota: paleo-biodiversity, paleoecology, biomarkers, paleoclimate,
stratigraphy, biogeography, altitudinal zonation
2.3 The hominoids
3. Evolution of Biodiversity – the genomic record
3.1 Plant groups
3.2 Invertebrate groups
3.3 Vertebrate groups
4. Present-day diversity distribution in Yunnan – a macroecology and modelling analysis
of present-day patterns
5. Future threats

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
13
B. Drafts of Superordinated Project Proposal, Group Proposals and Abstracts of
Subprojects
How orogenesis drives evolution: Processes, status & trends of geo-biodiversity in
the Himalayan-Yunnan Hotspot.
Joerg Bendix, Volker Mosbrugger, Georg Miehe, Lars Opgenoorth
Summary
On a global scale mountains are the most important harbor of biodiversity best exemplified
by the fact that all six major diversity centers surpassing ca. 5000 vascular plant species per
10 000 km² are located in mountain areas(Barthlott et al., 1996). There are two possibilities
for this link – either mountains act as refugia for species from the surrounding area or they
are themselves species pumps. In most cases it will be a combination of both. Nevertheless,
the role of orogenesis as a driver for evolutionary processes has long been accepted in
evolutionary ecology. The drivers behind this connection are manifold and include (i)
topographic complexity which increases geographic and thus genetic barriers, as well as
area, hydrological regime and others, (ii) climatic complexity including microclimatic effects
such as exposition and slope, as well as larger scale effects such as LEE effect and MEE
effect, all of which directly increase the available niche space, (iii) geological complexity by
driving habitat structure e.g. through influencing soil conditions, and erosional processes, (iv)
orogen stability – or more generally speaking – habitat stability. In this proposal we will
summarize all these effects under the term geodiversity. The effects of geodiversity vary in
time – depending on how long orogenesis has been taken place with what specific features –
as well as in space – as the latitudinal position has significant influence on how orographic
complexity acts upon biological processes and thus biodiversity.
Geodiversity is an abstract concept that is not easily measured and even less easily
translated into biotic processes because so many different drivers are superimposed over
each other. Furthermore, the effect each of these drivers have on different biological units is
diverse as well. For example, while relatively small elevational differences can be significant
barriers for wingless ground beetles directly driving evolutionary processes and eventually
speciation, flying beetles, birds or mammals will not be affected by them. Thus topographic
complexity will have much different effects and effect-thresholds depending on the life history
traits of a given taxon. One of the overarching goals of this research proposal therefore is to
disentangle important parts of these complex mechanisms and ultimately to derive measures
for geodiversity that will allow to model ‘across taxa biodiversity’ in mountain systems of
the world and moreover, determine how biodiversity as well as evolutionary processes can
be preserved in the future. More specifically, we want to

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
14
Derive high resolution descriptors of current geodiversity through remote sensing and
spatial modelling
Derive indices for past orographic complexity by analysing the biologically relevant
history of the orogenesis of the Himalaya-Tibet Orogen
Disentangle the specific drivers responsible for the creation of the Himalaya-Yunnan
Hotspot specifically comparing diverse evolutionary and spatial histories of diverse taxon
groups
Date when the Himalaya-Yunnan Hotspot first appeared in Earth History
Characterize the extant large scale biodiversity patterns by means of macroecology
and compare them to the historical processes.
Assess future threats to diverse groups of biodiversity and ecosystems in the
Himalaya-Yunnan Hotspot

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
15
0. Synthesis and integration
Developing a Memorandum of Understanding / Cooperation Agreement between German
institutions, and XTBG (CAS), KIB (CAS) of PR China (Volker Mosbrugger, Zhe-Kun Zhou,
Sun Hang, Georg Miehe)
Developing a common database.
0.1. Additional overview presentations
Evolution and extinction of some plants and their response to Asia Monsoon climate.
Zhe-Kun Zhou, Xishuangbanna Tropical Botanical garden, Chinese Academy of Sciences,
Yunnan, PR China
Seasonal precipitation is the main characteristic of the Eastern Asian monsoon: almost 80
percent of the annual precipitation is concentrated during the rainy season, usually from May
to October. The Asian monsoon region experiences wet summers and very dry winters and
springs. The Asian monsoon system was established during the Late Miocene in association
with the uplift of the Qinghai-Xizang Plateau. The following scientific questions are
addressed: how does the Asian monsoon impact on biodiversity and how do the plants
respond to the dry winter and spring. We propose the following hypothesis: Due to the
seasonality of the monsoon climate, some physiological processes of plants such as
germination and growth of seedlings would suffer from dryness in winter and spring, causing
their distribution area to shrink or even leading to their extinction. In order to answer these
questions, five research goals are designed in the current project: First, to reconstruct the
paleoclimate and intensity of the monsoon using stable isotopes from sediments of middle
Miocene to Pleistocene age. Concurrently the paleoclimate will be reconstructed using plant
fossil fruits and seeds. These independent approaches will be used to cross-check one
another. Second, the fossil histories of Metasequoia and other relict plants will be studied
and their paleoclimatic signal reconstructed. Third, research into the differentiation of Pinus
yunnanensis and Pinus kesiya var. langbianensis and their response to the monsoon climate
will be carried out. Molecular analysis, wood anatomy, stable isotopes of wood and leaves
will be involved in this mission. Fourth, seed biology and experiments of controlled cultivation
of Cedrus, Metasequoia, oaks with wider distributions and limited distributions will be
undertaken. Fifth, fossil history and phenology of Quercus subgenus Cyclobalanopsis will be
carried out in order to test if extinction of some oaks such as Quercus sichouensis is caused
by the dry winter.

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
16
1. The physical basis
1.1. Current state of geo- and climate diversity
High resolution digital elevation model“
Meteorological data of Yunnan (incl. data from MOST Project 973: Three River Gorges; and
data from China Meteorological Forcing Dataset)
Volker Mosbrugger and Joerg Bendix will develop a concept
Zhou Zhekun will find out, which data are publicly available

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
17
1.2. Uplift history and climate reconstruction
Cenozoic climate and uplift in Yunnan – evidence from the palaeobotanical record
Torsten Utescher1, Volker Mosbrugger1, Su Tao2, Zhou Zhe-Kun2
1Senckenberg Research Institute, Senckenberganlage 25, 60325 Frankfurt am Main,
Germany
2Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences, Menglun,
Mengla, Yunnan 666303, China
Climate data obtained from a total of 29 Yunnan palaeofloras (macro-remains, pollen) by
using the Coexistence Approach (29 floras, new calculations) and CLAMP (8 floras, literature
data) provide a first insight into Cenozoic temperatures in general, inferred altitudinal
changes and time resolution of the macro-records. Temperature data compiled for the
southern part of Yunnan reflect the large scale global trend of the late Neogene Cooling. The
record obtained for northern Yunnan can hardly be interpreted, most probably due to
differential uplift processes in that area. When relating Cenozoic temperatures inferred from
sites presently located at low and mid- altitudes to modern values it is shown that this
continental part was not significantly warmer compared to today, partly it might have been
even cooler. For most sites, presently located at higher altitudes pointed positive temperature
anomalies with respect to present are obtained. Thus, it has to be assumed that various
parts of Yunnan were subject to more recent uplift.
As is shown by first preliminary results the palaeobotanical record of Yunnan can contribute
substantial data on the uplift history of Yunnan while the high degree of overlapping of CA
and CLAMP-derive data testifies the coherence of the data obtained. However, there still
exist large gaps in the macrofloral record so far available comprising most of the
Palaeogene, the Middle Miocene, and the Early Pliocene. In order to study regional climate
dynamics in the context of landscape evolution at higher resolution suitable pollen records
are required and other proxies have to be included.

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
18
2. Evolution of Biodiversity – the fossil record
2.1. Standard section/drill core to study vegetation (pollen), climate (incl. glacial
history) using a multiproxy approach (incl. isotopes, biomarkers, etc.)
Perspectives on Late Cenozoic vegetation and climate study in Yunnan based on
standard sections and cores
Wei-Ming Wang
Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, Nanjing
210008, P. R. China, [email protected]
The Yunnan-Guizhou Plateau, located adjacent to the southeast of the Tibetan Plateau, has
a complicated topography, diverse climate conditions, and high plant diversity. Its geological
history has been strongly affected by the uplift of the Tibetan Plateau in the Cenozoic. Late
Cenozoic is a crucial period in the development of the local vegetation in Yunnan.
During the past decades, many studies focused on the the late Miocene deposits, such as
Xiaolongtan, Lincang and Xianfeng floras. There were severe environmental changes
occurred both on the lands and in the oceans at that time, for example, the development and
spread of C4 grasses, the aridification of the interiors of continents, the expansion of open
landscapes, and the development and evolution of the East Asian monsoon.
The high-resolution pollen study of the lake sediment in Heqing Basin is the only long core
which displays the vegetation history of the mountains around the basin since 2.78 Ma B.P. It
shows that the increase or decrease of vertical vegetational belts and the changes in the
components of vegetational belts are controlled by the tectonic uplift of mountains and the
climatic changes. Core from Lake Erhai also exhibits the local vegetaion and climate
changes since the latest Pleistocene.
Yunnan was once suggested to be one of the most possible areas for the origin of rice
agriculture. During the past decade, many Palaeolithic sites were excavated, which gives us
chances to reconstruct the vegetation and climate background for the early human beings.
Case studies were carried out on several sites. Among them, the site at Xiangbidong is the
first Palaeolithic cave relic, which is located in the Hengduan Mountains with complex
landforms and diverse natural landscapes. Pollen study reveals vegetation and environment
changes in relation with human activies.
It is anticipated that we will have a much more complete knowlege on the Late Cenozoic
vegetation and climate history in Yunnan, if we get further improved dating for these
Neogene floras, and some continued long profiles for comprehensive study with multi-
approaches.

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
19
Improving chronostratigraphic frame by palaeomagnetic and rock magnetic
investigations
Christian Rolf, Leipniz Institute for Applied Geophysics, Hannover, Germany
Ulrich Hambach, University of Bayreuth, Germany
The potential of rock and palaeomagnetism in stratigraphic and chronometric investigations
was intensely discussed during the previous meetings in Xishuangbanna (2013) and
Kunming (2015) where we have reported jointly with our Chinese colleagues about the broad
application possibilities of rock and palaeomagnetic investigations. All sub-projects with
stratigraphic approaches in the biodiversity hotspot program need a chronostratigraphic
frame of the pre-hotspot world in Yunnan. The palaeomagnetic methods should be able to
serve this request, as our Chinese colleagues have already shown in excellent publications
concerning magnetic polarity stratigraphy (MPS) of the late Neogene in Yunnan.
Not only the changing polarity of the Earth’s Magnetic Field (EMF) but also its variation in
intensity is recorded in rocks. In lithologically homogenous sediments the so called relative
palaeointensity (RPI) is recorded. Correlating these RPI data with the quite well-known
variation of the EMF during the Upper Neogene provides an independent tool for
stratigraphic dating with a temporal resolution in the order of ten thousand to thousands of
years. Great variability in lithology and magnetic mineralogy are limiting factors. Suitable
outcrops or drill cores with homogenous lithology are mandatory.
Time series analyses of down-section/down-hole rock-magnetic proxies -like magnetic
susceptibility-can provide a temporal frame in much higher resolution than magnetic polarity
stratigraphy (e.g. Milanković cycles). Cyclostratigraphy alone provides no absolute data, but
relative time estimates and need anchor points. This approach may be applied on large
outcrop sections or drill cores with wider time windows (> 100 ka).
In addition the changing properties of magnetic minerals give information about the temporal
and spatial environmental dynamics (climate, provenance, depositional systems and
diagenesis, etc.). Rock magnetic investigations are also mandatory to back the quality of
MPS and RPI recording.
Furthermore, we think it could be worth to investigate rock magnetic parameters as proxy
indicators for volcanic crypto-tephras. They will be excellent time markers, if recorded, and if
a tephra chronology of the area under investigation can be established. Following the
statements of Chenglong up to date tephra layers were found only in few sedimentary basins
of late Cenozoic age in Yunnan. Suggested volcanic activities of late Cenozoic age in
western Yunnan, however, makes the search of volcanic tephra layers in Yunnan basins
highly promising.

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
20
Cheng-Long Deng proposed acooperation on magnetostratigraphy (including rock
magnetism as proxies for quality of the magnetostratigraphic results) secular variation,
“palaeointensities” and the search for tephra layers. In addition we would like to focus on
younger (Plio-/Pleistocene) stratigraphic intervals in outcrops or drill cores in order to explore
the record of the Plio-/Pleistocene environmental change.
In case drilling projects will be realised we can offer the LIAG downhole logging equipment to
get geophysical borehole data in high resolution (for examples magnetic susceptibility,
natural gamma ray etc.). These data can be analysed by time series/cyclostratigraphic
methods to improve the age depth models of the sediments recovered.
Our experience in applying for funding from the DFG for new drilling projects tells us, that it is
advisable to have good quality data in advance. In case already existing and accessible
boreholes are available, our team would come on own funds to China to acquire data and
test their quality.

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
21
2.2. Neogene biota: paleo-biodiversity, paleoecology, biomarkers, paleoclimate,
stratigraphy, biogeography, altitudinal zonation
Deciphering a “pre-hotspot” world: the palaeoenvironmental perspective on Neogene
biota
Key words: palaeobiodiversity, palaeoherbivory, molecular biomarkers, palaeoecology,
palaeoclimate fluctuations, palaeoatmospheric composition
Group members:
Chinese side: Su Tao, Huang Yong-Jiang, Xie Sanping, Shi Gongle , Deng Cheng-Long
German side: Lutz Kunzmann, Karolin Moraweck, Torsten Wappler, Eva Niedermeyer, Ulf
Linnemann, Anita Roth-Nebelsick
Outline:
- Project group proposal: key questions and summary
- Project 1: Evolution of ecophysiological “fingerprints” of Neogene vegetation in
Yunnan (China) under multicausal influence of monsoon, mountain uplift, climate
change, atmospheric composition and forming topographies. (Lutz Kunzmann, Su
Tao, Karolin Moraweck, Gongle Shi, Yong-Jiang Huang, Eva Niedermeyer, Anita
Roth-Nebelsick)
- Project 2: Structural changes of herbivory since the Neogene in response to climatic
fluctuations from Yunnan (China). (Torsten Wappler, Su Tao)
- Project 3: Neogene climate change in Yunnan (China) inferred from molecular
biomarkers. (Eva Niedermeyer/Karolin Moraweck/Lutz Kunzmann/???)
- Project 4: “Dating/stratigraphy”. (Ulf Linnemann/Cheng-Long Deng/ …)
Project group proposal: key questions and summary
The present biodiversity in Yunnan’s mountain regions and its proximities in other Chinese
provinces as well as in neighboring countries is discussed to be the result of a multicausal
evolution. Main drivers certainly were mountain uplift, global climate change, establishment
of a monsoonal regime and constitution of the landscape after/during the Pleistocene by
glacigenous processes. The first key question concerns the crucial driver of the
establishment of the Yunnan Biodiversity Hotspot.
Several results of biodiversity studies in extant plant and animal groups tentatively give rise
to the hypothesis that the biodiversity hotspot isn’t that old as one could perhaps conclude on
the age of the appearance of the monsoonal regime, i.e. at least early Miocene or late
Eocene. In fact, by studying Neogene biota and environments the “pre-hotspot” scenario

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
22
could be deciphered as well as the underlying circumstances for the evolution of a
biodiversity hotspot.
Therefore, another key question of the project group is to test whether pre-Pleistocene floras
and herbivory data reveal any information of the rising biodiversity hotspot. With respect to
the megafloras several (late) Miocene and Pliocene floras will be involved in the studies.
Based on data on their systematic diversity and data of plant-arthropod interactions their
palaeoenvironments, palaeoclimatic conditions and palaeoatmospheric background will be
reconstructed. These reconstruction will be condacted based on published data as well as by
new studies. Molecular biomarkers are an essential tool for reconstructing ancient climate
from horizons where fossil biota are absent. The coupling of these results, i.e. proxy data
derived from several floras from two distinct time intervals, data from plant-arthropod
interactions and from fossil molecular biomarkers, is a novelty and allows the
consideration of the different factors in a multiproxy analysis.
If the hypothetical scenario is true, the project bundle can presumably explain the “pre-
hotspot world” of that region as the starting point and place of the evolution of a hotspot. In
particular we want to solve these questions:
(1) Why was this region predestined for the evolution of a hotspot?
(2) Which biotic and abiotic factors were essential for the “rise” of the hotspot?
(3) What can we learn from fossil floras and fossil herbivory about the hotspot’s starting
point and early evolution?
(4) Which climatic conditions and/or fluctuations and paleo-atmospheric conditions
favored the evolution of the hotspot?
Selected Neogene floras of Yunnan in the project
Tao Su, Xishuangbanna Tropical Botanical Garden, CAS, Yunnan, PR China
The tectonic activities in southeastern margin of the Qinghai-Tibetan Plateau created more
than 200 Neogene basins in Yunnan Province. Among them, plenty of well preserved fossil
floras have been uncovered especially during the last decade, which provide good
opportunity to study the biodiversity history under paloenvironmental changes in Yunnan.
Previous studies with focus on Neogene floras indicated that, the monsoon intensified
gradually since the Neogene in Yunnan and had profound influence on shaping modern
biodiversity. However, our understanding about the diversification pattern is far from being
enough comparing to the abundant fossil floras as well as extant high diversity here. In this
project, we will select several megafloras ranging from the early Miocene to the late Pliocene
as materials for study, i.e., the early Miocene Jinggu flora (S Yunnan), the middle Miocene
Wenshan flora (SE Yunnan) and Zhenyuan flora (SW Yunnan), the late Miocene Xiaolongtan

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
23
flora (SE Yunnan) and Lvhe flora (C Yunnan), the Pliocene Heqing flora (NW Yunnan) and
Weixi flora (NW Yunnan). According to the preliminary investigation, all these floras are not
only in excellent preservation condition, but also rich in plant diversity preserved as leaves,
fruits/seeds, flowers, and wood. Fossils from these floras will be collected, and their
systematic positions will be determined with morphological comparison to modern plants.
Moreover, new fossil sites in Yunnan will be explored according to this project.
Project interactions:
Coupling of morphometric, anatomical and ecophysiological plant traits: Climatic and
environmental driven changes in biodiversity patterns are often preserved in fossil floras.
Studying the composition of these plant assemblages as well as focusing on single taxa and
their adaptions in anatomy and morphology to extant climatic and environmental conditions
will help to understand the development and distribution of this hotspot region. The
morphometric reconstruction of fragmented fossil leaves delivers quantitative data about leaf
area, length, width and related data, which enable to conclude environmental data, like water
availability or temperature and heat conditions on the leaves surface. Changing climatic and
environmental conditions might have been drivers for changing compositions of plant
assemblages and perhaps led to specification of single genuses. Alongside the consideration
of morphometric traits, also anatomical adaptions can be evident for ecological and climatic
changes, as trichome density (TD) and stomata density (SD) are coupled with water use
efficiency and palaeoatmospheric conditions, which can be reconstructed using the gas
exchange model. The procedure for collecting and evaluating plant–insect associational data
is derived from an explicit classification of insect-mediated damage used by the PIs and
colleagues. For the Yunnan data, we will examine on slabs each identifiable plant fragment
greater than ca. 0.5 cm2, followed by a database record of unique specimen number, plant
identification, DTs (if any), image or line drawings of the damage (if any), repository location
and comments for each specimen. This spectrum of damage types (DTs) is based on life-
plant response to insect feeding using four diagnostic criteria. The presence or absence and
type of DT data are tabulated for each leaf per locality, together with host-plant identification,
habitat type and stratigraphic position, for the Neogene floras in Yunnan. The data will be
expressed as stratigraphic plots listing DTs, the percent of leaves damaged (frequency data),
the number of DTs per leaf (diversity data), and will be partitioned by plant-host and
organ/tissue specificity using several lines of evidence. Attention will be directed to the
temporal and spatial damage-type composition of the dominant plant hosts to assess the
evolution of herbivore partitioning of tissues.
Connecting factors to other project groups:

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
24
Observed variations in climate conditions probably represent intrinsic change and not shifts
resulting from tectonic uplift of the area. We expect that our findings will have major effects
on understanding interorganismic palaeoecological relationships and increase understanding
of the determinative role that past environmental changes have on those relationships, and
analogous, modern, ecological changes. The project is connected to the groups 3 and 7.
The knowledge of plants taxonomy and their composition in these fossil plant assemblages is
connected to group 5, analysing the diversity evolution and present day distribution. As
mountain uplift is connected with climate changes, the plant derived palaeoatmospheric and
palaeoclimate data can be coupled and verified using the uplift and climate data obtained by
group 3, using isotopes and other proxies.

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
25
Project 1: Evolution of ecophysiological “fingerprints” of Neogene vegetation in
Yunnan (China) under multicausal influence of monsoon, mountain uplift, climate
change, atmospheric composition and forming topographies
Among Neogene biota in Yunnan (China) plant megafossil assemblages occur widespread in
intracontinental sedimentary basins. Most of these floras are rather divers both in taxa and
plant parts / organs. They are therefore predestined for reconstruction of terrestrial
ecosystems through time. Main factors which influenced the vegetation are climate change
including monsoon regimes, mountain uplift, plate tectonics and constitution of the
landscape.
The project will identify an amount of morphological, morphometric, anatomical and
phytosociological data from the fossil leaf assemblages due to reveal an ecophysiological
“fingerprint” of a flora. These “fingerprints” show taxon-independent signals of environmental
and atmospheric changes through time. Two stratigraphic levels are chosen for the study,
late Miocene and Pliocene, because from here well-preserved megafloras containing a
diverse leaf component are known.
The fossil leaf remains themselves reveal climatic and atmospheric signals which could be
quantified by using several approaches. Climatic signals are calculated by CLAMP and CA,
whereas atmospheric signals, i.e. pCO2, are calculated using the mechanistic gas exchange
model. From this a scenario for changing climatic and atmospheric conditions under the
influence of mountain uplift and plate tectonics will be developed. The most important
questions are: (1) whether an increase in biodiversity detectable from the floras is visible or
not; (2) could an increase in diversity possibly lead to the current hotspot; (3) which were the
main drivers of increasing diversity; (4) when did main increases in diversity occur and if
vegetation changes did not necessarily coincide with an increase in diversity of plants (5)
what factors prevent the evolution of a hotspot.
Presumably an integrated analyses of Neogene floras lead to the conclusion that that the
main biodiversity rise appeared later than Pliocene. Thus, the evolution of the Neogene
vegetation gives evidence for a pre-hotspot diversity.

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
26
Project 2: Structural changes of herbivory since the Neogene in response to climatic
fluctuations from Yunnan (China)
Vascular plants together with insects contribute substantially to the Earth's rich biodiversity.
Their interactions constitute a complex and intricate trophic network in terrestrial ecosystems;
therefore, the patterns of plant-arthropod interactions in geological times are pivotal to
understand the evolution of terrestrial ecosystems. The origin of these trophic networks can
be traced back for more than 400 million years ago (Misof et al., 2014). However, research
on changes in the food web structures in ecological systems on the basis of
paleaeontological data is only at its beginnings and underlines our need to understand the
impacts of climate change on these systems. Still some progress has been reached in the
last decades and by now, the fossil compressions of dicotyledonous plant leaves represent
our main source of evidence of changes of these interactions (e.g., Labandeira, 2013). Most
analyses and results presented in previous studies have shown that the relationships are
more complex than generally anticipated (e.g., Wappler et al., 2012; Su et al., 2015), and
that most observed processes act in network structures rather than in isolation (Wappler et
al., 2015).
The study of plant-arthropod interactions faces many fundamental challenges. The
plants have to be identified from often-fragmentary material, and the arthropods interacting
with plants have to be inferred even only from the traces they had left on leaves before they
fossilized. The records of fossil insects themselves are still relatively scarce (Wappler, pers.
observ. 2015). On the contrary, fossilized leaves of Neogene age are relatively common at
many sites (e.g., Su et al., 2013; Zhang et al., 2015) and frequently large proportions of
leaves bear well-preserved traces of arthropod feeding (Wappler, pers. observ. 2015).
The work employing the plant-arthropod interactions for better reconstruction of
palaeoenvironments and for study of reactions of palaeoecosystems to climatic fluctuations
poses more constraints on our choice of our study material (e.g., sites – palaeoecosystems).
The quantitative reconstructions require large collections of fossil leaves (and also sufficient
numbers of those with arthropod damage) and the compared palaeoecosystems have to
differ only in as few characteristics as possible. This means that they should be close to each
other geographically and host as similar taxa as possible, so that the differences in plant-
arthropod associations can be attributed to the differences in climate and not to other
palaeogeographical or palaeoecological conditions. The less we can comply with these
conditions the more drastic changes in climate we have to focus on in order to attain at least
some certainty about ascribing the change in palaeoecosystem biodiversity and structure to
the effect of climate. Finally, it is also important from the viewpoint of comparison to current
ecosystems to find a geological era with climate fluctuations sufficiently similar to our current
ecosystems.

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
27
The Neogene seems to be very perspective from the point of the above-mentioned
conditions for study of impacts of climatic fluctuations on past and recent ecosystems. For
example, Miocene climate fluctuations were found similar in form to later Plio-Pleistocene
fluctuations resulting in cycles known as the Ice Ages. Generally speaking, the main triggers
to Miocene climate fluctuations are thought to be the emergence of the Antarctic ice cap
following the formation of Circum-Antarctic current and concurrent decline of
circumequatorial circulation due to closing of Tethys Ocean. Regionally, more important
agents were the large continental uplift due to collision of Afro-Arabian and Indian plates in to
Eurasia (Sun et al., 2015), coupled with the evolution of the Asian monsoon system (Su et
al., 2013). Overall, this was also coupled by long-term fluctuations in CO2 concentration (e.g.,
Kürschner et al., 2009). The use of Neogene plant-arthropod interactions harbours one more
important advantage – patterns of DT diversity and agent host specificity are already very
similar to recent conditions as evidenced by insect damage on an oak group (Su et al.,
2015). Due to all these reasons, Neogene represents an extremely interesting and important
period in relation to study of the way the ecosystems cope with climate fluctuations.
Despite this suitability, the studies that focus on this period are still in its very
beginnings (e.g., Prokop et al., 2010; Wappler 2010; Knor et al., 2013, 2015). Some of the
world’s best-developed deposits of Neogene sediments are those of Yunnan Province,
located at the southeastern boundary of the Qinghai-Tibetan Plateau. Until now, more than
twenty Neogene megafloras have been reported from different localities of Yunnan (Li, 1995;
Ge and Li, 1999). More importantly, the intensification of the monsoonal climate caused by
the uplift of the QTP became more prominent since the Miocene, characterizing by the drier
winter and wetter summer. Early studies indicate that this climate trend has largely
influenced the plant diversity in Yunnan (Su et al., 2013; Zhang et al., 2015). However, the
plant-insect interactions in terrestrial systems are still largely unknown there.
The study of the plant-insect associations from Neogene sediments located in the
Yunnan Province gives an opportunity to confirm some important predictions with regard to
climatic fluctuations in the light of the elevation and extent of the Himalaya–Tibetan plateau
(HTE). Together with modern samples from this biodiversity hotspot, the project aims to
understand: (1) the evolutionary pattern of insect damage on specific plant groups, e.g.,
Fagaceae, and Betulaceae; (2) the plant-insect interactions among paleofloras in respose to
paleoenvironmental changes there, such as the intensification of the monsoonal climate; and
(3) the mechanism of plant-insect interactions in structuring terrestrial ecosystems under
dramatically Neogene paleoenvironmental changes.
References:
Currano, E.D., Labandeira, C. and Wilf, P., 2010. Fossil insect folivory tracks paleotemperature for six million
years. Ecological Monographs, 80, 547-567.

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
28
Ge, H.-R., and Li, D.-Y., 1999. Cenozoic Coal-Bearing Basins and Coal-Forming Regularity in West Yunnan.
Yunnan Science and Technology Press, Kunming.
Knor, S., Kvaček, Z., Wappler, T. and Prokop, J., 2015. Diversity, taphonomy and palaeoecology of plant-
arthropod interactions in the lower Miocene (Burdigalian) in the Most Basin in north-western Bohemia
(Czech Republic). Review of Palaeobotany and Palynology, 219, 52-70.
Knor, S., Skuhravá, M., Wappler, T. and Prokop, J., 2013. Galls and gall makers on plant leaves from the lower
Miocene (Burdigalian) of the Czech Republic: systematic and palaeoecological implications. Review of
Palaeobotany and Palynology, 188, 38-51.
Kürschner, W.M., Kvacek, Z. and Dilcher, D.L., 2008. The impact of Miocene atmospheric carbon dioxide
fluctuations on climate and the evolution of terrestrial ecosystems. Proceedings of the National Academy of
Sciences, 105(2), 449-453.
Labandeira, C.C., 2013. A paleobiologic perspective on plant-insect interactions. Current Opinion in plant biology,
16(4), 414-421.
Li, X.-X., ed., 1995. Fossil Floras of China through the Geological Ages. Guangdong Science and Technology
Press, Guangzhou.
Misof B, Liu S, Meusemann K, Peters RS, Donath A, Mayer C, et al., 2014. Phylogenomics resolves the timing
and pattern of insect evolution. Science, 346(6210), 763-767.
Prokop, J., Wappler, T., Knor, S. and Kvaček, Z., 2010. Plant-arthropod associations from the Lower Miocene of
the Most Basin in northern Bohemia (Czech Republic): A preliminary report. Acta Geologica Sinica, 84(4),
903-914.
Su T, Adams JM, Wappler T, Huang Y-J, Jacques FMB, Liu Y-S, et al., 2015. Resilience of plant-insect
interactions in an oak lineage through Quaternary climate change. Paleobiology, 41(1), 174-186.
Su, T., Jacques, F.M.B., Ma, H.-J. and Zhou, Z.-K., 2013. Fossil fruits of Ailanthus confucii from the Upper
Miocene of Wenshan, Yunnan Province, southwestern China. Palaeoworld, 22(3–4), 153-158.
Su, T., Jacques, F.M.B., Spicer, R.A., Liu, Y.-S., Huang, Y.-J., Xing, Y.-W., and Zhou, Z.-K., 2013. Post-Pliocene
establishment of the present monsoonal climate in SW China: evidence from the late Pliocene Longmen
megaflora. Climate of the Past, 9(4): 1911-1920.
Sun, B. et al., 2015. Early Miocene elevation in northern Tibet estimated by palaeobotanical evidence. Scientific
Reports, 5, 10379.
Wappler, T., Labandeira, C.C., Engel, M.S., Zetter, R. and Grimsson, F., 2015. Specialized and Generalized
Pollen-Collection Strategies in an Ancient Bee Lineage. Current Biology, 25(23), 3092-3098.
Wappler, T., Labandeira, C.C., Rust, J., Frankenhäuser, H. and Wilde, V., 2012. Testing for the Effects and
Consequences of Mid Paleogene Climate Change on Insect Herbivory. PLoS ONE, 7(7), e40744.
Zhang, J.-W. et al., 2015. Sequoia maguanensis, a new Miocene relative of the coast redwood, Sequoia
sempervirens, from China: Implications for paleogeography and paleoclimate. American Journal of Botany,
102(1), 103-118.

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
29
Project 3: Neogene climate change in Yunnan (China) inferred from molecular
biomarkers
Project 3 will address climate reconstruction in Yunnan (China) during the Neogene using a
set of molecular biomarkers. On the base of Project group 5 (dating & stratigraphy) and
complementing Project group 3, the proposed study will focus on reconstructing the two most
important climate variables, i.e. temperature and rainfall. Whereas macrofossils may be
restricted to distinct stratigraphic sections, biomarkers are present throughout a stratigraphic
sequence, allowing for a high-resolution (i.e. cm-scale sampling) reconstruction of ancient
climates. They are, however, less specific than macrofossils with respect to their organismal
source. Therefore, the proposed study will be conducted in close cooperation with Project 1.
Paleo-temperatures will be reconstructed using microbial membrane lipids, so called GDGTs
(Glycerol-Diphenyl-Glycerol-Tetraethers), produced by produced by anaerobic soil bacteria.
Comparison with CA and CLAMP based temperature estimates (Project 1) will lead to a
robust paleo-temperature reconstruction.
Information on past rainfall changes will be inferred form the analysis of the stable hydrogen
and carbon isotopic composition of sedimentary higher plant waxes (δDwax and δ13Cwax,
respectively). Together with the above mentioned temperature reconstruction and the result
of macrofossil-based environmental and floral reconstruction conducted in Project 1, we will
use δDwax to derive an estimate of total annual mean rainfall as well as fluctuations of
northwest vs southeast monsoonal flow. δ13Cwax, in turn, may contain valuable information on
changes in canopy closure/vegetation density throughout the study interval. The proposed
study aims furthermore at comparing leaf wax isotopes from individual fossil cuticles with the
integrated leaf wax signal in the enclosing sediment to capture “snapshots” of monsoonal
activity at the time of individual leaf formation vs. long-term changes in monsoonal flow.
In summary, our paleoclimatic data will be interpreted together with paleo-diversity
reconstructions of Project 1 and paleo-altimetry and –topographical changes inferred from
Project Group 3 to understand the interaction of topography and climate on creating the
biodiversity hotspot in Yunnan.
Project 4: “Dating/stratigraphy/provenance analysis” (Ulf Linnemann / Cheng-Long
Deng?)

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
30
2.3. The hominoids
Ecology and Diversity of Primate Communities in the Neogene of Yunnan Province
Christine Hertler1, Tina Lüdecke2, Ji Xueping3 and Andreas Mulch2
1 Research Center “The role of culture in early expansions of humans (ROCEEH)”,
Senckenberg Research Institute, Frankfurt am Main, Germany.
2 Senckenberg Biodiversity and Climate Research Centre, Frankfurt am Main, Germany
3 Yunnan Cultural Relics and Archaeology Institute, Kunming, Yunnan, China.
The evolution of Neogene mammal communities in the Yunnan province is well documented
by a sequence of faunas from several Miocene to Plio-Pleistocene localities. These large
mammal communities display an unusual large taxonomic diversity, which is apparently
related to changes in the geological and climatological setting. The assemblages represent
major episodes of faunal change and therefore provide a rich database for the study of
faunal responses in relation to changes in their environments.
The Neogene records both, the uplift of the Tibetian plateau and the establishment of the
modern configuration of the East Asian Monsoon System, which played a significant role in
environmental change. Previous paleoecological studies however, found a remarkable
homogeneity in the dietary spectrum of herbivores indicating the persistence of tropical
rainforest. E.g., Biasatti et al. (2012) focused on herbivore communities and noted an
increase in the diversity of the dietary signals only after 3.5 Ma. Surprisingly, primates were
not included, although the primate communities in this region represent an important source
of information particularly for forest habitats.
South China plays an important role in our present understanding of primate diversity. Some
of the localities in the Yunnan province represent type localities for enigmatic taxa like the
hominoids Lufengpithecus and Gigantopithecus. Although the taxonomy of the hominoids in
the Neogene of South China has been studied by several authors (among others Harrison et
al. 2002 and Qi Guoqin et al. 2006) and the relationship of taxonomic diversity and
environmental change is undisputed (Jablonski 2005), the ecology of the primate
communities in this region has not yet been studied in greater detail.
We will study the ecological diversity and variability of the Yunnan province primates and
associated herbivore guilds to gain insights in differential faunal responses to environmental
change. The collection of ecological diversity indices and ecomorphological data will be
complemented by analyzing stable carbon and oxygen isotopes of their tooth enamel, which
record nutrition (including food and drinking water) and foraging strategies of the individual

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
31
mammals, which in turn reflects environmental parameters such as vegetation, water supply
or seasonality.
References:
Biasatti, D, Wang Yang, Gao Feng, Xu Yingfeng, Flynn, Lawrence, 2012. Paleoecologies and palaeoclimates of
late Cenozoic mammals from Southwest China: Evidence from stable carbon and oxygen isotopes, Journal of
Asian Earth Sciences 44, 48-61.
Harrison, T., Ji Xueping, Su, Denise, 2001. On the systematic status of the late Neogene hominoids from Yunnan
Province, China. Journal of Human Evolution 43, 207-227.
Jablonski, N.G., 2005. Primate homeland: forests and the evolution of primates during the Tertiary and
Quaternary in Asia. Anthropological Science 113, 117-122.
Qi Guoqin, Dong Wei, Zheng Liang, Zhao Lingxia, Gao Feng, Yue Leping, Zhang Yunxiang, 2006. Taxonomy,
age and environment status of the Yuanmou hominoids. Chinese Science Bulletin 51(6), 704-712.

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
32
3. Evolution of Biodiversity – the genomic record
3.1. Plant groups
What are the specific plant diversity patterns of the SE Himalaya – Yunnan Hotspot?
Georg Miehe
The ‘Eastern Himalaya-Yunnan Centre’ (Barthlott et al.1996) is
(1) the only biodiversity hotspot outside the Tropical Realms, in the Holarctic Floristic Realm
of the northern hemisphere. In contrast to the other hotspots it has
(2) a uniquely high geo-diversity with (i) largest vertical extent, (ii) unique mountain ranges in
parallel with deep gorges (UNESCO World Heritage Site) and a (iii) unique archipelago of
sky mountains in the southeastern periphery of the world’s largest highlands.
The region host global maxima with respect to
(3) a highly diverse humidity pattern ranging between 296 mm/a (Dry River Gorges:
Benzilan) and probably exceeding 6000 mm/a (Hponyin Razi, NW Myanmar),
(4) the tremendous floristic contrasts of the highest and lowest biodiversity (Hotspot of the
southern slopes of the Southeastern Himalaya and the Coldspot (Dickore & Miehe 2001) of
the Tibetan highlands)
(5) the hosting of world’s probably richest alpine flora, and the occurrence of a snowbed in
direct vicinity to the tropics.
The project’s principal goals are
(1) to unveil general diversity structures of the hotspot (“where the hotspot is hot and where
is it cold?”/”dark diversity”), expecting that such patterns open insights to the evolution of
species diversity/richness,
(2) to unveil diversity structures of the supposed Tethys woodlands of the Dry River Gorges
as a set of the ‘Plant Museums’ (Lopez-Pujol et al. 2011), expecting that the species set and
its genetics contribute to the question of the relief evolution of the area,
(3) to record the alpine plant communities of sky mountains along their longitudinal chain
from 21°N towards the Southeastern Himalaya, expecting that latitudinal trends in species
numbers and species sets are related to evolutionary pathways (“Out of Tibet into the
Himalayan Periphery Exile” vs “Out of the Tropical/Holarctic contact zone into the emerging
mountain biota of the Himalayas/Tibet”; Plant Cradle of Alpine Flora: Lopez-Pujol et al.
2011),
(4) to record the cryptogamic and vascular plant species set of snowbeds and snow
avalanche areas.
Suggested work packages

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
33
1.1 Review of current knowledge (This is pre-submission work to pre-test hypothesis):
Diversity Patterns according to the regional accounts (e.g. Flora Yunnanica/Flora of
China/Seed Plants of the Alpine Subnival Belt…/Plant List Hengduan Shan/Plant List
Gaoligong Shan/ Animals??). Question: What are the distribution patterns according
to plant lists? Which biota are rich/poor, what is the share of alpine biota?
1.2 Plot-based elevational transects: where is it possible to record natural sites all along
from the river to the summit (Yunnan??/Myanmar!!, Vietnam??, Bhutan!!).
Question: How is diversity distributed (where is it rich/poor/are there humps, who
makes the humps?)
1.3 Plant communities of the Dry River Gorges.
Question: Does the similarity of species set in the 3 river gorges give insights in the
ecological stability/age of dry Tethys woodlands?
1.4 Alpine plant communities of longitudinal (21° - 28°N) Sky Mountains in the SE
Himalayan Periphery (Chin Hills/Myanmar, Gaoligong Shan, Daxue Shan, Wuliang
Shan, Ailao Shan).
Question: Cradle/Species pump of the Eurasian Alpine Flora or Exile out of
Tibet/Hengduan Shan?
1.5 Plant communities of snowbeds and snow avalanches.
Question: Is the southernmost snowbed flora the cradle of the Eurasian snowbed and
avalanche flora? How old is the snowbed flora?

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
34
Project 1: Role of topographically based habitat richness for the future and the past of
the biodiversity hotspot of Yunnan
Contributors (alphabetical): Wilfried Konrad, Georg Miehe, Christoph Neinhuis, Anita Roth-
Nebelsick, Christiane Ritz, Stefan Wanke
Introduction
Abiotic environmental parameters, such as soil and air humidity, type of soil, insolation, mean
temperature and its annual course, form a subset within all conditions which are limiting the
successful existence of a plant taxon. Accordingly, habitat shifts are expected to be a
consequence of climate change (and already partially demonstrated for current
anthropogenically caused global changes), as is – over longer time periods – evolution.
Speciation and species extinction were repeatedly linked to changing environmental
conditions. For instance, disappearance of biotopes are considered a major extinction cause.
Important for the formation of suitable habitats is, however, not only the macroclimate which
characterizes a large area, or the mesoclimate characterizing regional conditions. In fact, a
quite large number of microhabitats exist within a local climate, as is, for example, commonly
demonstrated for species with different insolation demands but living quite next to each other
on adjacent shady or fully insolated spots. A certain macro- or mesoclimate can thus
accommodate a number of different habitats with quite different conditions.
The number of different local habitats should increase with increasing topographic
complexity. For instance, in mountainous areas, slopes face different directions (north-facing
slopes, south facing slopes etc.), with very different daily climate. If such slopes include
further topographic structuring, such as rifts or smaller elevated areas, the number of
different microhabitats on the slopes will further increase, and expectedly with increasing
topographic complexity. When being subjected to climate change, such a mosaic of
microhabitats may serve as a “habitat buffer” that allows for local habitat shifts within its
boundaries, thus preventing extinction. Moreover, microhabitats may be fragmented and
isolated in the process, depending on topography and presence of geological processes,
such as erosion, facilitating speciation processes.
Complex topographies are expected to be particularly possible with mountainous
landscapes. The combination of topographically caused mosaic patterns of microhabitats
and additional climatic diversity superimposed by altitude gradients may thus make richly
structured mountainous areas particularly suitable for accumulating species (via habitat shifts
within the microclimatic buffers) as well as for speciation events. It is suggested to test this
idea for the Yunnan hotspot with a suit of cooperation subprojects. The results of this project
will be constantly discussed and updated with respect to the work of other groups.

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
35
Subproject Leaf traits and function along selected altitude gradients
Leaf traits have attracted considerable attention in ecophysiology as well as in palaeobotany,
based upon numerous correlations with environment and/or leaf functions. There are three
types of leaf traits: 1) architectural leaf traits, such as leaf shape and type of margin, 2)
(eco)physiological leaf traits, such as maximum assimilation rate Amax or leaf life span (LLS),
and 3) (bio)chemical traits such as nitrogen content or leaf dry mass. In ecophysiology, the
concept of the Leaf Economic Spectrum (LES) focuses on LLS, leaf mass per area (LMarea),
nitrogen content and Amax as traits that are substantially correlated with environment and with
each other (Wright et al., 2005). The underlying reason for these interrelationships is
explained as two different economic strategies, that of “slow” and “fast” resource acquisition
and processing (Reich, 2014). These functional traits are completed by venation
pattern/density and stomatal density and arrangement which are - as representing the leaf
supply and gas exchange system – connected to productivity and the LES (Blonder et al.,
2011; Fiorin et al., 2016; Roth-Nebelsick et al., 2001).
In palaeobotany, architectural leaf traits showing correlations with environment are
extensively used in palaeoclimate reconstruction (Yang et al., 2007). Possible functional
causes for many of these correlations are still not well understood (Royer et al., 2012), but
may be at least partially related to the LES. The adaptive character of various traits may,
however, be attenuated by phylogenetic signals, i.e. various lineages “stick” to taxon-specific
traits (Little et al., 2010; Nobis et al., 2012).
It is suggested to address the following questions, in cooperation with project partners:
Are there correlations between LES and other leaf traits and selected
altitude/microclimate gradients, with respect to lineages and communities?
How do leaf traits of lineages behave, i.e. do they show adaptive plasticity or reveal
phylogenetic signals?
What are the ecophysiological profiles of the lineage representatives?
How do the results relate to palaeobotanical results?
Based on:
Monitoring of environmental conditions along transects
Vegetation along the transects, correlations with microclimate and topography
Determination of microscopic and macroscopic leaf traits, according to the LES and
other concepts
Community-based as well as taxon-based
Correlations with microclimate and topography

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
36
Subproject Ploidy level variation across altitudinal gradients
Polyploidy is ubiquitous phenomenon in plants and is considered as rapid mechanism for
sympatric speciation (Adams and Wendel 2005). Since Stebbins (1950) and Löve and Löve
(1967) it has been generally assumed that polyploids are more frequent in stressful
environments at high latitudes and high altitudes. This view is supported by the observation
that formation of polyploids is most frequently triggered by the production of unreduced
gametes which is in turned enhanced by environmental stress (e.g. cold, nutrition, water
stress; (Mason and Pires, 2015; Ramsey and Schemske, 1998). However, the dogma about
distribution of polyploids has becomes recently more elusive because floristic approaches did
not account for differences in biogeographical and phylogenetic histories of lineages, and
due to the fact that tropical and subtropical regions remained understudied (Weiss-
Schneeweiss et al., 2013). In contrast to Brochmann et al. (2004) who found a high incidence
of polyploids in the artic flora (60.7%), European mountain ranges and the Hengduan
mountains in Yunnan contained an unexpected low proportion of polyploids in their flora
(about 20%; Nie et al. 2005; Loureiro et al. 2013), but these polyploids were indeed mostly
restricted to high altitudes (Loureiro et al. 2013). Such studies relied mainly on bibliographic
reviews and did not systematically analyse the distribution of cytotypes in relation to altitude
or other ecological factors. However, such knowledge is very useful because ploidy variation
may lead to ecological niche differentiation (Sonnleitner et al. 2010). Polyploidy might also
indirectly influence ecological diversification because lineages containing polyploids were
taxonomically more diverse than diploid lineages (Petite and Thompson 1999; Vamosi and
Dickinson 2006). Up to now case studies in species-rich genera from Hengduan mountains
revealed controversial results some implying the polyploidy played a major role in the
evolution of some genera (Chen et al. 2014; Meng et al. 2014, Luo et al. 2011); but not in
others (Deng et al. 2011, Liu et al. 2010; Yuan et al. 2008, Zhuo et al. 2008, Jin et al. 2007).
Possible research questions
Does polyploidy have an impact on plant diversification in Hengduan mountains? Are
there differences between lineages containing polyplpids than those containing
mainly diploids (e.g. Delphinium/Rosa: endemic Rosa praelucens highest naturally
occurring ploidy level in the genus (Jian et al., 2010)
Is there any correlation between the distribution of polyploids and altitude?
Does polyploidy effect functional traits (leaf characters etc.)?

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
37
Subproject Phylogenetic study of lineages occurring along selected altitude gradients
- Phylogenomic analyses of plant biodiversity
Potentially hundreds of loci are needed to resolve recalcitrant phylogenetic relationships
(Leaché and Rannala 2010, Wickett et al. 2014, Prum et al. 2015). Recalcitrant relationships
are very short internodes usually on shallow level, but also found on deep level. Those ‘‘short
branched clades’’ are found all over the tree of life and as such in many plant lineages. For
plants, it has early been recognized that these nodes are among the most challenging to
resolve (e.g. Richardson et al., 2004).
Whole genome sequencing would provide all available loci of an organism and researchers
would only need to select loci matching the requirements of the respective phylogenetic
question. Despite decreasing sequencing costs, whole genome sequencing is very
expensive if applied on a broad sampling and overstates the case as only a fraction of the
genome is potentially needed (Ruane et al. 2015). Furthermore, it requires a lot of
downstream (post sequencing) processing with still a lot of custom scripts and only a glimpse
of the obtained data is usable for phylogenetic questions (Carstens et al., 2012; Lemmon and
Lemmon, 2013). Consequently, if it is not possible to resort to multiple already sequenced
genomes, other methods have to be used to obtain a sufficient number of informative loci.
One recently developed technique is “Anchored Hybrid Gene Enrichment” (AHGE). AHGE
describes an approach where, prior to sequencing, selected loci are captured and enriched
with the help of “capture probes”, oligonucleotide sequences of ~60-120 bp that hybridize to
target regions (Lemmon and Lemmon 2013). Lemmon et al. (2012) designed a vertebrate
capture probe set and obtained a fully resolved and supported species tree for amniotes and
vertebrates. More recently, the same capture probe set was applied on Caenophidian snakes
(Pyron et al. 2014), African Agama lizards (Leaché et al. 2014 a), birds (Prum et al. 2015)
and Malagasy pseudoxyhophiine snakes (Ruane et al. 2015). However, this probe set only
works for vertebrates. For plants a capture probe set has been developed (Mast et al. 2013,
2014) and was successfully tested on the species used to design the probes (Buddenhagen
et al. in prep, Systematic Biology). Furthermore qur research group has applied the probe
set using closely related species/populations (Müller et al. in prep., Molecular Phylogenetics
and Evolution) and we developed a multiplexing strategy called Plant Anchored MetaPrep
(Grandos-Mendoza et al. in prep, Methods in Ecology and Evolution) to significantly reduce
the costs for densely sampled phylogenomic studies with multiple target taxa across the
angiosperms.
Possible taxa to be studied are (eight-twelve target lineages covering the angiosperm tree of
life)
Aristolochiaceae, Magnoliids

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
38
Piperaceae, Magnoliids
Hydrangeaceae, Asterids
Theaceae, Asterids
Cyperaceae (traits?) monocot
One additional monocot lineages (to be discussed) Bambus Gattungen?
Pedicularis, Lamiales
Rhododndron (Ericales)
Rosa, Rosids
Fabaceae, Rosids? (Christiane?)
Caryophylaceae? Primulaceae?
Subproject Evolution of Rosa
Out of 166 wild rose species native in China, 41 species are endemic to the Yunnan province
occurring at altitudes from 1800 to 3400 m (Cuizhi and Roberston, 2003). Some of these
species were analysed in genus-wide or section-wide phylogenies (Chen et al., 2014;
Fougère-Danezan et al., 2014) (Fougere-Danezan et al. 2015; Zhu et al. 2015) but a
biogeographical analyses of the roses from the region is missing to date. Comparisons
between fossil rose leaves from the late Miocene of the Yunnan Province (R. fortutita) with
extant species of the region revealed the presence of semicraspedodromous venation – a
rather rare character state in the genus – in both fossil and many extant taxa suggesting that
the general climatic requirements for the genus did not substantially change since the late
Miocene in the region. (Su et al., 2015). Moreover, the semicraspedromous venation was
also found in Rosa lignitum from Oligocene in Europe (Kellner et al., 2012) supporting the
hypothesis that the current climatic conditions in SW China match the European tertiary
climate.
In general, combined with genetic analyses leave traits are appropriate tools to investigate
ecological adaptations and/or evolutionary relationships in Rosa.
Possible research questions
Biogeographic origin of Rosa species in Yunnan
Do leaf characters correlate with ecological characters and/or with taxonomy
Preliminary work:
Kellner, A., Benner, M., Walther, H., Kunzmann, L., Wissemann, V., Ritz, C. M.
(2012) Leaf architecture of extant species of Rosa L. and the Paleogene species
Rosa lignitum Heer (Rosaceae). Int. J. Plant. Sci. 173 (3) 239-250.

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
39
DFG-LIS project: Digitisation / Cataloguing of non-textual objects: Development of
new digitization standards for the large-scale assessment of leaf venation traits from
herbarium collections using microradiographic imaging (Botanik der SGN: FR und
GLM)
References
Chen GF, Sun W-G, Hong, D-Y, Zhou Z, Niu Y, Nie ZL, Sun H, Zhang JW, Li ZM (2014) Systematic significance
of cytology in Cyananthus ( Campanulaceae) endemic to the Sino-Himalayan region. J. Syst. Evol. 52:
260-270.
Cuizhi, G.; Roberston, K. R. (2003) Rosa Linnaeus. In: Zhengyi, W.; Raven, P. H.; Deyuan, H. Flora of China.
Band 9 Pittosporaceae through Connaraceae. Science Press, Beejing, China; Missouri Botanic Garden
Press, St. Louis, USA.
Deng T, Meng Y, Sun H, Nie ZL (2011) Chromosome counts and karyotypes in Chaetoseris and Stenoseris
(Asteraceae-Cichorieae) from the Hengduan Mountains of SW China. . Syst. Evol. 49: 339-446.
Fougere-Danezan M, Joly S, Bruneau A, Gao XF, Zhang LB (2015) Phylogeny and biogeography of wild roses
with specific attention to polyploids. Ann. Bot. 115: 275-291
Blonder, B., Violle, C., Bentley, L. P., and Enquist, B. J., 2011, Venation networks and the origin of the leaf
economics spectrum: Ecology Letters, v. 14, no. 2, p. 91-100.
Chen, G.-F., Sun, W.-G., Hong, D.-Y., Zhou, Z., Niu, Y., Nie, Z.-L., Sun, H., Zhang, J.-W., and Li, Z.-M., 2014,
Systematic significance of cytology in Cyananthus (Campanulaceae) endemic to the Sino-Himalayan
region: Journal of Systematics and Evolution, v. 52, no. 3, p. 260-270.
Cuizhi, G., and Roberston, K. R., 2003, Rosa Linnaeus, Beijing, St. Louis, Science Press, Missouri Botanic
Garden Press, Flora of China - Pittosporaceae through Connaraceae.
Fiorin, L., Brodribb, T. J., and Anfodillo, T., 2016, Transport efficiency through uniformity: organization of veins
and stomata in angiosperm leaves: New Phytologist, v. 209, no. 1, p. 216-227.
Fougère-Danezan, M., Joly, S., Bruneau, A., Gao, X.-F., and Zhang, L.-B., 2014, Phylogeny and biogeography of
wild roses with specific attention to polyploids: Annals of Botany.
Jian, H., Zhang, H., Tang, K., Li, S., Wang, Q., Zhang, T., Qiu, X., and Yan, H., 2010, Decaploidy in Rosa
praelucens Byhouwer (Rosaceae) Endemic to Zhongdian Plateau, Yunnan, China: Caryologia, v. 63, no.
2, p. 162-167.
Kellner, A., Benner, M., Walther, H., Kunzmann, L., Wissemann, V., and Ritz, C. M., 2012, Leaf architecture of
extant species of Rosa L. and the Paleogene species Rosa lignitum Heer (Rosaceae): International
Journal of Plant Sciences, v. 173, no. 3, p. 239-250.
Little, S. A., Kembel, S. W., and Wilf, P., 2010, Paleotemperature Proxies from Leaf Fossils Reinterpreted in Light
of Evolutionary History: PLoS ONE, v. 5, no. 12, p. e15161.
Mason, A. S., and Pires, J. C., 2015, Unreduced gametes: meiotic mishap or evolutionary mechanism?: Trends in
Genetics, v. 31, no. 1, p. 5-10.
Nobis, M. P., Traiser, C., and Roth-Nebelsick, A., 2012, Latitudinal variation in morphological traits of the genus
Pinus and its relation to environmental and phylogenetic signals: Plant Ecology & Diversity, v. 5, no. 1, p.
1-11.
Ramsey, J., and Schemske, D. W., 1998, Pathways, mechanisms, and rates of polyploid formation in flowering
plants: Annual Review of Ecology and Systematics, v. 29, p. 467-501.
Reich, P. B., 2014, The world-wide ‘fast–slow’ plant economics spectrum: a traits manifesto: Journal of Ecology,
v. 102, no. 2, p. 275-301.

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
40
Roth-Nebelsick, A., Uhl, D., Mosbrugger, V., and Kerp, H., 2001, Evolution and Function of Leaf Venation
Architecture: A Review: Annals of Botany, v. 87, no. 5, p. 553-566.
Royer, D. L., Peppe, D. J., Wheeler, E. A., and Niinemets, Ü., 2012, Roles of climate and functional traits in
controlling toothed vs. untoothed leaf margins: American Journal of Botany, v. 99, no. 5, p. 915-922.
Su, T., Huang, Y.-J., Meng, J., Zhang, S.-T., Huang, J., and Zhou, Z.-K., 2015, A Miocene leaf fossil record of
Rosa (R. fortuita n. sp.) from its modern diversity center in SW China: Paleoworld.
Weiss-Schneeweiss, H., Emadzade, K., Jang, T. S., and Schneeweiss, G. M., 2013, Evolutionary Consequences,
Constraints and Potential of Polyploidy in Plants: Cytogenetic and Genome Research, v. 140, no. 2-4, p.
137-150.
Wright, I. J., Reich, P. B., Cornelissen, J. H. C., Falster, D. S., Garnier, E., Hikosaka, K., Lamont, B. B., Lee, W.,
Oleksyn, J., Osada, N., Poorter, H., Villar, R., Warton, D. I., and Westoby, M., 2005, Assessing the
generality of global leaf trait relationships: New Phytologist, v. 166, p. 485-496.
Yang, J., Wang, Y. F., Spicer, R. A., Mosbrugger, V., Li, C. S., and Sun, Q. G., 2007, Climatic reconstruction at
the Miocene Shanwang basin, China, Using leaf margin analysis, CLAMP, Coexistence approach, and
overlapping distribution analysis: American Journal of Botany, v. 94, no. 4, p. 599-608.
Subproject: Research activities in Vietnam National Museum of Nature an Hoang Lien
plant diversity hotspot in Northern Vietnam
Truong Van Do
Vietnam National Museum of Nature, [email protected]
Vietnam National Museum of Nature (VNMN) has been found in 2006, is a state-owned
cultural science museum, under Vietnam Academy of Science and Technology. Over past
ten years, the collection of specimens of VNMN has over 100,000 specimens, i.e. mammals,
reptiles and amphibians, fish, insects, plants, fossils, mineral, and including 300 type
specimens of these collections. The evolutionary history exhibition room has been also built
here for training and education to pupils and students. During past ten years, scientists of
VNMN have described approximately 65 species of reptiles and amphibians, insects, and
plants as new to science.
Additionally, we also present about Hoang Lien National Park as a plant diversity hotspot in
northern Vietnam. Hoang Lien NP locates in the Hoang Lien mountain range in northern
Vietnam where is linked to the Ailao Shan, Yunnan, China, in the southeast of the Himalaya.
Most of the area is over 1000 m above sea level, with a main peak is about 3143 m and is
considered to be the “Roof of Indochina”. Within its natural area of 29,845 ha, there are
21,894 ha of forest. Of this there are 14,678 ha of natural forest which only been slightly
impacted by human activities. Up to date botanists have investigated and listed about 2346
vascular plant species of 1,020 genera and 285 families occurring in the area. Of which,
many species are sharing the distribution with the Yunnan plant diversity hotspot.

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
41
Project 2: Evolutionary Ecology and phylogenomics of endemic plant taxa unravel
evolutionary cues of the HTO uplift
Jian-Quan Liu, School of Life Science, Lanzhou University, Lanzhou, PR China
Lars Opgenoorth, Philipp University of Marburg, Marburg, Germany
Project Description
Based on integrating phylogenomic, fossil and biogeographic analyses, we aim to clarify the
origin and evolutionary drivers of the Yunnan plant diversity. We will utilize groups with high
diversification (endemic species), varying ecological niches, and sufficiently small genomes
in order to be able to utilize genomic data to clarify identify genes or genomic regions with
high differentiation to derive their speciation patterns and drivers. Also we will use the data to
detect and partly date underlying changes of the effective population sizes by modeling and
testing demographic scenarios of speciation. Based on these results, we will recover the key
genes accounting for species divergences and adaptive differentiations as well as their
causal relationships. We will integrate all results to make clear the origin and evolution of the
Yunnan plant diversity.

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
42
3.2. Invertebrate groups
Project 1: Radiation into megadiversity: Ground beetles as phylogenetic model group
can unravel evolutionary principles of the Himalaya-Yunnan Hotspot
Joachim Schmidt, University Rostock, Rostock, Germany
Lars Opgenoorth, Philipp University of Marburg, Marburg, Germany
Project Description
We will utilize ground beetle phylogenies to unravel how orogenesis and the radiation in this
megadiverse taxa are linked. Ground beetles are an ideal biological indicator to derive
evolutionary processes, timing and consistence of the Himalaya-Yunnan Hotspot. Some of
the features of ground beetles that make them uniquely suited are, that they
1. provide a large spatial resolution (many species, populations, or genotypes) in the entire
research area.
2. have strong ties to a specific ecological niche, which will enable conclusions to be drawn
from the presence of that taxon regarding important abiotic factors of the environment
(e.g., temperature/elevation and humidity/precipitation).
3. populate all altitudinal belts of the high mountain range with characteristic and specialized
species assemblages.
4. are unspecialized predator or detritivore, which is independent of the existence and the
distribution of specific prey species.
5. are weak disperser in evolutionary timescales and thus are unable to fly or disperse over
large distances via vectors such as wind, large animals or humans, and thus exhibits
strongly differentiated lineages spatially.
6. have an availability of sufficient calibration points also extendable in the framework of the
project for the molecular-clock approach to date adequately the clades, taxa, and
genotypes.
7. provide a sufficient amount of taxa to fine-tune the molecular-clock approach and include
a coherent timeframe with coherent error bars.
8. have an availability of a comprehensive sample collection that is easily extendable by
additional fieldwork and for which a good systematic revision exists.
In mountains at lower latitudes as in the Himalaya-Yunnan Hotspot, almost all of the ground
beetle lineages adapt to the high mountain habitat by the reduction of their hind wings and
flight muscles as well as an irreversible transformation of their exoskeleton (distinct
shortening of the metathoracal plates). This evolutionary stage is termed primary
winglessness (winglessness by descent, Schmidt et al. 2011) to differentiate it from the
facultative winglessness stage (species with a normal metathorax and presumably with the

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
43
reversible development of hind wings in at least some individuals). Facultative winglessness
in ground beetles is a common phenomenon in mountains of mid and higher latitudes. In
contrast, our long-term study in the Himalaya and in Tibet demonstrated that the vast
majority of ground beetle lineages adapted to the cloud forests and to the alpine zone of the
HTO belong to the primary wingless species (e.g., Andrewesius: ca. 45 species; Ethira clade:
76 species; Lepcha: ca. 50 species; Meganebrius: 25 species; Xestagonum: > 250 species).
In these species groups, winglessness appears to have evolved during an early stage in their
evolutionary history. Winglessness induces an extreme reduction in ground beetle vagility.
The dispersal of primary wingless species can only occur on foot. Therefore, because of the
multitude of potential barriers in high mountains (mountain crests, glaciers, deep river
gorges, dry slopes), dispersal is only possible within a very restricted geographical range.
Thus, the current range position of a lineage of wingless ground beetles is closely related to
the geomorphological and climatological development of the mountain range they inhabit.
The fossil database of ground beetles that will be used for calibrating the molecular clock is
of high-quality although it has only been partly revised. Ober & Heider (2010) published an
overview of the most useful Carabidae fossil evidence in the literature. Additional fossils of
the genus Carabus from the Oligocene and Miocene of Europe and North America are
known (reviewed by Deuve, 2004), and recently, two modern ground beetle genera
(Bembidion and Calathus) in Baltic Amber were described by Ortuño & Arillo (2009, 2010)
and of our working group (Schmidt 2015, 2016). However, if one considers the catalogues of
insect inclusions in a number of Baltic Amber collections (e.g., Klebs 1910, Larsson 1978,
Hieke & Pietrzeniuk 1984, Bachofen-Echt 1996, Weitschat & Wichard 1998), the fossil data
pool of ground beetles is much higher than one would expect from the number of existing
descriptions of fossil taxa. Indeed, many fossil examples of many modern ground beetle
tribes or genera are preserved in Baltic Amber collections (e.g., Agonum, Lebia,
Pterostichus, Syntomus, and Trechus among others). However, to date, these fossils have
not been morphologically analyzed or systematically revised, and the true systematic
positions of most of the Baltic Amber fossils remain questionable. Morphologic-systematical
studies of the fossils are of major importance in paleo-carabidology and are a key element of
this application.

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
44
Project 2: Freshwater Invertebrates as models for studying the origin and the causes
of species diversity in Yunnan – Project outline
Martin Brändle, Roland Brandl
Philipps-Universität Marburg, Faculty of Biology, Department of Ecology – Animal Ecology
Due to their insular character and clear physical boundaries freshwaters are good model
systems to study species communities, species propensity for dispersal but also the
evolution of species diversity. Freshwaters in general provide habitats for a large number of
invertebrate species comprising several taxonomic groups. Species within and among
taxonomic groups differ in life history traits, diet utilization and environmental requirements.
Many freshwater invertebrates are restricted to use either standing (lentic) or running (lotic)
waterbodies. Strong differences of these habitat types in predictability and environmental
conditions provide versatile opportunities to study genetic population differentiation, species
propensity for dispersal and the evolution of species diversity.
The Yunnan landscape is covered by a large number and variety of freshwater habitats.
Specific habitat characteristics of these environments and their spatial distributions may
serve as important template for the origin of the Yunnan hotspot of species diversity. In our
project we will focus on two overarching core question that are central for the main project: (i)
When did the Yunnan hotspot first appear in Earth History, and how did diversity
evolve? (ii) What are the main drivers in the evolution of diversity?
(i) To identify temporal patterns of diversification and the timing of radiations we will
consider time calibrated molecular phylogenies and lineage through time plots of
selected lentic and lotic freshwater invertebrates. For both habitat types we will
also analyze taxonomic groups that differ a priori in propensity for dispersal, for
instance wingless versus winged taxonomic groups (e.g. flatworms, mollusks,
crustaceans versus insect orders). The spatial patterns of diversification will be
analyzed by contrasting taxon age and phylogenetic relatedness with geographic
settings and distribution and subsequently compared between lineages differing in
propensity for dispersal. Here we are in particular interested how mountain uplift
influenced the diversification and distribution of species differing in phylogenetic
age. For example, we expect for poor dispersers that phylogenetic older groups
are more restricted to the bottom of the mountains while younger groups should
be found rather at higher elevation. In addition, we expect that populations of lotic
and wingless invertebrate species (flatworms, mollusks and crustaceans) are
more strongly spatially structured than lentic species and species of insect orders.

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
45
(ii) To identify the main drivers of diversification in Yunnan we will compare patterns
of species richness and composition of the Yunnan freshwater invertebrate fauna
with those of another mountain region, namely Central Europe and Southern
Europe. By relating species richness among higher taxonomic groups against
each other it will be possible to identify groups that show a disproportionally
higher richness in Yunnan. After having identified these groups we will determine
what are the dominant life history traits or niche requirements of the species
within these groups? For instance, the case that taxonomic groups comprising
more lotic species show a much higher species number in Yunnan than in the
Alps would indicate that the structuring of lotic freshwaters may have served as
an important driver of diversification. In a similar vein, a finding that higher
richness of typically alpine or subtropical clades in Yunnan would provide hints
that the distinct relief and the heterogeneity of the landscape have to be
considered as an important factor driving the diversification of the Yunnan
hotspot.

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
46
Project 3: Biodiversity of Cenozoic insects from Yunnan (China)
Sonja Wedmann, Senckenberg Research Institute Frankfurt/M., Germany
Insects are the most diverse organisms in the history of life, with more than one million
described species, and millions of species still unknown. Insects also have a rich fossil
record, but although many studies have been done and are still ongoing, the course of
evolution of the biodiversity of insects is still poorly known in large parts, even for the
Cenozoic.
During the early Cretaceous there was a major turnover in the insect fauna, probably related
to the radiation of flowering plants (Ross et al. 2000). By the late Cretaceous, about 95% of
insect families were the same as they are today, with no significant decrease across the K/T
boundary (Grimaldi & Engel 2005). For the Paleocene, fossil sites with insect remains are
scarce, but the general trend for increasing insect biodiversity during the Cenozoic is well
documented by rich insect localities from the Eocene, e.g. Baltic Amber and Messel from
Europe. The high insect biodiversity in the Paleogene was attributed to high origination rates
and low extinction rates along with a nearly worldwide tropical to temperate climate (e.g.
Grimaldi & Engel 2005). A drastic climatic cooling during the Eocene-Oligocene transition
resulted in a “lesser” mass extinction that also affected the insects. Since modern types of
tropical forests and grasslands began to establish themselves in the Oligocene, this might
also be the beginning of our modern insect fauna (Grimaldi & Engel 2005). The implications
of the climatic shifts during the Neogene are poorly studied, the Miocene is a very interesting
time span in this respect, and there is still much to do.
Similar to the evolution of insect biodiversity, the interrelationships between climate change
and biogeographic diversity patterns are still under discussion. Fossils have been shown to
be important to redefine biogeographic hypotheses for many different insect groups, which
were believed to have restricted distributions when only extant taxa were considered; see
Eskov (2002) and Grimaldi & Engel (2005).
Considering this broad context, it seems very promising to study fossil insects from the
Yunnan province in China. The knowledge on fossil insects from this region is very scarce. If
collections of fossil insects from Yunnan can be made and are available for study, their
analysis would be the base for a comparative study of the evolution of insect biodiversity in
this region, and also the base for further biogeographical and comparative studies.
From the Shangyong village, Mengla County, Yunnan, a few unique insect compression
fossils have been described (Lin et al. 2010a, b), but it is recorded that fossil insects can be
found frequently (Lin et al. 2010 a). The age of these insects is considered to be upper

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
47
Paleocene or lower Eocene, but the dating of the sediments was based on Conchostracans
and it is not uncontroversial. But the time is certainly very interesting, and comparison with
Messel and other fossil sites would be promising.
From Maguan County, Wenschan City, Yunnan, besides plants (Zhang et al. 2015) several
insects were found, which will be studied soon (personal comm. T. Wappler October, 2015).
These insects probably have a late Miocene age (Zhang et al. 2015), and so they represent
a sample of a fauna which is much more recent. Especially analysis of a larger collection of
these insects, which probably would have to be assembled in the course of the project,
would permit interesting comparisons over time. There are also other Miocene sites from
which plants are known, and it would be interesting to investigate whether there can perhaps
also insects be found (Ma et al. 2005, Su et al. 2013).
Eskov, K. Y. 2002. Geographical history of insects. 427-435. In: Rasnitsyn, A. & Quicke, D. L. J. (Eds.): History of
insects. Kluwer Academic Publishers, Dordrecht, Boston, London, 517 pp.
Grimaldi, D & Engel, M. S. 2005. Evolution of the insects. Cambridge University press, Cambridge, New York, etc.
755 pp
Lin Qibin, Szwedo, J., Huang Di-Ying, Stroinski, A. 2010a. Weiwoboidae fam. nov. of 'Higher' Fulgoroidea
(Hemiptera: Fulgoromorpha) from the Eocene Deposits of Yunnan, China. Acta Geologica Sinica (English Edition)
84: 751-755.
Lin Qi-Bin, Petrulevicius, J.F., Huang Di-Ying, Nel, A., and Engel, M.S., 2010. First fossil Calopterygoidea from
Southeastern Asia (Odonata: Zygoptera): A new genus and species from the Paleogene of China. Geobios 43:
349-353.
Ma , Q. W., Li , F. L. & Li, C. S. 2005: The coast redwoods ( Sequoia ,Taxodiaceae) from the Eocene Heilongjiang
and the Miocene of Yunnan, China. Review of Palaeobotany and Palynology 135 : 117 – 129.
Ross, A. J., Jarzembowski, E. A. & Brooks, S. J. 2000. The Cretaceous and Cenozoic record of insects
(Hexapoda) with regard to global change. 288-302. In: Culver, S. J. & Rawson, P. F. (Eds.): Biotic response to
global change - The last 145 million years. Cambridge University Press, 501 pp.
Su , T. , F. B. Jacques , H. J. Ma , & Z. K. Zhou. 2013. Fossil fruits of Ailanthus confucii from the upper Miocene
of Wenshan, Yunnan Province, SW China. Palaeoworld 22 : 153 – 158 .
Zhang, J. W., D’Rozario, A., Adams, J. M. , Li, Y. ,Xiao-Qing Liang, X-Q., Jacques, F. M., Su, T. & Zhou, Z.-K.
2015. Sequoia maguanensis , a new Miocene relative of the coast redwood, Sequoia sempervirens , from China:
implications for paleogeography and Paleoclimate. American Journal of Botany 102: 103 – 118.

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
48
3.3. Vertebrate groups
Project 1: Evolutionary history and present phylogeographic patterns of passerine
birds of the Yunnan biodiversity hotspot
Martin Päckert1, Jochen Martens2, Yue-Hua Sun3, Patrick Strutzenberger1, Swen Renner4
1Senckenberg Naturhistorische Sammlungen, Dresden, Germany
2Institut für Zoologie, Johannes-Gutenberg-Universität, Mainz, Germany
3Key Laboratory of Animal Ecology and Conservation, Institute of Zoology, CAS, Beijing, China
4Institute of Zoology, University of Natural Resources and Life Sciences, Vienna, Austria
Background: The mountain forests along the southern and south-eastern margins of the
Qinghai-Tibetan Plateau (QTP) are one of the most prominent biodiversity hotspots of the
Northern Hemisphere. Local species richness is particularly high in the Eastern Himalayas
with no less than 358 species along a local elevational gradient. The Hengduanshan harbors
a similarly rich biodiversity including a high number of endemics. Phylogeny combined with
reconstructions of biogeographic history suggested a major burst of diversification in almost
all passerine families during the mid to late Miocene. In these times the Yunnan biodiversity
hotspot was a center of origin and diversification for several groups of passerine birds such
as rosefinches and leaf-warblers. Extant distributions and phylogeographic patterns along
the eastern QTP margin were predominantly triggered by Pleistocene range contractions and
Holocene range expansions. Cryptic diversity is high in the Yunnan biodiversity hotspot and
species identification or recent descriptions of taxa new to science were mainly based on a
combination of morphological, genetic and bioacoustics traits – so-called integrative
taxonomy. Though genetic barcodes of birds are available in dense regional samplings
worldwide, the QTP region is still greatly underresearched. First results from a DFG-funded
barcode initiative on Eurasian birds confirm a considerable percentage of ‘undetected
species’ in the QTP region and stress the need of further research particularly in the
biodiversity hotspots.
Perspective: So far, our BOLD database comprises more than 600 COI barcodes of
Eurasian birds with dense samplings along the eastern margin of the QTP and in the Central
Himalayas (Nepal). The database is steadily being enlarged and can be used as the basis for
further biodiversity research in the region. Our sampling is complemented by rich material
from northern Myanmar (Hkakabo Razi) at the frontier to China (Yunnan) – from a
zoogeographic perspective this region is still part of the Yunnan Biodiversity Hotospot. Based
on our bacording initiative our future research will focus on phylogeographic studies that shall
help uncovering intraspecific differentiation and cryptic biodiversity. The genetic studies will
be flanked by bioaoustic analysis of territorial songs.

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
49
4. Present-day diversity distribution in Yunnan – a macroecology and modelling
analysis of present-day patterns
Understanding changing patterns of diversity in Yunnan and beyond
Alice C. Hughes1 and Matthew Forrest2
1Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences and 2Biodiversity
and Senckenberg Biodiversity and Climate Research Centre (BiK-F)
Abstract
Southeast Asia represents a complex and dynamic region both in terms of ecology and
geography. It currently maintains rich biodiversity [[references]] against a historical backdrop
of many geophysical processes and events (and the emergence of the Sunda shelf during
considerable periods of history has a lasting influence on contemporary patterns of
biodiversity) which have a had a significant effect on the landscape [[references,
references]]As a result, understanding modern patterns of diversity, and disentangling
genuine habitat requirements and physiological drivers of modern distributions from the
historical biogeographic drivers of biodiversity is challenging. Paleontological data from
across much of the region is patchy in terms of spatial and temporal resolution, and as a
result there is still dispute as to the landcover and connectivity across much of the region
during crucial portions of the region’s history, such as the last Glacial maximum. Employing
process-based models such as General Circulation Models (GCMs) and Dynamics Global
Vegetation Models (DGVMs) to study previous geological periods in combination with a
synthesis of existing paleontological data offers a means to build more complete picture of
past landscapes. Furthermore, using such models and data in tandem with present-day
biodiversity, species distribution and trait data may provide insights into both the habitat
changes in region and its effects on biodiversity. In a further extension to this methodology,
empirical models of biodiversity could be constructed using similar techniques to, or build
upon Species Distributions Models (SDMs) which are widely applied to predict present-day
distributions from a range of environmental data layers. By using consistent and mechanistic
simulations of climate and vegetation fields across different geological periods, and using
models of biodiversity calibrated using present day data and model output, we intend to
predict biodiversity patterns of the past and future, and examine the role of geophysical
processes in today’s biodiversity.
Research questions
We propose to develop a framework of interlinking research packages and novel
methodologies to tackle the following broad research questions:

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
50
Research question 1. How have key geophysical events and processes affected the
evolution of biodiversity in Southeast Asia and in particular the Yunnan biodiversity hotspot?
Research question 2. How has regional climate changed over time, and how has the
changing configuration of the region effected long term climatic trends
Research question 3. Which factors (including habitat change effects) best explain present
day biodiversity patterns in several keys groups (bird, bats, mammals, amphibians, insects,
trees) in Southeast Asia?
Background
Modern distributions of any given species will necessarily reflect former distributions, and
especially for dispersal limited species the legacy of former distributions is of particular
importance in understanding current patterns of diversity. Species distribution models for the
present and past and calibrated with appropriate data, have provided insights into species
distributions and useful projects of how species distributions might change under global
change scenarios.Such models can also be applied to previous time periods to estimate
species distribution, and how these may have changed over time, the choice of input data is
critical in obtaining meaningful and useful outputs. For example climatic models of this highly
diverse and heterogenous region are normally based on the interpolation of very coarse raw
climate data over the topography of the area using various splines, and without calibration
and testing for many regions (Deblauwe et al., 2016). As a result, the differences between
the climate in any two regions may actually be exceeded by the variability and uncertainty for
any given climate parameter in those regions, and such variation and error is likely to be
further compounded by extrapolation over time. Other issues, such as the changing
topography of the region (i.e the emergence of the Sunda shelf for half of the regions
geophysical history since taking it’s current configuration) will also have effected air-currents,
and thus profoundly affected the climate and even monsoon regimes across the region.
Thus, for accurate models of the distribution of species of any region, and especially one with
such heterogeneity having accurate climatic maps is essential.
Another important consideration when studying species distributions and biodiversity
patterns is the biotic, as opposed to the abiotic conditions of the landscape: specifically the
vegetation cover. This can be described by discrete land cover categorisations, such as
biome type, or as continuous variables such as woody cover fraction, net primary
productivity, phenological indices, and many others. For the present day, many of these can
be estimated using remote sensing data, however for the past and the future no such
measurements are available. Thus if spatially extensive maps describing the key features of
vegetation cover are to be used for calculating species ranges and biodiversity outside of the
present day, they must be reconstructed using models. This is of particular importance for
Southeast Asia, as for much of the region an understanding of the distribution of different

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
51
landcover types over important periods of history (i.e the Last Glacial Maximum) are crucial
to understanding patterns of refugia, population connectivity and thus drivers behind current
distribution.
Vegetation can of course be mapped and modeled in a variety of ways, and using
appropriate data (i.e. pollen, speleotherms, etc) it is possible to test, calibrate and verify the
accuracy of such vegetation models across time. LPJ-GUESS is an eco-system model which
also provides an alternate means of mapping regional landcover ( Forrest et al 2015).
Once vegetation models have been developed and tested for various time periods, the
combination of climatic, vegetative and topographic data can be used to run biodiversity
models (using predictive techniques) for a suite of taxa for each time period under scrutiny.
These changing distributions can then be explored using molecular approaches (where data
is available, and for select taxa only) to explore genetic connectivity, and phylogenetic
endemism, and explore the changing connectivity of these groups over time and the
accuracy of glacial refugia projected.
The intersection of various forms of data to understand past and present distributions of
vertebrate and plant species across the region represents a challenge, which without the
modern availability of data, software and higher capacity to run computationally intensive
models would have been impossible. Species distribution data for the present, and to some
extent for the past has already been obtained, though the databases are still under the
process of development (so are growing) under a separate initiative also lead by the leaders
of this proposal. Broadly we intend to build high resolution environmental and climatic data
for a number of time periods using various forms of available data, to build onto this
vegetation models for the appropriate periods, and then to hindcast modern diversity onto
these projected landscapes using species distribution modeling techniques, and calibrating
the outputs through genetic analysis (for changes since the LGM) and through fossil data for
older analysis. Thus the outputs promise new insights into patterns of diversity and their
drivers across geophysical time, and provide a basis to better conserve current species and
genetic diversity. Additionally outputs should be capable of detecting, to at least some
degree, the sensitivity of different species to change and their probable reactions to future
climatic changes.
Furthermore by examining the effects of climate stability and variability across such a
heterogeneous landscape across time, and relating it to modern diversity and endemism,
calibrated with molecular data will provide new perspectives on how species respond to such
changes over time. It is probable that areas with moderate levels of stability have the highest
diversity, whereas highly unstable regions may have a combination of highly endemic
dispersal poor taxa (which have survived through change) and highly generalist good
dispersers which have since colonized those regions. Modern patterns of distribution,

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
52
combined with population genetics will elucidate the changing distributions of each,
combined with the rate of movement and therefore carry useful implications for species
future responses to changing climate. The distribution of refugia across different areas of the
landscape, and the relative “velocity of climate change” related to the areas topography may
also carry important lessons for how species may respond to future change
Work Packages
WP 1. Paleoclimate modelling. Personnel ???
This depends on Johan Liakka's involvement. Feeds into WP 2 and 3. Alternate plan 1, use
data from/bring in Todd Ehlers? He and Jingmin Li are doing Middle Miocene stuff with the
explicit intention of looking of looking at East Asian monsoon. It might be easy(ish) for them
to do simulations other time periods. Alternate plan 2, Hui Tang’s simulations, and
collaborations, he seems to being similar things to Todd. But I think Johan is preferred ;-)
WP 2. Vegetation modeling. Personnel: One student in Frankfurt?
First we develop a regional parameterization for the model, also including paleo taxa. From
this we can offer information to about biomes, productivity, and other environmental data to
the other projects and, importantly, to WP3. Optionally, we can also consider starting to put
mechanism to allow trait variation within one PFT. Maybe, if this is done right, it also can be
used to explain biodiversity? Maybe then this is a hypothesis:
WP 3. Biodiversity modeling: Past, Present and future. Personnel ?? A student? In China
or Germany?
I think that this should start with mining the trait databases and biodiversity data (whilst the
other WP are getting the modelling going). The trait data can be used for a) A data driven
approach for testing hypotheses and b). Validating the trait-based improvements to the veg
model. Then, once the modelling is ready, they can use the model results and the data to
build the biodiversity model and answer some hypotheses. It is better for more informed
people to work these out, but initially I see possibilities like:
Hypotheses concerning drivers of present day biodiversity patterns

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
53
5. Future threats
Global Warming and changing monsoon? (Modelling/A. Hughes + M. Forrest)
Conservation strategies? (D. Cicuzza)
Fire? (Li Shu-Feng)
Land use / medicinal plant harvesting / mushroom economy (KIB, Yunnan University)
Abstracts/project outlines still required

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
54
Field trips
Day 1: Vereinigtes Schleenhain opencast mine, NW Saxony, Germany.
Day 2: Welzow-Süd opencast mine, S Brandenburg, Germany.

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
55
Field trip destinations opencast mine Vereinigtes Schleenhain and Welzow-Süd (map:
Susann Stiller).
S a
x
o
n
y
Bra
nd
en
bu
rg 2
0km

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
56
Practical palaeontological field work part 1
Eocene to Oligocene vegetation and climate change in the central German
Weißelster Basin (Vereinigtes Schleenhain opencast mine, Saxony)
Lutz Kunzmann & Karolin Moraweck
(text compiled from Kunzmann 2012, Kunzmann and Walther 2012, Kunzmann et al. 2016)
The Vereinigtes Schleenhain opencast mine is situated in the center of the Weißelster Basin,
which is a lignite lagerstätte south of the city of Leipzig in Saxony and Saxony-Anhalt, central
Germany (Meyer, 1950; Fig. 1). The name Weißelster is also applied to the important
palaeontological site in the strata (Walther and Kunzmann, 2008). However, in geological
and structural terms, the Weißelster Basin is not a distinct or isolated sedimentary
depositional area, but rather the southern part of the Leipzig Embayment, which pertains to
the North German Depression (Fig. 2; Standke, 2008a, 2008b). This name has therefore
gradually disappeared from the geological literature, whereas in the palaeobotanical
literature, it remains in use.
The Leipzig Embayment contains middle Eocene to middle Miocene terrestrial and marine
sedimentary sequences (Standke et al., 2010). Its southern end, the so-called Weißelster
Basin, is characterized by a series of fluvial, estuarine, and marine strata, including the
occurrence of several currently economically important lignite seams (Standke et al., 2010).
Lignite has been excavated here for several centuries and is nowadays mined by the
MIBRAG mbH in the Vereinigtes Schleenhain and in the Profen mines (Fig. 3).
The general lithostratigraphic section of the Weißelster Basin (Fig. 4) shows five main
sedimentation units, (1) the late middle to late Eocene Profen Formation with lignite seam
complex 1, (2) the late Eocene Borna Formation with lignite seam complex 2/3, (3) the latest
Eocene to earliest Oligocene Böhlen Formation with lignite seam complex 4, (4) the early
Oligocene “Rupelian” marine sediments, and (5) the late Oligocene Cottbus Formation
without lignite seams. Units (1) – (3) and (5) are predominated by clastic, non-consolidated
terrestrial sediments but (4) is brackish to fully marine. The field trip will guide to fossiliferous
horizons within the units (2) and (3). These sediments represent deposits of a shallow marine
tidal embayment and a coastal alluvial plain (Kunzmann 2012). A series of cyclic
transgressions and regressions caused a formation of large coastal mires accumulating peat
in the flat alluvial plain (Kunzmann et al. in press).

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
57
This area is well-known for its rich and diverse plant assemblages that cover the rather long
time span from the late middle Eocene to the earliest Miocene (Walther & Kunzmann 2008).
Here one can find a series of taphocoenoses of plant fossils, intensively studied, that touch
on the Mid-Eocene Climatic Optimum, the Eocene-Oligocene turnover as well as the late
Oligocene cooling (Zachos et al. 2008). Consecutively, these plant assemblages were used
to establish phytosociological and phytostratigraphic concepts, namely the “Florenkomplexe”
(floristic complex; Mai & Walther 1983, Mai 1995). By a definition, a floristic complex is a
temporary stage in the floristic evolution of regional vegetation independently from the
sedimentary facies type, the soil moisture and the type of plant communities (Mai and
Walther 1983, Mai 1995). Therefore, a floristic complex represents a kind of stasis in the
evolution of regional vegetation over a certain time span. Simultaneously, rich palynofloras
from the same horizons were the basis for another phytostratigraphic tool, the spore-pollen
zonation (e.g. Krutzsch 2011 and references therein). More recently, fossil floras from the
Weißelster Basin have been frequently used as an excellent database to estimate
palaeoclimatic and palaeoatmospheric changes (e. g. Mosbrugger et al. 2005; Grein et al.
2013; Roth-Nebelsick et al. 2014, Moraweck et al. 2015, Steinthorsdottir et al. 2015).
The fossiliferous horizon from unit (2) which is visited during the field trip contains floras from
the late Eocene floristic complex Zeitz (Mai and Walther 2000), and the fossiliferous horizon
from unit (3) contains a flora from the earliest Oligocene floristic complex Haselbach
(Kunzmann 2012). Between these two complexes an important floristic turnover from
predominantly notophyllous evergreen broad-leaved vegetation to mixed evergreen
deciduous vegetation could be observed (Kvaček & Walther 2001, Kvaček et al. 2014). This
change coincides with important climatic changes, i.e. temperature drop from “subtropical” to
warm-temperate (Kvaček et al. 2014), and decreasing palaeo-pCO2 values in the
atmosphere (Steinthorsdottir et al. 2016). In short, here a regional vegetation change is
recognized as a response to global change at the Eocene-Oligocene turnover (Fig. 5).
Late Eocene megafloras. Fossil floras from the late Eocene Borna Formation [unit (2)]
belong to the Zeitz floristic complex (Fig. 6; Mai & Walther 1985, 2000; Hennig & Kunzmann
2013; Ferdani 2014). This complex covers a considerable period of ca. 3 Ma from ~ 38.5 –
35 Myr (Text-fig. 1; Moraweck et al. 2015). The respective time slice is characterized by a
global cooling trend detected from the marine isotope record (Zachos et al. 2008) from nearly
‘subtropical’ to warm-temperate conditions. At the moment this trend could not be traced
adequately by various macrofloras in this area. Moraweck et al. (2015) concluded that floras
of the Zeitz complex are most likely unsuitable to detect gradual climatic changes based on
the limited species diversity in the individual taphocoenoses and high fragmentation rates of
the fossil plant material, which lowers the significance of the derived results by quantitative

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
58
palaeoclimate reconstruction methods. However, recent phytosociological investigations on
late Eocene macrofloras from Northern Bohemia and central Germany (Kvaček 2010;
Kvaček et al. 2014; Teodoridis and Kvaček 2015; Kunzmann et al. in press) provide hints
about gradual restructuring of late Eocene vegetation in central Europe that is most likely
caused by climate change.
Plant assemblages from about 35 floras of the Zeitz complex represent primarily riparian
vegetation of mainly azonal environments and to minor extent swamp forests of floodplains
and pioneer vegetation in highly disturbed habitats such as river banks, and mesophytic
forests. Kvaček (2010) named the vegetation type notophyllous evergreen broad-leaved
(riparian) forest with palms.
In more detail, Mai and Walther (2000) reconstructed for the Zeitz floristic complex several
azonal vegetation units, i.e. the Steinhauera-Rhodomyrtophyllum riparian forest, and the
Lauraceae-conifer swamp forests. These plant assemblages are usually predominated by
taxa such as Steinhauera subglobosa, Rhodomyrtophyllum reticulosum and Lauraceae (e.g.
Daphnogene cinnamomifolia, Laurophyllum syncarpifolium), which are associated with
evergreen Eotrigonobalanus furcinervis (Fagaceae), several Theaceae, palms and conifers.
Conifers are represented by Quasisequoia couttsiae (Cupressaceae) - a typical back-swamp
conifer element, and by Doliostrobus taxiformis (Doliostrobaceae), which is more
characteristic for riparian associations. Mai and Walther (2000) also concluded on the
presence of zonal Eotrigonobalanus-oak-Lauraceae forests with conifers, which is a matter
of interpretation.
Several plant taphocoenoses derived from the interbedding horizon between two measures
of the main lignite seam complex (Mai & Walther 2000). Some of these taphocoenoses have
been described recently (Hennig & Kunzmann 2013; Ferdani 2014). The siliciclastic
interbedding horizon consists of alluvial plain sediments (lacustrine, fluvial) and tidal
sediments (Rascher et al. 2008; pers. comm. Gerda Standke, Freiberg/Sa., Germany).
Fruits, seeds, leaves and wood are preserved as coalified remains (compressions, Ferdani
2014) and come from silts and silty clays.
Early Oligocene megaflora. The Haselbach horizon of unit (3) is assigned to the lowermost
part of the Böhlen Formation, recently named the Gröbers Member (Standke et al., 2010). In
particular, the Gröbers Member consists of brackish (tidal), fluvial, lacustrine, and palustrine
sediments (Standke et al., 2010) that in general represent a coastal plain environment during
a transgression setting. The Haselbach horizon is intercalated between a series of tidal
sands and silts and the Böhlen lignite seam complex. In its type area, the horizon consists
primarily of a 10–15-m-thick clay stratum (clay in the technological sense) that has been
mined commercially. However, the Haselbach horizon represents a series of fluvial,

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
59
lacustrine, and palustrine strata that were mainly deposited by anastomosing and
meandering rivers onto a heterogenous floodplain and by floodplain lakes. Abandoned
channel deposits, levee sediments, crevasse splay deposits, and floodplain mudstone are
recognized as common facies types (Kunzmann and Walther 2012). Recently, a split of the
Haselbach horizon into a lower and an upper stratum has been observed in Schleenhain.
The two units are separated from one another by a sand/silt layer. The age of the Haselbach
horizon is most likely earliest Oligocene based on pollen assemblages. Floristically, it is a
post Eocene-Oligocene turnover flora (Steinthorsdottir et al. 2016).
The megafossil plant assemblages were mostly excavated from abandoned channel deposits
at the base of the Haselbach horizon. These abandoned channel sediments are
characterized as generally planar-laminated, fine-grained sand and/or silt that alternate with
silty clay and phytoclasts (Figs. 7). Fruits, seeds, leaves and wood are preserved as coalified
remains (compressions, Kunzmann 2012). The fossil vegetation was characterized as mixed
deciduous-evergreen that were growing under warm-temperate and humid climate. In
particular, waterplant associations, riparian gallery forests, backswamp forests, and
mesophytic forests were reconstructed from the fossil assemblages (Kunzmann and Walther
2012).
In the last decades several taphonomical and phytosociological studies were used for
reconstructing the following fossil plant communities (see Kunzmann and Walther 2012).
(1) Water plant association: Lacustrine deposits of smaller floodplain lakes contain rarer
autochthonous carpo-assemblages such as a Salvinia-Azolla community including Azolla,
Salvinia, Lemna, Eichhornia, Hydrocharis, Ottelia, Stratiotes and Desembaya, and a
submersed-plant community including Aldrovanda, Ceratophyllum, Ludwigia, Potamogeton
and Vallisneria.
(2) Fern–monocot–shrubby angiosperm association from floodplain mud indicating alluviation
of still water: including the ferns Osmunda lignitum and Pronephrium stiriacum and
(deciduous) subshrubs such as Apocynophyllum neriifolium (Kunzmann and Walther 2007)
and Zingiberoideophyllum liblarense (Kunzmann 2012).
(3) Swamp forests (Fig. 8) are often dominated by a single conifer species such as
Quasisequoia couttsiae and Taxodium dubium. Myrica longifolia, several Lauraceae and
evergreen Fagaceae (Eotrigonobalanus furcinervis), palms, and Apocynophyllum neriifolium
also belong to this community that comes from organic-rich clays (Kunzmann and Walther
2012). A Taxodium-Nyssa swamp forest (Kunzmann et al. 2009) was a quite characteristic
association in the coastal lowlands consisting of Taxodium dubium, Nyssa altenburgensis,
Myrica longifolia and Sabal raphifolia.
(4) Riparian or gallery forest (Fig. 9): a softwood is comprised of Populus germanica,
Populus zaddachii, Alnus spp., Salix varians, Salix breunsdorfensis, Liquidambar europaea,

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
60
and Taxodium dubium, whereas Ulmus fischeri, Acer haselbachense, Carpinus grandis and
Platanus neptuni dominate the hardwood type. In more diversified taphocoenoses
Pyracantha kräuselii, Rosa lignitum, Lauraceae and the fern Lygodium kaulfussii come
along.
(5) Communities on sandy river banks have consisted of Pinus spp., Comptonia acutiloba,
Engelhardia orsbergensis, Liquidambar europaea, Pyracantha kräuselii, Pinus spp. and
palms.
(6) The zonal vegetation was represented by a Mixed Mesophytic Forest. This vegetation
unit is often dominated by evergreen species. Eotrigonobalanus furcinervis,
Trigonobalanopsis rhamnoides, Distylium fergusoni, Daphnogene cinnamomifolia,
Laurophyllum acutimontanum, L. pseudoprinceps and Illicium sp. are recognized in the leaf
component of the taphocoenoses. A divers range of deciduous plants such as Acer
haselbachense, Carpinus grandis, Engelhardia orsbergensis, Matudaea menzelii, Platanus
neptuni, and Ulmus fischeri were abundant but less frequent. Conifers including Pinus spp.
Tsuga plicata, Sequoia abietina and Cephalotaxus sp. occurred scattered. Shrubs and lianas
were recognized by Ampelopsis rotundata, Lygodium kaulfussii, Rosa lignitum,
Majanthemophyllum petiolatum.
References
Ferdani, F. 2014. Obereozäne Floren aus dem zentralen Weißelsterbecken (Mitteldeutschland) und
ihre paläoökologische Position. – Altenburger Naturwiss. Forsch. 16, 1-115.
Grein, M., Oehm, C., Konrad, W., Utescher, T., Kunzmann, L., Roth-Nebelsick, A. 2013. Atmospheric
CO2 from the late Oligocene to early Miocene based on photosynthesis data and fossil leaf
characteristics. – Palaeogeogr. Palaeoclimatol. Palaeoecol. 374, 41-51.
Hennig, D. and Kunzmann, L. 2013. Taphonomy and vegetational analysis of a late Eocene flora from
Schleenhain (Saxony, Germany). – Geologica Saxonica 59, 75-88.
Krutzsch, W. 2011. Stratigrafie und Klima des Palaogens im Mitteldeutschen Astuar im Vergleich zur
marinen nordlichen Umrahmung. – Z. dt. Ges. Geowiss. 162(1), 19-46.
Kunzmann, L. 2012. Early Oligocene plant taphocoenosis with mass occurrence of
Zingiberoideophyllum (extinct Musaceae). – Palaios 27, 765-778.
Kunzmann, L., Kvaček, Z., Mai, D.H., Walther, H. 2009. The genus Taxodium (Cupressaceae) in the
Palaeogene and Neogene of Central Europe. – Rev. Palaeobot. Palynol. 153(1-2), 153-183.
Kunzmann, L. and Walther, H. 2002. Eine obereozäne Blätterflora aus dem mitteldeutschen
Weißelster-Becken. – Paläont. Z. 76(2), 261-282.
Kunzmann, L. and Walther, H. 2007. A noteworthy plant taphocoenosis from the Early Oligocene
Haselbach member (Saxony, Germany) containing Apocynophyllum neriifolium Heer (cf.
Lythraceae). – Acta Palaeobot. 47(1), 145-16.

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
61
Kunzmann, L. and Walther, H. 2012. Early Oligocene plant taphocoenoses of the Haselbach
megafloral complex and the reconstruction of palaeovegetation. – Palaeobiodiversity and
Palaeoenvironments 92(3), 295-307.
Kvaček, Z. 2010. Forest flora and vegetation of the European early Palaeogene – a review. – Bull.
Geosci. 85(1), 63-76.
Kvaček, Z. and Walther, H. 2001. The Oligocene of Central Europe and the development of forest
vegetation in space and time on the basis of megafossils. – Palaeontographica Abt. B 259,
125-148.
Mai, D.H. 1995. Tertiäre Vegetationsgeschichte Europas. – 691 pp. (Fischer) Jena, Stuttgart, New
York
Mai, D.H. and Walther, H. 1978. Die Floren der Haselbacher Serie im Weißelster-Becken (Bezirk
Leipzig, DDR). – Abh. Staatl. Mus. Mineral. Geol. Dresden 28, 1-101.
Mai, D.H. and Walther, H. 1983. Die fossilen Floren des Weißelster-Beckens und seiner Randgebiete.
– Hallesches Jb. Geowiss. 8, 59-74.
Mai, D.H. and Walther, H. 1985. Die obereozänen Floren des Weißelster-Beckens (Bezirk Leipzig,
DDR). – Abh. Staatl. Mus. Mineral. Geol. Dresden 33, 1-260.
Mai, D.H. and Walther, H. 2000. Die Fundstellen eozäner Floren NW-Sachsens und des Bitterfelder
Raumes. – Altenburger Naturwiss. Forsch. 33, 3-59.
Meyer, G. 1950. Der Einfluss der geologischen Strukturen im Meuselwitzer–Bornaer
Braunkohlenrevier auf Planung und Abbau. – Freiberger Forsch.-H. 1, 49-51.
Moraweck, K., Uhl, D., Kunzmann, L. 2015. Estimation of late Eocene (Bartonian-Priabonian)
terrestrial palaeoclimate: contributions from megafloral assemblages from central Germany. –
Palaeogeogr. Palaeoclimatol. Palaeoecol. 433, 247-258.
Mosbrugger, V., Utescher, T., Dilcher, D.L. 2005. Cenozoic continental climate evolution of central
Europe. – Proc. Natl. Acad. Sci. 102, 14964–14969.
Rascher, J. Escher, D., Fischer, J. 2008. Zur stratigraphischen Gliederung des obereozänen
Hauptflözkomplexes (Thüringer und Bornaer Hauptflöz) in der Leipziger Bucht. – Z. dt. Ges.
Geowiss. 159(1), 105-116.
Roth-Nebelsick, A., Oehm, C., Grein, M., Utescher, T., Kunzmann, L., Friedrich, J.-P., Konrad, W.
2014. Stomatal density and index data of Platanus neptuni leaf fossils and their evaluation as
a CO2 proxy for the Oligocene. – Rev. Palaeobot. Palynol. 206, 1-9.
Standke, G. 2008a. Tertiär. – In: Pälchen, W., WALTER, H. (eds) Geologie von Sachsen. Geologischer
Bau und Entwicklungsgeschichte. – pp. 358-419 (E. Schweizerbart’sche
Verlagsbuchhandlung) Stuttgart,
Standke, G. 2008b. Paläogeografie des älteren Tertiärs (Paläozän bis Untermiozän) im
mitteldeutschen Raum. – Z. dt. Ges. Geowiss. 159/1, 81-103.
Standke, G., Escher, D., Fischer, J., Rascher, J. 2010. Das Tertiär Nordwestsachsens. Ein
geologischer Überblick. – 156 pp. (Sächsisches Landesamt für Umwelt, Landwirtschaft und
Geologie) Freiberg.

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
62
Steinthorsdottir, M., Porter, A., Holohan, A., Kunzmann, L., Collinson, M. E., McElwain, J. C. (2015):
Fossil plant stomata indicate decreasing atmospheric CO2 prior to the Eocene–Oligocene
boundary. – Clim. Past 12, 439-454.
Teodoridis, V. and Kvaček, Z. 2015. Palaeoenvironmental evaluation of Cainozoic plant assemblages
from the Bohemian Massif (Czech Republic) and adjacent Germany. – Bull. Geosci. 90(3),
695-720.
Walther, H. and Kunzmann, L. 2008. Zur Geschichte der paläobotanischen Forschung im
Weißelsterbecken. – Z. dt. Ges. Geowiss. 159(1), 13-21.
Zachos, J.C., Dickens, G.R., Zeebe, R.E. 2008. An early Cenozoic perspective on greenhouse
warming and carbon-cycle dynamics. – Nature 451, 279–283.

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
63
Figures
Fig. 1: Distribution of Paleogene sediments in central Germany – the Weißelster Basin (southern part
of the Leipzig embayment), arrow: Vereinigtes Schleenhain opencast min (from Standke 2008a).

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
64
Fig. 2: Distribution of marine Oligocene sediments in northern central Europe, maximum extension of
the “Rupelian North Sea”, arrow: the Leipzig embayment (from Standke 2008a).

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
65
Fig. 3: Central German mining district Weißelster Basin with two active opencast mines (Profen and
Vereinigtes Schleenhain) of the MIBRAG company (from www.mibrag.de).

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
66
Fig. 4: General lithostratigraphic chart of the Paleogene and Neogene in central Germany, arrows
indicate the position of the fossiliferous horizons in the Vereinigtes Schleenhain opencast mine, i.e.
main interbedding horizon of lignite seam complex 2/3 and Haselbach Clay complex (from Standke
2008a).

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
67
Fig. 5: Two leaf assemblages from the Weißelster Basin: the late Eocene assemblage Stedten of the
Zeitz floristic complex (left, from Mai 1995) and the early Oligocene assemblage Schleenhain of the
Haselbach floristic complex (right, from Kunzmann and Walther 2012).

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
68
Fig. 6 (next page): Predominant, key and accessory taxa of the floras of the Zeitz floristic complex in
the Cenozoic of the Weißelster Basin (central Germany) and in North Bohemian basins (Czech
Republic) (from Kunzmann et al. in press).
6/1 Doliostrobus taxiformis (Sternb. 1833) Kvaček 1971, foliage shoot, Klausa, Germany, MMG
PB Kl 11.
6/2 Rhodomyrtophyllum reticulosum (Rossm. 1840) Knobloch & Kvaček in Knobloch, Kvaček &
Konzalová 1996, leaf with characteristic emarginated apex, Schleenhain 1, Germany, MMG PB
SchleOE 755 (orig. Hennig & Kunzmann 2013: pl. 1, fig. F; orig. Moraweck et al. 2015: fig. 3I).
6/3 Polyspora saxonica Walther & Kvaček in Kvaček & Walther 1984, leaf, Klausa, Germany,
MMG PB Kl 423a (orig. Mai & Walther 1985: pl. 24, fig. 6; orig. Moraweck et al. 2015: fig. 3D).
6/4 Daphnogene cinnamomifolia (Brongn. in Cuvier 1822) Unger 1850, leaf, Staré Sedlo
Formation, Czech Republic, MMG PB Ss 579.
6/5 Trigonobalanopsis rhamnoides (Rossm. 1840) Kvaček & Walther 1988, leaf, Staré Sedlo
Formation, Czech Republic, MMG PB Ss 35 (orig. Knobloch et al. 1996: pl. 23, fig. 4).
6/6 Steinhauera subglobosa C. Presl in Sternb. 1838, two infructescences (left: surface view,
right: in cross section), Schleenhain 1, Germany, MMG PB SchleOE 709:1a, b (orig. Hennig &
Kunzmann 2013: pl. 1, fig. E; orig. Moraweck et al. 2015: fig. 3J).
6/7 Platanus neptuni (Ettingsh. 1866) Bůžek, Holý & Kvaček 1967, compound leaf, Kučlín, Czech
Republic, MMG PB Kin 84 (orig. Kvaček & Manchester 2004: fig. 7b, orig. Kvaček & Teodoridis 2011:
pl. 5, fig. 10).
6/8 Eotrigonobalanus furcinervis (Rossm.1840) Walther & Kvaček in Kvaček & Walther 1989, leaf,
Schleenhain 1, Germany, MMG PB SchleOE 720e (orig. Hennig & Kunzmann 2013: pl. 1, fig. A; orig.
Moraweck et al. 2015: fig. 3A).
6/9 Sabal raphifolia (Sternb. 1822) Knobloch & Kvaček in Knobloch, Kvaček & Konzalová 1996,
leaf, Schleenhain 2, Germany, MMG PB SchleOE 1:1 (orig. Moraweck et al. 2015: fig. 3K).
6/10 Laurophyllum (“Neolitsea”) syncarpifolium (Friedrich 1883) Wilde 1989 f. gardneri Kvaček in
Knobloch, Kvaček & Konzalová 1996, leaf, Profen-Süd 1, Germany, MMG PB Pf 297g.
6/11 "Laurophyllum“ fischkandelii L. Kunzmann & Walther 2002, leaf, Profen-Süd 1, Germany,
MMG PB Pf 1379a.

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
69

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
70
Fig. 7: Typical bedding plane with leaf compressions from an abundoned channel fill deposit from the
Haselbach Clay complex, Gröbers Member (from Kunzmann and Walther 2012).

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
71
Fig. 8: Vegetation scheme of a typical early Oligocene Quasisequoia couttsiae swamp forest (modified
from Kunzmann and Walther 2012).
Fig. 9: Vegetation scheme and taxa of a typical early Oligocene riparian softwood and hardwood
riparian forest from the Weißelster Basin (unpublished conference contribution).

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
72
Practical palaeontological field work part 2
Middle Miocene sealevel changes documented in plant taphocoenesis in the
2nd Lusatian lignite complex in East Germany
Franziska Ferdani & Lutz Kunzmann
During the field trip the Vattanfall Europe Mining opencast mine Welzow-Süd in the
Bundesland Brandenburg will be visited. The characteristic succession of plant
taphocoenoses within a middle Miocene lignite seam complex in relation to sealevel changes
and sedimentation processes in a coastal plane will be the focal point in the field. The
following pages contain useful diagrams and maps as an introduction to the geology,
stratigraphy and paleobotany of the East German mining area Lausitz (Lusatia).
Some important references:
Autorenkollektiv (2010): Die Geologische Entwicklung der Lausitz. – 193 pp. (Vattenfall
Europe Mining AG) Cottbus, Germany.
Dolezych, M. & Schneider, W. (2006): Inkohlte Hölzer und Cuticulae dispersae aus dem 2.
Miozänen Flözhorizont im Tagebau Welzow (Lausitz) – Taxonomie und
vergleichende feinstratigraphisch-fazielle Zuordnung. – Z. geol. Wiss., 34 (3-4): 165-
259; Stuttgart.
Dolezych, M. & Schneider, W. (2007): Taxonomie und Taphonomie von Koniferenhölzern
und Cuticulae dispersae im 2. Lausitzer Flözhorizont (Miozän) des Senftenberger
Göthel, M. (2004): Stratigraphie des Känozoikums in Brandenburg mit spezieller
Berücksichtigung des Braunkohlenreviers Lausitz. – Brandenburgische Geowiss.
Beitr., 11 (1-2): 149-168; Kleinmachnow.
Mai, D. H. (1999a): Die untermiozänen Floren aus der Spremberger Folge und dem 2.
Flözhorizont in der Lausitz. Teil I: Farnpflanzen, Koniferen und Monokotyledonen. –
Palaeontographica Abt. B, 250: 1-76; Stuttgart.
Mai, D. H. (1999b): Die untermiozänen Floren aus der Spremberger Folge und dem 2.
Flözhorizont in der Lausitz Teil II. - Palaeontographica Abt. B, 251: 1-70; Stuttgart.
Mai, D. H. (2000a): Die untermiozänen Floren aus der Spremberger Folge und dem 2.
Flözhorizont in der Lausitz. Teil IV: Fundstellen und Paläobiologie. –
Palaeontographica Abt. B, 254: 65-176; Stuttgart.
Mai, D. H. (2000b): Die mittelmiozänen und obermiozänen Floren aus der Meuroer und
Raunoer Folge in der Lausitz Teil I. – Palaeontographica Abt. B, 256: 1-68; Stuttgart.

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
73
Mai, D. H. (2001a): Die mittelmiozänen und obermiozänen Floren aus der Meuroer und
Raunoer Folge in der Lausitz Teil II. – Palaeontographica Abt. B, 257: 35-174;
Stuttgart.
Mai, D. H. (2001b): Die mittelmiozänen und obermiozänen Floren aus der Meuroer und
Raunoer Folge in der Lausitz Teil III. – Palaeontographica Abt. B, 258: 1-85; Stuttgart.
Nowel, W., Bönisch, R., Schneider, W. & Schulze, H. (1995): Geologie des Lausitzer
Braunkohlenreviers, 2. Aufl. – 104 pp. (Lausitzer Braunkohlengesellschaft)
Senftenberg, Germany.
Roselt, G. & Schneider, W. (1969). Cuticulae dispersae, ihre Merkmale, Nomenklatur und
Klassifikation. – Paläont. Abh., B III (1): 1-128; Berlin.
Schneider, W. (1966): Beziehungen zwischen Pflanzeninhalt und petrographischer
Beschaffenheit von Weichbraunkohlen am Beispiel der miozänen Braunkohlen der
Oberlausitz. – Ber. deutsch. Ges. geol. Wiss., A Geol. Paläont., 11 (5): 615-633;
Berlin.
Schneider, W. (1992): Floral Successions in Miocene swamps and bogs of Central Europe. –
Z. geol. Wiss., 20 (5/6): 555-570; Berlin.
Schneider, W. (1995): Palaeohistological studies on Miocene brown coals of Central Europe.
– Int. J. Coal Geol., 28 (2-4): 229-248; Amsterdam.
Schneider, W. (2004): Eine blätterführende Taphocoenose im 2. Miozänen Flöz von Nochten
(Lausitz): Taxonomie, Taphonomie und Phytostratigraphie. – Palaeontographica Abt.
B, 268 (1-3): 1-74; Stuttgart.
Schneider, W. (2007): Magnolia L. in peat-forming associations of the Miocene seams in
Lower Lusatia (East Germany). – Acta Palaeobotanica, 47 (1): 217-235; Kraków.
Standke, G. (2006): Paläogeographisch-fazielle Modellierung des Unter-/Mittelmiozän-
Grenzbereiches in der Lausitz (Briesker Folge/Formation). – Schriftenreihe für
Geowissenschaften, 14: 1-130; Berlin.
Standke, G. mit einem Beitrag von P. Suhr (2008): Tertiär. – In: Pälchen, W. & Walter, H.
(Hrsg.): Geologie von Sachsen. Geologischer Bau und Entwicklungsgeschichte. –
358-419 (E. Schweizerbart’sche Verlagsbuchhandlung) Stuttgart.
Standke, G., Rascher, J. & Strauss, C. (1993): Relative sea-level fluctuations and brown coal
formation around the Early-Middle Miocene boundary in the Lusatian Brown Coal
District. – Geol. Rundschau, 82: 295-305; Stuttgart.

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
74
Fig. 10: General lithostratigraphic chart of the Paleogene and Neogene in central Germany, arrows
indicate the position of the fossiliferous horizons in the Vereinigtes Schleenhain opencast mine, i.e.
main interbedding horizon of lignite seam complex 2/3 and Haselbach Clay complex (from Standke
2008a).

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
75
Fig. 11: East German mining area Lausitz (Lusatia) of the Vattenfall Europe Mining AG.

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
76
Fig. 12: Distribution of the lower Miocene sediments of the Drebkau Member (Brieske Formation), blue
coloured = distribution of marine sediments, brown coloured = distribution of the complex of
underlaying sediments (from Autorenkollektiv 2010).

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
77
Fig. 13: Figure above: Distribution of the middle Miocene sediments of the Welzow Member (Brieske
Formation) included the 2nd
Miocene lignite complex; Figure below: Distribution of regional
interburdens from the 2nd
Miocene lignite complex (from Autorenkollektiv 2010).

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
78
Fig. 14: Distribution of the middle Miocene sediments of the Greifenhain Member (Meuro Formation),
blue coloured = distribution of mainly marine sediments, brown coloured = distribution of the complex
of overlaying sediment, yellow coloured = supposed distribution of the “Sands of Seese” (from
Autorenkollektiv 2010).

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
79
Fig 15. Lithostratigraphic correlation chart of the Lausitzer (Lusatian) Paleogene and Neogene. Global
sea level changes and local sediment records (from Autorenkollektiv 2010).

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
80
Fig. 16: Model of successions of plant taphocoenosis wothin the middle Miocene 2nd
Lignite Seam
Complex of the Lausitz, east Germany (Schneider in Nowell et al. 1995).

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
81
Useful information

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
82

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
83

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
84

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
85

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
86

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
87

International Symposium, Dresden, April 02–06 2016 __________________________________________________________________________________________________________________________________________________________________
88

The Yunnan Biodiversity Hotspot Project __________________________________________________________________________________________________________________________________________________________________
89