the subeng vertebrate tracks: stratigraphy,...
TRANSCRIPT

Bollettino della Società Paleontologica Italiana, 56 (2), 2017, 181-198. Modena
ISSN 0375-7633 doi:10.4435/BSPI.2017.12
The Subeng vertebrate tracks: stratigraphy, sedimentology and a digital archive of a historic Upper Triassic palaeosurface (lower Elliot Formation),
Leribe, Lesotho (southern Africa)
Emese M. Bordy, Miengah ABrAhAms & Lara sciscio
E.M. Bordy, Department of Geological Sciences, University of Cape Town, University Avenue 13, ZA-7701 Cape Town, South Africa; [email protected]
M. Abrahams, Department of Geological Sciences, University of Cape Town, University Avenue 13, ZA-7701 Cape Town, South Africa; [email protected]
L. Sciscio, Department of Geological Sciences, University of Cape Town, University Avenue 13, ZA-7701 Cape Town, South Africa; [email protected]
KEY WORDS - 3D digital model, dinosaur, cynodont, southern Africa, palaeoenvironment, trace fossils.
ABSTRACT - Dinosaur vertebrate body and ichnofossils are relatively abundant in the Upper Triassic-Lower Jurassic Elliot Formation (Stormberg Group, Karoo Supergroup) in the main Karoo Basin in southern Africa. Herein we present the results of our sedimentological, stratigraphical and ichnological investigations at a historic ichnosite in NE Lesotho that is among the first documented vertebrate track-bearing palaeosurfaces in southern Gondwana. After decades of neglect, the Subeng ichnosite is restudied in this paper in light of the advances in ichnological methods and the formalised stratigraphy of the Stormberg Group. Documentation of this ichnosite was conducted using a mix of field-based sedimentological and ichnological methods and photogrammetry, which collectively allowed us to place the site within the lowermost Elliot Formation (Upper Triassic). Our detailed palaeoenvironmental reconstruction, based on palaeocurrent measurements and sedimentary facies analysis of the host rocks, suggests that this diverse Upper Triassic assemblage of vertebrate ichnofossils, which is an integral part of the history of life on land just before the end-Triassic mass extinction event, formed near a shallow oxbow lake or drying up watering hole on the floodplain of a meandering river system. This water source attracted numerous bipedal and quadrupedal animals as attested by the presence of tridactyl, tetradactyl and pentadactyl tracks on the palaeosurface. We have also produced a photogrammetric digital 3D model (available online) of the Subeng ichnosite to serve as a digital replica and archive for further ichnological study. This digital documentation is important not only for the preservation of this rapidly eroding palaeosurface (situated in an active streambed), but also for the provision of a visually stimulating tool for community outreach and science education in rural African communities.
RIASSUNTO - [Le impronte di vertebrati di Subeng: stratigrafia, sedimentologia ed archivio digitale di una paleosuperficie del Triassico Superiore di interesse storico (parte inferiore della Formazione di Elliot), Leribe, Lesotho (Africa meridionale)] - Resti fossili, impronte e piste di dinosauro sono relativamente abbondanti nella Formazione di Elliot (Triassico Superiore-Giurassico Inferiore; Gruppo di Stormberg, Supergruppo del Karoo; Africa meridionale). In questo studio presentiamo i risultati di ricerche sedimentologiche, stratigrafiche ed icnologiche sull’icnosito di Subeng (coordinate GPS 28°49’23.53”S, 28°4’29.82”E). Si tratta di un icnosito di fondamentale importanza situato a nordest del villaggio di Hlotse (Distretto di Leribe) in Lesotho, rappresentato da una paleosuperficie attribuibile alla parte inferiore della Formazione di Elliot. Questa superficie è esposta ed attivamente erosa dall’azione del torrente Subeng. Dal punto di vista storico, l’icnosito di Subeng rappresenta una delle prime superfici ad impronte di vertebrati ad essere state documentate in Africa meridionale. Tuttavia, dopo la prima perizia nel 1955 ed il successivo studio nei primi anni ‘70, l’icnosito ha ricevuto poca attenzione. Le revisioni icnotassonomiche delle icniti di Subeng si sono concentrate su pochi calchi realizzati a partire da materiale originale. Pertanto, era necessario rivalutare l’icnosito di Subeng, specialmente alla luce degli avanzamenti metodologici in icnologia e dell’aggiornamento della stratigrafia del Gruppo di Stormberg.
Lo studio dell’icnosito di Subeng è stato realizzato combinando metodi fotogrammetrici con metodi icnologici e sedimentologici tradizionali. Sulla base della sedimentologia e dei resti fossili di vertebrati, collochiamo l’icnosito di Subeng nel Triassico Superiore. Più precisamente, collochiamo l’icnosito nella porzione superiore della parte inferiore della principale unità arenitica della Formazione di Elliot, ed a meno di 15 metri dalla Formazione di Molteno. Viene fornita anche una dettagliata ricostruzione paleoambientale del sito, coadiuvata dalla misurazione delle paleocorrenti e dall’analisi di facies. Il primo approccio indica una corrente diretta approssimativamente verso E-NE, mentre il secondo indica che la paleosuperficie si colloca al culmine di una sequenza di fining upward, tipica dei fiumi a meandri. Questo contesto sedimentologico è tipico per una superficie ad impronte. La principale caratteristica icnologica dell’icnosito di Subeng è la presenza di prominenti impronte pentadattile, tetradattile e tridattile. È importante sottolineare che tre piste sono presenti sulla superficie attualmente esposta, la più lunga delle quali è costituita da 14 grandi impronte pentadattile; queste indicano che l’autore della pista si muoveva verso sud. Due piste tetradattile indicano che due animali quadrupedi, possibilmente un esemplare adulto e uno giovane, camminavano uno a fianco all’altro verso est. Impronte tridattile di bipedi, sporadicamente distribuite sulla paleosuperficie, sono attribuite ad animali di dimensioni comparabili; queste non formano piste. Sui ripple marks asimmetrici e tra le fratture di disseccamento si osservano comunemente tracce di invertebrati, preservate come tane superficiali con parziale backfill.
Sulla base delle caratteristiche sedimentologiche, osteologiche ed icnologiche, il paleoambiente viene interpretato come una lanca poco profonda oppure una pozza che fungeva da abbeveratoio, presente in un’ampia piana alluvionale di un sistema fluviale a meandri. Questa fonte d’acqua temporanea attirava numerosi animali bipedi e quadrupedi, sia erbivori (cinodonti e sauropodomorfi) che carnivori.
Il modello fotogrammetrico 3D costituisce una durevole replica digitale dell’icnosito ed un archivio per futuri studi icnologici. Questa documentazione digitale è importante non solo per la preservazione della paleosuperficie (il moderno torrente Subeng la erode intensamente, causando una rapida ed inevitabile perdita di dettaglio icnologico) ma anche per fornire alle comunità rurali africane uno strumento visuale stimolante per l’educazione scientifica. In questa zona persiste una cronica mancanza di risorse per l’educazione scientifica. Questo fenomeno si traduce in un limitato accesso a risorse quali materiale educativo, economico o gratuito, e musei di scienze naturali. Ci auguriamo che il modello digitale renda la superficie di Subeng più visibile sia alla scala locale che globale, e che permetta alla comunità locale di esibire l’importanza scientifica del sito durante le loro attività geoturistiche.
S. P. I.
SOC
IETA
' P
A
LEON TO L OGICA I T
AL
IANA
Bollettino della Società Paleontologica Italiana, 56 (2), 2017182
INTRODUCTION
The Subeng ichnosite in the Upper Triassic portion of the lower Elliot Formation exposes a variety of trace fossils of vertebrates and invertebrates as well as
sedimentary rocks suitable for the palaeoenvironmental reconstruction and stratigraphical analysis (Figs 1-3). The Subeng ichnotaxa mentioned in the literature (e.g., Ellenberger, 1955, 1970, 1972; Ellenberger & Ellenberger, 1958) are summarized in Fig. 2 and Tab. 1. The ichnosite
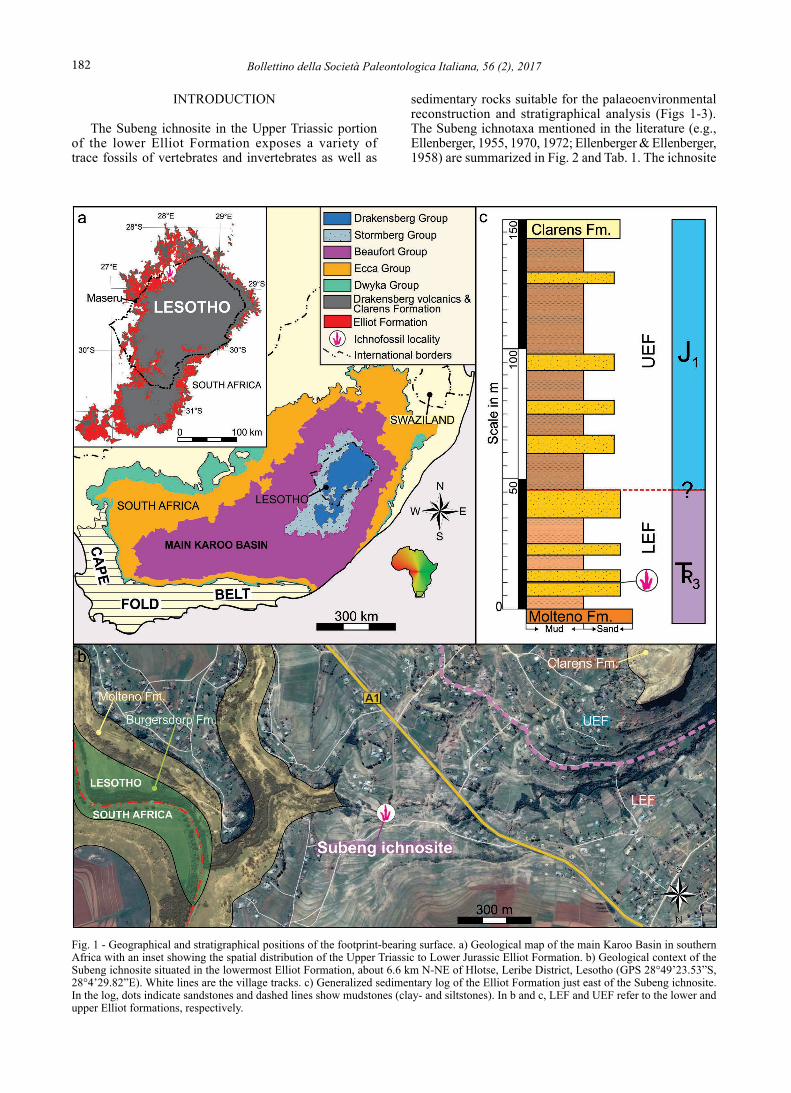
Fig. 1 - Geographical and stratigraphical positions of the footprint-bearing surface. a) Geological map of the main Karoo Basin in southern Africa with an inset showing the spatial distribution of the Upper Triassic to Lower Jurassic Elliot Formation. b) Geological context of the Subeng ichnosite situated in the lowermost Elliot Formation, about 6.6 km N-NE of Hlotse, Leribe District, Lesotho (GPS 28°49’23.53”S, 28°4’29.82”E). White lines are the village tracks. c) Generalized sedimentary log of the Elliot Formation just east of the Subeng ichnosite. In the log, dots indicate sandstones and dashed lines show mudstones (clay- and siltstones). In b and c, LEF and UEF refer to the lower and upper Elliot formations, respectively.

183E.M. Bordy et alii - Upper Triassic vertebrate tracks and trackways at Subeng in Lesotho, southern Africa
is located in the Subeng (Seobeng) stream bed (GPS 28°49’23.53”S, 28° 4’29.82”E), about 6.6 km N-NE of Hlotse (Leribe District, Lesotho; Fig. 1).
The Subeng ichnosite is among the first documented vertebrate footprint palaeosurfaces in southern Africa, having been briefly described and sketched for the first time by its discoverer, Paul Ellenberger in 1955 (“dalle de Subeng”) (Fig. 2). Subsequently, individual Subeng tracks and trackways have been documented by Ellenberger & Ellenberger (1958) and Ellenberger (1970, 1972) (Fig. 2; Tab. 1), but a complete illustration of all the tracks on the surface was never produced. More recently, D’Orazi Porchetti & Nicosia (2007) described the ichnotaxonomy of some of the Subeng ichnites (e.g., based on the direct examination track replicas deposited by P. Ellenberger in the Laboratoire de Paléontologie of the University of Montpellier II). It is noteworthy that just like Olsen & Galton (1984) and Lockley et al. (2006), D’Orazi Porchetti & Nicosia (2007) too have only studied the Lesotho tracks from significant but non-primary material (i.e., only drawings in the former study and plaster cast replicas in the latter two studies). While we genuinely acknowledge the absolute value of these ichnotaxonomic revisions, in our view, their study materials are problematic for ichnotaxonomic work, because they rely on incomplete
material that does not have the detail visible in the actual specimen. Furthermore, the potential human error associated with mislabelled or miscataloged casts and drawings (see below) introduces additional complications. Furthermore, ichnological assessment (that includes but goes beyond sound ichnotaxonomy) aimed at the understanding of a trackmaker’s behaviour, in the palaeoenvironmental milieu, is impossible without the accurate documentation of the track- and trackway-bearing palaeosurfaces in their original field context. Such contextual analysis is necessary for morphometric and shape-difference analyses, and biomechanics.
The aims of this paper are: 1) provide a long overdue palaeoenvironmental context for the Subeng site; 2) revise its position within the southern Africa stratigraphy; 3) generate a digital 3D model of this historic footprint-bearing palaeosurface, from which a distinctive ichnotype (holotype) had been identified. The latter will aid further work related to the ichnotaxonomy of the tracks and trackways, the behaviour and biomechanics of the various trackmakers, and the Subeng community-driven geotourism. The community is actively and enthusiastically involved in geotouristic promotion of the site, but not in the preservation of the palaeosurface due to lack of resources, and more specifically because it is
Fig. 2 - Outline drawings of the Subeng tracks. Six ichnotaxa were described from the site and illustrated in Ellenberger (1970, 1972); see Tab. 1 for details. a) Redrawing of the first illustration of the Subeng ichnosite (“dalle de Subeng”) published by Ellenberger in 1955. Redrawings of the first illustrations of the Subeng trackways published by Ellenberger & Ellenberger (1958) are as follows: b) Trackway E, c) Trackway D, d) Trackway I and e) Trackway H. Note that the original trackway labels of Ellenberger & Ellenberger (1958) are kept for ease of cross-reference.

Bollettino della Società Paleontologica Italiana, 56 (2), 2017184
situated in an active riverbed in a subtropical, seasonally-flowing Subeng stream. We used the 3D model in this work (Supplementary data at https://dx.doi.org/10.6084/m9.figshare.4737535) to document our observations of the physical parametres of the tracks and trackways as well as their spatial relationship. The current contribution forms part of our ongoing research program dedicated to the stratigraphic revision of vertebrate track distribution in the Upper Triassic-Lower Jurassic Stormberg Group of southern Africa.
GEOLOGICAL BACKGROUND
The Subeng ichnosite is surrounded by clastic sedimentary rocks of the Upper Triassic-Lower Jurassic Stormberg Group, which forms part of the Karoo Supergroup in southern Africa and contains the Molteno, Elliot and Clarens formations (Fig. 1). The Molteno Formation is a coal-bearing, wedge-shaped siliciclastic unit with a maximum thickness of about 600 m in the south and minimum thickness of ≤ 10 m in the north (Johnson et
Ichnotaxa
Trackmaker animal Illustration Primary reference (figure/plate numbers)Original name Current
ichnogenera
Pentasauropus incredibilis (Ellenberger, 1970)
Pentasauropus
(D’Orazi Porchetti &
Nicosia, 2007)
Quadrupedal or facultative biped: ?pseudosuchian;
?theromorph; ?large amphibian;
?prosauropod; ?dicynodont
- Fig. 52 A in Ellenberger, 1970
- Trackway I in Ellenberger & Ellenberger, 1958
- Planche I and XIV in Ellenberger, 1972
Deuterosauropodus minor (Ellenberger, 1970)
Invalid, undertrack -
(D’Orazi Porchetti &
Nicosia, 2007)
Quadrupedal: degenerate chirotherian;
?pseudosuchian; a small ?sauropod;
?sauropodomorpha Quadrupedal:
degenerate chirotherian; ?pseudosuchian; a small ?sauropod;
?sauropodomorpha
- Fig. 51 in Ellenberger, 1970
- Trackway H in Ellenberger & Ellenberger, 1958
- Planche XIII in Ellenberger, 1972
Deuterotrisauropus socialis (Ellenberger, 1970)
Grallator
(Olsen & Galton, 1984)
Tridactyl bipedal: ?saurischians
Fig. 44 in Ellenberger, 1970
Planche XIII in Ellenberger, 1972
Mafatrisauropus errans (Ellenberger, 1970)
indeterminate
(Olsen & Galton, 1984)
Tridactyl bipedal: ?saurischians Fig. 45 in Ellenberger, 1970
Pseudotrisauropus subengensis (Ellenberger, 1970) or Psilotrisauropus subengensis (Ellenberger, 1972)
indeterminate
(Olsen & Galton, 1984)
Tridactyl bipedal: ?saurischians
- Fig. 49 A Ellenberger, 1970
- Trackway E in Ellenberger & Ellenberger, 1958
- Planche XII in Ellenberger, 1972
Pseudotrisauropus dieterleni (Ellenberger, 1970)
indeterminate
(Olsen & Galton, 1984)
Tridactyl bipedal: ?saurischians
- Fig. 49 B in Ellenberger, 1970
- Trackway D in Ellenberger & Ellenberger, 1958
- Planche XII and XIII in Ellenber-ger, 1972
Tab. 1 - Summary of the Subeng ichnotaxa identified and depicted by Ellenberger (1970, 1972). All scale bars are equal to 5 cm.

185E.M. Bordy et alii - Upper Triassic vertebrate tracks and trackways at Subeng in Lesotho, southern Africa
al., 2006). Deposition occurred in broad, shallow braided rivers and extensive floodplains under cool and wet palaeoclimatic conditions as suggested by its abundant Carnian fossil plant and insect assemblages (Anderson et al., 1998). While bone fossils are extremely rare (to date, only fossil fishes have been found), Grallator-like footprints, a possible chelonian track and a tail mark of unknown vertebrate have been recorded from two localities in the Formation (Raath et al., 1990; Raath, 1996; Anderson et al., 1998).
The Elliot Formation is a succession of red sandstone and mudrock that reaches a maximum thickness of about 550 m in the south, near Elliot and thins progressively to the north, where the minimum thickness is < 50 m (Bordy et al., 2004a, b; Bordy & Eriksson, 2015). It has an unconformable lower contact with the underlying Molteno and gradual, conformable contact with overlying Clarens formations (Bordy et al., 2005). Based on sedimentary and geochemical data, the deposition occurred under increasing aridity, initially in high-energy, mixed-load meandering rivers and laterally continuous floodplains, and then in ephemeral fluvial systems with flash floods (Bordy et al., 2004a, b; Sciscio & Bordy, 2016). Although the age of the Elliot Formation is generally accepted to be Upper Triassic to Lower Jurassic (Olsen & Galton, 1984; Lucas & Hancox, 2001; Knoll, 2004), unfortunately, the absolute age dates for the Elliot Formation are lacking, and our geochronological dating of the detrital zircon samples collected from the ichnosite are not yet available. The Elliot Formation therefore straddles the Triassic-Jurassic boundary (Fig. 1c) and contains a rich terrestrial vertebrate body and trace fossil record from before and after the end-Triassic biotic crisis (Bordy & Eriksson, 2015; Sciscio et al., in press).
The Lower Jurassic Clarens Formation represents the last stages of sedimentation in the main Karoo Basin before the outpouring the voluminous continental flood basalts and intrusion of the associated subvolcanic complexes about 183±1 Ma ago (Karoo Large Igneous Province - Duncan et al., 1997; Johnson et al., 2006). The deposition of the yellowish-brown, fine- to medium-grained Clarens (previously Cave) sandstones occurred in a wet desert environment that was associated with playa lakes, sheet floods and ephemeral streams. In spite of the overall arid conditions, the palaeobotanical, osteological and ichnological records show that the Clarens desert was capable of supporting a relatively diverse ecosystem of plants, conchostracans, insects, lungfish, bony fish, synapsids and dinosaurs (Kitching & Raath, 1984; Bordy, 2008).
In addition to the ichnofossils (Tab. 1, Figs 1-3), vertebrate fossil remains have also been found in the lower Elliot Formation in the Subeng (aka Leribe-Subeng) area, which is the type locality of Basutodon ferox (Huene, 1932) that has been identified from a large tooth (type specimen: NMB R.610). This tooth and its taxonomic affinities have not been well-studied, and it may possibly belong to a theropod or an archosauriform (rauisuchian archosaur) taxa (Knoll, 2004). Other bone fossils from the vicinity of the ichnosite have been assigned to sauropodomorphs (Melanorosaurus sp.) (Haughton, 1924), “pseudosuchians” and cynodonts
(“Belesodon leribeensis”) (Kitching & Raath, 1984). The latter taxon has been restudied by Battail (2005) who assigned it to a large traversodont cynodont, Scalenodontoides macrodontes (Crompton & Ellenberger, 1957), which is exclusive to the lower Elliot Formation in southern Africa. Finally, vertebrate ichnofossils are not only limited to the Elliot Formation near Subeng, because small tetradactyl and pentadactyl tracks and one tetradactyl bipedal trackway have been apparently recovered from the Lower Jurassic Clarens Formation (see tracks M and L and trackway G in Ellenberger & Ellenberger, 1958).
METHODS
Field evidence was collected in the form of macroscopic observations of the host sedimentary rocks in the immediate vicinity as well as few kilometre radius around the Subeng ichnosite. We applied standard field techniques by recording the following: lithology, geometry (including thickness and lateral continuity), primary and secondary sedimentary structures, grain size variations in vertical and lateral directions, colour, and density and spacing of ichnofossils.
We also obtained 15 palaeocurrent measurements from the host rock of the Subeng ichnosite, which in combination with other sedimentological observations, can assist in reconstructing the mean flow direction and magnitude of the palaeocurrents (Tucker, 2011). In our study, we only used the most reliable palaeocurrent indicators, namely the foresets of trough cross-bedded sandstones, because other measurements, such as crest orientation of ripple marks, are known to deviate from the regional (and locally strongest) flow trends (Tucker, 2011; Bordy et al., 2004c). The palaeocurrent data required no additional corrections, because the regional dip of the strata ranges between 5° to 10°. The results are presented in a rose diagram in Fig. 3d.
We also generated a photogrammetric digital 3D model of the Subeng ichnosite, largely using the photogrammetric technique explained in Mallison & Wings (2014). Here, we only highlight some steps that we adopted. Over 300 photographs were taken using a Canon EOS 1200D (Focal length 28 mm, 5184 x 3456 pixel resolution) were automatically stitched together in the Agisoft Photoscan (standard version 1.1.4) software. We manually excluded photographs that were of low quality or contained foreign objects (e.g., foot of photographer) from the input data set, because these introduce errors and reduce the overall quality of the model. Agisoft provides an automatic image quality filter, which examines the sharpness level of the most focused point in a photograph, and all photographs with a quality less than below a certain threshold were removed from further processing. Subsequently, from the photographs, a dense point cloud was generated. The latter is scale-less, therefore an object of known length (e.g., photo with a scale bar on the surface) was used to calibrate the model. Lastly, an “orthomosaic” digital elevation model was applied. Three-dimensional model as well as all the photographs used for generating the model can be downloaded from https://dx.doi.org/10.6084/m9.figshare.4737535.

Bollettino della Società Paleontologica Italiana, 56 (2), 2017186
RESULTS
Stratigraphy and sedimentology of the ichnositeBased on our field investigation of the local geology,
the Subeng ichnosite is located < 15 m above basal contact of the Elliot Formation, within the upper portion of the lowermost major sandstone unit in the lower Elliot Formation (Fig. 1b-c). Locally, the basal contact of the Elliot Formation is about 1575-1590 m above sea level, below which the coarse- to very coarse-grained sandstones of the Molteno Formation are exposed (N of the ichnosite in the Caledon River) both in Lesotho and neighbouring South Africa (Fig. 1b). The upper contact of the Elliot Formation is about 1710-1740 m above sea level in the region, above which the conspicuous, beige to white, fine to medium-grained sandstones of the Clarens Formation occur as cliff forming features (SE and E of the ichnosite; Fig. 1b-c). The contact of the lower and upper Elliot Formation is difficult to identify here due to the lack of semi-continuous outcrops of these two, sedimentologically contrasting units (Bordy et al., 2004b, d). However, based on limited outcrop information (e.g., grain size, sandstone architecture, pedogenic alteration) and the position of the geomorphological break along the slopes that are underlined by the Elliot Formation, it is very tentatively positioned about 1620-1630 m above sea level (Fig. 1b-c).
In our assessment, the coarse- to very-coarse grained, tabular, sheet-like, and mostly planar cross-bedded sandstones of the Molteno Formation cannot be identified at the ichnosite. Conversely, the most common rocks in the immediate vicinity of the ichnosite are about 40-50 cm thick, medium-grained, strongly trough cross-bedded sandstones (Fig. 3a-c) that are found in the lower part of a well-developed upward-fining and thinning succession, which is < 10 m in thickness (Fig. 1c). Localized mudstone-clast conglomerates are confined to < 50 cm wide and < 20 cm thick lenses (Fig. 3f). Towards the top of the about 4-5 m thick succession, the sandstones are medium- to fine-grained, 10-30 cm thick, massive to ripple cross-laminated. In addition to the abundant vertebrate ichnites, the upper bedding planes expose abundant asymmetrical ripple marks and desiccation cracks as well as rare invertebrate traces (Fig. 3e-f). The crest orientation of the ripple marks is E-W and shows flow direction to the N. The sandstones are directly overlain by red and light purple mudstones (Fig. 3g) that lack pedogenic alteration features and appear to be unfossiliferous in the immediate vicinity of the ichnosite.
The locally measured mean palaeoflow direction is from about WSW to about ENE at the Subeng ichnosite,
which is the regional palaeoflow direction in Elliot Formation (Bordy et al., 2004c, d). The low consistency ratio of the palaeoflow data does not match that of meandering rivers but this discrepancy may be related to the small size of the outcrop where foreset measurements were obtained (< 700 m2). The orientation of sandstone outcrops at the Subeng ichnosite did not allow the 3D assessment of sandstone architecture, but nearby sites at a similar elevation have revealed well-developed lateral accretion surfaces indicative of meandering rivers (see Bordy et al., 2004d, fig. 4A). In summary, the dominance of trough cross-bedded, medium-grained sandstones, and pedogenic alteration-free, red and light purple mudstones, as well as the lack of diagnostic upper Elliot Formation facies (e.g., carbonate nodule conglomerates, clast-rich sandstones; Bordy et al., 2004b, c) collectively argue for the Subeng ichnosite as being part of lower Elliot Formation.
Ichnological e-archive - the digital 3D model of the Subeng ichnosite
In our field inspection of the Subeng ichnosite in 2003, 2012, 2015 and 2016, we repeatedly noted that the original surface as described by Ellenberger in 1955 is no longer exposed to its fullest extent (compare Fig. 4a and c with Fig. 4b and d). Furthermore, we noted that the remaining northern part of the surface is being severely eroded by stream action (Fig. 5), which has already visibly removed important ichnological detail (e.g., positive epirelief of invertebrate traces, expulsion rims around digits, depth variation of tracks). In order to preserve the ichnological detail on the remaining palaeosurface, we generated a digital 3D model of the currently exposed surface (Figs 5-6). On the current surface, we were able to detect 33 pes tracks, of which 16 are pentadactyl, 8 are tetradactyl and 10 are tridactyl. Fourteen of pentadactyl tracks form the longest trackway, showing a locomotion direction towards south (Figs 5-6). A partially preserved, second pentadactyl trackway comprises two pentadactyl track (Figs 5-6). The tetradactyl tracks make up two trackways and show movement from west to east (Figs 5-7). The tridactyl tracks (Figs 5-6, 8) do not form any trackways on the currently exposed surface (compare to Fig. 2a).
Subeng tracks and potential trackmakersPentAdActyl trAcks - The most spectacular track at
the Subeng ichnosite is the pentadactyl, crescent-shaped Pentasauropus incredibilis (Figs 2, 4-6; Tabs 1-2), which was discovered by Ellenberger in the 1950s and named in 1970 (Ellenberger, 1970). Because it has recently been reviewed ichnotaxonomically by D’Orazi Porchetti &
Fig. 3 - Sedimentological context of the Subeng ichnosite. Trough cross-bedded sandstone beds (St - a, b & c) are common, especially in the lower part of the upward-fining succession, which contains the track-bearing palaeosurface (see Fig. 1c). a) Trough cross-bedded sandstone (St) in map view. b) Outline of troughs and palaeoflow direction (compare to a). c) Trough cross-bedded sandstone in mainly side view. d) Rose diagram of the trough cross-bedded, medium-grained sandstone beds indicate palaeoflow to ~ENE. Chief sedimentary characteristics of the footprint-bearing palaeosurface in the Subeng stream are listed as follows in e-f. e) Asymmetrical ripple marks (Sr - palaeoflow direction to the north) that are locally bioturbated by shallow, partially backfilled invertebrate traces. f) Desiccation cracks, which initially formed in a thin mud layer and were filled with sand subsequently. The casts of the desiccation cracks adhere to the top of massive sandstone (Sm) that is underlain by massive, mudstone-clast conglomerates (Gcm). g) Light red to purple, massive mudstone (Fm) directly overlies the track-bearing palaeosurface and are conspicuously free of pedogenic alteration features. Note the bulk of the exposure shows the light yellow-pink recent, unconsolidated sediments of the Subeng stream.

187E.M. Bordy et alii - Upper Triassic vertebrate tracks and trackways at Subeng in Lesotho, southern Africa

Bollettino della Società Paleontologica Italiana, 56 (2), 2017188
Fig. 4 - Changes in the size and relief of the Subeng palaeosurface over 40 years in the Subeng stream. a) Photograph from Planche XIV in Ellenberger (1972), modified after Ellenberg (1972). b) Photograph taken by Fabien Knoll in 2012. The upper (southern) part of the Subeng palaeosurface as was seen in the 1970s is now completely obscured (middle area between the concrete bridge and foreground) by recent stream sediments and water. Note the difference in size and distribution of the large boulders in the streambed, which is an indication of the occasional extremely high energy levels of the Subeng stream. c-d) Photographs of the northern part of the Subeng palaeosurface taken over 40 years apart. c) Photograph from Planche I in Ellenberger (1972). d) Part of the photogrammetric image of the same part of the surface. Note the well-developed expulsion rims around the digit impressions in the Pentasauropus tracks in the pre-1972 image, which, have been planated by streambed erosion by 2016. Also see Fig. 4b.

189E.M. Bordy et alii - Upper Triassic vertebrate tracks and trackways at Subeng in Lesotho, southern Africa
Fig. 5 - Ortophoto and interpretive outline drawing of the tridactyl, tetradactyl and pentadactyl ichnites at the Subeng ichnosite. a) Detailed overview of the tracks on a single 21 m2 sandstone slab derived from the 3D digital model of the surface (see https://dx.doi.org/10.6084/m9.figshare.4737535). b) Individual tracks, some of which make up the three trackways (labelled P, T1 and T2; see Fig. 6), are colour coded for clarity: red for pentadactyl tracks; green for tetradactyl tracks and blue for tridactyl tracks. The latter tracks do not form part of any trackway. Note the well-developed current ripple marks (palaeoflow to south) and desiccation cracks in the southwestern and northwestern part of the surface, respectively.

Bollettino della Società Paleontologica Italiana, 56 (2), 2017190
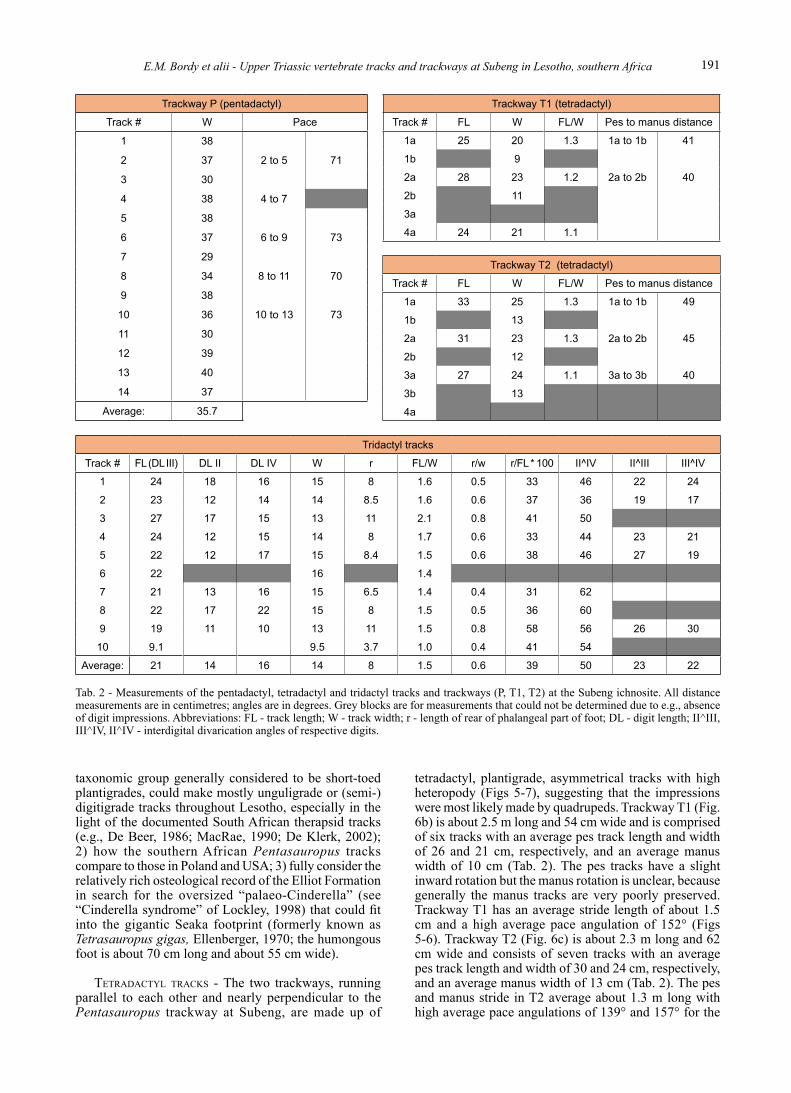
Nicosia (2007), the full description of Pentasauropus will not be repeated here. It should suffice to say that at the Subeng ichnosite, the type locality for Pentasauropus, these unique, pentadactyl tracks currently form a about 3.7 m long trackway (compare to Fig. 2a) comprised of 14 tracks, three of which are only partially preserved (Figs 5-6; Tab. 2). The average width of the complete tracks is 37 cm with strides ranging from 47 to 76 cm and an average pace angulation of 50° (Fig. 6; Tab. 2). Each pair of tracks (e.g., 2-3, 4-5; Fig. 6) is orientated in the same direction. On the preservation scale of Belvedere & Farlow (2016), Pentasauropus at Subeng would fall into the quadruped grade 1 to 2 categories.
These pentadactyl, crescent-shaped, mostly unguligrade or digitigrade and occasionally plantigrade tracks of manus and pes (with rounded sole if preserved) have been attributed to a variety of trackmakers (e.g., prosauropod, pseudosuchian, theromorph, large amphibian, dicynodont - e.g., Ellenberger & Ellenberger, 1958; Ellenberger, 1972). Pentasauropus-like tracks are
worldwide known, with the oldest having been described from the Lower Triassic Wiory Formation (Poland), where these ichnites were labelled as cf. Therapsipus by Klein & Niedzwiedzki (2012) and attributed them to therapsids. Other occurrences of the similar pentadactyl ichnites are mentioned by Lockley & Hunt (1995) and Hunt-Foster et al. (2017) from the Late Triassic Chinle Group of Utah (USA). D’Orazi Porchetti & Nicosia (2007) also argue that dicynodonts are the most plausible trackmakers of Pentasauropus due to the close match between the tracks and their skeletal autopodia. Body fossils of large therapsids have been reported from stratigraphically equivalent strata (i.e., the Upper Triassic lower Elliot Formation) in southern Africa, and from Subeng in particular (e.g., Battail, 2005), but we prefer to be cautious and only discuss the identity of the trackmaker after our investigation of all six different Pentasauropus localities in the lower Elliot Formation (i.e., Maseru, Seaka-Falatsa, Morobong, Maphutseng-Majakaneng, Phuthiatsana). In particular, we wish to: 1) probe how therapsids, a
Fig. 6 - Sketches and associated measurements of the three trackways at the Subeng ichnosite. a) Trackways P comprises pentadactyl tracks (Pentasauropus incredibilis). b-c) Trackways T1 and T2 comprise tetradactyl tracks (Deuterosauropodopus minor). Stride/pace measurements are indicated in centimetres. TL: trackway length; TW: trackway width; PA: pace angulation.
191E.M. Bordy et alii - Upper Triassic vertebrate tracks and trackways at Subeng in Lesotho, southern Africa
taxonomic group generally considered to be short-toed plantigrades, could make mostly unguligrade or (semi-)digitigrade tracks throughout Lesotho, especially in the light of the documented South African therapsid tracks (e.g., De Beer, 1986; MacRae, 1990; De Klerk, 2002); 2) how the southern African Pentasauropus tracks compare to those in Poland and USA; 3) fully consider the relatively rich osteological record of the Elliot Formation in search for the oversized “palaeo-Cinderella” (see “Cinderella syndrome” of Lockley, 1998) that could fit into the gigantic Seaka footprint (formerly known as Tetrasauropus gigas, Ellenberger, 1970; the humongous foot is about 70 cm long and about 55 cm wide).
tetrAdActyl trAcks - The two trackways, running parallel to each other and nearly perpendicular to the Pentasauropus trackway at Subeng, are made up of
tetradactyl, plantigrade, asymmetrical tracks with high heteropody (Figs 5-7), suggesting that the impressions were most likely made by quadrupeds. Trackway T1 (Fig. 6b) is about 2.5 m long and 54 cm wide and is comprised of six tracks with an average pes track length and width of 26 and 21 cm, respectively, and an average manus width of 10 cm (Tab. 2). The pes tracks have a slight inward rotation but the manus rotation is unclear, because generally the manus tracks are very poorly preserved. Trackway T1 has an average stride length of about 1.5 cm and a high average pace angulation of 152° (Figs 5-6). Trackway T2 (Fig. 6c) is about 2.3 m long and 62 cm wide and consists of seven tracks with an average pes track length and width of 30 and 24 cm, respectively, and an average manus width of 13 cm (Tab. 2). The pes and manus stride in T2 average about 1.3 m long with high average pace angulations of 139° and 157° for the
Trackway P (pentadactyl)
Track # W Pace
1 38
2 37 2 to 5 71
3 30
4 38 4 to 7
5 38
6 37 6 to 9 73
7 29
8 34 8 to 11 70
9 38
10 36 10 to 13 73
11 30
12 39
13 40
14 37
Average: 35.7
Trackway T1 (tetradactyl)
Track # FL W FL/W Pes to manus distance
1a 25 20 1.3 1a to 1b 41
1b 9
2a 28 23 1.2 2a to 2b 40
2b 11
3a
4a 24 21 1.1
Trackway T2 (tetradactyl)
Track # FL W FL/W Pes to manus distance
1a 33 25 1.3 1a to 1b 49
1b 13
2a 31 23 1.3 2a to 2b 45
2b 12
3a 27 24 1.1 3a to 3b 40
3b 13
4a
Tridactyl tracks
Track # FL (DL III) DL II DL IV W r FL/W r/w r/FL * 100 II^IV II^III III^IV
1 24 18 16 15 8 1.6 0.5 33 46 22 24
2 23 12 14 14 8.5 1.6 0.6 37 36 19 17
3 27 17 15 13 11 2.1 0.8 41 50
4 24 12 15 14 8 1.7 0.6 33 44 23 21
5 22 12 17 15 8.4 1.5 0.6 38 46 27 19
6 22 16 1.4
7 21 13 16 15 6.5 1.4 0.4 31 62
8 22 17 22 15 8 1.5 0.5 36 60
9 19 11 10 13 11 1.5 0.8 58 56 26 30
10 9.1 9.5 3.7 1.0 0.4 41 54
Average: 21 14 16 14 8 1.5 0.6 39 50 23 22
Tab. 2 - Measurements of the pentadactyl, tetradactyl and tridactyl tracks and trackways (P, T1, T2) at the Subeng ichnosite. All distance measurements are in centimetres; angles are in degrees. Grey blocks are for measurements that could not be determined due to e.g., absence of digit impressions. Abbreviations: FL - track length; W - track width; r - length of rear of phalangeal part of foot; DL - digit length; II^III, III^IV, II^IV - interdigital divarication angles of respective digits.

Bollettino della Società Paleontologica Italiana, 56 (2), 2017192
pes and manus tracks, respectively (Figs 5-6). Tab. 2 and Figs 5-6 also show that while the overall morphology of the tracks in the two tetradactyl trackways is similar, the wider trackway T2 comprises larger (longer and wider) pes and wider manus impressions, and therefore it is likely that the two trackways were made at the same time by the same kind, but different size quadrupedal animals (i.e., an adult and a juvenile), possibly moving adjacent to one another towards east. On the preservation scale of Belvedere & Farlow (2016), tetradactyl tracks at Subeng would fall into the quadruped grade 1 category.
The tetradactyl tracks at the Subeng were assigned to Deuterosauropodopus minor by Ellenberger (1970, 1972). Although, he referred this ichnotaxa to a basal sauropodomorph trackmaker, D’Orazi Porchetti & Nicosia (2007), who assessed a single cast of D. minor (specimen LES 053), concluded that D. minor is invalid and the specimen is an undertrack of a chirotherian [cf. Brachychirotherium (Beurlen, 1950)]. This is a curious case, because in the explanation of D’Orazi Porchetti & Nicosia (2007, p. 221), the inventory number in the Ellenberger specimen labels with the abbreviation LES (Lesotho) ought to correspond to the original number in the Ellenberger (1970) ichnotaxa list. In Ellenberger (1970), figure 53A, B, C depicts Pentasauropus erectus, and figure 53D shows Paratrisauropus equester, none of which are similar to D. minor mentioned from Subeng by Ellenberger (1970, 1972) or to LES 053 specimen shown in D’Orazi Porchetti & Nicosia (2007, fig. 21). Irrespective of this confusion, we agree with Ellenberger’s view that D. minor at Subeng is more probably made by a basal sauropodomorph, because we note the similarity of the footprint morphology and position of manus in all Deuterosauropodopus ichnites (Ellenberger, 1970, figs 50A-B, 51A-B) to some other Lesotho ichnites that are linked, though not exclusively, to basal sauropodomorphs. These morphologically similar ichnites are Sauropodopus
(Ellenberger, 1970, fig. 40), Paratetrasauropus seakensis (Ellenberger, 1972, fig. 37), Pseudotetrasauropus elegans (Ellenberger, 1970, fig. 30), P. francisci (Ellenberger, 1970, fig. 32) and P. curtus (Ellenberger, 1972, fig. 10B). The latter three are considered nomen dubium by D’Orazi Porchetti & Nicosia (2007). The further in-depth assessment of the tracks made by putative basal sauropodomorphs at Subeng is not possible before the conclusion of (or at least further advance in) our ongoing ichnological research in Lesotho. In this case, it requires the assessment of the morphological resemblance among all the above listed ichnites after the full documentation of their field context.
tridActyl trAcks - Ongoing erosion at the Subeng ichnosite has most severely affected the tridactyl bipedal tracks (Figs 2, 4-5) since their first in depth documentation by Ellenberger (1970, 1972; Tab. 1). Of the remaining tridactyl tracks, some are preserved only as digit impressions, others as connected digit traces and some merged with inflated proximal regions (Figs 5, 8). Quantitative analysis of the tracks (Tab. 2) suggests that they are morphometrically similar to each other, with an average length and width of 21 and 14 cm, respectively (average L/W ratio of 1.5). Anomalous to these averages is track G10 (Figs 5, 8), which is half of the average length and two-thirds of the average width, suggesting that is only partially reserved track. The average of the divarication angles is 50° and have an average mesaxonic index of 0.6 (i.e., anterior triangle L/W as in Lockley, 2009), with digit III being the longest digit, projecting on average 39% relative to the tip of digits II and IV (Tab. 2, Fig. 8). Track G9 is the exception projecting at 58%; the excessive projection is most likely due to the drag of the digit. The digits are straight, with slight divergent curvature along their extent, and often end in pointed tips (e.g., G1, G2, G9, G10 in Fig. 8). None of the tracks preserve additional
Fig. 7 - Comparison of the various illustrations of the same Deuterosauropodopus minor ichnite on the Subeng palaeosurface. a) Planche XIII in Ellenberger (1972). b) Fig. d in Ellenberger (1955). c) Track in trackway H in Ellenberger & Ellenberger (1958). d) Fig. 51 in Ellenberger (1970). e) Tracing of photo in f) taken in this study (see track 1a in the T2 trackway in Figs 5-6). All scale bars are equal to 10 cm.
Fig. 8 - Photographs and interpretive outlines of the individual tridactyl tracks currently exposed at the Subeng ichnosite. Tracks G1, G5 and G6 (a, b, j) were shown on the first sketch of the surface (Ellenberger, 1955) and were identified as Pseudotrisauropus dieterlini in Ellenberger (1970, 1972). Note the resemblance of tracks G2 and G10 (g, h) to P. dieterlini. Tracks G7, G8, G9 and G10 (d, e, f, h) are further to the east, outside the area documented in Ellenberger (1955). Tracks G2, G3 and G4 (g, i, c) are not shown on the first sketch of the surface. Note that tracks G4, G7, G8 and G9 (c, d, e, f) contain desiccation cracks of different sizes. For measurements of the individual tracks, see Tab. 2. All scale bars are equal to 5 cm.

193E.M. Bordy et alii - Upper Triassic vertebrate tracks and trackways at Subeng in Lesotho, southern Africa

Bollettino della Società Paleontologica Italiana, 56 (2), 2017194
morphological features such as digital pad impressions or hallux impressions. Desiccation cracks are preserved within numerous tracks (e.g., G7, G8, G9 in Fig. 8). On the preservation scale of Belvedere & Farlow (2016), tridactyl tracks at Subeng would fall into the biped grades 0 to 1 categories.
These observations are similar to diagnostic features of the ichnogenus Grallator (Hitchcock, 1858), which forms part of the continuum of Grallator-Anchisauripus-Eubrontes defined by Olsen et al. (1988), and which are commonly attributed to theropod dinosaurs. Grallator ichnogenera are described as small (length < 15 cm; FL/W ratio of 2), tulip-shaped footprints, where III digit is the longest and shows excessive projection, but never exceeding 50% of total footprint length. Outer digit divarication angles are narrow (i.e., can be as narrow as 10-30°). Grallator is distinguished from similar looking ichnogenera Carmelopodus, which lacks metatarsophalangeal pads (Lockley et al., 1998) and Jialingpus (Zhen et al., 1983), which has an inflated proximal area with digit IV metatarsophalangeal pad being twice as large as in Grallator (Xing et al., 2014). While no pads are observed in the remaining tridactyl tracks at the Subeng ichnosite, the tracks are more likely to be Grallator-like, because there is no digit IV proximal enlargement in the tracks as would be expected for Jialingpus. From the osteological record of southern Africa, there are two possible theropod trackmakers of Grallator-like tracks: 1) the smaller Coelophysis rhodesiensis (Raath, 1969) and 2) the larger Dracovenator regent (Yates & Raath, 2005). Given the average length of the tracks being about 21 cm, the biped Coelophysis rhodesiensis is deemed more likely as the trackmaker of the tridactyl tracks at the Subeng ichnosite.
Since their first description by Ellenberger (1970, 1972), the tridactyl ichnospecies (Tab. 1) from the Subeng ichnosite have only been revised to some degree by Olsen & Galton (1984). The latter authors have assigned Deuterotrisauropus socialis to the Grallator ichnogenera, and grouped the Pseudotrisauropus (Psilotrisauropus) subengensis; Pseudotrisauropus dieterleni and Mafatrisauropus errans into an indeterminate category of taxa with the remark that if “better material” would become available, these ichnotaxa may in fact turn out to be distinct and valid ichnogenera. Of the remaining tridactyl tracks, only two, G1 and G5, are marked in Ellenberger’s (1955) depiction of the Subeng ichnosite. Ellenberger classified both of these tracks as Pseudotrisauropus dieterlini (Tabs 1-2), which Olsen & Galton (1984) classified as intermediate ichnotaxa. Both G1 and G5 show distinct, narrow, curved digits with a “webbed” outline as drawn by Ellenberger (1970, 1972; Tab. 1). Given our aforementioned observations, we suggest that a) P. dieterlini should be synonymised with ichnogenus Grallator, and, b) the features distinguishing P. dieterlini from Grallator are simply extra-morphological resulting from variable preservational conditions, such as the moisture- and clay-content of the substrate, and track-substrate interaction, amongst others. The other tridactyl ichnogenera that Ellenberger (1970, 1972) described at the Subeng ichnosite (Tab. 1) cannot be observed and therefore their ichnological validity or synonymy with other ichnogenera cannot be established here.
DISCUSSION
Stratigraphy and palaeoenvironmentOur sedimentological and lithostratigraphical
observations show that the Subeng ichnosite is located about 5 m above the regional contact of the Elliot Formation with the underlying Molteno Formation despite initial assertion by Ellenberger et al. (1967) and Ellenberger (1970) that the ichnite-bearing host rocks belong to the Molteno Formation (Fig. 1B). This stratigraphic reinterpretation is in line with the reassessment of the stratigraphic position of the Lesotho cynodonts from the Molteno to the lower Elliot Formation by Turner (1972) and supported, among others, by Battail (2005). Discrepancies in Ellenberger’s stratigraphic interpretation of the Stormberg Group have also been addressed in Olsen & Galton (1984). Given that the accepted age of the lower Elliot Formation is Late Triassic throughout southern Africa (e.g., Olsen & Galton, 1984; Lucas & Hancox, 2001), the age of the Subeng ichnosite is most probably Late Triassic (Norian-Rhaetian), and thus falls into the upper part of Brachychirotherium (sensu stricto) tetrapod footprint biochron of Klein & Lucas (2010). This proposed age can be confirmed by refining of the chronostratigraphy of the Elliot Formation and determining the absolute age the ichnosite in particular (work in progress by authors).
Based on the local sedimentological and ichnological observations, the palaeosurface at the Subeng ichnosite represents a snapshot of a shallow oxbow lake in the Late Triassic. The site possibly served as a watering hole for a variety of bipedal, quadrupedal and potentially facultative bipedal vertebrates that roamed the broad fluvial floodplain (Fig. 9). The latter was dissected by both abandoned channels and active, high-energy meandering rivers that migrated laterally (Bordy et al., 2004d; fig. 4A) across the floodplain, and formed part of the drainage pattern on the NE directed regional palaeoslope (Bordy et al., 2004c, d). At the time of track formation, the oxbow lake still contained some water (that was occasionally moving; Fig. 9), however its shallow nature and shrinking surface area are demonstrated by the multitude of small ripple marks and desiccation cracks that cover the palaeosurface.
Ichnological e-archive - the digital 3D model of the Subeng ichnosite
The photogrammetric digital 3D model of this historic track-bearing palaeosurface at Subeng is now in the public domain (https://dx.doi.org/10.6084/m9.figshare.4737535). Digital 3D models of ichnosites are essentially resilient, accurate replicas as well as archives of weighty palaeosurfaces, which are usually difficult to conserve for various legislative, bureaucratic, infrastructural, etc. reasons. Housing structures to protect tracksites (e.g., Moyeni in Lesotho, Glen Rose in USA) do not ensure their preservation and may limit scientific work. At Glen Rose, despite the enclosure, loss of detail and an overall deterioration of the tracksite is observed (Bates et al., 2009; Adams et al., 2010). At Moyeni, the structure is robust, but the interior design is so basic that ichnological work on the palaeosurface is rather difficult (e.g., light conditions being suboptimal for high quality photographs).

195E.M. Bordy et alii - Upper Triassic vertebrate tracks and trackways at Subeng in Lesotho, southern Africa
In addition to creating a durable replica, digital models can be evaluated, restudied remotely or re-assessed when new methods and analytical techniques become available. Digital 3D models of ichnosites also allow for the more quantitative analysis of measurements and comparisons of results (Petti et al., 2008; Bates et al., 2009). Furthermore, digital models offer an abundance of possibilities to assess the locomotor dynamics of trackmakers as well as formation and preservation of tracks (Bates et al., 2009). For example, due to the dynamic nature of 3D models that allow high magnification and examination of tracks from any perspective, Razzolini et al. (2014) were able to quantitatively link intra-track morphology to the sedimentology of the track-bearing substrate.
It is hoped that in addition to scientific work, the digital 3D model of the Subeng ichnosite will serve as a visually stimulating tool for community outreach and science education. This is especially true where in rural African communities access to natural science museums and other educational materials is very restricted or lacking. It is noteworthy that in the past few years, the local Basotho community who lives near the Subeng ichnosite has made great effort to showcase their geoscientific heritage. For example, two small African huts (Fig. 10) were built and are maintained as a tiny visitors’ centre adjacent to the ichnosite. The huts, which only contain a visitors’ book and a few fragmentary fossil bones from the Elliot Formation, and their maintenance are a major achievement for the on-the-ground custodians of the Subeng ichnosite, who unfortunately have little or
no disposable income. It is our wish therefore that this 3D model and its description will contribute to the aspiration of the local community to share the message from the past with their visitors and to highlight the need to protect the Subeng ichnosite for future generations.
CONCLUSION
The sedimentary properties and the close stratigraphical position of the Subeng ichnosite relative to the lower contact of the Elliot Formation allow its placement into the lowermost part of the lower Elliot Formation, which is generally accepted to be Upper Triassic (Olsen & Galton, 1984; Lucas & Hancox, 2001; Knoll, 2004). The Subeng ichnosite and associated host rocks near Leribe (NW Lesotho) record the dynamics of the fluvio-lacustrine depositional environment and its biota in the Late Triassic, including short-term palaeoenvironmental changes (e.g., abandonment of meandering river channels, wet and dry cycles resulting in desiccation cracks). The 21 m2 sandstone palaeosurface is dominated by the presence of several tridactyl, tetradactyl and pentadactyl ichnites. The former are single tracks, and the others mainly make up two tetradactyl trackways and one pentadactyl trackway. Our palaeoenvironmental reconstruction demonstrates that these tracks were made around an ancient oxbow lake on a broad floodplain on which meandering rivers migrated laterally and flowed towards east-north-east. The animals which made the
Fig. 9 - Palaeoenvironmental reconstruction of the Subeng palaeosurface. This Late Triassic snapshot captures the depositional dynamics in the fluvio-lacustrine lower Elliot Formation and the generation of tracks around oxbow lakes by moderate size (some juvenile) sauropodomorphs, small bipedal theropod dinosaurs as well as large herbivorous cynodonts (therapsids). Outlines of the theropod and traversodont cynodont adapted from Olsen (1988). Numbers refer to: 1 - pentadactyl trackways: Pentasauropus incredibilis; 2 - tetradactyl trackways: Deuterosauropodopus minor; 3 - tridactyl trackways: Deuterotrisauropus socialis; 4 - tridactyl trackways: Psilotrisauropus subengensis. See text and Tab. 1 for details regarding ichnotaxonomy. Artwork by Emese M. Bordy.

Bollettino della Società Paleontologica Italiana, 56 (2), 2017196
pentadactyl and tetradactyl tracks are considered to have been of moderate to large size based on the size and morphology of the tracks. Tetradactyl tracks are most likely made by sauropodomorphs, the fossils remains of which dominate the osteological record in the lower Elliot Formation (Knoll, 2004). Previously, the pentadactyl tracks have been considered to represent a herbivorous dicynodont (therapsid) trackmakers. The tridactyl tracks are most likely associated with small bipedal theropod dinosaurs. The 3D digital model of the Subeng ichnosite will allow for future study and more detailed quantitative analyses of this key ichnological palaeosurface in the Upper Triassic lowermost Elliot Formation. Importantly, this visual tool can aid the current and on-going promotion and, ultimately, preservation of the Subeng ichnosite while
showcasing the geoscientific heritage of Lesotho on a local and global scale.
SUPPLEMENTARY DATA ARCHIVING
All the Supplementary data of this work are available at the Figshare https://dx.doi.org/10.6084/m9.figshare.4737535.
ACKNOWLEDGEMENTS
This work is a contribution to the call for papers arising from the Fourth International Congress on Ichnology “Ichnia 2016 - Ichnology for the 21st century: (palaeo)biological traces towards sustainable development”, held in Idanha-a-Nova (Portugal), 6-9 May 2016.
For this call and for the excellently managed event, we are indebted to the organizers of Ichnia 2016.
We thank our colleagues and friends, Dr Fabien Knoll and Dr Blair McPhee for their guidance regarding the fauna of the Elliot Formation and their genuine interest in our work. The local community around the Subeng ichnosite is acknowledged for their hospitality, cheerful companionship, enthusiastic site cleaning efforts and interest in our work. We are also very grateful to the Editors, Carlos Neto de Carvalho and Andrea Baucon, as well as reviewers Matteo Belvedere, Martin G. Lockley and Gerard Gierlínski for their assistance in improving this manuscript. This work was supported by the National Research Foundation of South Africa Competitive Programme for Rated Researches [NRF Grant number 93544 Bordy]. LS and MA are recipients of postdoctoral and postgraduate funding, respectively, from the DST-NRF Centre of Excellence in Palaeosciences (CoE in Palaeosciences). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript. Opinions expressed and conclusions arrived at are those of the authors and are not necessarily to be attributed to the CoE in Palaeosciences or NRF CPRR. We receive no financial benefit from this research.
REFERENCES
Adams T.L., Strganac C., Polcyn M.J. & Jacobs L.L. (2010). High resolution three-dimensional laser-scanning of the type specimen of Eubrontes (?) glenrosensis Shuler, 1935, from the Comanchean (Lower Cretaceous) of Texas: Implications for digital archiving and preservation. Palaeontologia Electronica, 13: 1-12.
Anderson J.M., Anderson H.M. & Cruickshank A.R.I. (1998). Late Triassic ecosystems of the Molteno/Lower Elliot biome of Southern Africa. Palaeontology, 41: 387-421.
Bates T., Falkingham P.L., Hidgetts D., Farlow J.O., Breithaupt B.H., O’Brien M., Matthews N., Sellers W.I. & Manning P.L. (2009). Digital imaging and public engagement in palaeontology. Geology Today, 25: 134-139.
Battail B. (2005). Late Triassic traversodontids (Synapsida, Cynodontia) in South Africa. Palaeontologia Africana, 41: 67-80.
Belvedere M. & Farlow J.O. (2016). A numerical scale for quantifying the quality of preservation of vertebrate tracks. In Falkingham P.L., Marty D. & Richter A. (eds), Dinosaur tracks, the next steps. Indiana University Press, Bloomington, Indiana: 92-98.
Beurlen K. (1950). Neue Fährtenfunde aus der Fränkischen Trias [New track discoveries in the Franconian Triassic]. Neues Jahrbuch für Geologie und Paläontologie Monatshefte, 1950: 308-320.
Fig. 10 - The Subeng ichnosite is currently managed on a volunteer basis, with extremely limited and private resources, by the Basotho community living in the vicinity of the site. In spite of lacking of funds, a sign board to the ichnosite (a) was erected on the main A1 road (sadly this was totally destroyed in 2016) and two small African huts (b) were built that function as the visitors’ centre. Various groups of locals (c) proudly and enthusiastically showcase the geoscientific heritage of the site to all visitors.

197E.M. Bordy et alii - Upper Triassic vertebrate tracks and trackways at Subeng in Lesotho, southern Africa
Bordy E.M. (2008). Enigmatic trace fossils from the Lower Jurassic Clarens Formation, southern Africa. Palaeontologia Electronica, 11/3, 16A. 16 pp.
Bordy E.M. & Eriksson P. (2015). Lithostratigraphy of the Elliot Formation (Karoo Supergroup), South Africa. South African Journal of Geology, 118: 311-316.
Bordy E.M., Hancox P.J. & Rubidge B.S. (2004a). Basin development during the deposition of the Elliot Formation (Late Triassic - Early Jurassic), Karoo Supergroup, South Africa. South African Journal of Geology, 107: 395-410.
Bordy E.M., Hancox P.J. & Rubidge B.S. (2004b). Fluvial style variations in the Late Triassic - Early Jurassic Elliot Formation, main Karoo Basin, South Africa. Journal of African Earth Sciences, 38: 383-400.
Bordy E.M., Hancox P.J. & Rubidge B.S. (2004c). Provenance Study of the Late Triassic – Early Jurassic Elliot Formation, main Karoo Basin, South Africa. South African Journal of Geology, 107: 587-602.
Bordy E.M., Hancox P.J. & Rubidge B.S. (2004d). A description of the sedimentology and palaeontology of the Upper Triassic - Lower Jurassic Elliot Formation in Lesotho. Palaeontologia Africana, 40: 43-58.
Bordy E.M., Hancox P.J. & Rubidge B.S. (2005). The contact of the Molteno and Elliot formations through the main Karoo Basin, South Africa: a second-order sequence boundary. South African Journal of Geology, 108: 349-362.
Crompton A.W. & Ellenberger F. (1957). On a new cynodont from the Molteno Beds and the origin of the tritylodontids. Annals of the South African Museum, 44: 1-13.
De Beer C.H. (1986). Surface markings, reptilian footprints and trace fossils on a palaeosurface in the Beaufort Group near Fraserburg, C.P. Annals of the Geological Survey of South Africa, 20: 129-140.
De Klerk W.J. (2002). A dicynodont trackway from the Cistecephalus Assemblage Zone in the Karoo, east of Graaff-Reinet, South Africa. Palaeontologia Africana, 38: 73-91.
D’Orazi Porchetti S. & Nicosia U. (2007). Re-examination of some large early Mesozoic tetrapod footprints from the African collection of Paul Ellenberger. Ichnos, 14: 219-245.
Duncan R.A., Hooper P.R., Rehacek J., Marsh J.S. & Duncan A.R. (1997). The timing and duration of the Karoo igneous event, southern Gondwana. Journal of Geophysical Research Solid Earth, 102: 18127-18138.
Ellenberger F. & Ellenberger P. (1958). Main types of vertebrate footprints in the Stormberg beds of Basutoland (South Africa). (Preliminary note). Compte rendu sommaire des séances de la Société Géologique de France, 1958: 65-67.
Ellenberger P. (1955). Preliminary note on the tracks and bone remains of vertebrates in Basutoland South Africa. Comptes rendus des Séances de l’Academie des Sciences (FR), 240: 889-891.
Ellenberger P. (1970). The fossil bearing strata associated with the earliest appearance of mammals in South Africa and their ichnology: establishment of detailed stratigraphic zones in the Stormberg of Lesotho, South Africa, Upper Triassic to Jurassic). In Haughton S.H. (ed.), Second Gondwanaland Symposium on Gondwana Stratigraphy and Palaeontology: proceedings and papers, Pretoria: Council for Scientific & Industrial Research (1970): 343-379.
Ellenberger P. (1972). Contribution to the classification of Triassic vertebrate trackways: types found in the Stormberg Series of South Africa (I). Paleovertebrata, Memoire Extraordinaire 1972. 152 pp. Montpellier.
Haughton S.H. (1924). The fauna and stratigraphy of the Stormberg Series. Annals of the South African Museum, 12: 323-497.
Hitchcock E. (1858). Ichnology of New England: a report on the sandstone of the Connecticut Valley, especially its fossil footmarks. 220 pp. William White, Boston.
Huene F.V. (1932). Die fossile Reptil-Ordnung Saurischia, ihre Entwicklung und Geschichte [The fossil reptile order Saurischia,
their development and history]. Monographien zur Geologie und Palaeontologie, serie 14: 1-361.
Hunt-Foster R.K., Lockley M.G., Milner A.R.C., Foster J.R., Matthews N.A., Breithaupt B.H. & Smith J.A. (2017). Tracking dinosaurs in BLM Canyon Country, Utah. Geology of the Intermountain West, 3: 67-100.
Johnson M.R., Van Vauuren C.J., Visser J.N.J., Cole D.I., de Wickens H., Christie A.D.M., Roberts D.L. & Brandl G. (2006). Sedimentary Rocks of the Karoo Supergroup. In Johnson M.R., Anhaeusser C.R. & Thomas R.J. (eds), The Geology of South Africa. Geological Society of South Africa, Johannesburg, Council for Geoscience, Pretoria: 461-501.
Kitching J.W & Raath M.A. (1984). Fossils from the Elliot and Clarens Formations (Karoo Sequence) of the Northeastern Cape, Orange Free State and Lesotho, and a suggested biozonation based on tetrapods. Palaeontologia Africana, 25: 111-125.
Klein H. & Lucas S.G. (2010). Tetrapod footprints – their use in biostratigraphy and biochronology of the Triassic. In Lucas S.G. (ed.), The Triassic Timescale. Geological Society, London, Special Publications, 334: 419-446.
Klein H. & Niedzwiedzki G. (2012). Revision of the Lower Triassic tetrapod ichnofauna from Wiory, Holy Cross Mountains, Poland. New Mexico Museum of Natural History and Science Bulletin, 56: 1-62.
Knoll F. (2004). Review of the tetrapod fauna of the “Lower Stormberg Group” of the main Karoo Basin (southern Africa): implication for the age of the Lower Elliot Formation. Bulletin de la Societe Géologique de France, 175: 73-83.
Lockley M.G. (1998). The vertebrate track record. Nature, 396: 429-432.
Lockley M.G. (2009). New perspectives on morphological variation in tridactyl footprints: clues to widespread convergence in developmental dynamics. Geological Quarterly, 53: 415-432.
Lockley M.G. & Hunt A.P. (1995). Dinosaur tracks and other fossil footprints of the western United States. 338 pp. Columbia University Press, New York.
Lockley M.G., Hunt A.P., Paquette M., Bilbey S. & Hamblin A. (1998). Dinosaur tracks from the Carmel formation, northeastern Utah: implications from middle Jurassic paleoecology. Ichnos, 5: 255-267.
Lockley M.G., Lucas S.G. & Hunt A.P. (2006). Eosauropus, a new name for a Late Triassic track: further observations on the Late Triassic ichnogenus Tetrasauropus and related forms, with notes on the limits of interpretation. New Mexico Museum of Natural History and Science, Bulletin, 37: 192-198.
Lucas S.G. & Hancox P.J. (2001). Tetrapod-Based Correlation of the Nonmarine Upper Triassic of Southern Africa. Albertiana, 25: 5-9.
MacRae C.S. (1990). Fossil vertebrate tracks near Murraysburg, C.P. Palaeontologia Africana, 27: 83-88.
Mallison H. & Wings O. (2014). Photogrammetry in paleontology – a practical guide. Journal of Paleontological Techniques, 12: 1-31.
Olsen P.E. (1988). Paleontology and paleoecology of the Newark Supergroup (early Mesozoic, eastern North America). Triassic-Jurassic Rifting: Continental Breakup and the Origins of the Atlantic Ocean and Passive Margins. Elsevier, Amsterdam: 185-230.
Olsen P.E. & Galton P.M. (1984). A review of the reptile and amphibian assemblages from the Stormberg of southern Africa, with special emphasis on the footprints and the age of the Stormberg. Palaeontologia Africana, 25: 87-110.
Olsen P.E., Smith J.B. & McDonald N.G. (1998). Type material of the type species of the classic theropod footprint genera Eubrontes, Anchisauripus, and Grallator (Early Jurassic, Hartford and Deerfield basins, Connecticut and Massachusetts, USA). Journal of Vertebrate Paleontology, 18: 586-601.
Petti F.M., Avanzini M., Belvedere M., De Gasperi M., Ferretti P., Girardi S., Remondino F. & Tomasoni R. (2008). Digital 3D modelling of dinosaur footprints by photogrammetry and

Bollettino della Società Paleontologica Italiana, 56 (2), 2017198
laser scanning techniques: integrated approach at the Coste dell’Anglone tracksite (Lower Jurassic, Southern Alps, Northern Italy). Studi Trentini di Scienze Naturali, Acta Geologica, 83: 303-315.
Raath M.A. (1969). A new coelurosaurian dinosaur from the Forest Sandstone of Rhodesia. Arnoldia (Rhodesia), 4: 1-254.
Raath M.A. (1996). The earliest evidence of dinosaurs from central Gondwana. Memoirs of the Queensland Museum, 39: 703-709.
Raath M.A., Kitching J.W., Shone R.W. & Rossouw G.J. (1990). Dinosaur tracks in Triassic Molteno sediments: the earliest evidence of dinosaurs in South Africa? Palaeontologia Africana, 27: 89-95.
Razzolini N.L., Vila B., Castañera D., Flakingham P.L., Barco J.L., Canudo J.I., Manning P.L. & Galobart A. (2014). Intra-Trackway Morphological Variations Due to Substrate Consistency: The El Frontal Dinosaur Tracksite (Lower Cretaceous, Spain). Plos One, 9: 1-14.
Sciscio L. & Bordy E.M. (2016). Palaeoclimatic conditions in the Late Triassic-Early Jurassic of southern Africa: a geochemical assessment of the Elliot Formation. Journal of African Earth Sciences, 119: 102-119.
Sciscio L., De Kock M., Bordy E.M & Knoll F. (in press). Magnetostratigraphy cross the Triassic-Jurassic boundary in the main Karoo Basin. Gondwana Research.
Tucker M.E. (2011). Sedimentary rocks in the field: a practical guide. 4th edition. 275 pp. Wiley Blackwell, Sussex.
Turner B.R. (1972). Revision of the stratigraphic position of cynodonts from the upper part of the Karroo (Gondwana) System in Lesotho. Geological Magazine, 109: 349-360.
Xing L.D., Lockley M.G., Klein H., Gierlinski G.D., Divay J.D., Hu S-M., Zhang J-P., Ye Y. & He Y-P. (2014).The non-avian theropod track Jialingpus from the Cretaceous of the Ordos Basin, China, with a revision of the type material: Implications for ichnotaxonomy and trackmaker morphology. Palaeoworld, 23: 197-199.
Yates A.M. & Raath M.A. (2005). Preliminary report of a large theropod dinosaur trackway in Clarens Formation sandstone (Early Jurassic) in the Paul Roux district, northeastern Free State, South Africa. Palaeontologia Africana, 41: 101-104.
Zhen S., Li J. & Zhen B. (1983). Dinosaur footprints of Yuechi, Sichuan. Memoirs of the Beijing Natural History Museum, 25: 1-19.
Manuscript received 16 December 2016Revised manuscript accepted 13 March 2017Published online 4 August 2017Guest Editors Andrea Baucon & Carlos Neto de Carvalho