the roles of ion fluxes in skeletal muscle fatigue
TRANSCRIPT

The roles of ion fluxes in skeletal muscle fatigue1
MICHAEL I. LHNDPNGER~ AND GEORGE 9. Fo '.EICENHAUSER~
School of Humn Biology, University of Gueirph, Guelph, Ont. , Canada MdG 2 Wd and Department of Medicine, McMsster University Medical Centre, Hamilton, Ont., C m a h U N 3Z.5
Received February 2 1, 1990
LINDHNGER, ha. I., and HEHGENHAUSER, G. J. F. 1991. The roles of ion fluxes in skeletal muscle fatigue. Can. J . Physiol. fhamacol. 69: 246 - 253.
Intense muscle contractions result in large changes in the intracellular concentrations of electrolytes. The purpose of this study was to examine the contributions of changes in intracellular strong ions to calculated changes in steady-state membrane potential (Em) and muscle intracellular H+ concentration ([H+Ji). A physicochemical model is used to examine the origin of the changes in [H+Ii during intense muscle contraction. The study used the isolated perfused rat hindimb intermittently stimulated to contract at high intensity for 5 min. This resulted in significant K+ depletion of both slow (soleus) and fast (white gastrocnemius, WG) muscle fibers and a release of K+ and lactate (Lac-) into venous pefisate. The mjo r contributor to a 12- to 14-mV depolarization of Em in soleus and WG was the decrease in intracellular K+ concentration ([K+Ii). The major independent contributors to [H+Ii are changes in the concentrations of strong and weak ions and in CO,. Significant decreases in the strong ion difference ([SID],) in both soleus and WG contributed substantially to the increase in [H+], during stimulation. In WG the model showed that the decrease in [SID], accounted for 35% of the increase in [H+Ii (133 -312 nequivil; pH, = 6.88 -6.51) at the end of stimulation. Of the main contributors to decreased [SIB],, increased [Lac-], and decreased [KtIi contributed 40 and 6096, respectively, to increased [H+],, whereas a decrease in contributed to reduced [H+],. It is concluded that decreased muscle [K+$, during intense contractions is the single most important contributor to reduced Em and increased [N'],. Depletion of PCr2- simultaneous to the changes in [Lac-], and [K+Ii prevents larger increases in [H9Ii and helps maintain the intracellular acid-base state.
Key wora3: exercise, acid-base, membrane potential, potassium, lactate.
EINDINGER, M. I., et HEEGENHAUSER, G. J. E 1991. The roles of ion fluxes in skeletal muscle fatigue. Can. 9. Physiol. Phamacol. 69 : 246-253.
Bes contractions musculaires intenses provoquent de fortes variations de concentrations intracellulaires d'Clectrolytes. Le but de cette Ctude a CtC d'examiwer les contributions des variations ioniques (Clectrolytes forts) intracelIulaires aux variations calcul&s du potentiel de membrane (Em) de repos et de la concentration de H" intracellulaire musculaire ([W+Ii). Un modble physicschimique a CtC utilisC pour examiner l'origine des variations de [H+li durant une contraction muscdaire intense. L'Ctude a utilisC le membre postCrieur pe&sC, isolC, du rat, stimulC par intermittence, pour provsquer des contractions de haute intensite d'une durCe de 5 min. Ceci a provoquC une dCplCtion K' significative des fibres musculaires 2a contraction rapide (gastrocnCmiens blancs, GB) et lente (solCaires) et une liberation de K+ et de lactate (Lac-) dans 1e perfusat veinaeux. La facteur principal d'aane dkpolarisation Be 12- 14 mV de Em dans le solkaire et He GB a CtC la diminution de la concentration de Kt intracellulaire ([K'],). k s principaux paramktres participant aux variations de [H6li sont les changements de concen- trations ioniques (Clectrolytes faibles et forts) et de CO,. La chute en diffkrences ioniques (Clectrolytes forts) ([DIP;],) dans le solCaire et le GB a manifestement contribuC a 19augmentation Be [H+Ii durant la stimulation. Dans le GB, le modble a montrC que la chute de [DIP;], a CtC respsnsable de 35 % de l'augmentation de [H+Ii (133 -312 nequiv./L; pHi = 6,$$--6,511 la fin de la stimulation. Les principaux paramktres B avoir participC B la diminution de [DIP;Ii sont m e augmentation de [Lac-], et une baisse de [K+],, qui ont CtC responsables de 40 et gO%, respectivement, de l'augmentation de [H+],, dors qu'une diminution de [PCr2-I,, a participC ?i la baisse de [H+Ii. On conclut qu'une rauction de [K+Ii muscu- laire durant d9intemes contractions est le plus important facteur de rbduction de Em et d'augmentation de [H'],. La dCplCtion de PC$-, qui survient en meme temps que les variations de [Lac-]i et de [K+Ii, pr6vient de plus fortes augmentations de [H+Ii et aide B rnaintenir 1'Cquilibre acido-basique intracellulaire.
1Wsts e&&s : exercise, acide-base, potentiel de membrane, potassium, lactate. [Traduit par la RCdaction]
Introduction Severd physiological md biochemical characteristics of rest-
ing and fatigued skeletd muscle, suck as metabolism, contrac- tile properties, and membrane properties, are highly dependent on the ionic composition of the extracellular and intracellular fluids. This gaper focuses on how ionic changes in intensely contracting muscle may contribute to the decrement in skeletal muscle force production, i . e. , fatigue. Specific consideration is given to contraction-induced changes in the intracellular con-
'This paper was presented at the symposium Cellular Aspects of Skeletal Muscle Fatigue, held January 26 and 27, 1990, University of Guelph, Guelph, Ont., Canada, and has undergone the Journal's usual p e r review.
'Author for correspondence at the first address. 3Career Investigator of the Ontario Heart and Stroke Foundation.
centrations of strong ions (those fully dissociated in solution), i .e., potassium (K+), sodium (Na +), magnesium (Mg2+), chloride (6'1-), lactate (Lac - , pK' = 3.8), and phosphocrea- tine (P6'rs -, pK' = 4.5).
Sustained muscle function and force output during contrac- tion is dependent on the provision of sufficient energy supplied by hydrolysis of ATP. During contraction, high rates of energy consumption are required by the muscle cell to main- tain transmembrane ionic and chemical gradients and osmotic equilibria in the face of increased energy demand by the con- traction-relaxation cycle. When the rate of ATP utilization exceeds locally available supplies, intracellular homeostasis will be compromised (Jones 1986). Transsarcolemmd ion movements and changes in the intracellular strong ion conwn- trations have long been associated with the depletion of cellu- lar energy stores and the fatigue md recovery processes in
PPinted in Canada I Imprim6 au Canada
Can
. J. P
hysi
ol. P
harm
acol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
YO
RK
UN
IV o
n 08
/12/
14Fo
r pe
rson
al u
se o
nly.

LINDINGER AND HEIGENHAUSER 247
skeletal muscle (Fenn 1936; Sjodin 1982; Venosa 1979). Characteristics of skeletal muscle fatigue that may be directly
influenced by contraction-induced changes in the intracellular strong ion concentrations are a decrease of the steady-state membrane potential (Hanson 1 974; Westerblad and Lannergren 1986), a decrease in the amplitude of the action potential (Han- son 1974; Sandercock et d. 1985), a decrease in intracellular pH (Roos and Boron 1981), and the activites of enzymes involved in ion pumping (Clausen 1986; Kuwayama 1988), metabolism (Trivedi and Banforth 1966; Kasvinski and Meyer 1977), and contraction (Fabiato and Fabiato 1978). The pur- pose of this paper was to examine the contributions of ion fluxes to some of these fatigue characteristics in a rat hindlimb model of skeletal muscle fatigue.
Male Sprague-Dawley rats were randomly divided into two groups: resting perfused (RP) (421 f 8 g, n = 7) and stimulated (STIM) perfused (424 f 6 g, n = 7). The experimental protocol was approved by the Care of Animals Committee at McMaster University.
The surgical preparation and electrical stimulation protocol have k e n detailed previously (Lindinger et a1 . 1987). Rats were anaesthe- tized with sodium pentobarbid (6 mg/108 g body mass i.p.). The right hindlimbs (control rest) from 13 animls randomly picked from the groups were surgically prepared for sampling of soleus, plantaris, and white gastrocnemius (WG) muscles and used as a reference for peffused muscle. Excised muscles were immediately freeze clamped with aluminum tongs cooled in liquid nitrogen.
The left hindlimb was skinned and the sciatic nerve was exposed, severed, and attached to a shielded bipolar platinum electrode. The rat was placed on a platform and the left ankle and knee secured with brackets. The gastrocnemius -plantaris - soleus (GPS) tendon was severed from its insertion at the cdcaneus and secured to a force trans- ducer. The femoral artery and vein of the left hindlimb were exposed above the knee and c a m u l a ~ with 22 and 18 gauge teflon catheters, respectively. The hindlimb was flushed with 1-2 mL of heparinized saline and was perfused with a defined perfusion medium (Lindinger et A. 1986) containing the following: bovine red blood cells (hernato- crit = 38 f 0.3%, hemoglobin = 135 f 1 g/L), [Naf] = 151 f 0.4 mequiv./L, [Cl-] = 114 f 0.4 mequiv./L, [K+] = 5.43 f 0.02 mequiv./L, [Mg2+] = 2.1 f 0.1 mequiv./L, [Ca2+] = 2.58 f 0.02 mequiv./L, [Lac-] = 0.91 f 0.84 mequiv./L, klucose] = 48 f 0.7 g/L, [albumin] = 41 f 1 g/L, PC% = 38.7 f 0.3 mmHg, pH = 7-44 f 0.004, 0, saturation = 99.2 f 0.2%. Muscle extracellular fluid volume (ECFV) was measured using [aH]mnnitol (168 pCi/L; New England Nuclear, Mississauga, Ont.) (1 Ci = 37 GBq) added to the arterial perfusate.
Pe&sion and stimulation Perfusions were performed at 35°C. In WP, the left hindlimb was
perfused at a flow of 1.7 mL/fin for 20 min to establish a steady state between muscle and perfusate. Arterial and venous blood samples were obtained at 10, 15, and 20 min of perfusion for blood gases and ion and metabolite analyses. At the end of 20 min the entire soleus and plantaris and a portion of WG were individually removed and quickly freeze clamped.
S T I I hindlimbs were initially perfused for 20 min at a flow of 1.7 d / m i n while resting and then electrically stimulated to contract for 5 min. With the onset of stimulation, perfusate flow rate was increased to 7.4 f 0.3 mE/min, a Wow that did not limit oxygen delivery to the muscles (Lindinger et al. 1987; Spriet et al. 1986). Intennittent stimulation of the GPS muscles via the sciatic nerve (Grass Instruments SrC8 Stimulator) consisted of 108-ms trains (186 Hz) at a frequency of 0.5 Hz; individual trains consisted of 10 separate impulses in 188 ms, each 0.2 ms in duration at a supramaximal 5-V intensity. The perfusion pressure (Gould-Statham 48783 pressure transducer) and isometric tetanic tension (Statham 10713 force trans- ducer) developed by the GPS muscle group were continuously moni-
tored on a chart recorder (Hewlett-Packard 7754A). Imedia tdy following 5 min of stimulation, muscle samples were taken from the hindlimb and freeze clamped.
Analyses Muscle samples were analyzed for Lac- and PC8- and arterial and
venous blood were analyzed for Lac- and glucose using enzymatic fluorometry (Bergmeyer 1974). Plasma was analyzed for Pco,, Po,, and pH at 37'C using a Radiometer BMS3 MK 2 blood microsystem coupled to a Radiometer PHM72 M U digital acid -base analyzer. Hemoglobin and oxygen saturation were measured using a Radiom- eter OSM2 hemoximeter. Plasma [Naf] and [K+] were measured with a Radiometer KNAl sodium-potassium analyzer and C1- was determined on a Buchler-Cotlove chloridometer. Plasma protein was measured by refractometry and hemtocrit was measured using a high-speed microcentrifuge.
Muscle inorganic ions (Na+, K*, Cl-, Mg2', and Ca2+) were measured by instrumental neutron activation analysis (Lindinger and Heigenhauser 1987). This technique measures both total bound and total free muscle ions; however, only the total ion concentrations are reported in this study, i.e., no distinction is made between bound and free intracellular ions. Muscle total tissue water (TTW) was deter- mined by $raze drying muscle samples to a constant weight. Intra- cellular ion concentrations (mequiv. /L ICF; ICF, intracellular fluid) were calculated from total muscle ion content, ECFV, and TTW as described previously (Lindinger et al. 1987; Lindinger and Heigen- hauser 1987).
The steady-state membrane potential of perfused resting and stimu- lated muscles (Em, in mV) was calculated using the following q u a - tion (Woodbury 1982):
where RTIF = 26.7 mV at 37'C, [ionlo is the interstitial fluid ion activity (assumed equal to that of plasm and ECF), [ionIi is the ICF activity, a is the ratio of Naf permeability to IC+ permeability, and y is the ratio of C1- permeability to K+ permeability. The same values of a and y were used for resting and stimulated muscle. It is known that changes in internal and external ion concentrations affect membmne permeability to ions (Hodgkin and Horowicz 1959; Wood- bury 1982); however, changes in ionic permeability of the sarco- lemma have not been quantified in fatigued skeletal muscle. The value of cr was taken to be 0.018 and 0.009 in soleus (SL) and WG, respectively (estimated from work of Yonemura, 1967, and Dul- hunty, 1978) and y = 0.208 (Woodbury 1982) was used for both muscles. The measured plasm concentrations of K+, Na+, and Cl- and the corresponding activity coefficients of 8.76, 0.65, and 0.75 (from Walker and Brown 1977) were used to calculate interstitial activities of 3.8, 91, and 79 rnequiv./L, respectively. The intracellu- lar ion activities for K', Na+, and C1- were 0.65; 0.42, and 0.18, respectively (Dsnaldson and Leader 1984; Leader et al. 1984).
The equations describing the m s s action and chemical equilibria for weak acids, carbon dioxide, and strong ions in aqueous solutions may be combined to yield a single equation that may be solved for [H+] when the strong ion difference ([SIB]), the total concentration of weak acids and bases ([&,el), and Pco, are known (equation 7A.1.2 from Stewart, 1981):
where KA, the mean dissociation constant for weak acids (A,,), has been determined to be 1 .&? x and 1 .98 x equiv./l in WG of resting and fatigued rats, respectively, and 1.64 x lo-' equiv./l in both resting and fatigued soleaas (Lindinger and Heigenhauser 19W). WG and SL [bodi was determined to be 145 and 130 mequiv./L in resting rat muscle, and after 5 f i n of intense
Can
. J. P
hysi
ol. P
harm
acol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
YO
RK
UN
IV o
n 08
/12/
14Fo
r pe
rson
al u
se o
nly.

248 CAN. J. PHYSIOL. PHARMACOL. VBL. 69, 1991
swimming to exhaustion it was 170 mequiv./L in WG and unchanged in SL (Lindinger and Heigenhauser 1990). The vdues for the con- stants are as follows: water dissociation (Kk) = 4.40 x 10-l4 (equiv . /L) (Ham& and Owen 1958); carbonate and bicarbonate dissociation (K3) = 6.0 X 18-" equiv./l (Edsall and Wymnn 1958); and Kc, the product of the apparent dissociation constant for C8, (7.41 x 1W3 equiv./l) and the CO, solubility coefficient, 8.0351
g-l at 37OC (1 m H g = 133.32 Pa) (Siesjo and Thews 1962). Muscle intracellular Pco, was taken as 56 and 70 mmHg in resting and fatigued muscle, respectively (Bonde-Petersen and Lundsgaard 1983; Hogg et al. 1984).
Intracellular [SID], was calculated as
Stati&YtE'cs Results are expressed as means k SE. Data were andysed using
one-way ANOVA to cornpare intratreatment differences over time and differences between treatments. When a significant F ratio was obtained, the means were cornpard rasing the Student's t-test, unpaired design. Significance was accepted at p I 0.05.
Results Hindlimb O2 uptake was 0.46 & 0.03 1 pmol - g- l perhsed
muscle min- l at rest and 2.3 8 & 0.154 pmol . g - l perfused muscle min-I during stimulation. Glucose uptake and lac- tate release in resting muscle were 0.054 + 0.003 pmol . g- l perfused muscle min-I and 0.085 + 0.883 pequiv. - g-l perfused muscle - anin-'. Perfused muscle mass was previ- ously determined to average 1.54 g/lOO g body mass, with stimulated muscle mass comprising 82 % of perfused muscle and the GPS muscle contributing 44% to perfused muscle mass and 53.4% to stimulated muscle mass (Lindinger et al. 1987).
Initid tetanic tension production by the GPS group was 3367 f 104 g . Tension decreased linearly between 0 and 2 min to 2626 & 126 and was 15 15 +_ 98 g after 5 min of stim- ulation.
Muscle fluid volumes increased in SL and WG during stimulation (Table 8 ) . h SL, TTW increased 6.1 % from RP, with 63% of the increase in TTW being confined to the extracellular space. In WG, TTW increased 5.776, with 22% of the increase being extracellular. This resulted in 2.4 and 4.7 % increases in ICHV of SL and WG, respectively.
During the 5 anin of stimulation there was a 2 -5 % reduction in venous plasma volume compared with arterial as determined from measurements of plasma protein and blood hematocrit and hemoglobin. Venous plasma ion concentrations were corrected for hernoconcentration and net ion fluxes were calculated as the product s f arteriovenous difference and Wow rate (Fig. 1). The final 5 min of resting perfusion showed no net fluxes of ions between pefisate and muscle indicating that a steady state had been established. Stimulation resulted in significant releases of Lac- and K+ that were maximal at 3 -4 min. In contrast, Na+ and Cl- continued to enter the stimulated muscles at increasing rates throughout the period of stimulation. During the 5 min of stimulation, perfused tissues took up 25.5 f 1 -2 pequiv. /g wet weight of Na+ and 26.0 + 1.6 pequiv ./g wet weight of C1- . Total K + loss was 1.86 f 0.08 pequiv. /g wet weight and total Lac- release was 10.4 f 0.74 pequiv./g wet weight.
Intracellular ion concentrations at the end of rest perfusion armd after 5 min of stimulation are shown in Fig. 2. In general, the changes reflect those seen in the pefisate. Stimulation sig- nificantly decreased [PCr2-Ii, [K + Ii, [Mg2+Ii, and [SIDIi in
TABLE 1 . TTW, ECFV, and HCFV in soleus and white gastrocnemius muscle before and after 5 min stimulation
Condition TTW ECFV ICFV
Soleus Control 0.766f 0.004 0.049+0.008 0.688 k0.007 RP 0.798f 0.011 0.090f 0.007 8.709f 0.006 STIM 0.847k0.019" 0.121 f 0.015" 0.426f 0.014
White gastrocnemius
Control 0.758+0.003 0.06Qf 0.010 0.701 k0.013 RP 0.785f 0.808 0.065f 0.006 0.721 f8.003 STIM 0.$30+0.023* 0.075 f 8.022 0.755 +0.005*
NOTE: Values are means f SE (mL/g wet weight). TIW, total tissue water; BCFV, extraceIlular fluid volume; ICFV, intracellular fluid vdume; STIM, stimulated perfusd; RP, resting perfused.
"Indicates STIM mean significantly different from RP mean.
both muscles and increased [LacWli in WG but not SL (Fig. 2). The changes in [Na+li and [C1-li in both muscles were not significant. These concentrations were used to calculate Em and intracellular [H +Ii.
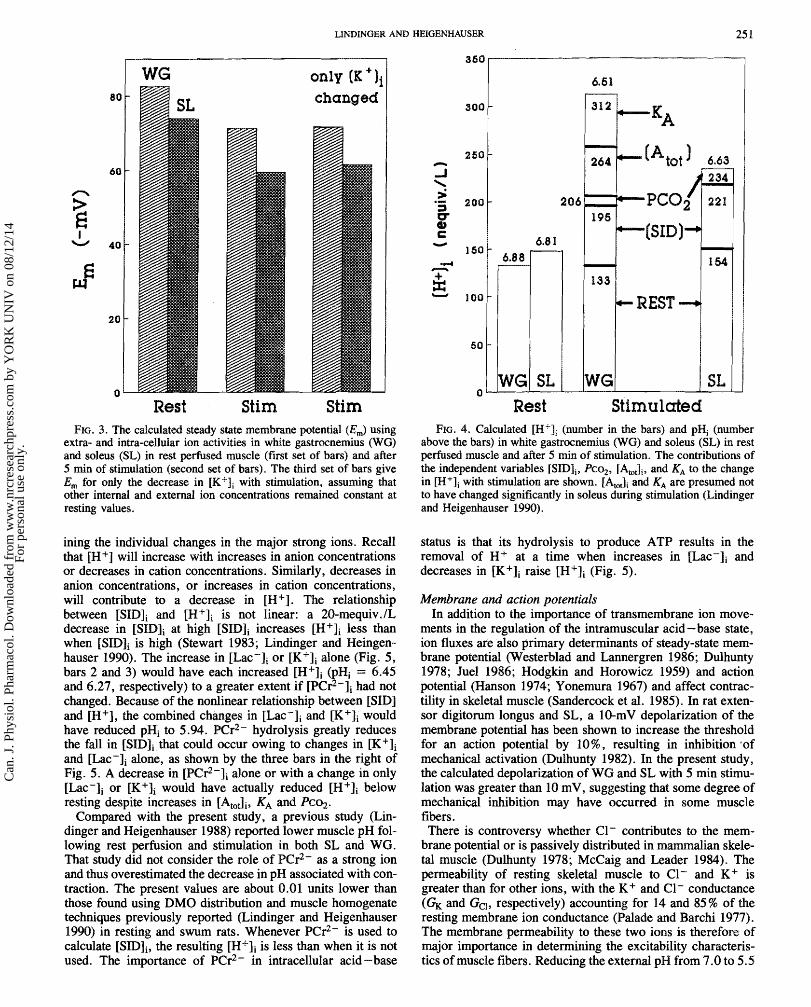
The calculated Em before and after stimulation are shown in Fig. '3. At rest, Em averaged 8.6 mV less in SL than in WG. Stimulation deplarized Em by 82 mV in WG and 14 rrmV in SL. Figure 3 (third series of bars) also shows that nearly all of the change in Em in both muscles was due to the reduction in [KfIi, since the changes in [Na+Ii and [Cl-Ii were minimal. The one-pass system maintained external ion concentrations constant. The potassium equilibrium potentials (Ed were - 85 and - $0 mV in W S and SL at rest, and -72 and -64 mV after stimulation.
Cdcdated [H + Ii in resting SL and WG and the contributions of changes in [SIDIi, Pco2, [AtoJi, and KA to [H+Ii in stimu- lated muscle are shown in Fig. 4. At rest, pHi averaged 6.81 f 0.608 and 6.88 f 0.087 in SL and WG, respectively, while pHi was 6.63 & 0.33 (WG) and 6.51 f 0.854 (SL) after stimulation. Increases in &02, [AtOtli, and KA and decreases in contributed to the increase in in WG (Fig. 4). In the SL of intensely swum rats there were no sig- nificant changes in and KA (Lindinger and Heigen- hauser 1990) and so these variables were kept constant for SL in this study to give the vdues shown in Fig. 4. The major contributor to the increase in [Hfqi was the decrease in [SIDIi, accounting for about 35 and 84% of the increase in
in W S and SL, respectively. An estimated 20 mmHg increase in Pco2 (from 50 to 70 m d g , see Methods) con- tributed 6- 16%. In WG the increases in KA and [A,Ji accounted for 27 and 30% of the increase in The con- tributors to the increases in [AtoJi in WG with stimulation include creatine, inorganic phosphate, and glycolytic phos- phates, which in turn raise the apparent KA for total weak acids in muscle (Lindinger and Heigenhauser 1988).
The contributions of individual and combined changes in the strong ions Lac-, K +, and PCr2- to [M + Ii in stimulated WG we shown in Fig. 5. The first bar in Fig. 5 shows the cdcu- lated WG [H+Ii (pHi = 6.51) at the end of stimulation and the other bars show the contributions to a change in only one or two of the major strong ions, with Pco2, [At&, and KA held at post-stimulation vdues. The increase in [Lac-Ii or the decrease in [K+Ii done could each have accounted for more than the entire increase in WG attributed to reduced [SIDIi (Fig. 5, bars 2 and 3). The change in [Lac-li con-
Can
. J. P
hysi
ol. P
harm
acol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
YO
RK
UN
IV o
n 08
/12/
14Fo
r pe
rson
al u
se o
nly.

LINDINGBR AND HBIGBNWAUSER
Time (min) Time ( m i d FIG. 1 . Ion fluxes in isolated gefised rat Rindlimb during the final 5 min of the 28-min equilibration period and during 5 min of intermittent
tetanic eleetrieal stimulation. Negative values represent an effaux from the hindlimb and positive values represent uptake by the hindlimb. All stimulation values (means f SEMI are significantly different from resting values.
tributed 40% to the combined effects of changed [Lac-ji and [K+li (Fig. 5, bar 4) which, in the absence of changes in the other variables, would have resulted in a pHi of about 5.94. However, the acidifying effects of Lac- accumulation and K+ depletion were counterbalanced to a large extent by the decrease in [PC$-ji, which, taken alone, would have con- tributed to a reduced [H+li (Fig. 5, bar 5). The influence of incorporating the decrease in [PCr2-Ii to the individual changes in and [K+ji (from bars 2 and 3, Fig. 5) on keeping [H+Ii low is shown in bars 6 and 7 of Fig. 5.
Discussion
During intense muscle contraction many closely associated events occur in skeletal muscle and in the blood perfusing the muscle, which lead to a downregulation of the contractile machinery and the metabolic inhibition characteristic of reduced force production.
It is the intent of this paper to demonstrate how changes in intracellular strong ion concentrations during muscle contrac- tion may influence cellular events associated with skeletal muscle fatigue. It was shown that changes in strong ions greatly affect the sarcolemal steady-state membrane poten- tial and [H+ji. These changes may in turn contribute to impaired force production at sites upstream from excitation- contraction coupling and energy supply systems (Bigland- Ritchie et al. 1986), while the intracellular ionic changes (including H+, protonated state of phosphates and proteins) may contribute more directly to a downregulation of glyco- genolytic energy production (Chasiotis et al. 1982; Trivedi
and Danforth 1966) and rate of actin-myosin cross-bridge cycling (Fabiato and Fabiato 1978).
The metabolic and ionic integrity of the isolated perfused rat hindlimb preparation used in this study has previously been demonstrated (Lindinger et d. 1987; Spriet et al. 1986). A major limitation of the present ionic analysis of skeletal muscle is that the physicochemical model treats the different intra- cellular compartments as one large compartment with homo- genous distribution of intracellular constituents. This is known not to be the case. Cellular organelles undergo discrete, though related, fluid and ion shifts during muscle contraction (Sembrowich et al . 1982; S o d yo et al . 1977). Another lirnita- tion is that free or diffusible ions (Walker and Brown 1977; Maughan and Godt 1989) are not distinguished from total intracellular ions. Microelectrode studies have shown that free, not total, K+, Naf and possibly C1- contribute directly to the membrane potential (Walker and Brown 1977; McCaig and Leader 1984) and the same is probably true with regard to intracellular acid -base state. However, it is unknown how the activity coefficients for the different ions change during exercise-induced alterations in subcellular and transsarcolem- mal ion distribution. The bound forms of these ions may also be physiologically important since they may be released during certain conditions, K+ release from glycogen during muscle contraction for example (Bergstrom and Hultman 1966). The results are also limited in that the same values for a and y were used in the calculation of Em for resting and stimulated muscle. It is known that the conductances of individual ions are affected by changes in extra- and intra-cellular ion concentra- tions (Woodbury 1982); however, fatigue-associated effects
Can
. J. P
hysi
ol. P
harm
acol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
YO
RK
UN
IV o
n 08
/12/
14Fo
r pe
rson
al u
se o
nly.

CAN. J. PHYSIOE. PHAWMACOL. VOL. 69, 1921
100 148
White Gastrocnemius
FIG. 2. Intracellular strong ion concentrations and [$ID], in rest perhsed muscle md after 5 min sf stimulation in soleus and white gastroc- nemius. Asterisks indicate stimulation means are significantly different from rest means.
on sarcolemal ion permeability have not been quantified in skeletal muscle. The advantage of the modelling technique used in this study lies in the ability to identify the variables that cause changes in muscle physicochemical status that accom- pany the fatigue and recovery processes.
A limitation of this study is that muscle [A,,,Ji and its dis- sociation constant KA were not measured. However, 5 nain of intermittent tetanic electrical stimulation in the rat hindirnb closely models the metabolic and ionic events observed in rat hindlimb during 5 min of swimming to exhaustion (Lindinger et d. 1987), so it is expected that the resultant changes in
and KA, which are determined by the metabolic and ionic events, would dso be similar. The main reason for the absence of change in SL [&&Ii and KA in the previous study in swum rats (Lindinger and Heigenhauser 1990) lies in the fact that changes in Per2- and ATP, which reflect changes in intracellular weak acids (Lindinger and Heigenhauser 1990), were small compared with those seen in the white gastroc- nemius.
jSHDji and pHi The biochemical mechanisms by which glycogenolytic
Lac- production, PCr2- hydrolysis - resynthesis, and ATP hydrolysis -synthesis may be involved in the metabolic production or consumption of H+ have been elegantly described by Hochachh and Momsen (1 983). Intracellular [H+li is dso dependent on intracellular buffering by weak acids and bases and by transport of ions across the cell mem- brane (Aickin and Thomas 11977; Woos and Boron 1981). At any point in time, therefore, [H+Ii may be determined by examining the physicoehemicd composition of the intra- cellular fluids, which reflects the net effect of metabolic, buffering, and transport processes acting on the cell. Changes in the intracellular concentrations of weak acids and bases ([A,&) and the apparent dissociation KA have been previ- ously described in detail (Lindinger and Heigenhauser 1990) and this discussion will focus on the strong ions.
With exercise the effects of increases in [Na+Ii9 [Mg2+Ii, and [C1-Ii are relatively small and tend to cancel each other. In contrast with the decrease in [K+ji and increase in [Lac-Ji9 which lowers [SIDIi and increases [H+li, the decrease in [PCr2-Ii raises [SID]; and lowers [H+Ii. Following intense exercise, PCr2- is rapidly regenerated and, together with Lac- removal and K+ repletion, is important in restoring intracellular ionic status during the recovery process.
The differences in the ionic status of skeletal muscle at rest and following exercise are related to muscle fiber type compo- sition and muscle function. The [SIDIi at rest and following exercise reflects the intracelluhr concentrations of strong ions (Fig. 2). Compared with fast twitch WG, the 15 mequiv./L lower [SZDIi in resting soleus results primarily from the sig- nificantly lower [K+Ii, in turn resulting in a higher resting [H+Ii (Fig. 4). The decrease in [SHDIi after stimulation in both muscles was similar; however, the increase in [P%+Ii in WG (177 nequiv./L) was greater than in SL (79 nequiv./L). This difference appears to be due to a significant increase in
of highly glycolytic fibers (WG) with intense muscular contraction (Lindinger and Meigenhauser B 990), which, together with the change in KA, contributes to an increase in
(Fig. 4). The high resting [SIDli in WG m y effectively prolong continued muscle function during exercise by prevent- ing even larger changes in [H+li and in the protonated form of muscle transport and metabolic proteins. Changes in intracellular strong ions may directly or indirectly alter the catalytic activity of enzymes h t are classically considered as "pH sensitive" (Dixon and Webb 1979). Such pH effects have been proposed for both the regulation of glycogenolysis at the level of phosphorylase and phosphofructokinase (Hultman and Sjoholm 1986; Trivedi and Banforth 1966) and also the impairment of contraction at the level of the sarcoplasmic reticulum (Fabiato and Fabiato 1978) and at the myofilaments (Cooke et al. 1988; Hibberd et al. 1985).
An understanding of the physicochehaaid Bbasis for the contri- bution of reduced [SHD] to increased [M+] is helped by exam-
Can
. J. P
hysi
ol. P
harm
acol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
YO
RK
UN
IV o
n 08
/12/
14Fo
r pe
rson
al u
se o
nly.
(K + I j changed
FIG. 3. The cdculatd steady state membrane potential (Ern) using extra- and intra-cellular ion activities in white gastrocnemius (We) and soleus (SL) in rest p e f i s d muscle (first set of bars) and after 5 min of stimulation (second set of bars). The third set of bars give Em for only the decrease in with stimulation, assuming that other internal and ex tend ion concentrations remained constant at resting values.
ining the individual changes in the major strong ions. Recall that [H+] will increase with increases in anion concentrations or decreases in cation concentrations. Similarly, decreases in anion concentrations, or increases in cation concentrations, will contribute to a decrease in [H+]. The relationship between [SIDIi and [HfIi is not linear: a 20-mequiv./L decrease in [SIDIi at high [SIH>ji increases [H+Ii less than when [SID]i is high (Stewart 1983; Lindinger and Heingen- hauser 1990). The increase in [Lac-Ii or [K'Ii done (Fig. 5, bars 2 and 3) would have each increased [H+Ii @Hi = 6.45 and 6.27, respectively) to a greater extent if [BCr2-1, had not changed. Because of the nonlinear relationship between [SID] and [H+], the combined changes in [Lac-Ii and [K+qi would have reduced pHi to 5.94. Per2- hydrolysis greatly reduces the fall in [SHDIi that could occur owing to changes in [K+Ii and [Lac-Ii alone, as shown by the three bars in the right of Fig. 5. A decrease in [P@r2-Ii done or with a change in only [Lac-]i or [K+Ii would have actually reduced [H+Ii below resting despite increases in [A&, KA and Bco2.
Compared with the present study, a previous study (Lin- dinger and Heigenhauser 1988) reported 'lower muscle pH fol- lowing rest perfusion and stimulation in both SL and WG. That study did not consider the role of Per2- as a strong ion and thus overestimated the decrease in pH associated with con- traction. The present values are about 8.01 units lower than those found using DM0 distribution and muscle homogenate techniques previously reported (Lindinger and Heigenhauser 1998) in resting and swum rats. Whenever Per2- is used to calculate [SHDIi, the resulting [HfJi is less than when it is not used. The importance of PGr2- in intracellular acid-base
206 I95
6.8 1 -6ID)- 1 54
134
~lc- REST --+.
SL WG SL Rest Stimulated
FIG. 4. Calculated [H'J, (number in the bars) and pHi (number above the bars) in white gastrocnemius w e ) and soleus (SL) in rest perfused muscle and after 5 knin s f stimulation. The csntributions of the independent variables [SIB],, Pcs,, [At&, and KA to the change in [H+], with stimulation are shown. [AtO,li and KA are presumed not to have changed significantly in soleus during stimulation (Lindinger and Heigenhsluser 1990).
status is that its hydrolysis to produce ATP results in the removal of H+ at a time when increases in and decreases in [K+Ii raise [HCIi (Fig. 5).
Membrane and action pokntials In addition to the importance of transmembrane ion move-
ments in the regulation of the intramuscular acid-base state, ion fluxes are also primary determinants of steady-state mem- brane potential (Westerblad and Lannergren 1986; Dulhunty 1978; Juel 1986; Hodgkin and Horowicz 1959) and action potential (Hanson 1974; Yonemura 1967) and affect contrac- tility in skeletal muscle (Sandercock et d. 1985). In rat exten- sor digitorum longus and SL, a 10-mV depolarization of eke membrane potential has been shown to increase eke threshold for an action potential by 1096, resulting in inhibition .of mechanical activation (DuHhunty 1982). In the present study, the calculated depolarization of WG and SL with 5 min stimu- lation was greater than 18 mV, suggesting that some degree of mechanical inhibition may have occurred in some muscle fibers. There is controversy whether Cl- contributes to the mem-
brane potential or is passively distributed in mammalian skele- tal muscle (Dulhunty 1978; McCaig and Leader 1984). The permeability of resting skeletal muscle to C1- and K+ is greater than for other ions, with the K+ and C1- conductance (& and GcI, respectively) accounting for 14 and 85 % of eke resting membrane ion conductance (Palade and Barchi 1977). The membrane permeability to these two ions is therefore of major importance in determining the excitability characteris- tics of muscle fibers. Reducing the external pH from 7.0 to 5.5
Can
. J. P
hysi
ol. P
harm
acol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
YO
RK
UN
IV o
n 08
/12/
14Fo
r pe
rson
al u
se o
nly.

252 CAN. I. PHYSIBL. BHARMACOL. V8L. 69, 1991
STSM Atac AK+ ALac ABCr ABCr APCr AK+ ALage Ah[+
FIG. 5. The contributions of changes in the major intracellular strong ions (Lac-, K+ , and PC$-), singly or in combination, to cal- culated [H+Ii and pHi (number above each bar) in white gastroc- nemius after 5 min of stimulation.
reduces Gel by 5076, indicating a pH sensitivity of the func- tional groups cdf the C1- channels (Pdade and Barchi 1977). It is reasonable to suggest that this apparent pH sensitivity of the Cl- channels is also present intracellularly so that decreases in pHi would reduce C1- conductance during intense exercise; however, this aspect of fatigued muscle does not appear to have been studied.
The increase in sarcolemal pemeability to K+ upon metabolic exhaustion in frog skeletd muscle has been sug- gested to be due to activation of ATP-sensitive K+ channels (Castle and Haylett 1987; Sjogaard 119911). Changes in the sarcolemmal permeabilities to K+ and C1- during fatigue only moderately affect calculation of the steady-state mem- brane potentid if resting values for the Na+/K+ and 61-/K+ permeability ratios were used. For example, a two-fold decrease in C1- permeability combined with a two-fold increase in K+ permeability would markedly decrease the Cl-/K+ permeability ratio, but this only polarizes Em by 1 -2 mV. Assuming constant Na+ permeability, a two-fold increase in K + permeability would only polarize Em by about 2-3 mV. Taken together, the effects of increased M+ and reduced C1- permeabilities polarize the membrane by approximately 5 mV and would negate approximately 60% of the stimulation-induced reduction in WG Em shown in Fig. 3. These mechanisms could contribute significantly to a mainten- ance of force production in the face of decreases in [K+qi.
Additional research is required to answer the hndamental questions of how ion permeabilities change in fatiguing skele- tal muscle, and what are the roles of bound and free ions in the regulation of muscle ion homeostasis during contraction, fatigue, and recovery.
Acknowledgments We gratefully acknowledge the technical assistance of Dr.
M. Ganagarajah and Sandra Peters. Thanks go to Donald Welsh for critical reading of the manuscript. This work was supported by grants from the Medical Research Council of Canada and the Natural Sciences and Engineering Research Council of Canada.
AICKIN, C. C., and THOMAS, R. C. 1977. An investigation of the ionic mechanism of intracellular pH regulation in mouse soleus muscle fibers. J. Physiol. (London), 273: 295 -4 16.
BERGMEYER, H. U. 1974. Methods of enzymatic analysis. Verlag Chemie, Weinheim.
BERGSTROM, J., and HULTMAN, E. 1966. The effect of exercise on muscle glycogen and electrolytes in normals. Scand. J. Clin. Lab. Invest. 18: 1-5.
BIGLAND-WITCHIE, B. R., DAWSON, N. J., JOWANSSON, R. S., and LIPPOLD, Q. C. J. 1986. Reflex origin for the slowing of motoneu- rone firing rates in fatigue of humn voluntary contractions. J. Physisl. (London), 379: 45 1 -459.
BONDE-PETERSEN, E, and LUNDSGAARD, J. S. 1983. Gas tensions (O,, CO,, Ar and N,) in human muscle during static exercise and occlusion. In Biochemistry of exercise. Edited by H. G. Knuttgen, J. A. Vogel, and J. Poortmans. Humn Kinetics, Champaign, IL. pp. 781 -786.
CASTLE, N. A., and HAYLETT, D. G. 1987. Effect of channel blockers on potassium efflux from metabolically exhausted frog skeletal muscle. J. Physiol. (London), 383: 31 -43.
CHASIBTIS, D., HULTMAN, .E., and SAWLIN, K. 1982. Acidotic depression of cyclic AMP accumulation and phosphorylase b to a transformation in skeletd muscle of man. J. Physiol. (London), 335: 197 -204.
CLAWSEN, 9. 1986. Regulation of active Na* -K+ transport in skeletal muscle. Physiol. Rev. 66: 542 -588.
CQOKE, R., FRANKS, K., LUCIANI, G. B., and PATE, E. 1988. The inhibition of rabbit skeletd muscle contraction by hydrogen ions and phosphate. J. Physiol. (London), 395: 77 -97.
DIXON, M., and WEBB, E. C. 1979. Evymes. 3rd ed. Academic Press, New York.
DONALIPSON, P. J., and LEADER, J. P. 1984. IntracelluIar ionic activi- ties in the EDL muscle of the mouse. Pfluegers Arch. $00: 166- 178.
DULPIUNTY, A. E 1978. The dependence of membrane potential on extracellular chloride concentration in mammalian skeletal muscle fibres. J. Physisl. (London), 276: 67 - 82.
1982. Effects of membrane potential on mechanical activa- tion in skeletal muscle. J. Gen. Physiol. 79: 233 -251.
EDSALL, J. To, and WYMANN, J. 1958. Biophysical chemistry. Vol. 1 . Academic Press, New York. pp. 578 -587.
FABIATO, A., and FABIATO, E 1978. Effects of pH on the myofila- ments and the sarcoplasmic reticulum of skinned cells from cardiac and skeletal muscles. J. Physiol. (London), 276: 234 -255.
FENN, W. 0. 1936. Electrolytes in muscle. Physiol. Rev. 16: 450-487.
HANSON, J. 1974. The effects of repetitive stimulation on the action potential and the twitch of rat muscle. Acta Physiol. Scand. 98: 387 -408.
HARNED, H. S . , and OWEN, Be B. 1958. The physical chemistry of electrolyte solutions. 3rd ed. Van Nostrand-Reinhold, New York.
HIBBERD, M. G. , DANTZIG, J. A., TRENTHAM, D. R., and GOLDMAN, Y. E. 1985. Phosphate release and force generation in skeletal mus- cle fibers. Science (Washington, D.C.), 228: 13 17 - 13 19.
WOGMACHKA, P. W,, and MOMMSEN, T. P. 1983. Protons and anaerobiosis. Science Washington, D.C.), 21 9: 1391 - 1397.
H O ~ K I N , A. L., and HOROWICZ, P. 1959. Movements of Na and K in single muscle fibres. J. Physiol. (London), 145: $05 -432.
HOGG, R. J., ~ C A C C O , L. R., CARTER, N. We, L A ~ O I , A. R., and KOKKO, J. P. 1984. In situ PCO, in the rend cortex, liver, muscle,
Can
. J. P
hysi
ol. P
harm
acol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
YO
RK
UN
IV o
n 08
/12/
14Fo
r pe
rson
al u
se o
nly.

LINDINGER AND HEIGENHAUSER 253
and brain of the New Zedand White rabbit. Am. J. Physiol. 247: F49 1 - F498.
HULTMAN, E., and SJOHOLM. H. 1986. Biochemical causes of fatigue. In Human Muscle Power. Edited by N. L. Jones, N. McCartney , and A. J. McComas. Human Kinetics, Champaign, IL. pp. 215-238.
JONES, D. P. 1986. Intracellular diffusion gradients of 0, and ATP. Am J. Physiol. 250: C663 -C675.
JUEL, C. 1986. Potassium and sodium shifts during in vitro isometric muscle contraction and the time course of the ion-gradient recov- ery. Pfluegers Arch. 406: 458 -463.
KASVINSKY, P. J., and MBYER, W. L. 1977. The effect of pH and tem- perature on the kinetics of native and altered glycogen phosphory- lase. Arch. Biochem. Biophys. 181: 616-63 1.
KWAYAMA, H. 1988. The membrane potential modulates the ATP- dependent Ca2+ pump of cardiac sarcolemma. Biochim. Biophys. Acta, 940: 295 - 299.
LEADER, J. R, BRAY, J. J., MACKNIGHT, A. D. C., MASON, D. R., MCCAIG, D., and MILLS, R. G. 1984. Cellular ions in intact and denervated muscles of the rat. J. Membr. Biol. 81: 19 -27.
LINDINGEW, M. I., and HEIGENHAUSER, G. J. F. 1987. Intracellular ion concentrations of rat hindimb muscles measured by instrumen- tal neutron activation analysis. J. Appl. Physiol. 63: 426-433.
1988. Ion fluxes during tetanic stimulation in isolated, per- fused rat hindlimb. Am. J. Physiol. 254: R117-R126.
1998. Acid-base systems in skeletal muscle and their response to exercise. In Biochemistry of exercise. VII. Edited by A. W. Taylor, P. D. Gollnick, H. J. Green, C. D. Ianuzao et al. Humn Kinetics, Champaign, IL. pp. 341 -357.
LINDINGER, M. I., HEIGENHAUSER, G. J. E, and JONES, N. L. 1986. Acid-base and respiratory properties of a buffered bovine ery- throcyte perh ion medium. Can. J. Physiol. Phamcol. 64: 55Q- 555.
LINDINGEW, M. I., HEIGEMHAUSER, G. J. F., and SPRIET, L. L. 1987. Effects of intense swimming and tetanic electrical stimulation on skeletal muscle ions and metabolites. J. Appl. Physiol. 63: 2331 -2339.
MAUGHAN, D. W., and Gom, R. E. 1989. Equilibrium distribution of ions in a muscle fiber. Biophys. J. 56: 717 -722.
MCCAIG, D., and LEADER, J. P. 1984. Intracellular chloride activity in the extensor digitorum longus (EDL) muscle of the rat. J. Membr. Biol. 81: 9-17.
PALADE, P. T., and BARCHI, R. L. 1977. Characteristics of the chlo- ride conductance in muscle fibers of the rat diaphragm. J. Gen. Physiol. 69: 325 -342.
Roos, A., and BORON, W. F. 1981. Intracellular pH. Physiol Rev. 61: 2%-434.
SANDERCOCK, T. G., FAULKNER, J. A., ALBERS, J. W., and
ABBRECHT, P. H. 1985. Single motor unit and fiber action poten- tials during fatigue. J. Appl. Physiol. 58: 1073 - 1879.
SEMBROWICH, W. L., JOHNSON, D., WANG, E., and HUTCHISON, T. E. 1982. Electron microprobe analysis of fatigued fast- and slow- twitch muscle. In Biochemistry of exercise. V. Edited by H. G. Knuttgen, J. A. Vogel, and J. Poortmans. Human Kinetics. Cham- paign, IL. pp. 571-576.
SIESJO, B. K., and THEWS, G. 1962. Ein verfahren zur Bestimung der C0,-Leitfaigkeit der C0,-Diffusionskoeffizienten und des scheinbaren C0,-bslichkeitkoeffiienten in Gehimgeweben. mue- gers Arch. 276: 192-210.
SJOBIN, R. A. 1982. Transport of electrolytes in muscle. J. Membr. Biol. 68: 161 - 178.
SJOGAARD, G. 1991. Role of exercise-induced potassium fluxes underlying muscle fatigue: a brief review. Can. J. Physiol. Phamacol. 69: This Issue.
SOMLYO, A. V., SHUMAN, H., and SOMLYO, A. P. 1977. Elemental distribution in striated muscle and the effects of hypertonicity: electron probe andysis of cry0 sections. J, Cell Biol. 71L: 828 - 857.
SPWIET, L. L., LINDINGER, M. I., HEIGENHAUSER, G. J. F., and JONES, N. L. 1986. Effects of alkalosis on skeletal muscle metabolism and performance during exercise. Am. J. Physiol. 251: R833 -R839.
STEWART, P. A. 1981. How to understand acid - base: a quantitative acid -base primer for biology and medicine. Elsevier-North Hol- land, New York.
1 983. Modern quantitative acid - base chemistry. Can. J. Physiol. Phamacol. 61: 144-4 - 1461.
WVEDI, B., and DANFORTH, W. H. 1966. Effect of pH on the kinetics of frog muscle phosphofructokinase. J. Biol. Chem. 241: 4110-4114.
VENOSA, R. A. 1979. Ionic movements across the plasma membrane of skeletal muscle fibers. In Membrane transport in biology. Vol. 11. Edited by G. Giebisch, D. C. Tosteson, and H. H. Ussing. Raven Press, New York. pp. 2 1 1 -242.
WALKER, J. L., and BROWN, H. M. 1977. Intracellular ionic activity measurements in nenie and muscle. Physiol. Rev. 57: 729-778.
WESTERBLAD, H., and LANNERGREN, J. 1986. Force and membrane potential during and after fatiguing, intermittent tetanic stimulation of single Xenopus muscle fibres. Acta Physiol. Scand. 128: 369-378.
WOODBURY, J. W. 1982. The cell membrane: ion fluxes and athe gen- esis of the resting potential. In Physiology and biophysics. Edited by T. C . Ruch and H. D. Patton. W. B. Saunders Co., Philadel- phia, PA. pp. 1-46.
YONEMURA, K. 1967. Resting and action potentials in red and white muscle of the rat. Jpn. J. Physiol. 17: 788 -7 19.
Can
. J. P
hysi
ol. P
harm
acol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
YO
RK
UN
IV o
n 08
/12/
14Fo
r pe
rson
al u
se o
nly.