the role of brain-gut peptides in the control of sodium appetite
TRANSCRIPT

192
The Role of Brain–Gut Peptides in the Control of Sodium Appetite
GAYLEN L. EDWARDSa AND JOYCE D. POWER
Department of Physiology and Pharmacology, College of Veterinary Medicine,The University of Georgia, Athens, Georgia 30602, USA
ABSTRACT: Ingestion of food and fluid stimulates release of a number of pep-tides from the gastrointestinal system. These peptides are recognized to act asneurotransmitters/neuromodulators and act at both peripheral and central re-ceptors. Many studies indicate that these peptides are important signals in ter-minating meals. Recent studies suggest that bombesin, a peptide related togastrin-releasing peptide, suppresses sodium appetite. We have investigatedthe role of cholecystokinin (CCK) in the control of sodium appetite. Our stud-ies indicate that CCK is effective at reducing saline intake. We found that ex-ogenous, intraperitoneal CCK octapeptide suppresses saline intake. Moreover,administration of trypsin inhibitor to stimulate endogenous CCK release re-sulted in suppression of saline intake. Finally, intraperitoneal administrationof the CCK receptor antagonist lorglumide resulted in increased saline intake.These observations extend the potential role of gastrointestinal peptides in themodulation of ingestive behavior.
Ingestion of sodium plays an important role in regulation of fluid balance. Inappro-priate regulation of sodium intake or excretion can contribute to pathologic disorderssuch as high blood pressure. Thus, studies examining factors that suppress sodiumappetite have focused on those factors related to controls of fluid balance. Notably,blood volume and blood pressure have received the most attention as variables thatcan influence sodium intake. Nonetheless, recent studies suggest that factors relatedto ingestion of food may also suppress sodium appetite. Since most sodium is ingest-ed with food, and not as a solution, it is reasonable to postulate that gastrointestinalfactors work to suppress sodium intake. In fact, initial reports suggest that gas-trointestinal factors, such as the peptide bombesin, may be important to the inhibi-tion of sodium appetite in rats.1,3,4 Thus, our laboratory has recently initiated studiesto examine the role of gastrointestinal peptides on the control of sodium appetite.
In our first studies we focused on the peptide bombesin. These studies were in-tended to utilize a short-term method of inducing sodium appetite and verify thatbombesin would suppress sodium appetite in this paradigm. Bombesin is a peptiderelated to gastrin-releasing peptide (GRP) that is released during ingestion of ameal.1,3,4 Earlier reports suggest that peripheral treatment with bombesin results insuppressed sodium appetite to sodium depletion-induced sodium appetite.1,3,4 Our
a706/542-5854 (voice);706/542-0261 (fax).e-mail: [email protected]
193EDWARDS & POWER: PEPTIDES CONTROLLING SODIUM APPETITE
studies used a modified paradigm developed by Fitts and colleagues2,8 to induce arapid-onset sodium appetite. In this paradigm the animals are treated with furosemi-de (10mg/kg) followed 10 min later by 5 mg/kg captopril. One hour later the animalsare presented with a concentrated sodium solution (2%). This paradigm has beendemonstrated to reduce blood volume as well as produce a marked sodium loss andinduce a robust sodium appetite.2,8
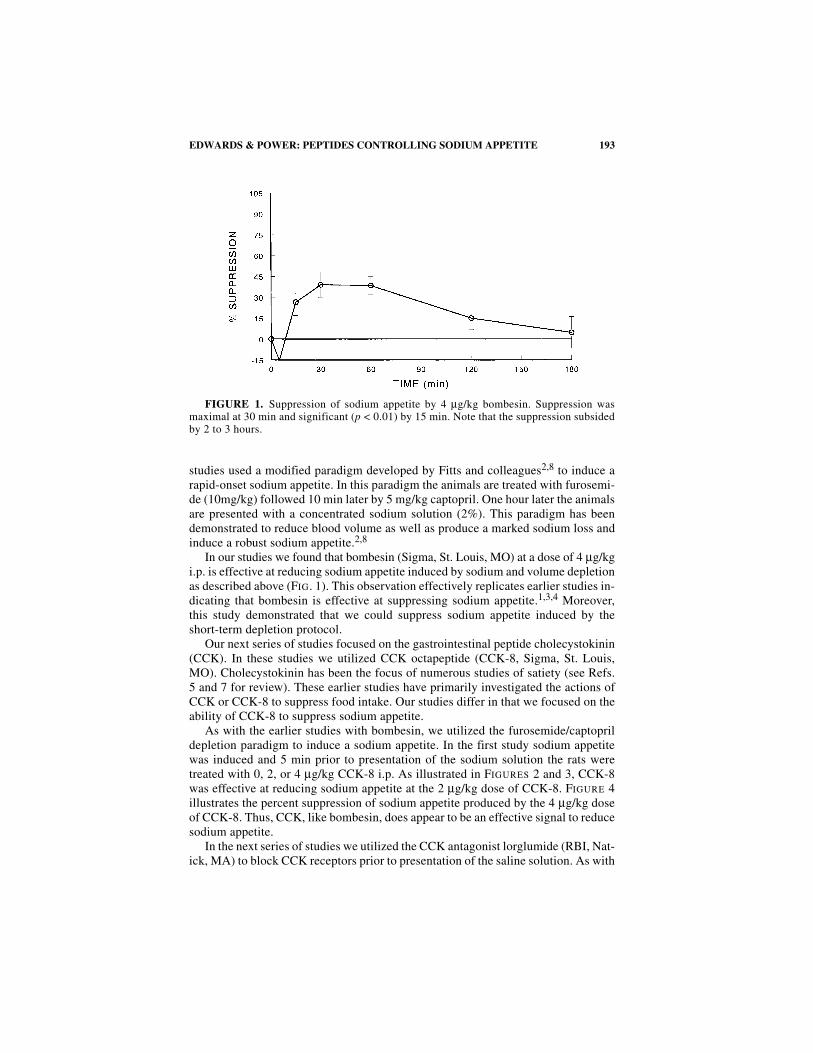
In our studies we found that bombesin (Sigma, St. Louis, MO) at a dose of 4 µg/kgi.p. is effective at reducing sodium appetite induced by sodium and volume depletionas described above (FIG. 1). This observation effectively replicates earlier studies in-dicating that bombesin is effective at suppressing sodium appetite.1,3,4 Moreover,this study demonstrated that we could suppress sodium appetite induced by theshort-term depletion protocol.
Our next series of studies focused on the gastrointestinal peptide cholecystokinin(CCK). In these studies we utilized CCK octapeptide (CCK-8, Sigma, St. Louis,MO). Cholecystokinin has been the focus of numerous studies of satiety (see Refs.5 and 7 for review). These earlier studies have primarily investigated the actions ofCCK or CCK-8 to suppress food intake. Our studies differ in that we focused on theability of CCK-8 to suppress sodium appetite.
As with the earlier studies with bombesin, we utilized the furosemide/captoprildepletion paradigm to induce a sodium appetite. In the first study sodium appetitewas induced and 5 min prior to presentation of the sodium solution the rats weretreated with 0, 2, or 4 µg/kg CCK-8 i.p. As illustrated in FIGURES 2 and 3, CCK-8was effective at reducing sodium appetite at the 2 µg/kg dose of CCK-8. FIGURE 4illustrates the percent suppression of sodium appetite produced by the 4 µg/kg doseof CCK-8. Thus, CCK, like bombesin, does appear to be an effective signal to reducesodium appetite.
In the next series of studies we utilized the CCK antagonist lorglumide (RBI, Nat-ick, MA) to block CCK receptors prior to presentation of the saline solution. As with
FIGURE 1. Suppression of sodium appetite by 4 µg/kg bombesin. Suppression wasmaximal at 30 min and significant (p < 0.01) by 15 min. Note that the suppression subsidedby 2 to 3 hours.

194 ANNALS NEW YORK ACADEMY OF SCIENCES
the earlier studies, sodium appetite was induced by furosemide/captopril treatment.Lorglumide (600 µg/kg) was administered 10 min prior to presentation of the salinesolution. As illustrated in FIGURE 5, lorglumide treatment significantly elevated sa-line intake in rats with depletion-induced sodium appetite. This observation comple-ments the earlier observation with CCK-8 as it suggests that antagonizingendogenous CCK receptors facilitates saline intake under these conditions.
The final set of studies utilized trypsin inhibitor (Sigma, St. Louis, MO) to stim-ulate release of endogenous CCK. Again sodium appetite was induced by the deple-tion method described above. Trypsin inhibitor (150 mg/kg) was administered bygavage 30 min prior to presentation of the saline tubes. Like exogenous CCK-8 treat-
FIGURE 3. Suppression of saline intake induced by short-term depletion by 2 µg/kgCCK-8. CCK significantly suppressed saline intake (p < 0.01) with maximal suppression by15 minutes.
FIGURE 2. Effect of 2 µg/kg CCK-8 on 2% saline intake induced by short-term deple-tion. Note that CCK significantly suppressed saline intake throughout the 3-hr period of thetest (p < 0.01).

195EDWARDS & POWER: PEPTIDES CONTROLLING SODIUM APPETITE
ment, trypsin inhibitor caused a significant suppression of saline intake (FIG. 6). Thisobservation strongly supports the observations with lorglumide implicating endoge-nous CCK in the control of sodium appetite.
Our data support a role for gastrointestinal signals in the control of sodium appe-tite. Particularly intriguing are the data with CCK. Our data indicate that CCK-8, aCCK receptor agonist, suppresses sodium appetite. Lorglumide, a CCK receptor an-tagonist, enhances intake, indicating that endogenous CCK is acting as a brake onsodium appetite. Finally, trypsin inhibitor acts to suppress sodium appetite, possiblyby releasing endogenous CCK. Together these data offer strong support for the hy-pothesis that CCK from the gastrointestinal tract acts to inhibit sodium appetite.
FIGURE 4. Suppression of saline intake induced by short-term depletion by 4 µg/kgCCK-8. CCK significantly suppressed saline intake (p < 0.01) with maximal suppression by15 minutes.
FIGURE 5. Enhanced saline intake induced by lorglumide (600 µg/kg). Lorglumidesignificantly elevated saline intake (p < 0.05) over the 2-hr test period.

196 ANNALS NEW YORK ACADEMY OF SCIENCES
The hypothesis that CCK acts to suppress sodium appetite is based on the fact thatmost sodium is ingested with food and not fluid. Thus, signals that suppress food in-take should also suppress sodium appetite. Our data strongly suggest that this is thecase.
The route by which CCK acts to suppress sodium appetite is unclear at this point.The most likely route is via vagal afferent fibers. Vagal afferent fibers have been dem-onstrated to convey the satietogenic signal of CCK to the brain for short-term controlof food intake.5,7 It is also possible that CCK acts by other routes to suppress sodiumappetite. These could include spinal afferent pathways, or circumventricular organs,although preliminary data suggest that the area postrema is not involved in CCK-in-duced suppression of sodium appetite (unpublished observation). Future studies invagally deafferented animals as well as in those with other central neural lesions willhelp elucidate the routes by which CCK acts to suppress sodium appetite.
ACKNOWLEDGMENTS
This work was supported by Grant DK42533 from the National Institutes ofHealth.
REFERENCES
1. DE CARO, G., C. POLIDORI, T.G. BELTZ & A.K. JOHNSON. 1998. Area postrema, lateralparabrachial nucleus, and the antinatriorexic effect of bombesin. Peptides 19: 1399–1406.
2. FITTS, D.A. & D.B. MASSON. 1989. Forebrain sites of action for drinking and saltappetite to angiotensin or captopril. Behav. Neurosci. 103: 865–872.
3. FLYNN, F.W. & R. RAMOS. 1994. Bombesin suppresses need-free and need-induced saltintake in rats. Behav. Neurosci. 108: 780–788.
FIGURE 6. Suppression of saline intake by trypsin inhibitor (150 mg/kg). Trypsin in-hibitor significantly suppressed saline intake (p < 0.03) over the 3-hr test period.

197 ANNALS NEW YORK ACADEMY OF SCIENCES
4. FRANKMANN, S.P. 1994. Bombesin, gastrin-releasing peptide, and neuromedin B atten-uate the salt appetite of sodium-depleted rats. Ann. N.Y. Acad. Sci. 739: 344–346.
5. GIBBS, J. & G.P. SMITH. 1986. Gut peptides and feeding behavior: The model of chole-cystokinin. In Feeding Behavior: Neural and Humoral Controls. R.C. Ritter, S. Ritterand C.D. Barnes, Eds.: 329–352. Academic Press. Orlando, FL.
6. MENANI, J.V. & A.K. JOHNSON. 1998. Cholecystokinin actions in the parabrachialnucleus: effects on thirst and salt appetite. Am. J. Physiol. 275: R1431–R1437.
7. SCHWARTZ, G.J. & T.H. MORAN. 1996. Sub-diaphragmatic vagal afferent integrationof meal-related gastrointestinal signals. Neurosci. Biobehav. Rev. 20: 47–56.
8. THUNHORST, R.L. & D.A. FITTS. 1994. Peripheral angiotensin causes salt appetite inrats. Am. J. Physiol. 267: R171–R177.