the river continuum redux: aquatic insect diets … · reveal the importance of autochthonous...
TRANSCRIPT

Loyola University ChicagoLoyola eCommons
Master's Theses Theses and Dissertations
2011
The River Continuum Redux: Aquatic Insect DietsReveal the Importance of AutochthonousResources in the Salmon River, IdahoKathryn VallisLoyola University Chicago
This Thesis is brought to you for free and open access by the Theses and Dissertations at Loyola eCommons. It has been accepted for inclusion inMaster's Theses by an authorized administrator of Loyola eCommons. For more information, please contact [email protected].
This work is licensed under a Creative Commons Attribution-Noncommercial-No Derivative Works 3.0 License.Copyright © 2011 Kathryn Vallis
Recommended CitationVallis, Kathryn, "The River Continuum Redux: Aquatic Insect Diets Reveal the Importance of Autochthonous Resources in theSalmon River, Idaho" (2011). Master's Theses. Paper 561.http://ecommons.luc.edu/luc_theses/561

LOYOLA UNIVERSITY CHICAGO
THE RIVER CONTINUUM REDUX:
AQUATIC INSECT DIETS REVEAL THE IMPORTANCE OF
AUTOCHTHONOUS RESOURCES IN THE SALMON RIVER, IDAHO
A THESIS SUBMITTED TO
THE FACULTY OF THE GRADUATE SCHOOL
IN CANDIDACY FOR THE DEGREE OF
MASTER OF SCIENCE
PROGRAM IN BIOLOGY
BY
KATHRYN LYNDSEY VALLIS
CHICAGO, ILLINOIS
DECEMBER 2011

Copyright by Kathryn Lyndsey Vallis, 2011
All rights reserved.

iii
ACKNOWLEDGEMENTS
Although the end result of a Master’s program is a degree celebrating an
individual’s achievement, the journey to get to that point is certainly not traveled alone. I
would first like to thank all of those that I had the privilege of working alongside in the
Rosi-Marshall lab at Loyola and the Cary Institute, particularly Dr. Heather Bechtold,
Holly Wellard Kelly, Dustin Kincaid, and Sarah Zahn Seegert, for their help with the
many and various aspects of this project, for their perspective and good humor, and for
their support and friendship both in and out of the lab. I would also like to thank Sylvia
Lee, for generously offering her time and diatom expertise when it was most needed.
I would also like to thank the Baxter lab at Idaho State University for their
accommodating and welcoming ways during the time I spent in Pocatello. I would
especially like to thank Jennifer Cornell, Kevin Donner, Hannah Harris, Jessica Lueders-
Dumont, Rebecca Martin, and Cameron Morris for teaching me the ways of the wild
West and for their help in the field and the lab. I must also thank Dr. G. Wayne Minshall,
for giving me access to the Idaho State University archives, and Dave Funk, for his insect
identification work and meticulously labeled specimen vials. This work was supported
by a grant from Idaho NSF-EPSCoR (EPS-0814387).
I would also like to thank the staff at the Cary Institute of Ecosystem Studies, for
facilitating my transition to New York and a new laboratory environment. I would
particularly like to thank Heather Malcolm and Deana Machtley for sharing their

iv
extensive scientific knowledge, lab space, and equipment, and for their continual
encouragement and support. I would like to thank Dr. David Strayer, for allowing me to
spend many microscope hours in his lab, and on his microscopes.
I’d also like to thank Dr. John Davis for all of the time and energy invested
toward me and this project, for his willingness to be a sounding-board and editor, for his
invaluable perspective, and for his constant encouragement and understanding.
I must also thank Dr. Colden Baxter for welcoming me into his lab at Idaho State
University, for his stream-side instruction and campfire-side tunes, and for his great
insight, thoughtful comments, and valued assistance throughout the course of this project.
I would like to thank my committee members Dr. Martin Berg, for his expertise in
all things aquatic insects and statistics, and for all of his time, effort, and support invested
in me and this project, and Dr. Christopher Peterson, for his help and constructive
remarks, for sharing his classroom with me, and for his assistance in the development and
writing of this work.
I thank my advisor, Dr. Emma Rosi-Marshall, for her endless enthusiasm, her
staunch support, her insightful contributions, and her constant guidance and
encouragement throughout my undergraduate and Master’s programs. For this, and for
all of the new experiences and incredible people I encountered as a result of her
mentorship and this project, I will always be thankful.
Finally, I must thank my family – especially my parents, George and Josephine,
and my brother, Christopher – and my friends for listening to me in times of exhilaration
and frustration, for offering invaluable advice and perspective, and for providing constant

v
and necessary reminders of the “bigger picture.” It was your support and love that saw
me through this journey, and I am incredibly grateful you were there to travel it with me.

To my parents, George and Josephine, and my brother, Christopher, for all of their love
and support. To the memory of my grandfather, Milton, who gave his family a place to
enjoy and appreciate all that our freshwater ecosystems have to offer.

No man ever steps in the same river twice…
Heraclitus

TABLE OF CONTENTS
ACKNOWLEDGEMENTS iii
LIST OF TABLES ix
LIST OF FIGURES x
ABSTRACT xii
CHAPTER ONE: INTRODUCTION 1
CHAPTER TWO: METHODS 6
Study sites 6
Taxa studied 9
Diet analysis 11
Statistical analysis 12
CHAPTER THREE: RESULTS 14
Community resource use and the predictions of the RCC 14
Functional feeding groups 17
Shredders 17
Collector-gatherers 18
Collector-filterers 18
Scraper-grazers 19
CHAPTER FOUR: DISCUSSION 26
The River Continuum Redux 26
Global climate change signals in macroinvertebrate diet composition 28
Functional feeding groups and gut content analysis 29
Facultative diet-switching may help stabilize food webs 32
Hidden treasures: Archived insects and the value of invertebrate
diet composition 33
Conclusions 35
LITERATURE CITED 36
VITA 44

ix
LIST OF TABLES
Table 1. Percent of (un-weighted) allochthonous (AL), autochthonous (AU),
amorphous detritus (AD), and animal (AN) materials in the diets of
individual taxa collected from Camp Creek (Site A) in summer and fall
1976 and 2009. 21
Table 2. Percent of (un-weighted) allochthonous (AL), autochthonous (AU),
amorphous detritus (AD), and animal (AN) materials in the diets of
individual taxa collected from Smiley Creek (Site B) in summer and fall
1976 and 2009. 22
Table 3. Percent of (un-weighted) allochthonous (AL), autochthonous (AU),
amorphous detritus (AD), and animal (AN) materials in the diets of
individual taxa collected from Obsidian (Site C) in summer and fall 1976
and 2009. 23
Table 4. Percent of (un-weighted) allochthonous (AL), autochthonous (AU),
amorphous detritus (AD), and animal (AN) materials in the diets of
individual taxa collected from Casino (Site D) in summer and fall 1976
and 2009. 24

x
LIST OF FIGURES
Figure 1. The Salmon River basin is located in central Idaho (inset). My sampling
sites were located in the upper Salmon River, in the Sawtooth National
Recreation area near Stanley, Idaho. Adapted from Minshall et al.
1982. 8
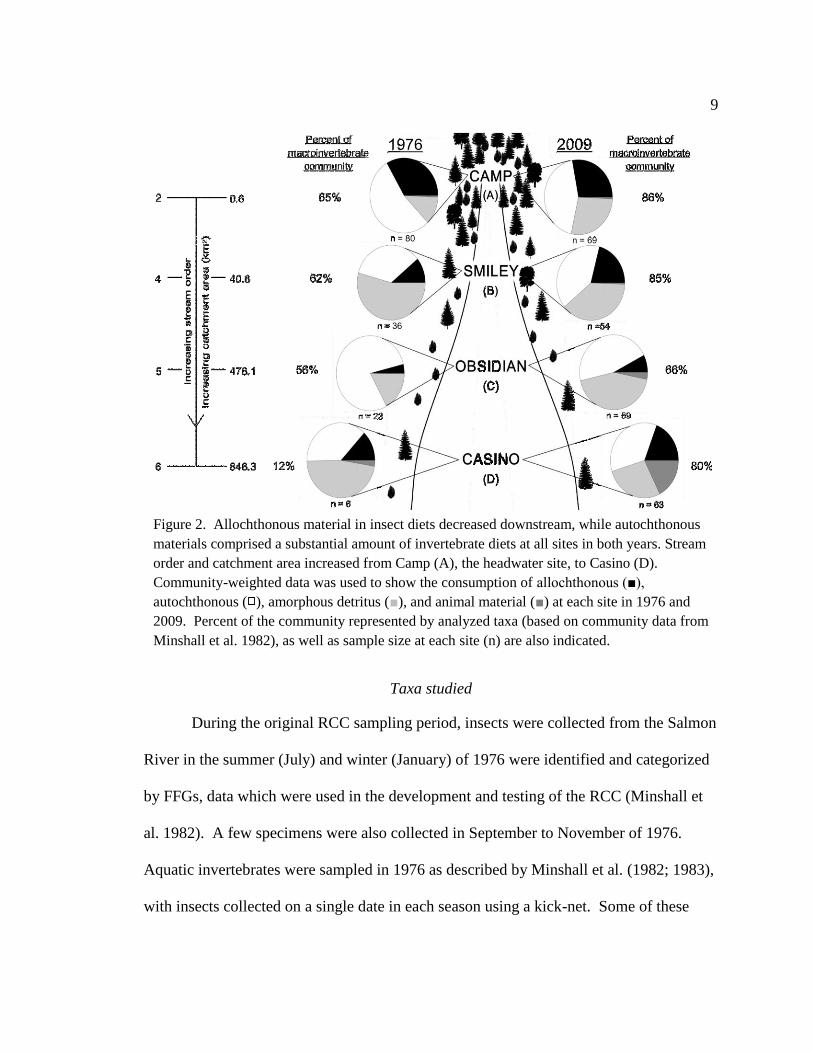
Figure 2. Allochthonous material in insect diets decreased downstream, while
autochthonous materials comprised a substantial amount of invertebrate
diets at all sites in both years. Stream order and catchment area increased
from Camp (A), the headwater site, to Casino (D). Community-weighted
data was used to show the consumption of allochthonous, autochthonous,
amorphous detritus, and animal material at each site in 1976 and 2009.
Percent of the community represented by analyzed taxa (based on
community data from Minshall et al. 1982), as well as sample size at
each site (n) are also indicated. 9
Figure 3. Variation in the proportion of (a) allochthonous materials (b)
autochthonous materials, and (c) amorphous detritus among individual
insect diets at each Salmon River, ID site in 1976 and 2009.
Consumption of all materials was similar at all sites in both years. 15
Figure 4. (a) Hypothetical changes in the presence of allochthonous resources and
autochthonous resources along the longitudinal gradient, as predicted by
the RCC: allochthonous resources should be highest in the headwaters,
decreasing downstream; autochthonous resources should increase
downstream. (b) Regression lines of allochthonous and autochthonous
resources in the diets of individual insects collected in 1976 and 2009
from the Salmon River, ID. Autochthonous material in diets is similar
across sites, and allochthonous material in diets decreases downstream;
however, consumption of allochthonous materials is not as dramatic as
originally predicted. 16

xi
Figure 5. Proportion allochthonous, autochthonous, amorphous detritus, and animal
materials found in the diets of (a) shredders, (b) collector-gatherers, (c)
collector-filterers, (d) scraper-grazers collected from the Salmon River, ID
in summer and fall 1976 and 2009. Overall, shredders consumed the most
allochthonous material, and autochthonous material was consumed by all
functional feeding groups (FFGs). In general, diets were similar across
sites, seasons, and years. 25

xii
ABSTRACT
Along a river network, changes in the physical characteristics of a stream –
changes articulated by the River Continuum Concept (RCC) – have been predicted to
influence stream food webs, particularly the resources supporting aquatic
macroinvertebrates. Although diets of macroinvertebrates can track resource availability
along the longitudinal gradient of streams, insect diets were not assessed in the original
RCC study; rather, insects were grouped into functional feeding groups (FFGs). In
addition, global climate change is increasing the frequency and spatial extent of wildfire
and beetle outbreaks throughout the western United States and could affect riparian
vegetation and the amount of allochthonous inputs entering the system. Based on gut-
content analysis of insects collected and archived from the Salmon River (SR), ID in
1976 and used in the development of the RCC, I first tested whether insect diets followed
the original predictions of the RCC. Based on the predictions of RCC, I predicted that
consumption of allochthonous inputs of terrestrial leaves would be greatest in the
headwaters and decline along the continuum. In addition, by resampling these sites in
2009, I assessed whether the relative importance of allochthonous and autochthonous
resources have shifted along this continuum, indicating potential shifts in riparian
vegetation since 1976. Insect diets in 1976 and 2009 were similar across years and
seasons, and these data did not indicate changes in riparian vegetation. In addition, insect

xiii
diets did not follow the predictions of the RCC, as SR insects consumed primarily
autochthonous material in the forested headwaters. Allochthony decreased along the
longitudinal gradient, as predicted by the RCC; however, this trend was not as dramatic
as predicted by FFGs alone. Insects in the headwaters, including the dominant shredder
Yoraperla (Insecta: Plecoptera), consumed mostly autochthonous material. Autotrophic
production, then, was shown to be an important food resource, even in this forested
headwater stream. As this reliance on autochthonous resources was high for those
invertebrates (e.g. shredders) thought to rely mostly on allochthonous material, my results
indicate the importance of gut content analysis and cautions against relying on FFGs to
explore which resources fuel stream food webs.

1
CHAPTER ONE
INTRODUCTION
The River Continuum Concept, developed by Vannote et al. (1980), provides a
framework by which one can predict biological, physical, and chemical changes along a
longitudinal gradient in stream systems. It was proposed that terrestrial and aquatic
ecosystems are inherently linked, as are downstream and upstream reaches (Minshall et
al. 1983, Vannote et al. 1980, Rosi-Marshall and Wallace 2002, Li and Dudgeon 2008),
and that the importance of these linkages can change along the continuum. While the
general applicability of the RCC has been debated (Thorp and Delong 1994 & 2002,
Statzner and Higler 1985, Li and Dudgeon 2008) and some elements have been amended
over time (Minshall et al. 1983), the general concept is still useful, especially in
temperate forested stream networks.
Drawing from observations and data collected from sites along a number of river
networks, the RCC proposes that to understand the biological dynamics of streams and
riverine systems, one must consider the gradient of physical factors formed by the
drainage network (Vannote et al. 1980). Based on the idea of dynamic equilibrium, the
authors propose that a stream ecosystem achieves a balance between the physical factors
– stream size, width, velocity, depth, sediment load – and the biological factors – primary
production, organism composition. Such shifts in the physical and chemical
characteristics of a stream are also reflected in resident macroinvertebrate assemblages of

2
stream systems. Aquatic insects are organized into functional feeding groups (FFGs)
based upon their behavioral and morphological adaptations for food acquisition
(Cummins 1973, Cummins and Klug 1979, Vannote et al. 1980). These groups include
shredders, collector-gatherers, collector-filterers, scraper-grazers, and predators. As food
resources shift along the longitudinal gradient of a river, the relative proportion of these
groups are also predicted to shift, reflecting food availability along the longitudinal
gradient (Wallace et al. 1997, Stone & Wallace 1998, Baer et al. 2001, Rosi-Marshall &
Wallace 2002). Specifically, headwater reaches in forested river networks are typically
heavily shaded, shallow, and cooler than downstream reaches. This riparian vegetation
can limit primary production and increase allochthonous inputs of coarse particulate
organic matter (CPOM, defined as organic matter >1mm), subsequently increasing the
reliance of macroinvertebrate consumers on terrestrial detritus (Vannote et al. 1980). As
macroinvertebrates in headwater streams consume this terrestrial input and convert it to
fine particulate organic matter (FPOM, defined as organic matter <1mm), it can increase
FPOM transport to downstream habitats, where organisms exploit this energy and to
maintain their productivity (Cummins et al. 1989, Vannote et al. 1980).
Although shifts in FFGs may be useful in determining the relative health of a
system, shifts in the diets of aquatic insects can provide even more insight into stream
function (Mihuc & Minshall 2005). However, FFG designation is confused with the
assumption that an insect definitively consumes a specific resource (e.g. all shredders eat
leaves). To the contrary, despite their FFG assignment, many insects have been found to
be generalist feeders (eating any abundant and available resource), or will switch

3
resources between seasons, during a disturbance event, or based on their developmental
stage (Cuffney & Minshall 1981, Mihuc & Minshall 1995, Wallace et al. 1997, Plague et
al. 1998, Rosi-Marshall & Wallace 2002). As a result, the diets of aquatic insects have
also been shown to track the availability of resources (Mihuc & Minshall 1995, Wallace
et al. 1997, Rosi-Marshall & Wallace 2002). Thus, the diets of aquatic insects should
reflect the available resources along the river continuum.
Analysis of the diets of aquatic insects could also provide insight into the effects
of disturbance on stream ecosystems, particularly changes in available food resources.
Global climate change (GCC), for example, has increased the frequency and spatial
extent of large-scale terrestrial disturbances (e.g., wildfire and beetle outbreaks)
(Westerling et al. 2003, 2006, Kurz et al. 2008). As a result, GCC may have
subsequently altered the structure of riparian vegetation and the relative importance of
allochthonous and autochthonous resources along the continuum. Such shifts may also
be reflected in the diets of insects along the river network. For instance, more frequent
wildfire that reduces riparian canopy cover may increase light flux to the stream surface,
increasing autotrophic production. In addition, reduced riparian cover may have
decreased inputs of terrestrial detritus, so that GCC may have increased the availability of
autochthonous resources along the continuum. Such habitat modifications linked to
global climate change have been observed in the western United States (Westerling et al.
2003, 2006, Kurz et al. 2008), and altered riparian vegetation may decrease allochthonous
inputs and increase in-stream production (Dwire & Kaufman 2003, Minshall 2003).
Accordingly, the diets of aquatic insects may track such changes, representing an

4
important tool for assessing how GCC may alter energy flows along the continuum.
Based on gut content analysis of specimens collected from Salmon River, ID sites
used in the testing of the RCC in 1976, I assessed whether aquatic insects in this system
followed the original predictions of the RCC. Based on these predictions, as well as
previous river continuum and diet analysis studies, I hypothesized that the diets of aquatic
insects collected in 1976 (archived specimens) and in 2009 would follow the predictions
of the RCC based on gut content analysis. That is, consumption of allochthonous
resources would be highest in the headwaters with consumption of autochthonous
resources increasing downstream. Second, the diets of aquatic insects would generally be
reflective of their FFG assignment (i.e. shredders would primarily consume leaf detritus
and grazers would consume algae). I also hypothesized that the diets of aquatic insects
from 1976 and 2009 would differ as a result of disturbances influenced by GCC,
specifically, increased incidence of fire and beetle kill would reduce the availability of
allochthonous resources in the headwaters (Mihuc & Minshall 2005). Thus, diet analysis
of aquatic insects from the past and present allowed me to be the first to actively test for
the predicted changes in resource use along the river continuum in 1976 (the original
RCC study) and 2009 (the RCC Redux study). It also allowed me to explore if aquatic
insect diets showed the effects of disturbances influenced by climate change in a three-
decade time span. To test my hypotheses, preserved specimens from the original RCC
study were obtained from Idaho State University’s archives. Aquatic insects were then
collected from the same RCC sites throughout summer and fall 2009, and individuals
from each site, season, and year were used for gut content analyses. These data provide

5
the first exploration of resource use in the Salmon River and examination of long-term
changes in resource use.

6
CHAPTER TWO
METHODS
Study sites
The Salmon River is located in the mountainous region of central Idaho (Figure
1). It flows 684 km from its headwaters to its confluence with the Snake River, making it
not only the largest tributary of the Snake, but also the longest free-flowing river in the
lower 48 States (Bureau of Land Management-Idaho 2010). Located in federally
designated wilderness areas, the Salmon River is unimpounded over its entire length and
has experienced minimal channelization, diversion, land use change, and anthropogenic
influence (Minshall et al. 1982). From its headwaters to its confluence with the Snake
River, there is an overall elevation drop of approximately 2000 m (Minshall et al. 1982).
Study sites were located in the Upper Salmon River basin in the Sawtooth
National Recreation Area near Stanley, Idaho in the Sawtooth Mountain Range of the
Idaho Rockies (Figure 1). Four sites were selected and sampled in 1976, with the
resulting data eventually used in the development and testing of the RCC (Vannote et al.
1980, Minshall et al. 1982, Minshall et al. 1983). We re-sampled these sites for the first
time in 2009. Distributed along 65 km of the river, the four sites were situated at the
headwaters of the Salmon River to just upstream of its confluence with the Yankee Fork
of the Salmon River (Minshall et al. 1982). Camp Creek (catchment area = 0.6 km2), a
second-order stream at approximately 2500 m elevation, served as the forested headwater

7
site (width of 1.1 m and a depth of 0.1 m) (Minshall et al. 1983). Fourth, fifth, and sixth-
order sites were also selected and are referred to as Smiley Creek, (catchment area = 40.6
km2, elevation: 2200 m), Obsidian (catchment area = 478.1 km
2, elevation: 2000 m), and
Casino (catchment area = 846. 3 km2, elevation: 1900 m). The stream width and depth of
Smiley is 6.3 m and 0.23 m, respectively. Stream width and depth of Obsidian and
Casino is 27.1 m and 0.39 m, and 42.9 m and 0.46 m, respectively (Minshall et al. 1983).
We were unable to sample at the exact location of the 1976 sampling at Obsidian in 2009,
due to expansion of the Sawtooth Fish Hatchery. As a result, we sampled approximately
50 m upstream of the original site, now the location of a water intake for the hatchery.
Camp Creek, Smiley Creek, Obsidian, and Casino will hereafter be referred to as Sites A,
B, C, and D, respectively, in regards to increasing stream order and catchment area from
A to D (e.g. Figure 2).
Snow accounts for the main source of precipitation from November to March and
enters the stream from about May to July (Minshall et al. 1983). Canopy cover was
greatest in the headwaters and progressively decreased downstream as the stream
widened. The riparian vegetation of the Upper Salmon River basin is largely dominated
by Pinus contorta (lodgepole pine), Pseudotsuga menziesii (Douglas fir), Salix sp.
(willow), and Populus tremuloides (quaking aspen) in the headwaters, with sagebrush
steppe ecosystem dominating the riparian zones of the lower sites. The Upper Salmon
River Basin has been affected by wildfire in recent years, and beetle kill has been
observed in riparian vegetation along our study reach (Bureau of Land Management-
Idaho 2010).

8
Figure 1. The Salmon River basin is located in central Idaho (inset). My sampling sites were
located in the upper Salmon River, in the Sawtooth National Recreation area near Stanley,
Idaho. Adapted from Minshall et al. 1982. Photo Credits: Satellite imagery: Google Earth
2011. Idaho/Salmon River Basin: USGS.
9
Taxa studied
During the original RCC sampling period, insects were collected from the Salmon
River in the summer (July) and winter (January) of 1976 were identified and categorized
by FFGs, data which were used in the development and testing of the RCC (Minshall et
al. 1982). A few specimens were also collected in September to November of 1976.
Aquatic invertebrates were sampled in 1976 as described by Minshall et al. (1982; 1983),
with insects collected on a single date in each season using a kick-net. Some of these
Figure 2. Allochthonous material in insect diets decreased downstream, while autochthonous
materials comprised a substantial amount of invertebrate diets at all sites in both years. Stream
order and catchment area increased from Camp (A), the headwater site, to Casino (D).
Community-weighted data was used to show the consumption of allochthonous (■),
autochthonous ( ), amorphous detritus (■), and animal material (■) at each site in 1976 and
2009. Percent of the community represented by analyzed taxa (based on community data from
Minshall et al. 1982), as well as sample size at each site (n) are also indicated.

10
specimens were archived at Idaho State University in the Stream Ecology Center’s
archives, and were viable for gut content analysis. Thus, although these insects were
collected from the Salmon River in 1976, their diet data was not examined until 33 years
later, in 2009. As only a limited number of taxa were archived, the number of taxa and
individuals used for gut content analysis was restricted to the specimens available from
the archived collection. The number of available taxa was highest for Camp Creek and
was more limited at downstream sites. Although preserved taxa were not consistently
available from Camp Creek to Casino, individuals used in the gut content analysis, in
general, comprised the majority of the macroinvertebrate assemblage (12%-65% of the
community; Figure 2). Samples collected in the summer (25-29 July 1976) and the fall
(23 September - 20 November 1976) were used to assess longitudinal patterns in insect
diets from the original RCC sampling data.
To assess whether potential shifts in terrestrial disturbance regimes (due to
wildfire or beetle kill) may have affected macroinvertebrate resource use, I collected
additional insect samples from each site in summer (July 16 to August 1) and fall
(September 29 to October 2) of 2009 using a coarse-mesh kick-net (800/900 µm) by
disturbing the substrate upstream of the net. Insects were preserved in Kahle’s solution
(Pennak 1978) in the field until the time of gut content analysis. Taxa were chosen for
diet analysis to provide both a means of comparison to the 1976 insects and a
longitudinal representation of present-day resource use from sites A to D. All individuals
were identified according to Merritt et al. (2008). When taxonomic classifications had

11
changed between the original 1976 sampling dates and 2009, recent taxonomic
classifications were used.
Overall, six shredder taxa, eleven collector-gatherer taxa, two collector-filterer
taxa, and three scraper-grazer taxa were analyzed in this study (Tables 1- 4). In total, gut
contents from twenty-two taxa preserved and/or found variably along the longitudinal
gradient were analyzed. Predators from 1976 and 2009 were excluded from this analysis,
as our main question concerned allochthonous and autochthonous resource use along the
river continuum and predators were presumed to have predominately animal material in
their diets (Minshall et al. 1982, Rosi-Marshall & Wallace 2002; personal observation).
Diet analysis
Gut content analysis was performed on individual insects collected in 1976 and
2009 (Benke & Wallace 1980). The foreguts of 1 to 15 individuals were dissected from
each taxon collected at each site in both years, depending upon availability. Gut contents
were sonicated and filtered onto 25mm, 0.45 um membrane filters that were then dried in
a drying oven at 60°C for at least 10 minutes. The dried filters were then affixed with
immersion oil to slides for gut content analysis. The number of individuals per slide
ranged from 1 to 5, based on the amount of material that was present in the individual
insects. Food particles were identified as leaf, wood, algae, amorphous detritus, fungi, or
animal, and ultimately assigned to the more general categories of allochthonous,
autochthonous, or amorphous detritus (Benke & Wallace 1980, Rosi-Marshall & Wallace
2002). The area of each food particle was then measured using ImageJ (National

12
Institutes of Health, Bethesda, Maryland, USA; http://imagej.nih.gov/ij/) to calculate the
relative proportion of food items in the diet(s) of those insect(s).
Statistical analysis
To assess how aquatic insect diets shifted along the longitudinal gradient and if
diets reflected FFG assignment, I grouped insects by FFG and compared their diets at
each sampling site. This comparison was based on both non-weighted diet data and
community weighted data. Using community composition data based on abundances
from Minshall et al. 1982, diet data were community-weighted to account for the relative
contribution of each taxa to the macroinvertebrate community of sites A-D and to
determine what proportion of the community was represented by the taxa analyzed
(Figure 2). Community-weighted data were used to compare seasonal resource use in
1976 and 2009 and explore the possibility of decadal shifts in the diets of aquatic insects.
Non-weighted data were used for statistical analyses to explore variation between sites
and individual taxa. To assess how diets shifted along the longitudinal gradient, I
compared the four FFGs (shredders, collector-gatherers, collector-filterers, and scraper-
grazers) at each site. Statistical analyses were used to analyze 2009 data; 1976 data were
precluded from statistical analyses due to insufficient sample size. I used statistical
analyses on the non-weighted seasonal 2009 data to explore diet differences between
seasons. 2009 allochthonous material diet data of each FFG and on the entire 2009
sampled community (p<0.05) were compared using one- or two-way ANOVAs in
SYSTAT, with site and season as the main effects. Site B was excluded from the
shredder and collector-filterer FFGs’ two-way ANOVA, due to lack of seasonal data

13
(Table 2). Separate one-way ANOVA were run on seasonal data to verify significance.
Only allochthonous material in gut contents was examined, as I was primarily concerned
with differences between allochthonous and autochthonous consumption, and the relative
proportions of the two resources are directly related to one another. When necessary, I
used the appropriate transformations (arcsine square-root) to meet statistical assumptions
(i.e. of ANOVA).

14
CHAPTER THREE
RESULTS
Community resource consumption and the predictions of the RCC
Overall, diets of individuals collected in 1976 and 2009 were similar between
years and across seasons (when samples were available and such a comparison could be
made) (Figure 2). However, in both years, the relative importance of allochthonous
materials in diets was significantly different among sites (F= 5.29, p=0.002, df=3),
whereby it was greatest at Camp Creek and decreased along the longitudinal gradient
(Figure 3a). Allochthonous materials in the diets of headwater insects were substantially
lower than the predicted presence of allochthonous resources in the headwater streams
(Figure 4a and b), representing about 40% of the overall diet. Similarly, insects in the
headwaters consumed large amounts of autochthonous materials (Fig. 3b). This pattern
was similar across all sites (Figure 3b) and was substantially greater than what was
predicted from the RCC (Fig. 4a and b). Amorphous detritus was found in similar
amounts in diets from sites B to D, with the lowest amounts of amorphous detritus found
in gut contents of insects collected from site A, Fall 2009 (Figure 3c, 4a, and 4b).
Consumption of animal material was highest at sites C and D in both seasons, mostly due
to the large amounts of animal material in the guts of the filter-feeding trichopteran
Brachycentrus (Tables 3 & 4).

15
Figure 3. Variation in the proportion of (a) allochthonous materials (b) autochthonous
materials, and (c) amorphous detritus among individual insect diets at each Salmon River, ID
site in 1976 (■) and 2009 (■). Consumption of all materials was similar at all sites in both
years. Sites are arranged from most upstream (Camp – A) to most downstream (Casino – D).
The solid line indicates the median, and the dotted line represents the mean. Dots indicate 5%
and 95% percentiles and whiskers show the range of data. The absence of whiskers or dots is
due to insufficient data.

16
Camp (A) Smiley (B) Obsidian (C) Casino (D)
% r
eso
urc
e found
in a
qua
tic insect die
ts
0
20
40
60
80
100
b Camp (A) Smiley (B) Obsidian (C) Casino (D) e
% p
resence o
f re
sourc
e p
redic
ed b
y R
CC
0
20
40
60
80
100a)
b)
Figure 4. Sites are arranged from most upstream (Camp – A) to most downstream (Casino –
D). (a) Hypothetical changes in the presence of allochthonous resources ( ) and
autochthonous resources ( ) along the longitudinal gradient, as predicted by the RCC:
allochthonous resources should be highest in the headwaters, decreasing downstream;
autochthonous resources should increase downstream. (b) Regression lines of allochthonous
(▬) and autochthonous (▬) resources in the diets of individual insects collected in 1976 and
2009 from the Salmon River, ID. Autochthonous material in diets is similar across sites, and
allochthonous material in diets decreases downstream; however, consumption of allochthonous
materials is not as dramatic as originally predicted. Dotted lines in 2b represent 95%
confidence intervals.

17
In terms of the community-weighted data comparisons, trends were similar
between 1976 and 2009 (Figure 3). Insects analyzed represented 12% (site D) to 65%
(site A) of the community in 1976, to 66% (site C) to 86% (site A) of the community in
2009; in all cases but one (i.e. site D in 1976), the majority of the assemblage was
sampled and analyzed. In diets, allochthonous material was highest in the headwaters
and decreased downstream, and autochthonous material was an important resource for the
macroinvertebrate community at all sites in both years (Figure 2), in a pattern loosely
reflective of the RCC’s predicted changes in the proportion of dominant resources along
the longitudinal gradient.
Functional feeding groups
Shredders
Compared to other FFGs, shredders consumed the most allochthonous material,
but they still relied substantially on autochthonous material, even in the most upstream
sites (sites A and B). Reliance on allochthonous material did not differ significantly
between sites (F=0.499, p-value=0.687, df=3) or seasons (F=3.81, p-value =0.062, df=1).
At least half of the shredder diets were composed of autochthonous material and
amorphous detritus, and this reliance did not appear to differ with sampling site.
Moreover, this overall reliance did not differ between 1976 and 2009.
Despite the overall trend in FFG diets, the relative reliance on allochthonous and
autochthonous material varied among taxa. Allochthonous material comprised roughly
half to the entire diet composition of Zapada, Eucapnopsis, and Capnia in 1976 and
2009, but the dominant headwater shredder, Yoraperla, consumed primarily algal

18
material (Table 1). In 1976, autochthonous material accounted for 69% of Yoraperla
diets (20% allochthonous) and 66% in 2009 (21% allochthonous). The sole shredder taxa
found at C and D in 2009, Pteronarcys, consumed 51% and 70% allochthonous material
at each site. Animal material also comprised a portion of their diets, particularly at site C
(27%) (Table 3).
Collector-gatherers
The diets of collector-gatherers were primarily composed of amorphous detritus
and autochthonous materials (Figure 5b), with limited contribution of allochthonous
material. This overall trend did not differ between 1976 and 2009 (Figure 2, Figure 3).
Although allochthonous material in the diets of collector-gatherers did not differ between
seasons (F=2.92, p=0.092, df=1), reliance on allochthonous material varied with
sampling location, whereby collectors consumed more allochthonous materials at site A
than at downstream sites (F= 4.72, p=0.005, df=3). Reliance on amorphous detritus and
autochthonous material was similar between sites. Despite these similarities in the
overall diets of collector-gatherers, diets did vary at the genera-level. Ephemerella
consumed the greatest amount of allochthonous materials at site A: 58% in fall 1976 and
69% in summer 2009 (Table 1). At the downstream sites, the closely related Drunella
(4%) and Serratella (8%) ate primarily autotrophic material and amorphous detritus
(Tables 2-4).
Collector-filterers
Collector-filterers exhibited high variability in the composition of their diets, but
in general had the highest amount animal material compared to other FFGs.

19
Allochthonous material in diets differed significantly across sites (F=5.70, p=0.004,
df=3), with significantly higher amounts of allochthonous material consumed at site A.
However, consumption of allochthonous material was not significantly different between
seasons (F=0.792, p=0.382, df=1). In 1976 and 2009, diets of Simulium at site A, the
only headwater collector-filterer analyzed, included 32% allochthonous materials
annually (Table 1). In simulid diets, allochthonous material in summer 2009 was not
significantly different across sites (F=2.30, p=0.194, df=3). Simulium gut contents
contained some animal material, and Brachycentrus diets included high amounts of
animal material (Tables 3 & 4). Collector-filterer diets at site C in 1976 differed from
those in 2009, with autochthonous material accounting for 73% of 1976 diets and animal
material comprising 65% of the 2009 collector-filters (Figure 2). This may be due, in
part, to Brachycentrus’ contribution of animal materials to the FFG collector-filterer
analysis, and the fact that Simulium was the only 1976 collector-filterer taxa available for
analysis at site C. At site D, allochthonous, autochthonous, and animal materials and
amorphous detritus were found in similar amounts in Brachycentrus gut contents (Table 3
& 4).
Scraper-grazers
The dominant resources comprising scraper-grazer diets were amorphous detritus
and autochthonous materials in similar proportions (Figure 5d), with limited contribution
of allochthonous material. Consumption of allochthonous materials was not significantly
different among sites (F=2.30, p=0.091, df=3), but did differ significantly between
seasons (F=4.96, p=0.032, df=1), with higher amounts of allochthonous materials

20
consumed in the fall. A comparison between scraper-grazer diet compositions in 1976
and 2009 was only possible at sites A and C. At site A in summer 1976, scraper-grazers
diets were comprised primarily of autochthonous materials and amorphous detritus
(Figure 5d). At site C in summer 1976 and 2009, the presence of allochthonous materials
in the gut contents of scraper-grazer was similar (5% and 8% respectively), but
consumption of autochthonous resources and amorphous detritus differed between the
two summers (Figure 5d, Table 3). Unlike the previous FFGs, consumption of
allochthonous materials by scraper-grazers was highest at site A only in summer 1976
(36%); the greatest amount of allochthonous materials was found in the diets of scraper-
grazers at site D in fall 2009 (42%).

21
Table 1. Percent of (un-weighted) allochthonous (AL), autochthonous (AU), amorphous
detritus (AD), and animal (AN) materials in the diets of individual taxa collected from Camp
Creek (Site A) in summer and fall 1976 and 2009. - - indicates unavailable data.
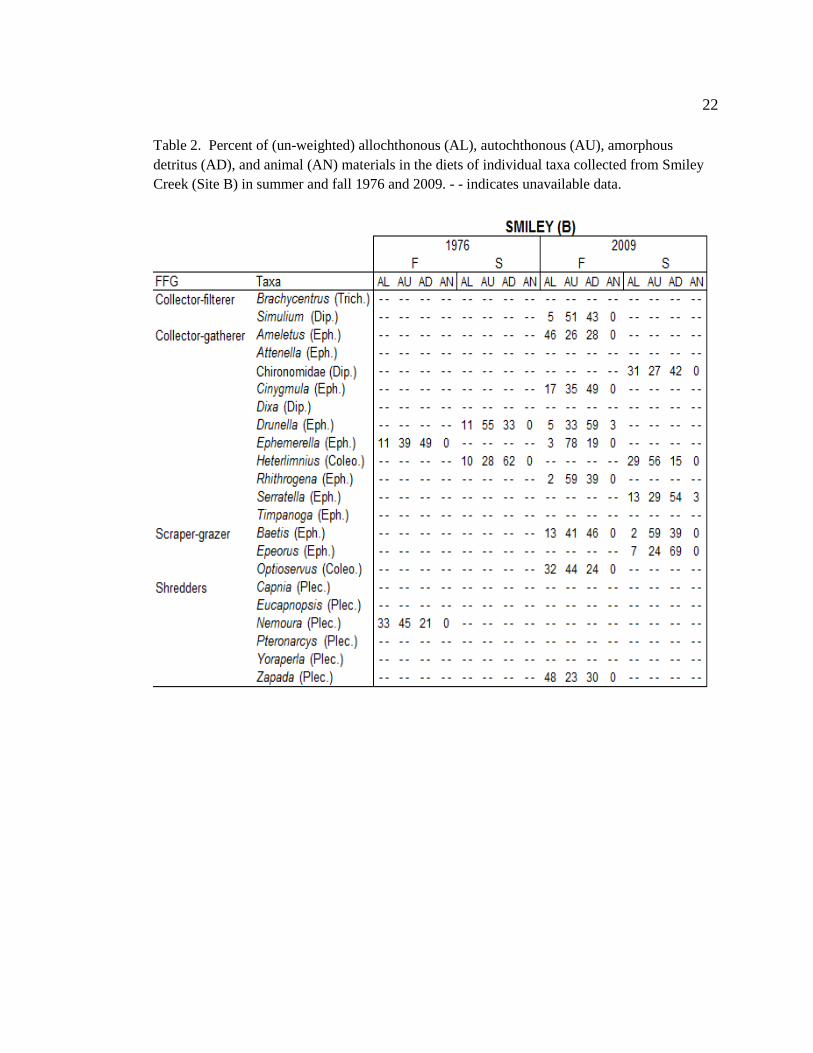
22
Table 2. Percent of (un-weighted) allochthonous (AL), autochthonous (AU), amorphous
detritus (AD), and animal (AN) materials in the diets of individual taxa collected from Smiley
Creek (Site B) in summer and fall 1976 and 2009. - - indicates unavailable data.

23
Table 3. Percent of (un-weighted) allochthonous (AL), autochthonous (AU), amorphous
detritus (AD), and animal (AN) materials in the diets of individual taxa collected from
Obsidian (Site C) in summer and fall 1976 and 2009. - - indicates unavailable data.

24
Table 4. Percent of (un-weighted) allochthonous (AL), autochthonous (AU), amorphous
detritus (AD), and animal (AN) materials in the diets of individual taxa collected from Casino
(Site D) in summer and fall 1976 and 2009. - - indicates unavailable data.

25
Figure 5. Sites are arranged from most upstream (Camp – A) to most downstream (Casino –
D). Proportion allochthonous (■), autochthonous ( ), amorphous detritus (■), and animal (■)
materials found in the diets of (a) shredders, (b) collector-gatherers, (c) collector-filterers, (d)
scraper-grazers collected from the Salmon River, ID in summer and fall 1976 and 2009.
Overall, shredders consumed the most allochthonous material, and autochthonous material was
consumed by all functional feeding groups (FFGs). In general, diets were similar across sites,
seasons, and years. Striped bars with “NA” indicate the absence of data. The * in (a) Camp
(A) F’76 represents the diet of one individual Capnia (Plec.).

26
CHAPTER FOUR
DISCUSSION
The River Continuum Redux
My results provide evidence that longitudinal patterns in macroinvertebrate diets
in the Salmon River are not consistent with the predictions of the RCC regarding changes
in dominant resources (Figure 4a and b). The RCC predicts that allochthonous resources
will support organisms in the headwaters and autochthonous resources will increase as
the river widens along the longitudinal gradient, peaking in mid-order reaches (Vannote
et al. 1980). However, gut content analysis of archived and present-day insects revealed
a different pattern. Although allochthonous resources were consumed in the greatest
amounts at the headwater site – an observation seemingly corresponding with the
predictions of the RCC – consumption of allochthonous materials was not as dramatic as
described in the RCC (Figure 4a and b), especially given that the dominant shredder
(Yoraperla) was highly algivorous. Although allochthonous inputs should be highest in
autumn when availability of leaf material is the highest, consumption of allochthonous
material was similar in the summer and post-leaf fall in 2009. Thus, this resource is
apparently not supporting macroinvertebrates to the extent that was originally presumed,
possibly due to a lack of retention of detrital resources (Minshall et al. 1992, Entrekin et
al. 2007, McNeely et al. 2007).
In contrast, autochthonous resources supported macroinvertebrate food webs at all

27
sites, in both years and seasons (Figure 2). These data, in conjunction with other studies
(Minshall 1978, Mayer & Likens 1987, Rosenfeld & Roff 1991, Thorp & Delong 1994 &
2002, McCutchan & Lewis 2002, McNeely et al. 2007), provide empirical evidence that
the importance of autochthony may be underestimated in stream ecosystems, particularly
in the headwaters (but see Vannote et al. 1980). The headwaters are presumed to have the
largest amounts of detrital material available to the macroinvertebrate community
(Vannote et al. 1980) and have been shown to support macroinvertebrate food webs in
some river basins (Rosi-Marshall & Wallace 2002). However, the diets of aquatic insects
collected in the headwaters of the Salmon River contained autochthonous material and
this resource often far exceeded the amount of allochthonous material consumed (Figure
3). Although not predicted by the RCC, macroinvertebrates in the headwaters may be
supported by algae (McNeely et al. 2007) particularly algae that are adapted to low-light
conditions. In addition, algae are considered to be a higher quality food resource than
detrital material (McNeely et al. 2007); in the presence of a thriving algal community,
aquatic insects may preferentially consume algae if possible, regardless of their FFG
classification. Thus, in the Salmon River, algae are not only a high quality food resource,
but are also fairly abundant, making it a viable food resource for much of the
macroinvertebrate community. Indeed, Minshall et al. (1983) reported that periphyton
chl-a was fairly high at all sites, and particularly high at the headwater site. Algal
resources appear to be extremely important to the Salmon River food webs and are of
greater importance in the headwaters than originally predicted by the RCC.

28
Global climate change signals in macroinvertebrate diet composition
Results from this study did not reveal a substantial shift in diet composition
between 2009 and 1976. This was contrary to my original prediction that more frequent
disturbances (e.g. fire) or increased unpredictability and severity of disturbance events
(e.g. flooding) associated with climate change would reduce riparian cover and the diets
of aquatic insects, particularly those in the headwaters (Dwire & Kaufman 2003,
Minshall 2003, Mellon et al. 2008). However, as insects in the Salmon River generally
appeared to be facultative feeders, this may have been a confounding factor in detecting a
climate signal. Additionally, insects may have preferentially been feeding on high-quality
algal material rather than potentially lower-quality detrital material (Ward & Cummins
1979, McNeely et al. 2007), even if it was not as readily available in 1976. If the
community was affected by environmental change, though, inflexible diets may have
caused the loss of certain taxa, leaving only generalist feeders to persist. Conversely,
riparian vegetation along the Salmon River simply may not have shifted to an extent that
may currently be measureable; therefore, a diet signal would not be expected to exist.
Because insect diets typically track resource availability (Cuffney & Minshall 1981,
Rosi-Marshall & Wallace 2002, McNeely et al. 2007), future comparisons of insect diets
to the data presented here may be useful.
Shifts in community composition associated with wildfire and loss of riparian
cover (Mihuc & Minshall 1995, Stone & Wallace 1998) have been observed in other
stream ecosystems, suggesting the potential for similar shifts associated with GCC-
induced shifts in these disturbance regimes (Mihuc & Minshall 1995, Stone & Wallace

29
1998, Gresswell 1999, Mihuc & Minshall 2005). Although I did not see shifts in overall
invertebrate diets, these disturbances may have altered the community composition of
macroinvertebrates. Analysis of community composition may provide additional insight
into how disturbances related to climate change affect invertebrate dynamics in the
Salmon River. My data on diets of macroinvertebrates collected 33 years apart provide a
strong framework upon which to develop hypotheses about which taxa may be sensitive
to change. For instance, some shredder taxa in the Salmon River headwaters consistently
ate leaves (i.e. Capnia and Eucapnopsis, Table 1); therefore, one may predict that these
taxa may be sensitive to changes in the amounts of allochthonous input or retention that
may occur as a result of climate change. Conversely, Yoraperla, which ate primarily
diatoms, and Zapada, which exhibit facultative plasticity (Mihuc & Minshall 1995), may
not be sensitive indicators.
Functional feeding groups and gut content analysis
In general, using FFGs to classify insects is useful inasmuch as they are used to
classify modes of feeding. FFGs should not be expected to accurately reflect of actual
resource consumption, nor should they be used as a replacement for gut content analysis
or stable isotope analysis when describing food webs. Previous studies have found that
FFGs are not reliable predictors of food resources consumed for many aquatic insects
(Benke & Wallace 1997, Mihuc & Minshall 1995, Plague et al. 1998, Rosi-Marshall &
Wallace 2002). Although some invertebrates can exhibit inflexible dietary preferences
(Eggert et al. 2003), other insects can be opportunistic feeders, exhibiting functional
plasticity and are capable of switching resources in the face of environmental

30
perturbation or disturbance (Mihuc & Minshall 1995; Dangles 2002). Possessing the
capacity to switch resources could provide a source of stability in an otherwise changing
ecosystem.
Similar to other studies, I hypothesized that invertebrate diets reflect functional
feeding group classification, (e.g. shredders consume mainly leaf material and grazers
consume mainly autotrophs) (Vannote et al. 1980). Although this was the case for a few
taxa, as has been found in prior studies (Cummins et al. 1973, Mihuc & Minshall 1995,
Baer et al. 2001), functional feeding group classification does not accurately predict diet
composition. In contrast to the predictions of the RCC, generalist feeders may be more
common along the continuum than insects that typify their assigned FFG than initially
thought “because mouthpart specialization does not necessarily indicate obligate resource
utilization” (Mihuc & Minshall 1995, Minshall 1988, Palmer et al 1993, Hall & Meyer
2008). That is, some insects may not be limited to a specific resource, despite their
morphology. For example, I found that shredders in the headwaters consumed large
amounts of autochthonous material in 1976 and 2009 (Figure 2, Table 1). Shredders ate
the most leaf material of all FFGs, but they still consumed large amounts of
autochthonous resources (Figure 5). Some shredder taxa (i.e. Capnia and Eucapnopsis)
ate large amounts of allochthonous material (Table 1), but the dominant shredder,
Yoraperla consumed primarily diatoms in 1976 and 2009. Previous studies have
demonstrated that Yoraperla consume detrital material (Dudley & Anderson 1982,
Hughes et al. 1999), but this study suggests they are facultative detritivores. The reliance
on autochthonous resources throughout the Salmon River by multiple FFGs strongly

31
indicates the potential problems of relying on FFGs to predict which resources support
macroinvertebrate food webs.
The diets of collector-gatherers, collector-filterers, and scraper-grazers were
similar among sites and between years, with autochthonous resources supporting the food
web. In addition, amorphous detritus was frequently consumed throughout the basin,
with the highest amounts found in diets from sites B, C, and D. Consumption of
amorphous detritus has been shown to increase along the longitudinal gradient in the
Little Tennessee River (Rosi-Marshall & Wallace 2002). Animal material was an
important resource at site D, which was primarily due to the influence of Brachycentrus,
a collecting-filtering trichopteran. According to Minshall et al. (1983), Brachycentrus
made 30% of the community composition at site D. Brachycentrus are generally
omnivores, grazing on algae and detritus from rocks and collecting seston from the water
column with their legs (Ross & Wallace 1981). It has been suggested that, due to the
structure of their highly-specialized legs, Brachycentrus is best suited to capture large
particles (e.g. animal material) from the water column, which would account for the large
amount of animal material found in their gut contents (Ross & Wallace 1981).
Food switching has been observed in many taxa analyzed in this study, including
Baetis, Drunella, and Zapada (Mihuc & Minshall 1995, Dangles 2002). Functional
plasticity is a common occurrence among aquatic insects, particularly in the event of
environmental disturbance (e.g. post-fire streams). Diet composition of 1976 and 2009
insects was similar among FFGs, sites, and seasons, which many indicate that many taxa

32
in the Salmon River are opportunistic feeders capable of facultative plasticity. In contrast
to other studies, these insects exhibited facultative plasticity under natural conditions.
Facultative diet-switching may help stabilize food webs
Opportunistic feeders that are tolerant of a wide range of conditions are well-
suited to surviving and thriving in the event of disturbance will help to provide stability
to a disturbed system (Mihuc & Minshall 1995, Gresswell 1999). Although assessments
of food web stability were beyond the scope of my study, theoretical and empirical
evidence suggests that my gut content results may provide insight into the potential
stability of the macroinvertebrate food web in the Salmon River. In the event of dramatic
changes to riparian vegetation (changes which may occur as a result of global climate
change), the biotic community, particularly in the headwaters, could be greatly affected.
However, if the macroinvertebrate community is composed of insects capable of
functional plasticity, those insects may not only survive, but thrive (Mihuc & Minshall
1995). Insects that are restricted in regards to food resources are more vulnerable and far
less resilient in the face of major environmental changes (Mihuc & Minshall 2005).
Indeed, dynamic food webs tend to encourage stability in an ecosystem (McCann 2000).
A food web composed solely of invertebrates with inflexible diets may be less likely to
persist when confronted with disturbances such as fire and beetle kill of riparian
vegetation. However, as has been demonstrated with Yoraperla in this study, more
insects may be capable of food switching than originally thought. This may indicate that
taxonomic diversity in post-disturbance streams may be more resilient than expected
(Stone & Wallace 1998), and taxa diversity has been shown to lend to the stability of a

33
system (McCann 2000); however, in such a potentially stressed system, there are
obviously other factors that must be considered (e.g. competition, changing stream
habitat). Regardless, in the western United States, where observable effects of climate
change are beginning to be recorded (Westerling et al. 2003, 2006, Kurz et al. 2008),
these opportunistic insects may eventually play an important role in contributing to a
thriving post-disturbance macroinvertebrate community and, in turn, the stability of the
stream ecosystem.
Hidden treasures: Archived insects and the value of invertebrate diet composition
While archived animal specimens, including organisms from fish to snakes, are
commonly utilized in diet composition analysis (Rodriguez-Robles 1998, Vander Zanden
et al. 2003, Shirey et al. 2008, Lavoie & Campeau 2010), this is, to my knowledge, the
first study to analyze the gut contents of archived aquatic insects. The ability to analyze
the diets of archived macroinvertebrates is an important tool when trying to determine the
effects of short or long-term environmental changes. It is also especially useful in the
event that isotope analysis is not possible, as was the case in this study. There were two
factors which precluded the use of isotope analysis in this study. First, isotope analysis
requires the destruction of the specimen in order to analyze it. As there were so few
archived 1976 taxa to begin with, the destruction of rare specimens and the resulting
depletion of the RCC archives were not feasible in this study. Second, the preservation
of the archived specimens may have affected the accuracy of isotope analysis. The 1976
specimens were first preserved in formalin, and some were later moved to ethanol for
long-term storage. Formalin is known to influence C and N isotopes (Hobson et al. 1997,

34
Bosley & Wainright 1999, Kaehler & Pakhomov 2001), although other studies have
found such effects to be negligible (Edwards et al. 2002) or correctable (Sarakinos et al.
2002). In addition, we were unable to definitively ascertain how the 1976 specimens
were prepared prior to preservation and subsequently treated in the years following.
Certain methods of preparation (i.e. acidification) have been shown to affect N (Jardine et
al. 2003, Jacob et al. 2005). Because of these factors, isotope analysis was not viable.
Such issues are not unique to this study; rather, they are issues that many
investigators may encounter when using archived specimens for diet composition
analysis. Museum specimens may be rare and therefore inappropriate to destroy, or the
means of preparation or preservation may interfere with isotope analysis. In light of
these issues, the advantage of gut content analysis when examining diet composition is
clear. Gut content analysis does not require destruction of the entire specimen, nor does
the type of preservative affect the analysis. The digestive tract of the specimen must be
sacrificed, but the rest of the specimen will mostly remain intact. Gut content analysis
also allows the investigator to see and measure the actual food resources in an
individual’s diet (Rosi-Marshall & Wallace 2002, Vander Zanden et al. 2003, Cross et al.
2007). Archived specimens, then, are not only important in their own right, but
individual specimens are also essentially archives in and of themselves. Insects, such as
those in this study, are collectors of diatoms and fungi and, much like diet composition
and macroinvertebrate community composition, diatom community composition may be
a sensitive indicator of environmental change (McCormick & Cairns 1994, Gregory-
Eaves et al. 1999, Saros et al. 2003). In this study, diatom community composition in

35
1976 was similar to 2009 in the Salmon River (Sylvia Lee, Personal correspondence,
2011). Of course, when possible, gut content analysis and isotope analysis used in
conjunction with one another may provide more insight into food resource utilization
than either analysis used alone.
Conclusions
Gut content analysis showed that diets did not follow the predictions of the RCC
and, in addition, FFGs did not prove to be reliable indicators of resources consumed.
Although the implementation of the RCC and FFGs in examining stream ecosystems and
the relative importance of basal resources may be useful in principle, they may be
misleading when used alone. Broad generalizations based on the RCC and FFGs may
lead us to underestimate the importance of certain resources in stream ecosystems and
should be used with caution. In contrast, gut content analysis can provide useful insights,
and such analyses are critical in understanding energy flow in an ecosystem.
Comparisons between past and present-day specimens can provide a means of
establishing long-term data sets. Although I was unable to detect a climate signal in this
study, long-term data sets may reveal the effects of environmental changes on stream
ecosystems. Care must be taken to consider other factors (e.g. extent of change over
time, individual characteristics of a system) when using diet analysis to detect signals of
change. Diet analysis and archived specimens can be important tools in exploring energy
flow and the effects of environmental change on ecosystems, particularly in the changing
global environment in which we live today, and especially when these changes span
multiple decades.

36
LITERATURE CITED
Allan, J. David. 2007. Stream Ecology: Structure and function of running waters. 2nd
ed.
New York: Springer.
Baer, Sara G., Edward R. Siler, Susan L. Eggert, and J. Bruce Wallace. 2001.
Colonization and production of macroinvertebrates on artificial substrata:
upstream-downstream responses to a leaf litter exclusion manipulation.
Freshwater Biology 46: 347-365.
Bates, B.C., Z.W. Kundzewicz, S. Wu and J.P. Palutikof, (Eds.), 2008: Climate Change
and Water. Technical Paper of the Intergovernmental Panel on Climate Change,
IPCC Secretariat, Geneva, 210 pp.
Benke, A.C. and J. B. Wallace. 1980. Trophic basis of production among net-spinning
caddisflies in a Southern Appalachian stream. Ecology 61: 108-118.
Boise State University Environmental Science and Public Policy Research Institute.
Wildfire, bugs, and you: An informational brochure, Sawtooth National Forest.
Ketchum, ID: USDA, 2004.
Bosley, Keith L. and Sam C. Wainright. 1999. Effects of preservatives and acidification
on the stable isotopes (15N:14N, 13C:12C) of two species of marine animals.
Can. J. Fish. Aquat. Sci. 56: 2181-2185.
Carpenter, Stephen R., Stuart G. Fisher, Nancy B. Grimm, and James F. Kitchell. 1992.
Global change and freshwater ecosystems. Annual Review of Ecology and
Systematics 23: 119-139.
Cross, Wyatt F., J. Bruce Wallace, and Amy D. Rosemond. 2007. Nutrient enrichment
reduces constraints on material flows in a detritus-based food web. Ecology 88:
2563-2575.
Cuffney, Thomas E. and G. Wayne Minshall. 1981. Life history and bionomics of
Arctopsyche grandis (Trichoptera) in a Central Idaho stream. Holarctic Ecology
4: 252-262.
Cummins, Kenneth W. 1973. Trophic relations of aquatic insects. Annual Review of
Entomology 18: 183-206.

37
Cummins, Kenneth W. and Michael J. Klug. 1979. Feeding ecology of stream
invertebrates. Annual Review of Ecological Systems 10: 147-172.
Cummins, Kenneth W., Margaret A. Wilzbach, Donna M. Gates, Joy B. Perry, W. Bruce
Taliaferro. 1989. Shredders and riparian vegetation. Bioscience 39: 24-30.
Dallas, Helen. 2008. Water temperature and riverine ecosystems: An overview of
knowledge and approaches for assessing biotic responses, with special reference
to South Africa Water SA 34: 393-405.
Dangles, Olivier. 2002. Functional plasticity of benthic macroinvertebrates: implications
for trophic dynamics in acid streams. Can. J. Fish. Aquat. Sci. 59: 1563-1573.
Dudley, Tom and Norman H. Anderson. 1982. A survey of invertebrates associated with
wood debris in aquatic habitats. Melanderia 39: 1-22.
Dwire, K.A. and J.B. Kauffman. 2003. Fire and riparian ecosystems in landscapes of the
western USA. Forest Ecology and Management 178: 61-78.
Edwards, Melanie S., Thomas F. Turner, Zachary D. Sharp, and W.L. Montgomery.
2002. Short- and long-term effects of fixation and preservation on stable isotope
values (ɗC, ɗN, ɗS) of fluid-preserved museum specimens. Copeia 4: 1106-1112.
Entrekin, Sally A., Emma J. Rosi-Marshall, Jennifer L. Tank, Timothy J. Hoellein, and
Gary A. Lamberti. 2007. Macroinvertebrate secondary production in 3 forested
streams of the upper Midwest, USA. J. N. Am. Benthol. Soc. 26: 472-490.
Gregory-Eaves, Irene, John P. Smol, Bruce P. Finney, and Mary E. Edwards. 1999.
Diatom-based transfer functions for inferring past climatic and environmental
changes in Alaska, U.S.A. Arctic, Antarctic, and Alpine Research 31: 353-365.
Gresswell, Robert E. 1999. Fire and aquatic ecosystems in forested biomes of North
America. transactions of the American Fisheries Society 128: 193-221.
Hall, Robert O. and Judy L. Meyer. 1998. The trophic significance of bacteria in a
detritus-based food web. Ecology 79: 1995-2012.
Hawkins, Charles P. and James R. Sedell. 1981. Longitudinal and seasonal changes in
functional organization of macroinvertebrate communities in four Oregon
streams. Ecology 62: 387-397.

38
Hobson, Keith A., Mark L. Gloutney, and H. Lisle Gibbs. 1997. Preservation of blood
and tissue samples for stable-carbon and stable-nitrogen isotope analysis. Can. J.
Zool. 75: 1720-1723.
Hogg, I.D., D.D. Williams, J.M. Eadie, S.A. Butt. 1995. The consequences of global
warming for stream invertebrates: A field simulation. Journal of Thermal Biology
20: 199-206.
Hughes, Jane M., Peter B. Mather, Andrew L. Sheldon, Fred W. Allendorf. 1999.
Genetic structure of the stonefly, Yoraperla brevis, populations: the extent of gene
flow among adjacent montane streams. Freshwater Biology 41: 63-72.
Jardine, T. D., S.A. McGeachy, C.M. Paton, M. Savoie, and R.A. Cunjak. 2003. Stable
isotopes in aquatic systems: Sample preparation, analysis, and interpretation.
Can. Manuscr. Rep. Fish. Aquat. Scie. No. 2656: 39 p.
Jacob, Ute, Katja Mintenbeck, Thomas Brey, Rainer Knust, and Kerstin Beyer. 2005.
Stable isotope food web studies: a case for standardized sample treatment.
Marine Ecology Progress Series 287: 251-253.
Kaehler S. and E.A Pakhomov. 2001. Effects of storage and preservation on the δ13
C
and δ15
N signatures of selected marine organisms. Marine Ecology Progress
Series 219: 299-304.
Karl, Thomas R. and Kevin E. Trenberth. 2003. Modern global climate change. Science
302: 1719-1723.
Kurz, W.A., C.C. Dymond, G. Stinson, G.J. Rampley, E.T. Neilson, A.L. Carroll, T.
Ebata, L. Safranyik. 2008. Mountain pine beetle and forest carbon feedback to
climate change. Nature 452: 987-990.
Lavoie, Isabelle and Stephane Campeau. 2010. Fishing for diatoms: fish gut analysis
reveals water quality changes over a 75-year period. J. Paleolimnol 43: 121-130.
Li, Aggie O.Y. and David Dudgeon. 2008. Food resources of shredders and other
benthic macroinvertebrates in relation to shading conditions in tropical Hong
Kong streams. Freshwater Biology 53: 2011-2025.
Mayer, Marilyn S. and Gene E. Likens. 1987. The importance of algae in a shaded
headwater stream as food for an abundant caddisfly (Trichoptera). J. N. Am.
Benthol. Soc. 6: 262-269.

39
McCann, Kevin S. 2000. The diversity-stability debate. Nature 45: 228-233.
McCormick, Paul V. and John Cairns, Jr. 1994. Algae as indicators of environmental
change. Journal of Applied Phycology 6: 509-526.
McCutchan, Jr. James H. and William H. Lewis, Jr. 2002. Relative importance of
carbon sources for macroinvertebrates in a Rocky Mountain stream. Limnol.
Oceanogr. 47: 742-752.
McPeek, Mark A. and Barbara L. Peckarsky. 1998. Life histories and the strengths of
species interactions: combining mortality, growth, and fecundity effects. Ecology
79: 867-879.
McNeely, Camille, Jacques C. Finlay, and Mary E. Power. 2007. Grazer traits,
competition, and carbon sources to a headwater-stream food web. Ecology 88:
391-401.
Mellon, Cassie D., Mark S. Wipfli, and Judith L. Li. 2008. Effects of forest fire on
headwater stream macroinvertebrate communities in eastern Washington, USA.
Freshwater Biology 53: 2331-2343.
Merritt, Richard W., Kenneth W. Cummins, Martin B. Berg. 2008. An Introduction to
the Aquatic Insects of North America. Dubuque, IA, USA: Kendall Hunt Pub Co.
Meyer, Judy L., Michael J. Sale, Patrick J. Mulholland, N. LeRoy Poff. 1999. Impacts
of climate change on aquatic ecosystem functioning and health. Journal of the
American Water Resources Association 35: 1373-1386.
Mihuc, Timothy B. and G. Wayne Minshall. 1995. Trophic generalists vs. trophic
specialists: Implications for food web dynamics in post-fire streams. Ecology 76:
2361-2372.
Mihuc, Timothy B. and G. Wayne Minshall. 2005. The trophic basis of reference and
post-fire stream food webs 10 years after wildfire in Yellowstone National Park.
Aquatic Sciences 67: 541-548.
Minshall, G. Wayne. 1978. Autotrophy in stream ecosystems. Bioscience 28: 767-771.

40
Minshall, G. Wayne, James T. Brock, Thomas W. LaPoint. 1982. Characterization and
dynamics of benthic organic matter and invertebrate functional feeding group
relationships in the Upper Salmon River, Idaho (USA). International Revue ges.
Hydrobiologie 67: 793-820.
Minshall, G. Wayne, Robert C. Petersen, Kenneth W. Cummins, Thomas L. Bott, James
R. Sedell, Colbert E. Cushing, Robin L. Vannote. 1983. Interbiome Comparison
of Stream Ecosystem Dynamics. Ecological Monographs 53: 2-25.
Minshall, G. Wayne. 2003. Responses of stream benthic macroinvertebrates to fire.
Forest Ecology and Management 178: 155-161.
O’Driscoll, Michael A. and David R. DeWalle. 2006. Stream-air temperature relations to
classify stream-ground water interactions in a karst setting, central Pennsylvania,
USA. Journal of Hydrology 1-2: 140-153.
Palmer, C. J. O’Keefe, A. Palmer, T. Dunne, and S. Radloff. Macroinvertebrate
functional feeding groups in the middle and lower reaches of the Buffalo river,
eastern Cape, South Africa. I. Dietary variability. Freshwater Biology 29: 441-
453.
Parmesan, Camille. 2006. Ecological and evolutional responses to recent climate
change. Annual Review of Ecology, Evolution, and Systematics 37:637-69.
Pennak, R.W. 1978. Freshwater invertebrates of the Unites States. 2nd
ed. New York:
John Wiley & Sons.
Plague, Gordon R., J. Bruce Wallace, Jack W. Grubaugh. 1998. Linkages between
trophic variability and distribution of Pteronarcys spp. (Plecoptera:
Pteronarcyidae) along a river continuum. American Midland Naturalist 139: 224-
235.
Powell, James A. and Jesse A. Logan. 2005. Insect seasonality: Circle map analysis of
temperature-driven life cycles. Theoretical Population Biology (in press).
Pritchard, Gordon, Lawrence Harder, Robert Mutch. 1996. Development of aquatic
insect eggs in relation to temperature and strategies for dealing with different
thermal environments. Biological Journal of the Linnean Society 58: 221-244.
Robinson, C.T., G.W. Minshall, T.V. Royer. 2000. Inter-annual patterns in
macroinvertebrate communities of wilderness streams in Idaho, USA.
Hydrobiologia 421: 1987-198.

41
Rodriguez-Robles, Javier A. 1998. Alternative perspectives on the diet of gopher snakes
(Pituophis catenifer, Colubridae): Literature records versus stomach contents of
wild and museum specimens. Copeia 2: 463-466.
Rosenfeld, J.S. and J.C. Roff. 1991. Primary production and the availability of
autochthonous carbon in southern Ontario streams. Hydrobiologia 224: 99-109.
Rosi-Marshall, Emma and J. Bruce Wallace. 2002. Invertebrate food webs along a
stream resource gradient. Freshwater Biology 47: 129-141.
Rosi-Marshall, Emma J. and Judy L. Meyer. 2004. Quality of suspended fine particulate
matter in the Little Tennessee River. Hydrobiologia 519: 29-37.
Ross, Douglas H. and J. Bruce Wallace. 1981. Production of Brachycentrus spinae Ross
(Trichoptera: Brachycentridae) and its role in seston dynamics of a southern
Appalachian stream (USA). Environ. Entomol. 10: 240-246.
Sarakinos, Helen C., Michael L. Johnson, and M. Jake Vander Zanden. 2002. A
synthesis of tissue-preservation effects on carbon and nitrogen stable isotope
signatures. Can. J. Zool. 80: 381-387.
Saros, Jasmine E., Sebastian J. Interlandi, Alexander P. Wolfe, and Daniel R. Engstrom.
2003. Recent changes in diatom community structure of lakes in the Beartooth
Mountain Range, U.S.A. Arctic, Antarctic, and Alpine Research 35: 18-23.
Shepard, Richard B. and G. Wayne Minshall. 1984. Role of benthic insect feces in a
Rocky Mountain stream: fecal production and support of consumer growth.
Holartic Ecology 7: 119-127.
Shirey, Patrick D., David E. Cowley, Rossana Sallenave. 2008. Diatoms from gut
contents of museum specimens of an endangered minnow suggest long-term
ecological changes in the Rio Grande (USA). J. Paleolimnol. 40: 263-272.
Solomon, S. D. Qin, M. Manning, Z. Chen, M. Marquis, K.B. Averyt, M. Tignor, and
H.L. Miller (eds.). 2007. Contribution of working group I to the fourth
assessment report of the Intergovernmental Panel on Climate Change. New York:
Cambridge UP.
Statzner, Bernhard and Bert Higler. 1985. Questions and Comments on the River
Continuum Concept. Canadian Journal of Fisheries and Aquatic Sciences 42:
1038-1044.

42
Stone, Michael K. and J. Bruce Wallace. 1998. Long-term recovery of a mountain
stream from clear-cut logging: the effects of forest succession on benthic
invertebrate community structure. Freshwater Biology 39: 151-169.
Sweeney, Bernard W. and Robin L. Vannote. 1978. Size variation and the distribution
of hemimetabolous aquatic insects: Two thermal equilibrium hypotheses. Science
200: 444-446.
Sweeney, Bernard W., John K. Jackson, J. Denis Newbold, and David H. Funk. 1992.
Climate change and the life histories and biogeography of aquatic insects in
Eastern North America. 143-176 in: Global Climate Change and Freshwater
Ecosystems. Eds. Penelope Firth and Stuart G. Fisher. New York: Spring-
Verlag.
Thorp, James H. and Michael D. Delong. 1994. The riverine productivity model: An
heuristic view of carbon sources and organic processing in large river ecosystems.
Oikos 70: 305-308.
Thorp, James H. and Michael D. Delong. 2002. Dominance of autochthonous
autotrophic carbon in food webs of heterotrophic rivers. Oikos 96: 543-550.
U.S. Department of the Interior: Bureau of Land Management (Idaho). Lower Salmon
River. Idaho Bureau of Land Management, 2010. Web. 2010.
Vannote, Robin L., G. Wayne Minshall, Kenneth W. Cummins, James R. Seddell,
Colbert E. Cushing. 1980. The River Continuum Concept. Canadian Journal of
Fisheries and Aquatic Sciences 37: 130-137.
Vander Zanden, M. Jake., Sudeep Chandra, Brant C. Allen, John E. Reuter, and Charles
R. Goldman. 2003. Historical food web structure and restoration of native
aquatic communities in the Lake Tahoe (California-Nevada) basin. Ecosystems
6: 274-288.
Wallace, J.B., S.L. Eggert, J.L. Meyer, J.R. Webster. 1997. Multiple trophic levels of a
forest stream linked to terrestrial litter inputs. Science 277: 102-104.
Walther, Gian-Reto, Eric Post, Peter Convey, Annette Menzel, Camille Parmesan, Trevor
J.C. Beebee, Jean-Marc Fromentin, Ove Hoegh-Guldberg, and Franz Bairlein.
2002. Ecological responses to recent climate change. Nature 416: 389-395.
Wang, Feiyue, Andre Tessier, and Landis Hare. 2001. Oxygen measurements in the
burrows of freshwater insects. Freshwater Biology 46: 317-327.

43
Ward, G. Milton and Kenneth W. Cummins. 1979. Effects of food quality on growth of
a stream detritivore, Paratendipes albimanus (Meigen) (Diptera: Chironomidae).
Ecology 60: 57-64.
Ward, James V. and Jack A. Stanford. 1982. Thermal responses in the evolutionary
ecology of aquatic insects. Annual Review of Entomology 27:97-117.
Westerling, A.L., A. Gershunov, T.J. Brown, D.R. Cayan, and M.D. Dettinger. 2003.
Climate and wildfire in the western United States. American Meteorological
Society 84: 595-604.
Westerling, A.L, H.G. Hidalgo, D.R. Cayan, T.W. Swetnam. 2006. Warming and earlier
spring increase western U.S. forest wildfire activity. Science 313: 940-943.
Wotton, Roger S (ed). 1994. The biology of particles in aquatic systems. Boca Raton,
FL: CRC Press.
Wotton, Roger S. 1996. Colloids, bubbles, and aggregates: a perspective on their role in
suspension feeding. Journal of the North American Benthological Society 15:
127-135.

44
VITA
Kathryn Vallis was born and raised in Calumet City, Illinois. After graduating
from Marian Catholic High School in 2003, she enrolled at Loyola University Chicago.
There, she volunteered in Dr. Emma Rosi-Marshall’s lab, sorting and identifying
caddisflies for a graduate research project. She graduated with a Bachelor of Science in
Biology with a minor in English in May 2007.
In fall 2008, Kathryn received a Teaching Assistantship at Loyola University
Chicago, where she began her Master’s thesis research investigating resource use of
aquatic insects, under the direction of Dr. Emma Rosi-Marshall. She served as a teaching
assistant for two semesters at Loyola University Chicago and participated in the
Fundamentals of Ecosystem Ecology course at the Cary Institute of Ecosystem Studies in
January 2009. Kathryn spent the summer of 2009 collecting insects and working on the
Salmon River, Idaho, with Dr. Colden Baxter and the Idaho State University Stream
Ecology Center. In January 2010, she moved with the Rosi-Marshall lab to the Cary
Institute of Ecosystem studies, where she finished her thesis work. Kathryn received the
“Best poster presentation in applied research (2009)” and “Best oral presentation in
applied research: Runner-up (2010)” awards and the “President’s Endowment Award
(2011)” from the North American Benthological Society for her Master’s work.
