the pathogenesis of fatal outcome in murine pulmonary
TRANSCRIPT

INFECTION AND IMMUNITY, Jan. 2005, p. 114–125 Vol. 73, No. 10019-9567/05/$08.00�0 doi:10.1128/IAI.73.1.114–125.2005Copyright © 2005, American Society for Microbiology. All Rights Reserved.
The Pathogenesis of Fatal Outcome in Murine PulmonaryAspergillosis Depends on the Neutrophil
Depletion StrategyShane D. Stephens-Romero,1† Aron J. Mednick,1† and Marta Feldmesser1,2,3*
Division of Infectious Diseases, Department of Medicine,1 Department of Microbiology &Immunology,2 and Department of Obstetrics & Gynecology and Women’s Health,3
Albert Einstein College of Medicine, Bronx, New York
Received 18 August 2004/Returned for modification 2 September 2004/Accepted 10 September 2004
Aspergillus fumigatus causes invasive disease in severely immunocompromised hosts but is readily clearedwhen host innate defenses are intact. Animal models for evaluation of therapeutic strategies to combat invasiveaspergillosis that closely mimic human disease are desirable. We determined optimal dosing regimens forneutrophil depletion and evaluated the course of infection following aerosol infection in mice by determiningsurvival, organ fungal burden, and histopathology in mice in which neutropenia was induced by three methods,administration of granulocyte-depleting monoclonal antibody RB6-8C5 (MAb RB6), administration of cyclo-phosphamide, and administration of both agents. Administration of either individual agent resulted in arequirement for relatively high conidial inocula to achieve 100% mortality in both BALB/c and C57BL/6 mice,although the infection appeared to be somewhat more lethal in C57BL/6 mice. Death following induction ofneutropenia with MAb RB6 occurred when a relatively low fungal burden was present in the lung and may havebeen related to the inflammatory response associated with neutrophil recovery. In contrast, administration ofboth agents reduced the lethal inoculum in each mouse strain by approximately 1 log10, and C57BL/6 mice thatreceived both agents had a higher fungal burden and less inflammation in the lung at the time of death thanBALB/c mice or mice of either strain that received MAb RB6 alone. Our data suggest that the relationshipamong fungal burden, inflammation, and death is complex and can be influenced by the immunosuppressionregimen, the mouse strain, and the inoculum.
Aspergillus fumigatus is a ubiquitous mold that causes inva-sive disease in severely immunocompromised patients. Theimportant risk factors for development of invasive aspergillosisinclude neutropenia, defective neutrophil function, the use ofcorticosteroids or other immunosuppressive therapies, such asthose used to prevent rejection following organ transplanta-tion, and late-stage human immunodeficiency virus infection(5, 12, 13, 30, 41, 54). The organism, aerosolized from soil, airfiltration, or water sources, is inhaled, and the most commonsite of primary infection is the lung (1, 40, 48, 58). The inoc-ulum required for disease production is unknown and may bedependent upon the immune defect of the affected host (14).Innate immune responses control infection in a normal host.Conidia are phagocytosed by alveolar macrophages, but in theabsence of effective macrophage function, the conidia germi-nate, and the organism resumes hyphal growth (51). Neutro-phils are rapidly recruited to the lung in response to experi-mental pulmonary infection with A. fumigatus (17) andeffectively kill hyphae that result from escape from conidialkilling by macrophages (51). The importance of NK cells aseffectors during the initial response to A. fumigatus has beendemonstrated recently (39).
Models for invasive aspergillosis have been established in
mice, rats, rabbits, and guinea pigs (3, 18, 31, 53). When pos-sible, these models have incorporated pulmonary rather thanintravenous (i.v.) deposition of conidia because of the route ofinfection in humans (6, 9, 35). Neutropenia, chemotherapy,and glucocorticoid use are important risk factors for invasiveaspergillosis in humans. Consequently, investigators haveestablished a number of experimental models with immuno-compromised mice using a granulocyte-depleting monoclonalantibody (MAb), MAb RB6-8C5 (MAb RB6), cyclophospha-mide, an alkylating agent that is commonly used for cancerchemotherapy, and a variety of glucocorticoid preparations (8,11, 36, 38, 55). The effects of these agents on host immuneresponses are different. To understand the way in which themethod of neutropenia induction alters the pathogenesis ofexperimental invasive pulmonary aspergillosis, we evaluatedthe effects of such agents on infection following aerosol expo-sure in a model patterned after that developed by Piggott andEmmons (47) and Brieland et al. (6).
MATERIALS AND METHODS
Immunosuppressive agents. The rat hybridoma producing MAb RB6 was agenerous gift from Robert L. Coffman (DNAX Research Institute of Molecularand Cellular Biology, Inc., Palo Alto, Calif.). Ascites containing MAb RB6 wasproduced by intraperitoneal (i.p.) injection of hybridoma cells into pristine-primed SCID mice (National Cancer Institute, Bethesda, Md.) and was sterilizedby filtration with a 0.2-�m-pore-size filter. The concentration of MAb RB6 wasdetermined by an enzyme-linked immunosorbent assay (ELISA) by using apurified rat immunoglobulin G2b (IgG2b) standard (PharMingen, BD Bio-sciences, San Diego, Calif.). Cyclophosphamide, triamcinolone, and methotrex-
* Corresponding author. Mailing address: Albert Einstein Collegeof Medicine, Forchheimer Building, Room 402, 1300 Morris ParkAvenue, Bronx, NY 10461. Phone: (718) 430-3487. Fax: (718) 430-8968. E-mail: [email protected].
† S.D.S.-R. and A.J.M. contributed equally to this study.
114
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 21
Jan
uary
202
2 by
260
4:3d
09:a
17c:
3100
:e0b
d:1d
7f:c
91a:
e019
.

ate were obtained from Sigma (St. Louis, Mo.). Dexamethasone was obtainedfrom ICN Biomedicals, Inc. (Irvine, Calif.).
Mice. Specific-pathogen-free female BALB/c, C57BL/6, A/JCr, and DBA/2mice were obtained from the National Cancer Institute. BALB/c and A/JCr micehave the Ly6.1 haplotype, while C57BL/6 and DBA-2 mice express the Ly6.2haplotype (34, 44). The mice weighed 16 to 18 g at the time of experimentation,except where indicated below. The mice were housed in microisolator cages in apathogen-free barrier facility at the Albert Einstein College of Medicine.
Mice were inoculated via the lateral tail vein or i.p. with MAb RB6 diluted insterile phosphate-buffered saline (PBS). The mice were bled from the orbitalsinus into Eppendorf tubes containing 2 �l of 10% EDTA (pH 7.4). The bloodwas diluted 1:20 in Turk’s solution (1% glacial acetic acid and 0.01% gentianviolet in distilled H2O), and the total number of white blood cells (WBCs) wasdetermined by using a hemacytometer (37). WBC differential counts were de-termined on smears of whole blood stained with the Hema3 staining system(Fisher Scientific, Biochemical Sciences, Inc., Swedesboro, N.J.) used accordingto the manufacturer’s instructions. At least 100 WBCs were counted per slide.The total WBC counts and differentials were used for calculation of the absoluteneutrophil count (ANC) and the absolute lymphocyte count (ALC).
Mouse serum was tested for MAb RB6-binding antibody by an ELISA. MAbRB6 obtained from concentrated cell supernatants was purified by protein Gchromatography. Polystyrene plates were coated with a solution containing 5 �gof MAb RB6 per ml, and following incubation for 1 h at 37°C, wells were blockedwith PBS containing 2% bovine serum albumin (BSA). Serum samples wereadded to the wells and then serially diluted, beginning with a 1:50 dilution. Afterincubation and washing, goat anti-mouse IgM or IgG was added at a concentra-tion of 1 �g/ml. p-Nitrophenyl phosphate in carbonate substrate buffer wasadded to the wells, and the A405 was determined by using a Multiskan microplatereader (Labsystems, Franklin, Mass.).
Flow cytometry. Five mice were treated with 25 �g of MAb RB6, five micewere treated with 150 mg of cyclophosphamide per kg, and five mice were treatedwith both agents and then killed 1 day later. Five naı̈ve mice served as a control.The spleens were removed, and single-cell suspensions were prepared by ho-mogenization in Hanks’ balanced salt solution. Following centrifugation at 290 �g for 10 min at 4°C and washing, the pellet was passed through a 70-�m-pore-sizefilter. Red blood cells were lysed by incubation of the cell suspension in 0.17 MNH4Cl (pH 7.0) at 4°C for 10 min. Fluorochrome- or biotin-conjugated MAbs tocell surface markers or isotype-matched controls were produced in rats, except asindicated below, and were obtained from Pharmingen (San Diego, Calif.) (CD3-allophycocyanin (APC) [hamster IgG], CD4-fluorescein isothiocyanate (FITC)[IgG2b], CD8a-phycoerythrin (PE) [IgG2a], CD19-biotin [IgG2a], Mac-3-PE[IgG1], CD11c-APC [hamster IgG]) or Caltag Laboratories (Burlingame, Calif)(IgG1-biotin, IgG2a-FITC, IgG2b-APC, and hamster IgG-biotin). MAb RB6was purified from concentrated cell supernatants by protein G chromatography,conjugated to Alexa-488 (Molecular Probes, Eugene, Oreg.) according to themanufacturer’s instructions, and quantified by ELISA. Cell suspensions weredistributed into four tubes, blocked with 10% goat, rat, and/or hamster serum,and then stained with the following combinations: (i) 1 �g of CD3 per ml, 2.5 �gof CD4 per ml, 10 �g of CD8a per ml, and 10 �g of CD19 per ml; (ii) 1 �g ofMac-3 per ml, 1 �g of CD11c per ml, 2 �g of MAb RB6 per ml, and 10 �g ofhamster IgG per ml; and (iii) 10 �g of rat IgG1per ml, 10 �g of rat IgG2a per ml,and 10 �g of rat IgG2b per ml. The cell suspensions were incubated for 20 minon ice, and then 1 ml of PBS with 1% BSA was added and the cells werecentrifuged for 5 min, suspended in a solution containing 1 �g of streptavidinconjugated to peridinin chlorophyll (Pharmingen) per ml, and incubated on icefor 20 min. The cells were washed again in 1 ml of buffer consisting of PBS with1% BSA and suspended in 1 ml of PBS with 1% BSA. A fluorescence-activatedcell sorting analysis was performed by using a FACSCalibur (Becton Dickenson,Franklin Lakes, N.J.). The analysis was performed by using the CellQuest soft-ware (Becton Dickinson).
A. fumigatus. Strain ATCC 90906 was obtained from the American TypeCulture Collection (Manassas, Va.). This strain was isolated from the blood of apatient with invasive aspergillosis (15). Stock solutions of conidia were main-tained at �80°C. Sabouraud dextrose agar slants were inoculated with a loopfulof frozen stock and grown at room temperature for 7 days. PBS with 0.05%Tween 20 was added to the slants, which were then gently scraped, and 4-mlportions of the resulting conidial suspensions were transferred to eight-armmouse inhalation flasks (Ace Glass, Vineland, N.J.) containing 150 ml of Sab-ouraud dextrose agar (6). The flasks were incubated at room temperature for 14days. For intratracheal (i.t.) infection experiments, conidia from cultures grownon agar as described above were collected in 0.1% Tween 20 and then passedtwice through 12-�m-pore-size Isopore membrane polycarbonate filters (Milli-pore, Billerica, Mass.) and washed in Tween 20 prior to quantification by hema-
cytometer counting. Microscopic examination of organisms collected in thismanner revealed that conidiophores and hyphal fragments were removed.Conidial viability was confirmed by plating appropriate dilutions on Sabourauddextrose agar. Colony counts were determined after growth at room temperaturefor 4 days. To determine the effect of preparation of conidia in solutions con-taining Tween 20 on the surface charge, 20-day-old cultures were collected in0.01 M NaCl with or without 0.2% Tween 20. The zeta potential of 10 randomlyselected cells passing through the detection meter was measured for each sampleby using a Zeta-Meter 3.0� (Zeta-Meter, Inc., Staunton, Va.), which calculatedthe zeta potential, as described previously (43).
Infection models. For i.t. infection, mice were anesthetized by i.p. injection ofketamine and xylazine, and then conidia (volume, 50 �l) were administered viaa midline neck incision by using a bent 26-gauge needle attached to a tuberculinsyringe, as described previously (19). The incisions were closed with Nexabandadhesive (Closure Medical Corporation, Raleigh, N.C.). For aerosol infection,mice were placed in the side arms of an inhalation flask, and then a cloud ofconidia was created by pumping air into the flask by using a 60-ml syringeattached to Tygon tubing. After 1 min, the mice were removed from the flask.The inoculum was varied by altering the force with which air was pumpedthrough the syringe. Before the flask was used for infection, a trial run wasperformed to evaluate the size of the conidium cloud produced by a given force.To determine the reproducibility of the infecting inoculum for mice infected ina flask, initially two separate experiments were performed, in which two flaskswere used and two trials were done per flask. All eight mice in each trial werekilled 2 h after infection, their lungs were homogenized in sterile PBS, anddilutions were plated for determination of the inoculum that reached the lung.The CFU were counted after incubation of plates at room temperature for 4days. In all subsequent experiments, two mice from each flask run were killed 2 hafter infection for inoculum determination based on the CFU in the lungs. Forall groups that were compared to each other, equal numbers of mice from eachgroup were included in each run of the flask. In experiments in which definedinocula reaching the lung were desired, when this determination was outside 0.5log10 of the targeted range, the data were excluded from further analysis. In allexperiments, four additional mice received each immunosuppression treatmentin conjunction with sham infection, in which flasks with agar that was not seededwith A. fumigatus were used.
Chitin measurement. Lung chitin was measured as described previously (33).The A650 was measured after 25 min. Lungs from uninfected mice were used asblanks. Glucosamine and distilled H2O were used as standards. The limit ofdetection of this assay was 0.002 �g of glucosamine.
Histopathology. In some experiments, the right upper lobe of the lung wasused for histopathology. Formalin-fixed tissues were embedded in paraffin. Five-micrometer sections were stained with hematoxylin and eosin (H&E) or Gomorimethenamine silver (GMS) and were examined by using an Axioplan 2 orAxiophot microscope (Carl Zeiss, Inc., Thornwood, N.Y.). At least six sectionswere examined for each stain for each mouse.
Morphometry. Each lung section was viewed with a Stemi SV11 microscope(Zeiss) at a magnification of �15.75 for determination of the total lung areaexamined. All areas of each section then were examined at a magnification of�25 or �90, and all areas with visible fungi were photographed. The area of alung containing A. fumigatus was quantified by using ImageJ, version 1.31t, basedon the black color of organisms on GMS-stained slides. The threshold of eachphotograph was adjusted to determine the number of pixels that were black andrepresented areas where there was fungus. Areas that were black but not asso-ciated with identifiable fungi were excluded. After determination of the relation-ship between pixel number and distance at each magnification, the area of lungcontaining hyphae was calculated for each section and divided by the total areaof the lung examined.
Statistical analysis. The statistical analysis was performed by using Sigmastat,version 3.0 (SPSS, Inc., Chicago, Ill.). For two-group comparisons, we usedStudent’s t test for normally distributed data or the Mann-Whitney rank sum testfor data that were not normally distributed. For multiple comparisons, pairwisecomparisons were made by using the Holm-Sidak test after demonstration ofstatistical significance by one-way analysis of variance (ANOVA) or Dunn’smethod for comparison after one-way ANOVA on ranks. The correlation coef-ficient for chitin and morphometry was determined by Spearman rank ordercorrelation. Significance was defined by a P value of �0.05.
RESULTS
Aerosol infection model. In initial experiments to assess re-producibility, in each of eight trials the standard deviation for
VOL. 73, 2005 PATHOGENESIS OF ASPERGILLOSIS IN NEUTROPENIC MICE 115
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 21
Jan
uary
202
2 by
260
4:3d
09:a
17c:
3100
:e0b
d:1d
7f:c
91a:
e019
.

the CFU retrieved from the lungs of infected mice was lessthan 0.25 log10, demonstrating that mice were evenly infectedin the different positions around the flasks. In these eight trials,the mean inocula ranged from log10 6.2 to 6.8, which is a rangewithin which pathogenesis is unlikely to be altered. Similarresults were obtained with delivery of smaller inocula, rangingfrom log10 4.7 to 5.7 per mouse (Fig. 1). In only 2 of 14experiments did the mean inoculum deviate from the targetinoculum by more than log10 0.5, and in none of the experi-ments was the deviation greater than log10 0.6. Inocula thatwere log10 4 or less could not be delivered reliably. In sum-
mary, our data show that reproducible results can be obtainedwith the aerosol model and that two of the eight mice in eachtrial can be used to establish the inoculum that reaches thelung.
MAb RB6-induced neutropenia. As noted previously, MAbRB6 may cross-react with other members of the Ly-6 family(20). Therefore, we sought to determine the smallest amountthat induced absolute neutropenia and retained selectivity.The results of four representative experiments are shown inFig. 2. We found that in BALB/c mice, administration of 1 �gof MAb RB6 significantly reduced the neutrophil counts 1 daylater. Absolute neutropenia, defined as an ANC of less than5 � 105 cells/ml (500 neutrophils/�l), was induced by doses of�7.5 �g. At doses less than 50 �g, absolute neutropenia per-sisted for 1 day. Administration of 10 �g of MAb RB6 or morealso resulted in significant ALC reductions, although variabilitywas seen when mice were treated with doses ranging from 10 to25 �g (Fig. 2). Mouse weight, the route of administration (i.v.or i.p.), and the mouse strain (C57BL/6, A/JCr, or DBA/2) didnot influence the reproducibility, degree, or duration of neu-tropenia when 25 �g of MAb RB6 was given (data not shown).To assess other cell populations depleted by this MAb, wedetermined its effect on splenocytes (Table 1). As expected,the number of MAb RB6� splenocytes was significantly re-duced. Significant differences in other splenocyte populationswere not detected despite a reduction in the peripheral ALC.
To determine whether sustained neutropenia could be in-duced by repeated injection of MAb RB6, mice were given 25�g of MAb or PBS i.v. every other day for four doses, andone-half of the mice were bled on each subsequent day. The
FIG. 1. Reproducibility of aerosol infection, demonstrating the re-lationship between the target inoculum and the inoculum achieved.Each circle represents the mean log10 inoculum attained in experi-ments that included 2 to 10 trials. The horizontal lines indicate targetinocula.
FIG. 2. Summary of representative experiments to determine effect of dose of MAb RB6 on leukocyte populations. The bars indicate means,and the error bars indicate standard deviations. All experiments were performed with BALB/c mice, and treatments were administered i.v. Tenmice per group were used for the experiments whose results are shown in panel A, and five mice per group were used for the experiments whoseresults are shown in panels B, C, and D. In the experiment whose results are shown in panel D, two additional groups were treated with 25 or 200�g of polyclonal rat IgG. In these groups, neither the ANCs nor ALCs differed from those obtained for groups that received PBS. Day 4 valueswere not obtained for mice that received 25 �g of MAb RB6. a, P � 0.05, as determined by the Holm-Sidak test, for comparison to the ANC onday 0 after one-way ANOVA; b, concomitant significant reduction in the ALC compared to the ALC on day 0 (P � 0.05, as determined by theHolm-Sidak test). c, P � 0.05, as determined by Dunn’s method for comparison to the ANC on day 0 after one-way ANOVA on ranks; d,concomitant significant reduction in the ALC compared to the ALC on day 0 (P � 0.05, as determined by Dunn’s method).
116 STEPHENS-ROMERO ET AL. INFECT. IMMUN.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 21
Jan
uary
202
2 by
260
4:3d
09:a
17c:
3100
:e0b
d:1d
7f:c
91a:
e019
.

ANC was 4.5 � 105 � 4.2 � 105 cells/ml in MAb RB6-treatedmice by day 2 and returned to the baseline value by day 5.Thus, administration of repeated doses did not induce sus-tained neutropenia. To determine whether our inability toinduce sustained neutropenia was due to the development ofmouse anti-rat IgG2b, serum from these mice was used for anELISA. No antibody binding to MAb RB6 was detected insamples from mice that received repeated injections of PBS orin mice that received MAb RB6 on days 0 through 3. However,IgM binding to MAb RB6 became detectable on day 4 and wasdetected in all mice from day 5 to day 10, by which time thetiters were declining (data not shown). Low levels of murineIgG binding to MAb RB6 were detected in serum on day 10 inall three mice tested but not in serum from earlier times orfrom control mice. To determine whether a single injection ofMAb RB6 induced an anti-rat IgM response, a subsequentexperiment was performed in which four mice each received 25�g of MAb RB6 i.v. and then were bled 7 and 14 days later.Seven days after administration, naı̈ve mice had a geometricmean IgM titer of 1:60 (range, 1:50 to 1:100), while mice thatreceived MAb RB6 had a geometric mean IgM titer of 1:400(range, 1:200 to 1:800) (P � 0.03, as determined by the Mann-Whitney rank sum test). By 14 days, the IgM titer in these micewas not significantly different from that in naı̈ve mice.
Pathogenesis of invasive aspergillosis following administra-tion of MAb RB6. In BALB/c mice that received 25 �g of MAbRB6, the minimum inoculum that reproducibly resulted in100% mortality was �106 conidia reaching the lungs. Theinoculum required for mortality was the same in mice thatreceived 200 �g of MAb RB6; however, deaths did not occuruntil 5 days after infection, compared to 3 days when the lowerdose was administered. Mice that received PBS did not diefollowing comparable exposures, which were the maximumthat reproducibly could be delivered with a single pump whenA. fumigatus strain ATCC 90906 was used (data not shown).Determination of the lung chitin concentrations revealed thatthe amounts of chitin present in lungs of mice that received 5� 105 conidia on days 1, 2, and 3 were not significantly different(Fig. 4A). Mice that received 1.7 � 105 conidia had low levelsof chitin on day 1, and there was a trend toward an increase onday 2 (P � 0.1, as determined by Student’s t test) (Fig. 4B). Onday 3, despite 100% mortality (by day 4) for the mice moni-
tored for survival, in three of four mice only a low fungalburden was detected by either method, while the fourth mousehad a high fungal burden as determined by both methods,suggesting that there is heterogeneity in the ability of mice tocontrol fungal growth. Examination of H&E-stained lung sec-tions from these mice indicated that by day 3 inflammatory fociwere frequently centered on large airways and bronchial lu-mens contained densely packed inflammatory infiltrates com-posed predominantly of neutrophils (Fig. 5). In some mice,thrombosis of nearby large blood vessels was observed. In mostmice, intense mixed inflammatory cellular infiltrates, in whichneutrophils were prominent, obliterated large portions of thedistal airspace. GMS-stained lung sections demonstrated thatfoci of infection were predominantly bronchocentric. Collec-tions of hyphae were present inside bronchi and invaded thesurrounding parenchyma through the bronchial wall. However,in regions where there was intense inflammation in the termi-nal airways, hyphae were rarely observed. Faint GMS-positivematerial was present diffusely in these regions, as were dys-morphic hyphae. Occasionally, infectious foci bordered thepleura and were associated with pleural thickening with in-flammatory cells.
In C57BL/6 mice that received MAb RB6 1 day prior to
FIG. 3. Survival of mice immunosuppressed with 25 �g of MAbRB6 and infected 1 day later in aerosol flasks. (A) BALB/c mice;(B) C57BL/6 mice. Each group contained six mice. The target inoculawere 105, 5 � 105, and 106 conidia. The actual inocula, determined byplating lung homogenates of two mice 2 h after infection, are indicated.The P value was 0.005 for a comparison of BALB/c mice infected with6 � 105 and 1.4 � 106 conidia in panel A, as determined by log rankanalysis. The data in panels A and B are representative of mortalityrates obtained in five and four additional experiments, respectively.
TABLE 1. Mean leukocyte populations in the spleens ofimmunosuppressed micea
Population
Treatment group
Naı̈ve MAb RB6 CPA MAb RB6� CPA
Total splenocytes (107) 4.0 � 1.4 3.9 � 0.5 0.9 � 0.2b 1.1 � 0.1CD3� (106) 16.2 � 4.6 17.4 � 1.4 4.7 � 0.9b 6.5 � 0.6CD4� CD8� (106) 9.2 � 2.7 10.2 � 1.1 2.7 � 0.5b 3.8 � 0.5CD8� CD4� (106) 6.9 � 2.0 7.2 � 0.5 2.0 � 0.4b 2.6 � 0.2CD4� CD8� (105) 2.7 � 2.1 3.3 � 0.9 0.4 � 0.1b 0.9 � 0.4CD19� (106) 19.6 � 8.2 19.3 � 2.9 3.5 � 0.7b 4.0 � 0.5MAC-3� (106) 1.9 � 0.9 2.4 � 0.5 0.7 � 0.1 0.8 � 0.1Ly6-G� (105) 5.0 � 1.5 0.2 � 0.1b 1.5 � 0.4 0.1 � 0.0b
a Mice were treated 1 day prior to determination with 25 �g of MAb RB6 i.v.,with 150 mg of cyclophosphamide (CPA) per kg i.p., or with both agents. Thevalues are means � standard deviations per spleen. Each group contained fourmice.
b P � 0.05, as determined by Dunn’s method for multiple comparisons afterKruskal-Wallis one-way analysis of variance on ranks.
VOL. 73, 2005 PATHOGENESIS OF ASPERGILLOSIS IN NEUTROPENIC MICE 117
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 21
Jan
uary
202
2 by
260
4:3d
09:a
17c:
3100
:e0b
d:1d
7f:c
91a:
e019
.

infection, an inoculum consisting of 5 � 105 conidia resulted in100% mortality (Fig. 3B). The lung fungal burden rose some-what more consistently in this mouse strain during the courseof infection than in BALB/c mice (Fig. 4A and C). The histo-logical appearance of sections from these mice was similar tothe appearance in BALB/c mice treated with MAb RB6, andextensive inflammation was seen on day 3. No deaths occurredin sham-infected mice in any of the experiments describedabove for either mouse strain.
i.t. infection. We determined the minimum inoculum thatreproducibly resulted in 100% mortality following i.t. infection
and the number of conidia retrieved from the lung 2 h afterinfection. With strain ATCC 90906, 5 � 106 conidia was theminimum lethal inoculum in BALB/c mice that had received25 �g of MAb RB6 (data not shown). Plating of the lungs oftwo mice infected with 5 � 106 conidia demonstrated that themean number of retrievable conidia was 9.4 � 105 � 0.4 � 105.Thus, administration of minimal consistently lethal inocula bythe i.t. and aerosol routes resulted in retrieval of comparablenumbers of CFU from the lungs. To determine whether thelack of demonstrable fungal growth was dependent on theinfection method, lung fungal burden was determined by bothchitin measurement and morphometry. Again, heterogeneityin the response to infection with a lethal inoculum was seen(Fig. 6).
Administration of cyclophosphamide and corticosteroids. Asingle i.p. injection of 150 or 200 mg of cyclophosphamide perkg resulted in severe leukopenia, and the neutrophil nadir wasreached on day 4 after inoculation (Table 2). By 5 days afteradministration of a single dose, the total WBC counts andANCs were at or near normal. Repeated administration of 150mg of cyclophosphamide per kg produced neutropenia in mostmice that was sustained for 16 days. Neutropenia induced bycyclophosphamide showed significant interanimal variation,while ALC depression was both more consistent and sustained.Administration of 200 mg/kg resulted in less interanimal vari-ation (data not shown). Repeated administration of 150 mg/kgresulted in loss of �10% of the body weight, followed byweight stabilization, while administration of 200 mg/kg resultedin loss of 25% of the body weight (data not shown), whichprecluded use of this dose. Administration of 150 mg/kg mark-edly reduced all of the splenic lymphocyte populations tested(Table 1). A trend toward reduced numbers of Mac-3� andLy6-G� splenocytes also was seen, but the differences were notstatistically significant after correction for multiple compari-sons.
When cyclophosphamide was administered on days �4, 0, 4,and 8 after infection, an inoculum of 106 conidia that reachedthe lungs was required to reproducibly induce 100% mortality(data not shown), which was comparable to the results ob-served with MAb RB6. To identify a model in which deathoccurred with a lower inoculum and with a high lung fungalburden, we combined a single 35-mg/kg dose of triamcinolone,administered 1 day prior to infection, with repeated cyclophos-phamide doses administered on days �4, 0, and 4. While 100%mortality occurred in mice that received �104 conidia by day 4or 5 after infection (data not shown), we consistently observedsome deaths in mice in this model by day 7 after sham infec-tion, despite addition of trimethoprim-sulfamethoxazole to thedrinking water to prevent bacterial infection. Coadministrationof dexamethasone at i.p. doses ranging from 2 to 10 mg/kg withcyclophosphamide did not render mice more susceptible todeath from infection with lower numbers of conidia comparedto administration of cyclophosphamide alone (data not shown).
Coadministration of MAb RB6 and cyclophosphamide. ForBALB/c mice, coadministration of 25 �g of MAb RB6 and 150mg of cyclophosphamide per kg on days 0, 3, 6, and 9 resultedin significantly more severe neutropenia on day 3 than didadministration of MAb RB6 or cyclophosphamide alone (theANCs were 12.7 � 105 � 5 � 105, 9.8 � 105 � 2.8 � 105, and0.5 � 105 � 0.3 � 105 for groups treated with MAb RB6,
FIG. 4. Lung fungal burden in MAb RB6-treated BALB/c orC57BL/6 mice infected with an aerosol. (A) Sixteen mice of each strainreceived 25 �g of MAb RB6 1 day prior to infection with A. fumigatus.Four mice of each strain were killed 1, 2, or 3 days after they received5.3 � 105 conidia. Each symbol represents a single mouse. (B) Exper-iment with BALB/c mice, performed as described above for panel A,except that on day 3 the fungal burden was also determined by mor-phometry. The mean inoculum was 1.7 � 105 conidia. (C) Ten MAbRB6-treated C57BL/6 mice received 2.6 � 105 conidia, and the fungalburden was determined by chitin measurement (F) or by morphometry(E) in six of these mice on day 3. The mortality rate was 100% foradditional mice infected with A. fumigatus that were monitored forsurvival (four, eight, and four mice in the experiments whose resultsare shown in panels A, B, and C, respectively). No sham-infected micedied in these experiments (four mice per experimental group).
118 STEPHENS-ROMERO ET AL. INFECT. IMMUN.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 21
Jan
uary
202
2 by
260
4:3d
09:a
17c:
3100
:e0b
d:1d
7f:c
91a:
e019
.
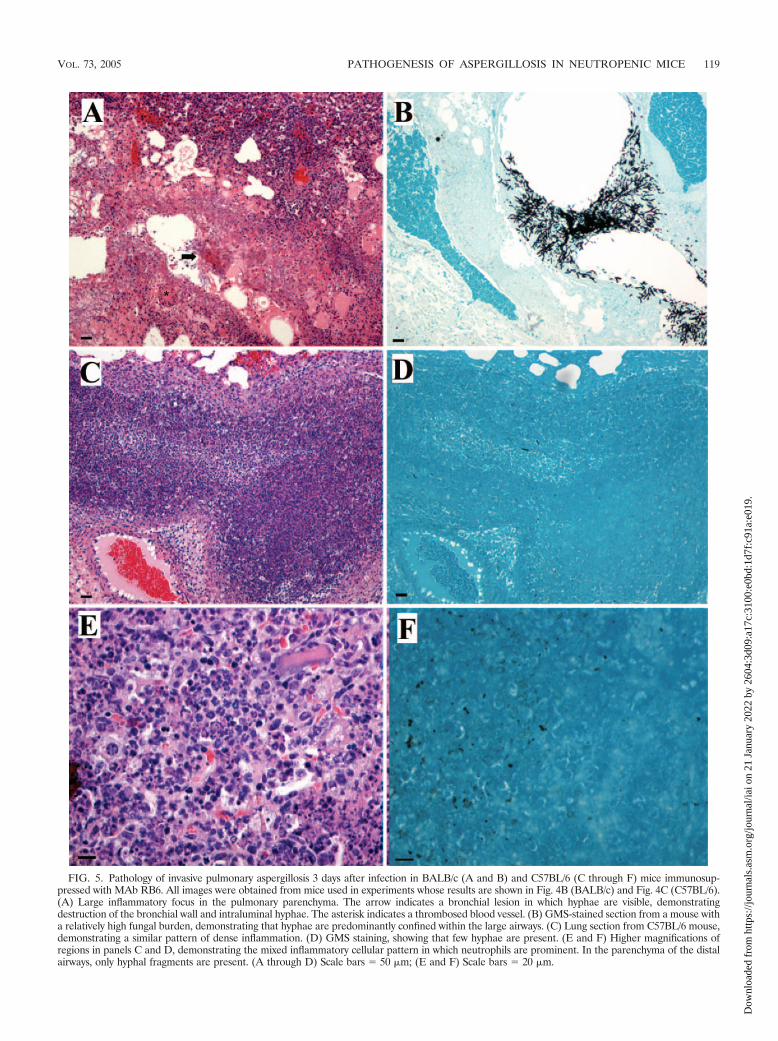
FIG. 5. Pathology of invasive pulmonary aspergillosis 3 days after infection in BALB/c (A and B) and C57BL/6 (C through F) mice immunosup-pressed with MAb RB6. All images were obtained from mice used in experiments whose results are shown in Fig. 4B (BALB/c) and Fig. 4C (C57BL/6).(A) Large inflammatory focus in the pulmonary parenchyma. The arrow indicates a bronchial lesion in which hyphae are visible, demonstratingdestruction of the bronchial wall and intraluminal hyphae. The asterisk indicates a thrombosed blood vessel. (B) GMS-stained section from a mouse witha relatively high fungal burden, demonstrating that hyphae are predominantly confined within the large airways. (C) Lung section from C57BL/6 mouse,demonstrating a similar pattern of dense inflammation. (D) GMS staining, showing that few hyphae are present. (E and F) Higher magnifications ofregions in panels C and D, demonstrating the mixed inflammatory cellular pattern in which neutrophils are prominent. In the parenchyma of the distalairways, only hyphal fragments are present. (A through D) Scale bars � 50 �m; (E and F) Scale bars � 20 �m.
VOL. 73, 2005 PATHOGENESIS OF ASPERGILLOSIS IN NEUTROPENIC MICE 119
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 21
Jan
uary
202
2 by
260
4:3d
09:a
17c:
3100
:e0b
d:1d
7f:c
91a:
e019
.

cyclophosphamide, and both agents, respectively [means �standard deviations; four mice per group; P � 0.05 for bothcomparisons, as determined by Dunn’s method for multiplecomparisons after Kruskal-Wallis one-way ANOVA onranks]). By day 6, the ANCs were comparable for mice treatedwith cyclophosphamide alone and mice that received bothagents, and by day 9, the ANCs were comparable to the ANCson day 0 for all treatment groups (data not shown). Coadmin-istration of 9 mg of methotrexate per kg did not alter the ANCin the peripheral blood of mice treated with MAb RB6 on day
3 (data not shown). In mice treated with both cyclophospha-mide and MAb RB6, the trend toward a reduction in splenicleukocyte populations 1 day later paralleled the trend observedin mice treated with cyclophosphamide alone, but the differ-ences were not statistically significant (Table 1). The numberof Ly6-G� cells was significantly reduced compared to thenumber in naı̈ve mice. These results suggest that the effects ofMAb RB6 and cyclophosphamide are additive. The anti-MAbRB6 titers in these mice were not different from those in naı̈vemice either 7 or 14 days after administration of a single dose ofMAb RB6 with cyclophosphamide (data not shown).
To determine whether administration of cyclophosphamideallowed a response to repeat doses of MAb RB6, four C57BL/6mice were given MAb RB6, cyclophosphamide, or both on day0 and then repeated injections of MAb RB6 on days 3 and 6after bleeding. Although, as in BALB/c mice, more severeneutropenia was seen on day 3 in mice treated with bothagents, by day 6 there was no difference between the groups,and the ANCs were not different from the values obtained onday 0 (data not shown). Significant reductions in ALCs wereseen in mice treated with cyclophosphamide alone or in micetreated with both agents on days 3, 6, and 9 compared to thevalues obtained on day 0.
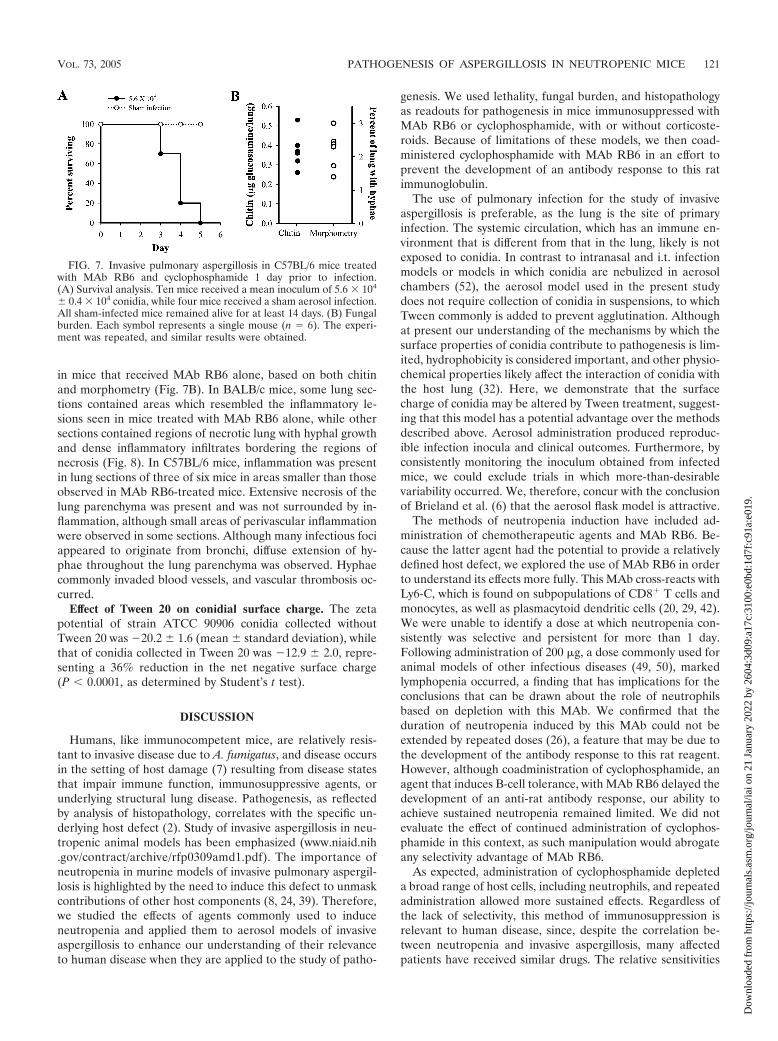
Pathogenesis of invasive aspergillosis following coadminis-tration of MAb RB6 and cyclophosphamide. An inoculum of105 conidia that reached the lung resulted in 100% mortality inBALB/c mice (data not shown), while C57BL/6 mice died afterthey received 5 � 104 conidia (Fig. 7A). In BALB/c mice thatreceived an inoculum of 1.9 � 105 conidia, heterogeneity in thefungal burden was seen (data not shown). In C57BL/6 micethat received 9 � 104 conidia, the fungal burden 3 days afterinfection was more consistently higher than the fungal burden
FIG. 6. Lung fungal burden in BALB/c mice immunosuppressedwith 25 �g of MAb RB6 and infected i.t., expressed as the amount ofchitin per lung (F) or by morphometry (E). Twenty mice were in-fected, and the lungs of two mice were plated for determination of theinoculum that reached the lung 2 h after infection. Four mice weremonitored for survival, and the remainder were killed on days 1, 2, and3 (five, five, and four mice, respectively) for determination of thefungal burden. The mortality was 100% in the group monitored forsurvival by day 4.
TABLE 2. Effect of cyclophosphamide on leukocyte populationsa
Expt DayWBC (106/ml) ANC (105/ml) ALC (106/ml)
Distilled H2O CPA Distilled H2O CPA Distilled H2O CPA
I 0 4.5 � 1.4 4.6 � 1.1 5.6 � 2.9 6.2 � 2.9 3.8 � 1.0 3.7 � 1.02 5.3 � 1.1 2.3 � 0.8b 4.7 � 3.0 6.9 � 5.3 4.8 � 1.1 1.5 � 0.6b
3 4.5 � 2.2 2.0 � 0.5 2.7 � 1.4 3.1 � 2.4 4.1 � 2.0 1.7 � 0.64 10.7 � 2.5 1.1 � 0.7b 12.5 � 4.4 0.4 � 0.5c 9.3 � 2.3 1.0 � 0.7b
II 0 6.3 � 2.6 3.5 � 1.5 9.4 � 7.2 5.0 � 2.9 5.1 � 1.8 2.9 � 1.23 6.7 � 3.0 2.0 � 1.1 9.3 � 5.6 2.2 � 1.8 5.8 � 2.6 1.7 � 0.9b
4 5.1 � 1.6 1.3 � 0.6b 5.4 � 2.3 0.1 � 0.2c 4.4 � 1.4 105 � 0.8b
5 9.9 � 3.3 2.6 � 0.9c 12.1 � 8.2 3.1 � 1.0c 8.5 � 2.5 2.1 � 0.8c
6 8.5 � 1.7 5.0 � 0.9c 11.9 � 4.9 10.8 � 4.3 7.2 � 2.0 3.8 � 0.8c
III 0 NDf 4.8 � 1.1 ND 6.3 � 2.0 (6.5) ND 4.0 � 0.94 2.9 � 1.0d 0.6 � 0.5 (0.7)e 2.8 � 1.0d
5 0.6 � 0.3d 2.9 � 5.0 (0.4)e 1.2 � 0.3d
8 1.7 � 0.8d 2.5 � 1.6 (3.2) 1.4 � 0.6d
12 2.0 � 1.1d 2.0 � 4.6 (0)e 1.8 � 0.8d
16 2.4 � 0.5d 1.2 � 1.7 (0)e 2.2 � 0.5d
20 3.3 � 0.8d 6.6 � 4.2 (6.6) 2.5 � 0.7d
a The data are representative of the results obtained in three experiments. In experiments I and II, mice received 150 mg of cyclophosphamide (CPA) per kg on day0, and each group contained four mice. The control mice received a comparable volume of distilled H2O. In experiment III, the cyclophosphamide-treated mice receivedcyclophosphamide on days 0, 4, 8, 12, and 16, and each group contained five mice. The numbers in parentheses are the median values obtained for five mice.
b P � 0.05 for a comparison to the group that received distilled H2O on the same day as determined by Student’s t test.c P � 0.05 for a comparison to the group that received distilled H2O on the same day, as determined by the Mann-Whitney rank sum test.d P � 0.05 for a comparison to the day 0 value, as determined by the Holm-Sidak method for multiple comparisons after one-way ANOVA.e P � 0.05 for a comparison to the day 0 value, as determined by Dunn’s method for multiple comparisons after Kruskall-Wallis one-way analysis of variance on ranks.f ND, not determined.
120 STEPHENS-ROMERO ET AL. INFECT. IMMUN.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 21
Jan
uary
202
2 by
260
4:3d
09:a
17c:
3100
:e0b
d:1d
7f:c
91a:
e019
.
in mice that received MAb RB6 alone, based on both chitinand morphometry (Fig. 7B). In BALB/c mice, some lung sec-tions contained areas which resembled the inflammatory le-sions seen in mice treated with MAb RB6 alone, while othersections contained regions of necrotic lung with hyphal growthand dense inflammatory infiltrates bordering the regions ofnecrosis (Fig. 8). In C57BL/6 mice, inflammation was presentin lung sections of three of six mice in areas smaller than thoseobserved in MAb RB6-treated mice. Extensive necrosis of thelung parenchyma was present and was not surrounded by in-flammation, although small areas of perivascular inflammationwere observed in some sections. Although many infectious fociappeared to originate from bronchi, diffuse extension of hy-phae throughout the lung parenchyma was observed. Hyphaecommonly invaded blood vessels, and vascular thrombosis oc-curred.
Effect of Tween 20 on conidial surface charge. The zetapotential of strain ATCC 90906 conidia collected withoutTween 20 was �20.2 � 1.6 (mean � standard deviation), whilethat of conidia collected in Tween 20 was �12.9 � 2.0, repre-senting a 36% reduction in the net negative surface charge(P � 0.0001, as determined by Student’s t test).
DISCUSSION
Humans, like immunocompetent mice, are relatively resis-tant to invasive disease due to A. fumigatus, and disease occursin the setting of host damage (7) resulting from disease statesthat impair immune function, immunosuppressive agents, orunderlying structural lung disease. Pathogenesis, as reflectedby analysis of histopathology, correlates with the specific un-derlying host defect (2). Study of invasive aspergillosis in neu-tropenic animal models has been emphasized (www.niaid.nih.gov/contract/archive/rfp0309amd1.pdf). The importance ofneutropenia in murine models of invasive pulmonary aspergil-losis is highlighted by the need to induce this defect to unmaskcontributions of other host components (8, 24, 39). Therefore,we studied the effects of agents commonly used to induceneutropenia and applied them to aerosol models of invasiveaspergillosis to enhance our understanding of their relevanceto human disease when they are applied to the study of patho-
genesis. We used lethality, fungal burden, and histopathologyas readouts for pathogenesis in mice immunosuppressed withMAb RB6 or cyclophosphamide, with or without corticoste-roids. Because of limitations of these models, we then coad-ministered cyclophosphamide with MAb RB6 in an effort toprevent the development of an antibody response to this ratimmunoglobulin.
The use of pulmonary infection for the study of invasiveaspergillosis is preferable, as the lung is the site of primaryinfection. The systemic circulation, which has an immune en-vironment that is different from that in the lung, likely is notexposed to conidia. In contrast to intranasal and i.t. infectionmodels or models in which conidia are nebulized in aerosolchambers (52), the aerosol model used in the present studydoes not require collection of conidia in suspensions, to whichTween commonly is added to prevent agglutination. Althoughat present our understanding of the mechanisms by which thesurface properties of conidia contribute to pathogenesis is lim-ited, hydrophobicity is considered important, and other physio-chemical properties likely affect the interaction of conidia withthe host lung (32). Here, we demonstrate that the surfacecharge of conidia may be altered by Tween treatment, suggest-ing that this model has a potential advantage over the methodsdescribed above. Aerosol administration produced reproduc-ible infection inocula and clinical outcomes. Furthermore, byconsistently monitoring the inoculum obtained from infectedmice, we could exclude trials in which more-than-desirablevariability occurred. We, therefore, concur with the conclusionof Brieland et al. (6) that the aerosol flask model is attractive.
The methods of neutropenia induction have included ad-ministration of chemotherapeutic agents and MAb RB6. Be-cause the latter agent had the potential to provide a relativelydefined host defect, we explored the use of MAb RB6 in orderto understand its effects more fully. This MAb cross-reacts withLy6-C, which is found on subpopulations of CD8� T cells andmonocytes, as well as plasmacytoid dendritic cells (20, 29, 42).We were unable to identify a dose at which neutropenia con-sistently was selective and persistent for more than 1 day.Following administration of 200 �g, a dose commonly used foranimal models of other infectious diseases (49, 50), markedlymphopenia occurred, a finding that has implications for theconclusions that can be drawn about the role of neutrophilsbased on depletion with this MAb. We confirmed that theduration of neutropenia induced by this MAb could not beextended by repeated doses (26), a feature that may be due tothe development of the antibody response to this rat reagent.However, although coadministration of cyclophosphamide, anagent that induces B-cell tolerance, with MAb RB6 delayed thedevelopment of an anti-rat antibody response, our ability toachieve sustained neutropenia remained limited. We did notevaluate the effect of continued administration of cyclophos-phamide in this context, as such manipulation would abrogateany selectivity advantage of MAb RB6.
As expected, administration of cyclophosphamide depleteda broad range of host cells, including neutrophils, and repeatedadministration allowed more sustained effects. Regardless ofthe lack of selectivity, this method of immunosuppression isrelevant to human disease, since, despite the correlation be-tween neutropenia and invasive aspergillosis, many affectedpatients have received similar drugs. The relative sensitivities
FIG. 7. Invasive pulmonary aspergillosis in C57BL/6 mice treatedwith MAb RB6 and cyclophosphamide 1 day prior to infection.(A) Survival analysis. Ten mice received a mean inoculum of 5.6 � 104
� 0.4 � 104 conidia, while four mice received a sham aerosol infection.All sham-infected mice remained alive for at least 14 days. (B) Fungalburden. Each symbol represents a single mouse (n � 6). The experi-ment was repeated, and similar results were obtained.
VOL. 73, 2005 PATHOGENESIS OF ASPERGILLOSIS IN NEUTROPENIC MICE 121
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 21
Jan
uary
202
2 by
260
4:3d
09:a
17c:
3100
:e0b
d:1d
7f:c
91a:
e019
.

FIG. 8. Lung pathology 3 days after infection of mice immunosuppressed with MAb RB6 and cyclophosphamide 1 day prior to infection. (A) H&E-stained section from a BALB/c mouse that received an inoculum of 1.8 � 105 conidia, showing bronchial lesions with necrotic lung surrounded byinflammatory infiltrate. (B) GMS staining of the region shown in panel A, showing invasion through the bronchus into the surrounding lung parenchyma.(C) In C57BL/6 mice, extensive hyphal spread through the lung parenchyma resulted in severe necrosis with inflammation perivascularly but not in thedistal airspace and large areas of necrotic lungs. (D) GMS-stained section of the region shown in panel C. (E) H&E staining of an infectious focus inthe lung of a C57BL/6 mouse, showing vascular thrombosis near a large bronchus. (F) GMS staining of the region shown in panel E, showing extensivehyphal penetration of the vessel. C57BL/6 sections were obtained from the mice described in the legend to Fig. 6. The arrows in panels E and F indicatea smaller, thrombosed blood vessel in which hyphae were visible. (A through D) Scale bars � 0.2 �m; (E and F) scale bars � 0.1 �m.
122 STEPHENS-ROMERO ET AL. INFECT. IMMUN.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 21
Jan
uary
202
2 by
260
4:3d
09:a
17c:
3100
:e0b
d:1d
7f:c
91a:
e019
.

of lymphocyte populations to cyclophosphamide-induced de-pletion vary, so that B cells are more susceptible than T cellsand CD4� T cells are more susceptible than CD8� T cells.However, cyclophosphamide also can deplete regulatory T-cellpopulations (reviewed in reference 21) and, therefore, mayhave proinflammatory actions. The known effects of cyclophos-phamide on local pulmonary immunity include inhibition ofcomplement synthesis and reduction of chemotactic activity byguinea pig bronchoalveolar macrophages (45, 46). The contri-bution of each of these extended effects to the pathogenesis ofinvasive pulmonary aspergillosis in murine models is unknown.
The fungal inoculum is an important determinant of lethalityin murine models (16). The inoculum required for productionof invasive disease in immunocompromised hosts is unknown,but it is probably smaller than the inocula commonly admin-istered intranasally or i.t. in murine infections, which typicallyrange from 5 � 106 to 2 � 107 conidia. We found that inBALB/c mice immunosuppressed with MAb RB6, the lethalinoculum in this aerosol model was comparable to that in thei.t. model and the same as that required following cyclophos-phamide administration. The inoculum required for 100%mortality in C57BL/6 mice was fivefold less in mice that re-ceived either MAb RB6 or both MAb RB6 and cyclophos-phamide, suggesting that there are subtle differences betweenthe mouse strains. Administration of a higher dose (200 �g) ofMAb RB6 did not reduce the inoculum required for 100%mortality, although deaths did not occur until 5 days afterinfection (data not shown), which is consistent with the longertime to neutrophil recovery when this dose is given. Coadmin-istration of MAb RB6 and cyclophosphamide resulted in a�10-fold decrease in the lethal inoculum in both mousestrains. Thus, this approach offers a less problematic alter-native to the combined cyclophosphamide-corticosteroid reg-imens that have been used to develop lower-inoculum models(55). Although cyclophosphamide appeared to delay or pre-vent the development of the anti-rat antibody response, asnoted above, we did not observe continued efficacy of repeatedMAb RB6 doses. Rather, enhanced susceptibility may be re-lated to the ability to sustain maximal neutropenia for the first3 days after infection, as the nadir counts with either individualagent lasted only 1 to 2 days, despite a somewhat longer sta-tistically significant duration of reduction. Again, given thebroad range of immune cells affected by this agent, the possi-bility of contributions by depletion of other cells cannot beexcluded.
The course of disease in both BALB/c and C57BL/6 micemade neutropenic with all of the immunosuppression regimensexamined was acute, as all deaths occurred within 3 to 5 daysafter infection. When inocula smaller than those resulting in100% mortality were administered, only occasional mice thatsurvived longer than 5 days after infection subsequently died.Plating of lung homogenates from such mice obtained 7 daysafter infection only occasionally yielded growth of small num-bers of A. fumigatus colonies (data not shown), suggesting thatthe surviving mice cleared the infecting inoculum.
Following induction of neutropenia with MAb RB6, deathcoincided with a return of the neutrophil counts to the baselinevalue, a variable pulmonary fungal burden, and a massive in-flammatory response in the lung parenchyma in the absence ofdisseminated disease (data not shown). These features suggest
that mice died as a result of the inflammatory response. Fatalpulmonary complications are associated with rapid neutrophilrecovery, and in human immunodeficiency virus-negative pa-tients, more than one-third of reported cases of immunoresti-tution disease occur in patients with invasive aspergillosis (10,57). Thus, the time of restoration of immunity is a time ofvulnerability in human disease. In some mice, infectious fociwith intact hyphae were observed in large bronchi. Therefore,it is possible that these lesions were responsible for the deathof some mice. Disease confined to large airways, a pattern thatis observed less commonly than bronchopneumonia, may re-sult in acute airway obstruction in humans and has been re-ported after resolution of neutropenia (4, 22). However, thepresence of more distal lesions in the mice at earlier times afterinfection (data not shown) and the consistent finding of exten-sive parenchymal inflammation in which only hyphal remnantswere observed suggest that these inflammatory lesions weresufficient to cause death. Furthermore, although significantbronchial infectious foci could have been missed by micro-scopic examination of limited numbers of sections, the strongcorrelation of whole lung chitin content with morphometryresults supports the finding that death occurred in mice withundetectable fungal burdens. The correlation of the valuesobtained by these methods for the samples used in this studywas highly significant (P � 0.0001; Spearman rank order cor-relation coefficient � 0.52; n � 68). Use of both methods forevaluation of the fungal burden provides additional confidencein the results obtained with a system in which CFU determi-nations are of questionable relevance.
In mice in which neutropenia was induced with both MAbRB6 and cyclophosphamide, infectious foci extended outwardfrom bronchi and bronchioles. Bronchopneumonia is the mostcommon pattern observed in human invasive pulmonary as-pergillosis (58). Distal lung disease results either from aspira-tion of conidia into the respiratory bronchioles and alveolarducts with direct spread or from colonization of the bronchialmucosa by invasion through the wall (25). Thus, the pathologyobserved in this model is consistent with that of human diseasein neutropenic hosts. The pathogenesis of the target lesion, ahallmark of invasive aspergillosis, reflects centrifugal expan-sion of a small focus of bronchitis or bronchopneumonia, re-sulting in a spherical nodule of necrotic lung in which vascularinvasion is present, while invasion of larger vessels may resultin classic wedge-shaped infarcts (23). Angioinvasion, a featurethat can occur in any pattern of invasive aspergillosis but thatis particularly associated with disease in neutropenic patients(23), also was observed. Necrosis may result from vascularinvasion or toxic fungal products (25, 28). In BALB/c mice,inflammatory infiltrates bordered necrotic lungs, and the fun-gal burden in lethally infected mice was inconsistent. However,in C57BL/6 mice, death appeared to occur independent of avigorous inflammatory response and coincided with a higherfungal burden. The difference in the responses of these mousestrains could represent subtle variations in the degree or per-sistence of immunosuppression or species-related disparities inthe inflammatory response to A. fumigatus products, such asthose that translate into marked differences in cell-mediatedimmune responses to other pathogens (27, 56).
In summary, as in human disease, the method of neutrope-nia induction is an important determinant of pathogenesis in
VOL. 73, 2005 PATHOGENESIS OF ASPERGILLOSIS IN NEUTROPENIC MICE 123
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 21
Jan
uary
202
2 by
260
4:3d
09:a
17c:
3100
:e0b
d:1d
7f:c
91a:
e019
.

murine models of invasive pulmonary aspergillosis. The hostresponse appears to be the primary cause of damage in miceimmunosuppressed with MAb RB6, and models in which thisagent is used may be the best models for studying the dynamicsof pathogenesis of invasive aspergillosis in the setting of neu-trophil recovery and the accompanying increase in host re-sponse. In C57BL/6 mice treated with a combination of MAbRB6 and cyclophosphamide, fungal growth may play a moreprominent role. Differences in pathogenesis must be borne inmind when these models are used to study the role of eitherfungal molecules or host inflammatory responses to infectionor during the evaluation of pharmacologic or immune modu-latory agents.
ACKNOWLEDGMENTS
This work was supported in part by NIH grant 1R03 AI53623 (toM.F.) and by a grant-in-aid of research from the American LungAssociation of New York (to M.F.).
We thank Anthony Cacciapuoti and David Loebenberg for assis-tance with establishing the aerosol model, Robert Coffman for provid-ing the MAb RB6-producing hybridoma cell line, Mark Jutila forproviding ammonium persulfate-precipitated MAb RB6, and Jim Cut-ler for helpful discussions. We also thank Histopathology Shared Re-source of the Albert Einstein Cancer Center (NIH CA 13330-33) forslide preparation, Carlos Taborda and Joshua Nosanchuk for perform-ing the zeta potential measurements, Liise-anne Pirofski for criticalreading of the manuscript, and Betty Diamond and Elena Peeva forhelpful discussions.
REFERENCES
1. Anaissie, E. J., and S. F. Costa. 2001. Nosocomial aspergillosis is waterborne.Clin. Infect. Dis. 33:1546–1548.
2. Barth, P. J., C. Rossberg, S. Koch, and A. Ramaswamy. 2000. Pulmonaryaspergillosis in an unselected autopsy series. Pathol. Res. Pract. 196:73–80.
3. Berenguer, J., M. C. Allende, J. W. Lee, K. Garrett, C. Lyman, N. M. Ali, J.Bacher, P. A. Pizzo, and T. J. Walsh. 1995. Pathogenesis of pulmonaryaspergillosis granulocytopenia versus cyclosporine and methylprednisolone-induced immunosuppression. Am. J. Respir. Crit. Care Med. 152:1079–1086.
4. Berlinger, N. T., and T. J. Freeman. 1989. Acute airway obstruction due tonecrotizing tracheobronchial aspergillosis in immunocompromised patients:a new clinical entity. Ann. Otol. Rhinol. Laryngol. 98:718–720.
5. Bodey, G. P. 1966. Fungal infections complicating acute leukemia. J. ChronicDis. 19:667–687.
6. Brieland, J. K., C. Jackson, F. Menzel, D. Loebenberg, A. Cacciapuoti, J.Halpern, S. Hurst, T. Muchamuel, R. Debets, R. Kastelein, T. Churakova, J.Abrams, R. Hare, and A. O’Garra. 2001. Cytokine networking in lungs ofimmunocompetent mice in response to inhaled Aspergillus fumigatus. Infect.Immun. 69:1554–1560.
7. Casadevall, A., and L. Pirofski. 2003. The damage-response framework ofmicrobial pathogenesis. Nat. Rev. Microbiol. 1:17–24.
8. Cenci, E., A. Mencacci, D. C. Fe, G. Del Sero, P. Mosci, C. Montagnoli, A.Bacci, and L. Romani. 1998. Cytokine- and T helper-dependent lung muco-sal immunity in mice with invasive pulmonary aspergillosis. J. Infect. Dis.178:1750–1760.
9. Cenci, E., A. Mencacci, A. Spreca, C. Montagnoli, A. Bacci, K. Perruccio, A.Velardi, W. Magliani, S. Conti, L. Polonelli, and L. Romani. 2002. Protectionof killer antiidiotypic antibodies against early invasive aspergillosis in amurine model of allogeneic T-cell-depleted bone marrow transplantation.Infect. Immun. 70:2375–2382.
10. Cheng, V. C., K. Y. Yuen, W. M. Chan, S. S. Wong, E. S. Ma, and R. M. Chan.2000. Immunorestitution disease involving the innate and adaptive response.Clin. Infect. Dis. 30:882–892.
11. Chiller, T. M., J. C. Luque, R. A. Sobel, K. Farrokhshad, K. V. Clemons, andD. A. Stevens. 2002. Development of a murine model of cerebral aspergil-losis. J. Infect. Dis. 186:574–577.
12. Cohen, M. S., R. E. Isturiz, H. L. Malech, R. K. Root, C. M. Wilfert, L.Gutman, and R. H. Buckley. 1981. Fungal infection in chronic granuloma-tous disease. The importance of the phagocyte in defense against fungi.Am. J. Med. 71:59–66.
13. DeGregorio, M. W., W. M. Lee, C. A. Linker, R. A. Jacobs, and C. A. Ries.1982. Fungal infections in patients with acute leukemia. Am. J. Med. 73:543–548.
14. Denning, D. W. 1998. Invasive aspergillosis. Clin. Infect. Dis. 26:781–803.15. Dermoumi, H. 1994. In vitro susceptibility of fungal isolates of clinically
important specimens to itraconazole, fluconazole and amphotericin B. Che-motherapy 40:92–98.
16. Dixon, D. M., A. Polak, and T. J. Walsh. 1989. Fungus dose-dependentprimary pulmonary aspergillosis in immunosuppressed mice. Infect. Immun.57:1452–1456.
17. Duong, M., N. Ouellet, M. Simard, Y. Bergeron, M. Olivier, and M. G.Bergeron. 1998. Kinetic study of host defense and inflammatory response toAspergillus fumigatus in steroid-induced immunosuppressed mice. J. Infect.Dis. 178:1472–1482.
18. Eisenstein, D. J., P. W. Biddinger, and J. C. Rhodes. 1990. Experimentalmurine invasive pulmonary aspergillosis. Am. J. Clin. Pathol. 93:510–515.
19. Feldmesser, M., and A. Casadevall. 1997. Effect of serum IgG1 to Crypto-coccus neoformans glucuronoxylomannan on murine pulmonary infection.J. Immunol. 158:790–799.
20. Fleming, T. J., M. L. Fleming, and T. R. Malek. 1993. Selective expression ofLy-6G on myeloid lineage cells in mouse bone marrow. RB6-8C5 mAb togranulocyte-differentiation antigen (Gr-1) detects members of the Ly-6 fam-ily. J. Immunol. 151:2399–2408.
21. Fox, D. A., and W. J. McCune. 1989. Immunologic and clinical effects ofcytotoxic drugs used in the treatment of rheumatoid arthritis and systemiclupus erythematosus. Concepts Immunopathol. 7:20–78.
22. Franquet, T., F. Serrano, A. Gimenez, J. M. Rodriguez-Arias, and C. Puzo.2002. Necrotizing aspergillosis of large airways: CT findings in eight patients.J. Comput. Assisted Tomogr. 26:342–345.
23. Fraser, R. S. 1993. Pulmonary aspergillosis: pathologic and pathogeneticfeatures. Pathol. Annu. 28:231–277.
24. Garlanda, C., E. Hirsch, S. Bozza, A. Salustri, M. De Acetis, R. Nota, A.Maccagno, F. Riva, B. Bottazzi, G. Peri, A. Doni, L. Vago, M. Botto, R. DeSantis, P. Carminati, G. Siracusa, F. Altruda, A. Vecchi, L. Romani, and A.Mantovani. 2002. Non-redundant role of the long pentraxin PTX3 in anti-fungal innate immune response. Nature 420:182–186.
25. Gowing, N. F., and I. M. Hamlin. 1960. Tissue reactions to Aspergillus incases of Hodgkin’s disease and leukaemia. J. Clin. Pathol. 13:396–413.
26. Han, Y., and J. E. Cutler. 1997. Assessment of a mouse model of neutropeniaand the effect of an anticandidiasis monoclonal antibody in these animals.J. Infect. Dis. 175:1169–1175.
27. Heinzel, F. P., M. D. Sadick, B. J. Holaday, R. L. Coffman, and R. M.Locksley. 1989. Reciprocal expression of interferon gamma or interleukin 4during the resolution or progression of murine leishmaniasis. Evidence forexpansion of distinct helper T cell subsets. J. Exp. Med. 169:59–72.
28. Henrici, A. T. 1939. An endotoxin from Aspergillus fumigatus. J. Immunol.36:319–338.
29. Jutila, M. A., F. G. Kroese, K. L. Jutila, A. M. Stall, S. Fiering, L. A.Herzenberg, E. L. Berg, and E. C. Butcher. 1988. Ly-6C is a monocyte/macrophage and endothelial cell differentiation antigen regulated by inter-feron-gamma. Eur. J. Immunol. 18:1819–1826.
30. Kaiser, L., T. Huguenin, P. D. Lew, B. Chapuis, and D. Pittet. 1998. Invasiveaspergillosis. Clinical features of 35 proven cases at a single institution.Medicine (Baltimore). 77:188–194.
31. Kirkpatrick, W. R., R. K. McAtee, A. W. Fothergill, M. G. Rinaldi, and T. F.Patterson. 2000. Efficacy of voriconazole in a guinea pig model of dissemi-nated invasive aspergillosis. Antimicrob. Agents Chemother. 44:2865–2868.
32. Latge, J. P. 1999. Aspergillus fumigatus and aspergillosis. Clin. Microbiol.Rev. 12:310–350.
33. Lehmann, P. F., and L. O. White. 1975. Chitin assay used to demonstraterenal localization and cortisone-enhanced growth of Aspergillus fumigatusmycelium in mice. Infect. Immun. 12:987–992.
34. Malek, T. R., E. M. Shevach, and K. M. Danis. 1989. Activation of Tlymphocytes through the Ly-6 pathway is defective in A strain mice. J. Im-munol. 143:439–445.
35. Mehrad, B., R. M. Strieter, and T. J. Standiford. 1999. Role of TNF-alphain pulmonary host defense in murine invasive aspergillosis. J. Immunol.162:1633–1640.
36. Mehrad, B., M. Wiekowski, B. E. Morrison, S. C. Chen, E. C. Coronel, D. J.Manfra, and S. A. Lira. 2002. Transient lung-specific expression of thechemokine KC improves outcome in invasive aspergillosis. Am. J. Respir.Crit Care Med. 166:1263–1268.
37. Miale, J. B. 1982. Laboratory medicine hematology, p. 864. The C. V. MosbyCompany, St. Louis, Mo.
38. Monga, D. P. 1983. Studies on experimental aspergillosis in immunodeficientmice. Zentrbl. Bakteriol. Mikrobiol. Hyg. A 254:552–560.
39. Morrison, B. E., S. J. Park, J. M. Mooney, and B. Mehrad. 2003. Chemo-kine-mediated recruitment of NK cells is a critical host defense mechanismin invasive aspergillosis. J. Clin. Investig. 112:1862–1870.
40. Mullins, J., R. Harvey, and A. Seaton. 1976. Sources and incidence ofairborne Aspergillus fumigatus (Fres). Clin. Allergy 6:209–217.
41. Mylonakis, E., T. F. Barlam, T. Flanigan, and J. D. Rich. 1998. Pulmonaryaspergillosis and invasive disease in AIDS. Chest 114:251–262.
42. Nakano, H., M. Yanagita, and M. D. Gunn. 2001. CD11c(�)B220(�)Gr-1(�) cells in mouse lymph nodes and spleen display characteristics of plas-macytoid dendritic cells. J. Exp. Med. 194:1171–1178.
43. Nosanchuk, J. D., and A. Casadevall. 1997. Cellular charge of Cryptococcus
124 STEPHENS-ROMERO ET AL. INFECT. IMMUN.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 21
Jan
uary
202
2 by
260
4:3d
09:a
17c:
3100
:e0b
d:1d
7f:c
91a:
e019
.

neoformans: contributions from the capsular polysaccharide, melanin, andmonoclonal antibody binding. Infect. Immun. 65:1836–1841.
44. Ortega, G., P. E. Korty, E. M. Shevach, and T. R. Malek. 1986. Role of Ly-6in lymphocyte activation. I. Characterization of a monoclonal antibody to anonpolymorphic Ly-6 specificity. J. Immunol. 137:3240–3246.
45. Pennington, J. E., and E. A. Harris. 1981. Influence of immunosuppressionon alveolar macrophage chemotactic activities in guinea pigs. Am. Rev.Respir. Dis. 123:299–304.
46. Pennington, J. E., W. J. Matthews, Jr., J. T. Marino, Jr., and H. R. Colten.1979. Cyclophosphamide and cortisone acetate inhibit complement biosyn-thesis by guinea pig bronchoalveolar macrophages. J. Immunol. 123:1318–1321.
47. Piggott, W. R., and C. W. Emmons. 1960. Device for inhalation exposure ofanimals to spores. Proc. Soc. Exp. Biol. Med. 103:805–806.
48. Rhame, F. S., A. J. Streifel, J. H. Kersey, Jr., and P. B. McGlave. 1984.Extrinsic risk factors for pneumonia in the patient at high risk of infection.Am. J. Med. 76:42–52.
49. Rogers, H. W., and E. R. Unanue. 1993. Neutrophils are involved in acute,nonspecific resistance to Listeria monocytogenes in mice. Infect. Immun.61:5090–5096.
50. Sayles, P. C., and L. L. Johnson. 1996. Exacerbation of toxoplasmosis inneutrophil-depleted mice. Nat. Immunol. 15:249–258.
51. Schaffner, A., H. Douglas, and A. Braude. 1982. Selective protection againstconidia by mononuclear and against mycelia by polymorphonuclear phago-cytes in resistance to Aspergillus. J. Clin. Investig. 69:617–631.
52. Sheppard, D. C., G. Rieg, L. Y. Chiang, S. G. Filler, J. E. Edwards, Jr., and
A. S. Ibrahim. 2004. Novel inhalational murine model of invasive pulmonaryaspergillosis. Antimicrob. Agents Chemother. 48:1908–1911.
53. Shibuya, K., M. Takaoka, K. Uchida, M. Wakayama, H. Yamaguchi, K.Takahashi, S. Paris, J. P. Latge, and S. Naoe. 1999. Histopathology ofexperimental invasive pulmonary aspergillosis in rats: pathological compar-ison of pulmonary lesions induced by specific virulent factor deficient mu-tants. Microb. Pathog. 27:123–131.
54. Singh, N. 2001. Trends in the epidemiology of opportunistic fungal infec-tions: predisposing factors and the impact of antimicrobial use practices.Clin. Infect. Dis. 33:1692–1696.
55. Smith, J. M., C. M. Tang, S. Van Noorden, and D. W. Holden. 1994. Viru-lence of Aspergillus fumigatus double mutants lacking restrictocin and analkaline protease in a low-dose model of invasive pulmonary aspergillosis.Infect. Immun. 62:5247–5254.
56. Stevenson, M. M., T. K. Kondratieva, A. S. Apt, M. F. Tam, and E. Skamene.1995. In vitro and in vivo T cell responses in mice during bronchopulmonaryinfection with mucoid Pseudomonas aeruginosa. Clin. Exp. Immunol. 99:98–105.
57. Todeschini, G., C. Murari, R. Bonesi, G. Pizzolo, G. Verlato, C. Tecchio, V.Meneghini, M. Franchini, C. Giuffrida, G. Perona, and P. Bellavite. 1999.Invasive aspergillosis in neutropenic patients: rapid neutrophil recovery is arisk factor for severe pulmonary complications. Eur. J. Clin. Investig. 29:453–457.
58. Young, R. C., J. E. Bennett, C. L. Vogel, P. P. Carbone, and V. T. DeVita.1970. Aspergillosis. The spectrum of the disease in 98 patients. Medicine49:147–173.
Editor: T. R. Kozel
VOL. 73, 2005 PATHOGENESIS OF ASPERGILLOSIS IN NEUTROPENIC MICE 125
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 21
Jan
uary
202
2 by
260
4:3d
09:a
17c:
3100
:e0b
d:1d
7f:c
91a:
e019
.