the p110 isoform of the cdp/cux transcription factor accelerates entry into s phase
TRANSCRIPT

MOLECULAR AND CELLULAR BIOLOGY, Mar. 2006, p. 2441–2455 Vol. 26, No. 60270-7306/06/$08.00�0 doi:10.1128/MCB.26.6.2441–2455.2006Copyright © 2006, American Society for Microbiology. All Rights Reserved.
The p110 Isoform of the CDP/Cux Transcription FactorAccelerates Entry into S Phase
Laurent Sansregret,1,2 Brigitte Goulet,1,2 Ryoko Harada,1,2 Brian Wilson,1,2
Lam Leduy,1 Jacques Bertoglio,5 and Alain Nepveu1,2,3,4*Molecular Oncology Group, McGill University Health Center,1 and Departments of Biochemistry,2 Oncology,3 and Medicine,4
McGill University, 687 Pine Ave. West, Montreal, Quebec, Canada H3A 1A1, and INSERM U461, Faculte de Pharmacie,Paris XI, 5 rue Jean Baptiste Clement, 92296 Chatenay-Malabry, France5
Received 26 September 2005/Returned for modification 31 October 2005/Accepted 29 December 2005
The CDP/Cux transcription factor was previously found to acquire distinct DNA binding and transcriptionalproperties following a proteolytic processing event that takes place at the G1/S transition of the cell cycle. Inthe present study, we have investigated the role of the CDP/Cux processed isoform, p110, in cell cycleprogression. Populations of cells stably expressing p110 CDP/Cux displayed a faster division rate and reachedhigher saturation density than control cells carrying the empty vector. p110 CDP/Cux cells reached the next Sphase faster than control cells under various experimental conditions: following cell synchronization in G0 bygrowth factor deprivation, synchronization in S phase by double thymidine block treatment, or enrichment inG2 by centrifugal elutriation. In each case, duration of the G1 phase was shortened by 2 to 4 h. Geneinactivation confirmed the role of CDP/Cux as an accelerator of cell cycle progression, since mouse embryofibroblasts obtained from Cutl1z/z mutant mice displayed a longer G1 phase and proliferated more slowly thantheir wild-type counterparts. The delay to enter S phase persisted following immortalization by the 3T3protocol and transformation with H-RasV12. Moreover, CDP/Cux inactivation hindered both the formation offoci on a monolayer and tumor growth in mice. At the molecular level, expression of both cyclin E2 and A2 wasincreased in the presence of p110 CDP/Cux and decreased in its absence. Overall, these results establish thatp110 CDP/Cux functions as a cell cycle regulator that accelerates entry into S phase.
CDP/Cux (CCAAT-displacement protein/cut homeobox)belongs to a family of transcription factors present in all meta-zoans and involved in the control of proliferation and differ-entiation (reviewed in reference 47). In Drosophila melano-gaster, mutations within cut coding sequences caused embryoniclethality late during larval development (28). In addition, alarge number of viable mutations were shown to cause de-velopmental alterations in the wings (“cut wing”), legs, ex-ternal sense organs, Malpighian tubules, tracheal system,and some structures in the central nervous systems (6–9, 13,21, 24–27, 35, 36, 43). At least in the wing margin, Cut wasshown to be a downstream effector of Notch (40, 41, 47, 49).In higher vertebrates, there are two Cut-like genes, calledCut-like 1 (CUTL1) and Cut-like 2 (CUTL2) in human andCut-homeobox 1 (Cux1) and Cut-homeobox 2 (Cux2) in mouseand chicken (48, 54, 67). While Cux1 is expressed in most tissues,Cux2 expression is restricted primarily to nervous tissues. ThecDNA for the human CUTL1 gene was originally cloned fol-lowing the purification of the CCAAT-displacement protein,CDP (48). Hereafter in the text, the term CDP/Cux will beused to describe the protein encoded by the human or mousegene. Genetic inactivation of the Cux1 locus in mice results inseveral phenotypes including perinatal lethality, curly whiskers,growth retardation, delayed differentiation of lung epithelia,altered hair follicle morphogenesis, male infertility, and a de-ficit in T and B cells (14, 39, 60, 66). In contrast to the small
size of the Cux1 mutant mice, Cux1 transgenic mice displayedmultiorgan hyperplasia and organomegaly, raising the possibil-ity that constitutive expression of Cux1 stimulated the prolif-eration of stem cells or the transient amplifying cells thatderive from them (32). Thus, from genetic studies with Dro-sophila and the mouse, the CDP/Cux/Cut gene plays an impor-tant role in the development and homeostasis of several tissues.
The full-length CDP/Cux protein, p200, contains four DNAbinding domains: three Cut repeats (CR1, CR2, and CR3) anda Cut homeodomain (2, 3, 19, 20, 48). Two DNA bindingactivities in cells have been characterized (19, 20, 44). p200CDP/Cux binds only transiently to DNA via CR1CR2 andcarries the CCAAT displacement activity (44). At the end ofthe G1 phase of the cell cycle, proteolytic cleavage of p200generates p110 CDP/Cux, which contains CR2CR3HD andexhibits distinct DNA binding specificity and kinetics (45). Inparticular, p110 but not p200 was shown to activate a DNApolymerase � gene reporter in transient-transfection assaysand to stimulate expression of the endogenous DNA polymer-ase � gene following the infection of cells with a high-titerretrovirus (45, 65).
CDP/Cux was found to function in precursor cells of variouslineages as a transcriptional repressor that down-modulatesgenes which later become expressed in terminally differenti-ated cells (34, 52, 61–63). This function was ascribed to theability of CDP/Cux to prevent the interaction of various tran-scriptional activators with their binding sites, probably via its“CCAAT displacement activity” (38, 44). More recently, CDP/Cux has been implicated as a downstream effector of trans-forming growth factor beta (TGF-�) in the promotion of cell
* Corresponding author. Mailing address: McGill University HealthCenter, Molecular Oncology Group, 687 Pine Avenue West, roomH5.21, Montreal, Quebec H3A 1A1, Canada. Phone: (514) 934-1934,ext. 35842. Fax: (514) 843-1478. E-mail: [email protected].
2441
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 13
Jan
uary
202
2 by
119
.237
.105
.46.

motility and invasion (42). Expression of CDP/Cux was in-creased following the treatment of cells with TGF-�, and CDP-specific small interfering RNA not only prevented the promi-gratory effects of TGF-� but also impaired the ability of tumorcells to form lung colonies in an experimental metastasis modelin vivo (42). In addition, a role for CDP/Cux specifically in theS phase of the cell cycle has been inferred from a number ofreports. Histone nuclear factor D (HiNF-D), which was laterfound to include CDP/Cux as its DNA binding partner, wasshown to be up-regulated in S phase in normal cells (22, 69,71–73). Up-regulation of CDP/Cux DNA binding at the G1/Stransition was found to result from at least two posttransla-tional modifications: dephosphorylation of the Cut homeodo-main by the Cdc25A phosphatase (11) and proteolytic cleavageof p200 CDP/Cux between CR1 and CR2 to generate N-ter-minally truncated p110 CDP/Cux (16, 45). The protease re-sponsible for proteolytic processing of CDP/Cux was shown tobe a nuclear isoform of cathepsin L that is devoid of a signalpeptide (16). The processed isoform, p110, was found to par-ticipate in the transcriptional activation of the DNA polymer-ase � gene and, at least in reporter assays, of a number of genesthat are up-regulated in S phase, like the dihydrofolate reduc-tase, carbamoyl-phosphate synthase/aspartate carbamoyl-trans-ferase/dihydroorotase, B-Myb, and cyclin A genes (65). The ac-tivity of the p110 isoform is confined primarily to the S phase ofthe cell cycle. Not only is p110 generated at the G1/S transition,but as cells progress into the G2 phase, DNA binding is even-tually down-modulated following phosphorylation by cyclinA/Cdk1 of serine 1237 in the region of the Cut homeodomain(58). CDP/Cux was found also to interact with cyclin A/Cdk2during S phase. In contrast to cyclin A/Cdk1, however, cyclinA/Cdk2 does not efficiently phosphorylate p110 CDP/Cux onserine 1237 and does not inhibit its DNA binding and tran-scriptional activities (59).
In the present study, we have generated populations of NIH3T3 fibroblastic cells and Kit225 T cells stably expressing p110CDP/Cux, and we have isolated mouse embryo fibroblasts fromCutl1�/� and Cutl1z/z mutant mice. Using these cells, we haveapplied functional tests to investigate whether the processedisoform of CDP/Cux, p110, has an impact on cell cycle pro-gression and cellular proliferation. Our study provides for thefirst time functional evidence that p110 CDP/Cux functions asa regulator of cell cycle progression that can accelerate entryinto S phase.
MATERIALS AND METHODS
Cell culture. NIH 3T3, mouse embryonic fibroblasts (MEFs), 3T3s, andHs578T cells were maintained in Dulbecco’s modified minimum essential me-dium (DMEM) supplemented with penicillin-streptomycin, glutamine, and 10%fetal bovine serum (FBS) (Gibco) (5% FBS for Hs578T). Kit225 T lymphocyteswere grown in RPMI 1640 (Wisent) containing 10% FBS, glutamine, penicillin-streptomycin, and 75 ng/ml of recombinant human interleukin 2 (IL-2) (PeproTechCanada, Inc.).
MEFs and 3T3 cells. Cutl1 mutant mice in the albino OF1 outbred strain wereobtained from the laboratory of Meinrad Busslinger and were maintained in theOF1 genetic background (14). Primary MEFs were prepared from 13.5-day-oldembryos, and heads were used for genotyping. Limbs and internal organs wereremoved, and the body was minced and incubated for 10 min in trypsin. Cellswere then washed once in complete medium and seeded in a 100-mm dish(passage 1). Two days later, the cells were trypsinized and seeded at 2 � 106
cells/100-mm dish for another 48 h before experiments were performed. Immor-
talization was done following the 3T3 protocol, where the cells were trypsinizedand seeded at the same density until immortalization was achieved (64).
Retroviral infection and stable cell lines. Retroviruses were produced bytranfecting 293VSV cells with plasmids encoding CDP/Cux (Myc tagged at the Nterminus and hemagglutinin (HA) tagged at the C terminus) inserted in thepREV/TRE plasmid (Clontech) or H-RasV12 inserted in pBabe (a kind gift fromScott Lowe). The supernatant was applied on cells at an equivalent titer, alongwith 8 �g/ml of polybrene (Roche), and plates were centrifuged at 300 � g for1 h. Stably infected cells were selected for 5 days in hygromycin (pREV/TRE)(Roche), and at least 500 resistant clones were pooled together for each popu-lation. Cell lines were infected with an empty vector or vectors expressing thefollowing proteins: p110 CDP/Cux (amino acids (aa) 878 to 1505) for NIH 3T3,p110 CDP/Cux (aa 747 to 1505) for Kit225, p110 CDP/Cux/TAP tag (aa 612 to1336-TAP tag) for Hs578T cells, and H-RasV12 for Cutl1 3T3 cells.
Proliferation curves. Cells were seeded at 5.5 � 103 cells/cm2 or 1.4 � 104
cells/cm2 for NIH 3T3 cells and MEFs, respectively. Each day, cells weretrypsinized and counted on a hemocytometer. The medium was replaced every 3days. Each time point was done in triplicate, and the averages � standarddeviations were calculated. Population doubling time (t2) was determined usingthe formula N(t) � N(0)X er t. The proliferation rate (r) was calculated betweendays 0 and 4.
Immunofluorescence. Cells were plated on glass coverslips and fixed in 3.7%paraformaldehyde. The cell membrane was solubilized in phosphate-bufferedsaline (PBS) containing 5% FBS and 0.5% Triton X-100. 5-Bromo-2�-deoxyuri-dine (BrdU) staining was done using a polyclonal sheep anti-BrdU polyclonalantibody (Biodesign International). For BrdU detection, the samples were incu-bated for 1 h in the solubilizing solution containing the anti-BrdU antibody and0.5 mg/ml DNase. Secondary detection was done with an Alexa Fluor 488-conjugated donkey anti-sheep antibody (Molecular Probes). Visualization wasdone using a Retiga 1300 digital camera (QIMAGING) and a Zeiss AxioVert135 microscope (Carl Zeiss Canada). Image analysis was carried out using North-ern Eclipse version 6.0 (Empix Imaging).
EMSA. Electrophoretic mobility shift assays (EMSA) were performed with 1�g of nuclear extract from mammalian cells. The samples were incubated atroom temperature for 5 min in a final volume of 30 �l of 25 mM NaCl, 10 mMTris, pH 7.5, 1 mM MgCl2, 5 mM EDTA, pH 8.0, 5% of glycerol, 1 mM ofdithiothreitol, with 100 ng of poly(dIdC) and 30 �g of bovine serum albumin asnonspecific competitors. End-labeled double-stranded oligonucleotides (5�-TCGAGACGATATCGATAAGCTTCTTTTC-3�) were added and further incubatedfor 15 min at room temperature. Samples were loaded on a 4% polyacrylamidegel (30:1) and separated by electrophoresis at 8 V/cm in Tris-glycine. Gels weredried and visualized by autoradiography.
Fluorescence-activated cell sorting (FACS) analysis. Cells were trypsinized,fixed in 75% ethyl alcohol, and stored at 20°C until analysis. For analysis, 50 �lof FBS was added to each sample. The cells were then centrifuged, washed inPBS, and resuspended in 300 �l of PBS containing 200 �g/ml of RNase (Sigma)and 5 �g/ml of propidium iodide (Sigma). Samples were incubated for 15 min at37°C and analyzed using a FACScan instrument (Becton Dickinson), using dou-blet discrimination to gate single cells. Cell cycle profiles were analyzed withFlowJo (Tree Star softwares). BrdU-labeled cells were fixed in 3.7% para-formaldehyde and incubated as described for immunofluorescence, except that amouse monoclonal anti-BrdU antibody conjugated to Alexa 488 was used (Mo-lecular Probes). Cells were then washed once and resuspended in a solutioncontaining propidium iodide and RNase.
Western blot analysis. Nuclear extracts were prepared according to the pro-cedure of Lee et al. (33), except that nuclei were obtained by submitting cells tothree freeze/thaw cycles in buffer A (10 mM HEPES, pH 7.9, 10 mM KCl, 1.5mM MgCl2, 1 mM dithiothreitol) (10) along with a protease inhibitor tablet(Roche). Sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) was performed with 8 �g of nuclear extract, and after electrophoretictransfer to polyvinylidene difluoride, membranes were washed in Tris-bufferedsaline–0.1% Tween 20 (TBS 0.1%T) and blocked in TBS 0.1%T containing 5%milk. Membranes were probed with antibodies directed against HA (1:2,000,MMS-101R; Covance,), p21Cip1 (1:1,000, anti-Waf1 Ab5; Calbiochem), cyclin A(1:700, Ab-5; Neomarkers), CDP/Cux (1:3,000, anti-1300; 1:2,000, anti-861) (45),cyclin E (1:1,000, Ab-1; NeoMarkers), cyclin E2 (1:500; Epitomics), Cdk2 (1:700,Ab-4; NeoMarkers), actin (1:1,000, I-19; Santa Cruz Biotechnology), CBP p300(1:2,000, C-20; Santa Cruz Biotechnology), Ras (1:2,000, C-20; Santa Cruz Bio-technology). Primary antibodies were incubated in TBS 0.1%T, and detectionwas done using a horseradish peroxidase-conjugated antirabbit or antimousesecondary antibody in TBS 0.1%T. Immunoreactive proteins were visualized bychemiluminescence with an ECL Western blotting detection kit (AmershamPharmacia Biotech).
2442 SANSREGRET ET AL. MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 13
Jan
uary
202
2 by
119
.237
.105
.46.

Kinase assay. Two milligrams of total cell lysates were immunoprecipitatedwith anti-cyclin E1 (NeoMarkers Ab-1) or anti-cyclin E2 (NeoMarkers Ab-1)antibody using protein A-Sepharose beads. Kinase reactions were done using 1�g histone H1, [32P]ATP in kinase buffer (50 mM HEPES, pH 7.5, 10 mMMgCl2, 1 mM NaF, and 0.5 mM NaVO4). Samples were resolved on a 12%SDS-PAGE gel and visualized by autoradiography.
Cell synchronization and centrifugal elutriation. For serum starvation exper-iments, cells were plated in 60-mm dishes in DMEM–10% FBS. The next day,the medium was replaced with serum-free DMEM and cells were incubated for84 h. Cells were stimulated to enter the cell cycle with DMEM–10% FBS andharvested at the indicated time points. For Fig. 3B, BrdU was added at a finalconcentration of 100 �M 8 h poststimulation in every plate, and the 0-h timepoint was pulsed for 2 h before fixation. Double thymidine block was done byincubating the cell in DMEM–10% FBS containing 2 mM thymidine overnight.Cells were then washed and incubated in fresh DMEM–10% FBS for 8 h. Asecond overnight incubation in 2 mM thymidine was done before washing andreleasing the block in DMEM–10% FBS. Centrifugal elutriation was carried outusing an Avanti J-20 centrifuge equipped with a JE 5.0 rotor and a standardchamber (Beckman Coulter) connected to a Pump Masterflex L/S pump (Cole-Parmer instrument company) (29). All steps were performed at room tempera-ture in PBS–1% FBS, and centrifugation was carried out at 2,460 rpm. Condi-tioned medium was kept at 37°C, and cells were trypsinized and counted on ahemocytometer. Cells (5 � 107) were loaded into the chamber, and equilibrationwas done at a flow rate of 23 ml/min for 25 min. The flow rate was increasedsuccessively by 2 ml/min for each 100-ml fraction. Cells from fractions collectedat flow rates of 65, 67, and 69 ml/min (the most highly enriched fractions for cellswith a G2/M DNA content) were centrifuged at 800 � g, counted, and plated in60-mm dishes containing conditioned medium. Synchronization of Kit225 cellswas done by rinsing the cells twice in PBS and culturing 5 � 105 cells/ml withoutIL-2 for 48 h. Cells were then seeded at a density of 2 � 105 cells/ml inIL-2-containing medium and collected at different time points. MEFs at passage3 were rendered quiescent by seeding 3 � 106 cells/100-mm dish, and 48 h laterthe medium was replaced with DMEM–0.1% FBS for an additional 72 h. MEFswere then trypsinized and seeded in DMEM–10% FBS at a density of 3.3 � 104
cells/cm2 to follow entry in S phase.Focus formation assays. 3T3 cells were infected with a pBabe or pBabe/H-
RasV12, pLXSN or pLXSN/p200, and pRev or pRev/p110 retroviruses. Forty-eight hours later, cells were split 1:2 and left for focus formation in DMEMsupplemented with 5% FBS or 10% FBS plus antibiotics (puromycin [1 �g/ml]),for a total of 12 10-cm dishes per condition. Medium was changed every 3 to 4days. Foci were fixed with buffered formalin phosphate (Fisher scientific) for 1 hand stained with Giemsa.
Tumor formation in vivo. Following infection, cells were selected for 5 dayswith puromycin and trypsinized once before injection. Tumor formation wasmeasured periodically. Four- to five-week-old female nude mice (CD1 nu/nu;Charles River Breeding Laboratories) were subcutaneously injected with 104 3T3cells.
RNA extraction, reverse transcriptase PCR (RT-PCR), and real-time PCR.RNA was extracted using TRIzol reagent (Invitrogen), and cDNA was preparedusing the Superscript II RNase H-reverse transcriptase kit (Invitrogen) followingthe manufacturer’s instructions. Real-time PCR was performed on a LightCyclerinstrument (Roche) using the FastStart DNA Master SYBR Green kit (Roche).The following primers were used: cyclin D1, 5�-TGCATGTTCGTGGCCTCTAA-3� and 5�-CAGTCCGGGTCACACTTGAT-3�; cyclin E1, 5�-CCAAAGTTGCACCAGTTTGC-3� and 5�-CTCTATGTCGCACCACTGAT-3�; cyclinE2, 5�-CCTCCATTGAAGTGGTTAAGAAAGC-3� and 5�-AGTGTTTTCCTGGTGGTTTTTCAGT-3�; cyclin A2, 5�-CACCATTCATGTGGATGAAGC-3�and 5�-TGCAGGGTCTCATTCTGTAG-3�; Cdc25A, 5�-TTCTCCAAGGGTTATCTCTT-3� and 5�-AAGAACTCCTTGTATCCCC-3�; DNA polymerase �,5�-GGGGTAATGGAGTTTGAAGACGG-3� and 5�-GGAAGGAAGTAGAGTGTTCGCTC-3�; glyceraldehyde-3-phosphate dehydrogenase, 5�-ACCACAGTCCATGCCATCAC-3� and 5�-TCCACCACCCTGTTGCTGTA-3�; p27Kip1, 5�-CCCGCCCGAGGAGGAAGATGTCAAAC-3� and 5�-CCCTTTTGTTTTGCGAAGAAGAATCT-3�; p21Cip1, 5�-CCGTGGACAGTGAGCAGTTG-3� and 5�-TGGGCACTTCAGGGTTTTCT-3�.
ChIP and ChAP. For each immunoprecipitation, 3 � 108 cells were cross-linked in the presence of 1% formaldehyde for 10 min at room temperature andthen quenched in 0.125 M glycine for 5 min at room temperature. Cells werewashed twice in PBS and were scraped. The pelleted cells were resuspended inLysis I buffer (50 mM HEPES-KOH, pH 8, 2 mM EDTA, 2 mM EGTA, 140 mMNaCl, 10% glycerol, 0.5% NP-40, 0.25% Triton X-100, 1 mM phenylmethyl-sulfonyl fluoride [PMSF], protease inhibitors [Roche Molecular Biochemicals])and incubated at 4°C for 10 min. Nuclei were isolated by centrifugation, washed
once in Lysis II buffer (10 mM Tris-HCl, pH 8; 2 mM EDTA; 2 mM EGTA; 200mM NaCl, 1 mM PMSF, protease inhibitors). Nuclei were then lysed in RIPA-Mbuffer (10 mM Tris-HCl, pH 8, 1 mM EDTA, 0.5 mM EGTA, 150 mM NaCl, 1%Triton X-100, 0.5% DOC, 0.1% SDS, 1 mM PMSF, protease inhibitors) andsonicated on ice to obtain 250- to 800-bp-long DNA fragments. Cells debris wasremoved by centrifugation, and supernatants were precleared for 1 h with 25 �lof protein A-agarose beads containing salmon sperm DNA (Upstate Biotech-nology). After brief centrifugation, supernatant was removed and incubated with2 �g of the purified CDP/Cux antibody overnight at 4°C. The immune complexwas collected the following day by incubating with the 25 �l of protein A-agarosebeads at 4°C for 1 h. Complexes were washed three times in low-salt buffer (20mM Tris-HCl, pH 8, 2 mM EDTA, 2 mM EGTA, 150 mM NaCl, 1% NP-40,0.5% deoxycholate (DOC), 0.2% SDS), three times in high-salt buffer (20 mMTris-HCl, pH 9, 2 mM EDTA, 2 mM EGTA, 500 mM NaCl, 1% NP-40, 0.5%DOC, 0.1% SDS), three times in LiCl buffer (50 mM Tris-HCl, pH 7.5, 2 mMEDTA, 1 mM EGTA, 0.5 M LiCl, 1% NP-40, 0.7% DOC,) and then washed oncein Tris-EDTA. DNA was eluted in 100 �l of 1% SDS, 10 mM Tris-HCl, pH 8, 10mM EDTA at 65°C for 30 min. The eluates were incubated at 65°C overnight toreverse the cross-links. After proteinase K digestion, DNA was recovered byphenol-chloroform extraction and ethanol precipitation and then used in PCR.For chromatin affinity purification (ChAP), we used stably infected Hs578T cellsexpressing a recombinant CDP/Cux protein fused to a tandem affinity purifica-tion tag. Cross-linked cells (3 � 108) were used for each purification. Cell pelletswere lysed in RIPA-M buffer and sonicated on ice to obtain 250- to 800-bp-longDNA fragments. After centrifugation, the supernatant was used for tandemaffinity purification as described previously (56a). Briefly, complexes containingDNA cross-linked to the CDP/Cux fusion protein were recovered on an immu-noglobulin G (IgG) matrix. After tobacco etch virus protease digestion, thereleased protein-DNA complexes were purified by affinity on calmodulin beadsin the presence of calcium and then eluted with EGTA. Eluates were incubatedat 65°C overnight to reverse the cross-links. After proteinase K digestion, DNAwas recovered by phenol-chloroform extraction and ethanol precipitation andthen used in PCR. Immunoprecipitated DNA by chromatin immunoprecipita-tion (ChIP) or ChAP was subjected to quantitative PCR amplification. PCR wasperformed using human cyclin A2 promoter-specific primers (5�-GCAGGGCCGAGGAGGT-3� and 5�-GCGCTTTCATTGGTCCATTT-3�. To normalize sam-ples by the amount of nonspecific DNA, we amplified a region of the humanglucose-6-phosphate dehydrogenase gene (5�-GGATGATCCCAAATTCATCG-3� and 5�-AGGTCAGTTCCTCCACCTTG-3�). PCR products were re-solved on a 6% polyacrylamide gel.
RESULTS
Generation of NIH 3T3 cells overexpressing p110 CDP/Cux.To study the effect of p110 CDP/Cux on cell cycle progression,we established a population of NIH 3T3 cells stably expressinga recombinant p110 protein from a retroviral vector. As acontrol, we generated a population of NIH 3T3 cells contain-ing the empty retroviral vector. To limit the effect of endoge-nous regulatory sequences at the integration sites, populationsof more than 500 clones were isolated. From indirect immu-nofluorescence analysis, ectopic p110 CDP/Cux was detectedas a nuclear signal in 100% of cells in the NIH 3T3/p110population (Fig. 1A). The expression and DNA binding activ-ity of recombinant p110 CDP/Cux were investigated in con-tinuously proliferating cells and following serum starvationand restimulation. From Western blot analysis, elevated p110CDP/Cux expression was detected at all time points and didnot vary significantly (Fig. 1B, lanes 2 to 7). In contrast, ex-pression of endogenous p200 CDP/Cux did not change signif-icantly in NIH 3T3/p110 cells (Fig. 1B, lanes 8 and 9). Anelectrophoretic mobility shift assay was performed using 1 �gof nuclear extract and a probe containing the CDP/Cux con-sensus binding site, ATCGAT (Fig. 1C). Note that DNA bind-ing by endogenous proteins cannot be detected with this lim-ited amount of nuclear extract, as shown by the absence ofretarded complex in the control NIH 3T3/vector lane (Fig. 1C,
VOL. 26, 2006 p110 CDP/Cux ACCELERATES CELL CYCLE PROGRESSION 2443
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 13
Jan
uary
202
2 by
119
.237
.105
.46.
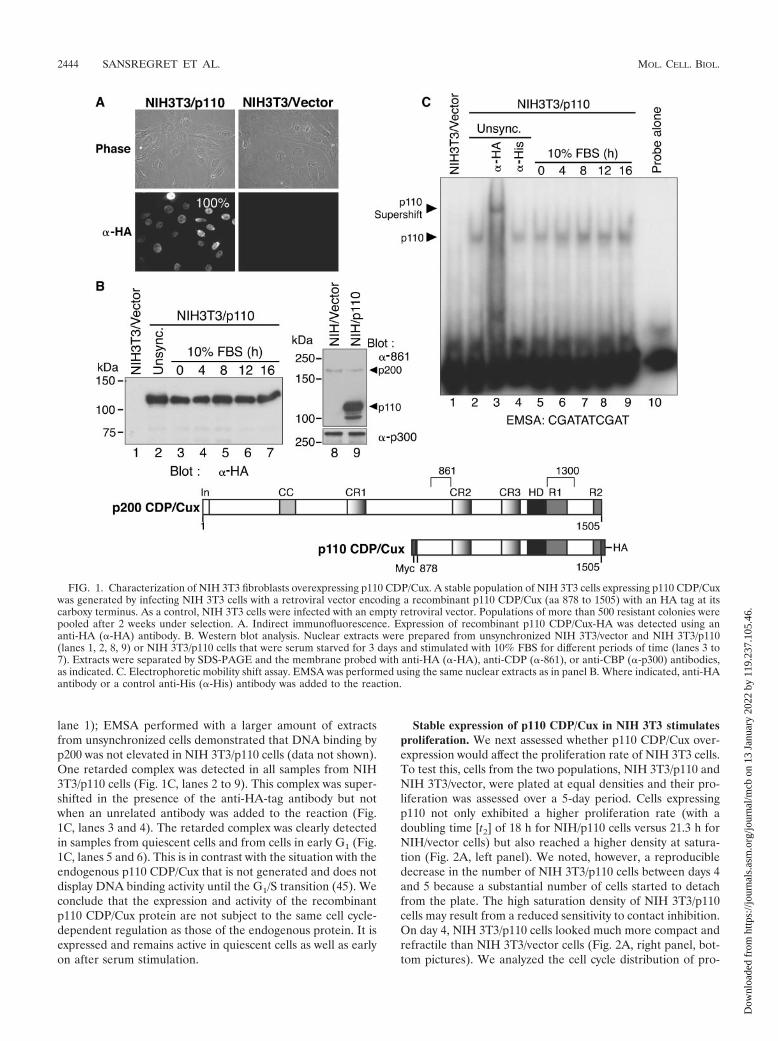
lane 1); EMSA performed with a larger amount of extractsfrom unsynchronized cells demonstrated that DNA binding byp200 was not elevated in NIH 3T3/p110 cells (data not shown).One retarded complex was detected in all samples from NIH3T3/p110 cells (Fig. 1C, lanes 2 to 9). This complex was super-shifted in the presence of the anti-HA-tag antibody but notwhen an unrelated antibody was added to the reaction (Fig.1C, lanes 3 and 4). The retarded complex was clearly detectedin samples from quiescent cells and from cells in early G1 (Fig.1C, lanes 5 and 6). This is in contrast with the situation with theendogenous p110 CDP/Cux that is not generated and does notdisplay DNA binding activity until the G1/S transition (45). Weconclude that the expression and activity of the recombinantp110 CDP/Cux protein are not subject to the same cell cycle-dependent regulation as those of the endogenous protein. It isexpressed and remains active in quiescent cells as well as earlyon after serum stimulation.
Stable expression of p110 CDP/Cux in NIH 3T3 stimulatesproliferation. We next assessed whether p110 CDP/Cux over-expression would affect the proliferation rate of NIH 3T3 cells.To test this, cells from the two populations, NIH 3T3/p110 andNIH 3T3/vector, were plated at equal densities and their pro-liferation was assessed over a 5-day period. Cells expressingp110 not only exhibited a higher proliferation rate (with adoubling time [t2] of 18 h for NIH/p110 cells versus 21.3 h forNIH/vector cells) but also reached a higher density at satura-tion (Fig. 2A, left panel). We noted, however, a reproducibledecrease in the number of NIH 3T3/p110 cells between days 4and 5 because a substantial number of cells started to detachfrom the plate. The high saturation density of NIH 3T3/p110cells may result from a reduced sensitivity to contact inhibition.On day 4, NIH 3T3/p110 cells looked much more compact andrefractile than NIH 3T3/vector cells (Fig. 2A, right panel, bot-tom pictures). We analyzed the cell cycle distribution of pro-
FIG. 1. Characterization of NIH 3T3 fibroblasts overexpressing p110 CDP/Cux. A stable population of NIH 3T3 cells expressing p110 CDP/Cuxwas generated by infecting NIH 3T3 cells with a retroviral vector encoding a recombinant p110 CDP/Cux (aa 878 to 1505) with an HA tag at itscarboxy terminus. As a control, NIH 3T3 cells were infected with an empty retroviral vector. Populations of more than 500 resistant colonies werepooled after 2 weeks under selection. A. Indirect immunofluorescence. Expression of recombinant p110 CDP/Cux-HA was detected using ananti-HA (�-HA) antibody. B. Western blot analysis. Nuclear extracts were prepared from unsynchronized NIH 3T3/vector and NIH 3T3/p110(lanes 1, 2, 8, 9) or NIH 3T3/p110 cells that were serum starved for 3 days and stimulated with 10% FBS for different periods of time (lanes 3 to7). Extracts were separated by SDS-PAGE and the membrane probed with anti-HA (�-HA), anti-CDP (�-861), or anti-CBP (�-p300) antibodies,as indicated. C. Electrophoretic mobility shift assay. EMSA was performed using the same nuclear extracts as in panel B. Where indicated, anti-HAantibody or a control anti-His (�-His) antibody was added to the reaction.
2444 SANSREGRET ET AL. MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 13
Jan
uary
202
2 by
119
.237
.105
.46.

liferating cells from each population at two different densities(Fig. 2B). We labeled S-phase cells with BrdU and measuredDNA content of cells by staining with propidium iodide andFACS analysis. At subconfluent density (corresponding to day2 in Fig. 2A), the NIH 3T3/p110 population had twice as manycells actively synthesizing DNA as the control cells. Of note,this shift in cell cycle distribution was at the expense of theG0/G1 phase, suggesting that overexpression of p110 CDP/Cuxmay shorten the time cells spend in G1 while weakly affectingcompletion of mitosis.
p110 CDP/Cux accelerates entry into S phase upon exit fromquiescence. We next investigated cell cycle progression aftersynchronization in G0/G1. Cells were rendered quiescent by a3-day incubation in serum-free medium and then stimulated toreenter the cell cycle with DMEM containing 10% FBS. Cellswere collected at various time points and their DNA contentmeasured by propidium iodide staining and FACS analysis.Two differences were noted between the cell populations.First, NIH 3T3/p110 cells were not as efficiently synchronizedin G0/G1 as the control NIH 3T3/vector cells. After 3 days inlow-serum medium (0 h), 16% of NIH 3T3/p110 cells were stillin S phase compared to 7% of NIH 3T3/vector cells (Fig. 3Aand C). In order to verify whether those cells were activelysynthesizing DNA, we measured how serum starvation was
affecting DNA replication by measuring BrdU incorporationduring the starvation process (Fig. 3C). The NIH 3T3/p110population displays twice as many cells actively synthesizingDNA, suggesting that p110 CDP/Cux might alter the responseof cells to the scarcity in growth factors. Secondly, NIH 3T3/p110 cells consistently entered S phase earlier than controlcells upon stimulation with 10% FBS. The fraction of NIH3T3/p110 cells in G0/G1 decreased faster and the fraction in Sphase increased faster than for the control cells (Fig. 3A). Thefirst significant increase in the S-phase fraction was observedbetween 10 h and 12 h for NIH 3T3/p110 cells and between12 h and 14 h for NIH 3T3/vector cells. Interestingly, many ofthe NIH 3T3/p110 cells appeared to complete the whole cellcycle in 20 h, whereas the NIH 3T3/vector cells were stillreaching G2/M (Fig. 3A, note the increase in G0/G1 between18 h and 20 h).
As an alternative approach to monitor the onset of DNAsynthesis following serum stimulation, BrdU was added to themedium 8 h poststimulation and cells were continuously la-beled thereafter. The percentage of cells having incorporatedBrdU at each time point was scored by indirect immunofluo-rescence (Fig. 3B). Comparison of the results at the 10-h and12-h time points shows that NIH 3T3/p110 cells started syn-thesizing DNA faster than control cells. The 50% labeling time
FIG. 2. p110 CDP/Cux stimulates proliferation in NIH 3T3 fibroblasts. A. Proliferation curves. Cells (5.5 � 103/cm2) were seeded in triplicateand counted daily for 5 days. Each point represents the average � standard deviation. The graph is a representative example of three independentexperiments. The doubling time (t2) is for the period from day 0 to day 4. On the right, cells were photographed at different days following seeding.B. Cell cycle distribution of unsynchronized cells. Cells were seeded at equal densities in triplicate and harvested after 48 h (subconfluent) or 96 h(confluent). For each time point, the cells were pulsed for 1 h with 100 �M BrdU, fixed, and stained with propidium iodide. The percentage ofcells in each phase was determined by FACS analysis using an anti-BrdU antibody conjugated to Alexa 488 and propidium iodide. Error barsrepresent standard deviations (Std. Dev.).
VOL. 26, 2006 p110 CDP/Cux ACCELERATES CELL CYCLE PROGRESSION 2445
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 13
Jan
uary
202
2 by
119
.237
.105
.46.

was reached slightly after 14 h in the case of NIH 3T3/p110 cellsand at 18 h for NIH 3T3/vector cells. We note that the differencesbetween the two cell populations persisted at the later timepoints. Altogether, these results indicate that stable expression ofp110 CDP/Cux accelerates entry into S phase following exit fromquiescence.
To determine whether the effect of p110 CDP/Cux on cellcycle progression was limited to fibroblasts, we did a similarexperiment using the IL-2-dependent human T-cell line, Kit225.Populations of cells stably carrying the empty vector or thep110 CDP/Cux vector were generated by retroviral infection(Fig. 4A). Synchronization in G0/G1 was achieved by incuba-tion in IL-2-free medium for 48 h, after which cells werecentrifuged and resuspended at a density of 2 � 105 cells/ml inIL-2-containing RPMI. Similar to what we observed with NIH3T3 cells, Kit225/p110 cells entered S phase faster upon exitfrom quiescence (Fig. 4B). These findings indicate that theeffects of p110 CDP/Cux on cell cycle progression are notlimited to one cell type.
Cells fractionated by counterflow centrifugal elutriation tra-verse the G1 phase faster upon ectopic expression of p110CDPCux. The first cycle following exit from quiescence isdifferent from the subsequent cycles in continuously prolif-erating cells. In particular, the G1 phase is normally longerduring the first cycle after an arrest in G0 than during thesubsequent cycles. Counterflow centrifugal elutriation hasthe advantage of separating cells based on their sizes andtheir densities in a short period of time (29). It allows theisolation of fractions enriched for cells in the same phase ofthe cell cycle without the use of drugs. We used this methodto obtain a fraction of cells enriched in the G2 phase of thecell cycle. Cells were replated, and their progressionthrough the cell cycle was monitored by FACS analysis.Cells from the two populations efficiently completed mitosisand moved into G1 with similar kinetics (Fig. 5, 8 h and 9 h).However, NIH 3T3/p110 cells entered the S and G2 phasesfaster than control cells (Fig. 5, S phase at 10 h and 11 h; G2
phase at 16 h and 18 h). These results establish that in
FIG. 3. p110 CDP/Cux accelerates entry into S phase upon exit from quiescence. A. Serum starvation and restimulation. Cells were seeded atequal densities for 24 h in DMEM–10% FBS and synchronized by serum starvation and restimulation as described in Materials and Methods.Nuclei were stained with propidium iodide, and DNA content was determined by FACS analysis. Cell cycle profiles were analyzed using FlowJo.B. Induction of DNA synthesis after serum starvation and restimulation. Cells were seeded on glass coverslips and treated as for panel A. Eighthours after stimulation, BrdU (100 �M) was added to the medium to continuously label the cells as they reach S phase. Coverslips were fixed in3.7% paraformaldehyde at each time point and stained using an anti-BrdU antibody for indirect immunofluorescence. At least 450 cells for eachtime point were randomly chosen to score for BrdU incorporation, and the average of three independent experiments is represented � standarddeviation. C. Effect of serum withdrawal on DNA synthesis. The cells were seeded at equal densities in triplicate for 24 h, and medium was replacedfor DMEM–0.1% FBS on day 0. Cells were pulsed with 100 �m BrdU for 1 h for each time point and analyzed by FACS to determine thepercentage of BrdU-positive cells.
2446 SANSREGRET ET AL. MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 13
Jan
uary
202
2 by
119
.237
.105
.46.

continuously proliferating cells, ectopic expression of p110CDP/Cux accelerates entry into S phase without compro-mising progression into G2 phase.
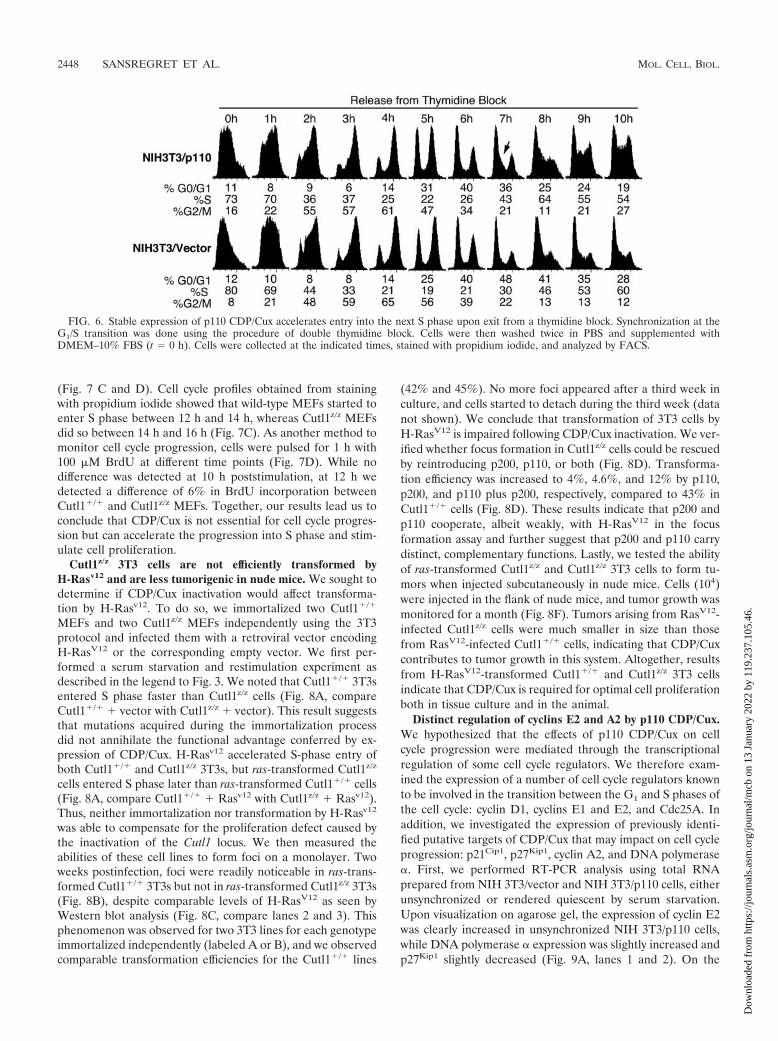
p110 CDP/Cux accelerates entry into the next S phase uponexit from a thymidine block. We wanted to verify whetherectopic expression of p110 CDP/Cux would affect progressionthrough the last phases of the cell cycle progression followingsynchronization by another method. The double thymidineblock method was used to obtain cells synchronized at the G1/S
transition, and cell cycle progression was monitored upon re-moval of thymidine. We found that progression through Sphase and G2/M was not altered in NIH 3T3/p110 cells, sincethey reached G1 of the next cycle at the same time as controlcells (Fig. 6, 1 h to 6 h). However, NIH 3T3/p110 cells reachedS phase of the following cycle before control cells (Fig. 6, 7 hand 8 h.). Thus, ectopic expression of p110 CDP/Cux acceler-ates passage through the G1 phase following both serum star-vation and a thymidine block, yet progression through the Sand G2 phases was not affected.
Lack of a functional CDP/Cux does not preclude prolifera-tion but slows it down. Since ectopic expression of p110 CDP/Cux stimulates proliferation and S-phase entry, we investigatedwhether the absence of functional CDP/Cux would affect cel-lular proliferation. To do so, we derived primary MEFs fromCutl1�/� and Cutl1z/z littermates. In the Cutl1z/z mice, theCutl1 locus expresses a CDP/Cux-LacZ fusion protein thatincludes the N-terminal region of Cux up to the start of the Cutrepeat 3. Therefore, this protein remains in the cytoplasm (14).In agreement with previous reports, disruption of the Cutl1locus merely affected the cell cycle distribution of primaryMEFs (Fig. 7A) (14, 39). On the other hand, we found thatCutl1z/z MEFs were slightly retarded in their proliferationcompared to wild-type MEFs. Since this result contrasted withwhat was originally reported, we performed the experimentwith four different litters for a total of seven wild-type andeight Cutl1z/z embryos (Fig. 7B). Although we observed differ-ences between separate experiments in the overall rate of di-vision and maximal density of cells at saturation, in all exper-iments the proliferation of Cutl1z/z MEFs was reproduciblyslower than that of wild-type MEFs (Fig. 7B). We also testedthe ability of Cutl1z/z MEFs to enter S phase following exitfrom quiescence. Essentially, MEFs were made quiescent bycultivating them at high density in 0.1% FBS, and reentry intothe cell cycle was induced by trypsinizing and seeding cells atlower density in 10% FBS. Using this protocol, we found thatCutl1z/z MEFs did not reach S phase as fast as wild-type MEFs
FIG. 4. p110 CDP/Cux accelerates S-phase entry following IL-2starvation and restimulation of human Kit225 T cells. IL-2 deprivationand stimulation of Kit225 T lymphocytes. A population of Kit225 cellsexpressing a recombinant p110 CDP/Cux protein was generated usingthe same retrovirus as for Fig. 1. A. Western blot analysis: ectopicCDP/Cux protein was detected using an anti-Myc (�-Myc) antibody. B.Proliferating Kit225 cells were cultured without IL-2 for 48 h, afterwhich the cells were seeded in IL-2-containing medium. Cells werecollected at the indicated time points, stained with propidium iodide,and analyzed by FACS.
FIG. 5. NIH 3T3/p110 cells fractionated by elutriation display a shorter G1 phase. Unsynchronized NIH 3T3/p110 and NIH 3T3/vector weretrypsinized and fractionated by counterflow centrifugal elutriation as described in Materials and Methods. Fractions enriched for cells with a 4NDNA content were seeded at equal density in prewarmed conditioned medium. Cells were collected at the indicated times, stained with propidiumiodide, and analyzed by FACS.
VOL. 26, 2006 p110 CDP/Cux ACCELERATES CELL CYCLE PROGRESSION 2447
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 13
Jan
uary
202
2 by
119
.237
.105
.46.
(Fig. 7 C and D). Cell cycle profiles obtained from stainingwith propidium iodide showed that wild-type MEFs started toenter S phase between 12 h and 14 h, whereas Cutl1z/z MEFsdid so between 14 h and 16 h (Fig. 7C). As another method tomonitor cell cycle progression, cells were pulsed for 1 h with100 �M BrdU at different time points (Fig. 7D). While nodifference was detected at 10 h poststimulation, at 12 h wedetected a difference of 6% in BrdU incorporation betweenCutl1�/� and Cutl1z/z MEFs. Together, our results lead us toconclude that CDP/Cux is not essential for cell cycle progres-sion but can accelerate the progression into S phase and stim-ulate cell proliferation.
Cutl1z/z 3T3 cells are not efficiently transformed byH-Rasv12 and are less tumorigenic in nude mice. We sought todetermine if CDP/Cux inactivation would affect transforma-tion by H-Rasv12. To do so, we immortalized two Cutl1�/�
MEFs and two Cutl1z/z MEFs independently using the 3T3protocol and infected them with a retroviral vector encodingH-RasV12 or the corresponding empty vector. We first per-formed a serum starvation and restimulation experiment asdescribed in the legend to Fig. 3. We noted that Cutl1�/� 3T3sentered S phase faster than Cutl1z/z cells (Fig. 8A, compareCutl1�/� � vector with Cutl1z/z � vector). This result suggeststhat mutations acquired during the immortalization processdid not annihilate the functional advantage conferred by ex-pression of CDP/Cux. H-Rasv12 accelerated S-phase entry ofboth Cutl1�/� and Cutl1z/z 3T3s, but ras-transformed Cutl1z/z
cells entered S phase later than ras-transformed Cutl1�/� cells(Fig. 8A, compare Cutl1�/� � Rasv12 with Cutl1z/z � Rasv12).Thus, neither immortalization nor transformation by H-Rasv12
was able to compensate for the proliferation defect caused bythe inactivation of the Cutl1 locus. We then measured theabilities of these cell lines to form foci on a monolayer. Twoweeks postinfection, foci were readily noticeable in ras-trans-formed Cutl1�/� 3T3s but not in ras-transformed Cutl1z/z 3T3s(Fig. 8B), despite comparable levels of H-RasV12 as seen byWestern blot analysis (Fig. 8C, compare lanes 2 and 3). Thisphenomenon was observed for two 3T3 lines for each genotypeimmortalized independently (labeled A or B), and we observedcomparable transformation efficiencies for the Cutl1�/� lines
(42% and 45%). No more foci appeared after a third week inculture, and cells started to detach during the third week (datanot shown). We conclude that transformation of 3T3 cells byH-RasV12 is impaired following CDP/Cux inactivation. We ver-ified whether focus formation in Cutl1z/z cells could be rescuedby reintroducing p200, p110, or both (Fig. 8D). Transforma-tion efficiency was increased to 4%, 4.6%, and 12% by p110,p200, and p110 plus p200, respectively, compared to 43% inCutl1�/� cells (Fig. 8D). These results indicate that p200 andp110 cooperate, albeit weakly, with H-RasV12 in the focusformation assay and further suggest that p200 and p110 carrydistinct, complementary functions. Lastly, we tested the abilityof ras-transformed Cutl1z/z and Cutl1z/z 3T3 cells to form tu-mors when injected subcutaneously in nude mice. Cells (104)were injected in the flank of nude mice, and tumor growth wasmonitored for a month (Fig. 8F). Tumors arising from RasV12-infected Cutl1z/z cells were much smaller in size than thosefrom RasV12-infected Cutl1�/� cells, indicating that CDP/Cuxcontributes to tumor growth in this system. Altogether, resultsfrom H-RasV12-transformed Cutl1�/� and Cutl1z/z 3T3 cellsindicate that CDP/Cux is required for optimal cell proliferationboth in tissue culture and in the animal.
Distinct regulation of cyclins E2 and A2 by p110 CDP/Cux.We hypothesized that the effects of p110 CDP/Cux on cellcycle progression were mediated through the transcriptionalregulation of some cell cycle regulators. We therefore exam-ined the expression of a number of cell cycle regulators knownto be involved in the transition between the G1 and S phases ofthe cell cycle: cyclin D1, cyclins E1 and E2, and Cdc25A. Inaddition, we investigated the expression of previously identi-fied putative targets of CDP/Cux that may impact on cell cycleprogression: p21Cip1, p27Kip1, cyclin A2, and DNA polymerase�. First, we performed RT-PCR analysis using total RNAprepared from NIH 3T3/vector and NIH 3T3/p110 cells, eitherunsynchronized or rendered quiescent by serum starvation.Upon visualization on agarose gel, the expression of cyclin E2was clearly increased in unsynchronized NIH 3T3/p110 cells,while DNA polymerase � expression was slightly increased andp27Kip1 slightly decreased (Fig. 9A, lanes 1 and 2). On the
FIG. 6. Stable expression of p110 CDP/Cux accelerates entry into the next S phase upon exit from a thymidine block. Synchronization at theG1/S transition was done using the procedure of double thymidine block. Cells were then washed twice in PBS and supplemented withDMEM–10% FBS (t � 0 h). Cells were collected at the indicated times, stained with propidium iodide, and analyzed by FACS.
2448 SANSREGRET ET AL. MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 13
Jan
uary
202
2 by
119
.237
.105
.46.

other hand, cyclin A2 expression was higher in quiescent NIH3T3/p110 cells (Fig. 9A, lanes 3 and 4). Since down-regulationof p27Kip1 and up-regulation of cyclin A2 or E-type cyclins canall promote entry in S phase, their expression was measured bysemiquantitative real-time PCR. Significant differences in theexpression of cyclins E2 and A2 were observed in p110-ex-pressing cells but in different situations. While cyclin E2 expres-sion was increased 18-fold in unsynchronized cells, cyclin A2 ex-pression was increased 2-fold in quiescent cells only (data notshown). Western blot analysis confirmed the distinct regulation ofcyclin E2 and cyclin A2 downstream of p110 CDP/Cux. In unsyn-chronized cells, cyclin E2 levels were much higher in NIH 3T3/
p110 cells than NIH 3T3/vector cells, whereas cyclin A2, cyclinE1, or Cdk2 levels did not change appreciably (Fig. 9B). In agree-ment with these results, cyclin E2-associated kinase activity to-wards histone H1 was higher in NIH 3T3/p110 cells, whereascyclin E1-associated kinase activity was not significantly changed(Fig. 9B). Following serum starvation and restimulation, we ob-served a significant increase in cyclin A2 protein expression in thep110 cells both at 0 h and at 4 h following serum addition (Fig. 9C,compare lanes 1 and 2 with 8 and 9). In contrast, increasedexpression of cyclin E2 was observed at the 12-h, 14-h, and 16-htime points (Fig. 9C, compare lanes 5 to 7 with lanes 12 to 14).Thus, the timing of cyclin E2 induction was not significantly ac-
FIG. 7. Inactivation of the Cutl1 locus affects the proliferation and the rate of entry in S phase of primary mouse embryonic fibroblasts. Mouseembryo fibroblasts were isolated from Cutl1 mutant mice and wild-type littermates. A. Cell cycle distribution of proliferating cells. Cells wereseeded at equal densities, grown for 48 h, pulsed with 100 �M BrdU for 1 h, and fixed in 3.7% paraformaldehyde. Cell cycle distribution wasdetermined by FACS using an Alexa 488-conjugated anti-BrdU antibody and propidium iodide. The histogram represents the average � standarddeviation of MEFs from three separate embryos for each genotype. B. Proliferation curves. MEFs from two different embryos for each genotypewere seeded at 1.4 � 104 cells/cm2 in triplicate and counted every day on a hemocytometer. Each value represents the average for two MEFs �standard error. C. Serum starvation at high density and replating. MEFs from three embryos for each genotype were pooled together and seeded(3 � 106 cells per 100-mm dish). Forty-eight hours later, the medium was replaced with DMEM–0.1% FBS, and cells were incubated for anadditional 72 h. MEFs were trypsinized and seeded at a density of 3.3 � 104 cells/cm2 and collected for FACS analysis with propidium iodide. D.MEFs from a different litter than for Fig. 7C were serum starved for 3 days, trypsinized, and plated at a lower density. One hour before fixation,MEFs were pulsed with 100 �M BrdU and analyzed by FACS. Values represent the average for two separate MEFs � standard deviation for eachgenotype.
VOL. 26, 2006 p110 CDP/Cux ACCELERATES CELL CYCLE PROGRESSION 2449
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 13
Jan
uary
202
2 by
119
.237
.105
.46.

celerated in NIH 3T3/p110 cells, but its expression was morerobust at these time points.
Two additional approaches were used to verify whether ex-pression of cyclins A2 and E2 was affected by the level of
CDP/Cux. First, we compared mRNA expression in early-pas-sage embryo fibroblasts from Cutl1z/z and wild-type mice.While expression of cyclin E1 was not affected, that of cyclinA2 was reduced slightly and that of cyclin E2 was markedly
FIG. 8. Proliferation defect caused by inactivation of the Cutl1 locus persists after immortalization and following transformation by H-RasV12. A.Inactivation of the Cutl1 locus causes a delay in S-phase entry both in immortalized 3T3 cells and in H-RasV12-transformed 3T3 cells. Cutl1�/� andCutl1z/z cells were immortalized following the 3T3 protocol and were then infected with a retrovirus expressing either activated H-RasV12 or an emptyvector. Cells were rendered quiescent by incubation in 0.1% FBS for 3 days, stimulated with 10% FBS, and prepared for FACS analysis. B. H-RasV12 infectionfails to stimulate formation of foci in Cutl1z/z 3T3 cells. Two 3T3 lines were independently immortalized from Cutl1z/z and Cutl1�/� MEFs (labeled Aand B) and were then infected with H-RasV12 or an empty vector. After 2 weeks in culture, foci were fixed and stained with Giemsa. The transformationefficiency is expressed as a percentage (no. of foci/no. of resistant colonies) for each condition. C. Western blot analysis with anti-Ras (top panel) oranti-CDP/Cux antibodies (labeled 1300) was done using 20 �g of total extracts from infected cells. Equal loading was confirmed by reblotting themembrane with antiactin antibody. D. Reintroduction of CDP/Cux in Cutl1z/z 3T3 cells partially rescues transformation by H-RasV12. Cutl1z/z 3T3 cellswere infected with combinations of vectors encoding H-RasV12, p200 CDP/Cux, p110 CDP/Cux, or their respective empty vectors. After successiveinfections with the viruses, cells were kept in culture for 2 weeks, fixed, and stained. E. Western blot analysis of Cutl1z/z 3T3 cells shown in panel D usinganti-Ras, anti-CDP (1300), or antiactin antibodies. F. Cutl1z/z 3T3/RasV12 cells are less tumorigenic in vivo. 10,000 infected cells were injectedsubcutaneously for each genotype, and tumor growth was monitored every 3 days. Shown is a photograph of representative tumors from each cell type.Each data point indicates the average size from four injection sites � standard deviation and is representative of two independent experiments.
2450 SANSREGRET ET AL. MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 13
Jan
uary
202
2 by
119
.237
.105
.46.

FIG. 9. p110 CDP/Cux stimulates cyclin A2 and cyclin E2 expression. A. Total RNA was extracted from NIH 3T3/vector and NIH 3T3/p110cells that were either unsynchronized or rendered quiescent by serum deprivation for 3 days. cDNA was prepared and served to measure theexpression of several regulators of the G1/S transition and putative CDP/Cux targets. B. Cyclin E2 protein and its associated kinase activity areelevated in NIH 3T3/p110 cells. Whole-cell lysates were prepared from unsynchronized NIH 3T3/vector and NIH 3T3/p100 cells and analyzed bySDS-PAGE and Western blotting using the indicated antibodies. Kinase assays were performed on histone H1 in the presence of [32P]ATP andfollowing immunoprecipitation of 2 mg of cell lysates with either cyclin E1 or E2 antibodies. Phosphorylated histone H1 was detected byautoradiography. C. Cyclin A2 and cyclin E2 protein expression are increased in NIH 3T3/p110 cells following exit from quiescence. NIH3T3/vector and NIH 3T3/p110 cells were serum starved for 3 days in 0.1% FBS and stimulated with 10% FBS. For each time point, total proteinextracts were prepared and subjected to SDS-PAGE and Western blot analysis with the indicated antibodies. D. Cyclin E2 mRNA expression isimpaired in Cutl1z/z MEFs. Total mRNA was extracted from MEFs at passage 2 for each genotype and analyzed by RT-PCR for cyclin E1 andE2 (E1, E2), cyclin A2 (A2), or glyceraldehyde-3-phosphate dehydrogenase (G). E. Cyclin E2 mRNA expression is stimulated following transientexpression of p110 CDP/Cux. NIH 3T3 and Hs578t cells were infected with a retroviral vector encoding p110 CDP/Cux. mRNA was isolated 48 hpostinfection and analyzed by RT-PCR. F. CDP/Cux binds to the cyclin A2 gene promoter in vivo. Chromatin immunoprecipitations (left panel)were performed with Hs578T cells using either the anti-861 CDP/Cux antibody (lane 3) or an IgG control (lane 2). The immunoprecipitated DNAwas used as a template in PCR using the indicated primers from the cyclin A2 gene promoter or from the glucose-6-phosphate dehydrogenasepromoter (G6), as indicated. As a control, the PCRs were performed in parallel using total chromatin (lane 1). Shown below is a map of the cyclinA2 gene promoter indicating the positions of primers used for PCR. Recombinant p110 CDP/Cux binds to the cyclin A2 gene promoter in vivo(right panel). Binding of p110 CDP/Cux to the cyclin A2 promoter in vivo was verified by chromatin affinity purification. Hs578T cells were infectedwith a retrovirus expressing a recombinant CDP/Cux protein that contains amino acids 612 to 1328 tagged at its C terminus with the calmodulinbinding domain and protein A. After 2 weeks under selection, a stable population was obtained by pooling more than 1,000 resistant colonies.Following cross-linking with formaldehyde, the chromatin was purified by tandem affinity purification (Tap tag) on IgG Sepharose beads andcalmodulin beads, as described in Materials and Methods. The purified DNA was used as a template in PCR using the same primers as for ChIP.
VOL. 26, 2006 p110 CDP/Cux ACCELERATES CELL CYCLE PROGRESSION 2451
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 13
Jan
uary
202
2 by
119
.237
.105
.46.

reduced (Fig. 9D, compare lanes 1 and 2). Second, continu-ously proliferating NIH 3T3 and Hs578T cells were infectedwith a retrovirus expressing p110 CDP/Cux or an empty cas-sette. While cyclin E1 expression was not affected by p110CDP/Cux, cyclin E2 was robustly induced in both cell lines(Fig. 9E, lanes 1 and 2 and lanes 3 and 4). Interestingly, cyclinA2 levels did not vary upon transient expression of p110 CDP/Cux, a result that is reminiscent of the situation in unsynchro-nized cells from the stable NIH 3T3/p110 population (Fig. 9Aand B). Together, these results indicate that p110 CDP/Cuxstimulates cyclin A2 expression in serum-starved cells and cy-clin E2 expression in proliferating cells.
Cyclin A2 is a direct target of CDP/Cux. Previous studiesimplicating cyclin A2 as a potential target of CDP/Cux werelimited to reporter assays (65). To verify whether CDP/Cux ispresent on the endogenous cyclin A2 promoter, we performedchromatin immunoprecipitation assays using cells from theHs578T breast tumor cell line, which is known to express highlevels of several CDP/Cux isoforms (17). We found that im-munoprecipitation with a anti-CDP antibody greatly enrichedfor a region of the cyclin A2 gene promoter (Fig. 9F, lane 3).This result indicates that a CDP/Cux protein resides on thecyclin A2 gene promoter; however, whether the p110 CDP/Cuxisoform is involved in the regulation of this gene could not bedetermined by this method, since our antibodies do not distin-guish between the full-length and short CDP/Cux isoforms. Toinvestigate whether p110 CDP/Cux could bind to the cyclin A2promoter, we performed ChAP using a population of cellsstably expressing a modified p110 CDP/Cux protein which wastagged at its C terminus with the calmodulin binding domainand protein A. Following cross-linking with formaldehyde, thechromatin was purified by tandem affinity purification (Taptag) on IgG Sepharose beads and calmodulin beads. PCRamplification showed that the cyclin A2 gene promoter wasgreatly enriched in the chromatin obtained by affinity purifica-tion (Fig. 9F, lanes 2 and 4). We conclude that p110 CDP/Cuxcan bind to the cyclin A2 gene promoter and that constitutiveexpression of p110 leads to the elevated expression of cyclin A2in certain situations. In contrast, in the case of the cyclin E2gene, we performed scanning ChIP using 11 primer pairs de-rived from the 5� flanking sequences, exon 1, intron 1, and 3�flanking sequences, but we did not obtain any evidence thatCDP/Cux can bind to this gene. This result suggests either thatcyclin E2 is not a direct target or that CDP/Cux binds toregulatory sequences that are located at a distance from thetranscription start site.
DISCUSSION
Previous studies demonstrated that the expression and ac-tivity of CDP/Cux are regulated in a cell cycle-dependent man-ner and provided circumstantial evidence that CDP/Cux couldplay a role in cell cycle progression. The full-length CDP/Cuxprotein was shown to be proteolytically processed at the G1/Stransition, and DNA binding by the processed isoform, p110,was found to peak in S phase as a result of dephosphorylationby the Cdc25A phosphatase (11, 16, 45). DNA binding de-creased later in the cell cycle following phosphorylation ofserine 1237 by cyclin A/Cdk1 (58). Moreover, a large body ofevidence demonstrated that the HiNF-D complex, of which
CDP/Cux is the primary DNA binding protein, is active in Sphase (22, 69, 71–73).
In the present study, we monitored cell cycle progression incells stably expressing p110 CDP/Cux. Cells were synchronizedeither in G0/G1 using serum starvation or at the G1/S transitionusing the double thymidine block. A third method, counter-flow centrifugal elutriation, was used to separate continuouslyproliferating cells and obtain a fraction of cells enriched inG2/M. In each of the three approaches, cells traversed the G1
phase faster than control cells (Fig. 3 to 6). Yet progressionthrough the S and G2 phases did not seem to be impaired.These results indicate that the accelerating effect of p110 CDP/Cux on the cell cycle is not limited to the first cycle followingexit from quiescence or release from a cell cycle block but cantake place in continuously proliferating cells. Moreover, weobserved that cellular proliferation was also accelerated in cellsexpressing p110 CDP/Cux. This is what would be expected ifindeed the G1 phase was shortened while the other phases ofthe cell cycle were not affected.
On the other hand, inactivation of the Cutl1 locus causedMEFs to be slightly delayed in their progression from quies-cence to S phase and to proliferate more slowly in the presenceof mitogens. Moreover, the proliferation defect resulting fromthe inactivation of Cutl1 persisted following immortalizationby the 3T3 protocol and transformation with H-RasV12. Theseresults are in contrast with those of two previous studies thatdid not report a proliferation deficit in MEFs obtained fromCutl1 knockout mice (14, 39). We have considered the possi-bility that different results could be obtained depending on theserum used in the medium or the type of substratum on whichcells are cultivated. Indeed, we observed variations in cellularproliferation between different batches of serum, but in allcases proliferation was slower in the Cutl1z/z cells. We havenoticed, however, that to observe reproducibly small differ-ences in proliferation it was crucial to seed MEFs at exactly thesame densities in the first and second passages prior to theactual experiment. Our interpretation of this observation isthat the density at which MEF cells are cultivated has animpact on their proliferation capacity upon reseeding.
Constitutive (or transient) p110 CDP/Cux expression wasnot sufficient to induce cell cycle entry of quiescent cells in theabsence of growth factors (Fig. 3 and 4; also data not shown).This inability to induce S-phase entry in the absence of appro-priate growth conditions is consistent with our findings thatp110 CDP/Cux did not induce apoptosis in cells that weremaintained in a low serum concentration (data not shown).Thus, our results indicate that p110 CDP/Cux cannot on itsown drive cells into S phase, but it can accelerate cell cycleprogression without apparent negative consequences for thecells. These properties clearly distinguish p110 CDP/Cux fromsome members of the E2F family and suggest that overexpres-sion of CDP/Cux in various tumors and in particular of its shortisoforms must provide cancer cells with a growth advantage(17, 46). Indeed, transgenic mice expressing short CDP/Cuxisoforms display increased susceptibility to various types ofcancers (C. Cadieux, S. Fournier, A. C. Peterson, B. J. Bedell,and A. Nepveu, submitted).
Constitutive expression of p110 CDP/Cux in NIH 3T3 cellswas found to alter the regulation of two cyclins but in differentways. While cyclin A2 was overexpressed in serum-starved cells
2452 SANSREGRET ET AL. MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 13
Jan
uary
202
2 by
119
.237
.105
.46.

only, cyclin E2 was overexpressed at those times when it isnormally induced following serum restimulation and also incontinuously proliferating cells. Increased expression of bothcyclin A and cyclin E was reported to be sufficient to accelerateS-phase entry (1, 37, 50, 55–57). The maintenance of cyclin A2expression in the absence of growth factors probably did notcontribute to the shortening of the G1 phase upon reentry intothe cell cycle but could provide an explanation for the fact thata higher percentage of NIH 3T3/p110 cells remained in Sphase. On the other hand, the increased cyclin E2 expressionand associated kinase activity in continuously proliferatingcells and its increased induction following serum stimulationmost likely contributed to the acceleration in S-phase entry.However, it is unlikely that the proliferation defect of Cutl1Z/Z
MEFs could be explained by a reduction in cyclin E2 expres-sion, considering that normal MEF and 3T3 cells express verylow levels of cyclin E2 and the knockout of cyclin E2 had noeffect on the proliferation of MEFs (Fig. 9B and C) (23, 51).Future studies will attempt to identify the full repertoire ofCDP/Cux targets that can impact on cellular proliferation andcell cycle progression.
In contrast to cyclin E2, cyclin E1 expression was not af-fected by p110 CDP/Cux. Cyclins E1 and E2 display similarbiochemical activities and expression patterns, in terms of bothtissue specificity and timing of expression during cell cycleprogression (15, 18, 31, 53). Yet there is a precedent for thedistinct regulation of either gene, since the papillomavirusoncoprotein E6 was found to stimulate cyclin E2 expressionbut to repress cyclin E1 (74). Our finding that p110 CDP/Cuxspecifically induces cyclin E2 expression further supports thenotion that cyclins E1 and E2 are regulated by different com-binations of transcription factors.
The fact that ectopic expression of p110 CDP/Cux acceler-ates S-phase entry indicates that p110 itself must be one of thelimiting factors in the progression from G1 to S. Yet geneinactivation of the Cutl1 locus demonstrated that CDP/Cux isnot essential (14, 39, 60, 66). How can we reconcile thesebiological properties in terms of mechanism of action? Wepropose that p110 CDP/Cux plays a role in the establishmentof at least part of the S-phase-specific transcription programand that CDP/Cux does not act alone in the regulation ofS-phase-specific genes but rather functions in combinatorialfashion with other transcription factors. This mode of actionwould be entirely consistent with known features of theHiNF-D complex, which resides on the promoter of histonegenes during S phase (4, 5, 30, 68–72). Moreover, this mech-anism of action would explain why cells reach S phase faster inthe presence of p110 CDP/Cux but are still able to proliferatein its absence, albeit more slowly. In cells where p110 CDP/Cuxis expressed constitutively, the promoters of S-phase-specificgenes would be occupied more rapidly by cooperating activa-tors at the end of the G1 phase. In contrast, in the absence ofp110 CDP/Cux, the same promoters would be occupied later asthe concentration of the other activators reaches a thresholdconcentration that enables them to interact with their low-affinity binding sites.
The presence of four DNA binding domains within CDP/Cux makes it a complex protein that has the potential to carryseveral independent functions. The two CDP/Cux DNA bind-ing activities so far described appear to serve distinct, yet
compatible, functions in dividing cells. In precursor cells ofvarious lineages, the CCAAT displacement activity reportedlyserves to downmodulate genes which later become expressedin terminally differentiated cells (34, 52, 61–63). This activityinvolves the full-length p200 isoform. The partial rescue andthe additive effects of p200 and p110 observed in the focusformation assay with H-rasV12 suggest that p200 also plays anactive role in cell proliferation. An additional function of theprotein is activated following a proteolytic processing eventthat generates the p110 isoform (16, 45). This protein makes astable interaction with DNA and is able to stimulate the ex-pression of several genes whose products are required forDNA replication (45, 65). At a cellular level, p110 acceleratesentry into S phase and stimulates cell proliferation. Thus, over-all CDP/Cux appears to fulfill several functions in proliferatingcells: the p200 isoform prevents terminal differentiation andstimulates proliferation in a manner that remains to be de-fined, whereas the p110 isoform accelerates S-phase entry.Moreover, a recent study demonstrated that CDP/Cux is re-quired for cell motility and invasion (42). In a survey of humanbreast cancers, CDP/Cux expression was found to be signifi-cantly increased in high-grade carcinomas and was inverselycorrelated with survival. Microarray analysis also revealed thatCUTL1 was one of the most up-regulated genes in malignantplasma cells from multiple myelomas (12). The multiple rolesof CDP/Cux in cell proliferation, motility, and invasion suggestthat it may be involved in both tumor growth and tumor pro-gression.
ACKNOWLEDGMENTS
We thank Meinrad Busslinger for the kind gift of the Cutl1�/z miceand Scott Lowe for the pBabe-H-RasV12 plasmid.
A.N. is the recipient of a scholarship from the Fonds de la Recher-che en Sante du Quebec. L.S. is the recipient of studentships from theRoyal Victoria Hospital Research Institute. Exchanges between theBertoglio and Nepveu laboratories were made possible through a grantfrom FRSQ and INSERM. This research was supported by grant014288 from the National Cancer Institute of Canada to A.N.
REFERENCES
1. Alevizopoulos, K., J. Vlach, S. Hennecke, and B. Amati. 1997. Cyclin E andc-Myc promote cell proliferation in the presence of p16INK4a and of hypo-phosphorylated retinoblastoma family proteins. EMBO J. 16:5322–5333.
2. Andres, V., M. D. Chiara, and V. Mahdavi. 1994. A new bipartite DNA-binding domain: cooperative interaction between the cut repeat and homeodomain of the cut homeo proteins. Genes Dev. 8:245–257.
3. Aufiero, B., E. J. Neufeld, and S. H. Orkin. 1994. Sequence-specific DNAbinding of individual Cut repeats of the human CCAAT displacement/Cuthomeodomain protein. Proc. Natl. Acad. Sci. USA 91:7757–7761.
4. Aziz, F., A. J. van Wijnen, J. L. Stein, and G. S. Stein. 1998. HiNF-D(CDP-cut/CDC2/cyclin A/pRB-complex) influences the timing of IRF-2-de-pendent cell cycle activation of human histone H4 gene transcription at theG1/S phase transition. J. Cell Physiol. 177:453–464.
5. Aziz, F., A. J. Vanwijnen, P. S. Vaughan, S. J. Wu, A. R. Shakoori, J. B. Lian,K. J. Soprano, J. L. Stein, and G. S. Stein. 1998. The integrated activities ofIrf-2 (Hinf-M), Cdp/Cut (Hinf-D) and H4tf-2 (Hinf-P) regulate transcriptionof a cell cycle controlled human histone H4 gene—mechanistic differencesbetween distinct H4 genes. Mol. Biol. Rep. 25:1–12.
6. Blanc, R. 1942. The production of wing scalloping in Drosophila melano-gaster. Univ. Calif. Publ. Zool. 49:1–31.
7. Bodmer, R., S. Barbel, S. Shepherd, J. W. Jack, L. Y. Jan, and Y. N. Jan.1987. Transformation of sensory organs by mutations of the cut locus of D.melanogaster. Cell 51:293–307.
8. Braun, W. 1942. The effect of puncture on the developing wing of severalmutants of Drosophila melanogaster. J. Exp. Zool. 84:325–350.
9. Cai, H. N., and M. Levine. 1997. The gypsy insulator can function as apromoter-specific silencer in the Drosophila embryo. EMBO J. 16:1732–1741.
10. Chattopadhyay, S., C. E. Whitehurst, and J. Chen. 1998. A nuclear matrix
VOL. 26, 2006 p110 CDP/Cux ACCELERATES CELL CYCLE PROGRESSION 2453
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 13
Jan
uary
202
2 by
119
.237
.105
.46.

attachment region upstream of the T cell receptor beta gene enhancer bindsCux/CDP and SATB1 and modulates enhancer-dependent reporter geneexpression but not endogenous gene expression. J. Biol. Chem. 273:29838–29846.
11. Coqueret, O., G. Berube, and A. Nepveu. 1998. The mammalian Cut homeo-domain protein functions as a cell-cycle dependent transcriptional repressorwhich downmodulates p21WAF1/CIP1/SDI1 in S phase. EMBO J. 17:4680–4694.
12. De Vos, J., T. Thykjaer, K. Tarte, M. Ensslen, P. Raynaud, G. Requirand, F.Pellet, V. Pantesco, T. Reme, M. Jourdan, J. F. Rossi, T. Orntoft, and B.Klein. 2002. Comparison of gene expression profiling between malignant andnormal plasma cells with oligonucleotide arrays. Oncogene 21:6848–6857.
13. Dorsett, D. 1993. Distance-independent inactivation of an enhancer by thesuppressor of Hairy-wing DNA-binding protein of Drosophila. Genetics134:1135–1144.
14. Ellis, T., L. Gambardella, M. Horcher, S. Tschanz, J. Capol, P. Bertram, W.Jochum, Y. Barrandon, and M. Busslinger. 2001. The transcriptional repres-sor CDP (Cutl1) is essential for epithelial cell differentiation of the lung andthe hair follicle. Genes Dev. 15:2307–2319.
15. Geng, Y., Q. Yu, W. Whoriskey, F. Dick, K. Y. Tsai, H. L. Ford, D. K. Biswas,A. B. Pardee, B. Amati, T. Jacks, A. Richardson, N. Dyson, and P. Sicinski.2001. Expression of cyclins E1 and E2 during mouse development and inneoplasia. Proc. Natl. Acad. Sci. USA 98:13138–13143.
16. Goulet, B., A. Baruch, N. S. Moon, M. Poirier, L. L. Sansregret, A. Erickson,M. Bogyo, and A. Nepveu. 2004. A cathepsin L isoform that is devoid of asignal peptide localizes to the nucleus in S phase and processes the CDP/Cuxtranscription factor. Mol. Cell 14:207–219.
17. Goulet, B., P. Watson, M. Poirier, L. Leduy, G. Berube, S. Meterissian, P.Jolicoeur, and A. Nepveu. 2002. Characterization of a tissue-specific CDP/Cux isoform, p75, activated in breast tumor cells. Cancer Res. 62:6625–6633.
18. Gudas, J. M., M. Payton, S. Thukral, E. Chen, M. Bass, M. O. Robinson, andS. Coats. 1999. Cyclin E2, a novel G1 cyclin that binds Cdk2 and is aberrantlyexpressed in human cancers. Mol. Cell. Biol. 19:612–622.
19. Harada, R., G. Berube, O. J. Tamplin, C. Denis-Larose, and A. Nepveu.1995. DNA-binding specificity of the cut repeats from the human cut-likeprotein. Mol. Cell. Biol. 15:129–140.
20. Harada, R., D. Dufort, C. Denis-Larose, and A. Nepveu. 1994. Conserved cutrepeats in the human cut homeodomain protein function as DNA bindingdomains. J. Biol. Chem. 269:2062–2067.
21. Hertweck, H. 1931. Anatomie und variabilitaet des nerven-systems und dersinneorgane von Drosophila melanogaster. J. Exp. Zool. 139:559–663.
22. Holthuis, J., T. A. Owen, A. J. van Wijnen, K. L. Wright, A. Ramsey-Ewing,M. B. Kennedy, R. Carter, S. C. Cosenza, K. J. Soprano, J. B. Lian, J. L.Stein, and G. S. Stein. 1990. Tumor cells exhibit deregulation of the cell cyclehistone gene promoter factor HiNF-D. Science 247:1454–1457.
23. Ishii, H., K. Mimori, Y. Yoshikawa, M. Mori, Y. Furukawa, and A. Vecchione.2005. Differential roles of E-type cyclins during transformation of murineE2F-1-deficient cells. DNA Cell Biol. 24:173–179.
24. Jack, J., and Y. DeLotto. 1992. Effect of wing scalloping mutations on cutexpression and sense organ differentiation in the Drosophila wing margin.Genetics 131:353–363.
25. Jack, J., and Y. DeLotto. 1995. Structure and regulation of a complex locus:the cut gene of Drosophila. Genetics 139:1689–1700.
26. Jack, J., D. Dorsett, Y. Delotto, and S. Liu. 1991. Expression of the cut locusin the Drosophila wing margin is required for cell type specification and isregulated by a distant enhancer. Development 113:735–747.
27. Jack, J. W. 1985. Molecular organization of the cut locus of Drosophilamelanogaster. Cell 42:869–876.
28. Johnson, T. K., and B. H. Judd. 1979. Analysis of the cut locus of Drosophilamelanogaster. Genetics 92:485–502.
29. Kauffman, M. G., S. J. Noga, T. J. Kelly, and A. D. Donnenberg. 1990.Isolation of cell cycle fractions by counterflow centrifugal elutriation. Anal.Biochem. 191:41–46.
30. Last, T. J., A. J. van Wijnen, M. C. de Ridder, G. S. Stein, and J. L. Stein.1999. The homeodomain transcription factor CDP/cut interacts with the cellcycle regulatory element of histone H4 genes packaged into nucleosomes.Mol. Biol. Rep. 26:185–194.
31. Lauper, N., A. R. Beck, S. Cariou, L. Richman, K. Hofmann, W. Reith, J. M.Slingerland, and B. Amati. 1998. Cyclin E2: a novel CDK2 partner in the lateG1 and S phases of the mammalian cell cycle. Oncogene 17:2637–2643.
32. Ledford, A. W., J. G. Brantley, G. Kemeny, T. L. Foreman, S. E. Quaggin, P.Igarashi, S. M. Oberhaus, M. Rodova, J. P. Calvet, and G. B. VandenHeuvel. 2002. Deregulated expression of the homeobox gene cux-1 in trans-genic mice results in downregulation of p27(kip1) expression during nephro-genesis, glomerular abnormalities, and multiorgan hyperplasia. Dev. Biol.245:157–171.
33. Lee, K. A. W., A. Bindereif, and M. R. Green. 1988. A small-scale procedurefor preparation of nuclear extracts that support efficient transcription andpre-mRNA splicing. Gene Anal. Tech. 5:22–31.
34. Lievens, P. M. J., J. J. Donady, C. Tufarelli, and E. J. Neufeld. 1995.Repressor activity of CCAAT displacement protein in HL-60 myeloid leu-kemia cells. J. Biol. Chem. 270:12745–12750.
35. Liu, S., and J. Jack. 1992. Regulatory interactions and role in cell typespecification of the Malpighian tubules by the cut, Kruppel, and caudal genesof Drosophila. Dev. Biol. 150:133–143.
36. Liu, S., E. McLeod, and J. Jack. 1991. Four distinct regulatory regions of thecut locus and their effect on cell type specification in Drosophila. Genetics127:151–159.
37. Lukas, J., T. Herzinger, K. Hansen, M. C. Moroni, D. Resnitzky, K. Helin,S. I. Reed, and J. Bartek. 1997. Cyclin E-induced S phase without activationof the pRb/E2F pathway. Genes Dev. 11:1479–1492.
38. Luo, W., and D. G. Skalnik. 1996. CCAAT displacement protein competeswith multiple transcriptional activators for binding to four sites in the prox-imal gp91(phox) promoter. J. Biol. Chem. 271:18203–18210.
39. Luong, M. X., C. M. van der Meijden, D. Xing, R. Hesselton, E. S. Monuki,S. N. Jones, J. B. Lian, J. L. Stein, G. S. Stein, E. J. Neufeld, and A. J. vanWijnen. 2002. Genetic ablation of the CDP/Cux protein C terminus results inhair cycle defects and reduced male fertility. Mol. Cell. Biol. 22:1424–1437.
40. Majumdar, A., R. Nagaraj, and U. Banerjee. 1997. Strawberry notch encodesa conserved nuclear protein that functions downstream of Notch and regu-lates gene expression along the developing wing margin of Drosophila.Genes Dev. 11:1341–1353.
41. Micchelli, C. A., E. J. Rulifson, and S. S. Blair. 1997. The function andregulation of cut expression on the wing margin of Drosophila: Notch,Wingless and a dominant negative role for Delta and Serrate. Development124:1485–1495.
42. Michl, P., A. R. Ramjaun, O. E. Pardo, P. H. Warne, M. Wagner, R. Poulsom,C. D’Arrigo, K. Ryder, A. Menke, T. Gress, and J. Downward. 2005. CUTL1is a target of TGF(beta) signaling that enhances cancer cell motility andinvasiveness. Cancer Cell 7:521–532.
43. Modolell, J., W. Bender, and M. Meselson. 1983. Drosophila melanogastermutations suppressible by the suppressor of Hairy-wing are insertions of a7.3-kilobase mobile element. Proc. Natl. Acad. Sci. USA 80:1678–1682.
44. Moon, N. S., G. Berube, and A. Nepveu. 2000. CCAAT displacement activityinvolves Cut repeats 1 and 2, not the Cut homeodomain. J. Biol. Chem.275:31325–31334.
45. Moon, N. S., P. Premdas, M. Truscott, L. Leduy, G. Berube, and A. Nepveu.2001. S phase-specific proteolytic cleavage is required to activate stable DNAbinding by the CDP/Cut homeodomain protein. Mol. Cell. Biol. 21:6332–6345.
46. Moon, N. S., W. Rong Zeng, P. Premdas, M. Santaguida, G. Berube, and A.Nepveu. 2002. Expression of N-terminally truncated isoforms of CDP/CUXis increased in human uterine leiomyomas. Int. J. Cancer 100:429–432.
47. Nepveu, A. 2001. Role of the multifunctional CDP/Cut/Cux homeodomaintranscription factor in regulating differentiation, cell growth and develop-ment. Gene 270:1–15.
48. Neufeld, E. J., D. G. Skalnik, P. M. Lievens, and S. H. Orkin. 1992. HumanCCAAT displacement protein is homologous to the Drosophila homeopro-tein, cut. Nat. Genet. 1:50–55.
49. Neumann, C. J., and S. M. Cohen. 1996. A hierarchy of cross-regulationinvolving Notch, wingless, vestigial and cut organizes the dorsal/ventral axisof the Drosophila wing. Development 122:3477–3485.
50. Ohtsubo, M., A. M. Theodoras, J. Schumacher, J. M. Roberts, and M.Pagano. 1995. Human cyclin E, a nuclear protein essential for the G1-to-Sphase transition. Mol. Cell. Biol. 15:2612–2624.
51. Parisi, T., A. R. Beck, N. Rougier, T. McNeil, L. Lucian, Z. Werb, and B.Amati. 2003. Cyclins E1 and E2 are required for endoreplication in placentaltrophoblast giant cells. EMBO J. 22:4794–4803.
52. Pattison, S., D. G. Skalnik, and A. Roman. 1997. CCAAT displacementprotein, a regulator of differentiation-specific gene expression, binds a neg-ative regulatory element within the 5� end of the human papillomavirus type6 long control region. J. Virol. 71:2013–2022.
53. Payton, M., S. Scully, G. Chung, and S. Coats. 2002. Deregulation of cyclinE2 expression and associated kinase activity in primary breast tumors. On-cogene 21:8529–8534.
54. Quaggin, S. E., G. B. Vandenheuvel, K. Golden, R. Bodmer, and P. Igarashi.1996. Primary structure, neural-specific expression, and chromosomal local-ization of Cux-2, a second murine homeobox gene related to drosophila cut.J. Biol. Chem. 271:22624–22634.
55. Resnitzky, D., L. Hengst, and S. I. Reed. 1995. Cyclin A-associated kinaseactivity is rate limiting for entrance into S phase and is negatively regulatedin G1 by p27Kip1. Mol. Cell. Biol. 15:4347–4352.
56. Resnitzky, D., and S. I. Reed. 1995. Different roles for cyclins D1 and E inregulation of the G1-to-S transition. Mol. Cell. Biol. 15:3463–3469.
56a.Rigant, G., a. Shevchenko, B. Rutz, M. Wilm, M. Mann, and B. Seraphin.1999. A generic protein purification method for protein complex character-ization and proteome exploration. Nat. Biotechnol. 17:1030–1032.
57. Rosenberg, A. R., F. Zindy, F. Le Deist, H. Mouly, P. Metezeau, C. Brechot,and E. Lamas. 1995. Overexpression of human cyclin A advances entry intoS phase. Oncogene 10:1501–1509.
58. Santaguida, M., Q. Ding, G. Berube, M. Truscott, P. Whyte, and A. Nepveu.2001. Phosphorylation of the CCAAT displacement protein (CDP)/Cuxtranscription factor by cyclin A-Cdk1 modulates its DNA binding activity inG(2). J. Biol. Chem. 276:45780–45790.
2454 SANSREGRET ET AL. MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 13
Jan
uary
202
2 by
119
.237
.105
.46.

59. Santaguida, M. T., and A. Nepveu. 2005. Differential regulation of CDP/Cuxp110 by cyclin A/Cdk2 and cyclin A/Cdk1. J. Biol. Chem. 280:32712–32721.
60. Sinclair, A. M., J. A. Lee, A. Goldstein, D. Xing, S. Liu, R. Ju, P. W. Tucker,E. J. Neufeld, and R. H. Scheuermann. 2001. Lymphoid apoptosis andmyeloid hyperplasia in CCAAT displacement protein mutant mice. Blood98:3658–3667.
61. Skalnik, D. G., E. C. Strauss, and S. H. Orkin. 1991. CCAAT displacementprotein as a repressor of the myelomonocytic-specific gp91-phox gene pro-moter. J. Biol. Chem. 266:16736–16744.
62. Stunkel, W., Z. Huang, S. H. Tan, M. J. O’Connor, and H. U. Bernard. 2000.Nuclear matrix attachment regions of human papillomavirus type 16 repressor activate the E6 promoter, depending on the physical state of the viralDNA. J. Virol. 74:2489–2501.
63. Superti-Furga, G., A. Barberis, E. Schreiber, and M. Busslinger. 1989. Theprotein CDP, but not CP1, footprints on the CCAAT region of the gamma-globin gene in unfractionated B-cell extracts. Biochim. Biophys. Acta 1007:237–242.
64. Todaro, G. J., and H. Green. 1963. Quantitative studies of the growth ofmouse embryo cells in culture and their development into established lines.J. Cell Biol. 17:299–313.
65. Truscott, M., L. Raynal, P. Premdas, B. Goulet, L. Leduy, G. Berube, and A.Nepveu. 2003. CDP/Cux stimulates transcription from the DNA polymerasealpha gene promoter. Mol. Cell. Biol. 23:3013–3028.
66. Tufarelli, C., Y. Fujiwara, D. C. Zappulla, and E. J. Neufeld. 1998. Hairdefects and pup loss in mice with targeted deletion of the first cut repeatdomain of the Cux/CDP homeoprotein gene. Dev. Biol. 200:69–81.
67. Valarche, I., J. P. Tissier-Seta, M. R. Hirsch, S. Martinez, C. Goridis, andJ. F. Brunet. 1993. The mouse homeodomain protein Phox2 regulates Ncampromoter activity in concert with Cux/CDP and is a putative determinant ofneurotransmitter phenotype. Development 119:881–896.
68. van den Ent, F. M., A. J. van Wijnen, J. B. Lian, J. L. Stein, and G. S. Stein.
1994. Cell cycle controlled histone H1, H3, and H4 genes share unusualarrangements of recognition motifs for HiNF-D supporting a coordinatepromoter binding mechanism. J. Cell Physiol. 159:515–530.
69. van Wijnen, A. J., T. K. Choi, T. A. Owen, K. L. Wright, J. B. Lian, R.Jaenisch, J. L. Stein, and G. S. Stein. 1991. Involvement of the cell cycle-regulated nuclear factor HiNF-D in cell growth control of a human H4histone gene during hepatic development in transgenic mice. Proc. Natl.Acad. Sci. USA 88:2573–2577.
70. van Wijnen, A. J., C. Cooper, P. Odgren, F. Aziz, A. De Luca, R. A. Shakoori,A. Giordano, P. J. Quesenberry, J. B. Lian, G. S. Stein, and J. L. Stein. 1997.Cell cycle-dependent modifications in activities of pRb-related tumor sup-pressors and proliferation-specific CDP/cut homeodomain factors in murinehematopoietic progenitor cells. J. Cell. Biochem. 66:512–523.
71. van Wijnen, A. J., M. F. van Gurp, M. C. de Ridder, C. Tufarelli, T. J. Last,M. Birnbaum, P. S. Vaughan, A. Giordano, W. Krek, E. J. Neufeld, J. L.Stein, and G. S. Stein. 1996. CDP/cut is the DNA-binding subunit of histonegene transcription factor HiNF-D: a mechanism for gene regulation at theG1/S phase cell cycle transition point independent of transcription factorE2F. Proc. Natl. Acad. Sci. USA 93:11516–11521.
72. van Wijnen, A. J., K. L. Wright, J. B. Lian, J. L. Stein, and G. S. Stein. 1989.Human H4 histone gene transcription requires the proliferation-specificnuclear factor HiNF-D. Auxiliary roles for HiNF-C (Sp1-like) and HiNF-A(high mobility group-like). J. Biol. Chem. 264:15034–15042.
73. Wright, K. L., R. T. Dell’Orco, A. J. van Wijnen, J. L. Stein, and G. S. Stein.1992. Multiple mechanisms regulate the proliferation-specific histone genetranscription factor HiNF-D in normal human diploid fibroblasts. Biochem-istry 31:2812–2818.
74. Zariwala, M., J. Liu, and Y. Xiong. 1998. Cyclin E2, a novel human G1 cyclinand activating partner of CDK2 and CDK3, is induced by viral oncoproteins.Oncogene 17:2787–2798.
VOL. 26, 2006 p110 CDP/Cux ACCELERATES CELL CYCLE PROGRESSION 2455
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 13
Jan
uary
202
2 by
119
.237
.105
.46.