the of no. inc. in u. s.a. isozymes of bovine intestinal ... · isozymes of bovine intestinal...
TRANSCRIPT

THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1985 by The American Society of Biological Chemists, Inc. Vol. 260, No. 20, Issue of September 15, pp. 11191-11193,1985
Printed in U. S.A.
Isozymes of Bovine Intestinal Alkaline Phosphatase”
(Received for publication, December 31, 1984)
Marc BesmanS and Joseph E. Coleman From the Department of Molecular Biophysics and Biochemistry, Yale University, New Haven, Connecticut 06510
Alkaline phosphatases from calf and bovine small intestines have been isolated in homogeneous form from both mucosa and luminal contents. The detergent- solubilized calf enzyme resolves into two peaks of ac- tivity, C-1 and C-2, on chromatofocusing. Only one of these activity peaks is present in the enzyme from the adult animal. Amino acid compositions, N-terminal se- quences, and tryptic peptide maps show that C-1 and C-2 are isozymes of differing primary structure and that the adult form of the enzyme is identical to C-2. The developmentally controlled expression of the two isozymes reported here suggests a molecular basis for the previous indications that functional changes in intestinal alkaline phosphatase occur with tissue mat- uration. The sugar composition of the carbohydrate chains of these isozymes has been determined and en- zymatic deglycosylation with endo-8-N-acetylglucos- aminidase-F indicates two N-linked and one or more 0-linked glycoconjugates/monomer.
Alkaline phosphatase is present in large amounts in the liver, intestine, placenta, bone, and kidney of mammals (1). Despite its characteristic histochemical association with cell membranes in these tissues, its detailed function is unclear, although mutations demonstrate its essential role in such processes as bone calcification (1). The prototype alkaline phosphatase is that from Escherichia coli where the enzyme is a dimeric Zn(I1) metalloprotein of identical subunits, con- taining three closely spaced metal-binding sites at each active center, one of which is a typical Zn(I1)-metalloenzyme site, while the other two can be occupied by Zn(I1) or Mg(1I) (2). The crystal structure of the dimer, M, = 94,000, has recently been determined (3).
Studies up to the present time on mammalian alkaline phosphatases from several species and tissues show these enzymes are apparently also dimers, but are glycosylated and have larger subunits, dimer M , -120,000 (1). The mammalian enzymes also contain Zn(II), but detailed data on the primary structure, number of metal-binding sites, and possible do- mains corresponding to the smaller bacterial subunit are not available. Earlier studies of the intestinal enzyme have for the most part relied on cell extracts or commercial prepara- tions and assumed a homogeneous molecule (1,4). Our initial studies presented here on alkaline phosphatase prepared from fresh calves’ and cows’ small intestines show that the calf
* This work was supported by National Institutes of Health Grant AM 32067. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
$ Graduate trainee under Training Grant GM 5-07223 from the National Institutes of Health. Submitted this work in partial fulfill- ment for the requirements of the Ph.D. dissertation at the Graduate School of Yale University.
enzyme can be resolved into two structural isozymes, one of which is not expressed in the adult animal. The three proteins have been characterized by N-terminal sequences to residue 23, complete tryptic peptide maps, sugar content and linkages ( N uersw 0) of the carbohydrates, and metal ion stoichiom- etries.
MATERIALS AND METHODS’
RESULTS AND DISCUSSION
Separation of Alkaline Phosphatase Isozymes-Elution (ac- tivity) profiles for the chromatofocusing columns run on the calf and adult bovine enzymes are shown in Fig. 1. The Triton X-100 present prevented the determination of an Azso profile. For this and all subsequent columns, protein profiles of the fractions were followed by SDS-PAGE.’ While the activity peaks clearly differentiate two species of alkaline phosphatase for the calf and apparently only one for the adult bovine, SDS-PAGE at this stage shows several contaminating pro- teins to be present. If the activity-containing fractions from the columns in Fig. 1 are pooled and chromatographed on the phosphonate affinity column, the peaks labeled C-1 and C-2 give rise to proteins with different gel mobilities (Fig. 2 A ) which correspond to the different tryptic peptide maps and N-terminal sequences to be shown below. On the other hand, the adult bovine enzyme gives rise to a single electrophoretic species.
The amino acid compositions of C-1, C-2, and B-2 are all rather similar, and while significant deviations occur between them in numbers of several of the amino acids, the precision is not sufficient to definitively prove the existence of isoen- zymes (Table I). In agreement with earlier literature, these enzymes have a high content of Ala, a characteristic shared with the bacterial enzyme (2). In contrast, we have not found the abnormally high Arg content reported in the most com- plete previous study of the calf enzyme (4). The latter study was on commercially supplicd enzyme and was apparently a mixture of the two isozymes.
The homogeneous proteins from our preparations require the presence of 0.1% deoxycholate in order to remain in solution. We have removed the detergent on several occasions (e.g., in preparation of the apoenzyme), a procedure which
Portions of this paper (including “Materials and Methods,” Ta- bles I and 11, and Figs. l and 3) are presented in miniprint at the end of this paper. Miniprint is easily read with the aid of a standard magnifying glass. Full size photocopies are available from the Journal of Biological Chemistry, 9650 Rockville Pike, Bethesda, MD 20814. Request Document No. 84M-3936, cite the authors, and include a check or money order for $4.80 per set of photocopies. Full size photocopies are also included in the microfilm edition of the Journal that is available from Waverly Press.
* The abbreviations used are: SDS-PAGE, sodium dodecyl sulfate- polyacrylamide gel electrophoresis; HPLC, high performance liquid chromatography; TPCK, ~-1-tosylamido-2-phenylethyl chloromethyl ketone; endo-F, endo-(3-N-acetylglucosaminidase-F.
11 190

Intestinal Alkaline Phosphataw 11 191
A. M W
116 97 = 66 - 47 -
8. L- L
nI-7- c-1 B -2
FIG. 2. A , SIN-PACE of the two calf isozvmes, commercial calf enzvme. and adult hovine enzyme is shown. /,one A, C-1; lonr H , C- 2; lonc,s (’ and I ) , calf intestinal alkaline phosphatase (Roehringer Mannheim. immunoassay grade); lane E . H-2. Molecular weight markers are in the l ~ f t column as indicated. Note the heterogeneity in two different samples of commercial enzyme. The presence of structural isozvmes and varying degrees of glycosylation (see H ) can hoth give rise to multiple hands on gels. This is currently under investigation for the commercial enzyme. H , Coomassie-stained SDS- PAGE o f C-1. C-2, and H-2 before ( le f t ) and after enzymatic degly- cosylat ion with endo-ij-N-flcetvlglucosaminidase-F (rifht ). The mo- lecular weight standards as in A are in the rniddlp /one. All hands were Coomassie and Schiff positive. The most rapidly migrating hand after deglycosvlation was used as the present best molecular weight estimate for use in Tahle 1. Since this hand still contains O-linked carhohvdrate. there may vet he some anomalous migration as sug- gested hv the different mohilities of C-2 and H-2, even though their tryptic peptide maps and sequence data indicate identical primary st rrlct ures.
always results in aggregation and precipitation. Readdition of detergent resoluhilizes the enzyme which remains active.
Over 50% of the enzyme is found in the luminal contents. This does not reflect proteolysis since alkaline phosphatase from lumen or mucosa could not be distinguished by SDS- or nondenaturing-PAGE. The high content in the lumen may result from sloughing of cells after death or possibly be due to the vesiculation ofthe brush-border memhrane, a recently descrihed “normal” event (14). While we initially added phen- ylmethanesulfonyl fluoride and p-chloromercurihenzoate to homogenates, the presence or absence of protease inhihitors does not affect the final product.
Carbohydrate Analysis-An analysis of the sugars present shows galactose, mannose, N-acetylglucosamine, and lesser
amounts of N-acetvlgalactosamine, sialic acid. and fucose (Table 11). The presence of glucose, a rare constituent o f glycoproteins, is difficult t o interpret as i t is a frequent and tenacious contaminant. C-1. C-2, and H-2 have similar sugar types, but C-2 appears to contain significantly more c a r h - hydrate than the adult H-2.
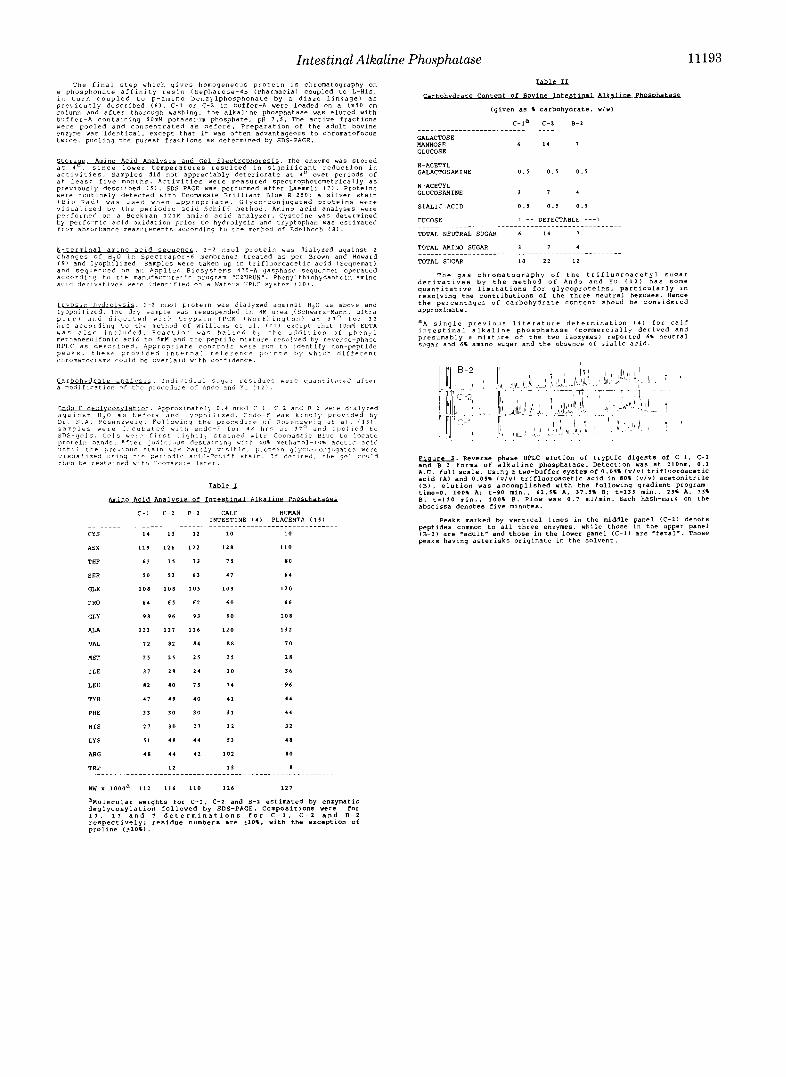
High I’wformance Liquid C‘hromntogrnph?, Annlysis Of T p p t i c Peptides-High performance liquid chromatography chromatograms of the tryptic digests of C - 1 and (“2 dernon- strate that approximatelvone-halfofthe peaks share identical retention times and relative intensities (Fig. : I ) . Since a single amino acid change in a given peptide would he expected to alter its elution, the different retention times o f the remaining half of the tr.yptic peptides establish that C-1 and C-2 are isozymes of alkaline phosphatase.
The tryptic digest of the adult hovine alkaline phosphatase, R-2, gives a chromatogram practically superimposahle on that of C-2 (Fig. 3) . Each peak common to hoth (’-1 and C-2 is also present in R-2, but all peptides unique to C-1 are ahsent, while those diagnostic of C-2 are present in the chromatogram of R-2 as well. There is one annmalv hetween C-2 and H-2 at 110-115 min which mav relate to differences in glvcosylation. The sugar analyses suggest that there is significant difference at least quantitatively.
N-terminal Amino Acid Sryurncrs nf thr Isoz~~mrs--N- terminal amino acid sequences of the two calf isozymes and the adult R-2 enzyme have been determined as far as residue 23 by automated Edman degradation using hoth gas- and solid- (data not shown) phase methods (Fig. 4 ) . T h e S- terminal sequences for C-2 and €3-2 are identical, while C-1 differs from the other two at three positions. All three amino acid substitutions, IIe -. Val at position 2 , Ala -. Val at position 4, and Asn Asp at position 8, can he accounted for by single hase changes. The onlv other puhlished sequence of a mammalian alkaline phosphatase is for the human placental enzyme (151, also shown in Fig. 4. The intestinal and placental proteins are extensively homologous at the N terminus.
The tryptic peptide maps clearlv implv that there are con- siderably more amino acid differences in other parts of the two isozymes. These data clearly rule out the mechanism hv which “isozymes” are generated in the harterial system: suh- units with two N-terminal sequences, NH,-Arg-Thr- and NH2-Thr-, are present due to variation in either the initial processing of the signal peptide or a later secondary proteol- ysis (21, giving rise to three “isozyme species” on gels hecause of homo-Thr dimers, homo-Arg dimers, and heterodimers.
The total numher of tryptic peptides ohserved, excluding the standard column artifacts (astrrisks, Fig. 3 1 , is -40 . Cal- culating from the numher of I,vs and Arg present in each isozyme, 39-43 tryptic peptides, depending on isozvme. are expected from complete digestion. Thus digestion is complete. The reason for adding EDTA to the denaturing digestion mix is that chelation of Zn(I1) renders the protein much more susceptible to tryptic digestion.
Endo- /j- N - a c e t ~ l ~ l u c o a a m i n i d a . ~ ~ - F Ikglycosylntion - C - 1, C-2, and R-2 each gave three similar handing patterns on SDS gels after enzymatic deglycosylation with endo-/j-.’V- acetylglucosaminidase-F (Fig. BR): the topmost band is the original intact protein while the two lower bands represent N-deglycosylated intermediates. The presence of two hands of lower apparent molecular weight armnes for two .V-linked glvcoconjugates/monomer. All of the hands stained Shiff- positive, thereby demonstrating at least one 0-linked oligo- saccharide/monomer. The removal of the 0-linked sugar hy alkaline hydrolysis is in progress.
Zinc Content-All three alkaline phosphatases descrihed

11192 Intestinal Alkaline Phosphatase , , ,5 , , 10 15 20 , 25 30 35 BOVINE INTESTINE B-2 L V P V E-€-E D P-A-F-W-N-R-Q-A-A-Q-A-L-D-V-A K-K-L-Q-P-I- - -A-A-K- CALF INTESTINE C-1 L I P A E-E-E N P-A-F- -N-R-Q-A-A-Q-A-L-D-V-A CALF INTESTINE C-2 L V P V E-E-E D P-A-F-W-N-R-Q-A-A-Q-A-L-D-V-A K-K-L-Q-P-I-Q- -A-A-K-N-V-I-L-F- HUMAN PLACENTA 1-I-P-V-E-E-E-N-P-D-F-W-N-R-E-A-A-E-A-L-G-A-A-K-K-L-Q-P-A-Q-T-A-A-K-N-L-I-I-F-
FIG. 4. N-terminal amino acid sequences of isolated bovine intestinal alkaline phosphatases, C- 1 , C- 2 , and B-2. After four separate determinations, residue 12 for C-1 is unknown; it may be cysteine, given that the protein was not carboxymethylated for this experiment. The N-terminal amino acid sequence for human placental alkaline phosphatase (15) is given for comparison.
here were found to contain, by atomic absorption spectrom- etry, 6 g atoms of Zn/dimer in their isolated forms. It was found that removal of the metals by treatment with ammo- nium sulfate as per Gettins and Coleman (16) resulted in loss of catalytic activity which was restored upon adding back zinc plus magnesium or calcium. Further studies concerning metal binding are in progress.
Conclusion-As C-1 is not present in the adult cow, we conclude that it is a fetal form which, although still being synthesized in 3-month-old calves, is not expressed in the adult. There are several examples of such developmental changeovers in the literature (17). In the case of alkaline phosphatase, this is the first evidence documenting the regu- lation of the enzyme at the transcriptional level.
Recent comparisons of human alkaline phosphatases iso- lated from fetal and adult tissues show the two enzymes to have different rates of migration on SDS-PAGE and to be distinguished by binding studies with monoclonal antibodies (18). While changes in post-translational glycosylation could explain some of the differences, at least one CNBr peptide that differs between the two enzymes does not contain de- tectable carbohydrate. The investigators suggest that the fetal and adult enzymes may be the products of two different genes (18).
Acknowledgments-We would like to thank Dr. K. R. Williams for his generosity in rendering protein sequencing, high performance liquid chromatography, and amino acid analysis instrumentation available to us, as well as his valued suggestions and advice. We are also grateful for the technical assistance of Kathy Stone. Finally we acknowledge Drs. T. Ariga and R. K. Yu who generously assisted us in the determination of carbohydrate contents.
Supplementary Material
to
Isozymes of Bovine Intestinal Alkaline Phosphatase
narc BeSman and Joseph E . Coleman
L&.ation o f h o m o a e n e o u s e DhosDh- tines (average length = 2Oml from freshly slaughtered calves were collected
. Four to eight small intes-
contents were extruded. The tissue w a s slit. cut into 20-30 Cm segments on ice. After removing connective tissue and extraneous fat. the lumen
and combined with the lumen pool. diluted 1 : l (vlvl with 25OmH SuCCOSe. homogenized. extracted With n-butanol and Selectively precipitated between
procedure is similar to that described by Brenna et al. ( 5 1 : however. it 4 0 % and 8 0 % saturated ammonium sulfate (Schwarz-Man". ultra pure). This
should be noted that the enzyme from fresh intestine is found wholly in a buoyant lipid-like layer at 8 0 % saturation, a layer which is Separate from the pellet. This layer was resuspended in lOmR Tris HC1 pH 8 . 0 . All
was dialyzed against lOmH Tris HC1 pH 8 . 0 . ImM Mg(II1. O.lmH ZnIII) (to subsequent purification steps were carried Out at bo. The pooled material
is HCI PH 8.0. containing 0.1% (wlv) sodium deoxycholate1 until equili- restoce metals to alkaline phosphatase) and then against buffer-A ( 1 0 M
brated.
( 4 x 3 0 cm). washed with buffer-A and eluted with a linear gradient of 0 to The dialyzed sample was loaded on a DE-52 (Whatman) ion-exchange Column
100 mH N a C l in buffer-A. Active fractions were pooled. concentrated by
against buffer-8 (15mH imidazole'HC1. pH 1 . 3 . containing 0 . 1 % ( v l v l Triton ultrafiltration using a Diaflo XH-50 or YM-30 membrane (Amiconl and dialyzed
x-1001.
1.
2.
3.
4.
5.
6.
7. 8. 9.
10.
11.
12. 13.
14.
15.
16.
17.
18.
REFERENCES
McComb, R. B., Bowers, G. N., Jr., and Posen, S. (1979) Alkaline Phosphatase, Plenum Press, New York
Coleman, J. E., and Gettins, P. (1983) Adu. Enzymol. Rel. Areas Mol. Biol. 5 5 , 381-452
Wyckoff, H. W., Handschumacher, M., Krishna Murthy, H. M., and Sowadski, J. (1983) Adv. Enzymol. Rel. Areas Mol. Biol.
Fosset, M., Chappelet-Tordo, D., and Lazdunski, M. (1974) Bio-
Brenna, O., Perrella, M., Pace, M., and Pietta, P. G. (1975)
Landt, M., Boltz, S. C., and Butler, L. G. (1978) Biochemistry
Laemmli, U. K. (1970) Nature 227,680-685 Edelhoch, H. (1967) Biochemistry 6, 1948-1954 Brown, W. E., and Howard, G. C. (1983) Methods Enzymol. 9 1 ,
Merrill, B. M., Williams, K. R., Chase, J. W., and Konigsberg,
Williams, K. R., Murphy, J. B., and Chase, J. W. (1984) J. Biol.
Ando, S., and Yu, R. K. (1979) J. Biol. Chem. 2 5 4 , 12224-12229 Rosenzweig, S. A., Madison, L. D., and Jamieson, J. D. (1984) J.
Misch, D. W., Giebel, P. E., and Faust, R. G. (1980) Eur. J. Cell
Ezra, E., Blacher, R., and Udenfriend, S. (1983) Biochem. Bio- phys. Res. Commun. 116 , 1076-1083
Gettins, P., and Coleman, J. E. (1983) J. Biol. Chem. 258 , 396- 407
Stamatoyannopoulos, G., and Nienhuis, A. W. (eds) (1981) Hemo- globins in Development and Differentiation, Alan R. Liss, New York
Vockley, J., D'Souza, M. P., Foster, C. J., and Harris, H. (1984) Proc. Natl. Acad. Sci. U. S. A. 8 1 , 6120-6123
55,453-480
chemistry 13, 1783-1788
Biochem. J. 151 , 291-296
17,915-919
36-41
W. H. (1984) J. Biol. Chem. 259, 10850-10856
Chem. 259, 11804-11811
Cell Biol. 99,1110-1116
Biol. 2 1 , 269-279
P B E - 9 1 (Pharrnacial equilibrated wlth buffer-B by eluting with Polybuffer The pooled sample from DE-52 was chromatofocused on a 1 x 3 s Cm bed of
assayed for activity. the two resolved isozymes, C - 1 and C - 2 . pooled 1 4 (Pharrnacial. pH 5 . 0 . containing 0.1% (vlvl Triton X - 1 0 0 . Fractions were
separately and dialyzed against buffer-A.
FRfiCTION NUMBER , , 7 m l l
. 1 . Activity elution profile for chromatofocused calf (lower *nd adult bovine (upper panel1 material, as described in the experimental section. The calf resolves into two peak8 of activity, C-1 and C-2. while the adult gives but one. The latter eluted as a broad envelope due to overloading: a narrow peak characteristic of C-2 was observed when less material was used.
Intestinal Alkaline Phosphatase
a Dhosnhonate affinitv ~ r s i n (Seuharose-ls (Pharmacial couoled to LkHiS. The final step which gives homogeneous protein is chromatography on
. . in turn coupled to p-amino benrylphosphonate by a diazo linkage) as previously described 1 6 1 . C- -1 01 C-2 in buffer-A were loaded on a 1 x 5 0 cm column and after thorough washing. the alkallne phosphatase vas eluted with
were pooled a n d concentrated a s before. Preparation of the adult bovine buffer-A containing 5OmH potassium phosphate. pH 7 . 5 . The active fractions
enzyme Y ~ E identical. except that it vas often advantageous to chromatofocus twice, poollng the purest fractions as determined by SDS-PAGE.
previously described i s ) . SDS-PAGE was performed after Laernrnli ( 1 1 . Proteins were routinely detected with Coomassie Brilliant Blue R-250; a silver Stain
vlsual~zed by the periodic acid-Schiff method. Amino acid analyses were IBio-Rad1 w a s u s e d w h e n appropriate. Glyco-conjugated proteins were
performer1 on a Beckman l 2 l M anino a c l d analyzer. Cysteine v a s determined by performic a c l d ox~dation prior to hydrolysis and tryptophan was estimated f rom a b s o r b a n c e measurements iccording to the rnett,od Of Edelhoch ( X I .
. .
N-terminal a-o a u d sea"- c h a n g e s of H 0 I P Spectrapor-6 membranes treated a s per Brown and Howard
. . 1 - 2 n n a l protein was d l a l y z e d against 2
a n d sequenced on an Applied BioSySteTS 470-A gasphase sequencer operated ( 9 1 and lyoph?llzed. Samples were taken up in trifluoroacetlc acid isequemat)
a c c o r d ~ n q to the manufacturer's program "OINRUN". Phenylthlohyda"toin~amino acid deriratlves w e ~ e identlfled on a Yaters HPLC 5ySten 1 1 0 ) .
h d 'I . 1 - 2 "mol protein was dialyzed against H 0 as above and ~ ~ ~ ~ ~ ~ ~ z e Y d . ~ ~ k T r y sample was resuspended in RN u r e a ,iSch:arr-Mann. ultra pure1 a n d d l q e s t e d with Lrypsin-TPCK lWorthinqton1 at 31' for 3 2 hrs according to the method Of l l i l l a m a et a l . 1 1 1 1 except that 10mM EDTA w a s a l s o 1nr:uded. R e a c t l o n w a s halted t , the ac::ditlan of o h e n v l
GALACTOSE MANNOSE GLUCOSE
N-ACETYL GALACTOSAMINE 0 . 5 0 . 5 0 . 5
GLUCOSAMINE N-ACETYL
6 1 4 1
3 1 4
SIALIC ACID 0 . 5 0 . 5 0 . 1
TOTAL AMINO SUGAR 3 1 4
TOTAL SUGAR 10 22 1 2
derivatives by the method of Ando a n d Yu ( 1 2 1 h a s some The g a s chromatography of the trifluoroacetyl sugar
quantitative limitations for glycoproteins, pa~ticularly ~n resolving the contributions of the three neutral hexoses. Hence the percentages of carbohydrate content shoud be considered approximate.
'A single previous literature determination (4) for calf
aresumablv a mixture of the two isozymes1 reported 6% neutral intestinal alkaline phosphatase (commercially derived and
s u g a r and 6% amino sugar and the absenc; Of sialic acid.
c
J"tli tt>e previous 5 t a l n b a s 3art :y s l s l t . l e . p r v t e 1 n g l j c o ~ ~ ~ , n ~ u g a t e 5 Yere v i s ~ a l l z e d using tile p e r l o r l x a c 1 ? - 3 c t , 1 f f s t a l o . I f desired. the ?el c o u l d -. Reverse phase HPLC elution of tryptic digests Of C-1. C-2 then be resta.nrd w l t t , C o o n a s ~ ~ e l a t e r . and 8-2 forms of alkaline phosphatase. Detection was at 21Onm. 0 . 1
1.0. full sca le . using a two-buffer systemof0.011 1 ~ 1 ~ 1 trifluoroacetic acid (AI and 0 . 0 5 % ( v l v ) trifluoroacetic acid in 80% i v l v ) acetonitrile ( 8 1 , elution was accomplished with the following gradient program: time=o, 1001 A: t-90 mi".. 62.31 A , 3 1 . 5 1 8; t-135 mi".. 251 A. 1 5 1
cld Annlysls of 1-e Phosokt.aS3A B: t-110 mi".. 100% 8. Flow was 0 . 1 mllnin. Each hash-mark 0" the abscissa denotes five minutes.
T W . .
C-l
CY s
ASX 113
THR 6 3
SER 3 0
GLX 1 0 8
PRO 64
GLY 9 8
ALA 123
VAL 1 2
MET 2 5
ILE 3 1
LEU 82
TY R 4 1
PHE 3 3
H I S 2 1
LYS 5 1
ARG 4 8
TRP
~.~...."~~.~... 1 4
"...."_.......
rm x lOOOa 112
..
C - 2
~~~
I 3
126
1 1
5 2
1 0 8
6 s
96
1 2 7
8 2
2 3
29
8 0
4 9
3 0
3 0
4 8
4 4
12
8-2 CALF HUMAN INTESTINE ( 4 1 PLACENTA ( 1 5 1
""
12
1 2 2
1 3
6 3
105
6 2
93
126
8 4
2 5
2 4
1 5
4 0
3 0
2 1
4 4
42
10 10
I28 110
1 1 8 0
4 7 64
103 1 2 0
6 0 6 6
9 0 1 0 8
1 2 0 1 3 2
8 8 1 0
I 5 1 8
30 36
1 4 96
4 1 4 4
31 4 4
3 1 3 1
53 4 8
1 0 2 8 0
I3 8
116 110 126 1 2 7
peptides common to a l l three enzymes. while those in the upper panel Peaks marked by vertical lines in the middle panel iC-11 denote
peaks having asterisks originate in the solvent. (8-11 are 'adult" and those in the lower panel ( C - 1 ) are -fetal'. Those
aHolecular weights for C-1. C-2 and 8-1 estimated by enzymatic dealvcosYlation followed by SDS-PAGE. Compositions were for 1 1 . 1 1 - a n d 1 d e t e r m i n b t i o n s f o r C-1. C-I and 8-2 respectively: residue numbers are ~ 1 0 % . with the exception of proline i t 2 0 1 1 .
"