the (leptothrix) buccalismmbr.asm.org/content/25/2/142.full.pdf · leptotrichia (leptothrix)...
TRANSCRIPT

THE CLASSIFICATION OF ORGANISMS TERMEDLEPTOTRICHIA (LEPTOTHRIX) BUCCALIS
II. REPRODUCTION OF Bacterionema matruchothi
MARION N. GILMOUREastman Dental Dispensary, Rochester, New York
INTRODUCTION
Cultures of Bacterionema matruchotii (5)exhibit a number of cell morphologies. Thepredominant form is a filament attached to abacillus-like body, the two generally beingseparated by a septum; other cell types arefilaments of varying lengths, bacillus-likebodies, and bacillus-like bodies with more thanone filament attached (1, 4, 7). The interrelation-ship of these cell forms is unknown. However,such information is necessary for the determina-tion of culture purity, for classification, and forthe initiation of genetic experiments. For thesereasons, observations of the mode of reproduc-tion of this microorganism were undertakenemploying time-lapse photography of thegrowth of single cells in slide cultures.
MATERIALS AND METHODSTen of the strains described by Gilmour and
Hunter (4) were maintained aerobically onslants of brain heart infusion agar (Difco)supplemented with 0.2 per cent yeast extract(BBL), and transferred at 3-week intervals.
Inocula and experimental media were asfollows. Three-day aerobically or anaerobicallyincubated streak plates of brain heart infusionagar supplemented with 0.2 per cent yeastextract were employed to inoculate the respectiveaerobic or anaerobic microcultures. Using 32Xmagnification, part of a colony was teased apartin 0.1 ml brain heart infusion broth supplementedwith 0.2 per cent yeast extract and 5 per centsheep serum to yield a suspension containingapproximately 500 to 1000 cells. Aerobic micro-cultures were prepared in brain heart infusion
1 This investigation was supported by a researchgrant, D-370, from the National Institute ofDental Research, U. S. Public Health Service. Apreliminary report of this work was presented atthe meeting of the International Association forDental Research, San Francisco, California,March 19, 1959.
broth supplemented with 0.2 per cent yeastextract and 2.5 per cent agar. Because anaerobicgrowth of single cells was poor on this medium,and its Eh was relatively high, a medium of thefollowing composition was employed for theanaerobic microcultures: brain heart infusionbroth (Difco), 18.5 g; soluble starch (Baker), 1 g;yeast extract (BBL), 2 g; glutathione (NutritionalBiochemicals Corporation), 1.23 g; agar, 25 g;distilled water, 1000 ml; pH 7.4.
Microcultures were prepared as described byKnaysi (9) with the following minor modifica-tions. One no. 2 (20 by 50 mm) cover slip wasemployed both for oil immersion objective useand to give a larger reservoir of medium. Topreclude the possibility of either an air space ora thin liquid boundary existing between themedium and the lower side of the cover slip,the cover slip was inoculated with a droplet ofthe inoculum suspension, and the cells wereembedded in place with a droplet of the samemelted and cooled agar medium to be employedin the microculture. This step not only held thecells close to the cover slip, but also increasedcell viability, as compared to that with theKnaysi cell. The cover slip was inverted over thecapillaries, and melted agar medium at 55 Cquickly run between the two inner capillaries,the slide, and the cover slip.For anaerobic incubation, the modifications
included: placing the inner two capillariescloser to the outer capillaries; running agarmedium to within 0.2 mm of the breadth of theslide; and completely sealing the cell and capil-laries with vaspar. Additionally in some experi-ments, yeast was cultured in the two areasexisting between the inner and outer capillariesfor a carbon dioxide source. To show thatanaerobic conditions existed in the anaerobicculture chambers, the aerobic microorganism,Nocardia asteroides (kindly supplied by Dr. A.Howell, Jr.) was inoculated in both aerobic andanaerobic microcultures. Growth occurred onlyunder aerobic conditions.
142
on July 6, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

CLASSIFICATION OF ORGANISMS TERMED LEPTOTRICHIA. II
Following its completion, the microculture wasplaced in a stage incubator regulated at 37 C, orobserved at room temperature of 29 to 32 C.Cells which were immediately on the underside ofthe cover slip and in one plane of focus werechosen for observation. Three pieces (2 by 5 by Y4in.) of heat absorbent glass (Libby Owens-Ford,Toledo, Ohio) were placed under the condenserto absorb heat and to serve as a blue-green filter.Initially, camera lucida drawings recorded timedobservations made with a 50 X American Opticalachromatic oil immersion objective, na 0.95, and10X oculars on a Bausch and Lomb researchmicroscope. Later observations employed aLeitz ortholux microscope fitted with 10 X pari-plan oculars and the 50X oil immersion objec-tive. Photomicrographs were made with LeitzIbsco and Photovolt attachments, a Leitz camera,and Kodak 35 mm panatomic X film.
RESULTSDifferences in cell size and the occurrence of
branching were found, dependent upon aerobic oranaerobic growth conditions. Growth underanaerobic conditions is described first becausethe larger cell size is preferable for detailed ob-servations. The development of a single cell underaerobic conditions is shown to illustrate branch-ng.
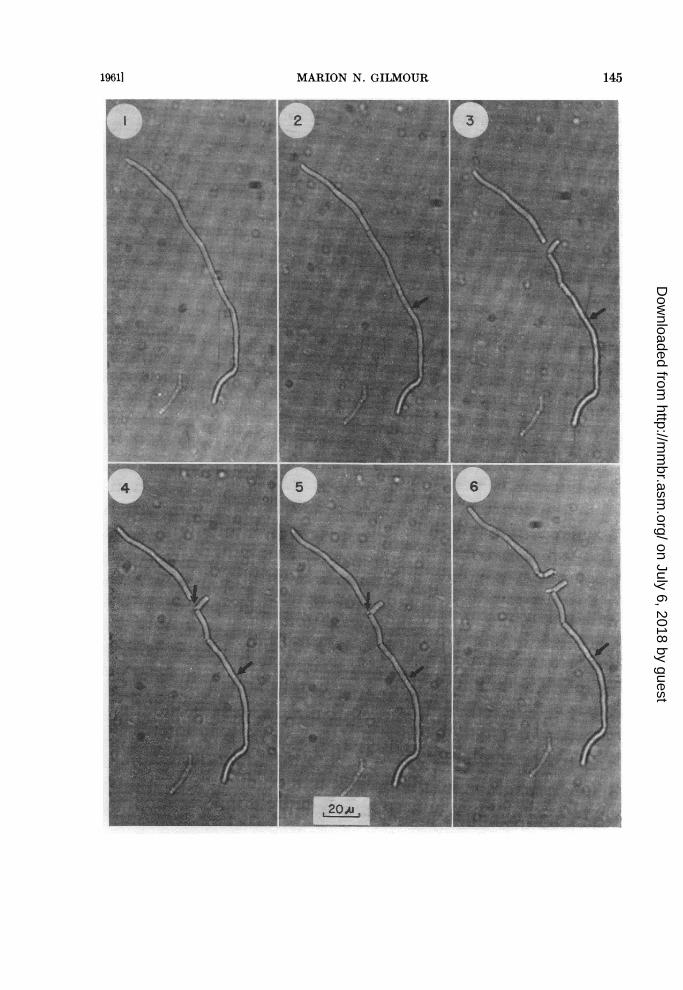
Anaerobic, starting with a filament. The se-quence of events, beginning with an apparentlynonseptate filament of a large-celled strain, isshown in figures 1 to 11. Figure 1 illustrates thefilament after 2 hr of observation during whichtime the sole change was a breadth increase of0.3 ju. After 2 hr, 30 min, the filament formed adark cross wall (figure 2), and 30 min later (figure3, at 3 hr), fragmentation occurred at the crosswall and in the segment below it resulting in theformation of a bacillus-like body. Furthermore, asecond cross wall had formed, the fragmentationof which is illustrated in figures 4 (at 3 hr, 10min), 5 (at 3 hr, 15 min), 6 (at 3 hr, 45 min), and7 (at 3 hr, 52 min). Further fragmentation of thelower segment occurred as shown in figures 8(at 4 hr), 9 (at 4 hr, 35 min), and 10 (at 5 hr, 10min), and fragmentation of the upper segment isshown in figures 3 and following. With highlycritical focussing and the transmitted light em-ployed, the point at which fragmentation oc-curred could sometimes be seen to be precededby the formation across the cell of an oval white
spot which subsequently became dark (see lowerarrows figures 2 to 10). The force of the ensuingfragmentation was such that the resulting bacil-lus-like body moved in 2.5 per cent agar, andformed a zig zagged row with other bacillus-likebodies if a series of consecutive septa fragmentedeither simultaneously or over a relatively shortinterim (figure 11).The separated bacillus-like body formed at
3 hr (figure 3) demonstrates the general fate ofthese cell forms. Ten minutes after separation,one corner had formed a protrusion (see upperarrow, figure 4, at 3 hr, 10 min) which 5 min laterwas clearly separated from its mother cell by aseptum (see arrow, figure 5, at 3 hr, 15 min).Thirty minutes later, the protrusion had elon-gated to form a tube (figure 6 at 3 hr, 45 min),and continued to elongate (figures 7 to 11) untilit was a true filament.
Rarely, the bacillus-like bodies formed fromfragmentation divided to form smaller bacillaryelements. The upper filament segment of figures3 (at 3 hr), 4 (at 3 hr, 10 min), and 5 (at 3 hr, 15min) had at its lower extremity a septum. Duringthe 30 min between figures 5 and 6, fragmentationoccurred at this septum to yield one large bacillus-like body which divided within 15 min to yieldthe two smaller bacillus-like bodies shown infigure 6 (at 3 hr, 45 min). The body on the rightin figure 6 immediately germinated, and 25 minafter its formation, the bacillus-like body on theleft formed a small protrusion (see upper arrow,figure 8 at 4 hr) which enlarged and was separatedby a septum from its mother cell within 35 min(see upper arrow, figure 9, at 4 hr, 35 min). Subse-quent elongation of both germination tubes isshown in figures 10 (at 5 hr, 10 min), and 11 (at10 hr, 25 min). Later observations, which arenot illustrated here, showed fragmentation ofthe remainder of the lower segment of the originalfilament and of the two new filaments attachedto bacillus-like bodies in figure 11. The upper re-maining segment of the original filament hadnot yet fragmented 6 hr after figure 11 had beenphotographed, but continued the elongationshown here (compare figures 6, at 3 hr, 45 min,and 11, at 10 hr, 25 min).
Anaerobic, starting with a bacillus-like body. Toshow the generality of the above mechanism, thedevelopment of a number of bacillus-like bodieshas been followed. A typical sequence of events isillustrated in figures 12 to 21, in which observa-
19611 143
on July 6, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

CLASSIFICATION OF ORGANISMS TERMED LEPTOTRICHIA. II
tions were made of the anaerobic growth ofanother large-celled strain. The original cell isshown in figure 12, the circle at the upper ex-tremity of the cell being caused by dust in theoil of the oil immersion objective. One hour andforty minutes later (figure 13), a germinationtube, separated from the mother cell by a septum,had formed at the upper end of the cell, and asecond protrusion (see arrow) could be seen atthe lower extremity. After 3 hr, 40 min (figure 14),both protrusions were larger, well defined, andseparated by septa from the mother cell. Sub-sequent elongation of both is illustrated in figures15 (at 5 hr, 55 min) and 16 (at 10 hr, 55 min); andfurther increases in both diameter and length, infigures 17 (at 16 hr, 23 min) and 18 (at 19 hr). At22 hr, 30 min (figure 19), the lower new fila-ment fragmented and the resultant uppermostbacillus-like body germinated 2 hr and 34 minlater forming a short filament separated fromits mother cell (figures 20 and 21). It should benoted that branching was not observed in any ofthe anaerobic cultures.
Aerobic, starting with a filament. To determinethe importance of the above method of reproduc-tion in colony formation and to illustrate thedevelopment of branches, observations were madeof the aerobic growth of single cells into largemicrocolonies. The essential features from atypical experiment are presented in figures 22 to33. Observations were initiated on two cellsseparated by a septum (see arrow, figure 22). Atotal of three germination tubes arose from thisseptum, one of which is first seen in figure 23,the other two in figure 26. The first germinationtube elongated to a filament (figures 24 and 25),which fragmented to yield a long segment andtwo bacillus-like bodies, of which one immediatelygerminated (figure 26). The latter cellular en-tities separated, the germination tube and thelong segment elongated to form filaments, and
the second bacillus-like body divided to form twonew bodies (figures 27, 28, and 29). One of thelatter germinated; the other disintegrated after44 hr of incubation. The disintegration of bacillus-like bodies prior to their germination is not arare phenomenon; the designated body in figure32 is another example of this. The developmentof other filaments, their fragmentation, and thegermination of the resultant bacillus-like bodiesto form new filaments is shown in figures 28 to32. The two types of branching which occur areillustrated in figure 33. Y forms occurring on theends of filaments are typical. The second type,which is relatively rare, originates from pro-trusions which arise on the sides of the filaments(poorly focussed protrusions are on the upper-most cell on the left, figure 33) and subsequentlyelongate (lower cell on the extreme right, figure33). Septa between the branches and the motherfilaments were not observed here but formedlater, often concomitantly with filament frag-mentation of the branches, which was followedby germination of the bacillus-like bodies andsubsequent filament formation.
Aerobic, starting with a bacillus-like body. Ob-servations of the aerobic growth of bacillus-likebodies gave results similar to those illustratedabove with anaerobic incubation (figures 12 to21) and with aerobic incubation (figures 30 to32).
Although it did not occur in this series of photo-graphs, three and four filaments can arise con-secutively from one bacillus-like body, yieldingthe type of configuration illustrated in figure34. Cell-wall staining has shown septa betweenthe filaments and the bacillus-like bodies. Thistype of configuration can be termed branched,but is unlike the two types of branching de-scribed above since septa are formed shortly afterthe germination of the mother cell.
Figs. 1-11. Anaerobic growth of a filament, Bacterionema matruchotiiFigure 1. At 2 hr after initial observation, showing nonseptate filament.Figure 2. At 2 hr, 30 min, showing one dark septum and at the arrow, a white spot which will later
become dark.Figure S. At 3 hr, showing fragmentation of the filament and new septa.Figure 4. At 3 hr, 10 min, showing germination of the separated bacillus-like body.Figure 5. At 3 hr, 15 min, showing a septum between the germ tube and the bacillus-like body from
which it was derived.Figure 6. At 3 hr, 45 min, showing fragmentation of the upper cell and germ tube development on
one of the resulting bacillus-like bodies.
144 [VOL. 25
on July 6, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from
19611 MARION N. GILMOUR
7TUEI-T
45
145
3
PI"4 6
on July 6, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

MARION N. GILMOUR [VOL. 25
I 8 9
O0
-ash.
146
on July 6, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

CLASSIFICATION OF ORGANISMS TERMED LEPTOTRICHIA. II
Figs. 12-21. Anaerobic growth of a bacillus-like body, Bacterionema matruchotiiFigure 12. At time zero, showing a bacillus-like body alone. The mark at one end of the cell is caused
by dust in the oil.Figure 13. At 1 hr 40 min, showing germ tube development at the upper end of the bacillus-like
body.Figure 14. At 3 hr, 40 min, showing a septum between the germ tube and the upper end of the bacillus-
like body and a new protrusion at the lower end of the bacillus-like body.Figure 15. At 5 hr, 55 min, showing growth of germ tubes, both clearly separated from the bacillus-
like body by septa. New cells at the top of the photograph arose from growth of a cell in a lower planeof focus.
Figure 16. At 10 hr, 55 min, showing enlargement of the germination tubes.Figure 17. At 16 hr, 23 min, showing increase in length and breadth of the germination tubes.Figure 18. At 19 hr, showing growth of lower germination tube into a short filament.Figure 19. At 22 hr, 30 min, showing fragmentation of lower filament into two bacillus-like bodies.Figure 20. At 25 hr, 14 min, showing development of a germination from one of the bacillus-like bodies
(see arrow).Figure 21. At 27 hr, 14 min, showing further growth of the germination tube.
Figure 7. At 3 hr, 52 min, showing growth of both germination tubes, and in the lower cell, separationof a bacillus-like body, and the formation of a new septum below it.
Figure 8. At 4 hr, showing further growth of both germ tubes and movement of the bacillus-like bodyimmediately below them.
Figure 9. At 4 hr, 35 min, showing development of a new germ tube on the bacillus-like body fromthe upper cell, movement of the central bacillus-like body from left to right, and separation of thebacillus-like body below it.
Figure 10. At 5 hr, 10 min, showing septum formation and breakage at the white spot indicated byarrow in previous photographs.
Figure 11. At 10 hr, 25 min, showing development of previously described germ tubes into filaments,and the three cell types.
19611 147
on July 6, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

MARION N. GILMOUR
Figs. 22-33. Aerobic growth of a filament, Bacterionema niatruchotiiFigure 22. At zero time, showing two cells separated by a septum indicated by the arro-w point.Figure 23. At 50 min, illustrating formation of a germination tube at septum.Figure 24. At 3 hr, 30 min, showing elongation of the germination tube.Figure 25. At 7 hr, 45 min, showing further elongation of the germination tube to form the filament
on the upper left.Figure 26. At 10 hr, 15 min, showing fragmentation of the filament on the left and development of
two new germination tubes from the septum originally seen.
Coccoidal elements and aerial hyphae havenot been seen in any cultures.
DISCUSSION
The above observations show that the variouscell morphologies found in smears of Bacterionemarnatruchotii are the consequence of the reproduc-tive mechanism. Filament fragmentation occur-ring shortly after septa formation yields bacillus-like bodies which upon germination form one ormore filaments. Thus, bacillus-like bodies alone,bacillus-like bodies attached to one or more fila-ments of varying length, and filaments whicharise by elongation of long fragments, are found.The general method of reproduction described
herein differs to some extent from that brieflydescribed by Baird-Parker and Davis (1) in adiagrammatic illustration based upon cell-wall-stained smears. Filament fragmentation occursnot only at regular intervals along the entirefilament length but also in segments of variouslengths, the shorter of which will germinate, thelonger of which will either elongate to filamentsor divide into smaller bacillus-like bodies. Addi-tionally, septa are formed between the mothercell and the germination tube shortly after germi-nation. It should be noted that although septaare not shown in the Baird-Parker and Davisdiagram, they appear to be present in their photo-graphs of cell-wall-stained smears.The method of reproduction and the branching
capabilities of this gram-positive filamentousorganism predicates classification of it in the
family Actinomycetaceae, which currently com-prises two genera, Nocardia and Actinomyces (2).In addition to other differences from these twogenera (6, 7), Bacterionema matruchotii appearsto differ in its mode of reproduction. Althoughthe literature on the reproduction of Actinomycesand Nocardia is confused and somewhat con-tradictory, the following points of difference ap-pear to hold. (a) The bacillus-like bodies resultingfrom filament fragmentation do not remain asdiscrete entities in Actinomyces and Nocardia, buteither themselves elongate to yield new filaments,or are converted into more coccoidal elementswhich are eventually absorbed during subsequentgermination (3, 8, 10-12). (b) Dichotomousbranching is more frequently found in Bacteri-onema than in Nocardia or Actinomyces (10, 11).(c) Two types of growth apparently exist inActinomyces and Nocardia. Species of both generaexhibit the "angular growth" described by Qrskov(10, 11). Additionally, in filaments of Actinomycesand perhaps some Nocardia species, septa areformed immediately prior to fragmentation,which often yields zigzagged rows of bacillus-likebodies similar to those described here for Bac-terionema matruchotii (8, 10). (d) Coccoidal ele-ments readily found in Nocardia, and also ob-served in Actinomyces (3, 10-12), have not beenseen in pure cultures of Bacterionema matruchotii.The above differences in the reproductivemethods substantiate the view (5) that Bac-terionema matruchotii cannot be classified ineither of the genera Nocardia or Actinomyces.
148 [VOL. 25
on July 6, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

2927
30 31
33
Figure £7. At 16 hr, 45 min, illustrating elongation of all germination tubes and more complete sepa-ration of the bacillus-like bodies.Figure 28. At 19 hr, showing growth of germination tubes to form short filaments.Figure 29. At 22 hr, 45 min, illustrating further elongation of germination tubes to form filaments.Figure 30. At 29 hr, 15 min, showing fragmentation of filaments on left.Figure 81. At 30 hr, 15 min, showing germination of bacillus-like bodies on the left, and fragmentation
of the filament on the right.Figure 8£. At 31 hr, 45 min, showing elongation of germination tubes on the left, germination of
bacillary elements from the fragmented filament on the right, and fragmentation of a third filament.Figure 88. At 40 hr, illustrating germination tube elongation to form filaments, and two types of
branching. Dichotomous branching can be seen at the upper portion of the microcolony, and develop-ment of branches from the sides of the mother filament is exemplified on the right.
149
on July 6, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

MARION N. GILMOUR
Figure 34. Bacterionema matruchotii. A bacillus-like body attached to four short filaments. Gramstained preparation from 5-day, anaerobically incubated plate of brain heart infusion agar supple-mented with 0.2 per cent yeast extract and 5 per cent sheep serum.
SUMMARY
These studies show the general method of re-production of Bacterionema matruchotii to con-sist of fragmentation and germination of theresulting bacillary elements. Nonseptate fila-ments form septa sequentially or simultaneouslyover part or all of their length, and fragment.When septum formation and the subsequentfragmentation occur simultaneously over theentire filament length, a zigzagged row of bacillus-like bodies will be formed. If septum formationand the consequent fragmentation occur se-quentially over all or part of the filament, somebacillary cells and other long fragments willarise. The bacillary cells form one to four pro-trusions which enlarge and are separated by aseptum from the mother cell. The protrusionselongate to form filaments which in turn repeatthe fragmentation-germination-filament forma-tion cycle. Long bacillary cells or segments mayeither fragment to form smaller bacillus-likebodies, or may elongate to form free filaments.Although this organism resembles the members
of the family Actinomycetaceae in being gram-positive, in branching particularly under somecultural conditions, and in reproducing by frag-
mentation of originally nonseptate filaments, theabove mechanism differs from those describedfor the species of Nocardia and Actinomyces.
REFERENCES
1. BAIRD-PARKER, A. C. AND DAVIS, G. H. G.1958 The morphology of Leptotrichiaspecies. J. Gen. Microbiol., 19, 446-450.
2. BREED, R. S., MURRAY, E. G. D., AND SMITH,N. R. 1957 Bergeys manual of deter-minative bacteriology, 7th ed. The Williams& Wilkins Co., Baltimore, 1094 pp.
3. CLARK, J. B. AND FRADY, J. J. 1957 Second-ary life cycle of Nocardia corallina. J.Bacteriol., 74, 698.
4. GILMOUR, M. N. AND HUNTER, P. A. 1958Isolation of an oral filamentous micro-organism. J. Bacteriol., 76, 294-300.
5. GILMOUR, M. N., HOWELL, A., JR., ANDBIBBY, B. G. 1961 The classification oforganisms termed Leptotrichia (Leptothrix)buccalis. I. Review of the literature andproposed separation into Leptotrichiabuccalis Trevisan, 1879 and Bacterionemagen. nov., B. matruchotii (Mendel, 1919)comb. nov. Bacteriol. Rev., 25, 131-141.
6. GILMOUR, M. N. AND BECK, P. A. 1961 Theclassification of organisms termed Lepto-
150 [VOL. 25
...
on July 6, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

CLASSIFICATION OF ORGANISMS TERMED LEPTOTRICHIA. II
trichia (Leptothrix) buccalis. III. Growthand biochemical characteristcs of Bac-terionema matruchotii. Bacteriol. Rev.,25, 152-161.
7. HOWELL, A., JR., AND ROGOSA, M. 1958Isolation of Leptrotrichia buccalis. J.Bacteriol., 76, 330-331.
8. HURST, V. 1950 Morphologic instability ofactinomycetes associated with enamel. J.Dental Research, 29, 571-582.
9. KNAYSI, G. 1957 New technique of growingmolds in slide culture under environmental
conditions similar to those prevailing in thepetri dish. J. Bacteriol., 73, 431-435.
10. MCCLUNG, N. M. 1949 Morphologicalstudies in the genus Nocardia. I. Develop-mental studies. Lloydia, 12, 137-177.
11. QRSKOV, J. 1923 Investigations into themorphology of the ray fungi. Levin andMunksgaard, Copenhagen.
12. WEBB, R. B., CLARK, J. B., AND CHANCE, H.L. 1954 A cytological study of Nocardiacorallina and other actinomycetes. J.Bacteriol., 67, 498-502.
1961] 151
on July 6, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from