the flood pulse concept: new aspects, approaches and
TRANSCRIPT

ABSTRACT
The flood pulse concept (FPC), published in 1989, was based on the scientific experience of the authorsand published data worldwide. Since then, knowledge on floodplains has increased considerably, creating alarge database for testing the predictions of the concept. The FPC has proved to be an integrative approach forstudying highly diverse and complex ecological processes in river-floodplain systems; however, the concepthas been modified, extended and restricted by several authors. Major advances have been achieved throughdetailed studies on the effects of hydrology and hydrochemistry, climate, paleoclimate, biogeography, biodi-versity and landscape ecology and also through wetland restoration and sustainable management of flood-plains in different latitudes and continents. Discussions on floodplain ecology and management are greatlyinfluenced by data obtained on flow pulses and connectivity, the Riverine Productivity Model and the MultipleUse Concept. This paper summarizes the predictions of the FPC, evaluates their value in the light of recentdata and new concepts and discusses further developments in floodplain theory.
117
Junk W.J. Wantzen K.M.
Max-Planck-Institute for Limnology, Working Group Tropical Ecology, P.O. Box 165, 24302Plön, Germany E-mail: [email protected]
THE FLOOD PULSE CONCEPT:NEW ASPECTS, APPROACHES
AND APPLICATIONS - AN UPDATE

INTRODUCTION
Rivers and floodplain wetlands are among themost threatened ecosystems. For example, 77 percentof the water discharge of the 139 largest river systemsin North America and Europe is affected by fragmen-tation of the river channels by dams and river regula-tion (Dynesius and Nilsson 1994). In recent reviews onwetlands, demographic trends, economic and politicaldevelopment, demand for hydroelectric energy andwater, agriculture and animal ranching, eutrophicationand pollution, fisheries, logging, recreation and eco-tourism and invasion by exotic species have been iden-tified as the most important current determinants forthe development of rivers and floodplain wetlands(Tockner and Stanford 2002; Junk 2002). The globalwater crisis and the threat to aquatic organisms, espe-cially riverine biota (Dudgeon 2000; Pringle 2001),increase the necessity to develop models that serveboth science and policy. The flood pulse concept (FPC)(Junk, Bayley and Sparks 1989) was primarilydesigned as a scientific concept, but it also outlinedsome strategies for use, recently specified in Junk et al.(2000). Here, the impact of advances in river ecologyon this and other contemporary concepts is criticallyanalyzed.
THEORETICAL BACKGROUND
HISTORICAL DEVELOPMENT
Limnologists classify inland waters as standingwaters (lakes, ponds) and running waters (streams andrivers). Both system types receive allochthonous sub-stances and produce autochthonous organic matter,both of which are metabolized and recycled. Standingwaters, however, are closed systems that store inorgan-ic and organic matter, circulate organic matter and dis-solved inorganic substances in characteristic internalcycles in the lake basin and eventually deposit them inthe sediments. These systems are characterized mainlyby thermal and/or chemical stratification. Runningwaters are open systems that transport water and dis-solved and suspended matter from the continents to thesea or to endorheic basins. This transport includesintermediate deposition and re-suspension of sedi-ments in the river channel or in the connected flood-
plain, where production and degradation of organicmatter also takes place.
These characteristics are reflected for lakes inthe “Seentypenlehre” (Lake typology), elaborated byThienemann and Naumann between 1915 and 1935(e.g. Thienemann 1925; Naumann 1932) and forstreams and rivers in articles by Illies (1961a) andIllies and Botosaneanu (1963) on the differentiationinto different zones. These zones, described in theseearly studies as rhithron and potamon with epi-, meta-and hypo-subzones, were mainly characterized by abi-otic patterns (current, temperature) and by the occur-rence of distinct animal and plant communities thatdepend on a given set of these abiotic patterns. Forexample, epirithral communities are those typical ofglacier outflows and that depend on low temperatures,high oxygen concentrations and fast current. In latitu-dinal comparisons, Illies (1961b) found evidence for agenerality of this zonation in mountain streams world-wide - high-elevation streams in the tropics have com-munities similar to those of low-latitude coldwaterstreams.
Later, Vannote et al. (1980) substituted thisrather static view of river classification with the RiverContinuum Concept (RCC), which introduced adynamic concept of continuously changing physicalconditions and biological components along the riverchannel, especially regarding the allochthonous andautochthonous inputs and the processing of organicmatter in the flowing water along the river continuum.The RCC predicts major changes in the load and qual-ity of organic matter and the biota in the stream/riverchannel from the headwaters to the lower river cours-es. The high allochthonous input from riparian vegeta-tion in the headwaters decreases with increasing chan-nel width (increasing stream order). Autochthonousprimary production in the headwaters is low because ofshading by trees (P/R <1), increases in the middlereaches because of low water depth and high irradia-tion (P/R >1) and decreases again in the lower reachesbecause of high water depth and increased turbidityand turbulence (P/R <1). In contrast to the zonationconcept, the RCC claims that occurring species are
118 The flood pulse concept: New aspects,

replaced continuously rather than in discrete stages.The percentage of shredders decreases and the numberof collectors increases with increasing stream orderbecause of decreasing input of coarse particulateorganic material and an increasing amount of fine par-ticles owing to the increasing level of processing.Headwater communities tend to optimize their use ofallochthonous matter, whereas an organism living inthe lower river reaches largely depend on the ineffi-ciency of organisms living in the upper reaches toprocess organic material. The interplay of processing,storage and leakage is predicted to reduce the diversi-ty of organic matter types and the maximization ofenergy utilization (i.e. adaptation to poorly degradableorganic matter) along the river continuum.
The RCC further predicts that biodiversity ofaquatic organisms is lower in the headwater regionsand in the lower parts of the rivers and that highestdiversity is found in the middle reaches of the streams,where the variability of temperature, riparian influenceand flow are highest and allow numerous different taxato find their thermal optima.
One of the major constraints of the RCC is thatit was originally based mostly on results from north-ern, temperate, low-order streams with dense treecanopies and steep gradients that flowed towardsmore-or-less-regulated rivers in long-term-managedareas. The hydrology of small streams is stronglyinfluenced by local rainfall and is rather erratic.Flooding of small streams occurs only for short periodsand is often altered by management of rivers in inten-sively used areas. Therefore, flooding events andfloodplains received little or no attention in the firstversion of the concept, but were considered later(Minshall 1985; Sedell, Richey and Swanson 1989).
Floodplains fall into the wetland category,which includes ecosystems at the interface of aquaticand terrestrial ecosystems and are therefore oftencalled ecotones. However, large wetlands have to beconsidered as specific ecosystems with unique proper-ties not adequately covered by present ecological par-adigms and by limnology, estuarine ecology and ter-
restrial ecology (Mitsch and Gosselink 2000).Floodplains are areas that are periodically inundatedby the lateral overflow of rivers or lakes and/or byrainfall or groundwater; the biota responds to theflooding by morphological, anatomical, physiological,phenological and/or ethological adaptations and char-acteristic community structures are formed (Junk et al.1989).
Until the 1970s, floodplains were studied sepa-rately by different disciplines: limnologists studiedfloodplain lakes treating them as classical lakes, ecol-ogists dealt with the terrestrial fauna and flora andhydrologists investigated water and sediment trans-port. An integrated approach was used by Welcomme(1979), who summarized data on floodplain fishery,limnology and hydrology and coupled fish productionwith the nutrient status of the parent rivers and theextent of flooding. The consequences of the fluctuatingwater level on fish have also been summarized byLowe-McConnell (1975, 1987). Bayley (1980) pointedto limits in limnological theory with respect to fishproduction in river floodplains. Junk (1980) describedthe multiple land-water interactions of the CentralAmazon River floodplain, analysed limnological con-cepts of rivers and lakes, pointed out a gap in limno-logical theory and described floodplains as specificecosystems.
During the first Large River Symposium inToronto in 1986, the discussion on the applicability ofthe RCC to large river-floodplain systems led to theformulation of the Flood Pulse Concept (FPC) (Junk etal. 1989). This concept focuses on the lateral exchangeof water, nutrients and organisms between the riverchannel (or a lake) and the connected floodplain. Itconsiders the importance of the hydrology and hydro-chemistry of the parent river, but focuses on theirimpact on the organisms and the specific processes inthe floodplain. Periodic inundation and drought (floodpulse) is the driving force in the river-floodplain sys-tem. The floodplain is considered as an integral part ofthe system that is periodically coupled and decoupledfrom the parent river by the aquatic/terrestrial transi-tion zone (ATTZ). The flood pulse can be monomodal
approaches and applications - an update 119

or polymodal, predictable or unpredictable and with ahigh or low amplitude. Predictable pulsing favors theadaptation of organisms and increases primary produc-tion and efficiency of nutrient use.
The FPC predicts that the nutrient status of thefloodplain depends on the amount and quality of dis-solved and suspended solids of the parent river; how-ever, it includes the premise that internal processes ofthe floodplain and nutrient transfer mechanismsbetween the terrestrial and the aquatic phase stronglyinfluence nutrient cycles, primary and secondary pro-duction and decomposition. At the same time, floodingis considered as a disturbance factor that leads to a reg-ular setback of community development and maintainsthe system in an immature, but highly productivestage.
Another tenet of the FPC is that in the river-floodplain system, a large part of the primary and sec-ondary production occurs in the floodplain, whereasthe river is mainly the transport vehicle for water anddissolved and suspended matter. The river is also therefuge for aquatic organisms during low-water periodsand serves as a route for active and passive dispersal.The “highway analogy” describing the river channel asa transport and migration corridor was used to visual-ize the different functions of the main river channeland its floodplain (Junk et al. 1989). The FPC wasbased on the personal experiences of the authors on theAmazon and Mississippi rivers but also on a vast liter-ature about other river systems. Therefore, the conceptwas not restricted to large tropical rivers, as is some-times cited (e.g. Benke et al. 2000), but was conceivedas a general concept for large river-floodplain systems.
Mostly limnologists, ichthyologists and fish-eries biologists study the ecology of floodplains andtheir organisms. They test the predictions of the FPConly for the aquatic phases of the system. However, ithas to be stressed that the FPC covers the river-flood-plain-system during the entire year and that its predic-tions are also valid for the terrestrial phases that are anintegral part of the river-floodplain system.
FURTHER CONCEPTS IN RUNNING-WATER ECOLOGY
Various conceptual approaches independent of(or complementary to) the RCC and FPC were devel-oped in the 1980s and 1990s on lotic ecosystem struc-ture and functioning. Many early seminal papers dealtwith the distribution of organisms within the lotic sys-tems. A key aspect was the description of hyporheiczones in which important ecosystem processes occur.Early work by Schwoerbel (1961) on the distributionof benthic and stygal fauna in bed sediments wasextended by the description of aquatic organisms farfrom the river channel area (Stanford and Ward 1988)and factors contributing to vertical distribution of theorganisms (Bretschko and Leichtfried 1988) and by theunderstanding of organic matter dynamics in this zone(e.g. Williams 1989; Triska, Duff and Avanzino 1993).Ward (1989) included the function and occurrence ofhyporheic zones in general stream theory by describ-ing streams as four-dimensional systems.
The importance of stream hydraulics (Statznerand Higler 1986) and of disturbance (Resh et al. 1988)for the distribution for benthic organisms has shownthat variation in water flow caused by climate and geo-morphology can set limits to the generalizations of theRCC since flow conditions typical of upstream anddownstream areas can change several times along theriver course (“discontinuum”, Poole 2002).Consequently, the template provided by the habitatconditions (Southwood 1977) and its alignment withspecies traits (Townsend and Hildrew 1994; Resh et al.1994) might be more important for the occurrence of aspecies than the position of the given site along thecontinuum. Since geomorphology is subject to non-continuous local variations, the distribution of streamhabitats appears as a mosaic (Pringle et al. 1988) ofhierarchically ordinated and dynamic patches(Townsend 1989; Poole 2002). Various hierarchicalconcepts have been developed for riverine landscapepatterns and their scale-dependent processes (e.g.Frissell et al. 1986; Townsend 1996; Petts and Amoros1996; Poff 1997; Montgomery 1999; Ward, Malardand Tockner. 2001; Poole 2002).
120 The flood pulse concept: New aspects,

Further conceptual approaches have dealt withthe production and processing of organic matter, suchas the nutrient spiraling concept (Elwood et al. 1983;Pinay et al. 1999), or with human impacts, e.g. theinterruption of natural flow pathways by dams [theSerial Discontinuity Concept (Ward and Stanford1983a; Ward and Stanford 1995)]. As an alternative tothe RCC, Montgomery (1999) proposed a multi-scalehypothesis in which spatial variability in geomorphicprocesses governs temporal patterns of disturbancesthat influence ecosystem structure and dynamics(Concept of Process Domains). Channel networks canbe divided into discrete regions in which communitystructure and dynamics respond to distinctly differentdisturbance regimes.
EXTENSIONS OF THE FPC
HYDROLOGY AND FLOW CHARACTERISTICS
The FPC was developed based upon data andlong-term observations of neotropical (Amazon) andtemperate zone (Mississippi) rivers. It provided a gen-eral outline and strengthened the premise that riversand their floodplains have to be considered as one unitand therefore cannot be treated separately in ecologicalstudies. The FPC has stimulated various studies onriver-floodplain ecosystems in, e.g. Lower Rhinefloodplain lakes (van den Brink et al. 1994), Missourifloodplain lakes (Knowlton and Jones 1997), theDanube (Tockner, Malard and Ward. 2000), theMurray-Darling (Humphries, King and Koehn 1999)and the Mississippi River (Sparks, Bayley and Kohlert1990). Various studies in which the FPC has beenapplied and its tenets tested have lead to proposals forsupplementation to the original conceptual framework.
Increasing knowledge on the hydrologicalcharacteristics of rivers has contributed much towardsthe understanding of their ecological processes. Thehydrographs of individual rivers are influenced by aseries of partly interacting factors, such as climate, gra-dients, landscape morphology, floodplain bufferingand human impacts, which together cause very com-plex patterns. Several authors have identified differentmeasures for identifying the hydrological variability ofrivers and have provided tools for classifying rivers
according to their hydrological signature (Richter,Baumgartener, Powell et al. 1996; Puckridge et al.1998). Providing more detail on the type of flow vari-ation, Puckridge et al. (1998) have stressed the gener-al importance of water level variations even below thebankfull stage (flow pulses), which might have signif-icant influence on the habitat size and characteristics.Irregular flood events, especially in streams (e.g.Winterbourn, Rounik and Cowie 1981) and arid zonerivers, have selected for resilient strategies of organ-isms to survive these events (Poff and Ward 1989;Lytle 2001) rather than to adaptations to profit fromthem (Junk et al. 1989). However, this view has recent-ly been modified because in seasonal climates, theperiod of flash flood events can be predicted andbecause flash-flood events mobilize organic matterresources from stream wetlands (Wantzen and Junk2000).
Tockner et al. (2000) extended the FPC by con-sidering that flooding resets temperature diversity inisolated aquatic floodplain habitats. Thus, aquatichabitats within floodplains might have a much broadertemperature range than the river itself, especially inrivers with wide and diversified beds, such as thealpine Fiume Tagliamento, where flow pulses occurfrequently.
Unpredictable flooding and decoupling of theflood pulse from the temperature pulse leads to lowtemperatures during floods and high temperatures thattrigger spawning of some fish species during low waterlevel in some parts of the Murray Darling River systemin Australia (Humphries et al. 1999, Low FlowRecruitment Hypothesis).
The fact that water level changes influenceriverine systems four-dimensionally in space and time(Ward 1989) is important. Rising water levels not onlyincrease the wetted surface of the channel and eventu-ally of the floodplain, but at the same time influencethe exchange between groundwater and surface watereither by allowing an up-welling of groundwater or byforcing a down-welling of the surface water into theaquifer vertically and laterally. The hyporheic zoneserves as an interface between groundwater and sur-face water (Schwoerbel 1961). Similarly, floodplains
approaches and applications - an update 121
act as interfaces for the interchange between the rivermainstreams and their tributaries or surface runofffrom rainwater. The flow direction of the interfaces isinfluenced by the fourth dimension, time, such that therecent and the past hydrological situations becomedecisive: elevated water levels can cause blocking oreven backflow of the tributaries and groundwater out-flows. In the northern Pantanal wetland, frequentchanges in the flow direction occur in floodplain chan-nels that connect water bodies that receive rain andriver water, depending on the respective water level(Wantzen and Da Silva, unpublished data). In thesouthern areas of the Pantanal, high levels of riverwater block the tributaries after the rainy season; there-fore, a large part of the inundation occurs after cessa-tion of the rainfall (Hamilton, Sippel and Melack1996).
ALLOCHTHONOUS AND AUTOCHTHONOUS PRODUCTION
AND NUTRIENTS
The FPC has focused on the productivity with-in the floodplain areas, in contrast to the RCC, whichhas focused on the import of more-or-less processedallochthonous matter from the upriver sections. Bothconcepts have been criticized by Thorp and Delong(1994) in the Riverine Productivity Model (RPM),which predicts that autochthonous production in theriver channel and allochthonous inputs in the lowerreaches provide a substantial portion of the organiccarbon used by river animals. While analyzing thepotential influence of the floodplain on the carbonbudget in a channel site and a floodplain region of theOhio River, Thorp et al. (1998) did not find significantdifferences in the isotopic C and N signatures, whichindicated a low floodplain contribution during a short-termed, unpredictable flood event at low water temper-atures in wintertime. This, however, does not necessar-ily contradict the predictions of the FPC.
Depending on temperature, light, nutrients andsubstrate conditions, river channels can show a consid-erable autochthonous primary production, which fuelsthe riverine food web as shown for Rhine River(Friedrich and Mueller 1984). Especially in thoserivers where these conditions are beneficial for algal
122 The flood pulse concept: New aspects,
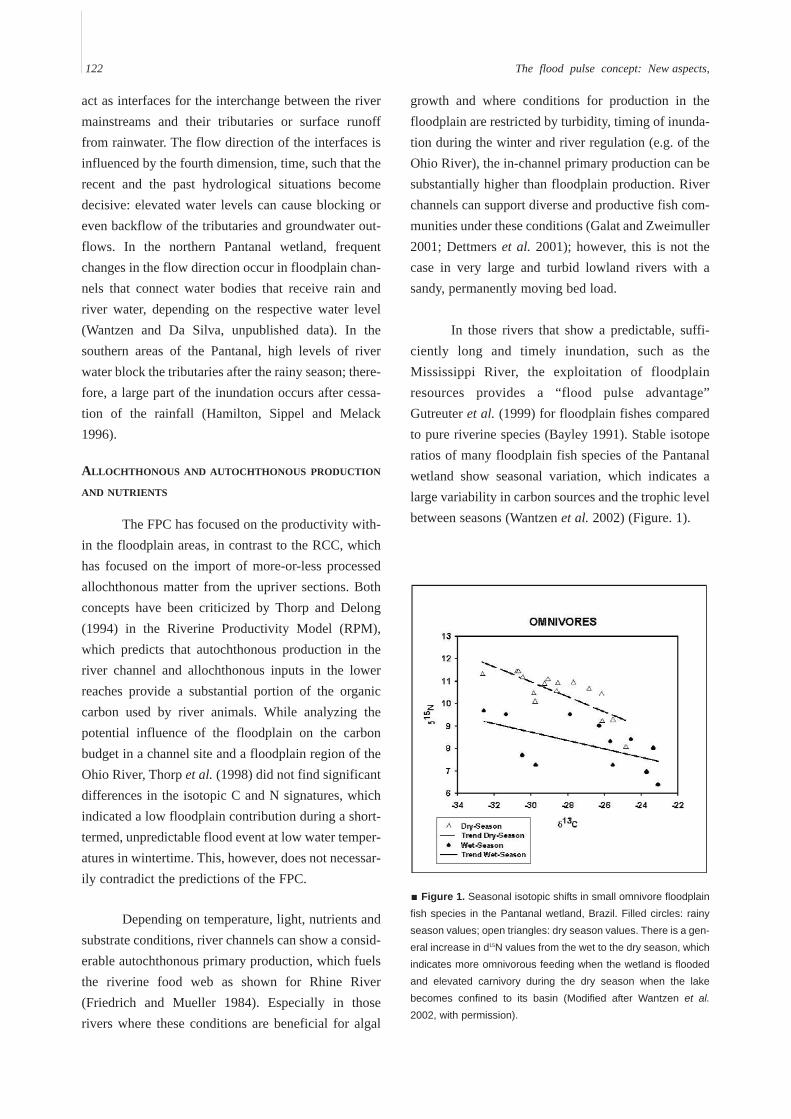
Figure 1. Seasonal isotopic shifts in small omnivore floodplainfish species in the Pantanal wetland, Brazil. Filled circles: rainyseason values; open triangles: dry season values. There is a gen-eral increase in d15N values from the wet to the dry season, whichindicates more omnivorous feeding when the wetland is floodedand elevated carnivory during the dry season when the lakebecomes confined to its basin (Modified after Wantzen et al.2002, with permission).
growth and where conditions for production in thefloodplain are restricted by turbidity, timing of inunda-tion during the winter and river regulation (e.g. of theOhio River), the in-channel primary production can besubstantially higher than floodplain production. Riverchannels can support diverse and productive fish com-munities under these conditions (Galat and Zweimuller2001; Dettmers et al. 2001); however, this is not thecase in very large and turbid lowland rivers with asandy, permanently moving bed load.
In those rivers that show a predictable, suffi-ciently long and timely inundation, such as theMississippi River, the exploitation of floodplainresources provides a “flood pulse advantage”Gutreuter et al. (1999) for floodplain fishes comparedto pure riverine species (Bayley 1991). Stable isotoperatios of many floodplain fish species of the Pantanalwetland show seasonal variation, which indicates alarge variability in carbon sources and the trophic levelbetween seasons (Wantzen et al. 2002) (Figure. 1).

In regulated rivers, where connected lakes rep-resent remains of a floodplain, e.g. in the lower RhineRiver in the Netherlands, the seepage and overflow ofnutrient-rich river water determines the productivityand composition of the phytoplankton such that lakeswith higher connectivity have a higher productivity(van den Brink et al. 1993). In most river-floodplainsystems, primary production in the floodplain is muchhigher than in the channel, (e.g. Australia, review byRobertson et al. 1999). We agree with Dettmers et al.(2001) that organic matter input and production derivefrom the upstream sites (RCC), from the floodplains(FPC) and from the river channel (RPM). The relativecontribution of these sites to the organic matter budgetof a river depends on the production and transport con-ditions in these three units (Figure 2).
approaches and applications - an update 123
Figure 2. Schematic interplay of variable carbon sources in dif-ferent segments of a river. Owing to the topographical variation inthe landscape, the sequence of the segments can vary along theriver course. Top: in catchment areas with strong aquatic-terres-trial interfaces, the floodplain extension is relatively small andinputs of terrestrially produced organic matter are high. Middle: innatural (mountainous, steep-bordered) or human-made (channel-ized) segments, the extension of the floodplains is restricted andcarbon fixation occurs largely by riverine plankton and aquaticmacrophytes. Below: in floodplain areas, carbon can contribute toriver carbon budget via water flow from the floodplain to the main-stream or via feeding migration of fish and other aquatic animalsbetween the floodplain and the main channel.
When considering the contribution of flood-plain carbon to the entire river carbon budget, twoaspects have to be stressed:
Flow conditions vary considerably betweendifferent river-floodplain systems. If geomorphologylimits the exchange between river and floodplain, thecontribution of the floodplain carbon to the main-stream carbon budget can be lower than expected.Lewis (1988) found that in the Orinoco and some trib-utaries, non-floodplain sources, including within-chan-nel and near-channel stagnant or slow-flowing areas,accounted for 63 percent of the annual transport ofphytoplankton carbon, while the floodplain accountedfor only 37 percent.
Mobile organisms such as fish actively seekfloodplain carbon in mass migrations as soon as flood-ing begins in order to feed in the floodplain(Welcomme 1985; Lowe-McConnell 1987;Winemiller 1989; Junk et al. 1997; Wantzen et al.2002). When small floodplain fish migrate back intothe main channel during the falling limb of the hydro-graph, they are preyed upon in large quantities byriverine predators (Wantzen et al. 2002). Therefore,floodplain carbon can contribute significantly to riverfood webs without strong hydrological connectivity.
THE MULTIPLE USE CONCEPT
Floodplain management should be based onconceptional considerations in order to avoid negativeside effects as much as possible (e.g. Nienhuis, Leuvenand Ragas 1998). The FPC predicts exchange of nutri-ents and energy between the aquatic and terrestrialphases. Human use of terrestrial resources will affectaquatic resources and vice versa. These impacts haveto be considered when developing management con-cepts.
The economic and ecological analysis of thedifferent utilization forms led to the formulation of anintegrated multiple-use concept for the central AmazonRiver floodplain (Junk 2000). It favours the optimiza-tion of the use of different resources instead of themaximization of the economic return of a single

resource. Priority is given to the sustainable use offishery resources because of low environmentalimpact, large requirement of labour force and high eco-nomic importance. Subsistence fisheries can be com-bined with smallholder agriculture and dairy farmingon the highest levees and floodplain-adapted sustain-able forest management by selective logging. Large-scale cattle and water buffalo ranching for beef pro-duction and agro-industries are considered destructivefor the ecosystem because of the destruction of flood-plain forests and socially unacceptable because of lowlabor force requirements (Junk et al. 2000).Decentralized administration of floodplain resourcesby local communities is considered essential to stimu-late the participation of the local population in thecomplex management processes (Isaac et al. 1998;McGrath et al. 1999). Multiple use concepts will varyconsiderably for different floodplains because of thelarge variety of floodplain types and related socio-eco-nomic parameters.
OPEN QUESTIONS IN FLOODPLAINRESEARCH
When river or lake water inundates the flood-plain via overspill or via floodplain channels, variouskey processes occur simultaneously: (1) pre-floodthermal and chemical heterogeneity between mainchannel and floodplain water bodies temporarily resets(Sabo et al. 1999a; Tockner et al. 2000); (2) consider-able inputs of mainstream (or lake) water-bound sub-stances (dissolved and suspended, organic and inor-ganic) flush into the floodplain (Fisher and Parsley1979; Lewis et al. 2000); (3) terrestrial habitats areflooded, large amounts of biomass decays and largeamounts of inorganic and organic matter depositedduring the terrestrial phase are mobilized by the over-laying water (Hamilton et al. 1997; Sabo et al. 1999b);(4) terrestrial organisms migrate into non-floodedhabitats or show adaptations to flooding (Adis 1984;Adis, Marques and Wantzen 2001); (5) aquatic organ-isms are flushed or migrate into the floodplain oreclode from resting stages (Welcomme 1985; Irmler1981) and (6) terrestrial carbon and floodplain prod-ucts from the canopy of the floodplain forest, such asterrestrial invertebrates, fruits and seeds, are incorpo-
rated in the aquatic food webs (Junk et al. 1989;Wantzen et al. 2002).
When the water level falls the following keyprocesses occur simultaneously: (1) water stored in thefloodplain with any dissolved and suspended matterenters the parent river or lake (Benke et al. 2000), (2)the ATTZ falls dry and becomes colonized by terrestri-al organisms (Junk and Piedade 1997; Adis and Junk2002), (3) large amounts of water-borne organic car-bon becomes stranded and incorporated in the terrestri-al food webs (Junk and Weber 1996), (4) aquaticorganisms move to permanent water bodies or showadaptations to periodic drought (Irmler 1981), (5) per-manent water bodies become increasingly isolatedfrom the parent river or lake and develop specificphysical and chemical characteristics and specificspecies assemblages (Furch 1984; Tockner et al.1999).
These changes either have a direct influence onaquatic and terrestrial flora and fauna in the flood-plains and related rivers and lakes, for examplethrough changes in the community composition andpopulation density (e.g. Heckman 1998; De-Lamônica-Freire and Heckman 1996; Sabo et al.1999b; de Oliveira and Calheiros 2000), or indirectlytrigger various behavioral traits, such as spawning andmigration of fish (Welcomme 1985; Junk et al. 1997),breeding of waterfowls (Petermann 1999; Magrath1992) and reproduction and migration of terrestrialinvertebrates (Adis and Junk 2002).
The complexity and the interdependence ofthese processes are yet not fully understood. Currently,questions arise about recent, past and future climaticimpacts, the importance of landscape connectivity anddynamics of the flooding on biodiversity and biogeo-chemical cycles and how to include the results offloodplain research into sustainable managementstrategies.
124 The flood pulse concept: New aspects,

THE IMPACT OF FLOOD PULSE ON WET-LANDS IN DIFFERENT CLIMATIC ZONES
The FPC states that the flood pulse is the maindriving force in river-wetland systems. This is true forthe humid tropics, but in lower latitudes, there areother driving forces that also affect the biota andprocesses in the floodplains and that can overlap withthe flood pulse. The FPC mentions these forces, buttheir impacts require more attention in comparativestudies. In semiarid and arid regions, drought and fireaffect the floodplains during the terrestrial phase, withconsequences for the aquatic phase. In temperateregions, biota react to day length and/or temperature(light/temperature pulse) and this cycle is superim-posed on the flood pulse (Junk 1999). Some effects (onfish fauna), because of the decoupling of the floodpulse from the temperature pulse, are discussed byHumphries et al. (1999) for the Murray-Darling Riverbasin. In high latitudes, prolonged ice cover and lowtemperatures strongly affect the biota; and the biotamight require as many adaptations to these events as tothe flood pulse.
PALEOCLIMATOLOGICAL HISTORY OF FLOODPLAINS
The predictions of the FPC also have to beinterpreted in the light of the paleo-ecological condi-tions that have influenced evolutionary processes andrates of speciation and extinction. For instance, theFPC states that predictable pulsing favors the develop-ment of adaptations of fauna and flora and increasesspecies diversity. This statement holds true for someriver floodplains, but not for others. The Amazon Riverfloodplain is very rich in plant and animal species thatare highly adapted to the predictable monomodal floodpulse. Approximately 1 000 flood-adapted tree speciesare found in the floodplains of the Amazon basin. Inthe floodplain of the Mamirauá Reserve near Tefé,about 800 km upstream of Manaus, which covers anarea of about 11 240 km2, until today approximately500 tree species have been recorded, about 80 percentof which are floodplain-specific (Wittmann 2002 andunpublished data). In comparison, the large majority ofthe about 250 tree species of the Pantanal of MatoGrosso, a wetland of approximately 140 000 km2, have
broad ecological amplitude; only about 5 percent arerestricted to regularly flooded areas (Nunes da Cunhaand Junk 2001 and unpublished data). The number offlood-resistant tree species in bottomland hardwoodforests of the USA approaches about 100 species,many of which also occur in the uplands (Clark andBenforado 1981). In northern European floodplains,only about a dozen flood-resistant tree species occur.
Many Amazonian soil arthropods are flood-plain-specific and have complex survival strategies(Adis and Junk 2002). First observations indicate thatflood-adapted soil arthropods in the Pantanal are lesscommon than in Amazonia (Adis et al. 2001).Terrestrial soil invertebrates in Europe are poorlyadapted to the flood pulse. Most are immigrants fromthe non-flooded uplands and suffer high losses duringfloods (Adis and Junk 2002).
Paleoclimatological history analysis ofAmazonia shows that during the last ice age the tem-perature was probably about 5o C cooler, the precipita-tion about 50 percent lower (Haffer and Prance 2001),the declivity greater (Müller et al. 1995) and the flood-plain area considerably smaller than today (Irion pers.comm.). However, the flood pulse continued to bemonomodal and predictable with respect to dry andrainy seasons and there was sufficient floodplain arealeft to guarantee survival of flood-adapted plant andanimal species. Despite the change in environmentalconditions in the Amazon basin, basic structures andfunctions of the large river floodplains situated northand south of the equator were comparatively littleaffected and extinction rates were low. In comparison,during the ice ages, the Pantanal of Mato Grosso, about2 500 km south of the equator, suffered from extreme-ly dry periods that eliminated most of the flood-adapt-ed plant and animal species.
Today’s wetland conditions became reestab-lished in the Pantanal only about 7 000 years ago andwetland organisms of the lower Paraguay River, thesurrounding Cerrado and Amazonia colonized the area(Ab’Saber 1988). Mobile animals, such as birds, whichare very diverse in the Pantanal, were most efficient at
approaches and applications - an update 125

colonization. However, pronounced annual and pluri-annual droughts in combination with frequent wild-fires led to additional stress for plants and animals. Abroad ecological amplitude was a better survival strat-egy for the organisms than adaptation to specific wet-land conditions, as shown by trees that occur over alarge range of habitats. The number of total species andthe level of adaptation are comparatively low andendemic species are rare because the time span afterthe dry glacial period was too short for genetic diversi-fication (da Silva et al. 2001). This holds true evenwith respect to genera that show high diversificationrates, as for instance, the tree genus Inga. Most of the300 species have developed in the last 2 million years(Richardson et al. 2001).
During the ice ages, European and NorthAmerican river floodplains suffered even larger cli-matic changes. The temperature was lower and gla-ciers covered most of Northern Europe and NorthAmerica. The discharge regime of the large rivers wasdetermined by snow and ice melt. The light-tempera-ture pulse strongly superimposed on the impact of theflood pulse. Today’s wetlands of these areas began todevelop about 10 000 years ago with deglaciation andthere was very little time for organisms to adapt to thenew conditions in the floodplains. However, during theice ages, North American floodplain species couldmigrate to a certain extent to lower latitudes and laterrecolonize the newly formed wetlands, an option thatwas blocked in northern Europe by high mountains(Alps and Pyrenees), which explains the relativelysmall number and low level of adaptations of organ-isms to flooding in the European floodplains. Theseexamples also illustrate the influence of the time scaleto the degree of specialization and the development offlood-adapted communities.
CONNECTIVITY AND LENTIC-LOTIC LINKAGES
Amoros and Roux (1988) introduced the tech-nical term “connectivity” from landscape ecology tolimnology in order to describe the level of connectionof the mainstream with floodplain lakes. Connectivitylevels vary from permanent connection to short-termconnection during extreme floods (Ward, Tockner, and
Schiemer 1999; Wantzen and Junk 2000). Withdecreasing connectivity, the impact of the river onfloodplain lakes diminishes and lakes develop theirown limnological characteristics. For the AustrianDanube River floodplain near Regelsbrunn, Tockner etal. (1999) have shown that species number and com-munity structure of many aquatic organisms changedepending on the connectivity level. The quality ofconnectivity changes when, at very high water levels,floodplain lakes change from water storage to watertransport systems, i.e. from a lentic to a lotic system(limnophase and potamophase, sensu Neiff 1990).Strong flow pulses (Puckridge et al. 1998) may lead todramatic resetting of the limnetic succession by clean-ing the lake of accumulated organic debris and pro-foundly modifying aquatic plant and animal communi-ties (Marchese, Escurra de Drago and Drago 2002). Onthe other hand, the establishment and the cessation offlow conditions are crucial for the oxygen budget indetritus-rich floodplains. Two processes becomeimportant: 1) When terrestrially accumulated organicmatter becomes flooded and decays, large amounts ofoxygen can be consumed, as shown for tropical flood-plains (Braum and Junk 1982; Junk, Soares andCarvalho 1983; Sabo et al. 1999b), which eventuallycause fish kills (Hamilton et al. 1997) and 2) whenflow ceases in floodplain water bodies, high-water-level stratification and hypoxia occur (Melack andFisher 1983; Junk et al. 1983; Sabo et al. 1999b),which affects aquatic organisms.
Very large floodplains have complex connec-tivity patterns. For example, lakes can become con-nected to the mainstream by other lakes. In this case,migrating aquatic organisms have free access to lakesand the river; however, input of dissolved and suspend-ed matter is concentrated in the lake near the river andis low in the remote lake, which maintains lacustrineconditions. Tributaries with different water quality cancause hydro-chemical disconnection of floodplainlakes, as shown in the Ria Lakes formed by clear waterand black water tributaries in the Amazon River flood-plain. These lakes can be permanently connected to theAmazon River, which transports white water. Whenthe water level falls, the black and clear water of the
126 The flood pulse concept: New aspects,

tributaries advances to the lake mouth; when the waterrises, the white water of the mainstream represses theriver water and dominates part of the lake. The mobilefrontier between river water and the water of the tribu-taries can become a hydro-chemical barrier for aquaticorganisms despite the hydrological connection, asshown by the growth of aquatic macrophytes and theoccurrence of water snails and bivalves that concen-trate in the whitewater-influenced area because of bet-ter nutrient and calcium supply and higher pH values(Junk, unpubl.). Detailed studies on the impact of thehydrological and hydro-chemical connectivity level onflora and fauna in tropical river floodplain systems arestill lacking.
Lentic-lotic linkages have so far been consid-ered mainly in interconnected rivers and lakes wherelentic and lotic conditions alternate along the continu-um of the river course. Similar linkages exist in river-
floodplains systems; however, they occur in a tempo-ral dimension (syntopically during different hydrolog-ical periods) rather than in a spatial dimension (syn-chronically at different sites Figure 3). Adopting thisperspective, spatially scaled processes in weakly ornon-pulsing systems (e.g. regulated rivers) can be con-sidered analogous to temporal scales in pulsing sys-tems. For example, regulated lakes and rivers are strat-ified into profundal and littoral zones all year round.Processes such as open-water plankton production andshallow-site plankton filtering by benthos are spatiallyseparated, but linked by the water movement. In river-floodplain systems, both processes can occur at thesame site, but at different water levels. The moving lit-toral follows the rising or sinking water level in theATTZ. In the same way, infralittoral and profundalzones migrate along the flooding gradient, providedthat the depth is sufficient. When water levels recede,the functional units of the deep water disappear infloodplain systems (Figure 3).
approaches and applications - an update 127
Figure 3. Schematic comparison between hydrologically stabilized (left) and pulsing (right) aquatic ecosystems at normal water lev-els (0), extremely high water levels (+1) and extremely low water levels (-1). Left: in the stabilized system (regulated lake or river or nat-ural water body without floodplain), water level 0 prevails most of the time, allowing the establishment of well defined littoral (L), pelagial(O) and profundal (P) communities that are well adapted to these environmental conditions and are optimized in using the locally occur-ring resources. Occasional extreme floods are catastrophic events that do not allow the use of the resources of the flooded epilittoral byflora and fauna. Right: in pulsing systems, the organisms are adapted to periodically changing water levels and profit from resources ofvarying origin. Flora and fauna move along the flooding gradient; therefore, the same place in the ecosystem can harbor littoral and pro-fundal communities at different water levels. Not shown in the graph: Extreme low water levels urge the profundal and pelagial organ-isms either to migrate into deeper water bodies or to estivate at the sites, whereas terrestrial (epilittoral) species have developed survivalstrategies during flooding.

TIMING AND SHAPE OF THE FLOOD PULSE
The FPC has drawn attention to the importanceof the timing of the flood pulse and the stage of the lifecycle of the organisms, but for many floodplains, dataare still insufficient for detailed predictions. Manyfloodplain organisms have a “physiological and pheno-logical window of susceptibility” to the benefits anddisturbances of the flooding. The timing decideswhether an organism can profit from the flood-borneresources or apply survival strategies or not. Winterflooding does not have such deleterious effects to non-flood-adapted trees as flooding during summer whentrees are physiologically fully active. Similarly, unpre-dicted winter flooding had no significant effect onfloodplain-carbon uptake by fish in a river in the USA(Delong et al. 2001), whereas predicted timely flood-ing in the Pantanal did (Wantzen et al. 2002). Most fishspecies of the upper Paraná River have adapted theirspawning to the flood pulse and are affected by themany reservoirs that in addition to interrupting con-nectivity between river reaches, modify timing andshape of the pulse. These changes influence spawningbehavior and affect recruitment success of somespecies but also affect community structure, forinstance by increased predator pressure (Agostinho etal. 2000; Agostinho, Gomes and Zalewski 2001).
A slowly rising water level of the AmazonRiver leads to interruption and/or delay of spawningmigration of many migrating fish species and inextreme conditions to gonad absorption (Junk pers.obs.). Different flood patterns lead to different macro-phyte assemblages, which in turn are important habi-tats and food sources for many fishes (Petr 2000). Theeffects of different flood patterns on fish populationshave been summarized by Welcomme and Halls(2001).
The impact of human induced hydrologicalchanges has been shown for seedling establishment ofpoplar (Populus spp.) in North American rivers (Roodand Mahoney 1990). Timing of floods for the manage-ment of grasses and herbs for ducks and geese is amajor tool in polders along the Mississippi River
(Fredrickson and Reid 1988; Reid et al. 1989).Comparative studies on aquatic macrophytes andwater birds in the central Amazon River floodplain andthe Pantanal of Mato Grosso point to the importance ofthe amplitude of the flood pulse for species composi-tion and life forms. In the Amazon River flood plain, ahigh flood amplitude of up to 15 meters hinders thegrowth of submersed plants and probably also the fooduptake of some wading birds. Both groups occur inlarge abundance and species numbers in the Pantanalof Mato Grosso, where the flood amplitude is only 1-3meters (Junk and Petermann, unpubl. data). However,for most plant and animal species and communitiessuch information is still missing. Considering theincreasing man-made changes of river discharge, stud-ies are required for a better understanding of theimpact of the quality of the flood pulse on the biota.
EXTREME CLIMATIC EVENTS AND GLOBAL CHANGE
The effect of extreme hydrologic and climaticevents on river-floodplain systems has been stressedby the FPC, but has been little studied. Long-term anddeep flooding affect the ecosystem through profoundlevels of hydraulic energy and/or by physiologicalstress. Studies on streams show that 80 percent of theannual transport of particulate organic matter canoccur during a single extreme flooding (Cummins etal. 1983; Hobbie and Likens 1973). Such an eventreshapes the entire channel bed and the floodplain ofrivers in mountainous regions, such as the TagliamentoRiver in the Alps (Arscott, Tockner and Ward 2000)and also strongly modifies the floodplain of lowlandrivers (Sparks, Nelson and Yin 1998).
Pluriannual dry and wet periods can have long-lasting effects on community structure in floodplains.The long flood period of the Amazon River in thebeginning of the 1970s led to the dieback of manyfloodplain trees in low-lying areas. These areas stillhave not yet been recolonized by trees; a pluriannualdry period is required for successful reestablishment invery low lying areas on the flood gradient (Junk 1989).The spread of Vochysia divergens, a flood-tolerant treespecies, in the Pantanal of Mato Grosso during the last30 years has been associated with a long-lasting wet
128 The flood pulse concept: New aspects,

period after a pluriannual dry period in the beginningof the 1960s (Nunes da Cunha and Junk unpublished).Fish catches in the central delta of the Niger Riverdeclined from 90 000 tonnes yr–1 to 45 000 tonnes yr–1
because of little rainfall in the 1980s (Lae 1994).
The study of the impacts of extreme climaticevents will be crucial for wetland ecosystem manage-ment and protection strategies. The IPCC (2001) indi-cates that the planet Earth will suffer considerable cli-mate changes during the next century, which will be, toa considerable extent, the result of a man-madeincrease in greenhouse gases, such as carbon dioxideand methane. A global average temperature increase of1.4 to 5.8
oC is predicted. Nearly all land masses, main-
ly those at northern high latitudes during the cold sea-son, will warm more rapidly than the global average.Global mean sea level is projected to rise by 0.09 to0.88 m because of temperature-related expansion ofthe water and melting of the glaciers of the northernpolar regions and high mountains. Changes in precipi-tation will occur in most regions - rainfall will increasein some regions and drought will increase in others.The strongest impact will be felt in northern sub-polarregions (permafrost regions), high mountains, coastalareas, deserts and savannas, where water is already alimiting factor. In many river floodplains, man-induced changes of hydrology, pollution and wetlanddestruction will be more important than the effects ofclimate change (Vörösmarty 2002), but extreme climatechange events will overlap with other human-inducedmodifications and aggravate the situation.
BIODIVERSITY
As stated by the FPC and other authors, flood-plains are hot spots of species diversity (Gopal andJunk 2000). They harbour not only many wetland-spe-cific plants and animals, but also many species fromadjacent terrestrial and deep-water habitats that canhave fundamental impacts on structures and functionsof floodplains. For instance, terrestrial plant speciessubstantially contribute to habitat diversity; primaryproduction and nutrient cycles and terrestrial ungulatesaffect plant community structure and increase second-ary production. However, inventories of floodplain
species are rare and incomplete because they requireinterdisciplinary approaches (Gopal, Junk and Davies2000, 2001).
One aspect of flooding is a variably strong dis-turbance that can modify or even reset environmentalconditions in the system. Therefore, the FPC has inte-grated the tenets of the intermediate-disturbancehypothesis (Connell 1978; Ward and Stanford 1983b)by predicting that floodplain areas with an intermedi-ate (and predictable) level of flooding are expected toprovide the highest diversity. The two extremes for thedisturbance-diversity relationship for a given flood-plain habitat are, therefore, (1) frequent-to-permanentchanges in the physical habitat structure caused byflooding (e.g. rainfall-driven floodplain habitats inlow-order streams) and (2) low number or lack ofhydrological changes with a continuous ecologicalsuccession of species, leading to a climax community(e.g. remote floodplain lakes during terrestrialization).By interrupting ecological succession in some patches,flooding causes the development of a mosaic of differ-ent successional stages at the same time on a small spa-tial scale. The intensity of multi-year wet and dry phas-es in floodplains, however, can provide additionalstressors. In the Pantanal of Mato Grosso, for instance,the occurrence of numerous life forms is limited by theextreme desiccation, combined with fires during thedry phase (Nunes da Cunha and Junk 2001; da Silva etal. 2001).
In riverine floodplains, hydrological variationshapes a high diversity of physical habitat structuresthat might be more heterogeneous across the flood-plain than along the main channel (Marchese andEzcurra de Drago 1992; Arscott et al. 2000), thus cre-ating the basis for a diverse flora and fauna. InAmazonia, forest diversity is related to river dynamics(Salo et al. 1986). However, flooding and drought canalso reduce spatial heterogeneity by linking aquaticpopulations that were separated in different water bod-ies during the low-water period (vice versa, isolatedterrestrial populations during a flooding period canmix genetically during drought). Similarly, the perma-nent drift of organisms from the catchment or upriver
approaches and applications - an update 129

areas inoculates the riverine or near-river populationsregularly and thus hinders the development of geneti-cally distinct populations.
Connectivity between the main channel and thefloodplain habitat has become a central theme in thebiodiversity debate (Ward, Tockner and Schiemer1999; Wantzen and Junk 2000; Amoros and Bornette2002). Lateral connectivity has been suggested todetermine the diversity patterns of many taxonomicgroups directly (Tockner et al. 1999). Flood-pulsingsystems encounter variable degrees and spatiotemporalpatterns of connectivity. Therefore, the diversity ofhydrological patterns is a key element for the mainte-nance of habitat and species diversity in river-flood-plain systems.
BIOGEOCHEMICAL CYCLES
According to the FPC, river floodplains can beconsidered as biogeochemical reactors that temporari-ly store and process organic and inorganic matter. Theflood pulse exerts hydraulic forces that erode, carryand deposit these substances. Long-term storage favorsin situ alteration, weathering and liberation of dis-solved substances, as shown for an Amazonian Várzealake (Weber, Furch and Junk 1996; Irion, Junk and deMello 1997). The water level fluctuations provokechanges in water chemistry by mixing water bodiesand resource input during the rising limb of the hydro-graph and by increasing stratification, oxygen con-sumption and concentration of ions in the restrictedwater bodies during the falling limb. In floodplainsthat widely dry out periodically, like the Pantanal, alarge part of the organic matter is turned over duringthe change of the hydrological phases.
Periodic flooding and drought of sedimentsleads to sequential occurrence of different redoxprocesses. For example, organisms like cyanobacteriaand legumes fix atmospheric nitrogen, but the changebetween anoxic and oxic conditions during the water-land transition and the availability of large amounts oforganic material favor denitrification (Kern, Darwich,Furch, et al. 1996; Kern and Darwich 1997).Wassmann and Martius (1997) estimate the methane
production of the Amazon River floodplain at 1-9 TgCH4 yr-1, corresponding to 1-8 percent of the globalsource strength of wetlands. High primary productionleads to considerable pulses in carbon dioxide uptakeand release, but also to carbon storage in the sedimentand carbon export to the oceans. About 10
14g of organ-
ic carbon is annually transported by the Amazon Riverto the Atlantic Ocean (Ritchey et al. 1980). A consid-erable part of it may derive from the floodplain (Junk1985).
Junk (1980) points to an underestimate of thetotal wetland area in tropical South America becausesmall wetlands are often not considered in inventories,although they might comprise about 50 percent of thetotal wetland area, most of them floodplains. Thismight also hold true to some extent for other tropicaland subtropical regions. We hypothesize that thesesmall floodplains and temporary wetlands also followthe predictions of the FPC. Mapping of these areas andinclusion of their impact on the budgets of biogeo-chemical cycles and the hydrological cycle and formaintenance of biodiversity are challenges for thefuture.
SUSTAINABLE MANAGEMENT AND RESTORATION OF
RIVER FLOODPLAINS
River floodplains have provided multiple bene-fits since early human settlement. Predictable floodsfavored the management of floodplain resources andthe development of ancient cultures, for example, onthe Euphrates and Tigris Rivers and the Nile River sev-eral thousand years ago. Pre-Columbian human densi-ty in the floodplain of the Amazon River was severaltimes higher than that in the adjacent upland. Rice cul-tivation started in China about 7000 years ago (Boulé1994) and continues to be the nutritional basis formuch of the human population worldwide.
The economic value of floodplains for buffer-ing extreme hydrological events has been underesti-mated for a long time. A dramatically increasinghuman population during the last two centuries led tolarge-scale floodplain destruction and deteriorationworldwide (Junk 2002, Tockner and Stanford 2002). In
130 The flood pulse concept: New aspects,

the past, large flood events led to heavy losses of goodsand humans in Europe and brought about major floodcontrol measures, such as the “correction” of the RhineRiver by Tulla in the nineteenth century (Friedrich andMueller 1984). The 500-year-old European tradition inriver regulation (Nienhuis et al. 1998) was first trans-ferred to North America and later applied worldwide.Only some decades ago did the negative ecological,economic and social side effects of floodplain destruc-tion become apparent, as recently shown by the cata-strophic floods along the Odra, Elbe, Rhine andDanube Rivers in 2001 and 2002 in Poland, the CzechRepublic, Germany and Austria.
Management plans are required for the sustain-able use of floodplain resources. The FPC providesgeneral outlines that can be used for the developmentof management strategies; however, consideringregional differences in the status of floodplain integri-ty, watershed management and demographic and eco-nomic development, there is a need for specific strate-gies for each floodplain and even for different stretch-es of large river floodplains. For instance, the impor-tance of floodplains for protein supply by fisheries islow in most Central European rivers; however, in mosttropical countries, floodplain fishery provides accessi-ble animal protein for millions of people and is one ofthe most important economic activities (Welcomme1985).
Knowledge on wetland restoration has beenincreasing rapidly for several decades and ambitiousrestoration projects are being undertaken in NorthAmerica and Europe (Mitsch and Jørgensen 2003).Some restoration projects have also been started in thetropics. These projects are excellent means of validat-ing predictions of the FPC, as shown by Heiler et al.(1995) for the Danube River. Creating and maintainingnatural variation of the pulsing hydrograph and theability of the landscape to develop a dynamic flood-plain appear to be the most important elements for con-servation and restoration concepts (Sparks et al. 1990,1998; Tockner et al. 1999).
CONCLUSIONS
Most freshwater systems are subjected to fluc-tuations in water levels. All systems that are notsteeply bordered by mountains, dykes, or regulatingchannels are fringed with floodable areas. Flooding iscontrolled by climate type (catchment rainfall patternsand evapotranspiration), landscape morphology(declivity and connectivity) and local effects (log jams,tributary inflows, recent local precipitation). With theknowledge of these variables, inundation-durationcurves can be plotted, as for instance, for a US coastalplain river (Benke et al. 2000), for a small alpine river(Arscott et al. 2000) and for the Pantanal wetland(Hamilton et al. 1996). This general pattern makes thecentral tenet of the FPC - that hydrological pulsing isthe driving force for the performance of organisms andfor patterns of ecological processes - a unifying themein limnological conceptualization.
Today, 17 years after its first presentation, theFPC is widely accepted and applied by most river ecol-ogists. It provides a conceptual framework for bothresearch and management in river-floodplain systems.Several researchers have refined its tenets. Even inupstream areas, unpredictable flood pulses can be prof-itable for the stream community (Wantzen and Junk2000), but this does not seem to be the case for regu-lated large rivers (Thorp et al. 1998). The characteris-tics of the pulse shape are crucial for the establishmentand survival of many aquatic organisms (Welcommeand Halls 2001; Wagner and Schmidt, unpublishedmanuscript). Flood pulses homogenize water qualityand habitat structure of formerly isolated water bodies(Marchese and Escurra de Drago 1992; Heckman1994; Tockner et al. 2000). It has also become clearthat there is no “either/or” distribution between pro-ductivity in the catchment, the river channel and thefloodplain, but rather a variable combination of thesethree sources for the food webs of the river-floodplaincontinuum (Figure 3).
Recent studies have shown that predictions ofthe FPC on the development of adaptations and sur-vival strategies of organisms have to be adjusted by
approaches and applications - an update 131

additional information on paleoclimatological history(Adis and Junk 2002). The interaction of the floodpulse with other environmental variables, such as thelight/temperature pulse, snow melting and prolongedice cover in high-latitude floodplain systems and rainyand dry seasons in arid regions, is not sufficientlyunderstood (Humphries et al. 1999). Also, the impactsof short- and long-term changes of the quality of theflood pulse on life history of organisms, communitiesand biogeochemical processes require additional stud-ies. The FPC also makes predictions about organismsand processes during the terrestrial phase at low waterperiods (Adis and Junk 2002; Parolin et al. in prep)that require additional studies. New techniques, suchas stable isotope determination, remote sensing, genet-ic tests and techniques for wetland restoration andmanagement provide powerful tools to test and refinethe FPC further.
ACKNOWLEDGEMENTS
We thank Rüdiger Wagner (MPIL-Schlitz) fordiscussions on the “phenological and physiologicalwindow of susceptibility”, Sabine Meier for editing themanuscript, Elke Bustorf for graphic design and KarenA. Brune for correction of the English. This paper isdedicated to the memory of Professor JürgenSchwoerbel (1927-2002), one of the pioneers in mod-ern running-water ecology.
132 The flood pulse concept: New aspects,
REFERENCES
Ab’Saber A.N. 1988. O Pantanal Mato Grossense e ateoria dos refugios. Revista Brasileira deGeographia, 50: 9-57
Adis J. 1984. Seasonal Igapo-forests of CentralAmazonian blackwater rivers and their terres-trial arthropod fauna. In The Amazon.Limnology and landscape ecology of a mightytropical river and its basin. pp. 245-268,Dordrecht, W. Junk Publishers.
Adis J. & Junk W.J. 2002. Terrestrial invertebrates inhab-iting lowland river floodplains of CentralAmazonia and Central Europe: A review.Freshwater Biology, 47: 711-731.
Adis J., Marques M.I., Wantzen K.M. 2001. First obser-vations on the survival strategies of terricolousarthropods in the northern Pantanal wetland ofBrazil - scientific note. Andrias, 15: 127-128.
Agostinho A.A., Gomes, L.C. & Zalewski M. 2001. Theimportance of floodplains for the dynamics offish communities of the upper river Paraná.Ecohydrology & Hydrobiology, 1: 209-217.
Agosthinho A.A., Thomaz S.M., Minte-Vera C.V. &Winemiller K.O. 2000. Biodiversity in the highParaná River floodplain. In B. Gopal, W.J.Junk & J.A. Davis eds. Wetlands Biodiversity.pp. 89-118. Leiden, The Netherlands,Backhuys Publishers.
Amoros C. & Bornette G. 2002. Connectivity and bio-complexity in waterbodies of riverine flood-plains. Freshwater Biology, 47: 761-776.
Amoros C. & Roux A.L. 1988. Interaction between waterbodies within the floodplains of large rivers:Function and development of connectivity.Münstersche Geographische Arbeiten, 29:125-130.

Arscott D.B., Tockner K. & Ward J.V. 2000. Aquatichabitat diversity along the corridor of anAlpine floodplain river Fiume Tagliamento,Italy. Archiv für Hydrobiologie, 149: 679-704.
Bayley P.B. 1980. The limits of limnological theory andapproaches as applied to river-floodplain sys-tems and their fish production. TropicalEcology & Development, 739-746.
Bayley P.B. 1991. The flood-pulse advantage and therestoration of river-floodplain systems.Regulated Rivers: Research & Management,6: 75-86.
Benke A.C., Chaubey I., Ward G.M. & Dunn, E.L. 2000.Flood pulse dynamics of an unregulated riverfloodplain in the southeastern US coastal plain.Ecology, 81: 2730-2741.
Boulé M.E. 1994. An early history of wetland ecology. InGlobal wetlands: Old world and new. pp. 57-74.
Braum E. & Junk W.J. 1982. Morphological adaptation oftwo Amazonian characoids Pisces for survi-ving in oxygen deficient waters. Int. Revue.Ges. Hydrobiol., 67: 869-886.
Bretschko G. & Leichtfried M. 1988. Distribution oforganic matter and fauna in a second order,alpine gravel stream Ritrodat-Lunz study area,Austria. Verh. Int. Verein. Limnol., 23: 1333-1339.
Clark J.R. & Benforado J. 1981. Wetland of bottomlandhardwood forest. Dev. Agric. Managed-ForestEcol. 11: Amsterdam, Oxford, New York,Elsevier Scientific Publishing Company.
Connell J.H. 1978. Diversity in tropical rain forests andcoral reefs. Science, 199: 1302-1309.
approaches and applications - an update 133
Cummins K.W., Sedell J.R., Swanson F.J., Minshall S.G.,Fisher S.G., Cushing C.E., Petersen R.C. &Vannote R.L. 1983. Organic matter budgets forstream ecosystems: Problems in their evalua-tion. In J.R. Barnes & G.W. Minshall eds.Stream ecology: Application and testing ofgeneral ecological theory. New York, PlenumPress. pp. 299-353.
Da Silva C.J., Wantzen K.M., Nunes da Cunha C. &Machado F.A. 2001. Biodiversity in thePantanal Wetland, Brazil. In Biodiversity inwetlands: Assessment, function and conserva-tion, Volume 2. pp. 187-215. Leiden, TheNetherlands, Backhuys Publishers.
De Oliveira M.D. & Calheiros D.F. 2000. Flood pulseinfluence on phytoplankton communities ofthe south Pantanal floodplain, Brazil.Hydrobiologia, 427: 101-112.
De Lamônica-Freire E. & Heckman C.W. 1996. The sea-sonal succession of biotic communities in wet-lands of the tropical wet-and-dry climatic ZoneIII: The algal communities in the Pantanal ofMato Grosso, Brazil, with a comprehensive listof the known species and revision of twodesmid taxa. Int. Rev. Ges. Hydrobiol., 81:253-280.
Delong M.D., Thorp J.H., Greenwood K.S. & MillerM.C. 2001. Responses of consumers and foodresources to a high magnitude, unpredictedflood in the upper Mississippi River Basin.Regulated Rivers: Research & Management,17: 217-234.
Dettmers J.M., Wahl D.H., Soluk D.A. & Gutreuter S.2001. Life in the fast lane: Fish and foodwebstructure in the main channel of large rivers.Journal of the North American BenthologicalSociety, 20: 255-265.
Dudgeon D. 2000. The ecology of tropical Asian riversand streams in relation to biodiversity conser-vation. Annual Review of Ecology andSystematics, 31: 239-263.

Dynesius M. & Nilsson C. 1994. Fragmentation and flowregulation of river systems in the northern 3rd
of the world. Science, 266: 753-762.
Elwood J.W., Newbold J.D., O’Neill R.V. & van WinkleW. 1983. Resource spiralling: An opearationalparadigm for analyzing lotic systems. In T.D.Fontaine III & S.M. Bartell eds. Dynamics oflotic ecosystems. Ann Arbor, MI, USA, AnnArbor Science Publishers. pp. 3-27.
Fisher T.R. & Parsley P.E. 1979. Amazon lakes: Waterstorage and nutrient stripping. Limnol.Oceanogr. L 24: 547-553.
Fredrickson L.H. & Reid F.A. 1988. Waterfowl use ofwetland complexes. U.S. Fish Wildlife Leaflet13.2.1. 6 pp.
Friedrich G. & Mueller D. 1984. Rhine West Germany. InEcology of European Rivers. pp. 265-316. ,Palo Alto, CA, USA, Blackwell ScientificPublications.
Frissell C.A., Liss W.J., Warren C.E. & Hurley M.D.1986. A hierarchical framework for streamhabitat classification: Viewing streams in awatershed context. EnvironmentalManagement, 10: 199-214.
Furch K. 1984. Interannuelle variation hydrochemischerparameter auf der Ilha de Marchantaria.Biogeographica, 19: 85-100.
Galat D.L. & Zweimuller I. 2001. Conserving large-riverfishes: Is the highway analogy an appropriateparadigm? Journal of the North AmericanBenthological Society, 20: 266-279.
Gopal B. & Junk W.J. 2000. Biodiversity in wetlands: Anintroduction. In Biodiversity in Wetlands:Assessment, function and conservation,Volume 1. pp. 1-10. Leiden, The Netherlands,Backhuys Publishers.
134 The flood pulse concept: New aspects,
Gopal B., Junk W.J. & Davies J.A. 2000. Biodiversity inwetlands: Assessment, function and conserva-tion, volume 1. 353 pp. Leiden, TheNetherlands, Backhuys Publishers.
Gopal B., Junk W.J. & Davies J.A. 2001. Biodiversity inWetlands: assessment, function and conserva-tion, volume 2. 311 pp. Leiden, TheNetherlands, Backhuys Publishers.
Gutreuter S., Bartels A.D., Irons K. & Sandheinrich M.B.1999. Evaluation of the flood-pulse conceptbased on statistical models of growth of select-ed fishes of the Upper Mississippi River sys-tem. Canadian Journal of Fisheries andAquatic Sciences, 56: 2282-2291.
Haffer J. & Prance G.T. 2001. Climatic forcing of evolu-tion in Amazonia during the Cenozoic: On therefuge theory of biotic differentiation.Amazoniana, XVI3/4: 579-607.
Hamilton S.K., Sippel S.J. & Melack J.M. 1996.Inundation patterns in the Pantanal wetland ofSouth America determined from passivemicrowave remote sensing. Arch. Hydrobiol.,137: 1-23.
Hamilton S.K., Sippel S.J., Calheiros D.F. & Melack,J.M. 1997. An anoxic event and other biogeo-chemical effects of the Pantanal wetland on theParaguay River. Limnol. Oceanogr., 42: 257-272.
Heckman C.W. 1994. The seasonal succession of bioticcommunities in wetlands of the tropical wet-and-dry climatic zone I: Physical and chemicalcauses and biological effects in the Pantanal ofMato Grosso, Brazil. Int. Revue. Ges.Hydrobiol., 79: 397-421.
Heckman C.W. 1998. The seasonal succession of bioticcommunities in wetlands of the tropical wet-and-dry climatic zone: V aquatic invertebrate

communities in the Pantanal of Mato Grosso,Brazil. Int. Revue. Ges. Hydrobiol., 83: 31-63.
Heiler G., Hein T., Schiemer F. & Bornette G. 1995.Hydrological connectivity and flood pulses asthe central aspects for the integrity of a river-floodplain system. Regulated Rivers: Research& Management, 11: 351-361.
Hobbie J.E. & Likens G.E. 1973. The output of phospho-rous, dissolved organic carbon, and fine partic-ulate carbon from Hubbard Brook watersheds.Limnol. Oceanogr., 18: 734-742.
Humphries P., King A.J. & Koehn J.D. 1999. Fish, flowsand flood plains: Links between freshwaterfishes and their environment in the Murray-Darling River system, Australia.Environmental Biology of Fishes, 56: 129-151.
Illies J. 1961a. Versuch einer allgemeinen biozönotischenGliederung der Fließgewässer. Int. Revue Ges.Hydrobiol., 462: 205-213
Illies J. 1961b. Gebirgsbäche in Europa und inSüdamerika - ein limnologischer Vergleich.Verh. Internat. Verein. Limnol., 14: 517-523
Illies J. & Botosaneanu L. 1963. Problemes et methodesde la classification et de la zonationecologiques des eaux courantes, consideressurtout du point de vue faunistique. Mitt.Internat. Verein. Limnol., 12: 1-57.
IPCC 2001. Technical summary, and summary for policy-makers. Third Assessment Report of WorkingGroup I of the Intergovernmental Panel onClimatic Change. (Available at http://www.ipcc.ch)
Irion G., Junk W.J. & de Mello J.A.S.N. 1997. The largecentral Amazonian river floodplains nearManaus: Geological, climatological, hydrolog-ical, and geomorphological aspects. pp. 23-46.In Junk W.J. ed. The Central Amazon
approaches and applications - an update 135
Floodplain - Ecology of a Pulsing System.Springer Verlag, Berlin.
Irmler U. 1981. Überlebensstrategien von Tieren imsaisonal überfluteten Amazonischen Über-schwemmungswald. Zool. Anz. Jena., 2061/2:26.38.
Isaac V.J., Ruffino M.L. & McGrath D. 1998. In search ofa new approach to fisheries management in themiddle Amazon. In T.J. Quinn II, F. Funk, J.Heifetz, J. Ianelli, J. Power, J. Schweigert, P.Sullivan & C.I. Zhang eds. Fishery stockassessment model. pp. 889-902. AL, USA,Faibanks, Alaska Sea Grant College ProgramUniversity of Alaska, As-SG-98-01.
Junk W.J. 1980. Áreas inundáveis - um desafio para lim-nologia. Acta Amazonica, 104: 775-795.
Junk W.J. 1985. The Amazon floodplain - a sink or sourcefor organic carbon? Mitt. Geol. Paläont. Inst.Univ. Hamburg.
Junk W.J. 1989. Flood tolerance and tree distribution incentral Amazonian floodplains. In L. B. Holm-Nielsen, I.C. Nielsen & H. Balslev eds.Tropical forests: Botanical dynamics, specia-tion and diversity. 47-64. London, AcademicPress.
Junk W.J. 1999. The flood pulse concept of large rivers:Learning from the tropics. Arch. Hydrobiol.Suppl., 1153: 261-280.
Junk W.J. 2000. The central Amazon River floodplain:Concepts for the sustainable use of itsresources. In W.J. Junk, J.J. Ohly, M.T.F.Piedade & M.G.M. Soares eds. The CentralAmazon floodplain: Actual use and options forsustainable management. pp. 75-94. Leiden,The Netherlands, Backhuys Publishers.
Junk W.J. 2002. Long-term environmental trends and thefuture of tropical wetlands. EnvironmentalConservation, 294. (forthcoming)

Junk W.J. & Piedade M.T.F. 1997. Plant life in the flood-plain with special reference to herbaceousplants. In W.J. Junk ed. The Central Amazonfloodplain: Ecology of a pulsing system. pp.147-186. Berlin, Springer Verlag.
Junk W.J. & Weber G.E. 1996. Amazonian floodplains: Alimnological perspective. Verh. Internat.Verein. Limnol., 26: 149-157.
Junk W.J., Bayley P.B. & Sparks R.E. 1989. The floodpulse concept in river-floodplain systems. Can.Spec. Publ. Fish. Aquat. Sci., 106: 110-127.
Junk W.J., Soares G.M., Carvalho F.M. 1983. Distributionof fish species in a lake of the Amazon Riverfloodplain near Manaus Lago Camelao withspecial reference to extreme oxgen conditions.Amazoniana, 7: 397-431.
Junk W.J., Soares G.M. & Saint-Paul U. 1997. The Fish.In W.J. Junk ed. The Central AmazonFloodplain - ecology of a pulsing system. pp.385-408. Berlin, Springer Verlag.
Junk W.J., Ohly J.J., Piedade M.T.F. & Soares M.G.M.2000. The Central Amazon floodplain: Actualuse and options for sustainable management.Leiden, The Netherlands, BackhuysPublishers. 584 pp.
Kern J. & Darwich A. 1997. Nitrogen turnover in thevárzea. In Junk W.J. ed. The Central Amazonfloodplain - ecology of a pulsing system, 119-136. , Berlin, Springer Varlag.
Kern J., Darwich A., Furch K. & Junk W.J. 1996.Seasonal denitrification in flooded andexposed sediments from the amazon flood-plain at Lago Camalão. Microbial Ecology, 32:47-57.
Knowlton M.F. & Jones J.R. 1997. Trophic status ofMissouri River floodplain lakes in relation tobasin type and connectivity. Wetlands, 17: 468-475.
Lae R. 1994. Effects of drought, dams, and fishing pres-sure on the fisheries of the central delta of theNiger River. International Journal of Ecologyand Environmental Sciences, 20: 119-128.
Lewis W.M. Jr. 1988. Primary production in the OrinocoRiver Venezuela. Ecology, 69: 679-692.
Lewis W.M. Jr., Hamilton, S.K., Lasi, M.A., Rodríguez &Saunders M.A. III. 2000. Ecological determin-ism on the Orinoco floodplain. BioScience, 50:681-692.
Lowe-McConnell R.H. 1975. Fish communities in tropi-cal freshwaters: Their distribution, ecologyand evolution. London, New York, Longman.337 pp.
Lowe-McConnel R.H. 1987. Ecological studies in tropi-cal fish communities. Cambridge, UK,Cambridge University Press.
Lytle D.A. 2001. Disturbance regimes and life-historyevolution. American Naturalist, 157: 525-536.
Magrath, M.J.L. 1992. Waterbird study of the lowerLachlan and Murrumbidgee Valley wetlands in1990/1991. A report prepared for the NSWDepartment of Water Resources.
Marchese M. & Ezcurra de Drago I.E. 1992. Benthos ofthe lotic environments in the middle ParanaRiver system: Transverse zonation.Hydrobiologia, 237: 1-13.
Marchese M., Escurra de Drago I. & Drago E. 2002.Benthic macroinvertebrates and physical habi-tat relationships in the Paraná river-floodplainsystem. In M. McClain ed. InternationalAssociation of Hydrological Sciences. Specialpublication No. 6. (forthcoming)
McGrath D., de Castro F., Câmara E. & Futema C. 1999.Community management of floodplain lakesand the sustainable development ofAmazonian fisheries. Advances in EconomicBotany, 13: 59-82.
136 The flood pulse concept: New aspects,

Melack J.M. & Fisher T.R. 1983. Diel oxygen variationsand their ecological implications in Amazonfloodplain lakes. Arch. Hydrobiol., 98: 422-442.
Minshall G.W. 1985. Developments in stream ecosystemtheory. Journal of Fisheries and AquaticScience, 42: 1045-1055.
Mitsch W.J. & Gosselink J.G. 2000. Wetlands. New York,USA, John Wiley & Sons.
Mitsch W.J. & Jørgensen S.E. 2003. Ecological engineer-ing and ecosystem restoration. New York,John Wiley & Sons. (forthcoming)
Montgomery D.R. 1999. Process domains and the rivercontinuum. Journal of the American WaterResources Association, 35: 397-410.
Müller J., Irion G., Nunes de Mello J. & Junk W.J. 1995.Hydrological changes of the Amazon duringthe last glacial-interglacial cycle in CentralAmazonia Brazil. Naturwissenschaften, 82:232-235.
Naumann E. 1932. Principles of regional limnology.Stuttgart, Germany, Schweizerbart Publishers.149 pp.
Neiff J.J. 1990. Ideas for an ecological interpretation ofthe Paraná. Interciencia, 156: 424-441
Nienhuis P.H., Leuven, R.S.E.W & Ragas, A.M.J. 1998.New concepts for sustainable management ofriver basins. Leiden, The Netherlands,Backhuys Publishers. 367 pp.
Nunes da Cunha C. & Junk W.J. 2001. Distribution ofwoody plant communities along the flood gra-dient in the Pantanal of Poconé, Mato Grosso,Brazil. International Journal of Ecology andEnvironmental Sciences, 27: 63-70.
approaches and applications - an update 137
Parolin P., de Simone O., Haase K., Waldhoff D.,Rottenberger S., Kuhn V., Kesselmeier J.,Schmidt W., Piedade M.T.F. & Junk W.J.Central Amazonian floodplain forests: Treesurvival in a pulsing system. (In preparation)
Petermann P. 1999. Biogeographie einer Insel-Avifaunain der Várzea des mittleren Amazonas, amBeispiel der Ilha de Marchantaria. Universityof Saarland. (Doctoral dissertation)
Petr T. 2000. Interactions between fish and aquaticmacrophytes in inland waters: A review. FAOFisheries Technical Paper, 396: 1-185.
Petts G.E. & Amoros C. 1996. Fluvial hydrosystems.London, Chapman & Hall. 322 pp.
Pinay G., Decamps, H. & Naiman, R.J. 1999. The spiral-ing concept and nitrogen cycling in large riverfloodplain. Archiv. Für Hydrobiologie, 143:281-291.
Poff N.L. 1997. Landscape filters and species traits:Towards mechanistic understanding and pre-diction in stream ecology. Jr. N. Am. Benth.Soc., 16: 391-409.
Poff N.L. & Ward J.V. 1989. Implications of streamflowvariability and predictability for lotic commu-nity structure - a regional-analysis of stream-flow patterns. Canadian Journal of Fisheriesand Aquatic Sciences, 46: 1805-1818.
Poole G.C. 2002. Fluvial landscape ecology: Addressinguniqueness within the river discontinuum.Freshwater Biology, 47: 641-660.
Pringle C.M. 2001. River conservation in tropical versustemperate latitudes. In Global Perspectives onRiver Conservation: Science, Policy andPractice. pp. 373-383. New York, John Wiley& Sons Ltd.

Pringle C.M., Naiman R.J., Bretschko G., Karr J.R.,Oswood M.W. Webster, J.R., Welcomme R.L.& Winterbourn M.J. 1988. Patch dynamics inlotic systems: The stream as a mosaic. Jr. N.Am. Benth. Soc., 7: 503-524.
Puckridge J.T., Sheldon F., Walker K.F. & Boulton A.J.1998. Flow variability and the ecology of largerivers. Marine and Freshwater Research, 49:55-72.
Reid F.R., Kelley J.R. Jr., Taylor T.S. & Fredrickson L.H.1989. Upper Mississippi Valley wetlands,refuges and moist-soil impoundments. In L.Smith, R. Pederson, R. Kaminski eds. Habitatmanagement for migrating and witeringwaterfowl in North America. pp, 181-202.Lubbock TX, USA, Technical UniversityPress.
Resh V.H., Hildrew A.G., Statzner B. & Townsend C.R.1994. Theoretical habitat templets, speciestraits and species richness: A synthesis of long-term ecological research on the Upper RhoneRiver in the context of concurrently developedecological theory. Freshwater Biology, 31:539-554.
Resh V.H., Brown A.V., Covich A.P., Gurtz L.H.W.,Minshall G.W., Reicem S.R., Sheldon A.L.,Wallace J.B. & Wissmar R.C. 1988. The roleof disturbance in stream ecology. J. N. Am.Benth. Soc., 7: 443-455.
Richardson J.E., Pennington R.T., Pennington T.D. &Hollinsworth P.M. 2001. Rapid diversificationof a species-rich genus of neotropical rain for-est trees. Science, 293: 2242-2245.
Richey J.E., Brock J.T., Naiman R.J., Wissmar R.C. &Stallard R.F. 1980. Organic carbon: Oxidationand transport in the Amazon River. Science,207: 1348-1351.
Richter B.D., Baumgartner J.V., Powell J. & Braun D.P.1996. A method for assessing hydrologic alter-ation within ecosystems. ConservationBiology, 10: 1163-1174.
Robertson A.I, Bunn S.E., Boon P. & Walker K.F. 1999.Sources, sinks and transformations of organiccarbon in Australian floodplain rivers. Marine& Freshwater Research, 50: 813-829.
Rood S.B. & Mahoney J.M. 1990. Collapse of riparianpoplar forests downstream from dams in west-ern prairies: Probable causes and prospects formitigation. Environ. Manage., 14: 451-464.
Sabo M.J., Bryan C.F., Kelso W.E. & Rutherford A.1999a. Hydrology and aquatic habitat charac-teristics of a riverine swamp: I. Influence offlow on water temperature and chemistry.Regulated Rivers: Research & Management,15: 505-523.
Sabo M.J., Bryan C.F., Kelso W.E. & Rutherford A.1999b. Hydrology and aquatic habitat charac-teristics of a riverine swamp: II. Hydrologyand the occurrence of chronic hypoxia.Regulated Rivers: Research & Management,15: 525-542.
Salo J., Kalliola R.J., Hakkinen I., Makinen Y., NiemalaP., Puhaka M. & Coley P.D. 1986. Riverdynamics and the diversity of Amazon lowlandforest. Nature, 322: 254-258.
Schwoerbel J. 1961. Über die Lebensbedingungen unddie Besiedlung des hyporheischenLebensraumes. Arch. Hydrobiol. Suppl, 25:162-214.
Sedell J.R., Richey J.E., Swanson F.J. 1989. The rivercontinuum concept: A basis for the expectedecosystem behavior of very large rivers? Can.Spec. Publ. Fish. Aquat. Sci., 106: 49-55.
138 The flood pulse concept: New aspects,

Southwood T.R.E. 1977. Habitat: The template for eco-logical strategies. Journal of Animal Ecology,46: 337-365.
Sparks R.E., Nelson J.C. & Yin Y. 1998. Naturalization ofthe flood regime in regulated rivers.BioScience, 48: 706-720.
Sparks R.E., Bayley P.B., Kohlert S.L., Osborne L.L.1990. Disturbance and recovery of large flood-plain rivers. Environmental Management, 14:699-709.
Stanford A. & Ward J.V. 1988. The hyporheic habitat ofriver ecosystems. Nature, 335: 64-66.
Statzner, B. & Higler B. 1986. Stream hydraulics as amajor determinant of benthic invertebratezonation patterns. Freshwater Biology, 16:127-139.
Thienemann A. 1925. Die BinnengewässerMitteleuropas. Die Binnengewässer 1. ,Stuttgart, Germany, Schweizerbart Publishers.
Thorp J.H. & Delong M.D. 1994. The riverine productiv-ity model: An heuristic view of carbon sourcesand organic-processing in large river ecosys-tems. Oikos, 70: 305-308.
Thorp J.H., Delong M.D., Greenwood K.S. & Casper A.F.1998. Isotopic analysis of three food web the-ories in constricted and floodplain regions of alarge river. Oecologia, 117: 551-563.
Tockner K. & Stanford J.A. 2002. Riverine flood plains:Present state and future trends. EnvironmentalConservation, 293: 308-330.
Tockner K., Malard F. & Ward J.V. 2000. An extension ofthe flood pulse concept. HydrologicalProcesses, 14: 2861-2883.
Tockner K., Schiemer F., Baumgartner C., Kum G.,Weigand E., Zweimuller, I. & Ward, J.V. 1999.The Danube restoration project: Species diver-sity patterns across connectivity gradients inthe floodplain system. Regulated Rivers:Research & Management, 15: 245-258.
Townsend C.R. 1989. The patch dynamics concept ofstream community ecology. Jr. N. Am. Bentho.Soc., 8: 36-50.
Townsend C.R. 1996. Concepts in river ecology: Patternand process in the catchment hierarchy. Archivfür Hydrobiologie Supplementband, 113 1-4:3-21.
Townsend C.R. & Hildrew A.G. 1994. Species traits inrelation to a habitat templet for river system.Freshwater Biology, 31: 265-275.
Triska F.J., Duff J.H. & Avanzino R. 1993. Patterns ofhydrological exchange and nutrient transfor-mation in the hyporheic zone of a gravel-bot-tom stream: Examining terrestrial-aquatic link-ages. Freshwater Biology, 29: 259-274.
Van den Brink F.W.B., Beljaards M.J., Botts N.C.A. &van der Velde G. 1994. Macrozoobenthosabundance and community composition in 3Lower Rhine floodplain lakes with varyinginundation regimes. Regulated Rivers:Research & Management, 9: 279-293.
Van den Brink F.W.B., de Leeuw J.P.H.M., van der VeldeG. & Verheggen G.M. 1993. Impact of hydrol-ogy on the chemistry and phytoplankton devel-opment in floodplain lakes along the LowerRhine and Meuse. Biogeochemistry Dordrecht,19: 103-128.
Vannote R.L., Minshall G.W., Cummins K.W., SedellK.W., Cushing C.E. 1980. The river continuumconcept. Can. J. Fish. Aquat. Sci., 37: 130-137.
approaches and applications - an update 139

Vörösmarty C.J. 2002. Global water assessment andpotential contributions from Earth SystemsScience. Aquatic Sciences, 64: 328-351.
Wantzen K.M. & Junk W.J. 2000. The importance ofstream-wetland-systems for biodiversity: Atropical perspective. In B. Gopal, W.J. Junk &J.A. Davies eds. Biodiversity in Wetlands:Assessment, function and conservation, 11-34,Leiden, The Netherlands, BackhuysPublishers.
Wantzen K.M., Machado F.A., Voss M., Boriss H. & JunkW.J. 2002. Seasonal isotopic changes in fish ofthe Pantanal wetland, Brazil. Aquatic Sciences,64: 239-251.
Ward J.V. 1989. The four-dimensional nature of loticecosystems. Jr. N. Am. Bentho. Soc., 8: 2-8.
Ward J.W. & Stanford J.A. 1983a. The serial discontinu-ity concept of lotic ecosystems. In T.D.Fontaine III & S.M. Bartell eds. Dynamics oflotic ecosystems, Ann Arbor, MI, USA, AnnArbor Science Publishing. pp. 29-42.
Ward J.V. & Stanford J.V. 1983b.The intermediate-distur-bance hypothesis: An explanation for bioticdiversity patterns in lotic ecosystems. In T.D.Fontaine III & S.M. Bartell eds. Dynamics oflotic ecosystems, Ann Arbor, MI, USA, AnnArbor Science Publishing. pp. 347-356.
Ward J.V. & Stanford J.A. 1995. The serial discontinuityconcept: Extending the model to floodplainrivers. Regulated Rivers: Research &Management, 11: 105-119.
Ward J.V., Malard F. & Tockner K. 2001. Landscape ecol-ogy: A framework for integrating pattern andprocess in river corridors. Landscape Ecology,17: 35-45.
Ward J.V., Tockner K. & Schiemer F. 1999. Biodiversityof floodplain river ecosystems: Ecotones andconnectivity. Regulated Rivers: Research &Management, 15: 125-139.
Wassmann R. & Martius C. 1997. Methane emissionsfrom the Amazon floodplain. In W.J. Junk ed.The Central Amazon Floodplain - Ecology of aPulsing System. pp. 137-143. Berlin, Springer-Verlag.
Weber G.E., Furch K. & Junk W.J. 1996. A simple mod-elling approach towards hydrochemical sea-sonality of major cations in a CentralAmazonian floodplain lake. EcologicalModelling, 91: 29-56.
Welcomme R.L. 1979. Fisheries ecology of floodplainrivers. London, Longman. 317 pp.
Welcomme R.L. 1985. River fisheries., FAO FisheriesTechnical Paper 262. Rome, Food andAgricultural Organization of the UnitedNations. 330 pp.
Welcomme R.L. & Halls A. 2001. Some considerations ofthe effects of differences in flood patterns onfish populations. Ecohydrology andHydrobiology, 13: 313-321
Williams D.D. 1989. Towards a biological and chemicaldefinition of the hyporheic zone in twoCanadian rivers. Freshwat. Biol., 22: 189-208.
Winemiller K.O. 1989. Patterns of variation in life-histo-ry among South American fishes in seasonalenvironments. Oecologia, 81: 225-241.
Winterbourn M.J., Rounick, J.S. & Cowie, B. 1981. AreNew Zealand stream ecosystems really differ-ent? New Zealand Journal of Marine andFreshwater Research, 15, 321-328.
Wittmann F. 2002. Species distribution, community struc-ture, and adaptations of Amazonian várzeaforests depending on the annual flood-stress,using remote sensing methods. University ofMannheim. (Doctoral dissertation)
140
