the extravascular contractile system in the human placenta
TRANSCRIPT

Anat Embryol (1994) 190:541 548
Anato _my and Embryolo
�9 Springer-Verlag 1994
The extravascular contractile system in the human placenta Morphological and immunocytochemical investigations
R. Graf l, J.-U. Langer 1, G. Schiinfelder ~, T. Oney 2, S. Hartel-Schenk 3, W. Reutter 3, H.H.H.W. Schmidt 4 Institut flit Anatomie, Freie Universitfit Berlin, K6nigin-Luise-Strasse 15, D-14195 Berlin, Germany
2 Frauenklinik und Poliklinik, UK Steglitz, Freie Universitfit Berlin, Hindenburgdamm 30, D-12203 Berlin, Germany 3 Institut fiir Molekularbiologie und Biochemie, Freie Universit~it Berlin, Arnimallee 22, D-14195 Berlin, Germany 4 Institut ffir Biochemie und Pathobiochemie, Medizinische Universit~it Wfirzburg, Versbacher Strasse 5, D-97078 Wiirzburg, Germany
Accepted: 6 June 1994
Abstract. In the human placenta, besides the fetal blood vessel system a second extravascular contractile system exists. It is localized in the chorionic plate and runs in a longitudinal direction and adjacent to fetal blood vessels into the stem villi, where it forms perivascular contractile sheaths. Characteristically, cells of the extravascular con- tractile system are extremely long and spindle-shaped and give rise to fine cell processes, by which they obvious- ly contact each other or insert into the basement mem- brane of the trophoblast. They show immunoreactivity with desmin, vimentin, ~-actin, myosin, nitric oxide syn- thase type I (brain form) and dipeptidyl peptidase IV. The ultrastructure suggests that ceils of the extravascular contractile system are related to smooth muscle cells, in- cluding subpopulations with myofibroblastic features. In stem villi a few cells are nitric oxide synthase type I im- munoreactive. These cells are thought to be specialized smooth-muscle-like cells of the extravascular contractile system or cells of the extravascular contractile system related to paraneurons that generate nitric oxide, which, in turn, may modulate the tone of perivascular contrac- tile sheaths. The high dipeptidyl peptidase IV activity suggests that modulation of the extravascular contractile system may also occur by substance P.
Key words: Human placenta Smooth muscle-like cells - Perivascular contractile sheath Nitric oxide synthase type I - Dipeptidyl peptidase IV
Introduction
In 1932, Dubreui! and Rivi6re reported, on the basis of morphological studies, an extravascular cellular system in the human placenta, which they presumed to be con- tractile. They described it as a plexiform network that is formed by a fibro-muscular layer in the chorionic plate and "axes fibro-musculaires" running in stem and inter-
Correspondence to: R. Graf
mediate villi. However, in the same paper they noted that this system, although it was described as early as 1906 by Happe, obviously did not attract the attention of other authors. A very detailed description of the extravascular presumed smooth muscle was given by Spanner (1936), who summarized his findings in a coloured drawing of the placenta. Twenty years later, in 1963, Krantz and Parker confirmed the contractility of the extravascular system by physiological studies, followed by King and Gr6schel-Stewart (1965) and Huszar and Bailey (1979a, b) who used biochemical methods.
The functions of the human placenta under normal and pathological conditions are still not fully under- stood. Current studies deal with the modulation of utero- placental and fetoplacental blood flow, focusing on the blood vessel systems. With the exception of a few studies (stem and terminal villi: Feller et al. 1985'; chorionic plate and villi: Beham et al. 1988; stem villi: Demir et al. 1992; Graf et al. 1992, 1993; Kohnen et al. 1993), most recent investigations, however, neither discuss nor even mention that in the placenta, besides the fetal blood vessel system, there exists a second extravascular contractile system as described above. Even the ultrastructural demonstration of contractile cells in the chorionic plate by Bertolini et al. (1969) has been refuted (Becker et al. 1981). Du- breuil and Rivi+re (1932), Spanner (1936) and Krantz and Parker (1963) had already suggested that contraction of the extravascular contractile system might lead to au- tonomous movement of the placenta, thus affecting ma- ternal blood flow in the placental lobules and fetal blood flow in a propulsive way.
Systematic studies investigating the presence of inter- mediate and contractile filaments in these cells, the na- ture of the extravascular contractile cells', their hierarchy in the chorionic plate and placental villous tree and their regulation and function are still missing. Therefore, we reinvestigated the extravascular contractile system (EVCS) in parts of the chorionic plate and in stem villi, by using morphological, immunocytochemical and enzyme histochemical methods. The study covered contractile and intermediate filaments, dipeptidyl peptidase IV (DPP

542
IV, E C 3.4.14.5) a n d n i t r i c o x i d e s y n t h a s e t y p e I ( N O S I, b r a i n fo rm) . D P P IV, w h i c h c l eaves s u b s t a n c e P, a n d N O S I I I ( e n d o t h e l i a l cell fo rm) , g e n e r a t i n g N O , a r e b o t h a c t i v e in t h e h u m a n p l a c e n t a (Pf i sche l e t al. 1982; M y a t t 1992) a n d w e r e i n v e s t i g a t e d as p o t e n t i a l m e d i a t o r s o f t h e E V C S . D P P IV, a d d i t i o n a l l y , w a s u s e d as m a r k e r of t he E V C S .
Material and methods
7issue preparation
Human placentae of uncomplicated pregnancies were delivered by elective Caesarian sections for reasons of pelvic presentations be- tween the 39th and 41st week of gestation. Tissue samples 3-5 mm thick and 1 ~ cm long were collected within 2 cm of the umbilical cord. For studying morphology, the samples were fixed in Karnovsky's solution for 24 h to 3 days at 4 ~ C, dehydrated in a graded ethanol series and embedded in glycol methacrylate (GMA, Technovit) according to the prescriptions of the manufacturer (Kulzer, Wehrheim, Germany), or for conventional histology in paraffin. Plastic sections were cut at 2 gm with 2050 rotary microto- me (Jung, Heidelberg, Germany), mounted on glass slides, dried for 1 h at 80 ~ C and subsequently at 45 ~ C overnight. For enzyme histo- chemistry and light microscopic immunocytochemistry, tissue sam- ples were mounted on specimen holders, frozen in liquid nitrogen and stored at - 4 0 ~ C until use. Cryostat sections, 4-10 ~tm thick, were cut at 25 ~ C with a Reichert Jung cryostat (model 2700 and 2800 N, Heidelberg, Germany). Sections were pretreated with ace- tone/chloroform (1:1/v :v, Merck, Darmstadt, Germany) or acetone for 5 min at --25 ~ C and subsequently air dried. For immunocyto- chemistry, sections were previously mounted on chrome alum-gela- tinized glass slides (Pappas 1971). All incubations were performed in a humidified chamber.
Staining procedures
Plastic sections, 2 gm thick, were stained with a 0.2% toluidine blue solution for 2 min on a hot plate at 60 ~ C. Subsequently, the slides were rinsed in distilled water and after dehydration mounted in Eukitt. The toluidine blue stock solution was prepared as follows: 2.5 g NaHCO3 (Merck, Darmstadt , Germany) were added to a 1% aqueous solution of toluidine blue O (Merck No. 1273). After 24 h, 30-40 drops of 1 N NaOH were added. For staining, this stock solution was diluted 1 : 5 with distilled water. For conventional his- tology, Azan staining (Romeis 1968) was used on 7-pm-thick paraffin sections.
Immunoc ytochemistr y
For light microscopic immunocytochemistry, acetone-pretreated cryostat sections were incubated with antibodies against u-actin (1:10, monoclonal, clone asm-1, Boehringer, Mannheirn, Germany), myosin (1:20, monoclonal, clone 2F12.Ag, Dianova, Hamburg, Germany), desmin (1:50-1:100, polyclonal, Laboserv, Giessen, Germany) and vimentin (1:50, polyclonal, Laboserv). Polyclonal antibodies against dipeptidyl peptidase IV (DPP IV, EC 3.4.14.5, dilution 1:20)were prepared as described by Hartel et al. (1987), against nitric oxide synthase type I (brain form, NOS I, dilution 1:50-1:100) according to Schmidt etal . (1992). Reactivity was demonstrated using FITC- or TRITC-labeled goat anti-mouse or rabbit IgG (Nordic, Tilburg, Netherlands), or swine anti-rabbit IgG/TRITC (Dakopatts, Hamburg, Germany). All dilutions were carried out with phosphate-buffered saline (PBS) pH 7.6, incuba- tions were performed for 1 h (first antibody) or 45 min (second antibody) at room temperature.
For postembedding immunogold labelling, tissue samples of stem villi were cut into pieces of approximately 1 mm ~, fixed in 3%
paraformaldehyde and 3% glutaraldehyde for 12 h at 4 ~ C, rinsed in a solution containing 0.01 M PBS, 1% BSA and 0.5% Tween, dehydrated in ethanol and embedded in LR-White (Plano, Mar- burg, Germany). Ult ra thin sections were collected on fine-mesh, naked nickel grids and incubated at room temperature for 12 h with the same specific antibodies as for light microscopic immunocyto- chemistry. Dilutions were the same as for light microscopy except for anti-~-actin (1 : 100) and anti-myosin (1 : 5). Visualization of im- munoreactivity was carried out using gold-labelled goat anti-mouse IgG 10 nm, 1 : 50, or goat anti-rabbit IgG 5 and 10 nm, 1 : 50 (Amer- sham, UK). Grids were subsequently stained with 1% tannin (15 min) and 4% uranylacetate (20 min) and examined in a Zeiss EM 109 electron microscope. For light and electron microscopy, double incubations with combinations of all the specific antibodies described were performed.
Controls
Control reactions for immunostaining were carried out in PBS without specific antibodies. Control reactions for NOS I were addi- tionally performed by using preabsorbed specific antibody: 50 or 100 ng NOS I and 1 ~tl antiserum were adjusted to 100 p.1 PBS, stirred gently for 1 h at room temperature and subsequently stored for 14 h at 0-4 ~ C. After centrifugation with a bench centrifuge (Biofuge A, Heraeus-Christ, Osterode/Harz, Germany) for 10 min at 3,000 RPM the supernatant was used for immunoincubation.
Enzyme histochemistry
Acetone/chloroform-pretreated cryostat sections were used to demonstrate dipeptidyl peptidase IV (DPP IV, EC 3.4.14.5) with Gly-Pro-4-methoxy-2-naphthylamide as substrate (Bachem, Heidelberg, Germany) and Fast Blue B (high purity grade, Serva, Heidelberg) for simultaneous azocoupling according to Lojda et al. (1991). Incubation was carried out for 45 min at 37 ~ C.
Results
Morphology
A f t e r A z a n s t a i n i n g o f p a r a f f i n s ec t i ons , t h e c h o r i o n i c p l a t e a n d s t e m vil l i r e v e a l e d b r i g h t l y r e d - c o l o u r e d ex- t r a v a s c u l a r cells w i t h d i f f e r en t f ea tu re s . T o l u i d i n e b l u e - s t a i n e d s e m i t h i n p l a s t i c s e c t i o n s d e m o n s t r a t e d s i m i l a r cells in t h e s a m e p o s i t i o n in a b l u e c o l o u r , w h i l e c o n n e c - t ive t i s s u e f ib res w e r e u n s t a i n e d . I n t h e m i d d l e l a y e r o f t h e c h o r i o n i c p l a t e t h e s e cells w e r e i n t e r m i n g l e d w i t h c o n -
Fig. 1. Chorionic plate, NOS I immunoreactivity. Reactivity in sev- eral longitudinally and transverse arranged layers of cells of the extravascular contractile system (EVCS). x 350
Figs. 2--4. Stem villi, semithin plastic sections, toluidine blue stain- ing. Fig. 2. EVCS cells in the perivascnlar contractile sheath (PVCS, small arrows) running in longitudinal direction to a fetal blood vessel (X, lumen not cut). x 220. Figs. 3 and 4. Different features of EVCS cells, some of which are forming tong, thin cell processes (large arrows), others are thick with coarse and plain cell processes (arrowheads), ending between connective tissue fibres, x 350
Fig. 5. Villi, ~-actin immunoreactivity. Reactivity in a perivascular sheath of a large stem villus (left side; the central fetal blood vessels are not cut). Outside the stem villus, immunoreactivity only in me- dia smooth muscle cells (arrows). x 220
Fig. 6. Stem villus, c~-actin immunoreactivity. EVCS cell processes insert beneath the trophoblast (arrow). F, Fibrinoid. x 350

543
544
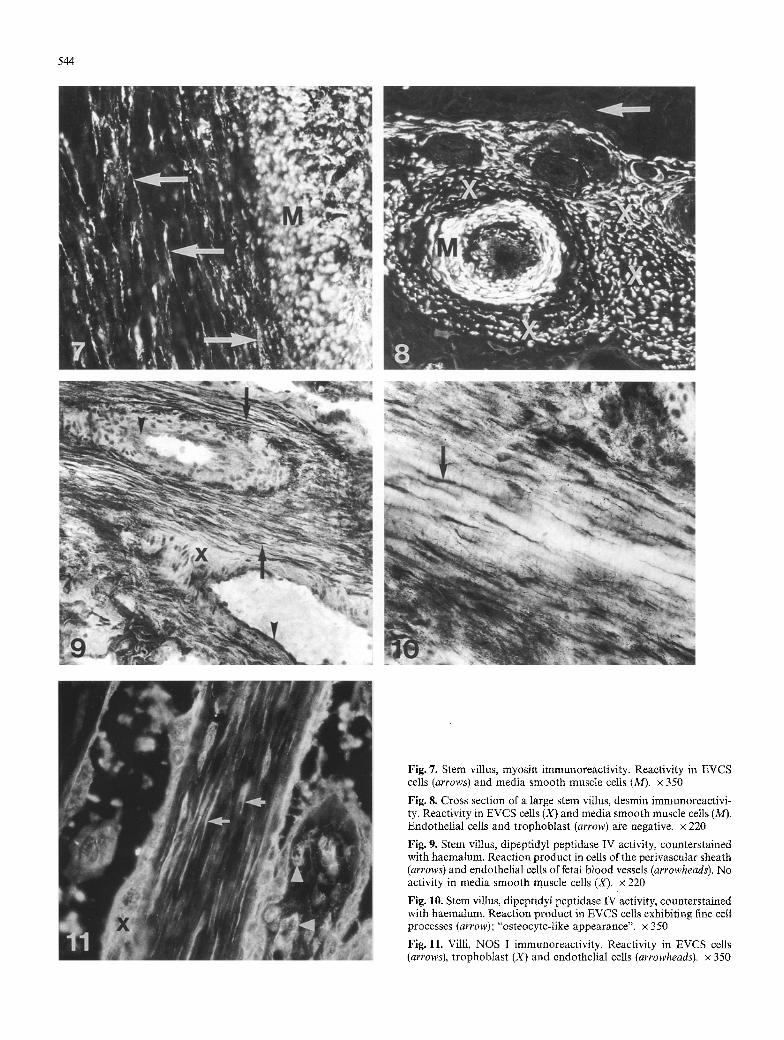
Fig. 7. Stem villus, myosin immunoreactivity. Reactivity in EVCS cells (arrows) and media smooth muscle cells (M). x 350
Fig. 8. Cross section of a large stem villus, desmin immunoreactivi- ty. Reactivity in EVCS cells (X) and media smooth muscle cells (M). Endothelial cells and trophoblast (arrow) are negative, x 220
Fig. 9. Stem ~dllus, dipeptidyl peptidase IV activity, counterstained with haemalum. Reaction product in cells of the perivascular sheath (arrows) and endothelial cells of fetal blood Vessels (arrowheads). No activity in media smooth muscle cells (X). x 220
Fig. 10. Stem villus, dipeptidyl peptidase IV activity, counterstained with haemalum. Reaction product in EVCS ceUs exhibiting fine cell processes (arrow); "osteocyte-like appearance", x 350
Fig. 11. Villi, NOS I immunoreactivity. Reactivity in EVCS cells (arrows), trophoblast (X) and endothelial cells (arrowheads). x 350

545
Fig. 12. Alpha-actin immunoreactivity. Immunogold labelling of a smooth-muscle-like EVCS cell. Large organelle-free areas (X). No gold particles in organelle-rich area at the pole of the nucleus (ar- rowheads). Cell process (arrow); collagen fibres (CF) in close connec- tion with the barely visible plasma membrane, x 61,200
Fig. 13. Dipeptidyl peptidase IV immunoreactivity. Immunogold labelling of a smooth-muscle-like EVCS cell, showing dense plaques (arrows), filamentous area (X) and caveolae (arrowheads). x 74,400

546
nective tissue fibres and had a rather compact structure, with two or three short cell processes. Towards the bor- der to the villous zone, they changed to spindle-shaped cells. From their whole length several thin and short pro- cesses could branch off nearly at right angles. Cells lying in parallel seemed to contact each other by these cell processes. At the border to the villous zone, spindle- shaped cells formed layers that were arranged in alternat- ing longitudinal and transverse directions (Fig. 1). Single cells branched off from these plates with their cell pro- cesses inserted just beneath the trophoblastic layer, obvi- ously in the basement membrane, others ended in sub- chorial fibrinoid. From the chorionic plate, closely ar- ranged spindle-shaped cells were running into the stem villi, and together with connective tissue fibres formed a sheath surrounding fetal blood vessels (Figs. 2, 5, 7, 8, 9). Some of the sheath cells seemed to insert beneath the trophoblastic layer (Fig. 6), while others ended in perivil- lous fibrinoid. According to the different segments of stem villi, the vascular sheath lost thickness, until in ra- muli chorii (according to the classification in Kaufmann et al. 1979), only a few long and slender spindle-shaped cells remained. Spindle-shaped cells in all stem villi char- acteristically often showed an extremely undulating ap- pearance.
In stem villi, besides the spindle-shaped form, other features of sheath cells were conspicuous. They could form extremely long, thin cell processes, by which they often seemed to contact each other. Other cells were thick and ended with coarse, plain cell processes between con- nective tissue fibres (Fig. 3). In semithin plastic sections, many of the sheath-forming cells revealed long filament- like structures (Figs. 3, 4). Next to large fetal blood ves- sels, it seemed that media smooth muscle cells were pro- ceeding into the extravascular cell bundles or vice versa.
munogold labelling was seen in the media and endothe- lial cells of fetal blood vessels and in the trophoblast (Fig. 11). Anti-~-actin reactivity was present in media smooth muscle cells (Figs. 5, 6). The same was true for myosin immunofluorescence (Fig. 7), which was addi- tionally detected in the trophoblast. Anti-desmin-reactiv- ity could also be detected in media smooth muscle cells, but not in endothelial cells and trophoblast (Fig. 8). Anti- DPP IV reactivity was localized in endothelial cells and trophoblast and was absent in media smooth muscle cells. Incubations without specific antibodies and incuba- tion with absorbed anti-NOS I were negative.
Enzyme histochemistry
DPP IV was highly active in all of the EVCS cells of the chorionic plate and stem villi described above (Figs. 9, 10). DPP IV-positive cells were often seen to end in sub- chorial and intervillous fibrinoid. In stem villi, sheaths of the EVCS were remarkable for their enzyme activity and their peculiar morphology. In the chorionic plate and stem villi, reaction product was also localized in fine cell processes branching off from the spindle-shaped cells at right angles. The cells seemed to contact each other via these processes and thus morphologically resembled os- teocytes (Fig. 10). Some of the sheaths had a centre de- void of DPP IV activity or contained only very few en- zyme-positive cells. DPP IV also generated large amounts of reaction product in the cytoplasm of cytotro- phoblasts and in the adventitia of fetal blood vessels, while small amounts were found in the cytoplasm of the syncytiotrophoblast and endothelial cells. Media smooth muscle cells never showed DPP IV activity (Fig. 9).
I mmunoc ytochemistr y
All of the above mentioned cell types in the chorionic plate and stem villi were immunoreactive for ~-actin (Figs. 5, 6), myosin (Fig. 7), desmin (Fig. 8), vimentin (not shown), DPP IV (Fig. 13) and NOS I (Figs. 1, 11). In com- parison with the numerous ~-actin immunofluorescent cells in sheaths of stem villi, only a few spindle-shaped sheath cells were NOS I-positive. Some NOS I-positive cells in stem villi had long, slender cell processes by which they seemed to cross or surround several of the sheath- forming cells. The presence of all investigated filament types, DPP IV and NOS I in the EVCS of the chorionic plate and stem villi was confirmed by double incubations or serial sections. Ultrastructurally, the immunogold-la- belled cells contained large amounts of filaments (Figs. 12, 13). Dense areas, numerous caveolae at the plasma membrane and few cell organelles, which often were restricted to the poles of the nuclei, were typical for these cells (Figs. 12, 13). Other immunogold-labelled cells contained more cell organelles and fewer filaments. Col- lagen fibres were arranged in direct contact with the EVCS cells and seemed to insert in or arise from the plasma membrane in an oblique direction (Fig. 12).
Besides the immunoreactivity in cells of the EVCS, a bright NOS I immunofluorescence and clearcut im-
Discussion
Our study clearly emphasizes that in the human placenta, besides the fetal blood vessel system, a second extravas- cular contractile system exists. This system obviously forms layers in the chorionic plate and sheaths surround- ing fetal blood vessels in stem villi (trunci chorii and rami chorii) and is therefore present in all cotyledons. Com- paring our results with the excellent and precise morpho- logical description of Spanner in 1936, there can be no doubt that the cells referred to in the two studies are identical. We have additionally confirmed the presumed contractility of these cells by visualizing their actin and myosin filament system, and thus in a way completed the physiological studies of Krantz and Parker (1963). These authors found spontaneous contractions in "muscle con- nective tissue" from anchoring villi, although it is not clearly stated in their study that the investigated "muscle connective tissue" was devoid of vascular smooth muscle.
In all biochemical and physiological studies of placen- tal smooth muscle, the differentiation between extravas- cular and vascular (media) smooth muscle seems to be a critical point. Referring to the extravascular smooth muscle study of Krantz and Parker in 1963, Huszar and Bailey (1979 a, b) isolated and characterized myosin from the human term placenta by removing all visible blood vessels and described a relationship between actin-

547
myosin interaction and myosin light chain phosphoryla- tion. In their study, however, they concede the possibility that a certain percentage of the isolated myosin might originate from the smooth muscle of blood vessel walls. Furthermore, King and Gr6schel-Stewart (1965) and Michael (1974) have reported the presence of actomyosin in the human term placenta, and related this to the ex- travascular smooth muscle. Although not stated by the authors, evidence exists from both studies that the inves- tigated tissues were not completely free of vascular smooth muscle. Therefore, at present, there exists no ob- vious physiological or biochemical proof for a sponta- neous contraction of the EVCS. Isolated strips from the periphery and centre of human term placentae, which were free of visible blood vessels up to x 25 magnifica- tion, showed no spontaneous contractions during iso- metric testing (Babcock 1969). Since extravascular smooth muscle cells were stained by classical histological techniques, Babcock (1969) explains his negative results by a failure of sensitivity in the isometric testing system. The demonstration of contractile filaments in EVCS cells in the present study supports this hypothesis.
In the chorionic plate, cells of the EVCS are loosely arranged between connective tissue fibres or form layers at the border to the villous zone. In stem villi, cells of the EVCS, in community with connective tissue fibres, form sheaths that enclose fetal blood vessels. It is obvious that EVCS cells of the sheaths arc part of the "fibrous par- avascular sheath", which, in placental pathology, is used as an index of maturity (Kloos and Vogel 1974; Vogel 1992). To emphasize the large numbers of contractile cells and their location within this sheath, we propose the common term "perivascular contractile sheath" (PVCS). By this definition, the perivascular contractile sheath consists of extravascular contractile cells and connective tissue, in contrast, the term EVCS (extravascular con- tractile system) defines extravascular contractile cells within the whole placenta (chorionic plate and stem villi) without connective tissue.
Some of the former studies refer to the EVCS as smooth muscle, others to myofibroblasts. From our study, the nature of the EVCS cells remains unclear. The filament distribution we described was shown to be typi- cal for myofibroblasts (Gown 1990). The fact that DPP IV is present in EVCS cells, also seems to justify their view of myofibroblasts rather than smooth muscle cells. DPP IV has never been demonstrated in smooth muscle cells, including those of the uterus (Graf, unpublished results). In human placental stromal cells, DPP IV was first described by Heymann and Mentlein (1984) and Feller et al. (1985). On the basis of their comparisons with tissue from Dupuytren's contracture, these authors argue that all DPP IV-positive stromal cells in all types of pla- cental villi, including terminal villi, are myofibroblasts. Using double immunogold labelling, we showed that in the EVCS contractile and intermediate filaments are co- localized with DPP IV. In terminal villi, however, apart from capillaries including pericytes, no ~-actin im- munoreactivity was demonstrated, only DPP IV enzyme activity and desmin and vimentin immunoreactivity (Graf et al. 1992). Thus, in the human placenta at least two different DPP IV-positive cell types exist.
In contrast, the ultrastructure of DPP IV-positive
EVCS cells, as shown in the present study, i.e. distribu- tion of filament areas, caveolae and cell organelles, rather point to smooth muscle than to myofibroblasts. DPP IV activity may correspond to a specialized or transitory form of smooth muscle. In order to identify contractile extravascular cells in stem villi of human placenta, Kohnen et al. (1993) used GB 42, a monoclonal antibody that binds to smooth muscle (Hsi and Yeh 1988). They showed a co-localization of GB 42 with anti-~-actin, - myosin, -desmin and -vimentin-positive cells in stem villi of more than 300 ~tm, while in smaller stem villi no GB 42-immunoreactivity was found.
The presence of DPP IV activity within the cytoplasm of EVCS cells, and cytotrophoblasts and the syncytiotro- phoblast (as previously reported, Graf et al. 1992) may astonish, because usually DPP IV is thought to be a plas- ma membrane-bound enzyme. However, cellular local- ization of DPP IV activity depends on the organ investi- gated. In kidney, intestine and liver, DPP IV is a brush- border respectively plasma membrane-bound enzyme, while in parotid and submaxillary gland, it is also present in serous secretory granules (McDonald and Barrett 1986). DPP IV activity, additionally, has been reported in human saliva, serum and cerebrospinal fluid (McDonald and Barrett 1986). DPP IV activity within the cytoplasm of EVCS cells of the human placenta, as reported in this study, is also in agreement with the results of Feller et al. (1985). These authors described strong DPP IV activity diffusely distributed throughout the fusiform or stellate cytoplasm and cytoplasmic extensions of stromal myofi- broblasts. Furthermore, our results after postembedding immunogold labelling confirm our light microscopical results. The plentiful enzyme activity throughout the cy- toplasm of EVCS cells and the predominance of filamen- tous areas within these cells may explain why gold parti- cles are also localized in the filamentous area of the cells. Further investigations may clarify the intracellular site of DPP IV activity in cells of the EVCS and its function in this respect.
Placental fetal blood flow is modulated by autacoids, which affect fetal vascular endothelial cells or media smooth muscle cells. A number of these autacoids are synthesized and metabolized by the placenta, for example angiotensins or substance P (Pfischel et al. 1982; Hey- mann and Mentlein 1984; McDonald and Barrett 1986; Boura and Walters 1991; Hagemann et al. 1994). It has been suggested that autacoids could modify cells of their origin or affect functions of adjacent or more distant cells (Boura and Walters 1991). They may reach the fetal ves- sels by the fetal blood stream or via maternal intervillous blood. Because of its localization just between both blood streams, it may be suggested that the PVCS in stem villi could also be modulated by autacoids. This modula- tion could occur via metabolizing enzymes, as in the case of substance P, which is metabolized by DPP IV (Hey- mann and Mentlein 1984). It could also occur via auta- cold-generating enzymes, as in the case of nitric oxide synthase (NOS).
NOS generates NO, which is thought to be a placental autacoid. Recently, isolation and characterization of hu- man placental NOS type III (endothelial form) has been performed by Myatt (1992). In the present study we were able to localize NOS type I (brain form) in smooth-mus-

548
cle-l ike cells of the chor ion ic p la te and in s tem villi. N o t all of the n u m e r o u s e -ac t in immunof luo re scen t cells in PVCS of s tem villi were N O S I immunoreac t ive . N O acts as a v a s o d i l a t o r by increas ing c G M P in m e d i a s m o o t h muscle cells (Schmid t a n d B6hme 1990; M o n c a d a et al. 1991). Thus, wi th in the PVCS, the few N O S I -pos i t ive EVCS cells cou ld be specia l ized smoo th -musc l e - l i ke cells or EVCS cells re la ted to p a r a n e u r o n s , genera t ing N O and so m o d u l a t i n g the tone of the PVCS (Sch6nfelder et al. 1993).
W h a t m a y be the func t ion of the EVCS in h u m a n p l a c e n t a ? As the EVCS is t hough t to be also presen t in ancho r ing villi (Spanner 1936), c o n t r a c t i o n of all cotyle- dons cou ld lead to an a p p r o a c h of the chor ion ic and basa l plate. This in tu rn cou ld affect b l o o d flow in the in terv i l lous space and fetal b l o o d vessel sys tem (Dubreu i l and Rivi6re 1932; Spanne r 1936; K r a n t z and Pa rke r 1963). However , it is no t clear, whe the r the capac i ty of the EVCS is h igh enough to induce a u t o n o m o u s m o v e m e n t of the whole p lacenta . A n o t h e r func t iona l role for the EVCS m a y be to regula te the muscle tone in s tem villi. In this way, the a rch i t ec tu re of the c o t y l e d o n m a y be kep t upr ight , i n d e p e n d e n t of b l o o d flow, pressure on o r tor - s ion of the co ty ledons . Final ly , the u p t a k e of nu t r i en t s in to p l acen ta l villi m a y be m e d i a t e d by vil lous con t r ac - t ions as in the smal l intest ine.
Acknowledgements. We are indebted to the nurses and midwives for excellent co-operation, M. Gutsmann, M. Khakpour, U. Sauerbier and H. Tersch for skilful technical assistance and H. Neudeck for helpful discussions. The authors are also thankful to Dr. M. Shak- ibaei for introduction to the technique of immunogold labelling. This work was supported by Deutsche Forschungsgemeinschaft (Sfb 174 and Re 523/3-4).
References
Babcock RJ (1969) Smooth muscle in the human placenta. Am J Obstet Gynecol 105:612-615
Becker V, Schiebler T, Kubli F (1981) Die Plazenta des Menschen. Thieme, Stuttgart, p 55
Beham A, Denk H, Desoye G (1988) The distribution of intermedi- ate filament proteins, actin and desmoplakins in human placen- tal tissue as revealed by polyelonal and monoelonal antibodies. Placenta 9:479-492
Bertolini R, ReiBig D, Schippel K (1969) Elektronenmikroskopis- che Befunde an den Zellen in der Chorionplatte der reifen men- schlichen Plazenta. Z Mikrosk-Anat Forsch 80:358-368
Boura ALA, Walters WAW (1991) Autacoids and the control of vascular tone in the human umbilical-placental circulation. Pla- centa 12:453-477
Demir R, Demir N, Kohnen G, Kosanke G, Mironov V, Ust/inel I (1992) Ultrastructure and distribution of myofibroblast-like cells in human placental stem villi. Electron Microsc 3:509-510
Dubreuil G, Rivi6re M (1932) Formations fibro-musculaires du chorion et des villositbs du placenta humain. CR Seances Soc Biol Fil 111:170-172
Feller AC, Schneider H, Schmidt D, Parwaresch MR (1985) Myofi- broblasts as a major cellular constituent of villous stroma in human placenta. Placenta 6:405-415
Gown AM (1990) The mysteries of the myofibroblast (partially) unmasked. Lab Invest 63:1-3
Graf R, Frank HG, 13hey T (1992) Histochemical and immunocyto- chemical investigations of the fetal extravascular and vascular contractile system in the normal placenta and during pre- eclampsia. In: Neubert D, Kavlock R J, Merker H J, Klein J (eds)
Risk assessment of prenatally-induced adverse health effects. Springer, Berlin Heidelberg New York, pp 537-550
Graf R, Langer J-U, Sch6nfelder G, Hartel-Schenk S, Reutter W (1993) The extravascular system of the human placenta. Placen- ta 14:A.25
Hagemann A, Nielsen AH, Poulsen K (1994) The uteroplacental renin-angiotensin system: a review. Exp Clin Endoerinol (in press)
Happe H (1906) Beobachtungen an Eihfiuten junger menschlicher Eier. Anat Hefte 32:173-212
Hartel S, Hanski C, Kreisel W, Hoffmann C, Mauck J, Reutter W (1987) Rapid purification of dipeptidyl peptidase IV from rat liver plasma membranes. Biochim Biophys Aeta 924:543-547
Heymann E, Mentlein R (1984) BeeinfluBt Dipeptidylpeptidase IV Blutdruck und Gerinnung? Klin Wochenschr 62:2-10
Hsi BL, Yeh CJG (1988) Monoclonal antibodies to placental vascu- lar structures. Trophoblast Res 3:139-148
Huszar G, Bailey P (1979a) Isolation and characterization of myosin in the human term placenta. Am J Obstet Gynecol 135:707-712
Huszar G, Bailey P (1979b) Relationship between actin-myosin in- teraction and myosin light chain phosphorylation in human placental smooth muscle. Am J Obstet Gynecol 135:718-726
Kaufmann P, Sen DK, Schweikhart G (1979) Classification of hu- man placental villi. I. Histology. Cell Tissue Res 200:409-423
King TM, Gr6schel-Stewart U (1965) Placental contractile protein. Am J Obstet Gynecol 93:253-258
Kloos K, Vogel M (1974) Pathologic der Perinatalperiode. Thieme, Stuttgart, pp 22-27
Kohnen G, Mironov V, Demir R, Castellucci M, Kaufmann P (1993) Immunhistochemie and Ultrastruktur kontraktiler Zellen im Zottenstroma der menschlichen Placenta. Ann Anat 175 [Sup- pl] :37
Krantz KE, Parker JC (1963) Contractile properties of the smooth muscle in the human placenta. Clin Obstet Gynecol 6:26-38
Lojda Z, Gossrau R, Stoward PJ (1991) Proteases. In: Stoward PJ, Pearse AGE (eds) Histochemistry. Theoretical and applied, vol 3. Enzyme histochemistry, 4th edn. Churchill Livingstone, Edinburgh, pp 305-309
McDonald JK, Barrett AJ (1986) Mammalian proteases: a glossary and bibliography, vol 2. Academic Press, London
Michael C (1974) Actomyosin content of the human placenta. J Obstet Gynaecol 81:307-310
Moncada S, Palmer RMJ, Higgs EA (1991) Nitric oxide: physiolo- gy, pathophysiology and pharmacology. Pharmacol Rev 43:109-142
Myatt L (1992) Control of vascular resistance in the human placen- ta. Placenta 13:329-341
Pappas PW (1971) The use of a chrome alum-gelatin (subbing) solution as a general adhesive for paraffin sections. Stain Teeh- nol 46:121-124
Pfischel G, Mentlein R, Heymann E (1982) Isolation and character- ization of dipeptidyl peptidase IV from human placenta. Eur J Biochem 126:359-365
Romeis B (1968) Mikroskopische Technik. Oldenbourg, Munich Schmidt HHHW, B6hme E (1990) NO, ein hormonaler Wirkstoff.
Med Mol Pharmacol 3:74-81 Sehmidt HHHW, Gagne GD, Nakane M, Pollock JS, Miller MF,
Murad F (1992) Mapping of neural nitric oxide synthase in the rat suggests frequent co-localization with NADPH-diaphorase but not with soluble guanylyl cyclase, and novel paraneuronal functions for nitrinergic signal transduction. J Histochem Cy- tochem 40:1439-1456
Sch6nfelder G, Graf R, Schmidt HHHW (1993) A possible regula- tion of the extravascular contractile system in human placenta by nitric oxide synthase immunoreactive cells. Placenta 14:A.69
Spanner R (1936) Mtitterlicher und kindlicher Kreislauf der men- schlichen Plazenta und seine Strombahnen. Z Anat Entwick- lungsgesch 105 : 163-242
Vogel M (1992) Atlas der morphologischen Plazentadiagnostik. Springer, Berlin Heidelberg New York, pp 4 and 80--91