the evolutionary study of the immunoglobulin heavy chain
TRANSCRIPT

The Evolutionary Study of the Immunoglobulin Heavy Chain Genes of a Bony Fish, Rainbow Trout
( Oncorhynchus my kiss)
c/>
AKADEMISK AVHANDLING
som för avläggande av doktorsexamen i medicinsk vetenskap vid Umeå Universitet, offentligen kommer att försvaras i föreläsningssalen,
Institutionen för Mikrobiologi, Umeå Universitet,Fredagen den 8 december 1995, kl 10.00
av
Elisabet Andersson
Institutionen för Tillämpad Cell och Molekylärbiologi Umeå Universitet
Umeå 1995
Fakultetsopponent: Prof. Gregory W. Warr, Charleston, USA

A b s tra c t
The Evolutionary Study of the Immunoglobulin Heavy Chain Genes of a Bony Fish, Rainbow Trout ( Oncorhynchus my kiss)Elisabet Andersson, Department of Cell and Molecular Biology University of Umeå, S-901 87 Umeå, Sweden. Fax: +46-90-77 14 20
The antibody (ab; or immunoglobulin, Ig) molecules1 of vertebrates constitute a major immune defence against microbes. The ab-diversity is generated through multitudes of mechanisms; e.g. recombination of germline encoded variable region genes (V), diversity (D) and joining (J) gene segments, junctional diversity and somatic mutations. A large number of V genes provides a basis for ab-diversity in the vertebrate genomes. This thesis focus on the aspects of evolutionary stability and rate and the complexity and diversity of the immunoglobulin genes in a teleost fish, rainbow trout ( Oncorhynchus mykiss).We have characterised many expressed V h genes and an IgM constant region gene (Cn) from rainbow trout. These Vhs can be divided into 11 different families based on the criterion of 80% DNA identity. Comparison of V h sequences of two fish species (rainbow trout and channel catfish) suggests that a V h family may last 150 million years (my) or longer in evolution. This is compatible with our study using Southern hybridisation of DNA from several different fish species. The Vh family evolution was further studied by extensive phylogenetic analysis involving a large number of V h families from all major vertebrate phyla. The "species-specific" residues were identified in many rainbow trout Vhs and are discussed in the context of multigene family evolution. Analysis of V h CDR3 regions suggest that rainbow trout utilises several D and Jh segments and nucleotide addition/deletion to generate antibody diversity.Studies using the Cn indicate that the Ig heavy chain organisation in rainbow trout is similar to the mammalian organisation (a Vh cluster followed by a D cluster, Uh cluster and one copy of the Ch gene). Rainbow trout probably generates membrane IgM lacking the Ch 4 domain through an unusual RNA splicing, which has also been found in other bony fish species.From amino acid comparisons of the Cn domains from many vertebrate phyla, we estimated the amino acid substitution rate for the Ch 4 domain to 1 .4x 1 0-9/site /year. This rate (molecular clock) was then used to estimate the divergence time between rainbow trout and several other fish species. The divergence time of salmonid fishes was also estimated (12 -1 6 my) using the nucleotide sequence in IgM intra-domain introns of rainbow trout and Arctic charr.
Keywords: immunoglobulin, heavy chain variable region gene, V h , rainbow trout, evolution, salmonid fish
New Series No 452 ISSN 0346-6612 ISBN 91-7191-116-2

The Evolutionary Study of the Immunoglobulin Heavy Chain Genes
of a Bony Fish, Rainbow Trout ( Oncorhynchus mykiss)
by
Elisabet Andersson
O•<
*
*cn
Umeå 1995

Cover Drawing:Francesco Colucci
Department of Cell and Molecular Biology Umeå University, Sweden
© Elisabet Andersson, 1995 New Series No. 452 ISSN 0346-6612
ISBN 91-7191-116-2
Printed in Sweden VMC
Fys. Bot. Umeå University
1995

In memory of my father, Lennart Andersson


Contents
A b s tra c t 1
A b b rev ia tio ns 3Abbreviations 3Encyclopaedia for non-fishers like myself 4
Publications 5
1. In tro d u c tio n 6
1:1. The antibody molecule: structural features 7
1:2. The immunoglobulin genes and their organisation 9The V gene 9The Ig heavy chain gene organisation 1 1The Ig light chain gene organisation 1 3
1:3. Generation o f antibody diversity 14Combinatorial diversity 1 4Gene conversion 1 4Junctional diversity 1 5Somatic hypermutation 1 5
1:4. The fish antibody response 1 6General features o f the fish immunity 1 6Fish antibodies 1 7Fish antibody response 1 8
1:5. Evolution of multigene families 18Definition o f a multigene family 1 8Evolutionary mechanisms acting on multigene families 1 9
1:6. The molecular clock of proteins 1 9
1:7. Fish phylogeny 20
2. Aims of this thesis 22

3. Results and Discussion 23
3:1. Rainbow trou t IgVH genes and family structure 23
3:2. Evolution of Vh genes and their families 24A. Stability o f a Vh gene family 2 4B. Evolution o f vertebrate Vh clans: identification o f a teleost and an archaic clan separated from the mammalian clans 2 6C. Species-specific amino acid residues: implicationon the IgV gene evolution 2 8Significance of the Vh family: a speculation 29
3:3. The IgH diversity in rainbow trout 29Vh diversity 3 0D-Jh sequences and CDR3 diversity 3 0The antibody response in fish and other low vertebrates-Why is it so restricted? 33
3:4. The IgM heavy chain constant region 34Genomic organisation o f the IgH locus in teleost fishes 34Peculiarity o f membrane IgM in bony fish 3 6
3:5. Speed of IgM constant region gene evolution 3 7The IgM protein 3 7The amino acid substitution rate o f IgM Ch 4 domain 3 8
3:6. Evolution of salmonid fishes 39
4. Concluding remarks 42
5. Acknowledgem ents 44
6. References 45
7. Papers I to V

Abstract
The Evolutionary Study of the Immunoglobulin Heavy Chain Genes of a Bony Fish, Rainbow Trout (Oncorhynchus my kiss)Elisabet Andersson, Department of Cell and Molecular Biology University of Umeå, S-901 87 Umeå, Sweden. Fax: +46-90-77 14 20
The antibody (ab; or immunoglobulin, Ig) molecules of vertebrates constitute a major immune defence against microbes. The ab-diversity is generated through multitudes of mechanisms; e.g. recombination of germline encoded variable region genes (V), diversity (D) and joining (J) gene segments, junctional diversity and somatic mutations. A large number of V genes provides a basis for ab-diversity in the vertebrate genomes. This thesis focus on the aspects of evolutionary stability and rate and the complexity and diversity of the immunoglobulin genes in a teleost fish, rainbow trout (Oncorhynchus mykiss).We have characterised many expressed Vh genes and an IgM constant region gene (Cn) from rainbow trout. These Vhs can be divided into 11 different families based on the criterion of 80% DNA identity. Comparison of Vh sequences of two fish species (rainbow trout and channel catfish) suggests that a Vh family may last 150 million years (my) or longer in evolution. This is compatible with our study using Southern hybridisation of DNA from several different fish species. The Vh family evolution was further studied by extensive phylogenetic analysis involving a large number of V h families from all major vertebrate phyla. The "species-specific" residues were identified in many rainbow trout Vhs and are discussed in the context of multigene family evolution. Analysis of Vh CDR3 regions suggest that rainbow trou t utilises several D and Jh segments and nucleotide addition/deletion to generate antibody diversity.Studies using the Cn indicate that the Ig heavy chain organisation in rainbow trout is similar to the mammalian organisation (a Vh cluster followed by a D cluster, Jh cluster and one copy of the Ch gene). Rainbow trout probably generates membrane IgM lacking the Ch4 domain through an unusual RNA splicing, which has also been found in other bony fish species.From amino acid comparisons of the Qi domains from many vertebrate phyla, we estimated the amino acid substitution rate for the Ch 4 domain to1.4x10-9/site/year. This rate (molecular clock) was then used to estimate the divergence time between rainbow trout and several other fish species. The divergence time of salmonid fishes was also estimated (12-16 my) using the nucleotide sequence in IgM intra-domain introns of rainbow trout and Arctic charr.
Keywords: immunoglobulin, heavy chain variable region gene, Vh , rainbow trout, evolution, salmonid fish
New Series No 452 ISSN 0346-6612 ISBN 91-7191-116-2
1

Referat av Elisabet Anderssons avhandling:En evolutionär studie av de gener som kodar fö r antikroppens tunga kedja i regnbågslax ( Oncorhynchus mykiss)
Antikroppar (ak) är mycket viktiga proteiner i vertebraters (ryggradsdjur) försvar mot mikroorganismer. Flera olika mekanismer; t.ex. kombinering av många gensegment (V-, D- och J-segment), insertioner/deletioner av nukleotider och somatiska mutationer genererar miljontals ak med olika specificiteter. En av de viktigaste faktorerna för denna ak diversitet är det stora antalet V-gener som finns i vertebraters arvsmassa. I denna avhandling presenteras olika aspekter av V-geners evolution med fokusering på den tunga kedjans gener i en primitiv vertebrat, regnbågslax (Oncorhynchus mykiss).Vi har karakteriserat många VH-gener samt den konstanta regionen av IgM (Cn) från regnbågslax. Dessa VH-gener kan delas in 11 olika VH-genfamiljer. Jämförande studier av VH-gener från två olika fiskar (regnbåge och kattfisk) visade att en VH-genfamilj kan överleva under evolutionen i mer än 150 miljoner år. Detta resultat bekräftades genom hybridiseringsförsök med DNA från olika mer eller mindre besläktade fiskar. Evolutionen av en VH-genfamilj studerades ytterligare i en omfattande fylogenetisk analys av VH-genfamiljer från de flesta större vertebrata klasser. De "art-specifika" aminosyra-positioner som identifierades i regnbågslaxens VH-gener diskuteras i samband med evolutionen av multigenfamiljer. Vidare indikerade våra resultat på att regnbågslax använder flera D- och Jh - gensegment samt insertioner/deletioner av nukleotider för att generera ak- diversitet på samma sätt som däggdjur.Våra resultat visade också att den genomiska organisationen av den tunga kedjans gener liknar den i möss, människor och benfiskar (d.v.s. en stor grupp av VH-gener följd av ett antal D- och JH-gener samt en kopia av C^). Resultaten indikerade dessutom a tt regnbågslax genererar den membranbundna formen av IgM genom att deletera CH4-domänen, e tt fenomen som observerats i andra benfiskar.Från jämförande studier av C^-domänerna från olika vertebrata klasser har vi kunnat uppskatta hur snabbt aminosyror byts ut i CH4-domänen (1.4x1 O*9 substitutioner/position/år). Denna hastighet (den molekylära klockan) kunde sedan användas för a tt beräkna när olika fiskar divergerade från varandra, t.ex. regnbågslax-lax divergerade för 10-16 miljoner år sedan. Vi använde även intradomän introner av Cn-gener för att uppskatta när olika laxfiskar divergerade. Regnbågslax och röding divergerade, enligt denna metod, för 12-16 miljoner år sedan.
2

Abbreviations
ig immunoglobulinigc immunoglobulin constant regionigH immunoglobulin heavy chainigL immunoglobulin light chainmlg membrane Igslg secreted IgTM transmembraneab antibodyH heavyL lig h tV variableD diversityJ joiningC constantFR frame work regionCDR complementarity determining regionTcR T cell receptorRSS recombination signal sequencesMHC major histocompatibility complexMLR mixed leukocyte reactionbp base pairnt nucleotideaa amino acidmy million yearKaa average number of aa substitutions per siteP0 percent aa differences between two peptidesT divergence time between two specieskaa average aa substitution rate per site per yeard expected number of nt substitutions per siteIEF isoelectric focusing measurementS.E. standard errorDNP dinitrophenyiTNP trinitrophenylKLH key hole limpet hemocyaninLPS lipopolysaccharide
3

"Encyclopaedia for non-fishers like myself.”
cyclostomeelasmobranch
teleosteuteleost
holosteancoelacanthdipnoitetrapod
lampreyhagfishsharkskatechimerasturgeonLa time rialongnose garbowfinladyfishcatfishgoldfishsalmonid fishrainbow troutsalmoncharrseatroutwhitefishpikeherringcodburbottunaperchsea basshalibutlungfishray-finned fishlobe-finned fish
lamprey and hagfishcartilaginous fishes such as sharks and skates (broskfiskar)bony fishes, e.g. rainbow trout, salmon and cod (benfiskar) a group of bony fishes represented in the thesis by: rainbow trout, salmon, charr, cod, ladyfish, catfish a group of bony fishes represent by longnose gar and bowfin a ” living fossil” represented today by Latimeria lungfishland living species e.g.: amphibian, reptile, bird, mammal, i.e. not fishes
nätting eller nejonögapirålhajrockahavsmusstörkvastfening bengädda bågfena, hundfisk elopidkattfisk, havskatt guldfisklaxfiskar, te.x. lax, regnbågslax, öring, röding, sikregnbågslaxiaxrödingöringsikgäddaströmmingtorsklaketonfiskabborrehavsabborrehelgeflundralungfiskstrålfenade fiskar, t.ex. bengädda, kattfisk, lax, torsk "rund"fenade fiskar, lungfiskar och kvastfeningar
vertebrate ryggradsdjurinvertebrate ryggradslösa djur

PublicationsThis thesis is based on the following publications and manuscript,which will be referred to by their roman numerals (l-V).
I . Andersson, E., Törmänen, V., and Matsunaga, T. Evolution of a Vh gene family in low vertebrates. Int. Immunol. 3:527-533, 1991.
I I . Andersson, E. and Matsunaga, T. Complete cDNA sequence of a rainbow trout IgM gene and evolution of vertebrate IgM constant domains. Immunogenetics 38:243-250, 1993.
I I I . Andersson, E. and Matsunaga, T. Evolutionary stability of immunoglobulin heavy chain variable region gene family: a Vh family can last for 150-200 million years or longer. Immunogenetics 41:18-28, 1995.
IV . Andersson, E., Peixoto, B., Törmänen, V. and Matsunaga, T. The evolution of the immunoglobulin M constant region genes of salmonid fish, rainbow trout (Oncorhynchus mykiss) and Atlantic salmon ( Salvelinus alpinus): Implications on the divergence time of species. Immunogenetics 41:312-315, 1995.
V. Andersson, E. and Matsunaga, T. Phylogeny of vertebrate immunoglobulin heavy chain variable region genes: Vh gene families of bony fish define independent clan lineages distinct from the three mammalian clans, (manuscript)
Published papers are reprinted with the permission from respective publishers.
5

1. Introduction
Ever since the multicellular organisms first appeared on earth, they have encountered the problem of pathogenic infections by various microorganisms. The development of effective defence mechanisms towards these microbes has been one of the major preoccupations of the host organisms. The complex and sophisticated adaptive immune system that has evolved in the vertebrates merely reflects the extent of constant battles fough t between microbes and host animals.
In most vertebrate phyla, adaptive immunity (or specific immunity) is characterised by specific responses involving antibody (ab) or immunoglobulin (Ig) and T cell receptor (TcR) molecules. These molecules are generated in a vast number of specific forms that enable them to recognise and bind molecules derived from microbes, generally called antigens (ag). For instance, if bacteria invade an animal, its immune system can recognise the signals from the foreign ag, select and amplify subpopulations of B lymphocytes that produce and secrete a large quantity of ab molecules specific for the ag of the invader (clonal selection). The ab molecules are effective in removal of bacteria from the body fluid by their ability to agglutinate bacterial cells leading to efficient phagocytosis by the host cells, or to lyse bacterial cells in collaboration with serum complement proteins (humoral immunity). The TcR molecules are expressed by thymus-derived cells (T lymphocytes) and are essential to eliminate intracellular pathogens (e.g. viruses) as well as to regulate the immune response by interacting with various immune competent cells (B lymphocytes, macrophages, other T lymphocytes). TcR molecules recognise microbe-derived peptides presented by the major histocompatibility complex (MHC) molecules that are expressed in all cell types of animals (MHC class I) or in certain cells in the immune system (MHC class II). The various types of cell-mediated immunity (e.g. graft rejection and delayed hypersensitivity) are the manifestation of T-cell responses.
The genes encoding the ab and TcR molecules constitute distinct and complex multigene families. Each gene member is splitted into gene segments that can somatically rearrange to generate diverse specificities for ag recognition (immunological repertoire or diversity). Although the capacity to generate an enormous ab and TcR repertoire is a necessity for the animal's survival, a certain portion of the ab and TcR specificities are bound to be directed against molecules of the host animals (self-ag) and
6

might cause autoimmunity. To avoid the problem of autoimmunity, the immune system has developed a number of mechanisms to discriminate self and non-self ag. There are mechanisms by which self-reacting lymphocytes are eliminated from the immune system (clonal deletion) or their activity is inhibited by other T cells (suppression).
The invertebrate immune system, on the other hand, seems to be constructed largely by non-specific immune responses (innate immunity). The innate immunity includes physical barriers (skin, mucous etc.), phagocytosis by macrophages and bacteria binding proteins and peptides (lysozyme, C-reactive proteins (CRP), hemolin, cecropin etc.) (1-3). Many of these non-specific mechanisms are present in vertebrates as well.
The enormous ab repertoire in vertebrate species has probably evolved to cope not only with the large number of diverse micro-organisms in the milieu, but also with the rapid antigenic adaptation of micro-organisms primarily due to their short generation time. In addition, some microbes have evolved highly sophisticated genetic systems which enable them to change their antigenic structure during an ongoing infection. For example, in the African trypanosomes such as Trypanosoma brucei, a single VSG gene expressed at a given time can rapidly be replaced by any of 1 000 different VSG genes, thereby changing its antigenic phenotype and evading the immune system (reviewed in 4). The somatic hypermutation of ab germline genes found in mammalian species can be understood in the context of this type of interplay between the hosts immune system and invading microbes.
On the other hand many ab genes and gene segments in the genome of animals (germline genes) seem to be more important to form the ab repertoire in lower vertebrates such as reptiles, amphibians and fishes (5, 6). This thesis is focused on the evolutionary aspect as well as genetic complexity of immunoglobulin genes of a teleost fish, rainbow trout ( Oncorhynchus mykiss, previously Salmo Gairdneri) (7).
1:1 The antibody molecule: structural features
The ab molecule has been studied in a variety of vertebrate species. It has a unit structure made of two identical heavy chains and two identical light chains held together by disulphide and non-covalent bonds (Figure 1, reviewed in 8). Each of the IgH and IgL chains are divided into a variable region and a constant region. The variable regions form the antibody
7

combining site that is responsible for recognition and binding of antigenic determinants or epitopes. The constant region of the heavy chain (Ch ) mediates different effector functions, such as complement fixation and binding to antibody receptors (or Fc receptors).
IgL IgH
I p a t
—I' 5i sapIA
_JO CH1
s-S-S
_
NXmlmU
wT -o
-T“
o
s-s
s-sf
frame work regions (FR)
hypervariable regions (CDR)
Figure 1. A schematic drawing of an IgM molecule. S-S denotes the disulphide bridges holding the H and L chains together. The domain structure and hypervariable regions are indicated in the figure. A tetrameric IgM characteristic for the bony fish secreted IgM is shown to the right.
The IgH and IgL chains are divided into several globular domains of 90 to 110 amino acids (aa). The IgL chain consists of one variable (V|_) and one constant (Cl ) domain, while the IgH chain is made of one variable domain and a varying number of constant domains (e.g. human IgM, 4 Ch domains; human IgG, 3 Ch domains (9); little skate, 2 Ch domains, (10)). Each domain is folded into two layers of anti-parallel ß-pleated sheets, one layer with four ß-strands and the other with three ß-strands. The two layers are held together and stabilised by an intra-domain disulphide bond, Figure 2 (9). This structure, also referred to as the immunoglobulin fold, is characteristic for the ab molecule as well as some other members of the Ig superfamily (e.g. TcR, MHC, CD4 and Thy-1, reviewed in e.g. 11). The IgH and
8
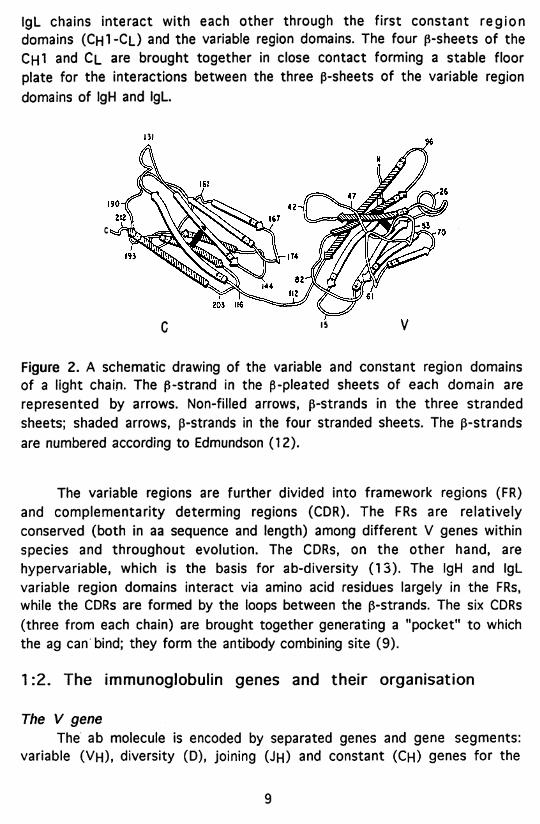
IgL chains interact with each other through the first constant region domains (Ch 1-C|_) and the variable region domains. The four ß-sheets of the Ch I and Cl are brought together in close contact forming a stable floor plate for the interactions between the three ß-sheets of the variable region domains of IgH and IgL
C is V
Figure 2. A schematic drawing of the variable and constant region domains of a light chain- The ß-strand in the ß-pleated sheets of each domain are represented by arrows. Non-filled arrows, ß-strands in the three stranded sheets; shaded arrows, ß-strands in the four stranded sheets. The ß-strands are numbered according to Edmundson (12).
The variable regions are further divided into framework regions (FR) and complementarity determing regions (CDR). The FRs are relatively conserved (both in aa sequence and length) among different V genes within species and throughout evolution. The CDRs, on the other hand, are hypervariable, which is the basis for ab-diversity (13). The IgH and IgL variable region domains interact via amino acid residues largely in the FRs, while the CDRs are formed by the loops between the ß-strands. The six CDRs (three from each chain) are brought together generating a "pocket" to which the ag can bind; they form the antibody combining site (9).
1:2. The immunoglobulin genes and their organisation
The V geneThe ab molecule is encoded by separated genes and gene segments:
variable (Vh), diversity (D), joining (Jh) and constant (Ch) genes for the
9

heavy chain and V|_, Jl and Cl for the light chain. Somatic recombination of the Vh-D-Jh genes and V l-JL genes during the B-cell differentiation form the complete variable regions of the IgH and IgL chains respectively (14). The V gene encodes the major part of the variable region, FR1-FR3 and CDR1 and CDR2, while the CDR3 and FR4 regions are encoded by D and Jh gene segments for the IgH and Jl for the IgL (14).
Studies of Ig molecules in a variety of different vertebrate phyla (including the cartilaginous fishes; sharks and skates) have shown that the overall genomic structure is conserved (Figure 3). The ab molecules might be absent in the cyclostomes (lampreys and hagfishes), the lowest representative of all vertebrates. The earlier claims (e.g. 15-17) that these species have ab molecules have not been supported by molecular cloning (18, 19).
TATA8mer box leader leader FRI CDR1 FR2 CDR2 FR3 7m er-9m er
+-D—H l 1 i m "H -Figure 3. A schematic figure of the genomic organisation of a single V gene. 8mer, octamer (5 'ATTTGCAT3 ); FR, frame work region; CDR, complementarity determining region; 7mer-9mer, heptamer-spacer-nonamer (5'CACAGTG-12/23 bp-ACAA AAACC3 ' ). Not drawn to scale.
Functional V genes encode an 18-22 amino acid leader peptide followed by the V gene segment, interrupted by an intron in between. The conserved regulatory regions (the octamer and TATA box) upstream of the V genes are found in all species except for most of the cartilaginous fishes (20-25). The conserved recombination signal sequences (RSS, the heptamer- 12/23 spacer-nonamer) are found in the 3 ' region of the V genes, with the exception of the germline joined V(D)J fusions found in the cartilaginous fishes (20). The D and J gene segments are also flanked by these sequences. The RSS are crucial for the rearrangement of V, D and J gene segments generating a complete variable region.
In contrast to the evolutionary conservation of each V gene, the genomic organisation of the IgH and IgL genes vary among vertebrate phyla as discussed below.
10
200
-1
OCX
) V
hs
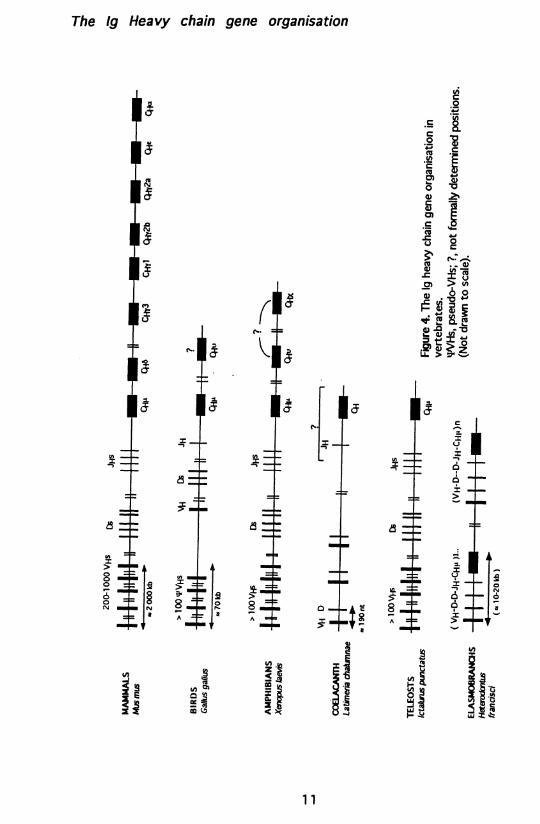
The lg Heavy chain gene organisation
11

In mammalian species, a large number of Vh genes are clustered together (spread over the chromosomal region of approximately 2000 kb in mouse) followed by small clusters of D gene segments (12 in mouse) and Jh gene segments (4 in mouse). The Ch genes are positioned, essentially as a single copy gene for each Ig class (e.g. m, ô, y, e, a in mouse), downstream of the Jh cluster, see Figure 4 (26). This IgH gene organisation will be referred to as the "mammalian type", which is also found in two other vertebrate phyla: amphibians (Xenopus laevis, 21) and several ray-finned bony fishes (channel catfish (Ictalurus punctatus 27); ladyfish (Elops saurus 28 ); Atlantic cod (Gadus morhua L. 29); Atlantic salmon (Salmo salar L. 30); longnose gar and bowfin (Lepisosteus osseus and Amia calva respectively 31)), thus representing the major evolutionary path of Ig gene organisation. The number of Vh genes varies from species to species, and can be divided into different families based on the DNA sequence identity between the Vh genes (>80% identity within a family). The mice Vhs are estimated to be more than one hundred, probably several hundred (32, 33), divided into 14 different Vh gene families (34-38, 197). In humans the number of Vhs is around one hundred divided into 7 families (39). The Xenopus and bony fishes seem to have eighty to a few hundred Vhs, also divided into several families (e.g. Xenopus: 11 Vh families (40); channel catfish: 6 Vh families (41, 42)).
A different type of IgH gene organisation is found in birds. In contrast to the many Vhs of the mammalian species, chicken has only one functional Vh gene located upstream of several D segments, one Jh segment and the Ch genes (Figure 4). However, about 80 pseudo-VH genes are found upstream of the functional Vh gene in the case of chicken (43).
The IgH gene organisation in the cartilaginous fishes differs dramatically from the mammalian type (44, 45). In these species, hundreds of unit clusters, each containing a single Vh, two D, one Jh and one Ch gene of Cn type, are repeated and spread throughout the genome, and is referred to as the "elasmobranch type" (Figure 4). The shark (Heterodontus francisci) has only one, or possible two, Vh family (20, 46). In rays, a second isotype (IgX or IgR) coupled to a second Vh family, not found in shark, has been identified (Raja erinacea) (10). The IgX genes are also organised as multiple clusters (47). The VDJ rearrangement is believed to occur only within a cluster (25). Another peculiarity of the cartilaginous fishes is the presence of already germline joined VDJ or VD clusters (20, 45) some of which are expressed (48).
12

The coelacanth, Latimeria chalumnae, may represent a fourth type of IgH gene organisation. In this lobe-finned fish (catfish, salmon, cod, ladyfish, rainbow trout etc. belong to the ray-finned fishes), Vh genes and D gene segments are separated by a small intron, thus resembling the elasmobranch type of IgH gene organisation. However, there are no Jh or Ch genes adjacent to the D genes, thereby excluding the elasmobranch type (49). Southern blot hybridisation indicates the presence of many Vh genes. Further studies are required to characterise this "coelacanth type" of IgH gene organisation.
The Ig light chain gene organisation
r r s m »N h|
BIRDSCallus galkis
VU VUt A VM JK
2SVVX*
ELASMOBRANCHS(Heterodontusfranósa)
( V L - J L * C L ) l „ . ( V L - J L - C l ) n
M- M JL( V L - J L - a ) l . . . ( V L - J L - a ) n M. a
I 1 —— »-1-1-■— - H— » I I I" -Cadus morhua Oncorhynchus myhss
Ictalurvs punctatus
Figure 5. The Ig light chain gene organisation in different vertebrates. wVXs, pseudo-VXs. (Not drawn to scale).
In most species the IgL genes are organised in the same way as the IgH gene organisation (Figure 5). This is true for the mammalian k light chain genes (50), bird X light chain genes (51, 52) and the two different IgL isotypes found in Xenopus (53, 54), the three different types found in elasmobranchs (type I, II and III) (23, 55-57). The mammalian X light chain locus is organised in a slightly different way. For instance, some laboratory mice strains have two clusters containing one \/X and two JX-CX complexes in the genome. The wild mice seem to have a more complex pattern (50, 58).
13

In contrast to the mammalian type of IgH gene organisation in channel catfish and Atlantic cod, the IgL loci in these species are organised in closely linked V-J-C clusters probably similar to the elasmobranch type (59-61).
1:3. Generation of antibody diversity
The ab repertoire is generated through several different mechanisms. The utilisation of these mechanisms can vary among vertebrate species and is to a certain extent influenced by the Ig gene organisation.
Combinatorial diversityAs previously mentioned, the complete Ig molecule is generated
through recombination events involving the V, D and J gene segments. This process is mediated by a complex enzymatic system involving recombinase (62, 63), which recognises the RSS (14). Two crucial components of the recombinase, RAG-1 and RAG-2 (recombination activating gene 1 and 2), have been isolated from cartilaginous and bony fishes (64, 65) indicating the evolutionary conservation of the recombinase system.
In mammals, amphibians and ray-finned bony fishes, Vh -(D )-Jh recombination as well as semi-random IgH/lgL pairing can generate several thousands of different combinations, contributing to the ab-diversity. If we assume that the IgH loci in mice contains 250 Vh , 12 D, 4 Jh gene segments, they can give rise to 250x12x4=1.2x104 different IgH chains. The IgL loci in mice have 2 Vk, 3JX, 250 Vk and 4 Jk gene segments, which can give rise to 2x3+250x4=1.0x103 different IgL specificities. Combination of different IgH and IgL chains would then give about 1.2x107 combinations. If we consider further diversity introduced by other mechanisms (see below), the potential ab repertoire in mice exceeds this estimate by the several orders of magnitude. The ab repertoire in lower vertebrates is smaller due to a number of reasons which are discussed later
In the cartilaginous fishes, the recombination is thought to occur within a cluster, which severely reduces the combinatorial diversity (25). This may also be true for the IgL in ray-finned bony fishes (61).
Gene conversionAs mentioned previously, chicken have only one functional Vh gene
which is recombined to one of the D gene segments and the single Jh gene
14

(V|_ and J|_ for the IgL). Ab diversity is then generated through gene conversion events, in which sequences of the upstream pseudo V hs or V ls donate to and modify the sequences of the functional Vh or V[_ genes (43, 52). This mechanism is also used in one of the mammalian species, rabbit, to generate the majority of the ab specificities despite the presence of many functional V h genes (66).
Gene conversion is also implicated in the homogenisation of V genes and other multigene families during evolution (see section 1:5). Whether gene conversion is used for diversification or homogenisation of multigene families seems to depend on various conditions.
Junctional diversityDuring the Vh-D-Jh recombination, nucleotides can be deleted due to
exonuclease activity (67), or added (68) in the Vh-Jh and/or D-Jh boundaries. There are two types of nucleotide additions: (A) non-templated sequences (N-sequences) which are added by terminal deoxynucleotidyl transferase (TdT) (68) and (B) template encoded additions (P-regions) (69). Low vertebrates use junctional diversity in a similar manner to the mammalian species (e.g. Xenopus, (21); Heterodontus, (20)).
Somatic hypermutationIn mammals, the ab-repertoire is further refined through a mechanism
introducing a high frequency of point mutations ( 10-3- 10-5 per base per cell division) in the variable regions of the ab-molecule. This phenomenon is called somatic hypermutation (70, 71). These ab-mutants are positively selected in specialised areas of the lymphoid organs, the germinal centres, and account for the 100-1000 fold increase in binding affinity during mammalian immune responses, i.e. affinity maturation (e.g. 72-74). Somatic hypermutations have also be shown to create the primary ab repertoire in sheep (75).
Somatic hypermutations of a similar rate to that in mammals have been observed in a low vertebrate, Xenopus (7-fold lower than the upper limit in mammals, (76). Despite this, Xenopus shows only weak affinity maturation (5) and low ab heterogeneity (77, 78). In higher vertebrates, the germinal centres are essential for selection of somatic mutants (reviewed in 79). The absence of these structures in low vertebrates has been proposed to explain the absence of significant affinity maturation during ab
15

responses in Xenopus (80). The physiological relevance, if any, of the hypermutation found in Xenopus remains to be resolved.
1:4. The fish antibody response
General features of the fish immunityAs in other vertebrates, the immune system in fish can be divided into
the non-specific defence (lysozyme, complement factors, interferon, phagocytosis by macrophages and non-specific cytotoxic cells, which are considered to be homologs of the mammalian NK cells) (81, 82) and the specific defence (B and T cells). The specific defence can be further divided into the humoral immunity (ab responses) and the cell mediated immunity characterised by phenomena such as graft rejection and mixed leukocyte reactions (MLR). The humoral responses, i.e. agglutination, complement mediated lysis and opsonisation mediated by ab molecules, have been shown in several different fish species (e.g. catfish, rainbow trout and shark) (81). The cell mediated immune defence has been studied through MLR (carp (83); catfish (84)) and mitogen stimulation. Different monoclonal abs have been used to characterise the cells involved in the adaptive defence (85, 86). Fish B and T cells can be distinguished by the following criteria: (A) The lg+ population, corresponding to B lymphocytes, is activated by the mammalian B-cell mitogen LPS (85), while the Ig* population, putative T cells, respond to ConA (87) and (B) Ab response to thymus dependent ag (TNP-KLH) requires both Ig* and lg+ lymphocytes and antigen presenting cells (APC, e.g. macrophages), while thymus independent ag (TNP-LPS) induces immune responses in the presence of lg+ cells and a small number of APC (86).
The MHC class I and II are essential molecules involved in the cellular immune responses and self-nonself recognition (88). The presence of MHC class I and II molecules in fish has been demonstrated both immunologically (MLR, graft rejections and through molecular cloning) (reviewed in 89, 90). The processing and presentation of ag during an immune response in fishes seems to be similar to the mechanisms of higher vertebrates (91, 92).
Recently the TcR a and ß genes have been isolated from shark and rainbow trout (93-95). This will provide good markers to study the function of fish T cells such as regulatory roles and cytotoxic activity mediated by T cells, TcR-MHC interactions, identification of accessory molecules (CD4, CD8) etc.
16

In summary, the major cellular and molecular players well characterised in the mammalian immune system (B and T lymphocytes, macrophages, APC, Ig, TcR, MHC) have also been shown to have homologs in the fish immune system above the level of cartilaginous fishes. As previously mentioned, there is no molecular evidence for the presence of ab as well as TcR molecules in the cyclostomes (lamprey and hagfish). There are, however, differences between the mammalian and fish immune system; the extent of ab-diversity, the absence of ab maturation (see below) and the absence of Ig classes other than IgM (see below).
Fish antibodiesThe fish ab proteins have been isolated and characterised from a
variety of fishes: horned shark, channel catfish, rainbow trout, Atlantic cod etc. (reviewed in 96, 97). The abs are of the IgM class and consist of 70 kD IgH chains and 22-25 kD IgL chains. Membrane IgM (mlgM) has two IgH and IgL chains (n,2L2), while the secreted IgM (slgM) forms polymers with several binding sites (Figure 1). The bony fish slgM occurs as tetramers, while pentameric slgM is present in the tetrapods (mammals, birds, amphibians) and cartilaginous fishes. In mammals several other Ig classes are known (IgD, IgG, IgA and IgE) and some of these are also found in other tetrapods such as birds (98) and amphibians (99-103). In cartilaginous fishes, a completely different isotype has been described, IgX or IgR (10, 104). Two non-IgM classes are found in a lobe-finned bony fish (lungfish) (96). In the ray-finned bony fishes, IgM is the only class fully characterised to date. However, some ray-finned bony fishes possess low molecular weight serum Igs corresponding to monomeric forms of IgM or a smaller form of monomeric IgM (97, 105). Whether the smaller forms of IgM are encoded by another gene or are the result of post-transcriptional modifications is not known.
Whether the ray-finned bony fishes have different classes or isotypes other than IgM remains to be seen. For example, a serological study suggested that the channel catfish has a different Ig class (106). However, this conclusion has not yet been supported by unambiguous biochemical studies or molecular cloning. More interesting is the recent finding of a gene encoding a non-(V molecule in the channel catfish (Dr. Melanie Wilson, personal communication). This gene is located downstream of the gene and is expressed through alternative splicing. A phylogenetic analysis suggests that this catfish Ig gene may be related to the mammalian Cô (IgD) genes.
17

Another peculiar feature of the bony fishes is the lack of the fourth constant region domain (Ch4) in membrane IgM (mlgM). This is due to an unusual splicing of the transmembrane exons (29, 30, 107; see section 3:4).
Fish antibody responseThe primary ab responses in fish is characterised by the production of
low affinity IgM abs (105, 108-111), as is seen in mammals. The secondary immune response in warm-blooded animals is much faster than the primary response and is characterised by increased titres of high affinity abs of non-IgM class (affinity maturation and class switch) (112). Fishes, on the other hand, do not seem to have affinity maturation (111, 113, 114). However, under certain conditions (e.g. type and dose of ag, route of immunisation), bony fishes give a significantly elevated ab response upon a secondary antigenic challenge (114-116). Vaccination trials have shown that fish species can develop protective immunity against certain bacteria (117). However, this immunological memory does not seem to sustain for a long period of time ( 6 months up to a year) (e.g. 118). The poor ability in fish to generate an enhanced secondary response may be due to the lack of other isotypes equivalent to mammalian IgG and/or no selection for high affinity abs. In addition, isoelectric focusing measurements (IEF) of the ab heterogeneity show that the ab repertoire in fish is much smaller compared to that of mammals (119, 120).
1:5. Evolution of multigene families
Definition of a multigene familyA multigene family is a group of nucleotide sequences or genes
characterised by multiplicity (ten to millions of copies), close linkage, sequence homology and overlapping or related functions (if any). Some of these families are without any known biological function, e.g. repetitive DNA such as Alu-families (121), while others are genes crucial for the animal's survival; ribosomal RNA (rRNA), transfer RNA (tRNA), histone 2A and 2B etc (e.g. 122-124).
The IgV multigene family is another example of a biologically important gene family (124). However, in contrast to other multigene families which are characterised by sequence identity or homogeneity among individual members, the IgV genes are diverse as well as conserved in sequence (CDR versus FR). To understand the evolution of the IgV multigene family, the following questions are of paramount importance: (1)
18

What mechanisms diversify the CDRs and conserve the FRs? (2) What selective force(s) is responsible for the V gene diversity during evolution and how does it act?
Evolutionary mechanisms acting on multigene familiesMechanisms such as gene duplication by unequal crossing-over and
gene conversion have been invoked to explain the sequence homogeneity of family members such as rRNA and tRNA (124-126). Homogenisation, also referred to as coincidental evolution (124) or concerted evolution (127), occurs constantly in these multigene families ensuring identical or near identical sequences of each member, counterbalancing mutations that diversify sequences.
During the homogenising process a rare mutant can spread and become the dominant member in the family of a species. This process can be identified by the presence of "species-specific" aa residues or sequences often found in multigene families (126, 128). Species-specific residues have also been identified in the IgV FRs of several vertebrate species (mammals (129-131); Xenopus, Caiman, rainbow trout and horned shark (24)), indicating that homogenising processes operate on the IgV multigene family as well. How is homogenisation of FRs and diversification of CDRs balanced during evolution of IgV genes? It has been suggested that the homogenising process might be slow, and the CDRs can be diversified through random mutation and positive Darwinian selection (132, 133).
1:6. The molecular clock of proteins
The phylogenetic relationship between different species has always intrigued scientists. Fossil records combined with the knowledge of comparative anatomy and physiology have been important sources for the study of phylogeny of species. It has, however, often been difficult to produce a clear picture of animal's evolutionary history due to lack of fossils. The development of molecular techniques introduced new tools for evolutionary studies. It became possible to compare amino acid and nucleotide sequences of homologous genes between species and to study phylogeny more quantitatively. These studies showed that for a given protein the number of aa substitutions between two species increases approximately linearly with the divergence time known from fossil records. In other words, the rate of aa substitutions is largely constant for a given
19

protein or gene and independent of the evolutionary lineages of species. This phenomenon was called the molecular clock (134, 135).
The neutral theory of population genetics (or neutral mutation-random drift hypothesis) (136) provides the explanation for the observed constancy of the evolutionary rate. In this theory, the majority of aa substitutions during the course of evolution is a product of random fixation by neutral or nearly neutral mutants rather than Darwinian selection. Consequently, the maintenance or disappearance of a new mutant allele, equally good as preexisting alleles, in a population is left to chance. This theory showed that if a mutant allele is selectively neutral or nearly so, the substitution rate per generation in a population is equal to the mutation rate per gamete per generation independent of the population size. Therefore a constant evolutionary rate of a protein can be observed. The Darwinian selection, on the other hand, seems preoccupied with the conservation of the functional region(s) of proteins found in modern species.
The molecular clock can be used to obtain approximate estimates of the divergence time between different species. To study phylogeny and evolution several mathematical algorithms and statistical methods have been developed (136, 137). We have used some of these to study the evolution of IgVH and IgCH genes (paper II and IV this thesis).
1:7. Fish phylogeny
The fishes represent a major group of modern vertebrate phyla; approximately 25 000 different fish species are known today (138). This phylum constitutes very heterogeneous groups of species living in various habitats influenced by different environmental factors such as temperature, light, pressure, salinity etc. Most of the different fish species (95-97%) belong to the group of modern bony fishes or teleosts (e.g. rainbow trout, catfish, goldfish, cod). The remaining species are represented by one major group, the Chondrichthyes (or cartilaginous fishes; shark, ray, ratfish, chimeras), and some minor groups such as Sarcopterygii (lungfish, coelacanth), Chondrostei (sturgeon, paddlefish), Amia (bowfin), Ginglymodi (gar) and Cyclostomes (lamprey, hagfish). These fishes, and also the ancestor of tetrapods, evolved during the Devonian era (350-400 my ago, Figure 6).
20
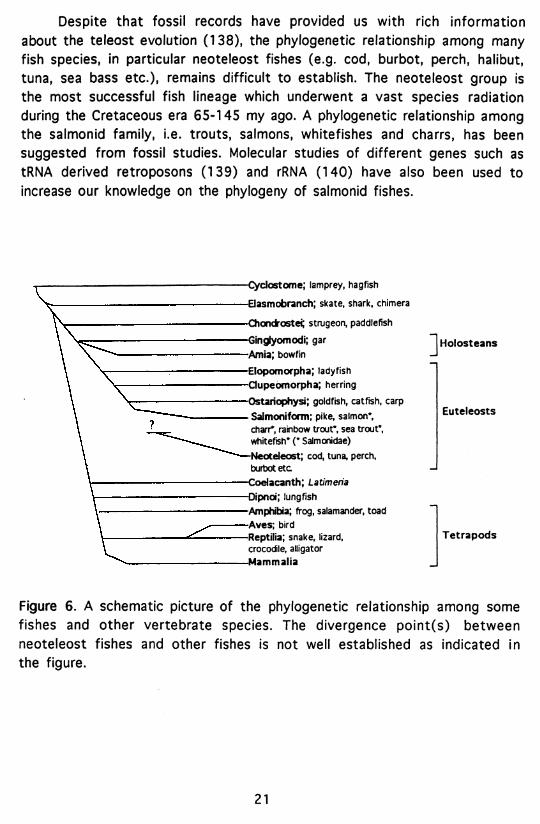
Despite that fossil records have provided us with rich information about the teleost evolution (138), the phylogenetic relationship among many fish species, in particular neoteleost fishes (e.g. cod, burbot, perch, halibut, tuna, sea bass etc.), remains difficult to establish. The neoteleost group is the most successful fish lineage which underwent a vast species radiation during the Cretaceous era 65-145 my ago. A phylogenetic relationship among the salmonid family, i.e. trouts, salmons, whitefishes and charrs, has been suggested from fossil studies. Molecular studies of different genes such as tRNA derived retroposons (139) and rRNA (140) have also been used to increase our knowledge on the phytogeny of salmonid fishes.
Cydostome; lamprey, hagfish
Elasmobranch; skate, shark, chimera
Oiondrostei; strugeon, paddlefish
Dinglyomodi; gar ■Amia; bowfin
Elopomorpha; ladyfish CJupeomorpha; herring
Ostariophysi; goldfish, catfish, carp Salmonifocm; pike, salmon*, charr*, rainbow trout*, sea trout*, whitefish* (* Saimonidae)Neoteleost; cod, tuna, perch, burbot etc
Coelacanth; Latimeria
Dipnoi; lung fish Amphibia; frog, salamander, toad Aves; birdReptilia; snake, lizard, crocodile, alligator Mammalia
]Holosteans
Euteleosts
Tetrapods
Figure 6. A schematic picture of the phylogenetic relationship among some fishes and other vertebrate species. The divergence point(s) between neoteleost fishes and other fishes is not well established as indicated in the figure.
21

2. Aims of this thesis
As previously mentioned, the Ig genes (V, D, J and C) and the diversifying mechanisms generating the ab-repertoire have been extensively studied in several tetrapod species, e.g. mice, humans and Xenopus (e.g. 14, 76). The fundamental question of how these Ig genes and diversifying mechanisms have evolved remains, however, to be elucidated. The aim of this thesis is to increase our understanding of the evolution of the IgVH multigene family, focusing on the comparative study of the IgH genes in a low vertebrate, rainbow trout (Oncorhynchus mykiss).
22

3. Results and Discussion
When I initiated my work, the first genomic teleost IgVn gene, RTVH431, had already been isolated from rainbow trout (24). This Vh gene has the structural features found in all functional IgVH genes (Figure 3), and shows 40-60% aa homology with Vhs from various vertebrate species. Southern hybridisation analysis shows that the RTVH431 family has approximately 20 members in the rainbow trout genome. The evolution of this Vh gene family was studied through Southern blot hybridisations using DNA from various fish species (Paper I). The evolutionary study focusing on the rainbow trout IgH genes was further extended by the characterisation of a complete IgM heavy chain cDNA (Paper H) and extensive comparison of IgVH cDNAs (Paper III, V). In addition, information concerning mechanisms generating ab-diversity, IgH organisation and unusual RNA splicing of mlg in rainbow trout, evolution of IgM constant domains and information about the divergence time between fish species was obtained. The divergence time of salmonid fishes was further studied using IgM intra-domain introns of rainbow trout and Arctic charr (Salvelinus alpinus, Paper IV).
3:1. Rainbow trout IgVH genes and family structure
In order to investigate the IgVH gene complexity of rainbow trout and its evolution, many V h genes were isolated and characterised (27 cDNA V h s , Paper II, III; two genomic IgVn genes (24); Paper I). According to the border criterion of 80% nt identity for V gene family classification (34), these 29 Vhs could be divided into eight or nine different families (figure 2 in Paper III). At about the same time a French group published a similar study of the rainbow trout Vh gene complexity, using three sequences from our group (24; paper I, II), one sequence from Lee et al (141) and seven additional Vh sequences (142), in which they identified nine Vh families.
To further clarify the number of different Vh gene families in rainbow trout, we made a thorough comparative analysis of all published rainbow trout V h s using Jukes-Cantor distance matrix (143) in PHYLIP package version 3.5c by Felsenstein (144). We came to the conclusion that rainbow trout has at least 11 different Vh families. Consequently, we jo intly proposed the following nomenclature for the IgVH families, Onmy Igh-V 7 to Onmy Igh-V 7 7 (after Oncorhynchus mykiss, table 1). The number of Vh families may increase if more genes are isolated. For instance, the lgh-V4 gene is an "outcast" gene in the Onmy Igh-V 7 family, and isolation of
23

additional genes might separate this gene into its own family. Southern blot hybridisation using family specific probes detects several unique bands for some of each family (4-20 bands, unpublished results). We speculate that there are 100-200 germline Vhs which can contribute to the ab-repertoire in rainbow trout. However, this method is crude in that it may also detect pseudo-VHs, and does not give a precise estimate on the functional germline Vh genes.
Family Previous family Reference(new nomenclature) name
Onmy Igh-V 1 RT VH la Paper IIIlgh-V4 Roman and Charlemagne (142)
Onmy Igh-V 2 RT VH VI Paper IIIlgh-V2 Roman and Charlemagne (142)
Onmy Igh-V 3 RT VH VIII Paper IIIlgh-V5 Roman and Charlemagne (142)
Onmy Igh-V 4 lgh-V6 Roman and Charlemagne ( 142)Onmy Igh-V 5 RT VH lb Paper III
lgh-V7 Roman and Charlemagne ( 142)Onmy Igh-V 6 RT VH II Paper III
lgh-V8 Roman and Charlemagne (142)Onmy Igh-V 7 lgh-V9 Roman and Charlemagne ( 142)Onmy Igh-V 8 RT VH III Paper IIIOnmy Igh-V 9 RT VH IV Paper IIIOnmy Igh-V 10 RT VH V Paper IIIOnmy Igh-V 17 RT VH VII Paper III
Table 1. Nomenclature of the rainbow trout Vh sequences. The eleven Vh families are designated Onmy Igh-V 7 to Igh-V 7 7 (manuscript submitted)
3:2. Evolution of Vh genes and their families
Although evolution of the IgVs has been extensively studied using mainly mammalian IgVs (e.g. 35, 38, 132, 145), information on the IgVs from low vertebrates may highlight new aspects of this issue. We carried out extensive analysis of the rainbow trout Vhs in order to study several aspects of IgVH gene evolution: (A) How long can a Vh family last during evolution? (B) How is the bony fish related to the mammalian Vh clan I, II and III? (C) What is the role of homogenisation for Vh family evolution?
A. Stability o f a Vh gene familyComparative analysis of mouse and human V genes has shown that
some V gene families share >80% nt identity, suggesting that a V gene
24

family can last for at least 65 my, the divergence time between mouse and human (35, 132, 145, 146). Can an IgV family last longer than 65 my? If so, how much longer? To investigate these questions, we made extensive interspecies comparisons between Vh genes from rainbow trout and channel catfish (Table 1 in Paper III). Several of the rainbow trout and catfish Vh gene families show over 70% nt identity (70.8-77.4%).
More striking is the nt identity between Onmy Igh-V 9 (previously RTVH IV, Paper III; Table 1) and catfish VH I family of over 80%. We assume these two Vh families evolved from a common ancestor gene(s) at the time of divergence of rainbow trout and catfish. Using the constant substitution rate of amino acid residues in lgM-CH4 domains (see section 3:5), we could estimate the rainbow trout-catfish divergence time to at least 150-200 my (Paper II and III). Consequently, we concluded that a Vh gene family can last for at least 150-200 my in two separate species.
The above study is based on cDNA sequences. It has been shown in Xenopus that somatic mutations of Vh genes can give rise to a range of 3% fluctuation (76). However, accounting for this range of fluctuation in our studies does not significantly affect our conclusion.
In Paper I, genomic DNA from salmonid species (rainbow trout, Atlantic salmon, Arctic charr, sea trout and whitefish) as well as distantly related fishes (pike, goldfish, herring, Atlantic cod, burbot and sea lamprey) was hybridised under stringent conditions with a rainbow trout Vh gene, RTVH431, as a probe. As expected several strong hybridisation bands (7 to 25 bands) were observed from the closely related salmonid fishes. We also observed consistent signals (4 to 10 bands) from pike, goldfish and herring, which had diverged from the salmonid fishes probably 100-200 my ago (138). In accordance with the hybridisation signals from goldfish DNA, a goldfish Vh gene (99A) shows an average of 80% nt identity with the rainbow trout Onmy Igh-V 9 family (previously RTVH IV, Paper III; Table 1). This goldfish Vh gene also shows >80% nt identity with the VH I family of the more closely related channel catfish. In summary, these Southern hybridisation results are compatible with our conclusion that a Vh family can last for at least 150 my.
Two plausible explanations for the conservation of V h families are: (A) certain Vh families have important functions and they have been conserved during evolution, or (B) all V h genes evolve more or less with a
25

constant substitution rate (approximately 1% DNA change/10 my based on the estimated life time of a V h family, Paper III). The first possibility was suggested to explain the 70 % nt homology between an ancient reptile, Caiman, and a mouse SI 07 V h gene (147), as well as the cross-hybridisation of mouse Vh probes with DNA from other species (148). The SI 07 Vh family belongs to the mammalian clan III (see next section). Some members of the clan III families (e.g. 7183) are preferentially expressed in the early stage of immunological development (149) and may have a special role in the establishment of the immunological repertoire (reviewed in 150). For this or other reasons the clan III V hs may have been evolutionary conserved. The alternative possibility is that the ancestral clan III V h s were present before the divergence between bony fish and tetrapods, and these V h s have evolved with a relatively constant rate (approximately 1% DNA change/10 my). Thus, the average 60% identity between rainbow trout-mouse V hs
(table 2 in Paper III) and the 70% identity between Caiman-mouse V h s , may merely reflect the evolutionary time passed since the species diverged (rainbow trout-mouse, 400 my; Caiman-mouse, 300 my) (138). These two possibilities are not mutually exclusive. We have proposed another explanation for the stability of a V gene family which will be discussed later in this section.
B. Evolution of vertebrate Vh clans: identification of a te le o s t and an archaic clan separated from the mammalian clans
As mentioned above, the mammalian V h families are further divided into different clans (I, II and III), which are defined by intervals in the FRI and FR3 (146) (see Figure 1 and 3). It has been suggested that these clans represent the major lineages in mammalian V h evolution. A more recent phylogenetic study (151) showed that there may be five lineages (Group A-E) of vertebrate V h families; three lineages mainly defined by mammalian V hs (clan I, II and III) and two other lineages constituting V hs from either bony fishes or cartilaginous fishes. Due to the small number of bony fish Vh families used in their study, it remained unclear whether the two fish lineages can be considered as independent clans and if they have Vh families from other vertebrate phyla. Therefore, we made a more thorough analysis including the 11 rainbow trout V h families as well as V h s from several other bony fishes.
The obtained phylogenetic tree is characterised by 5 major lineages, which largely overlap with those of Ota and Nei (151) and are therefore named Group A-E (Figure 1, Paper V). In summary, Group A, B and C cover all
26

the mammalian V h s , and correspond to the mammalian clan I, II and III respectively. Most of the amphibian Vhs are also found in these groups. In addition, V h s of bird, reptile and some of the bony fishes are found in Group C. Our study clearly confirms that Group D is a bony fish specific V h lineage; most of the bony fish V h s are found in this group which can be divided in two sublineages. The earliest branching lineage in our (as well as Ota and Nei's) tree is Group E, which has all V hs of the cartilaginous fishes. Our study shows, however, that several minor sub-groups of rainbow trout and amphibian V h s (XIVH8-Re20; AmlghVC9; RTVH24-RTVH2) are found in this group as well. This means that some of the bony fish Vh still retain sequences related to cartilaginous V h s . In addition, the two amphibian V hs
(XIVH8, AmlghVCI 9), the rainbow trout Onmy Igh-V 3 family and the skate Re20 VH codon 75 and codon 77 are deleted (Figure 4b, Paper V), indicating that these genes can form a sublineage with the cartilaginous families (e.g. Hf2809). Note that Group E has some irregularity in that branches do not form a single node. This may be due to the extent of diverged sequences in this group.
As the mammalian clan I, II and III are split up in Group A, B and C respectively, we asked if the two non-mammalian groups D and E could also be defined as clans. To address this question we constructed a phylogenetic tree using the codon 6-24 of FRI and codon 67-85 of FR3, which are the intervals defining the mammalian clans (146). Except for minor alterations within each major Group, the obtained tree is similar to the tree described above. It can be concluded that Group D and E are clans made of independent, non-mammalian lineages.
In conclusion, there are five major V h lineages or clans: Group A and B (or clan I and II), two tetrapod-specific lineages; Group C (or clan III), a lineage with V h s from all phyla except cartilaginous fishes; Group D (or teleost clan), a bony fish-specific lineage and finally Group E (or archaic clan), a lineage with V h s from cartilaginous fishes, bony fishes and amphibians.
Based on the phyla within each group and the ir evolutionary relationship, a possible order of appearance of each clan can be described. The archaic clan existed in the cartilaginous fishes before the divergence to bony fishes and tetrapods (450-500 my ago). The presence of bony fish and amphibian V h s in the archaic clan supports this point. The second clan to appear is clan III, whose members are found in bony fishes as well as
27

tetrapods, which diverged about 350 my ago. The teleost clan evolved at some point during the teleost evolution. Clan I and II appeared in the tetrapod lineage before the divergence of amphibians, reptiles, birds and mammals. During the evolution of different tetrapod phyla, certain clans have been lost. In birds, which only have one or a few functional Vhs, all clans except clan III have been lost. One might speculate that birds have eliminated all unnecessary Vhs. in the mammalian lineages, the archaic clan seems to have disappeared. It is clear that although a Vh family may last 150-200 my, some have been lost, probably due to contractions, during 450- 500 my of vertebrate history.
C. Species-specific amino acid residues: implication on the IgV gene evolution
Species-specific residues, i.e. markers for the homogenising process, have previously been identified in mammalian IgV proteins (124, 129-131) and more recently in V hs from lower vertebrates, including two V h s from rainbow trout (24, Paper I). We searched for additional rainbow trout-specific residues among the members of the 11 Onmy Igh-V families.Several rainbow trout-specific residues were found mainly in the FRI and FR3, but also in the CDR2 (boxed and shadowed in Figure 3 of Paper III, unpublished observation). These residues are clearly spread to most of the members within a Vh family but not to members in other Vh families, indicating that homogenising mechanisms are involved in shaping IgV gene families. "Species-specific" is a relative and conventional term. If the same residues are found in related species, they should perhaps be called lineage specific residues. For example, two residues (Met 10, Gin 83) occur only inthe rainbow trout Onmy Igh-V 6 (previously RTVH II, Paper II; Table 1) andthe channel catfish VH II families, which are very similar to each other (77.4% identity).
in summary, species-specific residues are found in the IgV proteins of all vertebrates, showing that homogenising processes have been operating during 450 my of vertebrate evolution. These mechanisms explain the sequence homogeneity of FRs, the conserved regions of the Vh where most of the species-specific residues are localised. The presence of such residues in the CDRs (34 Asn of CDR1 in RTVH431 and RTVH253 (24, Paper I); and 62 Val of CDR2 of Onmy Igh-V 8 (previously RTVH III, Paper III; Table 1)) suggests that homogenisation also acts on the CDR regions. This is not compatible with the general belief that the CDRs are highly variable in sequence and structure. It has, however, been suggested that only a small
28

number of aa in the CDRs are highly variable and in direct contact with the ag (152, 153). In addition, studies of mammalian IgVH CDRs showed a very limited number of CDR conformations, so called canonical structures (154, 155). Some of these canonical sequences are also present in fishes and amphibians (Paper I, 156, 157), suggesting that some structures of the combining site have been conserved over 450 my. Thus, homogenisation of the CDRs may play some role in the conservation of the general structure of the CDRs.
Significance of the Vh family: a speculationA Vh family can be stable for 150-200 my in evolution. What functions
may these Vh families then have, if any?
Because microbes can rapidly change their antigenic structure, the host immune system must find strategies to counteract the effects of these changes. One way may be to use ab specificities directed against relatively conserved epitopes of the micro-organisms. Certain Ig V genes may therefore have been selected for binding to these conserved ags (158). Effective selection may act on a Vh family as a whole, since the members are very similar to each other (80-100% identity) and have overlapping ag binding specificities. A Vh family could thus "help" an organism to maintain certain ab-specificities (Paper V): (I) The redundancy of specificities would protect against loss of a certain specificity through random mutations. (II) Gene conversion by remaining members within a family (159) could rescue mutationally defected genes. Mutation at certain critical positions of the CDRs (152, 154) would be enough to recover the original specificity. (Ill) Contraction of the Vh repertoire by unequal crossing-over may occasionally take place (148), and this event can be catastrophic for the animal. Other Vh families with overlapping ag-specificities may compensate for this loss.
In other words, we speculate that a Vh family may function as a buffer against deleterious processes acting on the V h s during evolution, and important for the maintenance of certain ab-specificities.
3:3. The IgH diversity in rainbow trout
Ab-diversity is fundamental for the survival of vertebrates, and is generated through several mechanisms: V-(D)-J recombination, junctional diversity and somatic (hyper)mutation (see section 1:3). The extent of sequence variation of different V, D and J germline gene segments are
29

primarily important for ab-diversity in low vertebrates. Our rainbow trout V h cDNAs containing Vh -D-Jh sequences provide essential information concerning the contribution of these gene segments to the diversity in the IgH variable region.
Vh d iversityAs discussed above, the rainbow trout has 11 Vh families and maybe
as many as 100-200 genomic V h s . Another bony fish, the channel catfish, has 6 families and approximately the same number of germline V h s (41, 42). Thus, the number of V hs contributing to IgH diversity seems to be similar in some bony fishes and some mammalian species.
Our sequence analysis of 40 Vh cDNAs gives the impression that the CDR1 and CDR2, which are part of the ab combining site, are relatively homogenous within a Vh family. If this is a common feature of the rainbow trout Vhs, the diversification by the Vhs would decrease. A similar tendency has also been observed in another low vertebrate, Xenopus, in which IgH diversity seems to be confined mainly to the CDR3, i.e. the Vh-D- JH junctions (160). Therefore it is also important to know the CDR3 diversity of Vh-D-Jh junctions in rainbow trout, this is described below.
D-Jh sequences and CDR3 diversityEight different rainbow trout Jh gene segments could be identified
through sequence comparison of trout Jhs and Jhs of other species (Paper III). The rainbow trout Jhs also have the evolutionary conserved region (Trp 103-Gly 104-Gly 106-Thr 107) found in most Jhs (161). There are, however, two exceptions (cRTVH4.1 and CRTVH11), in which Trp 103 and Gly 104 residues have been substituted to Ser and Pro respectively. These aa are not seen in any other vertebrate Jh segment and may affect the Jh-JL interaction in a negative way and not be expressed as intact IgM. Therefore, these two peculiar Jhs will be excluded in the following discussion. Another study reported the presence of six Jhs in rainbow trout (142, 162). Sequence comparison between our Jhs and their Jhs revealed eight different Jh gene segments, Igh-JA to \gh-JH according to their nomenclature (Figure 7). The Jhs isolated by two other groups belong to one of these eight Jh segments (Igh-JD (141); Igh-J (163)). Out of the total 122 Vh-D-Jh clones isolated, Igh-JD and Igh-JB seem to be preferentially used, while Igh-JE and Igh-JF segments occur less frequently. Four of the Jh segments were each used once or twice (the frequencies are shown in Figure 7). The most frequently
30

used Jh segments recombine randomly with Vh gene independently of Vh family.
Igh-J cons TTT GAC TAC TGG GGG AAA GGG ACA ATG GTC ACC GTT TCA TCAIgh-JA — C ------------------- G — A T-C C A A G A-A A-T — T 1/120Igh-JB — C --------------------------- C C A A G A-A A-T — T 30/120Igh-JC T -------T C-G — T ----- A — A ... 1/120Igh-JD C ------G -------G — C A— 45/120Igh-JE C ----------T ---------------- 22/120Igh-JF T — T ----------- 17/120Xgh-JG C ----------T AC 2/120Igh-JH------ C-------- T C-G — T A — A ----------- T — T 1/120
Figure 7. The nucleotide sequences of the eight rainbow trout Igh-J segments. The Igh-J segments correspond to amino acid residues 100 to 113 (FR4, Kabat et al, 1991). (-), identical residues; (.) absence of the last codon in Igh-JC. The Igh-JA-lgh-JF are from Roman and Charlemagne (142), the Igh-JG and Igh-JH are from Paper III. The frequency of each Jh is summarised and shown in the right (combined results from Paper III, 141, 142, 162, 163).
The Jh encodes the FR4 and is involved in the interaction between IgH and IgL chains. This region is, therefore, relatively conserved and does not make any major contribution to the diversity of the ag combining site.
In contrast to the Jhs, the D gene segments encode parts of the CDR3, a diverse region created when the Vh, D and Jh segments are combined. Furthermore, nt can be added or deleted in the junctions between each segment, i.e. N-diversity. Since no germline D gene segments have yet been isolated from rainbow trout, it is difficult to define distinct D segments. We could, however, identify four recurring sequences in the CDR3 of our Vh- D-Jh cDNAs (boxed in Figure 4, Paper III), which are candidates for separate D gene segments. In a more recent report, CDR3 regions of 88 rainbow trout Vh-D-Jh cDNAs were analysed (162). Prototype D segments were defined as sequences occurring in at least two clones, with a minimum of five consecutive nucleotides. They could identify ten prototype D gene segments ranging between 5-16 nt (DH1-DH10). As expected, some of our CDR3 regions overlap with some of their D segments, further supporting that these prototype D segments have genomic counterparts (Figure 8). Ten of our clones do not belong to any of the prototype D gene segments and may represent yet unidentified D gene segments.
31

In some vertebrates, two additional mechanisms are used to further increase the CDR3 diversity, namely D-D fusions and inverted D segments (e.g. 164, 165). Our results together with the results from Roman et al. imply that D-D fusions can also occur in rainbow trout (a putative DH1-DH2 fusion in cRTVH9, Figure 8; DH2-DH2 in the clone 85X3 (162)). Since no germline D gene segments have been isolated, we can not conclude whether or not inversions occur. The flanking regions of these putative Ds vary considerably in length and sequence (Figure 4, Paper III), suggesting that the mechanism for N-diversity is also present in rainbow trout.
N ND H l CAGAATAACGGC9 G . . ------------------• AGT AC AAC T AC *50 GTC • . . ----------------.GGCT
DH2 ATAACTACGGA6 GTGGG. • . . ------------• CTCCTACCGT8 TACT- . . ------------. . CCGCT9 GGAATAACGGAGTAC. . ------------*14 CTCGG------ ---------- • . ATGCT21 AACGAC.. ------------
DH3 TCGGTACACTGGGAGT3 --------- . CCGGGGTCCCCGCTATATCGAG255 GATTG.................... ...............-CAGCTAC1 CGAGCT.................. ................. -CCTA
DH4 AATAGCGGGTACCC271 GAGTATCC.. . ----------------------4 AGTCCAT.---------------------- • - TA
DH8 ATGGGGGC25 AAGGGAG. --------------GC TTC ACTAGCT22 G----------------AGCTAC24 ----------------GAACTAC
DH9 AACTAC GGGGGC7 CTAT....................... • • TACCGT11 GAG.. -------------------ATTGCT12 GGGCGACAA--------------------- . GGGTC GAATGCT15 GG---------------------- . TCCTACAACTAC16 A. . ------------------. GTGCT17 GGAGTTT • • --------------------CCACAACTAC26 GGGGCT. ----------------------CGATTACTAT
Figure 8. Nucleotide sequences of prototype D gene segments; DH 7, DH2, DH3, DH4, DH8, DH9 (162), are aligned with CDR3 regions (the numerical name 9 correspond to cRTVH9, 50 to CRTVH50 etc) from Paper III. The nucleotide sequence between amino acid position 94 and 100 (Figure 3 in Paper III) are shown. Hyphens (-) indicate identity; dots (.) ind icate deletions in the 5 ' and 3 ' of the D segments; N, N-region sequences. The putative D-D fusion in clone 9 is marked with a star (*).
32

We conclude that the rather extensive CDR3 diversity in rainbow trout is generated by mechanisms known from other vertebrates: (A) random recombination of several Vh , D and Jh gene segments, and (B) junctional diversity (i.e. deletion and /or addition of nucleotides).
The antibody response in fish and other low vertebrates-Why is it so restricted?
In mammals, the ab-response is very heterogeneous (e.g. 500 IEF spectrotypes in mouse ab responses against DNP) (166, 167) and undergoes several changes with time (isotype switch, a ffin ity maturation and generation of memory B cells). In contrast, low ab-heterogeneity (e.g. anti- DNP ab responses in different low vertebrates are as follows: 40 IEF spectrotypes in Xenopus (78); 10-30 IEF spectrotypes in bony fishes (119, 120)); a marginal increase, if any, in affinity and a poor generation of memory B-cells characterise the ab-response in fishes and amphibians. Several parameters have been proposed to explain these differences.
For example, the presence of multiple V-(D)-J-C clusters (Figure 4) in cartilaginous fishes has been invoked to explain the limited ab-repertoire in these species. The V(D)J recombination occurs within a cluster, thus eliminating the combinatorial diversity (e.g. 46). The low sequence heterogeneity between different V h , D and Jh gene segments and germline joined clusters in these species (e.g. 20) further limits the ab-diversity. The V l -Jl germline joined segments are, however, relatively heterogeneous in sequence, especially in the CDRs (48, 168).
On the other hand, in ray-finned bony fishes the number of Vh, D and Jh gene segments is comparable with that of mammals. In addition, V(D)J recombination and junctional diversity mechanisms seem to work well in these low vertebrates. Despite these the ray-finned bony fishes show a restricted ab-diversity. This is perhaps explained by the following reasons: (I) The germline V genes, including the CDR1 and CDR2, show limited sequence heterogeneity in Xenopus and maybe this is the case with bony fishes (160, 164, discussed above). (II) It has been suggested that the very diverse CDR3 regions found in mammals are essential for the ag recognition (154, 155, 169). The generally shorter IgH CDR3 regions in low vertebrate may reduce the ab-repertoire (162, 170). (Ill) A marginal degree or absence of affinity maturation in low vertebrates might be the major explanation for the limited ab-diversity in these species. As previously mentioned, affinity maturation is a product of somatic hypermutation followed by
33

selection of high affinity ab mutants. The germinal centres provide the environment for this selective process (reviewed in 79). The lack of these structures in low vertebrates has been invoked to explain the poor affinity maturation in Xenopus (80), where somatic hypermutations occur at a comparable rate to that of mammals (76). This situation might be the same in fishes. It has been suggested that somatic mutations introduce sequence variation in the IgV genes of adult shark (46). In summary, somatic hypermutations seem to occur in some low vertebrates, and the lack of affinity maturation can be ascribed to the lack of germinal centres. Whether or not somatic hypermutations occurs in fishes has yet to be determined.
Recently, a new putative ag-binding receptor was isolated from nurse shark (NAR, new antigen receptor) (173). This NAR molecule is encoded by V, D, J and C genes organised in clusters like the shark Ig and TcR genes (NAR shows a relationship with both Ig and TcR genes depending on the gene segments compared) and seems to undergo extensive diversification by somatic mutation. It is, however, possible that these somatic mutations have accumulated during the lifetime of the fish.
The T cell repertoire is also important for the selection of the abrepertoire, which should be considered when more detailed information onthe fish T cell is obtained.
In addition to genetic and physiological factors, the immune system is affected by the environmental factors. For example, the immune response by T cells and to lesser extent B cells is suppressed at low temperatures (171). Ig levels, lymphocyte number, specific cytotoxicity and phagocytosis is dependent on seasonal changes (172).
3:4. The IgM heavy chain constant region
In Paper II we isolated a complete rainbow trout IgM cDNA, which was identified as the secreted form of IgM, i.e. Ch 1-Ch 2-Ch 3-Ch 4 domainsfollowed by the C-terminal secretory peptide. This cDNA was used inseveral studies which will be discussed below and in the next section.
Genomic organisation o f the IgH locus in teleost fishesTwo major types of IgH gene organisations have emerged during the
vertebrate evolution: (A) the mammalian type and (B) the elasmobranch type (Figure 4). Southern blot hybridisations suggested that ray-finned bony
34

fishes also have the mammalian type, i.e. exons for V h , D and Jh gene segments grouped in separate chromosomal regions followed down stream by a single copy of the Cn (channel catfish (27); ladyfish (28); Atlantic cod (29); Atlantic salmon (30)). Our estimation of the Cn copy number in rainbow trout is consistent with these findings (Figure 3 in Paper II) Several other features are considered in favour of a mammalian type organisation of the ray-finned bony fish IgH locus: (I) As already discussed, some ray-finned bony fishes possess a large number of V h genes, but only several D and Jh
segments, i.e. similar to mammals (e.g. Paper III; Table 1). (II) Genomic mapping of the catfish IgH genes showed that the Vh genes are linked together forming a separate group in upstream region of putative D and Jh
gene segments (174). The Jh segments are grouped together close to the IgM ChI exon (175).
We conclude that the mammalian type of IgH gene organisation is found in most vertebrate phyla, most likely established in an ancestral fish before the divergence of the bony fish and tetrapod lineages. Both IgH and IgL gene organisation in cartilaginous fish are of a cluster type (23). However, in contrast to the mammalian type of IgH organisation, the IgL gene organisation in ray-finned bony fish resembles that of the elasmobranch type (59, 61). Two possible explanations for these findings are: (A) The elasmobranch type represents the primitive form of the IgH and IgL gene organisation. The mammalian type evolved in two successions, the first (IgH locus) after the divergence between cartilaginous fishes and ray-finned bony fishes and the second (IgL locus) after ray-finned bony fish-tetrapod divergence. Consequently the ray-finned bony fishes have both types of gene organisation, while the tetrapods only have the mammalian type. (B) The mammalian type represents the ancestral organisation and the elasmobranch type evolved later due to contraction and expansion events during the evolution. The finding that the coelacanth fish (a lobe-finned fish) has an intermediate type (Vh-D repeats with no adjacent Jh and Ch genes, Figure 4) suggests that the first possibility is more likely if this lobe-finned fish species diverged between cartilaginous fishes and ray-finned bony fishes.
As already discussed in section 1:4, the ray-finned bony fishes do not have functionally different isotypes other than IgM (perhaps with the possible exception of the IgD-like isotype in channel catfish, Dr Melanie Wilson, pers. communication). Even if ray-finned bony fishes have additional Ig isotypes, they are probably unable to utilise an isotype switching mechanism, as indicated by the lack of switch regions in front of the fish Cn
35

gene (141, 175). Two very similar Cn genes, named isotypes by the authors, are present in Atlantic salmon (30). We believe that these two Cn "isotypes" are either alleles or might represent duplicates from the tetraploidisation event which occurred in salmonid fishes (176).
Peculiarity of membrane IgM in bony fish.IgM can be either expressed on the surface of B-cells or secreted into
the extracellular surroundings. These two variants are generated through alternative mRNA splicing (177). The regulatory mechanism controlling the generation of slgM versus mlgM involves a splicing event mediated through a cryptic donor site in the boundary of C|-|4-C-terminal peptide (Figure 4 in Paper II, 178). This site is conserved in cartilaginous fishes and tetrapods, which all generate mlgM through splicing of the transmembrane exons (TM) to the Ch4 domain. However, several ray-finned bony fishes have a mutated, probably non-functional cryptic donor site, and mlgM is generated through splicing of the TM to the Ch3 domain, eliminating the entire Ch4 domain (channel catfish (107), Atlantic cod (29) and Atlantic salmon (30)). An early observation in rainbow trout was that the mlg protein was shorter than slg (179), and in Paper II (Figure 5) we and others (141, 163) provide additional evidence that this difference in size is due to the unusual splicing of the TM. Recently it was shown that some primitive bony fishes, longnose gar and bowfin, utilise both the cryptic Ch4 donor site and the Ch3 splice site in order to generate the mlgM (180). The difference in the mlgM splicing is summarised in Figure 9.
Gar and bowfin belong to the holostean fishes, and are regarded as more primitive than the euteleosts, represented here by ladyfish, catfish, salmon, rainbow trout and cod (Figure 6, 138). For an evolutionary explanation one might speculate that the first mutation leading to a deficient cryptic donor site in Ch 4 occurred before the divergence of holostean and euteleost fishes. One possibility is that the ancestor of these two fish lineages had undergone tetraploidisation (176), allowing the less efficient (or non-functional) variant of the Ch 4 donor-site to coexist with the functional one. In time, the splicing machinery started to use the Ch3 donor site as well as Ch4 donor site in the generation of mlgM. After their divergence, the ancestor(s) of euteleosts completely eliminated the splicing of TM to Ch4 in generating mlgM.
The mammalian mlgM associates with the lg-a/lg-ß co -recep to r required for signal transduction upon IgM-ag crosslinking and efficient
36

transport of mIgM to the B-cell surface (reviewed in 181). This co-receptor interacts with the TM region and the Ch 4 domain (182) Activation of catfish B-cells is very similar to the mammalian B-cell activation (183) suggesting that a similar co-receptor is present in the euteleosts. If so we can further speculate that the interaction between lg-a/lg-ß and IgM could have been modified during evolution, possibly using the Ch3 domain instead. The Ch 4 domain is, however, required for the tetramerisation of slgM protein and is thus also conserved in the bony fishes.
A. secreted IgM
birdamphibian euteteast hokstean cartilaginous fish
B. membrane IgMmammalbirdamphibian hokstean cartilaginous fish
C. membrane IgMeuteteasthokstean
OM S TM1 TM? CK3 CW S TM! TMZ
CHI o e CHJ Ok sI I I I ■
s lgM
CHI CH2 OB CH4 TM1, 2i i \
m lgM
CHI CH? CK3TM1.2
i i rmm lgM
Figure 9. Diagram of the mRNA splicing generating secreted IgM and membrane IgM. A. The generation of slgM in all vertebrates; B. The generation of mlgM in tetrapods, holosteans (gar and bowfin) and cartilaginous fishes (e.g. shark), where the TM exons are spliced to the Ch4 domain; C. The generation of mlgM in holosteans and euteleosts (ladyfish, catfish, salmon, rainbow trout and cod), where the TM exons are spliced to the Ch 3 domain deleting the Ch 4 domain. slgM, secreted IgM; mlgM, membrane IgM; Ch , heavy chain constant region domain; TM, transmembrane exon; S, C-terminal secretory peptide.
3:5 Speed of IgM constant region gene evolution.
The IgM protein.The constant region gene, despite its name, has been subject to many
amino acid substitutions, insertions and deletions during the vertebrate evolution, which is shown in the aa alignment of Cn regions from different species (Figure 1, Paper II). Some of these aa differences are associated with a certain phylogenetic lineage (the teleost-specific residues are bold
37

and shadowed in Figure 1, Paper II). The aa percentage identity between the rainbow trout Cn gene and those of other species is generally quite low (25.9-39.1%, Table I in Paper II), but naturally very high (95-97%) between rainbow trout and Atlantic salmon, another salmonid fish. Consistent with other studies (184, 185), we found that the Ch4 and Ch1 domains are more conserved than the Ch2 and Ch3 domains. This difference is probably due to varied functional importance among domains, e.g. (I) Ch4 is essential for the polymerisation of slgM, (II) Ch1 is important for the interaction between Ch and C l, which is demonstrated in a study where catfish ChI is shown to interact with mouse X IgL chain (186).
The amino acid substitution rate of IgM CH4 domain.Once the constancy of the amino acid substitution rate (kaa) of a
protein measured over a long evolutionary period is established, it can be applied to estimate the divergence time (T) between species. For this purpose, 11 IgM aa sequences from different vertebrate phyla were aligned for each domain. The average number of aa substitutions per site, Kaa, is given by:
Kaa=-ln(1-pd) equation 1 (136)
pd is the percent difference between the two peptides compared (136). Each species was compared to the human Cn domains from which the average Kaa was calculated for separate phyla, i.e. cartilaginous fish (shark and skate); bony fish (rainbow trout, Atlantic salmon, Atlantic cod, channel catfish, ladyfish); amphibian (Xenopus), bird (chicken) and mammal (mouse and rabbit). The Kaa values for each domain, as well as the whole IgM, were then plotted against T estimated from fossil records (138) between human and other phyla.
We obtained a highly significant linear relationship (regression coefficient=0.92) for the Ch4 domain, Figure 4 in Paper II. The Ch I and the whole IgM showed a linearity but with less precision than the Ch4 domain, while Ch2 and Ch 3 domains did not give a clear result. This is in accordancewith the notion that the Ch4 and Ch I domains are more conserved.
The average aa substitution rate per site per year (kaa) is then obtained by:
kaa=Kaa/(2xT) equation 2 (136)
38

We calculated a kaa=1 .4x10*9 substitutions per site per year for the Ch4 domain and kaa=2 .0x10 -9 for the whole IgM Qt region. These substitution rates are in the same range as the kaa estimated for the mammalian IgCX., IgCy and IgCK region genes (2 .7 x 1 0 '9 , 3 .1x10 ‘9 and 3 .7 x 1 0 ‘9 respectively) (1 8 7 ) and falls between those of cytochrome C (0 .2 2 x 1 0-9) (1 8 8 ) and fibrinopeptide (9 .0 x 10-9) (189 ), the fastest evolving protein known.
When the kaa for Ch4 is determined it can be used to estimate the unknown divergence time (T) between closely related species whose Cn sequence (therefore Kaa) is known. T is given by:
T=kaa/(2xKaa) from equation 2
Though this method is inherently susceptible to random fluctuations, it provides an additional, sometimes powerful, tool in determining the approximate divergence time between species when fossil information is insufficient. With a rapidly accumulating amount of molecular data on many genes from different species, no doubt these molecular methods will be used extensively to determine phylogenetic relationship and divergence time of species with increasing precision.
3:6 Evolution of salmonid fishes.
Using the above formula and the IgM Ch 4 molecular clock, we calculated the approximate divergence time between two salmonid fishes, rainbow trout and Atlantic salmon, to be 10 my and 16 my (Paper II).
We also used other genetic regions for the same purpose (Paper IV). It is known that introns are largely freed from natural selection, but their high substitution rates fall within a certain range (136, 137, 190). They can thus be used as an independent molecular clock when closely related species are studied. For this purpose we isolated the genomic IgM constant region genes from two salmonid species, rainbow trout and Arctic cha rr ( Salvelinus alpinus). The ChI, Ch2, Ch3 and Ch4 domains are encoded by separated exons followed by introns (intron 1, 2 and 3; Figure 1 in Paper IV) similar to the Ig genes of other vertebrate species. The rainbow trout and Arctic charr introns could be aligned with each other but not, as expected, with Cu introns of other phyla or even another fish (channel catfish) which diverged 150-200 my ago. The expected nucleotide substitution per site (d) in introns or exons is given by:
39

d=-3/4 ln(1-4pd/3 ) Jukes-Cantor equation (143)
where pd is the percent nt difference between the two sequences compared. The expected nt substitutions per site (expressed by d xlO2) in total introns (1846 bp) between rainbow trout and Arctic charr is d=5.39 (S.E.=0.55). In order to determine T between these two salmonid fishes, the d value was compared with that of introns (1362 bp) of the IgCa gene of several primate species, analysed by a method identical to ours (191). Our d value (5.39) is the same as that between orangutan and three other primates; human, chimpanzee and gorilla (5.64, 5.17 and 4.62 respectively, S.E.=0.49), considering the errors. The divergence time separating orangutan and the lineage for human-chimpanzee-gorilla is considered to be 12-16 my based on fossil records. Consequently, we may conclude that the divergence time of rainbow trout and Arctic charr is in the similar range of 12-16 my. Using the molecular clock of the Ch4 domain as described above, the estimated T between rainbow trout and Arctic charr is 18 my. This is not significantly different considering the large standard error.
The above data demonstrate that these two independent methods give rise to the same result. Both these methods may be useful for measuring the divergence time between any closely related species of vertebrates.
North American fossils suggest that three Oncorhynchus-Wke species (pink salmon, 0. gorbuscha; sockeye salmon, O. nerke\ chum salmon, O. keta) diverged before 5.5-7.6 my ago (192). The salmonid genera Oncorhynchus, Salmo, Salvelinus, Coregonus, Hucho and Thymallinus diverged much earlier(193). For example, the oldest fossil of Salvelinus is at least 10 my old(194). Salvelinus is considered to have diverged earlier from the Salmo- Oncorhynchus lineage (139, 140, 198). However, in a study using the mitochondrial cytochrome b gene, Salvelinus is grouped with Salmo rather than Oncorhynchus (195).
Our molecular estimates indicate that rainbow trout-salmon and rainbow trout-Arctic charr diverged 10-16 my ago. This suggests that the events generating the different genera Salvelinus (charr), Salmo (salmon) and Oncorhynchus (rainbow trout) occurred in short successions around mid- Miocene. This is in accordance with another study where they estimated the divergence time between the genus Oncorhynchus and Salmo using the growth hormone gene (11 my, 196). These conclusions, summarised in Figure
40

10, are tentative, and accumulation of additional sequence information may further revise and clarify the phylogenetic relations between different salmonid fishes.
6 my1
10-16 my
12-16 my
genera
chum salmon pink salmon
sockeye salmon
€ 1rainbow trout cutthroat trout
Oncorhynchus
-CAtlantic salmo7T|Sa/mobrown trout __ |
■ Arctic charr Salvelinus
Hucho5-25 mymiocene
■ whitefish Coregonus
Figure 10. Phylogenetic tree of several salmonid fishes. Fossil records show that divergence for some Oncorhynchus species occurred before 6 million years ago. Our estimates suggest that Oncorhynchus-Salmo and Oncorhynchus-Salvelinus diverged 10-16 and 12-16 million years ago respectively. (*), divergence time based on fossil records (193).
41

4. Concluding remarks
1. Comparison between rainbow trout and channel catfish Vh genes suggests that a V h gene family can last for 150 million years or longer during vertebrate evolution. This conclusion is compatible with Southern hybridisations using DNA from different fish species.
2. The phylogenetic study of V h gene families from all major vertebrate phyla show five major groups. Group A, B and C correspond to the mammalian clan I, II and III respectively. The majority of bony fish Vhs forms a group o f their own (Group D) representing a new clan, the teleost clan. Some rainbow trout Vhs branch out together with the Vhs of cartilaginous fishes in Group E (the archaic clan).
3. The presence of "species-specific" residues in several rainbow trout VHs as well as Vhs of other low vertebrates implies tha t homogenising mechanisms have been operating on the IgV multigene family over 500 million years of vertebrate evolution.
4. Rainbow trout has 100-200 Vh gene segments divided into 11 families and several D and Jh gene segments. These IgH genes are organised as the mammalian type, i.e. each exons for Vh , D and Jh gene segments are located in groups followed by a single copy of the gene.
5. Rainbow trout generates ab-diversity through random recombination ofmany V, D and J gene segments, junctional diversity and combination of IgL and IgH chains, mechanisms common to most other vertebrate species. The presence or absence of somatic hypermutations in fish Ig genes is not yet determined.
6. Rainbow trout generates mlgM by splicing of TM exons to the Ch3 domain, deleting the Ch 4 domain. This is also observed in other bony fishes, suggesting that the mutation causing this splicing pattern occurred in an early ancestor of bony fishes.
7 . The average substitution rate (kaa) for the Ch4 domain of vertebrate IgMis estimated to be 1.4x1 O'9 amino acids per site per year.
8. Two methods were used to determine the approximate divergence timebetween salmonid fishes: (1) The divergence time between Atlantic salmon
42

and rainbow trout was estimated at 10-16 million years using the constant rate of the Ch4 domain. (2) From the comparison of the nt substitutions per site in introns of two sets of species (Ch introns of rainbow trout-Arctic charr IgM and IgA-Ca introns of orangutan-human-chimpanzee-gorilla), the divergence time between rainbow trout and Arctic charr was estimated to be 12-16 million years. Both estimates are compatible with the divergence time of a few other salmonid fishes known from fossil records and molecular estimates of the growth hormone gene.
43

Acknowledgements
I wish to sincerely thank all of you that have helped me one way or another during my initiating years in the scientific world of unsolved mysteries. Especially I would like to thank:
My supervisor and friend Docent Takeshi Matsunaga, for all support and encouragement at all times.
Vuokko Törmänen and Ulf Dahl who helped me to get started in the very beginning.
All previous and present members of Group Dan Holmberg for being like a second group.
Francesco Colucci who drew the cover page in a very capturing way and Kate Huges for correcting the English into a more reader-friendly text.
Dr. Fredrik Ivars, Håkan Persson, Kristina Ruuth and a long list of others who have helped me with technical problems of all kinds.
Dr. Melanie Wilson for access to her unpublished data.
S-floor crew: Johnny Stenman (I'm going to miss our talks at lunch time), Bengt-Erik Nordlund (the computer survived my three day runs!), Karin Andersson, Mia Carlsson, Christina Thylen, Ethel Strömdahl, Karin Ekman (keep on singing), Karin Öberg and Gull-Britt Berggren; the previous members Vuokko T., Johanna Wahlquist, Ewa Lindström; and everyone else who has visit our corner.
Susanne Larsson for being the best of friends during these years in Umeå,especially when I needed it the most.
My friend and training partner Kristina Vallbo; we really have to go to London again.
Karin Andersson and Asa Edwinsson, I am so glad that you made sure that I survived my "snow-diving" experiences, despite foggy mountains.
My dear family Birgitta, Lasse and Per, for all support and love during theseyears. I love you all.
Thank you all.
This work was performed at the Department of Applied Cell and Molecular Biology, Umeå University and supported by NFR, SJFR, Kempe Foundation, Stiftelsen Lars Hiertas Minne and the Department of Applied Cell and Molecular Biology.
44

References
1. Steiner, H., Hultmark, D., Engström, A., Bennich, H. and Boman, H. G. (1981) Sequence and specificity of two antibacterial proteins involved in insect immunity, Nature 292:246-2482. Sun, S.-C., Lindström, I., Boman, H. G., Faye, I. and Schmidt, 0. (1990) Hemolin: An insect-immune protein belonging to the immunoglobulin superfamily. Science 250:1729-17323. Beck, G., Cooper, E. L., Habicht, G. S. and Marchalonis, J. J. (1 994) Primordial Immunity. Foundations for the Vertebrate Immune System. Annals o f the New York Academy o f Sciences Academy of Sciences vol. 7124. Vanhamme, L. and Pays, E. (1995) Control of gene expression in Trypanosomes. Microbiol. Rev. 59:223-2405. Du Pasquier, L. (1982) Antibody diversity in lower vertebrates-why is it so restricted? Nature 296:311-3136. Matsunaga, T. (1985) Evolution of antibody repertoire - Somatic mutation as a late comer. Dev. Comp. Immunol. 9:585-5967. Smith, G. R. and Stearly, R. F. (1989) The classification andscientific names of rainbow and cutthroat trouts. Fisheries 12:4-108. Davies, D. R. and Padlan, E. A. (1975) Three-dimensional structure of immunoglobulins. Annul. Rev. Biochem. 44:639-6679. Hasemann, C. A. and Capra, J. D. Immunoglobulins: Structure and Function. In W. E. Paul (ed.) Fundamental Immunology (2nd ed.), pp. 209-233, Raven Press Ltd, New York, 198910. Harding, F. A., Amemiya, C. T., Litman, R. T., Cohen, N. andLitman, G. W. (1990) Two distinct immunoglobulin heavy chain isotypes in a primitive, cartilaginous fish, Raja erinacea. Nucl. Acids Res. 18:6369- 637611. Williams, A. F. and Barclay, A. N. (1988) The immunoglobulinsuperfamily-domains for cell surface recognition. Ann. Rev. Immunol. 6:381- 40512. Edmundson, A. B., Ely, K. R., Abola, E. E., Schiffer, M. andPanagiotopoulos, N. (1975) Rotational allomerism and divergent evolution of domains in immunoglobulin light chains. Biochem. 14:3953-396113. Wu, T. T. and Kabat, E. A. (1970) An analysis of the sequences of the variable regions of Bence Jones proteins and myeloma light chains and their implications for antibody complementarity. J. Exp. Med. 132:211-25014. Tonegawa, S. (1983) Somatic generation of antibody diversity. Nature 302:575-58115. Marchalonis, J. J. and Edelman, G. M. (1968) Phylogenetic origins o f antibody structure. III. Antibodies in the primary immune response of the sea lamprey, Petromyzon marinus. J. Exp. Immunol. 127:891-89616. Raison, R. L., Hull, C. J. and Hildemann, W. H. (1978 ) Characterization of immunoglobulin from the Pacific hagfish, a primitive vertebrate. Proc. Natl. Acad. Sci. USA 75:5679-568217. Hanley, P. J., Seppelt, I. M., Gooley, A. A., Hook, J. W. and Raison, R. L. (1990) Distinct lg H chains in a prim itive vertebrate, Eptatretus stouti. J. Immunol. 145:3823-3828
45

18. Hanley, P. J., Hook, J. W., Raftos, D. A., Gooley, A. A., Trent, R. and Raison, R. L. (1992) Hagfish humoral defence protein exhibits structural and functional homology with mammalian complement components. Proc. Natl. Acad. Sci. USA 89:7910-791419. Ishiguro, H., Kobayashi, K., Suzuki, M., Titani, K., Tomonaga, S. and Kurosawa, Y. (1992) Isolation of a hagfish gene that encodes a complement component. EMBO J. 11:829-83720. Kokubu, F., Litman, R., Shamblott, M. J., Hinds, K. and Litman, G. W. (1988) Diverse organization of immunoglobulin Vh gene loci in a primitive vertebrate. EMBO J. 7:3413-342221. Schwager, J., Grossberger, D. and Du Pasquier, L. (1988) Organization and rearrangement of immunoglobulin M genes in the amphibian Xenopus. EMBO J. 7:2409-241522. Wilson, M., Miller, N., Clem, C. W. and Warr, G. W. Germline Vh genes of the catfish, Ictalurus punctatus. (1988) 4th ISDCI Meeting Abstract 1,3.23. Shamblott, M. J. and Litman, G. W. (1989) Genomic organization and sequences of immunoglobulin light chain genes in a primitive vertebrate suggest coevolution of immunoglobulin gene organization. EMBO J. 12:3733- 378924. Matsunaga, T., Chen, T. and Törmänen, V. (1990) Characterization of a complete immunoglobulin heavy chain variable region germ line of rainbow trout. Proc. Natl. Acad. Sci. USA 87:7767-777125. Litman, G. W., Rast, J. P., Shamblott, M. J., Haire, R. N., Hulst, M., Roess, W., Litman, R. T., Hinds-Frey, K. R., Zilch, A. and Amemiya, C. T. (1993) Phylogenetic diversification of immunoglobulin genes and the antibody repertoire. Mol. Biol. Evol. 10:60-7226. Du Pasquier, L. Evolution of the Immune System. In W. E. Paul (ed.) Fundamental Immunology (2nd ed.), pp. 139-165, Raven Press Ltd, New York, 198927. Ghaffari, S. H. and Lobb, C. J. (1989) Nucleotide sequence of channel catfish heavy chain cDNA and genomic blot analysis. Implications for the phylogeny of Ig heavy chains. J. Immunol. 143:2730-273928. Amemiya, C. T. and Litman, G. W. (1990) Complete nucleotide sequence of an immunoglobulin heavy-chain gene and analysis of immunoglobulin gene organization in a primitive teleost species. Proc. Natl. Acad. Sci. USA 87:811-81529. Bengtén, E., Leanderson, T. and P ilström , L. (1 9 9 1 ) Immunoglobulin heavy chain cDNA from the teleost Atlantic cod (Gadus morhua L.): nucleotide sequences of secretory and membrane form show an unusual splicing pattern. Eur. J. Immunol. 21:3027-303330. Hordvik, I., Voie, A. M., Glette, J., Male, R. and Endersson, C.(1992) Cloning and sequence analysis of two isotypic IgM heavy chain genes from Atlantic salmon, Salmo salar L. Eur. J. Immunol. 22:2957-296231. Wilson, M. R., van Ravenstein, E., Miller, N. W., Clem, L. W., Middleton, D. L. and Warr, G. W. (1995) cDNA sequences and organization of IgM heavy chain genes in two holostean fish. Dev. Comp. Immunol. 19:153- 164
46

32. Livant, D., Blatt, C. and Hood, L. (1986)One heavy chain region gene segment subfamily in the BALB/c mouse contains 500-1000 or more members. Cell 47:461-47033. Brodeur, P. H. Genes encoding the immunoglobulin variable regions InF. Calabi and M. S. Neuburger (eds.) Molecular Genetics o f Immunoglobulin, pp. 81-109, Elsevier, New York, 198734. Brodeur, P. H. and Riblet, R. (1984) The immunoglobulin heavy chain variable region (Igh-V) locus in the mouse. I. One hundred Igh-V genes comprise seven families of homologous genes. Eur. J. Immunol. 14:922-94035. Kroemer, G., Faessler, R., Strohal, R., Wick, G. and Kofler, R. (1987) Evidence for a new murine immunoglobulin heavy chain variable region gene family. Immunol. Invest. 16:107-11636. Kofler, R. (1988) A new murine Ig Vh gene family J. Immunol. 140:4031-403437. Reininger, L., Kaushik, A., Izui, S. and Jaton, J.-C. (1988) A member of a new Vh gene family encodes antibromelinized mouse red blood cell autoantibodies. Eur. J. Immunol. 18:1521-152638. Tutter, A., Brodeur, P., Shlomchik, M. and Riblet, R. (1991) Structure, map position, and evolution of two newly diverged mouse Ig Vh gene families. J. Immunol. 147:3215 - 322339. Cook, G. P. and Tomlinson, I. M. (1995) The human immunoglobulin Vh repertoire. Immunol. Today 16:237-24240. Haire, R. N., Amemiya, C. T., Suzuki, D. and Litman, G. W. (1990) Eleven distinct Vh gene families and additional patterns of sequence variation suggest a high degree of immunoglobulin gene complexity in a lower vertebrate, Xenopus laevis. J. Exp. Med. 171:1721-173741. Warr, G. W., Middleton, D. L., Miller, N. W., Clem, L. W. andWilson, M. R. (1991) An additional family of Vh sequences in the channel catfish. Eur. J. Immunogenetics 18:393-39742. Ghaffari, S. H. and Lobb, C. J. (1991) Heavy chain variable region gene families evolved early in phylogeny. Ig complexity in fish. J. Immunol. 146:1037-104643. Reynaud, C.-A., Dahan, A., Anquez, V. and Weill, J.-C. (1989)Somatic hyperconversion diversifies the single Vh gene of the chicken with a high incidence in the D region. Cell 59:171-18344. Hinds, K. R. and Litman, G. W. (1986) Major rearrangement ofimmunoglobulin Vh segmental elements during vertebrate evolution. Nature 320:546-54845. Harding, F. A., Cohen, N. and Litman, G. W. (1990) Immunoglobulin heavy chain gene organization and complexity in the skate, Raja erinacea. Nucl. Acid Res. 18:1015-102046. Hinds-Frey, K. R., Nishikata, H., Litman, R. T. and Litman, G. W.(1993) Somatic variation precedes extensive diversification of germline sequences and combinatorial joining in the evolution of immunoglobulin heavy chain diversity. J. Exp. Med. 178:815-82447. Anderson, M., Amemiya, C., Luer, C., Litman, R., Rast, J., Niimura, Y. and Litman, G. (1994) Complete genomic sequence and
47

patterns of transcription of a member of an unusual family of closely related, chromosomally dispersed Ig gene clusters in Raja. Int. Immunol. 6:1661-167048. Anderson, M. K., Shamblott, M. J., Litman, R. T. and Litman, G. W. (1995) Generation of immunoglobulin light chain gene diversity in Raja erinacea is not associated with somatic rearrangement, en exception to a central paradigm of B cell immunity. J. Exp. Med. 182:109-11949. Amemiya, C. T., Ohta, Y., Litman, R. T., Rast, J. P., Haire, R. N. and Litman, G. W. (1993) Vh gene organization in a relict species, the coelacanth Latimeria chalumnae: Evolutionary implications. Proc. Natl. Acad. Sci. USA 90:6661-666550. Max, E. E. Immunoglobulins: Molecular Genetics. In W. E. Paul (ed.) Fundamental Immunology (2nd ed.), pp. 235-290, Raven Press Ltd, New York, 198951. Magor, K. E., Higgins, D. A., Middleton, D. L. and Warr, G. W.(1994) cDNA sequence and organization of the immunoglobulin light chain gene of the duck, Anas platyrhynchos. Dev. Comp. Immunol. 18:523-53152. Reynaud, C.-A., Anquez, V., Dahan, A. and Weill, J.-C. (1985) A single rearrangement event generates most of the chicken immunoglobulin light chain diversity. Cell 40:283-29153. Schwager, J., Bürckert, N., Schwager, M. and Wilson, M. (1991) Evolution of immunoglobulin light chain genes: analysis of Xenopus IgL isotypes and their contribution to antibody diversity. EMBO J. 10:505-51154. Zezza, D. J., Stewart, S. E. and Steiner, L. A. (1992) Genes encoding Xenopus laevis lg L chains: Implications for evolution of k and X chains. J. Immunol. 149:3968-397755. Hohman, V. S., Schlüter, S. F. and Marchalonis, J. J. (1992) Complete sequence of a cDNA clone specifying sandbar shark immunoglobulin light chain: Gene organization and implications for the evolution of light chains. Proc. Natl. Acad. Sci. USA 89:276-28056. Greenberg, A. S., Steiner, L., Kasahara, M. and Flajnik, M. F.(1993) Isolation of a shark immunoglobulin light chain cDNA clone encoding a protein resembling mammalian k light chains: implications for th e evolution of light chains. Proc. Natl. Acad. Sci. USA 90:10603-1060757. Rast, J. P., Anderson, M. K., Ota, T., Litman, R. T., Margittai, M., Shamblott, M. J. and Litman, G. W. (1994) Immunoglobulin light chain class multiplicity and alternative organizational forms in early vertebrate phylogeny. Immunogenetics 40:83-9958. Blomberg, B., Traunecker, A., Eisen, H. and Tonegawa, S. (1981) Organization of four mouse X light chain immunoglobulin genes. Proc. Natl. Acad. Sci. USA 78:3765-376959. Daggfeldt, A., Bengtén, E. and Pilström, L. (1993) A cluster type organization of the loci of the immunoglobulin light chain in Atlantic cod (Gadus morhua L.) and rainbow trout (Oncorhynchus mykiss Walbaum) indicated by nucleotide sequences of cDNAs and hybridization analysis. Immunogenetics 38:199-209
48

60. Ghaffari, S. H. and Lobb, C. J. (1993) Structure and genomic organization of immunoglobulin light chain in the channel catfish. J. Immunol. 151:6900-691261. Bengtén, E. The Immunoglobulin Genes in Atlantic Cod (Gadus morhua) and Rainbow Trout ( Oncorhynchus mykiss). Uppsala University, Sweden, 199462. Schatz, D. G., Oettinger, M. A. and Baltimore, D. (1989) The V(D)J recombination activating gene, RAG-1. Cell 59:1035-104863. Oettinger, M. A., Schatz, D. G., Gorka, C. and Baltimore, D. (1990) RAG-1 and RAG-2, adjacent genes that synergistically activate V(D)J recombination. Science 248:1517-152364. Greenhalgh, P. and Steiner, L. A. (1995) Recombination activating gene 1 (RAG1) in zebrafish and shark. Immunogenetics 41:54-5565. Hansen, J. D. and Kaattari, S. L. (1995) The recombination activating gene 1 (RAG1) of rainbow trout (Oncorhynchus mykiss): cloning, expression and phylogenetic analysis. Immunogenetics 42:188-19566. Becker, R. S. and Knight, K. L. (1990)Somatic diversification ofimmunoglobulin heavy chain VDJ genes: Evidence for somatic geneconversion in rabbits. Cell 63:987-99767. Lewis, S., Gifford, A. and Baltimore, D. (1985)DNA elements are asymmetrically joined during the site-specific recombination of kappa immunoglobulin genes. Science 228:675-68568. Alt, F. W. and Baltimore, D. (1982)Joining of immunoglobulin heavy chain gene segments: Implications from a chromosome with evidence of three D-Jh fusions. Proc. Natl. Acad. Sci. USA 79:4118-412269. Lafaille, J. J., DeCloux, A., Bonneville, M., Takagaki, Y. andTonegawa, S. (1989) Junctional sequences of T cell receptor yò genes: Implications for yô T cell lineages and for a novel intermediate of V-(D)-J joining. Cell 59:859-87070. McKean, D., Huppi, K., Bell, M., Staudt, L., Gerhard, W. andWeigert, M. (1984) Generation of antibody diversity in the immuneresponse of BALB/c mice to influenza virus hemagglutinin. Proc. Natl. Acad. Sci. USA 81:3180-318471. Wable, M., Burrows, P. D., von Gabin, A. and Steinberg, C. (1985) Hypermutation at the immunoglobulin heavy chain locus in a pre-B-cell line. Proc. Natl. Acad. Sci. USA 82:479-48272. Griffiths, G. M., Berek, C., Kaartinen, M. and Milstein, C. (1984)Somatic mutation and the maturation of immune response to 2-phenyl oxazolane. Nature 312:271-27573. Berek, C., Griffiths, G. M. and Milstein, C. (1985) Molecular events during maturation of the immune response to oxazolane. Nature 316:412-41874. Weiss, U. and Rajewski, K. (1990) The repertoire of somaticantibody mutants accumulating in the memory compartment after primary immunization is restricted through affinity maturation and mirrors that expressed in the secondary response. J. Exp. Med. 172:1681-168975. Reynaud, C.-A., Garcia, C., Hein, W. R. and Weil, J.-C. (1995)Hypermutation generating the sheep immunoglobulin repertoire is an antigen-independent process. Cell 80:115-125
49

76. Wilson, M., Hsu, E., Marcuz, A., Courtet, M., Du Pasquier, L and Steinberg, C. (1992) What limits affinity maturation of antibodies in Xenopus - the rate of somatic mutation or the ability to select mutants? EMBO J. 11:4337-434777. Brandt, D. C., Griessen, M., Du Pasquier, L. and Jaton, J.-C. (1980) Antibody diversity in amphibians: evidence for the inheritance of idiotypic specificities in isogenic Xenopus. Eur. J. Immunol. 10:731-73678. Du Pasquier, L. and Wable, M. R. (1978) Antibody diversity in amphibians: inheritance of isoelectric focusing antibody patterns in isogenic frogs. Eur. J. Immunol. 8:428-43379. Berek, C. and Ziegner, M. (1993) The maturation of the immune response. Immunol. Today 14:400-40480. Nahm, M. H., Kroese, F. G. M. and Hoffmann, J. W. (1992) The evolution of immune memory and germinal centres. Immunol. Today 13:438- 44181. Ingram, G. A. (1980) Substances involved in the natural resistance of fish to infection - A review. J. Fish Biol. 16:23-6082. Moody, C. E., Serreze, D. V. and Reno, P. W. (1985) Non-specificcytotoxic activity of teleost leukocytes. Dev. Comp. Immunol. 9:51-6483. Caspi, R. R. and Avtalion, R. R. (1984) The mixed leukocyte reaction (MLR) in carp: bidirectional and unidirectional MRL responses. Dev. Comp. Immunol. 8:631-63784. Miller, N. W., Deuter, A. and Clem, L. W. (1986) Phylogeny of lymphocyte heterogeneity: The cellular requirements for the mixed leukocyte reaction with channel catfish. Immunol. 59:123-12885. DeLuca, D., Wilson, M. and Warr, G. W. (1983) Lymphocyte heterogeneity in the trout, Salmo gairdneri, defined with monoclonal antibodies to IgM. Eur. J. Immunol. 13:546-55186. Miller, N. W., Sizemore, R. C. and Clem, L. W. (1985) Phylogeny oflymphocyte heterogeneity: the cellular requirements for in vitro antibodyresponses of channel catfish leukocytes. J. Immunol. 134:2884-288887. Sizemore, R. C., Miller, N. W., Cuchens, M. A., Lobb, C. J. and Clem, L. W. (1984) Phylogeny of lymphocyte heterogeneity: the cellular requirements for in vitro mitogenic responses of channel catfish leukocytes. J. Immunol. 133:2920-292488. Klein, J. Natural History o f the Major Histocompatibility Complex. John Wiley & Sons, Inc. 198689. Stet, R. J. M. and Egberts, E. (1991) The histocompatibility system in teleostean fishes: from multiple histocompatibility loci to a major histocompatibility complex. Fish and Shellfish Immunol. 1:1-1690. Dixon, B., van Erp, S. H. M., Rodrigues, P. N. S., Egberts, E. and Stet, R. J. M. (1995) Fish major histocompatibility complex genes: an expansion. Dev. Comp. Immunol. 19:109-13391. Vallejo, A. N., Miller, N. W. and Clem, L. W. (1992) Antigen processing and presentation in teleost immune responses. Annu. Rev. Fish Diseases 73-8992. Vallejo, A. N., Miller, N. W., Harvey, N. E., Cuchens, M. A., Warr,G. W. and Clem, L. W. (1992) Cellular pathway(s) of antigen processing
50

and presentation in fish APC: endosomal involvement and cell-free antigen presentation. Dev. Immunol. 3:51-6593. Rast, J. P. and Litman, G. W. (1994) T-cell receptor gene homologs are present in the most primitive jawed vertebrates. Proc. Natl. Acad. Sci. USA 91:9248-925294. Partula, S., de Guerra, A., Fellah, J. S. and Charlemagne, J . T h e molecular cloning o f fish T-cell receptor genes: strategies and findings. The Nordic Symposium on Fish Immunology. 199595. Partula, S., de Guerra, A., Fellah, J. S. and Charlemagne, J.(1995) Structure and diversity of the T cell antigen receptor ß-chain in a teleost fish. J. Immunol. 699-70696. Litman, G. W. Physical properties of immunoglobulins of lower species: a comparison with immunoglobulins of mammals. In Marchalonis, J. J. (ed.) Comparative Immunology 239-275; 197697. Wilson, M. R. and Warr, G. W. (1992) Fish immunoglobulins and the genes that encode them. Annu. Rev. Fish Diseases 201-22198. Hädge, D. and Ambrosius, H. (1984) Evolution of low molecular weight immunoglobulins-IV. IgY-like immunoglobulins of birds, reptiles and amphibians, precursors of mammalian IgA. Mol. Immunol. 21:699-70799. Green, C. and Steiner, L. A. (1976) Isolation and preliminary characterization of two varieties of low molecular weight immunoglobulin in the bullfrog, Rana catesbeiana. J. Immunol. 117:364-373100. Warr, G. W., Ruben, L. N. and Edwards, B. F. (1982) Evidence for low-molecular weight antibodies in the serum of a urodele amphibian, Ambystoma mexicanum. Immunol, letters 4:99-102101. Hsu, E. and Du Pasquier, L. (1984) Studies on X e no pu s immunoglobulins using monoclonal antibodies. Mol. Immunol. 21:257-270102. Fellah, J. S. and Charlemagne, J. (1988) Characterization of an IgY-like low molecular weight immunoglobulin class in the Mexican axolotl. Mol. Immunol. 25:1377-1386103. Hsu, E., Flajnik, M. F. and Du Pasquier, L. (1985) A third immunoglobulin class in amphibians. J. Immunol. 135:1998-2004104. Kobayashi, K., Tomonaga, S. and Tanaka, S. (1992) Identification of a second immunoglobulin in the most primitive shark, the frill shark, Chlamydoselachus anguineus. Dev. Comp. Immunol. 16:295-299105. Clem, L. W. and Small, P. A. (1971) Phylogeny of immunoglobulin structure and function. IV. Immunoglobulins of the giant grouper, Epinephelus itaira. J. Biol. Chem. 246:9-15106. Lobb, C. J. and Olson, M. O. J. (1988) Immunoglobulin heavy H chain isotypes in a teleost fish. J. Immunol. 141:1236-1245107. Wilson, M. R., Marcuz, A., van Ginkel, F., Miller, N. W., Clem, L. W., Middleton, D. and Warr, G. W. (1990) The immunoglobulin M heavy chain constant region gene of the channel catfish, Ictalurus punctatus: an unusual mRNA splicing pattern produces the membrane form of the molecule. Nucleic Acid Res. 18:5227-5233108. Russel, W. J., Voss Jr, E. W. and Sigel, M. M. (1970) Some characteristics of anti-dinitrophenyl antibody of the grey snapper. J. Immunol. 105:262-264
51

109. Shankey, T. V. and Clem, L. W. (1980) Phylogeny of immunoglobulin structure and function. IX. Intramolecular heterogeneity of shark 19S IgM antibodies to the dinitrophenyl hapten. J. Immumol. 125:2690-2698110. Vilain, C., Wetzel, M.-C., Du Pasquier, L. and Charlemagne, J. (1984) Structural and functional analysis of spontaneous anti-nitrophenyl antibodies in three cyprinid fish species: carp (Cyrinus carpio), goldfish (Carassius auratus) and tench ( Tinca tinea). Dev. Comp. Immunol. 8:611-622111. Killie, J.-E., Espelid, S. and Jergensen, T. 0 . (1991) The humoral immune response in Atlantic salmon (Salmo salar L.) against the hapten carrier antigen NIP-LPH; the effect of determinant (NIP) density and the isotype profile of anti-NIP antibodies. Fish & Shellfish Immunol. 1:33-46112. Kishimoto, T. and Hirano, T. B lymphocyte activation, proliferation, and immunoglobulin secretion. In W. E. Paul (ed.) Fundamental Immunology (2nd ed.), pp. 385-411, Raven Press Ltd, New York, 1989113. Lobb, C. J. (1985) Covalent structure and affinity of channel catfish anti-dinitrophenyl antibodies. Mol. Immunol. 22:993-999114. Arkoosh, M. R.and Kaattari, S. L. (1991) Development of immunological memory in rainbow trout (Oncorhynchus mykiss). I. An immunochemical and cellular analysis of the B cell response. Dev. Comp. Immunol. 15:279-293115. O'Neill, J. G. (1979) The immune response of the brown trout, Salmo trutta L., to MS2 bacteriophage: immunogen concentration and adjuvants. J. Fish Biol. 15:237-248116. Cossarini-Dunier, M. (1986) Secondary response of rainbow trout (Salmo gairdneri Richardson) to DNP-haemocyanin and Yersinia ruckeri. Aquaculture 52:81-86117. Newman, S. G. (1993) Bacterial vaccines for fish. Annual Rev. o f Fish Diseases 145-185118. Thuvander, A., Hongslo, T., Jansson, E. and Sundquist, B . (1987) Duration of protective immunity and antibody titres measured by ELISA after vaccination of rainbow trout, Salmo gairdneri Richardson, against vibriosis. J. Fish Diseases 10:479-486119. Wetzel, M.-C. and Charlemagne, J. (1985) Antibody diversity in fish. Isoelectrofocalization study of individually-purified specific antibodies in three teleost fish species: tench, carp and goldfish. Dev. Comp. Immunol. 9:261-270120. Cossarini-Dunier, M., Desvaux, F.-X. and Dorson, M. (1986) Variability in humoral responses to DNP-KLH of rainbow trout ( Salmo gairdneri ) . Comparison of antibody kinetics and immunoglobulins spectrotypes between normal trouts and trouts obtained by gynogenesis of self-fertilization. Dev. Comp. Immunol. 10:207-217121. Jelinek, W. R., Toomey, T. P., Leinwand, L., Duncan, C. H., Biro, P. A., Choudary, P. V., Weissman, S. M., Rubin, C. M., Houck, C. M., Deininger, P. L. and Schmid, C. W. (1980) Ubiquitous, interspersed repeated sequences in mammalian genomes. Proc. Natl. Acad. Sci. USA 77:1398-1402
52

122. Brown, D. D. and Weber, C. S. (1968) Gene linkage by RNA-DNA hybridization. I. Unique DNA sequences homologous to 4 s RNA, 5 s RNA and ribosomal RNA. J. Mol. Biol. 34:661-680123. Birnstiel, M. L., Chipchase, M. and Speirs, J. (1971) The ribosomal RNA cistrons. Prog. Nucl. Acid Res. Mol. Biol. 11:351-389124. Hood, L., Campbell, J. H. and Elgin, S. C. R. (1 975 ) The organization, expression, and evolution of antibody genes and other multigene families. Annu. Rev. Genet. 9:305-353125. Smith, G. P. (1974) Unequal crossover and the evolution of multigene families. Cold Spring Harb. Symp. Quant. Bib. 38:507-513126. Dover, G. (1982) Molecular drive: a cohesive mode of species evolution. Nature 299:111-117127. Arnheim, N. Concerted evolution of multigene families. In M. Nei and R. K. Koehn (eds.) Evolution o f Genes and Proteins pp. 38-61, Sinauer Sunderland, Massachusetts, 1983128. Arnheim, N., Krystal, M., Schmickel, R., Wilson, G., Ryder, 0. and Zimmer, E. (1980) Molecular evidence for genetic exchanges among ribosomal genes on nonhomologous chromosomes in man and apes. Proc. Natl. Acad. Sci. USA 77:7323-7327129. Kabat, E. A. (1967) The paucity of species-specific amino acid residues in the variable regions of human and mouse Bence-Jones proteins and its evolutionary and genetic implications. Proc. Natl. Acad. Sci. USA 57:1345-1349130. Hood, L., Eichmann, K., Lackland, H., Krause, R. M. and Ohms, J. J. (1970) Rabbit antibody light chains and gene evolution. Nature 228:1040- 1044131. Kehoe, J. M. and Capra, J. D. (1972) Sequence relationships among the variable regions of immunoglobulin heavy chains from various mammalian species. Proc. Natl. Acad. Sci. USA 69:2052-2055132. Gojobori, T. and Nei, M. (1984) Concerted evolution of the immunoglobulin Vh gene family. Mol. Biol. Evol. 1:195-212133. Tanaka, T. and Nei, M. (1989) Positive Darwinian selection observed at the variable-regions genes of immunoglobulins. Mol. Biol. Evol. 6:447-459134. Zuckerkandl, E. and Pauling, L. Evolutionary divergence and convergence in proteins. In V. Bryson and H. J. Voger (eds.) Evolving Genes and Proteins, pp. 97-166, Academic Press, New York, 1965135. Wilson, A. C., Carlson, S. S. and White, T. J. (1977) Biochemical evolution. Ann. Rev. Biochem. 46:573-639136. Kimura, M. The Neutral Theory o f Molecular Evolution, Cambridge University Press, Cambridge, 1983137. Nei, M. Molecular evolutionary Genetics, Colombia university Press, New York, 1987138. Carroll, R. L. Vertebrate Paleontology and Zoology, W . H. Freeman, New York, 1988139. Kido, Y., Aono, M., Yamaki, T., Matsu moto, K., Murata, S., Saneyoshi, M. and Okada, N. (1991) Shaping and reshaping of salmonid genomes by amplification of tRNA-derived retroposons during evolution. Proc. Natl. Acad. Sci. USA 88:2326-2330
53

140. Phillips, R. B. and Pleyte, K. A. (1991) Nuclear DNA and salmonid phylogenetics. J. Fish Biol. 39:259-275141. Lee, M. A., Bengtén, E., Daggfeldt, A., Rytting, A. a n d Pilström, L. (1993) Characterisation of rainbow trout cDNAs encoding a secreted and membrane-bound Ig heavy chain and the genomic intron upstream of the first constant exon. Mol. Immunol. 30:641-648142. Roman, T. and Charlemagne, J. (1994) The im m unog lobu linrepertoire of the rainbow trout (Oncorhynchus mykiss): definition of nine Igh-V families. Immunogenetics 40:210-216143. Jukes, T. H. and Cantor, C. R. Evolution of protein molecules. In H. N. Munro (ed.) Mammalian Protein Metabolism, pp. 21-132, Academic Press, New York, 1969144. Felsenstein, J. (1989) PHYLIP-Phylogeny Inference Package ( version 3.2 ). Cladistics 5:164-166145. Rechavi, G., Bienz, B., Ram, D., Ben-Neriah, Y., Cohen, J. B.,Zakut, R. and Givol, D. (1982) Organization and evolution o f immunoglobulin Vh gene subgroups. Proc. Natl. Acad. Sci. USA 79:4405-4409146. Schroeder, H. W., Hilson, J. L. and Perlmutter, R. M. (1990)Structure and evolution of mammalian Vh families. Int. Immunol. 2:41-50147. Litman, G. W., Berger, L., Murphy, K., Litman, R., Hinds, K. andErickson, B. W. (1985) Immunoglobulin Vh gene structure and diversity in Heterodontus, a phylogenetically primitive shark. Proc. Natl. Acad. Sci. USA 82:2082-2086148. Tutter, A. and Riblet, R. (1989) Conservation of an immunoglobulin variable-region gene family indicate a specific, noncoding function. Proc. Natl. Acad. Sci. USA 86:7460-7464149. Yancopoulos, G. D., Desiderio, S. V., Paskind, M., Kearney, J.F., Baltimore, D. and Alt, F. W. (1984) Preferential utilization of the most JH-proximal Vh gene segments in pre-B cell lines. Nature 311:727-733150. Varela, F. J. and Coutinho, A. (1991) Second generation immunenetworks. Immunol. Today 12:159-166151. Ota, T. and Nei, M. (1994) Divergent evolution be the birth-and- death process in the immunoglobulin Vh gene family. Mol. Biol. Evol. 11:469- 482152. Ohno, S., Mori, N. and Matsunaga, T. (1985) Antigen-bindingspecificities of antibodies are primarily determined by seven residues of Vh- Proc. Natl. Acad. Sci. USA 82:2945-2949153. Matsunaga, T. and Ohno, S. The evolution of antibody and antibody repertoire. In G. Kelsoe and D. H. Schulze (eds.) Evolution and Vertebrate Immunity: the Antigen Receptor and MHC Gene Families, pp. 429-451, The University of Texas Press, Austin, 1987154. Chothia, C., Lesk, A. M., Tramontano, A., Levitt, M., Smith-Gill, S. J., Air, G., Sheriff, S., Padlan, E. A., Davies, D., Tulip, W. R., Colman, P. M., Spinelli, S., Alzari, P. M. and Poljak, R. J. (1989)Conformations of immunoglobulin hypervariable regions. Nature 342:877- 883
54

155. Chothia, C., Lesk, A. M., Gherardi, E., Tomlinson, I. M., Walter,G., Marks, J. D., Llewelyn, M. B. and Winter, G. (1992) Structural repertoire of the human Vh segments. J. Mol. Biol. 227:799-817156. Barré, S., Greenberg, A. S., Flajnik, M. F. and Chothia, C. (1 9 9 4 ) Structural conservation of hypervariable regions in immunoglobulins evolution. Nature Structural S/o/. 1:915-920; 1994157. Matsunaga, T., Törmänen, V., Karlsson, K. and Andersson, E. Evolution of teleost antibody genes. In G. Beck, E. L. Cooper, G. S. Habicht and J. J. Marchalonis (eds.) Primordial Immunity; Foundation for the Vertebrate immune System, pp. 42-56, Academy of Sciences, New York, 1994158. Langman, R. E. and Cohn, M. (1987) The E-T (elephant-tadpole) paradox necessitates the concept of a unit of B-cell function: the protecton. Mol. Immunol. 24:675-697159. Ferguson, S. E., Cancro, M. P. and Osborne B. A. (1989) Analysis of a novel VhS107 haplotype in CLA-2 and WSA mice. Evidence for gene conversion among IgVH genes in outbred populations. J. Exp. Med. 170:1811- 1823160. Schwager, J., BOrckert, N., Courtet, M. and Du Pasquier, L.(1989) Genetic basis of the antibody repertoire in Xenopus: analysis of the Vh diversity. EMBO J. 8:2989-3001161. Kabat, E. A., Wu, T. T., Perry, H. M., Gottesman, K. S. and Foeller, C. Sequences o f Immunological Interest (5th ed.), U. S. Department of Health and Human Servicies, Public Health Service National Institutes of Healths, Bethesda, 1991162. Roman, T., De Guerra, A. and Charlemagne, J. (1995) Evolution of specific antigen recognition: size reduction and restricted length distribution of the CDRH 3 regions in the rainbow trout. Eur. J. Immunol. 25:269-273163. Hansen, J., Leong, J.-A. and Kaattari, S. (1994) C o m p le te nucleotide sequence of rainbow trout cDNA encoding a membrane-bound form of immunoglobulin heavy chain. Mol. Immunol. 31:499-501164. Schwager, J., Bürckert, N., Courtet, M. and Du Pasquier, L. (1991) The ontogeny of diversity at the immunoglobulin heavy chain locus in Xenopus. EMBOJ. 10:2461-2470; 1991165. Yamada, M., Wasserman, R., Reichard, B. A., Shane, S., Caton, A. J. and Rovera, G. (1991) Preferential u tilization of specific immunoglobulin heavy chain diversity and joining segments in adult human peripheral blood B lymphocytes. J. Exp. Med. 173:395-407166. Pink, J. R. L. and Askonas, B. A. (1974) Diversity of antibodies to cross-reacting nitrophenyl haptens in inbred mice. Eur. J. Immunol. 4:426- 430167. Kreth, H. W. and Williamson, A. R. (1973) The extent of diversity of anti-hapten antibodies in inbred mice: anti-NIP (4-hydroxy-5-iodo-3- nitro-phenacetyl) antibodies in CBA/H mice. Eur. J. Immunol. 3:141-147168. Hohman, V. S., Schuchman, D. B.;, Schlüter, S. F. and Marchalonis, J. J. (1993) Genomic clone for sandbar shark X light chain: Generation of diversity in the absence of gene rearrangement. Proc. Natl. Acad. Sci. USA 90:9882-9886
55

169. Padlan, E. A. (1994) Anatomy of the antibody molecule. Mol. Immunol. 31:169-217170. Hsu, E. and Steiner, L. A. (1992) Primary structure of immunoglobulins through evolution. Current Opinion in Struct. Biol. 2:422- 431171. Bly, J. E. and Clem, L. W. (1991) Temperature-mediated processes in teleost immunity: in vivo low temperature immunisation does not induce tolerance in channel catfish. Fish & Shellfish Immunol. 1:229-231172. Zapata, A. G., Varas, A. and Torroba, M. (1992) Seasonal variations in the immune system of lower vertebrates. Immunol. Today 13:142-14 7173. Greenberg, A. S., Avila, D., Hughes, M., Hughes, A., McKinney, E. S. and Flajnik, M. F. (1995) A new receptor gene family that undergoes rearrangement an extensive somatic diversification in sharks. Nature 374:168-173174. Ventura-Holman, T., Jones, J. C., Ghaffari, S. H. and Lobb, C.J. (1994) Structure and genomic organization of Vh gene segments in the channel catfish: members of different Vh gene families are interspersed and closely linked. Mol. Immunol. 31:823-832175. Hayman, J. R., Ghaffari, S. H. and Lobb, C. J. (1993) Heavy chain joining region segments of the channel catfish. J. Immunol. 151:3587-3596176. Ohno, S. Evolution by Gene Duplication, Springer Verlag New York Inc., 1970177. Early, P., Rogers, J., Davis, M., Calarne, K., Bond, M., Wall, R. and Hood, L. (1980) Two mRNAs can be produced from a single immunoglobulin p gene by alternative RNA processing pathways. Cell 20:313-319178. Petersson, M. L. and Perry, R. P. (1989) The regulated production of urn an ^s poly(A) site usage and the cn4-to-M1 splice, Mol. Cell Biol. 9:726-738179. Warr, G. W., DeLuca, D. and Griffin, B. R. (1979) Membrane immunoglobulin is present on thymic and spleenic lymphocytes of the trou t Salmo gairdneri. J. Immunol. 123:910-917180. Wilson, M. R., Ross, D. A., Miller, N. W., Clem, L. W., M iddleton, D. L. and Warr, G. W. (1995) Alternative pre-mRNA processing pathways in the production of membrane IgM heavy chains in holostean fish. Dev. Comp. Immunol. 19:165-177181. Reth, M. (1995) The B-cell antigen receptor complex and coreceptors. Immunol. Today 16:310-313182. Hombach, J., Lottspeich, F. and Reth, M. (1990) Identification of the genes encoding the IgM-a and IgM-ß components of the IgM antigen receptor complex by amino-terminal sequencing. Eur. J. Immunol. 20:2795- 2799183. van Ginkel, F. W., Miller, N. W., Cuchens, M. A. and Clem, L. W.(1994) Activation of channel catfish B cells by membrane immunoglobulin cross-linking. Dev. Comp. Immunol. 18:97-107
56

184. Schwager, J., Mikoryak, C. A. and Steiner, L. A. (1988) Am ino acid sequence of heavy chain from Xenopus laevis IgM deduced from cDNA sequence: Implications for evolution of immunoglobulin domains. Proc. Natl. Acad. Sci. USA 85:2245-2249185. Fellah, J. S., Wiles, M. V., Charlemagne, J. and Schwager, J. (1992) Evolution of vertebrate IgM: complete amino acid sequence of the constant region of Ambystoma mexicanum n chain deduced from cDNA sequence. Eur. J. Immunol. 22:2595-2601186. Ledford, B. E., Magor, B. G., Middleton, D. L., Miller, R. L., Wilson, M. R., Miller, N. W., Clem, W. and Warr, G. W. (1993) Expression of a mouse-channel catfish chimeric IgM molecule in a mouse myeloma cell. Mol. Immunol. 30:1405-1417187. Dayhoff, M. O. Survey of new data and computer methods of analysis. In M. 0. Dayhoff (ed.) Atlas of Protein Sequences and Structure, vol. 5, supp. 3, pp. 2-8, National Biomedical. Research Foundation, Silver Springs, 1978188. McLaughlin, P. J. and Dayhoff, M. O. Evolution of species and proteins: a time scale. In M. 0. Dayhoff (ed.) Atlas o f Protein Sequence and Structure, pp. 47-52, National Biomedical Research Foundation, Washington, 1972189. Ohta, T. and Kimura, M. (1971) Functional organization of genetic material as a product of molecular evolution. Nature 233:118-119190. Miyata, T., Yasunaga, T. and Nishida, T. (1980) Nucleotide sequence divergence and functional constraint in mRNA evolution. Proc. Natl. Acad. Sci. USA 77:7328-7332191. Kawamura, S., Tanabe, H., Watanabe, Y., Kurosaki, K., Saito, N. and Ueda, S. (1991) Evolutionary rate of immunoglobulin alpha noncoding region is greater in hominoids than in old world monkeys. Mol. Biol. Evol. 8:743-752192. Smith, G. R., Swirdczuk, K., Kimmel, P. G. and Wilkinson, B. H. (1982) Fish biostratiography of late Miocene to Pleiocene sediments of the western Snake River plain, Idaho. Idaho Bureau of Mines and Geology Bull. 26:519-541193. Smith, G. R. and Miller, R. R. (1985) Taxonomy of fishes from Miocene Clarkia beds, Idaho. In C. J. Smiley (ed.) Late Cenozoic History i f the Pacific Northwest, pp. 75-83, American Association for the Advance of Science, San Francisco, 1985194. Cavender, T. M. (1980) Systematics of Salvelinus from the North Pacific Basin. In E. K. Balon (ed.) Charrs, Salmonid Fishes o f the Genus Salvelinus, vol. 1, pp. 295-322, 1980195. McVeigh, H. P. and Davidson, W. S. (1991) A salmonid phylogeny inferred from mitochondrial cytochrome b gene sequences. J. Fish Biol. 39 (Suppl. A):277-282196. Devlin, R. H. (1993) Sequence of sockeye salmon type-1 and 2 growth hormone genes and the relationship of rainbow trout with Atlantic and Pacific salmon. Can. J. Fish Aquat. Sci. 50:1738-1748197. Winter, E., Radbruch, A. and Krawinkel, U. (1985) Members of novel Vh gene families are found in VDJ regions of polyclonaliy activated B- lymphocytes. EMBO J. 4:2861-2867
57

198r Ferguson, A. and Flemming, C. C. Evolutionary and taxonomic significance of protein variation in the brown trout (Salmo trutta L.) and other salmonid fishes. In G. S. Oxford and D. Rollinson (eds.) Protein polymorphism: Adaptive and Taxonomic Significance, pp. 85-99, Academic Press, New York, 1983
58