the effect of manganous ion on the kinetics and … journal of biological chemistry vol. 239, no. 5,...
TRANSCRIPT

THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 239, No. 5, May 1964
Printed in U.S.A.
The Effect of Manganous Ion on the Kinetics and
Mechanism of the Yeast Hexokinase Reaction*
VIRGINIA ZEWE,~ HERBERT .I. FROMM, AND R. FABIASO$
From the Guy and Bertha Ireland Research Laboratory, Department of Biochemistry, University of North Dakota School of Medicine, Grand Forks, North Dakota
(Received for publication, September 3, 1963)
The mechanism (mode of enzyme and substrate interaction) of the yeast hexokinase reaction was the subject of a recent report from this laboratory (1). The conclusions alluded t,o in that study were made on the basis of kinetic data obtained from experiments carried out in both the presence and absence of reaction product. The suggestion was made that yeast hexoki- nase reacts with the substrates adenosine triphosphate and D-
glucose in a random fashion leading to ternary complexes, and that all steps in the sequence are in rapid equilibrium except for the interconversion of the ternary complexes (1). It was further suggested that the presence of either substrate on the enzyme does not influence the binding of the other substrate.
Before the publication of the previously mentioned report, Hammes and Kochavi (23) concluded from kinetic experiments that the yeast hexokinase reaction probably involves an ordered pathway of enzyme and substrate interaction, in which n-glucose must add to hexokinase before the nucleotide substrate. On the other hand, Kaji, Trayser, and Colowick (4) observed that six times recrystallized hexokinase exhibits adenosine triphosphatase activity and that this activity, although very small in relation to the phosphotransferase activity, parallels the hexokinase activity throughout the purification and recrystallization procedures. These investigators also noted that inhibitors of glucose inhibited the adenosine triphosphatase activity of their hexokinase prep- arations. Such observations support the view that ATP does in fact add to hexokinase in the absence of the sugar substrate.
Silverstein (5), using the technique of equilibrium isotope exchange kinetics (6), obtained data which implicated a random pathway of substrate addition to yeast hexokinase. Cohn (7) has recently investigated the mechanism of this same reaction and has concluded from nuclear magnetic resonance experiments that glucose is probably required for the binding of ATP by hexoki- nase. These studies were carried out with the paramagnetic ion Mn*+ rather than Mg”+.
Because of the obvious confusion regarding the mechanism of yeast hexokinase as well as our continuing interest in this prob- lem, we decided to extend our original kinetic investigations. It was also thought that the lack of agreement between the
* This investigation was supported by Public Health Service Research Program Award GM-K3-16,226 and by a Grant A-1678 from the United States Public Health Service.
t Predoctoral Fellow of the United States Public Health Serv- ice.
f National Institutes of Health Medical Student Summer Re- search Fellow.
findings of Cohn (7) and those presented earlier by us might be explained on the basis of differences between Mgzf and Mn*+.
Data are presented in this report which appear to support our original contention regarding the mechanism of hexokinase when studies were carried out with Mg”+. In the presence of Mn*+, there appears to be an alteration in the reaction mech- anism; however, this effect is of a quantitative rather than a qualitative nature. On the basis of the studies to be presented in this report, a more detailed mechanism of substrate inter- action with hexokinase is presented.
EXPERIMENTAL PROCEDURE
M&rids-D-Glucose and n-fructose were products of Mal- linckrodt Chemical Works; n-mannose was obtained from Pfan- stiehl Laboratories, Inc. Magnesium chloride and manganese chloride were Fisher Analytical Grade reagents. Glucose 6- phosphate dehydrogenase (type V) was obtained from the Sigma Chemical Company, and phosphoglucose isomerase (three times recrystallized from rabbit muscle) was kindly donated by Dr. Ernst Noltmann. Crystalline yeast hesokinase was purchased from C. F. Boehringer und Soehne, and nucleotides, adenine, and adenosine, from Pabst Laboratories.
Methods-Hexokinase was assayed and found to be free of contaminating enzymes which could affect either the substrates or products of the system. Kinetic measurements were made in a fluorometer according to the procedure developed by Dalziel (8). Reaction mixture samples, in 0.06 M Tris-chloride buffer, pH 7.6, were kept in a water bath at 28” until used. Reactions were initiated by the addition of enzyme. Initial reaction veloc- ity was recorded as the molar concentration of product formed per 80 seconds. All reaction vessels contained TPN at a final concentration of 2 X 10e5 M, and glucose B-phosphate dehydro- genase in excess (1). In those cases in which fructose was included as a substrate, measurement of the product, glucose 6- phosphate, was accomplished by the addition of excess phospho- glucose isomerase.
Immediately before each series of kinetic experiments, the hexokinase was diluted with ice-cold water containing 2 mg of bovine serum albumin (Armour Laboratories) per ml and glucose 6.phosphate dehydrogenase. In those instances in which fructose was a substrate, phosphoglucose isomerase (20 pg of three times recrystallized enzyme per reaction tube) was also included.
Protein fluorescence measurements were made in an Aminco-
1625
by guest on July 7, 2018http://w
ww
.jbc.org/D
ownloaded from

1626 Mn2 f and Hexolcinase Kinetics Vol. 239, No. 5
Bowman spectrofluorometer equipped with a 150-watt Hanovia lamp. The cell housing of the instrument was modified to contain a water jacket through which water from a temperature- controlled bath, maintained at 28”, was circulated. For the calculation of dissociation constants, readings were taken directly from the electrometer. Additions of reagents to 0.1 mg of enzyme in l.O-ml buffer solutions in cuvettes were made by means of lo-p1 micropipettes. All calculations were corrected for dilution of solutions and fluorescence of buffers. The slit widths of the spectrofluorometer were combinations of sizes rec- ommended by the manufacturer to be used with a &inch pho- tomultiplier slit width. The spectrofluorometer was equipped with an RCA IP28 photomultiplier tube, and the wave length scaIe of the fluorescence monochrometer was calibrated with a mercury vapor lamp (American Instrument Company, Inc.). Further details of the procedure followed in the fluorescence experiments are presented elsewhere (9).
In all kinetic measurements the metal (Mg2+ or M&) to ATP ratio was maintained at 2.5 : 1.
RESULTS
Experiments with Mgz+-In the previous report, the mechanism of the yeast hexokinase reaction was suggested to involve a pathway of random interaction of substrates with enzyme leading to ternary complexes where all steps except the interconversion of the ternary complexes equilibrate rapidly (1). Furthermore, the earlier kinetic results indicated that the presence of one substrate on hexokinase does not influence the binding of the second substrate. Qualitatively, very similar data were obtained when initial reaction velocities were measured fluorometrically rather than spectrophotometrically. KATp and Kglucose were estimated to be 1.61 X 10e4 M and 8.35 X 10h5 M, respectively, by the former determination. If the mechanism suggested
0
4 6 8 IO WATP) x10-3 M
FIG. 1. a, plot of reciprocal initial velocity (V) versus reciprocal cedure.” b, plot of reciprocal of initial reaction velocity (V) of molar concentration of ATP. The concentrations of n-fructose versus reciprocal of molar concentration of n-fructose. The are: 13, 8.75 X 10e4 M; v, 2.19 X 10e4 M; 0, 1.095 X 10e4 iv; A, concentrations of ATP are: 0, 1.11 X 10e3 M; 0, 2.22 X lo+ M;
5.48 X lo+ M; 0, 3.84 X lo+ M. V was determined as a function a, 1.48 X 1OV M; 0, 9.86 X 10-e M. V was determined as a func- of ATP concentration, which was varied in the range from 1.11 X tion of n-fructose concentration, which was varied in the range 10-3 M to 9.86 X 10m5 &r. Velocities are expressed as moles per from 8.75 X lo+ M to 3.84 X 10-s M. V is expressed as in Fig. 1. liter of product formed per 80 seconds. The activating ion was The activating ion was Mg”+. Other experimental details are Mgz+. Other details are described under “Experimental Pro- given under “Experimental Procedure.”
originally for hexokinase was correct, one might expect that KATr would be the same with glucose or any other sugar substrate. In Fig. 1, a and b, are shown results obtained when fructose was employed as a substrate with hexokinase. It can be seen from the figures that the curves of the Lineweaver-Burk plots (10) intersect at a common point on the l/substrate axis. The KATP and Kfruotose values were calculated to be 2.0 X lop4 M
and 6.28 X 10e4 M, respectively. The kinetic parameters were determined by the method of Florini and Vestling (11). The similarity between these data and those obtained with glucose as a substrate as well as the consistent value of KATp would appear to support our original contention regarding the mechanism of the considered reaction (1).
As was pointed out earlier, inhibition with respect to AMP in the presence of Mgzf led to the conclusion that substrate addition to hexokinase occurs in a random fashion (1). Similar experi- ments were conducted with adenosine and adenine. Fig. 2, a and b, depicts results of inhibition experiments in which adenine was employed. Similar results were obtained with adenosine.
For the random pathway mechanism outlined above, if adenine acts as a competitive inhibitor of ATP, it may interact with free enzyme or the enzyme-glucose complex. Glucose may also bind the enzyme-adenine complex. The rate equation for this mechanism with inhibitor is
V= l+~+Jg(l+~j+~(l+;J (l)
where v, I/‘,,,, Ka, Ke, A, B, 1, KI, and KII represent observed velocity, maximal velocity, Michaelis constant of glucose, Mi- chaelis constant of AT??, glucose, ATP, adenine, dissociation con- stant for enzyme-adenine, and dissociation constant for enzyme-
(I/FRUCTOSE) x lO-3 M
by guest on July 7, 2018http://w
ww
.jbc.org/D
ownloaded from

Nay 1964 V. Zewe, H. J. Fromm, and R. Fabian0 1627
2.5
2.0
1.5
(D ‘0 ;i 1.0
--I>
0.5 I 2 3 4 5 6 7 8 9
(l/ATP)xlO-3M
FIG. 2. a, plot of reciprocal of initial reaction velocity (V) versus reciprocal of molar concentration of ATP in the absence and presence of adenine. The concentrations of adenine are: v,nOne; q ,y.b x lO+M; A,l.% X lo-'M; 0,2X lo-'M. D-f&- case was maintained constant at 2.5 X lo+ M. I’ was determined as a function of ATP concentration, which was varied in the range from 1.05 X 1W3 M to 1.05 X 10m4 M and is expressed as in Fig. 1. The activating ion was Mg*+. Other details are described under “Experimental Procedure.” b, plot of reciprocal of initial reac-
glucose-adenine, respectively. Because the binding of either substrate should not influence binding of the other substrate to the enzyme, it follows that KI = KII and KIII = Ka, where KIII
is taken to be the dissociation constant for the reaction, enzyme- glucose-adenine = enzyme-adenine + glucose.
If one assumes an ordered mechanism (2, 3) where glucose adds first to hexokinase, the following rate law is obtained with adenine as inhibitor.
VIll,X V=
1+$++% l+il +z ( 1
(2)
In Equation 2 it is assumed that adenine can react with the enzyme-glucose complex but not with the free enzyme. This assumption seems reasonable as ATP is the second substrate in the compulsory binding mechanism. KI and KII were calculated from the data of Fig. 2, a and b, and Equation 1 to be 1.47 x 10” M and 1.21 X lop2 M, respectively. When adenosine was used as the inhibitor, KI was calculated to be 1.07 x 10e2 M,
and KII, 1.39 X 1OW’ M. From earlier experiments with AMP (I), KI and KII were found to be 3.30 X 1O-3 M.
The data of Fig. 2, a and b, would appear to exclude the or- dered pathway mechanism with glucose acting as first substrate. Had this mechanism been applicable, hexokinase inhibition by adenine with respect to glucose would be of the uncompetitive type. It would appear that the findings presented in Fig. 2, a and 5, are most consistent with Equation 1 and in basic agree- ment with the random sequence hypothesis.
Although the investigations with competitive inhibitors of ATP and n-fructose are in harmony with the proposal of a random sequence of enzyme and substrate interaction, another kinetic approach was employed to test this hypothesis. In the experiments with n-mannose inhibition cited elsewhere (l), initial reaction velocity was measured by assaying glucose 6- phosphate production. If, however, one measures total product
-16 -12 -8 -4 0 4 8 I2 16
(I/GLUCOSE) x IO-3 M
tion velocity (V) versus reciprocal of molar concentration of n-glucose in the absence and presence of adenine. The concen- trations of adenine are: 0, none; 0, 6.9 X 10e3 M; 0, 2.0 X lo+ M; A, 4.0 X 10e2 M. ATP was maintained constant at 2.08 X 10V4 M. V was determined as a function of D-ghCOSf? concentra- tion, which was varied in the range from 1.0 X 10-s M to 6.35 X lOA M and is expressed as in Fig. 1. The activating ion was Mg2+. Other experimental details are described under “Experimental Procedure.”
formation, i.e. either ADP formation or sugar phosphate ester production, a completely different rate law is obtained. The kinetic equations which one obtains from this type of experiment permits, theoretically at least, a choice to be made from among the three most usual types of pathways of enzyme and substrate interaction (12).
In the case of the rapid equilibrium random mechanism pos- tulated for hexokinase if A, B, X, C, D, and XZ are taken to be ATP, n-glucose, n-fructose, ADP, glucose 6-phosphate, and fructose 6-phosphate, respectively, the following reaction se- quence is possible.
E+A=EA;Kl EXZ = ES2 + C; KS E+B=EB;Kz EXZ = EC + &; Kg
EA + B = EAB;Kg ES2 = E + Sz; K,, EB + A = EAB; K, EXY = ED + C; K11
E+S= ES;Kg EXY = EC + D; Klz EA + S = EAS;Ks ED = E f D; K,z ES+A=EAS;K? EC = E + C; K,,
h EAB iyz=d EXY,
kz
h EAS <- EXZ
ka
I f the assumption is made that all steps of the pathway equili- brate rapidly relative to interconversion of the ternary complexes, the following rate expression is obtained when total sugar phos- phate product is used to measure initial reaction velocity.
v= l+:-+gl+~j+~(l+g (3)
+ l+~+$(l+;j+~(l+;)
where V,, and Ti,, represent kr Eo and k~ Eo, respectively, and E. is taken to be total enzyme concentration. For the case in
by guest on July 7, 2018http://w
ww
.jbc.org/D
ownloaded from

Mn2 + and Hexolcinase Kinetics Vol. 239, No. 5
0.2
0 2 4 6 8 IO I
0 4 8 12 16 20 24
(I/GLUCOSE) x10-3M
FIG. 3. a, plot of reciprocal of initial reaction velocity (V) versus reciprocal of the molar concentration of ATP in the absence
b, plot of reciprocal of initial reaction velocity (V) versus the
and presence of n-fructose. n-Glucose concentration was con- reciprocal of the molar concentration of n-glucose in the absence and presence of n-fructose. ATP concentration was maintained
stant at 2.19 X lo+ M. The concentrations of n-fructose are: constant at 4.44 X 10e4 M. The concentrations of n-fructose are: 0, none; A, 1.44 X 10e4 M; 0, 4.37 X 10-h M; 0, 8.75 X lo+ M.
The lines are theoretical assuming Eauation 3 to be valid: the 0, none; A, 1.44 X lO-4 M; 0, 4.37 X 1oe4 M; 0, 8.75 X 10-d M.
points are experimental. V (glucose* 6-phosphate + fr&tose The lines are theoretical assuming Equation 3 to be valid; the
6-phosphate) was determined as a function of ATP concentration, points are experimental. V (glucose 6-phosphate + fructose
which was varied in the range from 4.44 X 10e4 M to 9.86 X 1O-5 M 6-phosphate) was determined as a function of n-glucose concen-
and is expressed as in Fig. 1. The activating ion was Mgz+. tration, which was varied in the range from 8.75 X 10d4 M to 3.84 X
Other details are as described under “Experimental Procedure.” 1OV M and is expressed as in Fig. 1. The activating ion was Mgz+. Other details are as described under “Experimental Procedure.”
which the presence of either substrate on the enzyme does not influence binding of the other substrate, K1 = K4 = K7, KB = KS, and Kg = KG.
In the case of the Theorell-Chance (13) and ordered ternary complex (12) mechanisms, a rate law similar to that of Equation
3 is obtained if an alternate substrate for the substrate adding initially to the enzyme is used. On the other hand, if an al- ternate substrate for the second substrate of the ordered mech- anism is used, rate expressions are obtained which permit a choice of mechanism to be made from the three under discussion. Although a more complete treatment of this approach will be presented elsewhere,’ it can readily be shown that for the ordered mechanisms, a l/velocity versus l/(first substrate) plot, in the presence of the second, plus alternate substrate, yields a linear relationship.
In Fig. 3, a and b, are presented dat,a obtained from kinetic experiments carried out with n-fructose as the alternate substrate. Cursory examination of these findings implies that n-glucose cannot be the first substrate of any ordered mechanism. The curves drawn in Fig. 3, a and 6, are theoretical lines if Equation 3 is assumed to be valid. These values for the kinetic parameters were obtained from independent experiments (see Fig. 1, a and b). It would appear from these kinetic investigations that substrates and hesokinase in the presence of Mg2+ interact randomly.
Erfect of ilfd+ on Kinetics of Yeast liexokinase-In light of the report by Cohn (7) on the mechanism of the yeast hexokinase reaction, it was desirable t,o study the reaction kinetics of the system with MnZ+. It was observed that in the concentration range of substrates invest.igated (see below) a ratio of Mnzf to ATP below 2 : 1 or above 3 : 1 caused inhibition and alteration of the KATP. For this reason the ratio of MnZf to ATP was main- tained at 2.5:1. The method used to evaluate the proper act.ivating ion to nucleotide ratio was employed elsewhere (1,14).
1 Manuscript in preparation.
In Fig. 4, a and b, are shown Lineweaver-Burk double recip- rocal plots of kinetic data with n-glucose and ATP. The point to note is that the curves in Fig. 4, a and b, converge at a common point above the l/substrate axis. Such findings are consistent with three mechanisms of substrate and enzyme interaction (12). KATP and KD-rlucose were calculated to be 1.35 X lop4 M and 6.65 X lop5 M, respectively. When these experiments were repeated with n-fructose in place of n-glucose, qualitatively similar results were obtained. The results of Fig. 4, a and b, are consistent with the following rate law.
V max v= l+Ee+!g+K!c (4)
KATp and Kn-rruetose were found to be 1.28 x lop4 M and 5.0 X lop4 M, respectively, in this case. liAn is taken to be a complex constant defined by Alberty (12).
It was of interest to compare these data with those obtained with Mg*;‘+ and further to compare the affinities of ATP in the presence of n-glucose and n-fructose. From all of the experi- mental data to be presented, it was finally concluded that the mechanism of the hesokinase reaction in the presence of the activating ion M&, is of the rapid equilibrium random type. However, the’presence of either substrate on the enzyme does influence the binding of the second substrate by the enzyme. In other words (KATP) (KS,,,,) is not equal to KATp.sugar. This conclusion accounts for the intersect,ion of the Lineweaver- Burk plots above the l/substrate axis in the second quadrant in Fig. 4, a and b. Such data are also consistent with two types of ordered pathways of enzyme and substrate interaction.
If the random pathway under consideration does in fact apply to hexokinase in the presence of MnZf, the dissociation constants for MnATP*- and MgATP2- should be the same with glucose and fructose provided the divalent ion is not of consequence relative to nucleotide and enzyme interaction. If the metal ion is im-
by guest on July 7, 2018http://w
ww
.jbc.org/D
ownloaded from

May 1964 V. Zewe, H. J. Fromm, and R. Fabian0 1629
+x10-6
a 0
]
s A
0 l
2 4 6 8 IO 12 14
(VATPI x IO-3 M
+x10-6
-8 -4 0 4 8 I2 I6 20 24 28
WGLUCOSE)xlO-3 M
FIG. 4. a, plot of reciprocal of initial reaction velocity (V) versus the reciprocal of the molar concentration of ATP. The concentrations of n-glucose are: v, 8.35 X lO-4 M; l ,1.05 X 10e4 M;
A, 5.3 X 10-s M; 0, 3.7 X 10m5 M; 0, 2.5 X 1Om6 M. V was de- termined as a function of ATP concentration, which was varied in the range from 9.54 X 10M4 M to 7.16 X lo-& M and is expressed as in Fig. 1. The activating ion was Mn2+. Other details are de- scribed under “Experimental Procedure.” b, plot of reciprocal
of initial reaction velocity (V) versus the reciprocal of the molar concentration of n-glucose. The concentrations of ATP are: v, 9.54 X lo-* M; 0, 1.9 X lo-” M; A, 1.23 X lo+ M; 0, 8.9 X
low5 M; 0, 7.2 X low5 M. V’ was determined as a function of D-glU-
case concentration, which was varied in the range from 8.35 X lo+ M to 3.84 X 10W5 M. V is expressed as in Fig. 1. The activat- ing ion was Mn2+. Other details are as described under “Experi- mental Procedure.”
0.4)j 2 4 6 8 IO I2 -8 -4 0 4 8 12 I6 20 24 28
(I/ATP)xIO-3M
FIG. 5. a, plot of reciprocal of initial reaction velocity (V) versus reciprocal of the molar concentration of ATP in the absence and presence of adenine. The concentrations of adenine are: 0, none; 0, 7.42 X 1OW M; 0, 1.06 X 1OV M; A, 2.12 X lo-* M.
n-Glucose concentration was maintained constant at 2.19 X 10-4~. V was determined as a function of ATP concentration, which was varied in the range from 1.12 X 1OW l\r to 8.74 X lop5 M and is expressed as in Fig. 1. The activating ion was Mn*+. Other details are described under “Experimental Procedure.” b, plot
portant in ATP and enzyme interaction, KATf with n-glucose
and n-fructose should be the same; however, there would be a difference between these values for KATp with Mg2+ and M&.
From the data already presented, it would appear that the ion
does not affect binding of the nucleotide by the enzyme with either metal ion. This point will be elaborated upon when the experiments of protein fluorescence quenching are described. Such results serve to explain why one may attribute a rapid equilibrium random mechanism to the yeast hexokinase system in light of the findings of Cohn (7).
In order to ascertain whether the pathway of the hexokinase reaction is in fact ordered in the presence of M& with glucose
(I/GLUCOSE)xlCT3M
of reciprocal of initial reaction velocity (V) versus reciprocal of the molar concentration of n-glucose in the absence and presence of adenine. The concentrations of adenine are: V, none; 17, 6.93 X 10M3 M; A, 9.90 X lo+ l”r; 0, 1.98 X 10e2 M. ATP concen- tration was maintained constant at 2.2 X 1OW M. V was de- termined as a function of n-glucose concentration, which was varied in the range from 8.75 X low4 M to 3.84 X lo-’ M and is expressed as in Fig. 1. The activating ion was Mn2+. Other details are given under “Experimental Procedure.”
as first substrate, experiments with adenine, adenosine, and D-
mannose were undertaken. In Fig. 5, a and b, are shown data on the effect of adenine on the kinetics of the system under discussion. It can be seen from Fig. 50, that adenine functions as a competitive inhibitor of ATP with Mn2+ as activating ion. Such findings are consistent with results obtained in the presence of MgZ+; however, the data of Fig. 5b suggest an important difference between the two ions. If the random pathway were applicable, inhibition with respect to glucose should be noncom- petitive. On the other hand, with glucose as the initial substrate in the ordered mechanism, inhibition with respect to glucose, as suggested above, should be uncompetitive. Cursory exami-
by guest on July 7, 2018http://w
ww
.jbc.org/D
ownloaded from

1630 Mn2+ and Hexokinase Kinetics
+x10-6
I L -6 -3 0 3 6 9 12
(VATPhlO-3M
FIG. 6. a, plot of reciprocal of initial reaction velocity (V) versus reciprocal of the molar concentration of ATP in the absence and presence of n-mannose. The concentrations of o-mannose are: v, none; 0, 1.875 X IOm4 M; A, 3.75 X 10e4 M; Cl,?‘.5 X 10e4 M.
o-Glucose was maintained constant at 2.19 X 10e4 K V was determined as a function of ATP concent,ration, which was varied in the range from 1.11 X 1OW M to 8.75 X 10m5 IM and is expressed as in Fig. 1. The activation ion was Mn*+. Other details are as described under “Experimental Procedure.” b, plot of reciprocal
nation of the data of Fig. 5b indicates a mixed type of inhibit,ion, i.e. intermediate between the two types. The data of Fig. 5, a and b, are consistent with the ordered mechanism where adenine acts as a competitive inhibitor of ATP if, and only if, ATP func- tions as the first substrate. No one who has investigated the mechanism of the hexokinase reaction has made this suggestion to date.
The results of Fig. 5, a and b, can be explained two ot,her ways. The first of these assumes an ordered pathway with glucose as initial substrate where glucose and adenine compete for the same enzymatic site. Because of the structural dissimilarity between these two compounds, this suggestion would appear highly unlikely. The other possibility is one in which eit,her adenine or adenosine acts as a competitive inhibitor of ATP. The most probable mechanism would be of the rapid equilibrium type; however, binding of either substrate to the enzyme influences addition of the other substrate. If one assumes that adenine (1) reacts with hesokinase in the following fashion
E + I F? EI; KI EA + I F? EAI; KII EI + A F’t EAI; Krr~
the following rate equation is obtained.
v= l+e$+~(l+vn)+qk”(l+i,) C5)
where A and B represent glucose and ATP, respectively. Equa- tion 5 appears to account for the data of Fig. 5, a and b, and assumes only that adenine competes for the ATP-binding site of hesokinase. The inhibition constants are 3.23 X lop2 M and 5.46 x 10m2 M, respectively, for K1 and KI1 with adenosine, and 2.2 x 10e2 M and 2.95 X 1OW M, respectively, for K1 and KI1 with adenine. As previously defined, KIII = KAKII/KI.
By virtue of the fact that the random sequence of substrate and enzyme interaction is symmetrical, with respect to either
b 4 8 12 k
(I/GLUCOSE) x IO-3 M
of initial reaction velocity (V) versus reciprocal of the molar concentration of n-glucose in the absence and presence of n-man- nose. The concentrations of o-mannose are: 0, none; q , 6.9 X 10-K M; A, 1.04 X low4 M; v, 3.13 X 1OW M. ATP was maintained constant at 2.08 X 1OW 1~. V was determined as a function of n-glucose concentration, which was varied in the range from 1.0 X 10-s 1~ to 6.4 X 10e6 M and is expressed as in Fig. 1. The activating ion was Mn*+. Other details are as described under “Experimental Procedure.”
substrate, a similar effect should be observed with a competitive inhibitor for either substrate. u-Mannose, a competitive in- hibitor of n-glucose (15), was chosen to test further the hypothesis put forth to explain the kinetic findings with Mn2+. The data obtained from these experiments are shown in Fig. 6, a and b. It is apparent from these graphs that n-mannose acts, as might be expected, as a competitive inhibitor of n-glucose. Inhibition with respect to ATP appears mixed. Equation 5 can be utilized to evaluate the dissociation constants K1 and KI1. In this situation A represents ATP, B represents n-glucose, and I is taken to be n-mannose. The calculated values for K1 and KI1 are 1.49 X 10h4 M and 2.61 X lop4 M, respectively. It is to be noted that the dissociation constant of the mannose-enzyme complex (K,) agrees favorably with the value for this constant determined in the absence of glucose (15). It would appear from the data presented in Fig. 6, a and b, that the rapid equilibrium random mechanism is applicable to yeast hexokinase with Mn2+ as activating ion, where the presence of either substrate on the enzyme influences binding of the other substrate.
The mechanism of the hexokinase reaction in the presence of XI& was investigated with alternate substrate as outlined above. The results of these studies are presented in Fig. 7, a and b. Generally, the same conclusions can be drawn from the experi- ments with Mg2+ and Mn2+. The theoretical curves shown in Fig. 7 were obtained with the use of Equation 3 as suggested above. These alternate substrate data are consistent with the rapid equilibrium random mechanism already suggested.
The various kinetic parameters calculated from the data of Figs. 1 to 7 are presented in Table I.
Fluorescence Experiments with Hexokinase2-From the fore-
2 Trayser and Colowick (21) reported no change in the fluores- cence spectrum of hexokinase upon the addition of D-glUCOSe or MgATP*-. In a personal communication, Dr. Colowick has suggested that the opposing results might be due to the differences in assay pH (the earlier study was carried out at pH 8.2) or to the possibility that different chromatographic forms of the en- zyme were used.
by guest on July 7, 2018http://w
ww
.jbc.org/D
ownloaded from
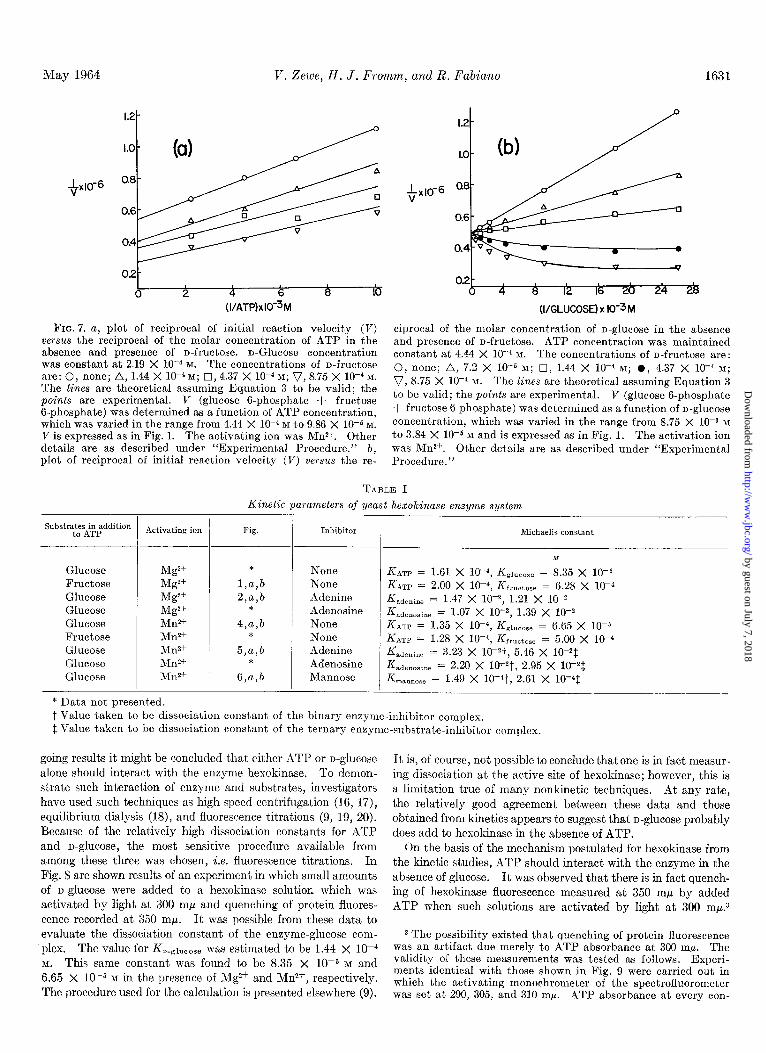
May 1964 V. Zewe, H. J. Fromm, and R. Fabian0 1631
+x10-6 +x10-6
02-
0 2 4 6 8 16 (I/ATP)xlO-3M (I/GLUCOSE) x IO-3 M
FIG. 7. a, plot of reciprocal of initial reaction velocity (V) ciprocal of the molar concentration of n-glucose in the absence versus the reciprocal of the molar concentration of ATP in the and presence of n-fructose. ATP concentration was maintained absence and presence of n-fructose. n-Glucose concentration constant at 4.44 X 10W4 K The concentrations of n-fructose are: was constant at 2.19 X 10e4 M. The concentrations of n-fructose are: 0, none; A, 1.44 X 10m4 M; cI,4.37 X 10m4 M; v, 8.75 X 10m4 M.
0, none; A, 7.2 X lo+ M; 0, 1.44 X 1OP M; 0, 4.37 X 10e4 M;
The lines are theoretical assuming Equation 3 to be valid; the v, 8.75 X 10V4 M. The lines are theoretical assuming Equation 3
points are experimental. V (glucose 6-phosphate + fructose to be valid; the points are experimental. V (glucose 6-phosphate
B-phosphate) was determined as a function of ATP concentration, + fructose 6-phosphate) was determined as a function of n-glucose
which was varied in the range from 4.44 X 1O-4 M to 9.86 X 10e5 M. concentration, which was varied in the range from 8.75 X 1OW M
V is expressed as in Fig. 1. The activating ion was Mn2+. Other to 3.84 X 10-b M and is expressed as in Fig. 1. The activation ion details are as described under “Experimental Procedure.” b, was Mn2+. Other details are as described under “Experimental plot of reciprocal of initial reaction velocity (V) versus the re- Procedure.”
TABLE I
Kinetic parameters of yeast hexokinase enzyme system
SubstytF.+paddition Activating ion Fig. Inhibitor Michaelis constant
Glucose Mg2+ Fructose Mg2+ Glucose Mg2+ Glucose Mg2+ Glucose Mn2+ Fructose Mn2+ Glucose Mn2+ Glucose Mn2+ Glucose Mn2+
* Data not presented.
*
l,a,b 2,a,b
*
4,a,b *
5,a,b *
6,a,b
Ax None Km = 1.61 X 10-4, i&~ueose = 8.35 X 1O-5 None KATP = 2.00 X W4, &ruetose = 6.28 X 1o-4 Adenine K adenine = 1.47 x 10-2, 1.21 x 10-Z Adenosine Kadenosine = 1.07 X 10e2, 1.39 X 1OW None KATp = 1.35 X 10-4, f&.~ucose = 6.65 X 10-j None K&p = 1.28 x 10-4, Kfructose = 5.00 x 10-4 Adenine K &den& = 3.23 X 10--2t, 5.46 X lOV$ Adenosine Kadenosine = 2.20 X KVt, 2.95 X 10-Z$,
Mannose KmannOse = 1.49 X IO-*?, 2.61 X lo-41
t Value taken to be dissociation constant of the binary enzyme-inhibitor complex. $, Value taken to be dissociation constant of the ternary enzyme-substrate-inhibitor complex.
going results it might be concluded that either ATP or n-glucose alone should interact with the enzyme hexokinase. To demon- strate such interaction of enzyme and substrates, investigators have used such techniques as high speed centrifugation (16, 17),
equilibrium dialysis (18), and fluorescence titrat.ions (9, 19, 20). Because of the relatively high dissociation constants for ATP
and u-glucose, the most sensitive procedure available from
among these three was chosen, i.e. fluorescence titrations. In Fig. 8 are shown results of an experiment in which small amounts
of o-glucose were added to a hexokinase solutior, which was activated by light at 300 rnp and quenching of protein fluores- cence recorded at 350 mp. It was possible from these data to evaluate the dissociation constant of the enzyme-glucose com- plex. The value for F&iucose was estimated to be 1.44 X 10e4 M. This same constant was found to be 8.35 X 1O-5 M and 6.65 X lop5 M in the presence of Mg2+ and M&, respectively. The procedure used for the calculation is presented elsewhere (9).
It is, of course, not possible to conclude thatone is in fact measur- ing dissociation at the active site of hexokinase; however, this is a limitation true of many nonkinetic techniques. At any rate, the relatively good agreement between these data and those obtained from kinetics appears to suggest that n-glucose probably does add to hexokinase in the absence of ATP.
On the basis of the mechanism postulated for hexokinase from the kinetic studies, ATP should interact with the enzyme in the absence of glucose. It was observed that there is in fact quench- ing of hexokinase fluorescence measured at 350 rnp by added ATP when such solutions are activated by light at 300 mp.3
3 The possibility existed that quenching of protein fluorescence was an artifact due merely to ATP absorbance at 300 rnp. The validity of these measurements was tested as follows. Experi- ments identical with those shown in Fig. 9 were carried out in whieh the activating monochrometer of the spectrofluorometer was set at 290, 305, and 310 rnp. ATP absorbance at every con-
by guest on July 7, 2018http://w
ww
.jbc.org/D
ownloaded from

1632 Mn2 + and Hexolcinase Kinetics Vol. 239, No. 5
96
n A
70 1 I 0 2 4 6 8 IO 12 14 16
GLUCOSE x 105M
FIG. 8 (left). Protein fluorescence quenching curve of hexo- kinase in the presence of n-glucose. The protein was activated with light at 300 mM, and fluorescence quenching was measured at 350 rnp. The enzyme (0.1 mg) was added to 1.0 ml of 0.06 M Tris-chloride buffer, pH 7.6. n-Glucose additions (10 ~1) were made with micropipettes, and the sample was corrected for dilu- tion. In the figure the points are experimental and the line is theoretical, assuming a dissociation constant for the enzyme- glucose complex of 1.44 X 10e4 M. At infinite n-glucose concentra- tion, hexokinase fluorescence is quenched 30%. Quenching at infinite substrate concentration was determined from a plot of l/fluorescence versus l/substrate concentration (9).4
FIG. 9 (right). Protein fluorescence quenching curve in the
The dissociation constants for MgATP2- and MnATP2- were determined to be 2.40 X 10d4 M and 2.06 X 10e4 M, respectively. It was quite surprising to find that ATP4- quenched hesokinase
fluorescence to the same extent as the metal chelate complexes of
centration employed in Fig. 9 was determined at 290,300,305, and 310 mp in the Beckman DU spectrophotometer. The only meas- urable decrease in transmittance was noted at 290 mp, and a suitable correction for this effect was made (9) for calculation of the dissociation constant of the ATP-enzyme complex. The values for &rp obtained with activating light at 290, 305, and 310 rnr were in good agreement with those determined from the data of Fig. 9. Similar experiments were carried out with adenine in place of ATP, but at twice the nucleotide concentrations shown in Fig. 9. There was no evidence of protein quenching upon the addition of adenine when a suitable correction was made for adenine absorbance. These findings appear to add credence to those data obtained in the quenching experiments with ATP.
4 The percentage of quenching at infinite substrate concentra- tion, referred to in the legends to Figs. 8 and 9, was determined from a double reciprocal plot of l/AF against l/substrate concen- tration by the equation l/AF = l/(0’ - @)Eo + K/Q’ - /~)Eo&, where AP = F - LEO. F, p, fl’, Ea, K, and & represent observed fluorescence, molar emittance of free enzyme, molar emittance of the enzyme-substrate complex, total enzyme concentration, dissociation constant of the enzyme-substrate complex, and total substrate concentration, respectively. This equation permits evaluation of K without a knowledge of 8, p’, and EO and is valid in cases where the amount of substrate in the form of the enzyme- substrate complex is very small compared to total substrate. The percentage of quenching was evaluated from the relationship (fluorescence in absence of substrate minus fluorescence at infinite substrate concentration) X lOO/(fluorescence in absence of sub- strate), where fluorescence at infinite substrate concentration was determined by extrapolation of the data shown in Figs. 8 and 9 to the l/AF axis.
I I 1 I 0 0.5 1.0 1.5 2.0 2.5 3.0
ATPx 104M
presence of ATPJ- (o), MgATP” (O), and MnATP2- (A), and hexokinase. The protein was activated with light at 300 rnp, and the fluorescence was measured at 350 rnN. The enzyme (0.1 mg) was added to 1.0 ml of 0.06 M Tris-chloride buffer, pH 7.6. Nucleo- tide additions were made with micropipettes (10 Al), and all sam- ples were corrected for dilution. In the figure the points are experimental, and the lines are theoretical. The upper curve was drawn assuming a dissociation constant for the enzyme-nucleotide complex of 2.4 X 10e4 M, while the Zmuer curve assumes a dissocia- tion constant of 2.06 X lo-” 1~. At infinite nucleotide concentra- t.ion, hexokinase fluorescence is quenched 23%. Quenching at infinite substrate concentration was determined from a plot of l/fluorescence versus l/substrate concentration (9) .4
TABLE II
Dissociation constants of hexokinase and substrates determined from jluorescence titrations
Substrate Fig. Dissociation constant
M
Glucose.................... 8 1.44 X 10-4
ATP4- 9 1.48 X 1O-4 MgATPz- 9 2.40 X 1O-4 MnATP2-. 9 2.06 X 1O-4
the nucleotide. Further, the KATpd- was very similar (1.48 x 10m4 M) to dissociation constants determined in the presence of metal ion. The divalent metal ions, Mg2+ and Mn&, had no effect on the hexokinase fluorescence. The data obtained from the fluorescence experiments are summarized in Table II. Such results lead to two rather tempting speculations: first, that ATP does not require n-glucose to interact with hexokinase, a con- clusion which is supported by both the kinetic and fluorescence data; and second, that the end, or divalent metal ion pyrophos- phate portion of ATP is not involved in the interaction with hexokinase in the absence of n-glucose. This latter extrapola- tion, in view of the fluorescence quenching experiments, would seem to explain Cohn’s (7) nuclear magnetic resonance results.
Possibly the pyrophosphate-Mn2+ portion of ATP reacts only with glucose, or with the protein after a glucose-induced con- formational protein alteration. At any rate, in light of these fluorescence data, it is not surprising that hexokinase did not induce a significant change in the nuclear magnetic resonance experiments reported by Cohn (7) in the absence of n-glucose.
by guest on July 7, 2018http://w
ww
.jbc.org/D
ownloaded from

May 1964 V. Zewe, Ii. J. Fromm, and R. Fabian0 1633
DISCUSSION
The purpose of the present investigation was to provide evidence that would either support or refute our original proposal (1) regarding the mode of substrate and yeast hesokinase inter- action. Data are available in the literature which appear to be in harmony with the rapid equilibrium random mechanism; however, other well documented findings tend to support an ordered sequence of enzyme and substrate interaction with glucose as initial substrate.
In the present report it is suggested that adenosine and adenine in the presence of Mg2+ act as competitive and noncompetitive inhibitors of ATP and glucose, respectively. The experimental methods used for these kinetic studies are technically simple, and the assumptions made to account for such inhibition minimal in number. These experiments probably provide the strongest evidence in support of the rapid equilibrium random mechanism proposed earlier (I). For an ordered mechanism with glucose as the leading substrate, a competitive inhibitor of ATP would give uncompetitive inhibition with respect to glucose.
illthough the observation that the L&weaver-&irk plots shown in Fig. 1, a and b, with fructose as substrate and analogous experiments with glucose exhibit, a common intercept on the abscissa may be considered fortuitous, data obtained from these and other experiments are consistent with the random pathway mechanism. Finally, the results obtained in experiments with fructose as the alternate substrate are in accord with the original thesis regarding the mode of enzyme and substrate interaction
(1). The only suggestion that can be offered to reconcile the kinetic
data from this laboratory and those presented by Hammes and Kochavi (2, 3) might possibly involve hesokinase itself. It is known, for example, that the enzyme exists in chromatographi- tally separable forms.” Possibly some of these isozymes may exhibit different kinetic properties.
Although recrystallized hesokinase is known to contain glucose (al), Trayser and Colowick were able to show that each mole of enzyme is associated with only 0.25 mole of sugar and that this glucose is probably not associated with the active site of the en- zyme. On the other hand, these workers observed that glucose was capable of protecting hesokinase against tryptic digestion (21). Such findings suggest that the sugar substrate may add to the enzyme in the absence of ATP.
That ATP may interact at the active site of hesokinase in t.he absence of glucose is suggested by the report that the highly puri- fied enzyme exhibits ATPase activity, and that this activity is inhibited by inhibitors of glucose (4). The assumption is made that water rather than glucose acts as phosphoryl acceptor in the absence of the sugar substrate. The fact that the ilTPase activ- ity is relatively very low compared to glucose does not appear to weaken the argument that the phosphatase activity is actually hesokinase activity with water taking over the function of glu- cose. A wide spectrum of activities has been observed for differ- ent sugars with yeast hesokinase (22). The findings relative to the independent binding of ATP and glucose by hesokinase would appear to support our original suggestion on the mecha- nism of hexokinase (1); however, Kaji et al. (4) have observed that KATP is 50 times great.er with ATPase than with herokinase. This would provide evidence against the random pathway mecha-
5 S. P. Colowick, personal communication.
nism; however, it should be pointed out that the ATPase deter- minations require much greater concentrations of enzyme than those employed for hesokinase measurements5 Possibly the difference in the kinetic paramet,ers can be explained on the basis of different states of enzyme association in the two assay pro- cedures.
The strongest evidence in opposition to the random pathway mechanism of substrate interaction with hexokinase was provided by Cohn (7) from nuclear magnetic resonance studies of metal activation. She observed that glucose was required for the binding of MnADP- to hesokinase. These findings would ap- pear to support the kinetic studies of Hammes and Kochavi (2,3), which implicated the ordered pathway of enzyme and sub- strate interaction with glucose as initial substrate.
The kinetics of hexokinase was reinvestigated as it was felt that the disagreement betwern our kinetic studies (1) and the re- port of Cohn (7) might have been attributable to the fact that an attempted comparison of the same mechanism was being under- taken with different divalent met.al ions @Mg2+ and Mn2+). The kinetic data presented in this report with Mn2+ still suggest a random pathway of hesokinase interaction with substrates with only minor modifications relative to results obtained with Mg2+.
It would seem that all of the kinetic data contained in the present report for the hesokinase system are consistent with the random sequence pathway. This is particularly true regarding the inhibition studies with adenosine, adenine, and mannose.
The results of hesokinase fluorescence quenching presented here are also in agreement with the random pathway mechanism under consideration. Quenching of hesokinase fluorescence was observed m-ith glucose, ATP-, MgATP*-, and MnATP2-, but not with either Mgz+ or Mn2+. Furthermore, the fact that the dissociation constants obtained from these experiments are con- sistent with the analogous constants obtained from kinetic esperi- ments would imply interact.ion of these compounds at the active site of hesokinase. It is of course not possible to say with cer- tainty that such is the case; however, most other nonkinetic approaches suffer from similar limitations (16, 18).
The most interesting result obtained from the fluorescence quenching experiments is the interaction of ATP4- with heso- kinase in the absence of either glucose or divalent metal ion. The KATp4- calculated was found to be essentially equal to K M~ATPP- and KM~ATP~-. From the inhibition studies with adenine, adenosine, and ilMP (I), it would appear that the ade- nine portion of A4TP can be bound to hesokinase. The fact that ITP is a relatively weak substrate for hesokinase (23) suggests that the ATP is probably bound through the amino group of C-6. &cause adenine and adenosine exhibit similar inhibition constants, it is tempting to suggest that the sugar moiety is very weakly bound to the enzyme.
The data obtained from the fluorescence experiments, which suggest that ATP4- may add to hesokinase, are consistent with the kinetic results, which imply that adenine and adenosine, compounds not associat.ed with divalent metal ions, compete wit.h MgATP2- and MnATP2- for the same enzymatic locus. It would appear from these studies that nucleotide substrate binding to the enzyme does not involve the participation of the metal ion directly.
It should be emphasized that the fluorescence quenching data present.ed in this report are subject to serious criticisms. In the case of hesokinase fluorescence quenching by glucose, only a IO y0
by guest on July 7, 2018http://w
ww
.jbc.org/D
ownloaded from

l/In2 + and Hexokinase Kinetics Vol. 239, No. 5
decrease in protein fluorescence was observed experimentally. The experimentally recorded decrease in fluorescence was ap- proximately 127, with ATP. These data were subsequently extrapolated (see footnote 4) to 30% and 23% at infinite glucose and ATP concentrations, respectively. Thus, these measure- ments represent small differences between two large and similar values. On the other hand, the experiments involving quenching of hexokinase fluorescence by glucose and ATP were repeated six different times. These changes, although small, appear to be reproducible. A similar effect was observed by Velick (19) when DPN was used to quench glyceraldehyde 3-phosphate dehy- drogenase fluorescence.
If one accepts the foregoing suggestions, it is possible to rationalize t,he results of Cohn (7) with the random pathway sequence. Here it may be assumed that MnATP2- interacts in the absence of glucose; however, the chelated portion of the molecule is not involved in enzyme binding directly. In the presence of sugar, the M& portion of MnATP2- interacts either with glucose or with a site on the enzyme which in turn gives rise to the active quaternary complex.
The recent findings of Silverstein (5) are in harmony with the mechanism for yeast hexokinase presented in this report. It would appear that enzyme-phosphate (24, 25) and enzyme-glu- cose (26) intermediates do not participate in the hexokinase reaction.
SUMMARY
The mechanism of the yeast hexokinase reaction was investi- gated kinetically with inhibitors of n-glucose and adenosine triphosphate, an alternate substrate of n-glucose, and fluorescence quenching of the hesokinase spectrum by substrates.
The mechanism of the reaction in the presence of Mg2+ appears to involve a random addition of substrates to hesokinase, in which the presence of either substrate on the enzyme does not influence binding of the second substrate. In the presence of M&, the same basic mechanism prevails; however, the presence of either substrate on the enzyme does appear to influence binding of the other substrate to hexokinase.
Neither divalent metal ion affects the fluorescence spectrum of hexokinase (measured at 350 mp); however, n-glucose and adenosine triphosphate quench protein fluorescence 30 y0 and 23%, respectively. This effect is approximately the same with free nucleotide substrate and in the presence of divalent metal ion. The dissociation constants calculated from these experi-
ment,s are consistent with the corresponding constants evalua,ted from kinetic experiments.
It is suggested that the nucleotide substrate is bound to hexo- kinase primarily through the adenosine 5’-phosphate moiety in the absence of n-glucose. In the presence of the sugar, there is a conformational change in the enzyme which gives rise to a quaternary complex involving divalent metal ion and the pyro- phosphate portion of adenosine triphosphate.
REFERENCES
1. FROMM, H. J., AND ZEWE, V., J. Biol. Chem., 237, 3027 (1962). 2. HAMMES. G. G.. AND KOCHAVI. D.. J. Am. Chem. Sot.. 84,
3. 2069 (i962). ’
I I ,
HAMMES,G. G., AND KOCHAVI,D., J. Am. Chem.Soc.,84,2073 (1962).
4. KAJI, A., TRAYSER, K. A., AND COLOWICK, S. P., Federation Proc., 20, 233 (1961).
5. 6. 7. 8. 9.
10.
SILVERSTEIN, E., Ph.D. thesis, University of Minnesota, 1963. BOYER, P. D., Arch. Biochem. Biophys., 82,387 (1959). COHN, .M., Biochemistry, 2, 623 (i963). DALZIEL. K.. Biochem. J.. 84. 244 (1962). FROMM, k. j., J. Biol. Chem.; 238, ‘2938’ (1963). LINEWEAVER, H., AND BURK, D., J. Am. Chem. Sot., 56, 658
(1934). 11.
12. 13.
14. 15.
FLORINI, J. R., AND VESTLING, C. S., Biochim. et Biophys. Acta, 25, 575 (1957).
ALBERTY, R. A., J. Am. Chem. SOL, 75, 1928 (1953). THEORELL, H., AND CHANCE, B., Acta Chem. &and., 6, 1127
(1951). FROMM, H. J., AND ZEWE, V., J. Biol. Chem., 237, 1661 (1962). SLEIN, M.W.,inS.P. COLOWICHANDN.O.KAPLAN (Editors),
Methods in enzymology, Vol. III, Academic Press, Inc., New York, 1957, p. 154.
16.
17.
18.
19. 20.
21.
22.
23. 24.
25.
26.
HAYES, J. E., JR., AND VELICK, S. F., J. Biol. Chem., 207, 225 (1954) .
TAKENIIK.4, Y., ANI) SCHWERT, G. W., J. Biol. Chem., 223, 157 (1956).
VELICK, S. F., AND HAYES, J. E., JR., J. Biol. Chem., 203,545 (1953).
VELICK, S. F., J. Biol. Chem., 233, 1455 (1958). WINER, A. D., SCHWERT, G. W., AND MILLAR, D. B. S., J.
Biol. Chem., 234, 1149 (1959). TRAYSER, K. A., AND COLOWICK, S. P., Arch. Biochem. Bio-
phys., 94, 169 (1961). SOLS. A.. DELAFUENTE.G..VILLAR-PALASI,C..ANDASENSIO.
C.,‘Bikhim. et Biophys. Acta, 30,92 (1958). ’ MARTINEZ, R. J., Arch. Biochem. Biophys., 93, 508 (1961). AGREN, G., AND ENGSTRBM, L., Acta Chem. Scand., 10, 489
(1956). AGREN, G., ENGSTR~M, L., AND EKLUND, S., Biochim. et Bio-
phys. Acta, 67, 143 (1963). NAJJAR, V. A., AND MCCOY, E. E., Federation Proc., 17, 1141
(1958).
by guest on July 7, 2018http://w
ww
.jbc.org/D
ownloaded from

Virginia Zewe, Herbert J. Fromm and R. FabianoHexokinase Reaction
The Effect of Manganous Ion on the Kinetics and Mechanism of the Yeast
1964, 239:1625-1634.J. Biol. Chem.
http://www.jbc.org/content/239/5/1625.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/239/5/1625.citation.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on July 7, 2018http://w
ww
.jbc.org/D
ownloaded from