the effect of folate-related snps on clinicopathological features, response to neoadjuvant treatment...
TRANSCRIPT

Gene 518 (2013) 397–404
Contents lists available at SciVerse ScienceDirect
Gene
j ourna l homepage: www.e lsev ie r .com/ locate /gene
The effect of folate-related SNPs on clinicopathological features, response toneoadjuvant treatment and survival in pre- and postmenopausal breastcancer patients
Nataliya Babyshkina a,⁎, Elena Malinovskaya a, Mariya Nazarenko c, Mariya Koval c, Polina Gervas a,Olga Potapova d, Elena Slonimskaya b, Nadejda Cherdyntseva a
a Department of Experimental Oncology, Cancer Research Institute of Siberian Branch of Russian Academy of Medical Sciences, Tomsk, 634050, Russian Federationb Department of General Oncology, Cancer Research Institute of Siberian Branch of Russian Academy of Medical Sciences, Tomsk, 634050, Russian Federationc Research Institute of Medical Genetics of Siberian Branch of Russian Academy of Medical Sciences, Tomsk, 634050, Russian Federationd Cureline, Inc., South San Francisco, CA 94080, USA
Abbreviations: BMI, body mass index; 95% CI, confidephamide, methotrexate and 5-fluorouracil; DNMT3b, Ddihydrofolate reductase; ER, estrogen receptor; FAC, 5cyclophosphamide; HER2, human epidermal growthmethylenetetrahydrofolate dehydrogenase; MTHFR, mettase; MTR, methionine synthase; MTRR, methionine sratios; PCR-RFLP, polymerase chain reaction restrictiophism; PFS, progression-free survival; PR, progesteronecarrier 1; SNP, single nucleotide polymorphism; TS, thy⁎ Corresponding author at: Cancer Research Institute of S
emy of Medical Sciences, Department of Experimental OTomsk, 634050, Russian Federation. Tel.: +7 3822 512529;
E-mail address: [email protected] (N. Babyshkin
0378-1119/$ – see front matter © 2013 Elsevier B.V. Allhttp://dx.doi.org/10.1016/j.gene.2012.12.095
a b s t r a c t
a r t i c l e i n f oArticle history:Accepted 20 December 2012Available online 4 January 2013
Keywords:Breast cancerFolate polymorphismMenopausal statusNeoadjuvant chemotherapySurvival
This study aimed to investigate the relationship of ten single nucleotide polymorphisms (SNPs) in the MTHFR,MTR,MTRR,DHFR,MTHFD1, TS,RFC1 andDNMT3bgeneswith cancer survival, therapeutic response to neoadjuvantchemotherapy and clinicopathological characteristics in 300 pre- and postmenopausal breast cancer patients of aRussianWestern Siberian population.We found that theMTHFR 677CT genotype aswell as combination ofMTHFR677CT and 677TT genotype was related to tumor size and estrogen-positive status in postmenopausal group. TheRFC1 80А allelewas associatedwith an increased risk of lymph nodemetastases among postmenopausal women.TheMTHFR 677TT genotypewas significantly correlatedwith a better progression-free survival in premenopausalpatients. In contrast, a worse outcomewas observed in this group patient withMTHFD1 1958AA genotype. In themultivariate analysis, the MTHFD1 1958AA genotype was identified as an independent prognostic factor forpremenopausal breast cancer survival. Our findings provide evidence for associations of breast cancer survivalwith folate-related SNPs in a population of Western Siberian region of Russia and the MTHFD1 (1958G>A) mayhave additional prognostic value especially among premenopausal patients.
© 2013 Elsevier B.V. All rights reserved.
1. Introduction
Breast cancer remains the most frequent malignant disease in theRussian Federation and it is one of the leading causes of cancer-relateddeath (Davydov and Aksel, 2011). A highly heterogeneous and differentoutcome of breast cancer patients, even among women with the samebiological characteristics and stage, is the greatest obstacle for the treat-ment of this disease. Identification of additional molecular markers as-sociated with more aggressive tumor behavior and a poor prognosis
nce intervals; CMF, cyclophos-NA methyltransferase; DHFR,-fluorouracil, adriamycin andfactor receptor 2; MTHFD1,hylenetetrahydrofolate reduc-ynthase reductase; OR, oddsn fragment length polymor-receptor; RFC1, reduce folate
midylate synthetase.iberian Branch of Russian Acad-ncology, 5 Kooperativny Street,fax: +7 3822 514097.a).
rights reserved.
could be instrumental for treatment choices and improve patients'survival.
The present study focuses on selected single nucleotide polymor-phisms (SNPs) in folate-related genes involved in the availability ofmethyl groups and in the synthesis and repair of DNA. Specifically,these genes include methylenetetrahydrofolate reductase (MTHFR)whose product regulates intracellular folate pool; methionine synthase(MTR) whose enzyme catalyzes the remethylation of homocysteineto methionine; methionine synthase reductase (MTRR) whose productmaintains the activity of MTR; dihydrofolate reductase (DHFR) whoseenzyme catalyzes the reduction reaction of dihydrofolate intotetrahydrofolate; methylenetetrahydrofolate dehydrogenase (MTHFD1)whose enzyme is involved in DNA biosynthesis; thymidylate synthetase(TS) whose product acts as a co-factor in folic acidmetabolism aswell asDNA synthesis; reduce folate carrier 1 (RFC1) which is an essential folatetransporter and DNA methyltransferase (DNMT3b) whose enzyme isrequired for de novo methylation. Although the various genetic poly-morphisms of these genes coding key enzymes in folate metabolismhave been a focus of many studies that have evaluated the risk of breastcancer development (Akram et al., 2012; DE Cássia Carvalho Barbosa etal., 2012; Diakite et al., 2012; Ericson et al., 2009; Hu et al., 2010; Lajinet al., 2012; Li et al., 2006; Marian et al., 2011; Mir et al., 2008;
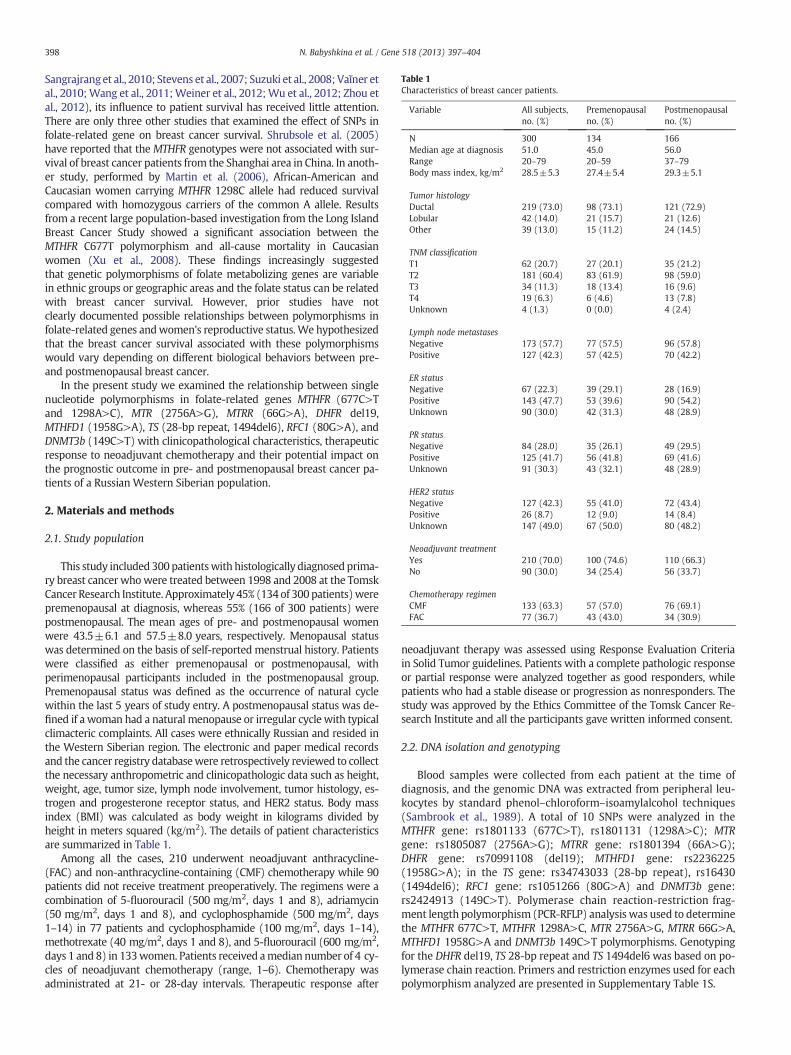
Table 1Characteristics of breast cancer patients.
Variable All subjects,no. (%)
Premenopausalno. (%)
Postmenopausalno. (%)
N 300 134 166Median age at diagnosis 51.0 45.0 56.0Range 20–79 20–59 37–79Body mass index, kg/m2 28.5±5.3 27.4±5.4 29.3±5.1
Tumor histologyDuctal 219 (73.0) 98 (73.1) 121 (72.9)Lobular 42 (14.0) 21 (15.7) 21 (12.6)Other 39 (13.0) 15 (11.2) 24 (14.5)
TNM classificationT1 62 (20.7) 27 (20.1) 35 (21.2)T2 181 (60.4) 83 (61.9) 98 (59.0)T3 34 (11.3) 18 (13.4) 16 (9.6)T4 19 (6.3) 6 (4.6) 13 (7.8)Unknown 4 (1.3) 0 (0.0) 4 (2.4)
Lymph node metastasesNegative 173 (57.7) 77 (57.5) 96 (57.8)Positive 127 (42.3) 57 (42.5) 70 (42.2)
ER statusNegative 67 (22.3) 39 (29.1) 28 (16.9)Positive 143 (47.7) 53 (39.6) 90 (54.2)Unknown 90 (30.0) 42 (31.3) 48 (28.9)
PR statusNegative 84 (28.0) 35 (26.1) 49 (29.5)Positive 125 (41.7) 56 (41.8) 69 (41.6)Unknown 91 (30.3) 43 (32.1) 48 (28.9)
HER2 statusNegative 127 (42.3) 55 (41.0) 72 (43.4)Positive 26 (8.7) 12 (9.0) 14 (8.4)Unknown 147 (49.0) 67 (50.0) 80 (48.2)
Neoadjuvant treatmentYes 210 (70.0) 100 (74.6) 110 (66.3)No 90 (30.0) 34 (25.4) 56 (33.7)
Chemotherapy regimenCMF 133 (63.3) 57 (57.0) 76 (69.1)FAC 77 (36.7) 43 (43.0) 34 (30.9)
398 N. Babyshkina et al. / Gene 518 (2013) 397–404
Sangrajranget al., 2010; Stevens et al., 2007; Suzuki et al., 2008; Vaĭner etal., 2010;Wang et al., 2011;Weiner et al., 2012;Wu et al., 2012; Zhou etal., 2012), its influence to patient survival has received little attention.There are only three other studies that examined the effect of SNPs infolate-related gene on breast cancer survival. Shrubsole et al. (2005)have reported that the MTHFR genotypes were not associated with sur-vival of breast cancer patients from the Shanghai area in China. In anoth-er study, performed by Martin et al. (2006), African-American andCaucasian women carrying MTHFR 1298C allele had reduced survivalcompared with homozygous carriers of the common A allele. Resultsfrom a recent large population-based investigation from the Long IslandBreast Cancer Study showed a significant association between theMTHFR C677T polymorphism and all-cause mortality in Caucasianwomen (Xu et al., 2008). These findings increasingly suggestedthat genetic polymorphisms of folate metabolizing genes are variablein ethnic groups or geographic areas and the folate status can be relatedwith breast cancer survival. However, prior studies have notclearly documented possible relationships between polymorphisms infolate-related genes andwomen's reproductive status.We hypothesizedthat the breast cancer survival associated with these polymorphismswould vary depending on different biological behaviors between pre-and postmenopausal breast cancer.
In the present study we examined the relationship between singlenucleotide polymorphisms in folate-related genes MTHFR (677C>Tand 1298A>C), MTR (2756A>G), MTRR (66G>A), DHFR del19,MTHFD1 (1958G>A), TS (28-bp repeat, 1494del6), RFC1 (80G>A), andDNMT3b (149C>T) with clinicopathological characteristics, therapeuticresponse to neoadjuvant chemotherapy and their potential impact onthe prognostic outcome in pre- and postmenopausal breast cancer pa-tients of a Russian Western Siberian population.
2. Materials and methods
2.1. Study population
This study included 300 patientswith histologically diagnosed prima-ry breast cancer whowere treated between 1998 and 2008 at the TomskCancer Research Institute. Approximately 45% (134 of 300 patients)werepremenopausal at diagnosis, whereas 55% (166 of 300 patients) werepostmenopausal. The mean ages of pre- and postmenopausal womenwere 43.5±6.1 and 57.5±8.0 years, respectively. Menopausal statuswas determined on the basis of self-reported menstrual history. Patientswere classified as either premenopausal or postmenopausal, withperimenopausal participants included in the postmenopausal group.Premenopausal status was defined as the occurrence of natural cyclewithin the last 5 years of study entry. A postmenopausal status was de-fined if a woman had a natural menopause or irregular cycle with typicalclimacteric complaints. All cases were ethnically Russian and resided inthe Western Siberian region. The electronic and paper medical recordsand the cancer registry databasewere retrospectively reviewed to collectthe necessary anthropometric and clinicopathologic data such as height,weight, age, tumor size, lymph node involvement, tumor histology, es-trogen and progesterone receptor status, and HER2 status. Body massindex (BMI) was calculated as body weight in kilograms divided byheight in meters squared (kg/m2). The details of patient characteristicsare summarized in Table 1.
Among all the cases, 210 underwent neoadjuvant anthracycline-(FAC) and non-anthracycline-containing (CMF) chemotherapy while 90patients did not receive treatment preoperatively. The regimens were acombination of 5-fluorouracil (500 mg/m2, days 1 and 8), adriamycin(50 mg/m2, days 1 and 8), and cyclophosphamide (500 mg/m2, days1–14) in 77 patients and cyclophosphamide (100 mg/m2, days 1–14),methotrexate (40 mg/m2, days 1 and 8), and 5-fluorouracil (600 mg/m2,days 1 and 8) in 133women. Patients received amedian number of 4 cy-cles of neoadjuvant chemotherapy (range, 1–6). Chemotherapy wasadministrated at 21- or 28-day intervals. Therapeutic response after
neoadjuvant therapy was assessed using Response Evaluation Criteriain Solid Tumor guidelines. Patients with a complete pathologic responseor partial response were analyzed together as good responders, whilepatients who had a stable disease or progression as nonresponders. Thestudy was approved by the Ethics Committee of the Tomsk Cancer Re-search Institute and all the participants gave written informed consent.
2.2. DNA isolation and genotyping
Blood samples were collected from each patient at the time ofdiagnosis, and the genomic DNA was extracted from peripheral leu-kocytes by standard phenol–chloroform–isoamylalcohol techniques(Sambrook et al., 1989). A total of 10 SNPs were analyzed in theMTHFR gene: rs1801133 (677C>T), rs1801131 (1298A>C); MTRgene: rs1805087 (2756A>G); MTRR gene: rs1801394 (66A>G);DHFR gene: rs70991108 (del19); MTHFD1 gene: rs2236225(1958G>A); in the TS gene: rs34743033 (28-bp repeat), rs16430(1494del6); RFC1 gene: rs1051266 (80G>A) and DNMT3b gene:rs2424913 (149C>T). Polymerase chain reaction-restriction frag-ment length polymorphism (PCR-RFLP) analysis was used to determinethe MTHFR 677C>T, MTHFR 1298A>C, MTR 2756A>G, MTRR 66G>A,MTHFD1 1958G>A and DNMT3b 149C>T polymorphisms. Genotypingfor the DHFR del19, TS 28-bp repeat and TS 1494del6 was based on po-lymerase chain reaction. Primers and restriction enzymes used for eachpolymorphism analyzed are presented in Supplementary Table 1S.

399N. Babyshkina et al. / Gene 518 (2013) 397–404
Each PCR amplification was performed in a 15 μl reaction volumecontaining 100 ng genomic DNA, 1× PCR buffer (1.5 mM MgCl2,60 mM Tris–HCl, 25 mM KCl; pH 8.5), 200 μM each of nucleotidesdATP, dTTP, dGTP, dCTP, 25 pM of each primer and 0.5 U Taq polymer-ase (Medigene, Novosibirsk, Russia). After initial denaturation at 94 °Сfor 5 min, the PCR reaction was carried out for 35 cycles of 94 °С for30 s, annealing for 30 s at a primer-specific temperature, 72 °С for30 s and a final cycle of 72 °С for 5 min. Amplified DNA was digestedusing the buffers and temperatures recommended by the manufac-turers. After incubation with restriction enzymes, DNA fragmentswere analyzed on 3.0% agarose gels stained with ethidium bromide.To confirm the genotyping results, selected PCR-amplifiedDNA samplesfor each polymorphism were examined by DNA sequencing.
2.3. Statistical analysis
STATISTICA version 6.1 software (1203d, StatSoft Inc., 2007) wasused for statistical analyses. Associations between genotypes of folatemetabolizing genes and clinicopathological characteristics, and re-sponse to chemotherapy were tested with chi-square or Fisher's exacttest. The odds ratios (OR) and 95% confidence intervals (CI) were alsocalculated.
Patients were followed up since the diagnosis until the end of Octo-ber, 2010. Theprimary outcomewas progression-free survival (PFS) andthe median follow-up time was 59.5 months (range, 5–201 months).PFS was calculated from the time of diagnosis until date of lastfollow-up appointment, disease progression or date of death from dis-ease. Patients who were alive at the last follow-up were censored atthat time. PFS estimation was performed according to the Kaplan–Meier method and the comparison of the groups was based on alog-rank test. Cox's proportional-hazard regression model was used toidentify independent prognostic factors. All tests were two-sided and aP-value of less than 0.05 was considered statistically significant.
3. Results
3.1. Associations of folate-related SNPs with clinicopathological factors ofbreast cancer patients
We first analyzed the relationship between each SNP and clinico-pathological factors including age, tumor size, lymphnode involvement,tumor histology, estrogen and progesterone receptor status, and HER2status in all subjects and the subjects stratified by menopausal status.The MTHFR (677C>T), MTHFR 1298A>C), MTR (2756A>G), DHFRdel19, MTHFD1 (1958G>A), TS (28-bp repeat), TS (1494del6), andRFC1 (80G>A) genotypes were successfully determined in all of the pa-tients, whereas MTRR (66G>A) and DNMT3b (149C>T) in 299 (99.7%)of cases. All ten polymorphisms were in Hardy–Weinberg equilibrium.
We found that MTHFR 677CT carriers were more frequent amongwomen with a tumor size less than 5 cm (41.2% versus 26.4%, P=0.04), especially in postmenopausal patients (48.1% versus 24.1%, P=0.03; Table 2). The association remains significant between the com-bined genotypes of this polymorphism (MTHFR 677CT+MTHFR677TT) and tumor size among all subjects (OR=0.54, P=0.04). Also,a significant difference was observed in the distribution of TS ins6/ins6 homozygous variant genotype among different pathological sub-groups of patients (20.8% in T3–T4 cases versus 8.2% in T1–T2; P=0.004). Moreover, this difference was pronounced in the postmeno-pausal subject group (P=0.006). Besides and as shown in Table 2, post-menopausal women carrying at least one variant allele for the TS(1494del6) polymorphism have a large tumor size (OR=2.78, CI:1.12–7.02, P=0.01). RFC1 (80G>A) rs1051266 but not the other SNPsexamined, showed a statistically significant difference in frequency be-tween patients with andwithout lymphnode involvement according tothemenopausal status. Thus, the RFC1 80А allelewas associatedwith anincreased risk of lymph node metastases among postmenopausal cases
in comparison to patients carrying the RFC1 80G allele (OR=2.7, CI:1.14–6.45, P=0.01; data not shown).
In all breast cancer subjects, patients with the MTHFR 677CT geno-type were significantly more likely to be ER-positive as opposed topatients with the MTHFR 677CC (P=0.03) and a similar relationshipwas observed for postmenopausal group (P=0.01). Additionally, theMTHFR 677T allele carriers were more frequent in ER-positive patients(OR=0.53, CI: 0.28–1.01, P=0.04), with a greater difference seen inpostmenopausal women (OR=0.31, CI: 0.11–0.83, P=0.009, Table 3).A significant association has been foundwith positive estrogen receptorstatus among premenopausal women carrying the TS del6/ins6 geno-type (49.1% versus 25.6% in ER-negative, P=0.02; Table 3). The associ-ation remains significant for the premenopausal women carrying thevariant TS ins6 allele (OR=0.35, CI: 0.14–0.91, P=0.02). No associationwith age, tumor histology, progesterone receptor status andHER2 statuswas observed (data not shown).
3.2. SNPs and therapeutic response to neoadjuvant chemotherapy
The breast cancer patients included in this study received eitheranthracycline-based chemotherapy (FAC) or CMF as neoadjuvant treat-ment. Among all cases, 203 patients were available for evaluation of re-sponse, including 128 (63.1%) CMF-treated and 75 (36.9%) FAC-treatedpatients. We then analyzed the role of the SNPs studied in the responseto neoadjuvant chemotherapy for these two distinct chemotherapeuticregimens together in breast cancer patients. Nine patients reached com-plete response (4.4%), ninety-three patients achieved complete re-sponse (45.8%), 96 patients (47.3%) experienced stable disease andfive patients (2.5%) had disease progression during treatment. Thus,good responders to neoadjuvant chemotherapy comprised 102 patients(50.3%), while nonresponders comprised 101 patients (49.7%).
No relationship was observed between the folate-related SNPs andresponse to neoadjuvant chemotherapy in all subjects (data notshown). The subgroup analysis by menopausal status showed a border-line association between the MTHFR (1298A>C) or DHFR del19 poly-morphisms and tumor response. Premenopausal patients with theMTHFR 1298AC genotype were more likely to have poor response thanthose carrying MTHFR 1298AA genotypes (OR=2.22, CI: 0.87–5.61,P=0.06; data not shown). In addition premenopausal group with theMTHFR 1298AC+MTHFR 1298CC variants was associated with higherdisease progression compared to the carriers of the wild-type genotypealthough the difference did not reach statistical significance (P=0.06).A similar trend was observed for the DHFR ins/del genotype whichwas associated with decreased chance of treatment response comparedwith DHFR ins/ins genotype in postmenopausal patients subgroup(OR=2.21, CI: 0.84–5.85, P=0.07). No significant association wasfound between the other polymorphisms analyzed and neoadjuvanttreatment response in premenopausal women or in postmenopausalwomen.
3.3. SNPs and breast cancer survival
We also investigated the association between the single nucleotidepolymorphisms in folate-related genes and survival of breast cancer pa-tients. At the time of the final analysis (October, 2010), 62 patientsshowed disease progression, 202 patients were alive and 36 were lostto follow-up due to migration.
In all patients, only genotypes of MTHFR (677C>T) (rs1801133)were statistically significantly associated with PFS. As shown in Fig. 1,the survival was significantly better in the patients carrying theMTHFR 677TT genotype than the survival of patients with thewild-type genotype (log rank=1.98, P=0.047).
When the cohort was divided into premenopausal and postmeno-pausal patients, the effect for PFS was found to be derived mostlyfrom the premenopausal rather than the postmenopausal patients.Premenopausal carriers of the 677TT variant of the MTHFR (677C>T)

Table 2Associations between polymorphisms in folate-related genes and clinical tumor size (T1–T2 vs.T3–T4) according to the menopausal status.
Genotypes All subjects, no. (%) Premenopausal, no. (%) Postmenopausal, no. (%)
T1–T2 T3–T4 OR (95% CI) P T1–T2 T3–T4 OR (95% CI) P T1–T2 T3–T4 OR (95% CI) P
MTHFR (677C>T)CC 124 (51.0) 35 (66.1) 1.00 (Ref.) 65 (59.1) 17 (70.8) 1.00 (Ref.) 59 (44.4) 18 (62.1) 1.00 (Ref.)CT 100 (41.2) 14 (26.4) 0.50 (0.24–1.02) 0.04 36 (32.7) 7 (29.2) 0.74 (0.25–2.14) 0.54 64 (48.1) 7 (24.1) 0.36 (0.13–0.99) 0.03ТT 19 (7.8) 4 (7.5) 0.75 (0.20–2.54) 0.79 9 (8.2) 0 (0.0) 0.00 (0.00–2.53) 0.20 10 (7.5) 4 (13.8) 1.31 (0.30–5.36) 0.73CT+TT 119 (49.0) 18 (33.9) 0.54 (0.27–1.04) 0.04 45 (40.9) 7 (29.2) 0.59 (0.20–1.69) 0.28 74 (55.6) 11 (37.9) 0.49 (0.20–1.19) 0.08
MTHFR (1298A>C)AA 107 (44.0) 24 (45.3) 1.00 (Ref.) 43 (39.0) 11 (45.8) 1.00 (Ref.) 64 (48.1) 13 (44.8) 1.00 (Ref.)AC 113 (46.5) 24 (45.3) 0.95 (0.48–1.85) 0.86 56 (51.0) 10 (41.7) 0.70 (0.25–1.97) 0.45 57 (42.9) 14 (48.3) 1.21 (0.49–3.02) 0.66CC 23 (9.2) 5 (9.4) 0.97 (0.29–3.06) 0.95 11 (10.0) 3 (12.5) 1.07 (0.20–5.25) 1.00 12 (9.0) 2 (6.9) 0.82 (0.11–4.65) 1.00AC+CC 136 (56.0) 29 (54.7) 0.95 (0.50–1.80) 0.87 67 (61.0) 13 (54.2) 0.76 (0.29–2.02) 0.54 69 (51.9) 16 (55.2) 1.14 (0.47–2.76) 0.75
MTR (2756A>G)AA 151 (62.2) 30 (56.6) 1.00 (Ref.) 66 (60.0) 13 (54.2) 1.00 (Ref.) 85 (63.9) 17 (58.6) 1.00 (Ref.)AG 79 (32.5) 21 (39.6) 1.34 (0.69–2.60) 0.36 37 (33.6) 10 (41.7) 1.37 (0.50–3.76) 0.50 42 (31.6) 11 (37.9) 1.31 (0.52–3.28) 0.53GG 13 (5.3) 2 (3.8) 0.77 (0.11–3.90) 1.00 7 (6.4) 1 (4.1) 0.73 (0.03–6.90) 1.00 6 (4.5) 1 (3.5) 0.83 (0.87–1.12) 1.00AG+GG 92 (37.8) 23 (43.4) 1.26 (0.66–2.39) 0.45 44 (40.0) 11 (45.8) 1.27 (0.48–3.36) 0.60 48 (36.1) 12 (41.4) 1.25 (0.51–3.05) 0.59
MTRR (66G>A)GG 82 (33.9) 18 (33.9) 1.00 (Ref.) 41 (37.3) 9 (37.5) 1.00 (Ref.) 41 (31.0) 9 (31.0) 1.00 (Ref.)GA 114 (47.1) 25 (47.2) 1.00 (0.49–2.06) 0.99 45 (41.0) 12 (50.0) 1.21 (0.42–3.53) 0.69 69 (52.3) 13 (44.8) 0.86 (0.31–2.41) 0.75AA 46 (19.0) 10 (18.9) 0.99 (0.39–2.50) 0.98 24 (21.8) 3 (12.5) 0.57 (0.11–2.64) 0.52 22 (16.7) 7 (24.1) 1.45 (0.41–5.04) 0.51GA+AA 160 (66.1) 35 (66.1) 1.00 (0.51–1.96) 0.99 69 (62.7) 15 (62.5) 0.99 (0.37–2.72) 0.98 91 (69.0) 20 (69.0) 1.00 (0.39–2.61) 0.99
DHFR del19ins/ins 58 (33.5) 39 (30.9) 1.00 (Ref.) 42 (38.2) 6 (25.0) 1.00 (Ref.) 40 (30.0) 9 (31.0) 1.00 (Ref.)ins/del 83 (47.9) 67 (53.2) 1.20 (0.69–2.08) 0.49 45 (40.9) 12 (50.0) 1.87 (0.58–6.21) 0.25 77 (57.9) 15 (51.7) 0.87 (0.32–2.37) 0.76del/del 32 (18.6) 20 (15.9) 1.93 (0.44–1.96) 0.84 23 (20.9) 6 (25.0) 1.83 (0.45–2.41) 0.33 16 (12.1) 5 (17.3) 1.39 (0.34–5.56) 0.75ins/del+del/del 115 (66.5) 87 (69.1) 1.13 (0.67–1.90) 0.64 68 (61.8) 18 (75.0) 1.85 (0.63–5.71) 0.22 93 (70.0) 20 (69.0) 0.96 (0.37–2.50) 0.92
MTHFD1 (1958G>A)GG 74 (30.5) 22 (41.5) 1.00 (Ref.) 31 (28.2) 11 (45.8) 1.00 (Ref.) 43 (32.3) 11 (37.9) 1.00 (Ref.)GA 124 (51.0) 22 (41.5) 0.60 (0.29–1.21) 0.12 58 (52.7) 10 (41.7) 0.49 (0.17–1.40) 0.14 66 (49.6) 12 (41.4) 0.71 (0.26–1.92) 0.46AA 45 (18.5) 9 (17.0) 0.67 (0.26–1.71) 0.36 21 (19.1) 3 (12.5) 0.40 (0.08–1.85) 0.32 24 (18.1) 6 (20.7) 0.98 (0.28–3.35) 0.97GA+AA 169 (69.5) 31 (58.5) 0.62 (0.32–1.19) 0.12 79 (71.8) 13 (54.2) 0.46 (0.17–1.25) 0.09 90 (67.7) 18 (62.1) 0.78 (0.32–1.95) 0.56
TS 28-bp repeat3R/3R 72 (29.6) 18 (33.9) 1.00 (Ref.) 32 (29.1) 8 (33.3) 1.00 (Ref.) 40 (30.0) 10 (34.5) 1.00 (Ref.)3R/2R 111 (45.7) 27 (50.9) 0.97 (0.48–2.00) 0.94 52 (47.3) 12 (50.0) 0.92 (0.31–2.80) 0.87 59 (44.4) 15 (51.7) 1.02 (0.38–2.73) 0.972R/2R 60 (24.7) 8 (15.1) 0.53 (0.20–1.41) 0.17 26 (23.6) 4 (16.7) 0.62 (0.14–2.62) 0.46 34 (25.6) 4 (13.8) 0.47 (0.11–1.85) 0.233R/2R+2R/2R 171 (70.4) 35 (66.1) 0.82 (0.42–1.62) 0.53 78 (70.9) 16 (66.7) 0.82 (0.29–2.34) 0.68 93 (70.0) 19 (65.5) 0.82 (0.32–2.09) 0.64
TS 1494 6-bp deletiondel6/del6 136 (55.9) 22 (41.5) 1.00 (Ref.) 57 (51.8) 12 (50.0) 1.00 (Ref.) 79 (59.4) 10 (34.5) 1.00 (Ref.)del6/ins6 87 (35.8) 20 (37.7) 1.42 (0.70–2.90) 0.29 43 (39.1) 8 (33.3) 0.88 (0.30–2.59) 0.80 44 (33.1) 12 (41.4) 2.15 (0.79–5.92) 0.09ins6/ins6 20 (8.2) 11 (20.8) 3.40 (1.32–8.74) 0.004 10 (9.1) 4 (16.7) 1.90 (0.42–8.72) 0.46 10 (7.5) 7 (24.1) 5.53 (1.49–20.8) 0.006del6/ins6+ins6/ins6 107 (44.1) 31 (58.5) 1.79 (0.94–3.42) 0.06 53 (48.2) 12 (50.0) 1.08 (0.41–2.84) 0.87 54 (40.6) 19 (65.5) 2.78 (1.12–7.02) 0.01
RFC1 (80G>A)GG 76 (31.2) 17 (32.1) 1.00 (Ref.) 43 (39.1) 11 (45.8) 1.00 (Ref.) 33 (24.8) 6 (20.7) 1.00 (Ref.)GA 119 (49.0) 22 (41.5) 0.83 (0.39–1.75) 0.59 47 (42.7) 8 (33.3) 0.67 (0.22–2.00) 0.42 72 (54.1) 14 (48.3) 1.07 (0.34–3.45) 0.90AA 48 (19.8) 14 (26.4) 1.30 (0.55–3.10) 0.51 20 (18.2) 5 (20.8) 0.98 (0.25–3.63) 0.97 28 (21.1) 9 (31.0) 1.77 (0.49–6.44) 0.33GA+AA 167 (68.8) 36 (67.9) 0.96 (0.49–1.92) 0.91 67 (60.9) 13 (54.2) 0.76 (0.29–2.02) 0.54 100 (75.2) 23 (79.3) 1.26 (0.44–3.81) 0.64
DNMT3b 149C>TCC 71 (29.3) 13 (24.5) 1.00 (Ref.) 30 (27.3) 2 (8.3) 1.00 (Ref.) 41 (31.1) 11 (37.9) 1.00 (Ref.)CT 126 (52.1) 28 (52.8) 1.21 (0.56–2.65) 0.60 54 (49.4) 14 (58.3) 3.89 (0.76–26.2) 0.13 72 (54.5) 14 (48.3) 0.72 (0.28–1.90) 0.47TT 45 (18.6) 12 (22.6) 0.46 (0.56–3.77) 0.39 26 (23.6) 8 (33.4) 4.62 (0.79–34.8) 0.08 19 (14.4) 4 (13.8) 0.78 (0.18–3.18) 1.00CT+TT 171 (70.7) 40 (45.5) 1.28 (0.62–2.69) 0.48 80 (72.7) 22 (91.7) 4.13 (0.86–27.1) 0.09 91 (68.9) 18 (62.1) 0.74 (0.30–1.85) 0.47
Note: values in bold indicate significant results.
400 N. Babyshkina et al. / Gene 518 (2013) 397–404
polymorphism had a significantly increased survival times thanwild-type homozygotes CC (log rank=2.21; P=0.027, Fig. 2). In post-menopausal women, this SNP was not significantly associated withprogression-free survival (log rank=1.00; P=0.317, data not shown).
In addition, the 1958AA variant of theMTHFD1 (1958G>A) SNPwasshown to be associated with a significantly increased risk of poor out-come in premenopausal patients (log rank=2.83; P=0.004, Fig. 2). Incontrast, the MTHFD1 1958G allele postmenopausal carriers, either
heterozygote (GA) or homozygote (GG), had non-significantly reducedsurvival, when compared to homozygote of the common A allele (logrank=0.93; P=0.354 and log rank=0.84; P=0.402 respectively,data not shown). No clear association was observed for the other SNPsin relation to progression-free survival.
Further multivariate Cox proportional hazard analysis adjusted forage, nodal status and tumor histology status indicated that onlyMTHFD1 (1958G>A) SNP was a statistically significant factor associated

Table 3Associations between polymorphisms in folate-related genes and ER status (negative vs. positive) according to the menopausal status.
Genotypes All subjects, no. (%) Premenopausal, no. (%) Postmenopausal, no. (%)
ER-positive ER-negative OR (95% CI) P ER-positive ER-negative OR (95% CI) P ER-positive ER-negative OR (95% CI) P
MTHFR (677C>T)CC 72 (50.3) 44 (65.7) 1.00 (Ref.) 33 (62.3) 24 (61.5) 1.00 (Ref.) 39 (43.3) 20 (71.4) 1.00 (Ref.)CT 63 (44.1) 19 (28.3) 0.49 (0.25–0.97) 0.03 18 (34.0) 12 (30.8) 0.92 (0.34–2.47) 0.85 45 (50.0) 7 (25.0) 0.30 (0.10–0.86) 0.01ТT 8 (5.6) 4 (6.0) 0.82 (0.19–3.24) 1.00 2 (3.7) 3 (7.7) 2.06 (0.25–19.4) 0.64 6 (6.7) 1 (3.6) 0.32 (0.19–3.24) 0.42CT+TT 71 (49.7) 23 (34.3) 0.53 (0.28–1.01) 0.04 20 (37.7) 15 (38.5) 1.03 (0.40–2.63) 0.94 51 (56.7) 8 (28.6) 0.31 (0.11–0.83) 0.009
MTHFR (1298A>C)AA 67 (46.9) 26 (38.8) 1.00 (Ref.) 24 (45.3) 15 (38.5) 1.00 (Ref.) 43 (47.8) 11 (39.3) 1.00 (Ref.)AC 63 (44.1) 32 (47.8) 1.31 (0.67–2.55) 0.40 23 (43.4) 18 (46.2) 1.25 (0.47–3.37) 0.62 40 (44.5) 14 (50.0) 1.37 (0.51–3.70) 0.49CC 13 (9.0) 9 (13.4) 1.78 (0.61–5.16) 0.24 6 (11.3) 6 (15.3) 1.60 (0.36–7.09) 0.52 7 (7.7) 3 (10.7) 1.68 (0.29–9.12) 0.68AC+CC 76 (53.1) 41 (61.2) 1.39 (0.74–2.62) 0.27 29 (54.7) 24 (61.5) 1.32 (0.53–3.35) 0.51 47 (52.2) 17 (60.7) 1.41 (0.55–3.67) 0.43
MTR (2756A>G)AA 88 (61.5) 44 (65.7) 1.00 (Ref.) 29 (54.7) 25 (64.1) 1.00 (Ref.) 59 (65.6) 19 (67.9) 1.00 (Ref.)AG 47 (32.9) 20 (29.9) 0.85 (0.43–1.68) 0.62 20 (37.7) 11 (28.2) 0.64 (0,23–1.74) 0.33 27 (30.0) 9 (37.7) 1.04 (0.38–2.82) 0.94GG 8 (5.6) 3 (4.4) 0.75 (0.15–3.33) 1.00 4 (7.6) 3 (7.7) 0.87 (0.14–5.26) 1.00 4 (4.4) 0 (0.0) 0.00 (0.00–5.31) 0.57AG+GG 55 (38.5) 23 (34.3) 0.84 (0.44–1.60) 0.56 24 (45.3) 14 (35.9) 0.68 (0.26–1.72) 0.37 31 (34.4) 9 (32.1) 0.90 (0.33–2.42) 0.82
MTRR (66G>A)GG 53 (37.3) 21 (31.3) 1.00 (Ref.) 24 (45.3) 15 (38.5) 1.00 (Ref.) 29 (32.6) 6 (21.4) 1.00 (Ref.)GA 58 (40.9) 39 (58.2) 1.70 (0.85–3.42) 0.11 17 (32.1) 22 (56.4) 2.07 (0.76–5.67) 0.11 41 (46.1) 17 (60.7) 2.00 (0.64–6.52) 0.19AA 31 (21.8) 7 (10.5) 0.57 (0.19–1.63) 0.25 12 (22.6) 2 (5.1) 0.27 (0.04–1.56) 0.18 19 (21.3) 5 (17.9) 1.27 (0.28–5.67) 0.74GA+AA 89 (62.7) 46 (68.7) 1.30 (0.67–2.54) 0.40 29 (54.7) 24 (61.5) 1.32 (0.53–3.35) 0.51 60 (67.4) 22 (78.6) 1.77 (0.59–5.50) 0.26
DHFR del19ins/ins 44 (30.8) 22 (32.8) 1.00 (Ref.) 21 (39.6) 12 (30.8) 1.00 (Ref.) 23 (25.6) 10 (35.7) 1.00 (Ref.)ins/del 73 (51.1) 31 (46.3) 0.85 (0.42–1.74) 0.63 22 (41.5) 17 (43.6) 1.35 (0.47–3.91) 0.53 51 (56.7) 14 (50.0) 0.63 (0.22–1.81) 0.34del/del 26 (18.1) 14 (20.9) 1.08 (0.43–2.67) 0.86 10 (18.9) 10 (25.6) 1.75 (0.49–6.31) 0.33 16 (17.7) 4 (14.3) 0.57 (0.12–2.52) 0.41ins/del+del/del
99 (69.2) 45 (67.2) 0.91 (0.47–1.77) 0.76 32 (60.4) 27 (69.2) 1.48 (0.56–3.89) 0.38 67 (74.4) 18 (64.3) 0.62 (0.23–1.68) 0.30
MTHFD1 (1958G>A)GG 49 (34.3) 24 (35.8) 1.00 (Ref.) 18 (34.0) 14 (35.9) 1.00 (Ref.) 31 (34.4) 10 (35.7) 1.00 (Ref.)GA 74 (51.8) 31 (46.3) 0.86 (0.43–1.71) 0.63 25 (47.2) 19 (48.7) 0.98 (0.35–2.71) 0.96 49 (54.4) 12 (42.9) 076 (0.27–2.18) 0.57AA 20 (14.0) 12 (17.9) 1.23 (0.47–3.17) 0.65 10 (18.8) 6 (15.4) 0.77 (0.19–3.10) 0.68 10 (11.2) 6 (21.4) 1.86 (0.46–7.60) 0.34GA+AA 94 (65.7) 43 (64.2) 0.93 (0.49–1.79) 0.83 35 (66.0) 25 (64.1) 0.92 (0.35–2.39) 0.85 59 (65.6) 18 (64.3) 0.95 (0.36–2.52) 0.90
TS 28-bp repeat3R/3R 45 (31.5) 19 (28.4) 1.00 (Ref.) 17 (32.1) 8 (20.5) 1.00 (Ref.) 28 (31.1) 11 (39.3) 1.00 (Ref.)3R/2R 64 (44.8) 33 (49.3) 1.22 (0.59–2.56) 0.57 24 (45.3) 19 (48.7) 1.68 (0.53–5.39) 0.32 40 (44.4) 14 (50.0) 0.89 (0.32–2.48) 0.812R/2R 34 (23.7) 15 (22.3) 1.04 (0.43–2.54) 0.92 12 (22.6) 12 (30.8) 2.13 (0.57–8.03) 0.20 22 (24.5) 3 (10.7) 0.35 (0.07–1.60) 0.133R/2R+2R/2R 98 (68.5) 48 (71.6) 1.16 (0.59–2.31) 0.65 36 (67.9) 31 (79.5) 1.83 (0.63–5.40) 0.22 62 (68.9) 17 (60.7) 0.70 (0.27–1.84) 0.42
TS 1494 6-bp deletiondel6/del6 70 (48.9) 39 (58.2) 1.00 (Ref.) 22 (41.5) 26 (66.7) 1.00 (Ref.) 48 (53.3) 13 (46.4) 1.00 (Ref.)del6/ins6 59 (41.3) 21 (31.3) 0.64 (0.32–1.26) 0.16 26 (49.1) 10 (25.6) 0.33 (0.12–0.90) 0.02 33 (36.7) 11 (39.3) 1.23 (0.45–3.38) 0.66ins6/ins6 14 (9.8) 7 (10.5) 0.90 (0.30–2.64) 0.83 5 (9.4) 3 (7.7) 0.51 (0.08–2.86) 0.46 9 (10.0) 4 (14.3) 1.64 (0.35–7.26) 0.48del6/ins6+ins6/ins6
73 (51.1) 28 (41.8) 0.69 (0.37–1.29) 0.21 31 (58.5) 13 (33.3) 0.35 (0.14–0.91) 0.02 42 (46.7) 15 (53.6) 1.32 (0.59–2.31) 0.52
RFC1 (80G>A)GG 40 (27.9) 26 (38.8) 1.00 (Ref.) 20 (37.7) 19 (48.7) 1.00 (Ref.) 20 (22.2) 7 (25.0) 1.00 (Ref.)GA 67 (46.9) 27 (40.3) 0.62 (0.30–1.27) 0.16 18 (34.0) 15 (38.5) 0.88 (0.31–2.46) 0.78 49 (54.4) 12 (42.9) 0.70 (0.21–2.32) 0.51AA 36 (25.2) 14 (20.9) 0.60 (0.25–1.41) 0.20 15 (28.3) 5 (12.8) 0.35 (0.09–1.32) 0.08 21 (23.4) 9 (32.1) 1.22 (0.33–4.57) 0.73GA+AA 103 (72.1) 41 (61.2) 0.61 (0.32–1.18) 0.11 33 (62.3) 20 (51.3) 0.64 (0.25–1.60) 0.29 70 (77.8) 21 (75.0) 0.86 (0.29–2.59) 0.76
DNMT3b 149C>TCC 45 (31.5) 15 (22.4) 1.00 (Ref.) 13 (24.5) 8 (20.5) 1.00 (Ref.) 32 (35.6) 7 (25.0) 1.00 (Ref.)CT 71 (49.7) 37 (55.2) 1.56 (0.73–3.37) 0.21 28 (52.8) 19 (48.7) 1.10 (0.34–3.62) 0.86 43 (47.8) 18 (64.3) 1.91 (0.65–5.78) 0.19TT 27 (18.8) 15 (22.4) 1.67 (0.65–4.30) 0.24 12 (22.7) 12 (30.8) 1.63 (0.42–6.36) 0.42 15 (16.6) 3 (10.7) 0.91 (0.16–4.80) 1.00CT+TT 98 (68.5) 52 (77.6) 1.59 (0.32–1.18) 0.17 40 (75.5) 31 (79.5) 1.26 (0.32–1.18) 0.65 58 (64.4) 21 (75.0) 1.66 (0.58–4.83) 0.30
Note: values in bold indicate significant results.
401N. Babyshkina et al. / Gene 518 (2013) 397–404
with RFS in premenopausal patients (Table 4). NeitherMTHFR (677C>T)norMTHFD1 (1958G>A) polymorphism status was associated with RFSin postmenopausal group (Table 4).
4. Discussion
Breast cancer is frequently a hormonally dependent cancer andmany findings support arguments from the literature showing that
differentmechanismsmay be involved in breast carcinogenesis process-es between postmenopausal and premenopausalwomen. In this regard,we analyzed the effect of 10 gene polymorphisms involving 8 relevantcandidate genes related to the folate metabolism, namely MTHFR, MTR,MTRR, DHFR,MTHFD1, TS, RFC1 and DNMT3b, on the clinicopathologicalfeatures, response to neoadjuvant treatment and survival of Russian pa-tients of the Western Siberian region according their reproductivestatus.

Fig. 1. Kaplan–Meier survival curves for all patients group in relation to MTHFR (677C>T)polymorphism. Survival in the MTHFR 677TT group was better compared to that in the CCgroup (log rank=1.98, P=0.047).
Fig. 2. Kaplan–Meier survival curves for premenopausal patients group in relation toMTHFR(677C>T) (A) and MTHFD1 (1958G>A) polymorphism (B). (A) Premenopausal patientswith the MTHFR 677TT genotype had a longer PFS time than patients' homozygote withthe wild-type CC genotype (log rank=2.21; P=0.027). (B) Premenopausal carriers of theMTHFD1 1958AA variant had shorter survival times than wild-type homozygotes GG (logrank=2.83; P=0.004).
Table 4Multivariate Cox proportional hazard analysis of the MTHFR (677C>T) and MTHFD1(1958G>A) SNPs.
Factor Premenopausal Postmenopausal
Progression-free survival
Hazardratioa
95% CI Pvalue
Hazardratioa
95% CI Pvalue
MTHFR 677C>T(TT vs CC)
0.04 0.04–0.29 0.087 0.58 0.49–1.10 0.455
MTHFD1 1958G>A(AA vs GG)
4.39 2.19–10.7 0.003 0.70 0.44–0.94 0.473
a Adjusted for age, nodal status and tumor histology status.
402 N. Babyshkina et al. / Gene 518 (2013) 397–404
Several studies have considered the relationship between folatepolymorphism and clinical/pathological characteristics in patientswith breast cancer (Grieu et al., 2004; Henríquez-Hernández et al.,2010; Li et al., 2006; Naushad et al., 2012). However, the correlationswere not significant and the results have been observed only in theoverall patient cohort without stratification according to menopausalstatus. Our data suggested that the association between the MTHFRSNP (rs1801133), RFC1 SNP (rs1051266) and clinicopathological datawas mainly evident in postmenopausal women, while the TS SNP(1494del6) was significant in both pre- and postmenopausal cases. Inpostmenopausal group, we found that theMTHFR 677CT genotype wasrelated to tumor size and ER-positive status. In addition, the frequencyof the combination of MTHFR 677CT and 677TT genotype was signifi-cantly higher in patients with ER-positive tumor. For the SNPrs1051266 in the RFC1 gene, the carriers of the 80А allele had a moreoften occurrence of lymph node metastasis. In our previous study, wehave reported that the frequency ofMTHFR 677CT genotype was higherin postmenopausal patients compared with premenopausal patientswith the same genotype although the trend P did not reach statisticalsignificance (Perel'muter et al., 2008). It is known that the T allele ofMTHFR (677C>T) polymorphism has been associated with decreasedenzyme activity of MTHFR and increased plasma levels of homocysteinewhichmay affect hypo- and hypermethylation of DNA (Ma et al., 1997).Interestingly, Chango et al. (2000) andDevlin et al. (2006) found a geno-type–genotype interaction in which healthy subjects with the combina-tion ofMTHFR 677TT and RFC1 80GG genotypes had significantly higherserum homocysteine concentrations compared with otherMTHFR/RFC1genotype combinations. In keeping with this, our data suggest that thevariation of specific genotype of MTHFR (677C>T) and RFC1 (80G>A)polymorphismsmay predispose both to amore or less tumor aggressivebehavior among postmenopausal women.
Apart from possible involvement as tumor aggressiveness factors forbreast cancer, folate polymorphisms might also be associated with theefficacy of treatments by influencing the activity of DNA synthesis andDNA repair process. We did not observe any significant difference in ge-notype frequencies among analyzed SNP types stratified by response toneoadjuvant chemotherapy according to reproductive patient's status.However, premenopausal patients with MTHFR 1298C allele tended tohave a poorer response to neoadjuvant chemotherapy than patientswith the other polymorphisms. Similar results were obtained for theDHFR ins/del genotype carrier but in postmenopausal women group.Thus, it can be hypothesized that the efficacy of breast cancer treatmentmight be modulated by specific effect of menopausal status and geneticvariations in folate enzymes. The association between the folate poly-morphism status and response to breast cancer treatment has beenreported by several authors (Henríquez-Hernández et al., 2010; Huanget al., 2008; Paré et al., 2007; Toffoli et al., 2000; Xu et al., 2008; Xu etal., 2009; Zárate et al., 2007). Most of these investigations involve poly-morphisms of MTHFR, DHFR, and TS and some of these studies aim atthe prediction of the efficacy of adjuvant treatment of breast cancer butnot of neoadjuvant treatment (Huang et al., 2008; Paré et al., 2007;Toffoli et al., 2000; Xu et al., 2009; Zárate et al., 2007). However,

403N. Babyshkina et al. / Gene 518 (2013) 397–404
Henríquez-Hernández et al. (2010) found that the response to5-fluorouracil and cyclophosphamide-based neoadjuvant chemotherapywas not significantly different among theMTHFR (677C>T) and TS 28-bprepeat SNPs butMTHFR allele 677T was strongly associated with adversereactions. In another study, there was a trend for patients homozygousfor 3 copies of the TS (28-bp repeat) polymorphismwhichwere overrep-resented in the progression disease response group compared to thecombined partial and stable disease groups of patients treatedneoadjuvantly with fluorouracil-based chemotherapy (Nordgard et al.,2008). In a recent report from India, the response to 5-fluorouracil waspoor in patients with 2R/3R and 2R/2R genotypes than in patients withthe triple-repeat homozygous; however, this study had small samplesizes (Kumar et al., 2010). The treatment protocol underlying the presentstudy consists of anthracyclines and nonanthracycline-containing regi-mens and the lack of difference in our results is likely related to differentregimens and different cycles of chemotherapy. It is possible that, withhomogeneous protocol of treatment and adequate numbers of patients,particularly in treatment subgroups, some differences would be evident.
It is plausible that the genetic variants of the genes coding key en-zymes in folate metabolism may influence their function, resulting inaberrant methylation or DNA synthesis inhibition and may also be im-plicated in the progression of tumors and consequently in the prognos-tic outcome.When evaluating the overall patients group, the associationwas observed only between MTHFR (677C>T) polymorphism andprogression-free survival, which is in accordance with the resultsrecently reported by Martin et al. (2006) and Xu et al. (2008). Whenwe conducted a subgroup analysis according to menopausal status, ahighly significant association with PFS was observed in premenopausalwomen, but not in postmenopausal women. We found that breast can-cer patientswith homozygous expression of theMTHFR 677TT genotypehad significant better progression-free survival than the677CChomozy-gous patients. In addition, a worse outcome was observed in patientswith MTHFD1 1958AA genotype compared with wild-type homozy-gotes GG for RFS. Multivariate analysis in our study indicated that theMTHFD1 (1958G>A) SNPmight be an independent predictor for surviv-al of premenopausal breast cancer patients. Interestingly, Martin et al.(2006)12 showed an opposite effect of MTHFR (677C>T) and theMTHFR (1298A>C) on breast cancer survival although both variant al-leles were found to decrease MTHFR enzyme activity. Our datasuggested an inverse association between the MTHFR (677C>T) andother evaluatedMTHFD1 (1958G>A) polymorphism on premenopausalbreast cancer survival.
It has been proposed that the effect of genetic variation in folate-related genes on breast cancer development may depend on the levelsof folate intake and estrogen status. Recent investigations have shownthat on theonehand, if dietary folate levels are low,MTHFR677TT variantcan induce a DNA hypomethylation and consequent DNA damage andmutations resulting in increased relapse and unfavorable outcome. Onthe other hand, in the presence of high folate levels the same genotypeleads to a great pool ofmethylenetetrahydrofolate available for DNA syn-thesis, and therefore prevents cancer (Friso et al., 2002; Levine et al.,2000; Ulrich et al., 1999). The poor survival conferred by the MTHFD11958AA genotype could be due to the result of DNA synthesis changesand consequent lack of cell control (Li et al., 2006). It may be hypothe-sized that the different mechanisms of action of estrogen with folatepolymorphism interaction could at least partly explain the difference ofsurvival data observed in premenopausal women compared with post-menopausal women. In particular, the prevalence of estrogen genotoxiccomponent pathway plays an important role in premenopausal breastcancer risk, as well as in progression that may modulate the outcomeof patients.
This study has several limitations. We could not consider extensiveinformation on life-style factors including tobacco smoking, alcoholdrinking, and dietary habits. However, the level of folate intake fromfoods is relatively low in Western Siberian region. Thus, dietary habitscould not have a significant influence on the biological behavior of
breast cancer. The lack of information on the blood folate concentra-tions and the plasma homocysteine levels also limits the interpretationof the results. Further studies with larger sample sizes should incorpo-rate these folate-related genes in order to understand the effect of spe-cific neoadjuvant treatment on the survival by these polymorphisms. Acombined genotype analysis of these SNPs could contribute to the selec-tion of premenopausal and postmenopausal patients who would bene-fit from anthracyclines and nonanthracycline-containing regimens oftreatment.
In conclusion, our study provides information about the role of vari-ous genetic polymorphisms of folate-related genes in outcome amongpre- and postmenopausal breast cancer patients and is the first reporton the studies of these SNPs in a homogeneous ethnic population inthe Western Siberian region of Russia. We found that the MTHFR(677C>T) and theMTHFD1 (1958G>A) polymorphisms were inverselyassociated with progression-free survival only among premenopausalbreast cancer patients and the MTHFD1 1958AA genotype may be aprognostic factor for PFS in the same patients. Further investigationwould be needed to understand the biological mechanisms throughwhich the folate-related SNP use impacts the progression ofpremenopausal breast cancer.
Conflict of interest
None declared.
Acknowledgments
This workwas supported by grants from the Siberian Branch of Rus-sian Academy of Medical Sciences. We thank Dr. Vladimir Perel'muterand Dr.Marina Zav'ialova from the Pathology and Cytology Departmentfor providing the clinicopathologic data. We are also grateful toViktoriya Stalbovskaya for her advice on the statistical analysis.
Appendix A. Supplementary data
Supplementary data to this article can be found online at http://dx.doi.org/10.1016/j.gene.2012.12.095.
References
Akram, M., Malik, F.A., Kayani, M.A., 2012. Mutational analysis of the MTHFR gene inbreast cancer patients of Pakistani population. Asian Pac. J. Cancer Prev. 13,1599–1603.
Chango, A., et al., 2000. A polymorphism (80G→A) in the reduced folate carrier geneand its associations with folate status and homocysteinemia. Mol. Genet. Metab.70, 310–315.
Davydov,M.I., Aksel, E.M., 2011. Cancer statistics in Russia and CIS in 2009. J. N. N. BlokhinRuss. Cancer Res. Cent. RAMS 22, 9–123.
DE Cássia Carvalho Barbosa, R., DA Costa, D.M., Cordeiro, D.E., Vieira, A.P., Rabenhorst,S.H., 2012. Interaction of MTHFR C677T and A1298C, and MTR A2756G Gene Poly-morphisms in Breast Cancer Risk in a Population in Northeast Brazil. AnticancerRes. 32, 4805–4811.
Devlin, A.M., Clarke, R., Birks, J., Evans, J.G., Halsted, C.H., 2006. Interactions amongpolymorphisms in folate-metabolizing genes and serum total homocysteine con-centrations in a healthy elderly population. Am. J. Clin. Nutr. 83, 708–713.
Diakite, B., Tazzite, A., Hamzi, K., Jouhadi, H., Nadifi, S., 2012. MethylenetetrahydrofolateReductase C677T polymorphism and breast cancer risk in Moroccan women. Afr.Health Sci. 12, 204–209.
Ericson, U.C., et al., 2009. Increased breast cancer risk at high plasma folate concentrationsamong women with the MTHFR 677T allele. Am. J. Clin. Nutr. 90, 1380–1389.
Friso, S., et al., 2002. A common mutation in the 5,10-methylenetetrahydrofolate re-ductase gene affects genomic DNA methylation through an interaction with folatestatus. Proc. Natl. Acad. Sci. U. S. A. 99, 5606–5611.
Grieu, F., Powell, B., Beilby, J., Iacopetta, B., 2004. Methylenetetrahydrofolate reductaseand thymidylate synthase polymorphisms are not associated with breast cancerrisk or phenotype. Anticancer. Res. 24, 3215–3219.
Henríquez-Hernández, L.A., Murias-Rosales, A., González-Hernández, A., de León, A.C.,Díaz-Chico, N., Fernández-Pérez, L., 2010. Distribution of TYMS, MTHFR, p53 andMDR1 gene polymorphisms in patients with breast cancer treated with neoadjuvantchemotherapy. Cancer Epidemiol. 34, 634–638.
Hu, J., Zhou, G.W., Wang, N., Wang, Y.J., 2010. MTRR A66G polymorphism and breastcancer risk: a meta-analysis. Breast Cancer Res. Treat. 124, 779–784.

404 N. Babyshkina et al. / Gene 518 (2013) 397–404
Huang, M.Y., et al., 2008. Multiple genetic polymorphisms of GSTP1 313AG, MDR13435CC, and MTHFR 677CC highly correlated with early relapse of breast cancerpatients in Taiwan. Ann. Surg. Oncol. 15, 872–880.
Kumar, K., Vamsy,M., Jamil, K., 2010. Thymidylate synthase gene polymorphisms effecting5-FU response in breast cancer patients. Cancer Biomark. 6, 83–93.
Lajin, B., Alhaj Sakur, A., Ghabreau, L., Alachkar, A., 2012. Association of polymorphismsin one-carbon metabolizing genes with breast cancer risk in Syrian women. Tu-mour Biol. 33, 1133–1139.
Levine, A.J., et al., 2000. Themethylenetetrahydrofolate reductase 677C→T polymorphismand distal colorectal adenoma risk. Cancer Epidemiol. Biomarkers Prev. 9, 657–663.
Li, S.Y., Rong, M., Iacopetta, B., 2006. Germ-line variants in methyl-group metabolismgenes and susceptibility to DNA methylation in human breast cancer. Oncol. Rep.15, 221–225.
Ma, J., et al., 1997.Methylenetetrahydrofolate reductase polymorphism, dietary interactions,and risk of colorectal cancer. Cancer Res. 57, 1098–1102.
Marian, C., Tao, M., Mason, J.B., Goerlitz, D.S., Nie, J., Chanson, A., Freudenheim, J.L.,Shields, P.G., 2011. Single nucleotide polymorphisms in uracil-processing genes,intake of one-carbon nutrients and breast cancer risk. Eur. J. Clin. Nutr. 65,683–689.
Martin, D.N., et al., 2006. Association of MTHFR gene polymorphisms with breast cancersurvival. BMC Cancer 6, 257.
Mir, M.M., et al., 2008. Combined impact of polymorphism of folate metabolism genes;glutamate carboxypeptidase, methylene tetrahydrofolate reductase and methioninesynthase reductase on breast cancer susceptibility in Kashmiri women. Int. J. HealthSci. (Qassim) 2, 3–14.
Naushad, S.M., Pavani, A., Rupasree, Y., Divyya, S., Deepti, S., Digumarti, R.R.,Gottumukkala, S.R., Prayaga, A., Kutala, V.K., 2012. Association of aberrations inone-carbon metabolism with molecular phenotype and grade of breast cancer.Mol Carcinog. 51, E32–E41.
Nordgard, S.H., et al., 2008. Pathway based analysis of SNPs with relevance to 5-FU therapy:relation to intratumoral mRNA expression and survival. Int. J. Cancer 123, 577–585.
Paré, L., et al., 2007. Influence of thymidylate synthase and methylenetetrahydrofolate re-ductase gene polymorphisms on the disease-free survival of breast cancer patients re-ceiving adjuvant 5-fluorouracil/methotrexate-based therapy. Anticancer Drugs 17,821–825.
Perel'muter, V.M., et al., 2008. Genetic and clinical and pathological characteristics ofbreast cancer in premenopausal and postmenopausal women. Adv. Gerontol. 21,643–653.
Sambrook, J., Fritsch, E.F., Maniatis, T., 1989. Molecular Cloning: A Laboratory Manual,second ed. Cold Spring Harbor Laboratory Press, New York.
Sangrajrang, S., Sato, Y., Sakamoto, H., Ohnami, S., Khuhaprema, T., Yoshida, T., 2010. Geneticpolymorphisms in folate and alcohol metabolism and breast cancer risk: a case–controlstudy in Thai women. Breast Cancer Res. Treat. 123, 885–893.
Shrubsole, M.J., et al., 2005. MTHFR genotypes and breast cancer survival after surgeryand chemotherapy: a report from the Shanghai Breast Cancer Study. Breast CancerRes. Treat. 91, 73–79.
Stevens, V.L., et al., 2007. Association of polymorphisms in one-carbon metabolismgenes and postmenopausal breast cancer incidence. Cancer Epidemiol. BiomarkersPrev. 16, 1140–1147.
Suzuki, T., et al., 2008. One-carbon metabolism-related gene polymorphisms and riskof breast cancer. Carcinogenesis 29, 356–362.
Toffoli, G., Veronesi, A., Boiocchi, M., Crivellari, D., 2000. MTHFR gene polymorphism and se-vere toxicity during adjuvant treatment of early breast cancer with cyclophosphamide,methotrexate, and fluorouracil (CMF). Ann. Oncol. 11, 373–374.
Ulrich, C.M., et al., 1999. Colorectal adenomas and the C677T MTHFR polymorphism:evidence for gene–environment interaction? Cancer Epidemiol. Biomarkers Prev.8, 659–668.
Vaĭner, A.S., et al., 2010. Polymorphic variants of folate metabolizing genes (C677T andA1298C MTHFR, C1420T SHMT1 and G1958A MTHFD) are not associated with therisk of breast cancer in West Siberian Region of Russia. Mol. Biol. 44, 816–823.
Wang, J., Wang, B., Bi, J., Di, J., 2011. The association between two polymorphisms in theTYMS gene and breast cancer risk: a meta-analysis. Breast Cancer Res. Treat. 128,203–209.
Weiner, A.S., Boyarskikh, U.A., Voronina, E.N., Selezneva, I.A., Sinkina, T.V., Lazarev, A.F.,Petrova, V.D., Filipenko, M.L., 2012. Polymorphisms in the folate-metabolizinggenes MTR, MTRR, and CBS and breast cancer risk. Cancer Epidemiol. 36, 95–100.
Wu, X.Y., Ni, J., Xu, W.J., Zhou, T., Wang, X., 2012. Interactions between MTHFR C677T–A1298C variants and folic acid deficiency affect breast cancer risk in a Chinese pop-ulation. Asian Pac. J. Cancer Prev. 13, 2199–2206.
Xu, X., Chen, J., 2009. One-carbon metabolism and breast cancer: an epidemiologicalperspective. J. Genet. Genomics 36, 203–214.
Xu, X., et al., 2008. B-vitamin intake, one-carbonmetabolism, and survival in a population-based study of women with breast cancer. Cancer Epidemiol. Biomarkers Prev. 17,2109–2116.
Zárate, R., et al., 2007. GSTP1 and MTHFR polymorphisms are related with toxicityin breast cancer adjuvant anthracycline-based treatment. Curr. Drug Metab. 8,481–486.
Zhou, J.Y., Shi, R., Yu, H.L., Zeng, Y., Zheng, W.L., Ma, W.L., 2012. The association be-tween two polymorphisms in the TS gene and risk of cancer: a systematic reviewand pooled analysis. Int. J. Cancer 131, 2103–2016.