the effect of dietary fibre on feed intake and growth in beagle puppies
TRANSCRIPT

The Effect of Dietary Fibre on Feed Intake andGrowth in Beagle Puppies
C.B. Delorme, D. Barrette, R. Mongeau and N. Lariviere*
ABSTRACT
We studied the growth, feed intake,feed efficiency and protein efficiencyratio of groups of female Beagle pup-pies fed 16% or 22% crude proteinrations to which 6% or 12% wheatbean was added at the expense of thetotal diet. The filnal neutral detergentfibre concentrations were 12%, 16%,22% and 23% (dry matter basis). Theaddition of wheat bran to puppyrations, bringing the neutral detergentfibre up to 16% in a 21% crude proteindiet had no deleterious effects on feedintake, feed and protein efficiency orgrowth in Beagle puppies. Over a suf-ficiently long period of time, thegrowth of this group did not differfrom that of the controls (12% neutraldetergent fibre, 23% crude protein)although it was higher at intermediatetimes. The effects of the high filbre (22or 23% neutral detergent fibre) dietson growth, feed intake, feed efficiencyand protein efficiency ratio are con-sistent with an energy deficit resultingfrom the animals' inability to adaptfully to the dilution of their rationsleading to lower growth, less efficientuse of feed and, in the case of group 3(22% crude protein, 22% neutraldetergent fibre), a lower protein effi-ciency ratio. The protein efficiencyratio of group 4 (16% crude protein,23% neutral detergent fibre) washigher than that ofgroup 3, most likelythe result of a more limiting amount ofdietary protein leading to a more effi-cient use for growth by the animal. Wehave concluded that intermediate lev-els of neutral detergent fibre (up to16%) were not deleterious even inpuppy rations. However, at least in the
presence of low levels of dietary pro-tein, there is a limit to the amount ofbran which could be added to puppyrations. This limit was surpassed withthe 22% crude protein, 22% neutraldetergent fibre diet and such levelsshould be avoided.
Key words: Diet dilution, dietaryfibre, puppy, feed intake, growth.
RESUME
Cette experience visait a determinerla vitesse de croissance, les taux deconsommation et d'efficacite alimen-taires, ainsi que le coefficient d'effi-cacite proteique, chez des groupes dejeunes chiennes Beagle qui recevaientdes rations contenant 16% ou 22% deproteine brute, auxquelles on avaitajoute 6% ou 12% de son de ble. Lesquatre rations utilisees contenaientrespectivement 12%, 16%, 22% et 23%de fibre, selon la methode d'extractionavec un detergent neutre, sur une basede matiere seche. L'ajout de son a laration du groupe #2 qui contenait 21%de proteine brute amena son taux defibre 'a 16%; une telle modificationn'exerVa aucune influence nocive surl'un ou l'autre des quatre parametresexperimentaux. Sur un intervalle suf-fisamment long, la vitesse de crois-sance des sujets de ce groupe se revelaequivalente a celle des temoins dont laration contenait 23% de proteine bruteet 12% de filbre; elle s'avera cependantplus rapide, sur un intervalle plus bref.Les effets des rations qui contenaientautant que 22% ou 23% de fibre, sur lesquatre parametres precites, seraientcompatibles avec un deficit energe-
tique attribuable a l'incapacite deschiennes experimentales de s'adapter 'ala dilution de leur ration avec du son. Ilen resulta une chute de croissance, uneutilisation moins efficace des alimentset, pour celles du groupe #3 qui rece-vaient 22% de proteine brute et 22% defibre, une baisse du coefficient d'effi-cacite proteique. Ce parametre affichacependant une hausse chez celles dugroupe #4 dont la ration contenait 16%de proteine brute et 23% de fibre,indice probable d'un deficit proteiqueplus marque qui avait amene une utili-sation plus efficace des proteines pourla croissance. Les resultats de l'ex-perience permettent de conclure queles rations destinees aux jeunes chienspeuvent contenir, sans inconvenient,jusqu'a 16% de fibre. II existe cepen-dant, du moins en presence d'un faibletaux de proteines alimentaires, unelimite a l'addition de son aux aliments.Cette limite se trouva depassee avec laration du groupe #3 qui contenait 22%de proteine brute et 22% de fibre; ilfaut par consequent eviter un tel exces.
Mots cles: dilution alimentaire, fibrealimentaire, chiot, consommationalimentaire, croissance.
INTRODUCTION
Before recently renewed public andscientific interest (1-3), dietary fibrewas considered to be of limited interestin monogastric nutrition. Much of thework published to date bears on theassociation of low-fibre diets with var-ious health problems, based on epide-miological data or clinical studies (1).There has been less attention focussed
*Departement d'Anatomie et Physiologie animales, Faculte de Medecine veterinaire, Universite de Montreal, C.P. 5000, Saint-Hyacinthe, QuebecJ2S 7C6 (Delorme, Barrette, Lariviere) and Banting Research Centre, Food Directorate, Division of Nutrition Research, Ottawa, Ontario K IA 0L2(Mongeau).The present work was supported by a grant from the Canadian Veterinary Research Trust Fund.
Submitted October 24, 1983.
Can J Comp Med 1985; 49: 278-285.278
on the inverse question of high-fibrediets and its implications for healthand in the utilization of essential nut-rients (2, 3).
Previous work in this laboratory,using rats as an experimental animal,revealed an interaction between die-tary cellulose and protein in the adap-tation to caloric dilution, in proteinutilization and in the growth of theexperimental animals (4, 5). Plantfood sources are used in formulatingcommercial canine rations and high-fibre diets are utilized in the treatmentof constipation and obesity in dogs.Despite this practice, comparativelylittle is known about the effects of die-tary fibre in this species or the limitsone should place on its concentrationin the diet. Since the dog is not adaptedto the consumption of dietary fibre inthe same way as the rat, it appeared tous to be of practical interest to verifythe degree to which this species cantolerate fibre in its rations. The pur-pose of this study was to examine theeffects of bran on the growth, feedintake and feed efficiency of growingdogs. We chose to use puppies in thisstudy since any deleterious effect ofdietary fibre would be likely to mani-fest itself in a situation of high need,such as growth.
500-
MATERIALS AND METHODS
Female Beagle puppies werereceived in groups of six. At time ofarrival they were seven weeks old.Their physical status was satisfactoryand they weighed 2.31 ± 0.06 kg. Thetrial was preceded by a three weekperiod for adaptation to their newenvironment and for observation, dur-ing which they were fed Purina puppychow and given tap water ad libitum.Because of health problems whichdeveloped during this period, twoanimals had to be eliminated fromgroup 1 and one each from groups 2and 4.
Following adaptation, the animalswere housed by group in runs, fed theexperimental diets and given tap waterad libitum for the duration of theexperiment (up to 18 weeks). At thistime, groups 1, 2, 3 and 4 weighedrespectively 4.38 ± 0.26 kg, 4.29 ± 0.17kg, 4.27 ± 0.23 kg and 4.61 ± 0.19 kg(means ± SEM). There were no signif-
icant differences between groups(P > 0.05). Feed intake was recordeddaily and body weights were measuredweekly. The control group (group 1)was fed powdered Purina dog chow(20% crude protein, 7% crude fibre).This protein concentration, which islow for puppies, was chosen so thatany effects of bran on protein utiliza-tion would be likely to modify parame-ters such as growth, feed efficiency andprotein efficiency. In the formulationof rations #2 and #3, powdered Purinadog chow was diluted with 6% and12% wheat bran, respectively. In ratstudies, we have previously found thismethod of addition to be the mostappropriate when working with puri-
fied fibre such as cellulose, since itmaintains the most constant ratios ofvarious nutrients to digestible energy(6). Though bran contains nutrientsother than fibre, we considered that itwould still be the most applicablemethod. In the formulation of diet #4(low protein, high fibre), we dilutedthe control diet with wheat bran(33.8%), corn meal (31.0%), tricalciumphosphate (0.6%) and powderedlimestone (0.9%).The moisture, crude protein and
dietary fibre contents of the experi-mental diets are reported in Table I.The feeds were taken to constantweights in a vacuum oven and themoisture content determined by dif-
TABLE I. Moisture, Crude Protein, Neutral Detergent Fibre, Acid Detergent Fibre and Hemicellu-lose Content of the Experimental Rations
Neutral AcidCrude Detergent Detergent
Moisture Protein a Fibrea Fibrea Hemicellulosea
(%) (%) (%) (%) (%)Ration #1 6.35 22.7 11.9 3.43 8.5Ration #2 6.54 21.3 16.1 5.03 11.1Ration #3 6.65 21.7 21.7 5.69 16.0Ration #4 7.25 16.1 22.9 6.53 16.3aReported on a dry matter basis
0
Groupno.I -- 23%CP, 12%NDF
no.2-Q-- 21% CP, 16% NDF
no.3 t 22%/CP,22% NDF
no.4--- 16% CP, 23/% NDF
400-
a
w4I-
z
0ww 300-U.
200-
2 3 4 5 6 7 8 9 10 12 13 14TIME (weeks)
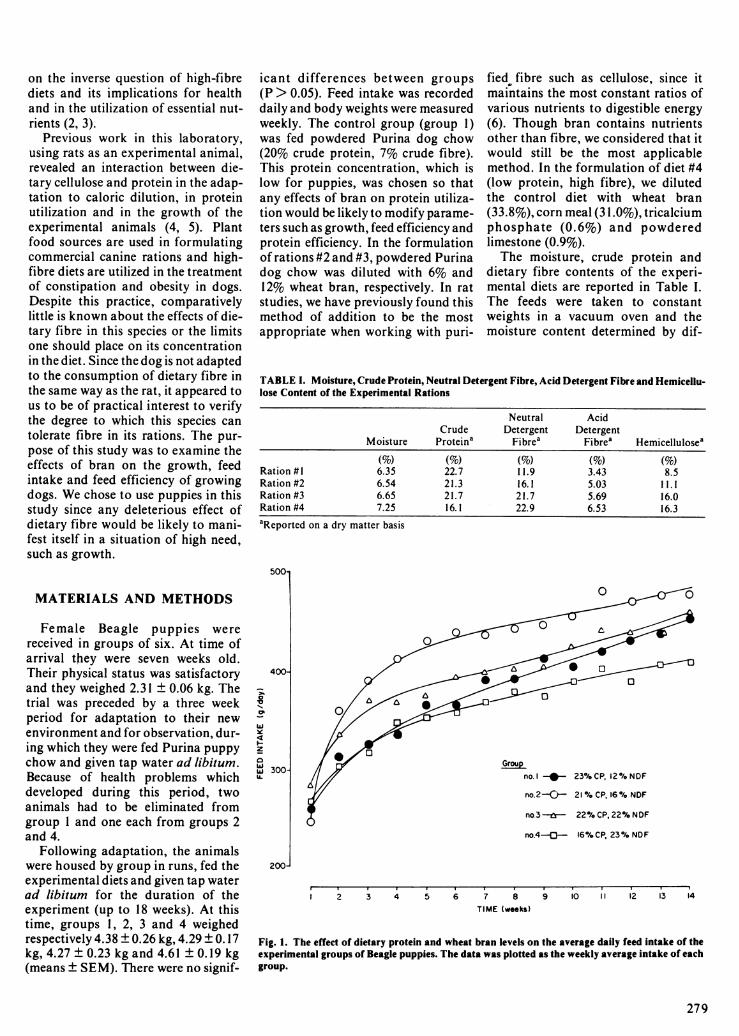
Fig. 1. The effect of dietary protein and wheat bran levels on the average daily feed intake of theexperimental groups of Beagle puppies. The data was plotted as the weekly average intake of eachgroup.
279
ference. The dietary crude protein wasassayed by the Kjeldahl procedure (7).The neutral detergent fibre (NDF) andacid detergent fibre (ADF) were mea-sured according to a modification ofthe method of Goering and Van Soest(8). The method incorporated a rapidtreatment with hog pancreatic amylase(Sigma, A6880), as described by Mon-geau and Brassard (9). This treatmentremoves residual starch and a largeproportion of denatured proteins.
90
80
0
04
zW 60-Li.
zToa:w
w0-J 40-
w
Hemicellulose was calculated as thedifference between NDF and ADF.Nonfibre feed intake was calculated asthe gross feed intake from which theNDF was subtracted.The feed efficiency was calculated
for the first and last 60 days of the trial,as well as for the overall period (thefull 120 days), as grams of body weightgain per gram of feed intake. It wascalculated on the basis both of grossfeed intake and nonfibre intake (over-
<) Groupno.1l-* 23% CP,12% NDF
/ ~~~~~~~~~~no.2-<-21% CP, 16% NDF
no-3-6- 22% CP, 22%NDFno.4-c- 16% CP,23%NDF
20J
1 2 3 4 5 6 7 8 9 10 I 12 13 14
TIME (weeks)
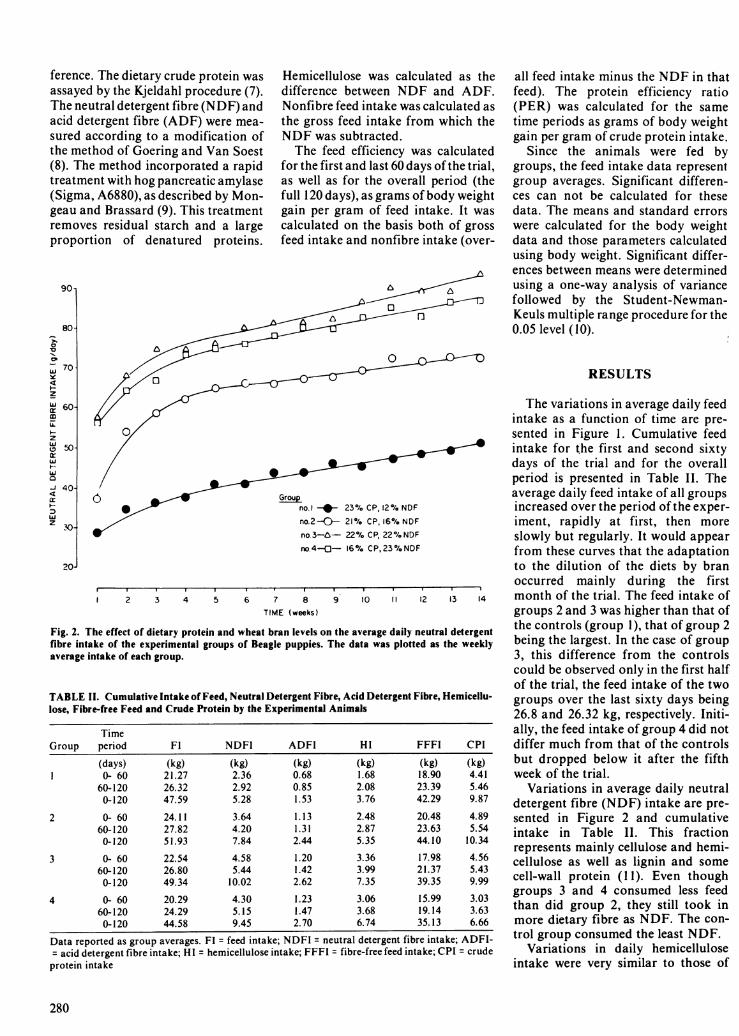
Fig. 2. The effect of dietary protein and wheat bran levels on the average daily neutral detergentflbre intake of the experimental groups of Beagle puppies. The data was plotted as the weeklyaverage intake of each group.
TABLE II. Cumulative Intake of Feed, Neutral Detergent Fibre, Acid Detergent Fibre, Hemicellu-lose, Fibre-free Feed and Crude Protein by the Experimental Animals
TimeGroup period Fl NDFI ADFI HI FFFI CPI
(days) (kg) (kg) (kg) (kg) (kg) (kg)I 0- 60 21.27 2.36 0.68 1.68 18.90 4.41
60-120 26.32 2.92 0.85 2.08 23.39 5.460-120 47.59 5.28 1.53 3.76 42.29 9.87
2 0- 60 24.11 3.64 1.13 2.48 20.48 4.8960-120 27.82 4.20 1.31 2.87 23.63 5.540-120 51.93 7.84 2.44 5.35 44.10 10.34
3 0- 60 22.54 4.58 1.20 3.36 17.98 4.5660-120 26.80 5.44 1.42 3.99 21.37 5.430-120 49.34 10.02 2.62 7.35 39.35 9.99
4 0- 60 20.29 4.30 1.23 3.06 15.99 3.0360-120 24.29 5.15 1.47 3.68 19.14 3.630-120 44.58 9.45 2.70 6.74 35.13 6.66
Data reported as group averages. Fl = feed intake; NDFI neutral detergent fibre intake; ADFI-= acid detergent fibre intake; HI = hemicellulose intake; FFFI = fibre-free feed intake; CPI = crudeprotein intake
all feed intake minus the NDF in thatfeed). The protein efficiency ratio(PER) was calculated for the sametime periods as grams of body weightgain per gram of crude protein intake.
Since the animals were fed bygroups, the feed intake data representgroup averages. Significant differen-ces can not be calculated for thesedata. The means and standard errorswere calculated for the body weightdata and those parameters calculatedusing body weight. Significant differ-ences between means were determinedusing a one-way analysis of variancefollowed by the Student-Newman-Keuls multiple range procedure for the0.05 level (10).
RESULTS
The variations in average daily feedintake as a function of time are pre-sented in Figure 1. Cumulative feedintake for the first and second sixtydays of the trial and for the overallperiod is presented in Table I1. Theaverage daily feed intake of all groupsincreased over the period of the exper-iment, rapidly at first, then moreslowly but regularly. It would appearfrom these curves that the adaptationto the dilution of the diets by branoccurred mainly during the firstmonth of the trial. The feed intake ofgroups 2 and 3 was higher than that ofthe controls (group 1), that of group 2being the largest. In the case of group3, this difference from the controlscould be observed only in the first halfof the trial, the feed intake of the twogroups over the last sixty days being26.8 and 26.32 kg, respectively. Initi-ally, the feed intake of group 4 did notdiffer much from that of the controlsbut dropped below it after the fifthweek of the trial.
Variations in average daily neutraldetergent fibre (NDF) intake are pre-sented in Figure 2 and cumulativeintake in Table II. This fractionrepresents mainly cellulose and hemi-cellulose as well as lignin and somecell-wall protein (11). Even thoughgroups 3 and 4 consumed less feedthan did group 2, they still took inmore dietary fibre as NDF. The con-trol group consumed the least NDF.
Variations in daily hemicelluloseintake were very similar to those of
280
NDF. Those of acid detergent fibre(ADF) were also quite similar,although there was less differencebetween groups 2, 3 and 4 than therewas for NDF or hemicellulose andgroup 4 appears to have consumedmore ADF than did group 3, theinverse of what was observed for NDF
and hemicelluloseof ADF and hensented in Table II.The variations
intake (overall fNDF) as a functisented in Figure 3intake in Table II.
Group
no -- 23% CP.;no 2-0-21% CP, 1(no 3 ---22%CP. 2
no.4 16%CP,2
I I
2 3 4 5 6 7 8 9 10TIME (weeks)
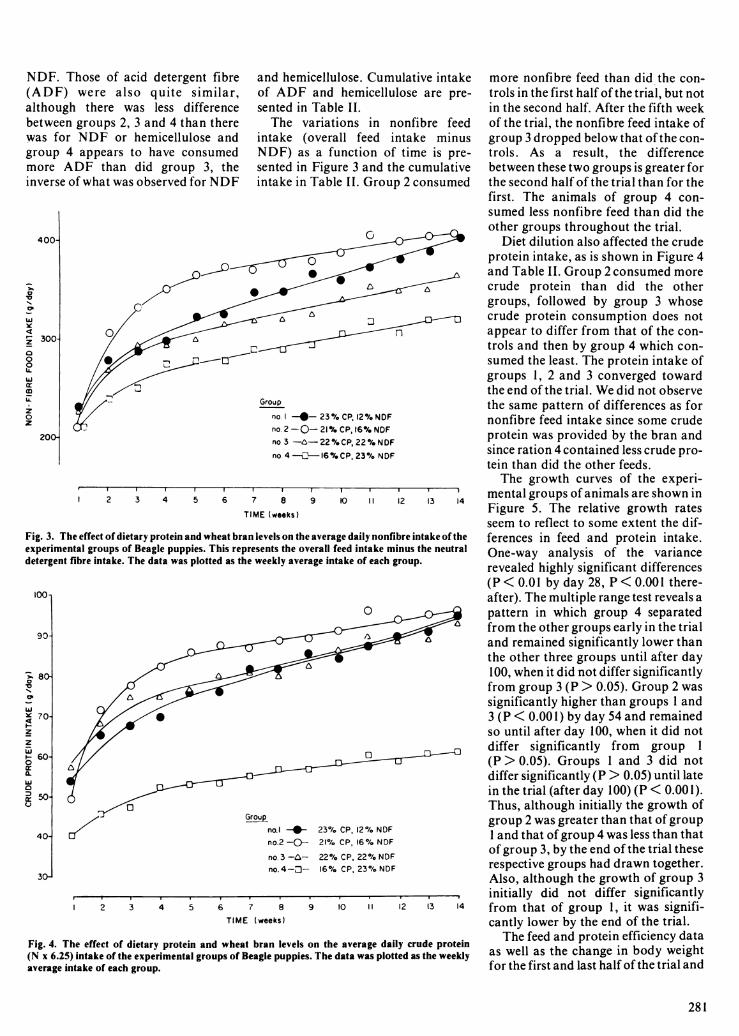
Fig. 3. The effect of dietary protein and wheat bran levels on the average daiexperimental groups of Beagle puppies. This represents the overall feed irdetergent fibre intake. The data was plotted as the weekly average intake c
100l
90-
> 80-a
-Tw
x 70-
zz
60-0
04350-
40-
,0Group
ol/~ no.1 + 23% CP, 14no.2-(- 21% CP, 11
no.3 -a- 22% CP, 2no.4-0- 16% CP, 2
2 3 4 5 6 7 8 9 10
TIME (weeks)
Fig. 4. The effect of dietary protein and wheat bran levels on the aver(N x 6.25) intake of the experimental groups of Beagle puppies. The data v
average intake of each group.
Cumulative intake more nonfibre feed than did the con-nicellulose are pre- trols in the first half of the trial, but not
in the second half. After the fifth weekin nonfibre feed of the trial, the nonfibre feed intake of
feed intake minus group 3 dropped below that of the con-ion of time is pre- trols. As a result, the differenceand the cumulative between these two groups is greater forGroup 2 consumed the second half of the trial than for the
first. The animals of group 4 con-sumed less nonfibre feed than did the
G other groups throughout the trial.Diet dilution also affected the crude
protein intake, as is shown in Figure 4and Table II. Group 2 consumed morecrude protein than did the othergroups, followed by group 3 whose
D3 0 crude protein consumption does notappear to differ from that of the con-trols and then by group 4 which con-sumed the least. The protein intake ofgroups 1, 2 and 3 converged towardthe end of the trial. We did not observethe same pattern of differences as for
2%NDF nonfibre feed intake since some crude2% NDF protein was provided by the bran and
3% NDF since ration 4 contained less crude pro-tein than did the other feeds.The growth curves of the experi-
I2 I3 I4 mental groups of animals are shown inII 12 13 14Figure 5. The relative growth ratesseem to reflect to some extent the dif-
ilynonfibreintakeofthe ferences in feed and protein intake.ntake minus the neutral One-way analysis of the variance)f each group. revealed highly significant differences
(P < 0.01 by day 28, P < 0.001 there-after). The multiple range test reveals a
0 pattern in which group 4 separatedfrom the other groups early in the trialthand remained significantly lower thanthe other three groups until after day100, when it did not differ significantlyfrom group 3 (P > 0.05). Group 2 wassignificantly higher than groups 1 and3 (P < 0.001) by day 54 and remainedso until after day 100, when it did notdiffer significantly from group 1(P> 0.05). Groups 1 and 3 did notdiffer significantly (P > 0.05) until latein the trial (after day 100) (P < 0.001).Thus, although initially the growth of
2% NDF group 2 was greater than that of group6% NDF l and that of group 4 was less than that?2% NDF of group 3, by the end of the trial these?3% NDF respective groups had drawn together.
Also, although the growth of group 3initially did not differ significantly
12 13 14 from that of group 1, it was signifi-cantly lower by the end of the trial.
*age daily crude protein The feed and protein efficiency datawas plotted as the weekly as well as the change in body weight
for the first and last half of the trial and
281
400-
0
w4z 300-a00U.w
U.
0z
200-
0w39_
0
zw0z4U
GroupNo --- 23% CP, 12 % NDF
No.2 --21% CP,16% NDF5- No.3 22% CP,22% NDF
No.4-O-16%CP,23% NDF
4t-1 1 >
O , , , .v _ s,~~~~1
10 20 30 40 50 60 70 80 90 100 110 120
TIME (toys)
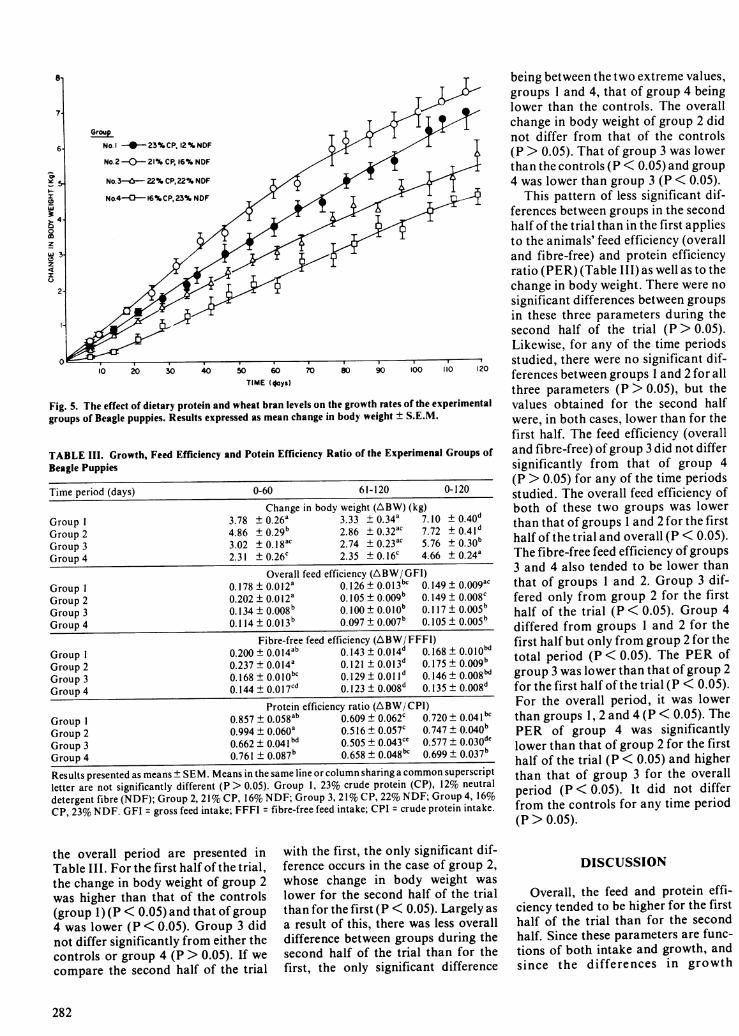
Fig. 5. The effect of dietary protein and wheat bran levels on the growth rates of the experimentalgroups of Beagle puppies. Results expressed as mean change in body weight ± S.E.M.
TABLE III. Growth, Feed Efficiency and Potein Efficiency Ratio of the Experimenal Groups ofBeagle Puppies
Time period (days) 0-60 61-120 0-120
Change in body weight (ABW) (kg)Group 1 3.78 ± 0.26a 3.33 ± 0.34a 7.10 ± 0.40dGroup 2 4.86 ± 0.29" 2.86 ± 0.32ac 7.72 ± 0.41dGroup 3 3.02 ± 0.18ac 2.74 ± 0.23ac 5.76 ± 0.30"Group 4 2.31 ± 0.26c 2.35 ± 0.16c 4.66 ± 0.24a
Overall feed efficiency (ABW/GFI)Group 1 0.178 ± 0.012a 0.126 ± 0.013bc 0.149 ± 0 oogacGroup 2 0.202 ± 0.012a 0.105 ± 0.009b 0. 149 ± 0.008cGroup 3 0.134 ± 0.008b 0.100 ± 0.010b 0.117 0.005bGroup 4 0.114 ± 0.013b 0.097 ± 0.007" 0.105 0.005b
Fibre-free feed efficiency (ABW/ FFFI)Group 1 0.200 ± 0.014"' 0.143 ± 0.014d 0.168 0.010bdGroup 2 0.237 ± 0.014a 0.121 ± 0.013d 0.175 0.009bGroup 3 0.168 ± O.O1Obc 0.129 ± 0.01ld 0.146 + 0.008bdGroup 4 0.144 ± 0.017 0.123 + 0.008 0.135 ± 0.008d
Protein efficiency ratio (ABW/CPI)Group 1 0.857 ± 0s058"a 0.609 ± 0.062c 0.720 ± 0.041"bGroup 2 0.994 ± 0.060a 0.516 ± 0.057c 0.747 ± 0.040bGroup 3 0.662 ± 0.041bd 0.505 ± 0.043ce 0.577 ± 0.030deGroup 4 0.761 ± 0.087" 0.658 ± 0.048bc 0.699 ± 0.037"
Results presented as means ± SEM. Means in the same line or column sharing a common superscriptletter are not significantly different (P > 0.05). Group 1, 23% crude protein (CP), 12% neutraldetergent fibre (NDF); Group 2, 21% CP, 16% NDF; Group 3,21% CP, 22% NDF; Group 4,16%CP, 23% NDF. GFI = gross feed intake; FFFI = fibre-free feed intake; CPI = crude protein intake.
being between the two extreme values,groups I and 4, that of group 4 beinglower than the controls. The overallchange in body weight of group 2 didnot differ from that of the controls(P > 0.05). That of group 3 was lowerthan the controls (P < 0.05) and group4 was lower than group 3 (P < 0.05).
This pattern of less significant dif-ferences between groups in the secondhalf of the trial than in the first appliesto the animals' feed efficiency (overalland fibre-free) and protein efficiencyratio (PER) (Table III) as well as to thechange in body weight. There were nosignificant differences between groupsin these three parameters during thesecond half of the trial (P > 0.05).Likewise, for any of the time periodsstudied, there were no significant dif-ferences between groups 1 and 2 for allthree parameters (P > 0.05), but thevalues obtained for the second halfwere, in both cases, lower than for thefirst half. The feed efficiency (overalland fibre-free) of group 3 did not differsignificantly from that of group 4(P > 0.05) for any of the time periodsstudied. The overall feed efficiency ofboth of these two groups was lowerthan that of groups 1 and 2 for the firsthalf of the trial and overall (P < 0.05).The fibre-free feed efficiency of groups3 and 4 also tended to be lower thanthat of groups 1 and 2. Group 3 dif-fered only from group 2 for the firsthalf of the trial (P < 0.05). Group 4differed from groups 1 and 2 for thefirst half but only from group 2 for thetotal period (P < 0.05). The PER ofgroup 3 was lower than that of group 2for the first half of the trial (P < 0.05).For the overall period, it was lowerthan groups 1, 2 and 4 (P < 0.05). ThePER of group 4 was significantlylower than that of group 2 for the firsthalf of the trial (P < 0.05) and higherthan that of group 3 for the overallperiod (P K 0.05). It did not differfrom the controls for any time period(P > 0.05).
the overall period are presented inTable III. For the first half of the trial,the change in body weight of group 2was higher than that of the controls(group 1) (P < 0.05) and that ofgroup4 was lower (P < 0.05). Group 3 didnot differ significantly from either thecontrols or group 4 (P > 0.05). If wecompare the second half of the trial
with the first, the only significant dif-ference occurs in the case of group 2,whose change in body weight waslower for the second half of the trialthan for the first (P < 0.05). Largely asa result of this, there was less overalldifference between groups during thesecond half of the trial than for thefirst, the only significant difference
DISCUSSION
Overall, the feed and protein effi-ciency tended to be higher for the firsthalf of the trial than for the secondhalf. Since these parameters are func-tions of both intake and growth, andsince the differences in growth
282
E

between the two periods were signifi-cant only in the case of group 2(P < 0.05), this may be taken as anindication that feed and protein intakewere in fact higher in the second half ofthe trial period than in the first andthat differences were more than justapparent. Since, because of growth,the animals were heavier on day 60than on day 0, this result was to beexpected. In fact, although the animalsconsumed more food during the latterpart of the trial, a greater proportionof the nutrients ingested would havebeen required for maintenance, result-ing in lower efficiency.
Differences in feed intake wereobserved between groups. Although itis difficult to evaluate the significanceof these differences, they cannot beattributed to differences in the initialweight of the experimental groups. Onday 0, mean weights did not differ sig-nificantly (P > 0.05). Thus, any differ-ences in intake, at least during theinitial part of the trial, would have tobe attributed to dietary factors. In theabsence of statistics, it is not certainhow much weight can be given toapparent differences in feed intakebetween groups. Nevertheless, at leastin some cases, the absence of signifi-cant differences in feed and proteinefficiency in the face of significant dif-ferences in growth could be taken astending to validate these differences infeed intake. Growth and intake appearto have changed proportionally inthese cases. Though not definite proof,we can view this as supportiveevidence.
Variations in feed intake can beassociated with modifications in theamount of dietary protein, in thecaloric density of the rations and in theratio of digestible protein to digestibleenergy of those rations. Animals adaptto caloric dilution of their diet byincreasing their total feed intake (12-17), although this may be influencedby the protein adequacy ofthe diet andby the ratio of dietary protein to dig-estible energy (4,6,18,19). Dietary pro-tein itself influences food intake.While the weight gained by young ratsis directly related to the amount andquality of protein consumed (20,21),the amount of food and thus of proteineaten is also influenced by the qualityand concentration of dietary protein(22-24). Egan has found that with
sheep fed roughage diets of low nitro-gen content, but not those whose pro-tein content was adequate, the infu-sion of casein or urea into theduodenum increased feed intake( 18,19). It thus appears that a low ratioof dietary digestible protein to digesti-ble energy depresses feed intake, andthat this applies to more than onespecies.
Except for diet 4, whose proteinconcentration was low, we tried tokeep this ratio as constant as possibleby adding the wheat bran to therations by total diet dilution (6). Thus,differences between groups 1, 2 and 3should be mainly due to changes in thefibre content of the diet, rather thanprotein. Likewise, differences betweengroups 3 and 4 should be due to theprotein content of the diet or to theratio of protein to digestible energyrather than to the NDF concentration.As we have seen, although over the
short term (60 days), the growth ofgroup 2 was higher than that of group1, over a longer term (120 days), theirgrowth did not differ. Their feed andprotein efficiency did not differ at anytime. Improved growth responses havebeen produced in various species bythe inclusion of different sources ofdietary fibre in their rations (25-28),improvement which may be due atleast in part to increased feed intake(13,17). The results obtained withgroup 2 are consistent with thesereports.Compared to group 1, group 3 had
lower growth, feed intake, overall feedefficiency and PER, although thefibre-free feed efficiency of the groupsdid not differ significantly. Theseresults are consistent with an energydeficit leading to less efficient use ofthe dietary protein for growth, sincepart would have been catabolized as asource of energy. If kept in mind that22% crude protein is low for puppyrations, any decrease in available foodenergy intake would not only decreasethe proportion of energy available forgrowth, but would also increase thecatabolism of dietary protein as asource of energy. The animal's growthrate would thus be lowered and alongwith it the protein and probably thefeed efficiency.
It appears from our results that thisis what occurred in the case of group 3.The most limiting factor in the growth
of these animals seems to have beentheir low digestible energy intake.Thus, either the dietary NDF hadreached high enough levels or the ratioof protein to digestible energy was lowenough so that these animals were lessable to adapt than were those of group2.The results of group 4 show that the
combination of a low dietary crudeprotein concentration (16% CP) with ahigh NDF level (23% NDF) mainlymodified protein utilization. Althoughtheir initial growth was lower than thatof group 3, their feed efficiency did notdiffer significantly and their growthcurves seemed to be drawing togethertoward the end of the trial. Althoughtheir energy deficit was at least asserious as that of group 3, as indicatedby growth, feed intake and feed effi-ciency, their protein intake was lowerand their overall PER higher. This isconsistent with a relatively more limit-ing amount of dietary protein leadingto a more efficient use for growth bythe animal.
It thus appears that the lower pro-tein content of diet 4 did not signifi-cantly further diminish the animals'longer-term growth or feed efficiency,as compared to group 3. However, thedifferential effect of these two diets onthe PER points out the importance ofthe protein-energy balance in deter-mining what use is made of dietaryprotein by the experimental animal.
It should be pointed out that, asso-ciated with the differences in bodyweight of the experimental groups ofanimals may be differences in bodycomposition. Not wishing to sacrificethe animals, we did not perform car-cass analyses. However, previouswork with rats (33) showed that,although cellulose has been reportedto diminish their fat content (34,35),we found no significant effect unlessthe high cellulose levels were com-bined with a low quality of dietary pro-tein. Under these conditions, the etherextract was diminished and the carcasswater increased (33). On the otherhand, Meyer (36,37) has found thatunder conditions of a lower ratio ofdietary protein to digestible energy,rats had a greater proportion of theirweight gain as fat. It is therefore possi-ble that one or more of the high-fibregroups might have leaner carcasses,particularly where their caloric intake
283

was inadequate. It is uncertain fromthis whether the body fat of group 4might be higher or lower than the con-trols and in the absence of experimen-tal data we can draw no conclusions.From the results of this study, it
seems safe to conclude that in the pres-ence of low concentrations of dietaryprotein, there is a limit to the amountof bran which can be safely added topuppy rations. This shows up clearlywith group 3 (22% NDF). No delete-rious effect could be observed withintermediate levels of NDF (16%).Indeed, depending on the period stu-died, one might even be led to believein some improvement in growth.Growth responses have been
reported to be improved by the inclu-sion of fibrous materials in the diets ofvarious species, such as rats, mice,guinea pigs, chicks and swine (25-29,38-41). In the light of the presentstudy, we may perhaps extend thisobservation to dogs, although theeffect observed depended not only onthe diet fed to the animals but also onthe duration of the trial and the timeperiod studied. At least part of sucheffects on growth may be due toincreased feed consumption (14,17)which itself may be related to anincreased rate of transit (30,31). Adap-tive change in the size of the digestivetract may also be involved (32). It hasalso been proposed that intestinalmicroflora might be involved. Fibredegradation in the gut is highly varia-ble, depending on the type of fibre, theanimal species used, their age andindividual variations (42). Individualdifferences in digestibility have beenrelated to transit time (30,42). Accord-ing to Cummings (43), there can belittle doubt that a fermentive systemsimilar to that of the rumen occurs inthe colon of many animals, includingman. Its importance would depend onthe amount and type of dietary fibreand of other substrates ingested whichreach the hind gut, as well as on theanimal species. It appears that in allmammalian species studied, consider-able amounts of short-chain fattyacids are produced, and most areabsorbed. With high-fibre diets, short-chain fatty acids may be making a sig-nificant contribution to energy bal-ance (43). In the case of diet 3, the fibredoes not seem to have contributed tothe energy balance. Since there is
competition between the transit timeand bacterial growth in the lowerintestinal tract, the additional fibreintake may have increased the rate oftransit to an extent where the lag phasefor bacterial growth preventedoptimal production and/or absorp-tion of volatile fatty acids from thelarge intestine.
In conclusion, the addition of wheatbran to puppy rations, bringing NDFup to 16% in a 21% crude protein diethad no deleterious effects on feedintake, feed and protein efficiency orgrowth in Beagle puppies. Over a suf-ficiently long period of time, thegrowth of this group did not differ sig-nificantly from that of the controls(12% NDF, 23% CP). Higher NDFlevels should be avoided, at least withlow-protein diets. The effects of thehigh fibre diets (22 or 23% NDF) ongrowth, feed intake, feed efficiencyand PER reflected to a large extent theenergy deficit resulting from theirinability to compensate for the dilu-tion of their diets, conjugated or notwith a low dietary crude protein level(22% versus 16% CP). Once again,over a sufficiently long period of time,the growth and feed efficiency of thesetwo groups did not differ, although thePER of the low (16%) protein groupwas higher than that of the 22% pro-tein group.
Considering the differences ob-served in results between the first andsecond half of the trial and for the fullperiod, it is clear that the results andthe conclusions drawn from themcould differ, depending on the dura-tion of the trial and on the time periodused. This brings up the usefulness oflonger-term trials which, even thoughmore onerous than studies of shorterduration, allow more time not only forchronic effects to appear but also foradaptation to take place.
REFERENCES
1. MENDELOFF AI. Dietary fiber. Nutr Rev1975; 33: 321-330.
2. SPILLER GA, AMEN RJ. Dietary fiber inhuman nutrition. CRC Crit Rev Food SciNutr 1975; 7: 39-70.
3. SPILLER GA, SHIPLEY EA, BLAKE JA.Recent progress in dietary fiber (plantix) inhuman nutrition. CRC Crit Rev Food SciNutr 1978; 10: 31-90.
4. DELORME CB, WOJCIK J. The interac-tion of dietary protein with cellulose in theadaptation to caloric dilution by weanlingrats. J Nutr 1982; 112: 21-28.
5. WOJCIK J, DELORME CB. The effect ofdietary cellulose level on the utilization ofamino acid-supplemented bread protein byweanling rats. Nutr Rep Int 1982; 25:709-720.
6. DELORME CB, WOJCIK J, GORDONC. Method of addition of cellulose to exper-imental diets and its effect on rat growth andprotein utilization. J Nutr 1981; 111:1522-1527.
7. ASSOCIATION OF OFFICIAL ANA-LYTICAL CHEMISTS, Official methodsof analysis. 12th ed. W. Horowitz, ed.AOAC, Washington, D.C. 1975.
8. GOERING HK, VAN SOEST PJ. Forageanalyses. In: US Dept Agric Handbook 379,USDA, Washington, D.C. 1970.
9. MONGEAU R, BRASSARD R. Determi-nation of neutral detergent fibre in break-fast cereals: Pentose, hemicellulose, cellu-lose and lignin content. J Food Sci 1982; 47:550-555.
10. NIE NH, HULL CH, JENKINS JG,STEINBRENNER K, BENT DH. Statisti-cal package for the social sciences. 2nd ed.Toronto: McGraw Hill, 1975.
11. VAN SOEST PJ, ROBERTSON JB. Whatis fibre and fibre in food? Nutr Rev 1977; 35:12-22.
12. JANOWITZ HD, GROSSMAN MI. Effectof variations in nutritive density on intakeof food of dogs and rats. Am J Physiol 1949;158: 184-193.
13. HILL FW, DANSKY LM. Studies of theenergy requirements of chickens. 1. Theeffect of dietary energy level on growth andfeed consumption. Poult Sci 1954; 33:112-119.
14. PETERSON DW, GRAY CR, PEEK NF.Growth and food consumption in relationto dietary levels of protein and fibrous bulk.J Nutr 1954; 52: 241-257.
15. SIBBALD IR, BERG RT, BOWLAND JP.Digestible energy in relation to food intakeand nitrogen retention in the weanling rat. JNutr 1956; 59: 385-392.
16. PETERSON AD, BAUMGARDT BR.Food and energy intake of rats fed dietsvarying in energy concentration and den-sity. J Nutr 1971; 101: 1057-1067.
17. BOHMAN VR, KIDWELL JF, McCOR-MICK JA. High levels of alfalfa in therations of growing-fattening swine. J AnimSci 1953; 12: 876-880.
18. EGAN AR, MOIR RJ. Nutritional statusand intake regulation in sheep. 1. Effects ofduodenally infused single doses of casein,urea and propionate upon voluntary intakeof a low-protein roughage by sheep. Aust JAgric Res 1965; 16: 437-449.
19. EGAN AR. Nutritional status and regula-tion in sheep. VIII Relationships betweenthe voluntary intake of herbage by sheepand the protein/energy ratio in the digestionproducts. Aust J Agric Res 1977; 28:907-915.
20. HEGSTED DM, CHANG Y. Protein utili-zation in rats. I. Relative growth index as a
bio-assay procedure. J Nutr 1965; 85:159-168.
284

21. HEGSTED DM, CHANG Y. Protein utili-zation in growing rats at different levels ofintake. J Nutr 1965; 87: 19-25.
22. OSBORNE TB, MENDEL LB, FERRYEL. A method of expressing numerically thegrowth-promoting value of proteins. J BiolChem 1919; 37: 223-229.
23. HARPER AE, BENEVENGA NJ, WOHL-HUETER RM. Effects of ingestion of dis-proportionate amounts of amino acids.Physiol Rev 1970; 50: 428-558.
24. HARPER AE. Effects of disproportionateamounts of amino acids. In: Improvementof protein nutriture. Washington, D.C.:National Academy of Sciences, 1974.
25. BELL JM, GIDYK M. The effects of frostdamage on the nutritional value of wheat.Can J Agric Sci 1954; 34: 305-312.
26. DAVIS F, BRIGGS GM. The growth-promoting action of cellulose in purifieddiets for chicks. J Nutr 1947; 34: 295-300.
27. DAVIS F. Sawdust in purified rations.Poult Sci 1948; 27: 117-118.
28. DAVIS F. On the use of cellulose in diets.Science 1948; 107: 292.
29. LEPP A, HARPER AE, ELVEHJEM CA.Cellulose in purified rations for chicks.Poult Sci 1949; 28: 372-374.
30. HUMMEL FC, SHEPHERD ML, MACYIG. Disappearance of cellulose and hemicel-
lulose from the digestive tracts of children. JNutr 1943; 25: 59-70.
31. BELL JM. A comparison of fibrous feed-stuffs in non-ruminant rations. Effects ongrowth responses, digestibility, rates of pas-sage and ingesta volume. Can J Anim Sci1960; 40: 71-82.
32. BOHMAN VR, HUNTER JE, McCOR-MICK J. The effect of graded levels ofalfalfa and aureomycin upon growing-fattening swine. J Anim Sci 1955; 14:499-506.
33. WOJCIK J, DELORME CB. Modificationby cellulose of the effect of dietary proteinquality on the carcass and liver compositionof weanling rats. Nutr Rep Int 1983; 27:745-755.
34. SUNDERAVALLI OE, SHURPALE-KAR KS, RAO MN. Effects of dietary cel-lulose supplements on the body composi-tion and cholesterol metabolism of albinorats. J Agric Food Chem 1971; 19: 116-118.
35. SUNDERAVALLI OE, SHURPALE-KAR KS, RAO MN. Inclusion of cellulosein calorie-restricted diets. J Am Diet Assoc1963; 62: 41-43.
36. MEYER JH. Interactions of dietary fiberand protein on food intake and body com-position of growing rats. Am J Physiol1958; 193: 488-495.
37. MEYER JH, HARGUS WA. Factorsinfluencing food intake of rats fed low-protein rations. Am J Physiol 1959; 197:1350-1352.
38. CHEEKE PR, PATTON NM. Effect ofalfalfa and dietary fiber on the growth per-formance of weanling rabbits. Lab AnimSci 1978; 28: 167-172.
39. HEGDE SN, ROLLS BA, TURVEY A,COATES ME. Effects on chicks of dietaryfibre from different sources: a growth factorin wheat bran. Br J Nutr 1978; 40: 63-69.
40. WILLIAMS JE, McLAREN GA, FAHEYGC Jr. Influence of acid-resistant hemicellu-lose degradation products on rat andmicrobial growth. Nutr Rep Int 1979; 19:491-498.
41. McLAREN GA, CUPPETT SL, WILLI-AMS JE, FAHEY GC Jr, SMITH TR.Acid-resistant hemicellulose fractions in ratnutrition. Nutr Rep Int 1974; 10: 163-172.
42. CUMMINGS JH. Dietary fibre. Br MedBull 1981; 37: 65-70.
43. CUMMINGS JH. Short-chain fatty acidsin the human colon. Gut 1981; 22: 763-779.
285