the edwardsiella piscicida type iii translocon protein ... · the edwardsiella piscicida type iii...
TRANSCRIPT

The Edwardsiella piscicida Type III Translocon Protein EseCInhibits Biofilm Formation by Sequestering EseE
Ying Li Liu,a,b Tian Tian He,a,b Lu Yi Liu,a Jia Yi,a,b Pin Nie,a,c,d Hong Bing Yu,e Hai Xia Xiea
aState Key Laboratory of Freshwater Ecology and Biotechnology, and Key Laboratory of Aquaculture Disease Control, Ministry of Agriculture, Institute of Hydrobiology,Chinese Academy of Sciences, Wuhan, Hubei, China
bUniversity of Chinese Academy of Sciences, Beijing, ChinacLaboratory for Marine Biology and Biotechnology, Qingdao National Laboratory for Marine Science and Technology, Qingdao, Shandong, ChinadCollege of Marine Science and Engineering, Qingdao Agricultural University, Qingdao, Shandong, ChinaeDepartment of Pediatrics, BC Children's Hospital Research Institute and the University of British Columbia, Vancouver, British Columbia, Canada
ABSTRACT The type III secretion system (T3SS) is one of the most important viru-lence factors of the fish pathogen Edwardsiella piscicida. It contains three transloconproteins, EseB, EseC, and EseD, required for translocation of effector proteins intohost cells. We have previously shown that EseB forms filamentous appendages onthe surface of E. piscicida, and these filamentous structures mediate bacterial cell-cellinteractions promoting autoaggregation and biofilm formation. In the present study,we show that EseC, but not EseD, inhibits the autoaggregation and biofilm forma-tion of E. piscicida. At 18 h postsubculture, a ΔeseC strain developed strong autoag-gregation and mature biofilm formation, accompanied by enhanced formation ofEseB filamentous appendages. This is in contrast to the weak autoaggregation andimmature biofilm formation seen in the E. piscicida wild-type strain. EseE, a proteinthat directly binds to EseC and also positively regulates the transcription of the escC-eseE operon, was liberated and showed increased levels in the absence of EseC. Thisled to augmented transcription of the escC-eseE operon, thereby increasing thesteady-state protein levels of intracellular EseB, EseD, and EseE, as well as biofilmformation. Notably, the levels of intracellular EseB and EseD produced by the ΔeseEand ΔeseC ΔeseE strains were similar but remarkably lower than those produced bythe wild-type strain at 18 h postsubculture. Taken together, we have shown that thetranslocon protein EseC inhibits biofilm formation through sequestering EseE, a posi-tive regulator of the escC-eseE operon.
IMPORTANCE Edwardsiella piscicida, previously known as Edwardsiella tarda, is aGram-negative intracellular pathogen that mainly infects fish. The type III secretionsystem (T3SS) plays a pivotal role in its pathogenesis. The T3SS translocon proteinEseB is required for the assembly of filamentous appendages on the surface of E. pi-scicida. The interactions between the appendages facilitate autoaggregation and bio-film formation. In this study, we explored the role of the other two translocon pro-teins, EseC and EseD, in biofilm formation. We have demonstrated that EseC, but notEseD, inhibits the autoaggregation and biofilm formation of E. piscicida, providingnew insights into the regulatory mechanism involved in E. piscicida biofilm forma-tion.
KEYWORDS Edwardsiella piscicida, biofilm, translocon protein, type III secretionsystem
Edwardsiella piscicida PPD130/91, previously known as Edwardsiella tarda PPD130/91(1, 2), is a Gram-negative intracellular pathogen that often infects fish, causing liver
hypertrophy, renal abscess, or splenic abscess (3, 4). One of its most important virulence
Citation Liu YL, He TT, Liu LY, Yi J, Nie P, Yu HB,Xie HX. 2019. The Edwardsiella piscicida type IIItranslocon protein EseC inhibits biofilmformation by sequestering EseE. Appl EnvironMicrobiol 85:e02133-18. https://doi.org/10.1128/AEM.02133-18.
Editor Hideaki Nojiri, University of Tokyo
Copyright © 2019 American Society forMicrobiology. All Rights Reserved.
Address correspondence to Hai Xia Xie,[email protected].
Received 4 September 2018Accepted 4 February 2019
Accepted manuscript posted online 15February 2019Published
GENETICS AND MOLECULAR BIOLOGY
crossm
April 2019 Volume 85 Issue 8 e02133-18 aem.asm.org 1Applied and Environmental Microbiology
4 April 2019
on January 24, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

factors is the type III secretion system (T3SS), which is also widely present in many otherGram-negative bacteria (5, 6).
The T3SS is located on bacterial surface and delivers bacterial proteins (effectors)from bacterial cytosol into host cells via a one-step effector translocation process. Thisstrategy enables bacteria to survive and replicate inside the host cell (7, 8). The E.piscicida T3SS locus contains 34 genes encoding the secretion system apparatus,chaperones, regulators, and effectors (5, 9–14). When E. piscicida cells are cultured invitro under a T3SS-inducing condition, three major proteins, EseB, EseC, and EseD, aresecreted into the supernatant (15). Upon secretion, these three proteins form a proteincomplex that induces pore formation on the host cell membrane and delivers T3SSeffectors into the host cytosol (15).
E. piscicida and many other bacteria use the biofilm formation as a strategy tosurvive and thrive in a hostile environment. Notably, biofilm formation and type IIIsecretion are not mutually exclusive (16). For example, we have shown that EseB isessential for biofilm formation of E. piscicida (17). Similar to this, Pseudomonas aerugi-nosa requires the T3SS translocon protein PopB to form cell-associated aggregation,which exhibits key characteristics of biofilm formation on the surface of polarizedepithelial cells (18).
Moreover, the elevated Salmonella pathogenicity island 1 (SPI-1) type III secretionmediates biofilm formation in Salmonella enterica serovar Typhimurium (19). In Xan-thomonas citri subsp. citri, the loss of a functional T3SS causes changes in the expres-sion of proteins involved in metabolic processes, exopolysaccharide (EPS) production,and bacterial motility, all of which contribute to biofilm formation (20).
In this study, we examined the role of the other two E. piscicida translocon proteins,EseC and EseD, in biofilm formation. We show that EseC inhibits biofilm formationthrough sequestering EseE, a positive regulator of the escC-eseE operon, whereas EseDdoes not play any roles in biofilm formation.
RESULTSEseC inhibits autoaggregation of E. piscicida at 18 h postsubculture. We have
previously shown that the translocon protein EseB forms filamentous appendages thatpromote autoaggregation in E. piscicida (17). Whether the other two translocon pro-teins (EseC and EseD) also contribute to autoaggregation remains unknown. To addressthis, E. piscicida wild-type (WT) and mutant strains that lack eseB, eseC, or eseD (i.e.,ΔeseB, ΔeseC, or ΔeseD strain, respectively) were cultured overnight, followed bysubculture in Dulbecco’s modified Eagle medium (DMEM) at 25°C under a 5% CO2
atmosphere. At 18 h postsubculture, cell clumps were formed at the bottom of theglass tube cultured with the ΔeseC strain but not the other strains (Fig. 1). Moreover,
FIG 1 Dynamic analysis of E. piscicida autoaggregation. E. piscicida strains were cultured in DMEM in glasstubes. Autoaggregation was recorded at 18 h and 24 h postsubculture (hps).
Liu et al. Applied and Environmental Microbiology
April 2019 Volume 85 Issue 8 e02133-18 aem.asm.org 2
on January 24, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

the supernatant of the ΔeseC culture was transparent, whereas the supernatants ofother cultures were cloudy. These findings suggest that the ΔeseC strain has a strongerability to form autoaggregation than does the WT strain. However, this difference wasnot seen at 24 h postsubculture, when all strains except for the ΔeseB strain formed cellclumps at the bottom of the tubes. To further confirm the role of EseC in inhibitingautoaggregation of E. piscicida at 18 h postsubculture, we generated a complementa-tion strain (ΔeseC/pJN-eseC) where a functional allele of eseC carried on a plasmid(pJN105) was introduced into the ΔeseC strain. Indeed, the autoaggregation levels (i.e.,cell clumps) of the ΔeseC/pJN-eseC complementation strain and the WT strain arecomparable at both 18 h and 24 h postsubculture (Fig. 1). The ΔeseD mutant strain andits complementation strain (ΔeseD/pJN-eseD) displayed the same level of autoaggre-gation as the WT strain at both time points, suggesting that EseD does not play a rolein the autoaggregation process. These findings demonstrate that EseC, but not EseD,inhibits autoaggregation of E. piscicida at 18 h postsubculture.
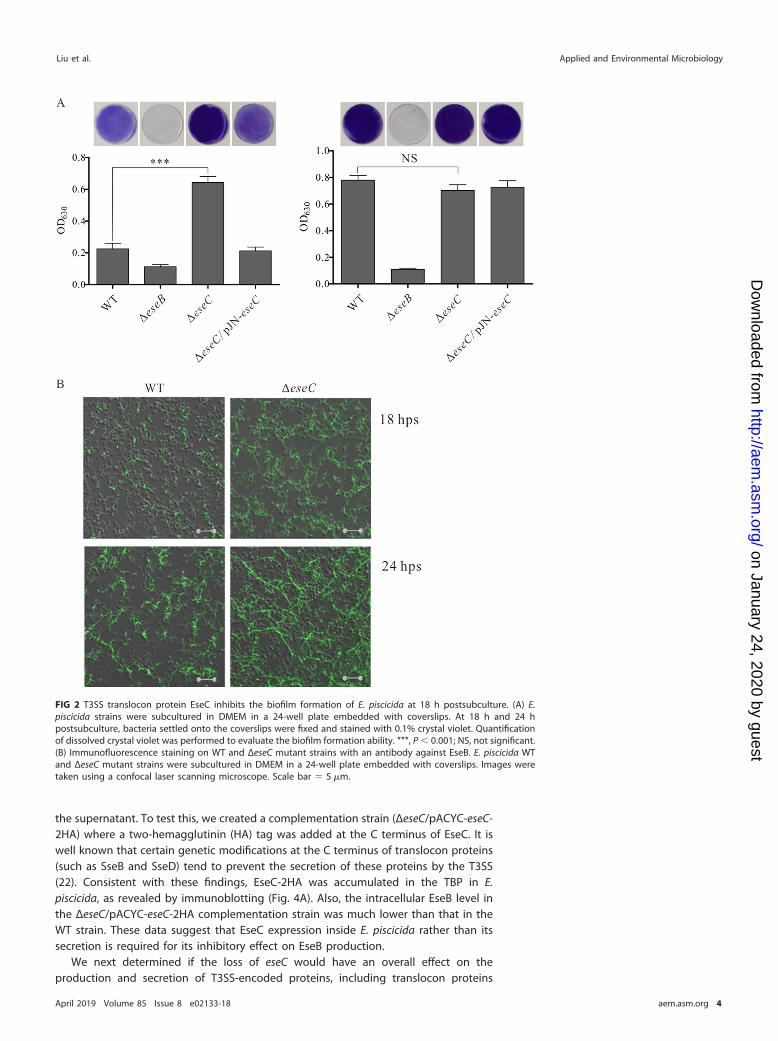
EseC inhibits biofilm formation at 18 h postsubculture. Autoaggregation andbiofilm formation are closely related in E. piscicida (17). We next sought to determineif EseC also plays a role in biofilm formation in E. piscicida. The WT and the ΔeseB, ΔeseC,and ΔeseC/pJN-eseC strains were subcultured into 24-well plates embedded withcoverslips for 18 h and 24 h. Biofilms formed on the coverslips were visualized with acrystal violet staining method and then solubilized into SDS to quantify biofilmformation (as reflected by the optical density at 630 nm [OD630] values). At 18 hpostsubculture, the ΔeseC strain formed a mature biofilm, with an OD630 value of 0.64,whereas the WT and the ΔeseC/pJN-eseC strains formed an immature biofilm (withOD630 values close to 0.2), and no biofilm was developed by the ΔeseB strain (OD630,0.11) (Fig. 2A, left). However, at 24 h postsubculture, the WT, ΔeseC, and ΔeseC/pJN-eseCstrains formed similar levels of mature biofilm, and the ΔeseB strain showed no biofilmformation, as expected (Fig. 2A, right). Thus, EseC also inhibits biofilm formation in E.piscicida at 18 h postsubculture.
EseC inhibits the assembly of EseB filaments on the surface of E. piscicida. Tounderstand the cellular mechanism by which EseC inhibits the biofilm formation of E.piscicida, WT and ΔeseC mutant strains attached to coverslips at 18 h and 24 hpostsubculture were probed with an anti-EseB antibody (17) and donkey anti-mouseIgG Alexa Fluor 488 antibody. As shown in Fig. 2B, much longer and more abundantEseB filamentous appendages were assembled on the surface of the ΔeseC strain thanon the WT strain at both time points (Fig. 2B), suggesting that EseC inhibits theassembly of EseB filaments on the surface of E. piscicida. We also saw brighterfilamentous appendages in both strains at 24 h postsubculture than those at 18 hpostsubculture. This suggests that higher levels of EseB are produced at a later timepoint, which potentially results in the saturation of biofilm formation in both strains asseen in Fig. 2A (right panel).
EseC suppresses the production of intracellular EseB. We next explored themolecular mechanism underlying the inhibition of EseB filament assembly by EseC. WT,ΔeseC mutant, and ΔeseC/pJN-eseC complementation strains were subcultured for 18 has described above, and their total bacterial protein (TBP) lysates were subjected toWestern blotting analyses. EvpC, a protein secreted via the type VI secretion system(T6SS) but not by the T3SS, was used as a protein loading control (21). As shown in Fig.3A, the ΔeseC strain did not show any EseC signals, as expected. Notably, intracellularlevels of EseB were increased by 3.3-fold in the ΔeseC strain compared to the WT strain,and the intracellular protein level of EseB in the complementation strain (ΔeseC/pJN-eseC) was slightly higher than that in WT strain (1.75-fold), while it was much lower thanthat in the ΔeseC mutant strain (Fig. 3B). These data show that EseC suppressesintracellular EseB production in E. piscicida.
EseC suppresses the production of proteins encoded by the escC-eseE operon,including eseB. While EseC suppresses intracellular EseB production, it is unclear if thisresults from the expression of EseC within the bacteria and/or the secretion of EseC into
EseC Inhibits Biofilm Formation through Sequestering EseE Applied and Environmental Microbiology
April 2019 Volume 85 Issue 8 e02133-18 aem.asm.org 3
on January 24, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from
the supernatant. To test this, we created a complementation strain (ΔeseC/pACYC-eseC-2HA) where a two-hemagglutinin (HA) tag was added at the C terminus of EseC. It iswell known that certain genetic modifications at the C terminus of translocon proteins(such as SseB and SseD) tend to prevent the secretion of these proteins by the T3SS(22). Consistent with these findings, EseC-2HA was accumulated in the TBP in E.piscicida, as revealed by immunoblotting (Fig. 4A). Also, the intracellular EseB level inthe ΔeseC/pACYC-eseC-2HA complementation strain was much lower than that in theWT strain. These data suggest that EseC expression inside E. piscicida rather than itssecretion is required for its inhibitory effect on EseB production.
We next determined if the loss of eseC would have an overall effect on theproduction and secretion of T3SS-encoded proteins, including translocon proteins
FIG 2 T3SS translocon protein EseC inhibits the biofilm formation of E. piscicida at 18 h postsubculture. (A) E.piscicida strains were subcultured in DMEM in a 24-well plate embedded with coverslips. At 18 h and 24 hpostsubculture, bacteria settled onto the coverslips were fixed and stained with 0.1% crystal violet. Quantificationof dissolved crystal violet was performed to evaluate the biofilm formation ability. ***, P � 0.001; NS, not significant.(B) Immunofluorescence staining on WT and ΔeseC mutant strains with an antibody against EseB. E. piscicida WTand ΔeseC mutant strains were subcultured in DMEM in a 24-well plate embedded with coverslips. Images weretaken using a confocal laser scanning microscope. Scale bar � 5 �m.
Liu et al. Applied and Environmental Microbiology
April 2019 Volume 85 Issue 8 e02133-18 aem.asm.org 4
on January 24, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

(EseB/C/D), effector proteins (EseG/J) (11, 12), and EseE (a chaperone protein for EseC)(23). EvpC, a T6SS-encoded protein, was used as a loading control. As shown in Fig. 4Aand B, intracellular levels of EseB, EseD, and EseE were increased by 1.6- to 3.2-fold inthe ΔeseC strain but decreased by 1.2- to 4.0-fold in the ΔeseC/pACYC-eseC-2HA straincompared to the WT strain. Accordingly, the secretion of EseB and EseD was increasedin the ΔeseC strain but decreased in the ΔeseC/pACYC-eseC-2HA strain. Unlike EseB/D/E,the intracellular and extracellular levels of EseG and EseJ were not altered in theabsence of eseC. Interestingly, eseB, eseD, and eseE are all located in the escC-eseEoperon, whereas eseG and eseJ are located in other operons (Fig. 4C) (23, 24). Thissuggests that EseC may selectively control the expression of proteins encoded by theescC-eseE operon.
EseC suppresses the transcription of the escC-eseE operon by sequesteringEseE. EseE is not only a chaperone protein that directly binds to EseC but also aregulator that promotes the transcription of the escC-eseE operon (23). This led us tohypothesize that EseC suppresses the production of proteins encoded by the escC-eseEoperon through its interaction with EseE. We therefore compared the amount of EseBand EseD in TBP and extracellular protein (ECP) fractions of the WT, ΔeseE mutant, andΔeseC ΔeseE mutant (a mutant lacking both eseE and eseC) strains. Compared to the WTstrain, both the ΔeseE and ΔeseC ΔeseE strains showed reduced EseB and EseD levels intheir TBP and ECP fractions (Fig. 4D). This is consistent with our previous report showing
FIG 3 Intracellular EseB production is decreased in the presence of EseC. (A) The TBP from equal amountsof E. piscicida wild-type, ΔeseC, and ΔeseC/pJN-eseC strains were probed with antibodies against EseC,EseB, and EvpC. EvpC, a T6SS-secreted protein, was used as the loading control. To induce the expressionof EseC, 10 mM L-arabinose was supplemented into the culture of the ΔeseC/pJN-eseC complementationstrain. (B) Quantitative analysis of EseB protein level from TBP. Proteins were quantified by densitometryand normalized to EvpC. The graphs show the relative ratios of intracellular EseB, which are averages ofthe results from at least three independent experiments. **, P � 0.01.
EseC Inhibits Biofilm Formation through Sequestering EseE Applied and Environmental Microbiology
April 2019 Volume 85 Issue 8 e02133-18 aem.asm.org 5
on January 24, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from
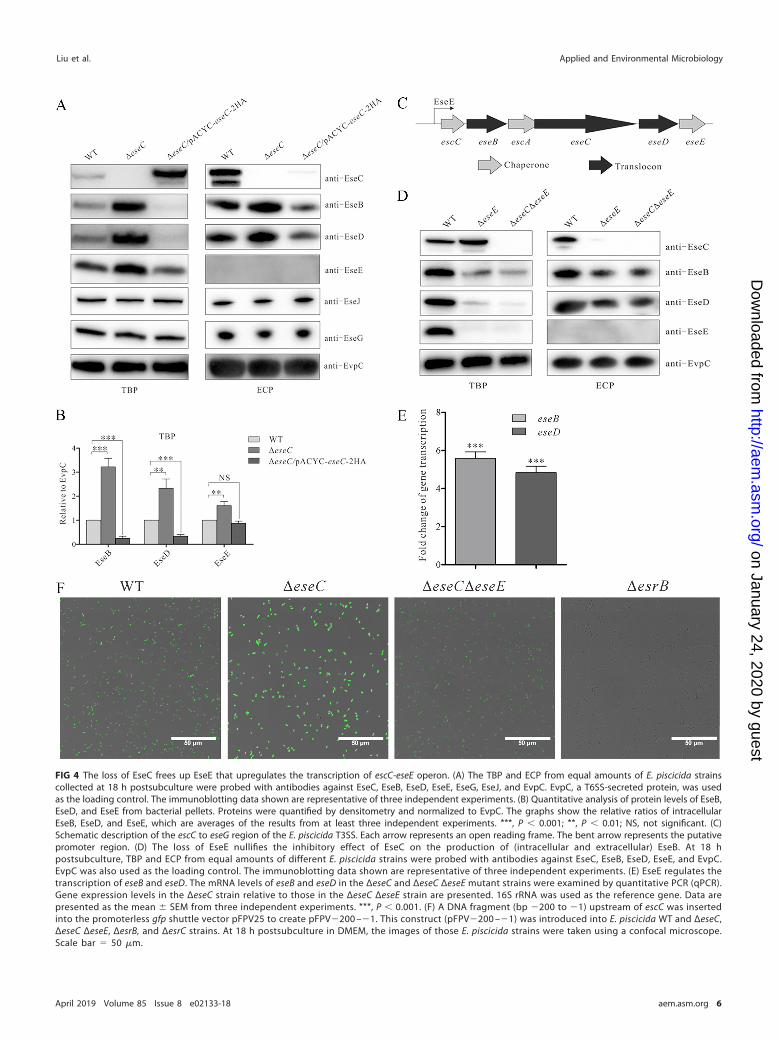
FIG 4 The loss of EseC frees up EseE that upregulates the transcription of escC-eseE operon. (A) The TBP and ECP from equal amounts of E. piscicida strainscollected at 18 h postsubculture were probed with antibodies against EseC, EseB, EseD, EseE, EseG, EseJ, and EvpC. EvpC, a T6SS-secreted protein, was usedas the loading control. The immunoblotting data shown are representative of three independent experiments. (B) Quantitative analysis of protein levels of EseB,EseD, and EseE from bacterial pellets. Proteins were quantified by densitometry and normalized to EvpC. The graphs show the relative ratios of intracellularEseB, EseD, and EseE, which are averages of the results from at least three independent experiments. ***, P � 0.001; **, P � 0.01; NS, not significant. (C)Schematic description of the escC to eseG region of the E. piscicida T3SS. Each arrow represents an open reading frame. The bent arrow represents the putativepromoter region. (D) The loss of EseE nullifies the inhibitory effect of EseC on the production of (intracellular and extracellular) EseB. At 18 hpostsubculture, TBP and ECP from equal amounts of different E. piscicida strains were probed with antibodies against EseC, EseB, EseD, EseE, and EvpC.EvpC was also used as the loading control. The immunoblotting data shown are representative of three independent experiments. (E) EseE regulates thetranscription of eseB and eseD. The mRNA levels of eseB and eseD in the ΔeseC and ΔeseC ΔeseE mutant strains were examined by quantitative PCR (qPCR).Gene expression levels in the ΔeseC strain relative to those in the ΔeseC ΔeseE strain are presented. 16S rRNA was used as the reference gene. Data arepresented as the mean � SEM from three independent experiments. ***, P � 0.001. (F) A DNA fragment (bp �200 to �1) upstream of escC was insertedinto the promoterless gfp shuttle vector pFPV25 to create pFPV�200 –�1. This construct (pFPV�200 –�1) was introduced into E. piscicida WT and ΔeseC,ΔeseC ΔeseE, ΔesrB, and ΔesrC strains. At 18 h postsubculture in DMEM, the images of those E. piscicida strains were taken using a confocal microscope.Scale bar � 50 �m.
Liu et al. Applied and Environmental Microbiology
April 2019 Volume 85 Issue 8 e02133-18 aem.asm.org 6
on January 24, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

that the transcription of eseB and eseD was downregulated by �4-fold in the absenceof EseE (23). Furthermore, the transcription of eseB and eseD in the ΔeseC ΔeseE mutantstrain was �5 times lower than that in the ΔeseC mutant strain (Fig. 4E). These findingssuggest that EseE plays a key role in mediating the upregulated transcription of eseBand eseD in the ΔeseC strain.
To further understand if EseE directly regulates the transcription of the escC-eseEoperon, a putative promoter region (200 bp) upstream of the escC start codon wascloned upstream of a promoterless gfp in pFPV25. The resulting construct, pFPV�200 –�1, was transformed into the WT and ΔeseC, ΔeseC ΔeseE, ΔesrB, and ΔesrC mutantstrains. This green fluorescent protein (GFP) reporter system can be used to measurethe activity of this putative promoter in the strains mentioned above. The ΔesrB andΔesrC mutant strains were used as controls, as EsrB and EsrC are known to positivelyregulate the transcription of T3SS genes (9). As seen in Fig. 4F, the intensity of GFPsignals was the strongest in the ΔeseC strain and modest in the WT strain but very weakor invisible in the ΔeseC ΔeseE, ΔesrB, and ΔesrC mutant strains. These data suggest thatthis promoter region is positively regulated by EseE and further confirm that EseCsuppresses its activity through EseE. Meanwhile, using purified EseE and an electro-phoretic mobility shift assay, we failed to show a direct interaction of EseE with thisputative promoter (data not shown). Whether EseE indirectly interacts with this pro-moter, or interacts with a longer promoter region, remains to be further explored.
Taken together, it appears that EseE is bound (i.e., sequestered) by EseC in the WTstrain, preventing its function as a positive regulator of the escC-eseE operon (includingeseB and eseD); however, in the absence of EseC (i.e., in the ΔeseC strain), EseE is freedup and becomes fully functional, leading to increased transcription of the escC-eseEoperon.
EseE mediates the inhibitory effect of EseC on biofilm formation. Finally, wetested if EseE would be essential for EseC to inhibit biofilm formation in E. piscicida. WTand ΔeseE, ΔeseC, ΔeseC ΔeseE, and ΔeseB mutant strains were cultured for 18 h and24 h, followed by the measurement of biofilm formation. While the ΔeseC strain formeda mature biofilm, the WT, ΔeseE, and ΔeseC ΔeseE strains only formed immature biofilmsat 18 h postsubculture (Fig. 5, left). At 24 h postsubculture, similar levels of maturebiofilm were formed by the WT and ΔeseC mutant strains, in contrast to immaturebiofilms formed by the ΔeseE and ΔeseC ΔeseE mutant strains (Fig. 5, right). The ΔeseBstrain showed no biofilm formation at either time point, as expected. Our results thusindicate that EseE mediates the inhibitory effect of EseC on biofilm formation at 18 hpostsubculture.
FIG 5 EseC inhibits biofilm formation through sequestering EseE. E. piscicida strains were subcultured in DMEM ina 24-well plate embedded with coverslips. At 18 h and 24 h postsubculture, bacteria settled onto the coverslipswere fixed and stained with 0.1% crystal violet. Quantification of dissolved crystal violet was performed to evaluatethe biofilm formation ability of the bacteria. ***, P � 0.001; *, P � 0.05; NS, not significant.
EseC Inhibits Biofilm Formation through Sequestering EseE Applied and Environmental Microbiology
April 2019 Volume 85 Issue 8 e02133-18 aem.asm.org 7
on January 24, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

DISCUSSION
Our previous study has demonstrated that the translocon protein EseB can assemblefilamentous appendages on the surface of E. piscicida, and the interactions betweenEseB filaments leads to autoaggregation and biofilm formation (17). In this study, weexplored the role of the other two translocon proteins, EseC and EseD, in autoaggre-gation and biofilm formation. We show that EseC, but not EseD, inhibits autoaggrega-tion. More importantly, EseC inhibits biofilm formation by sequestering EseE, a positiveregulator of the escC-eseE operon (23).
A previous study has suggested that EseC and EseD are required for autoaggrega-tion in E. piscicida (5). However, it should be noted that this conclusion was made basedupon eseC and eseD insertional mutants, which presumably had a polar effect on thefunction of downstream gene eseE, a critical regulator of the entire escC-eseE operon,including eseB. To confirm the role of EseC and EseD in autoaggregation, we createdin-frame deletion mutants of eseC or eseD as well as their complementation strains(ΔeseC/pJN-eseC and ΔeseD/pJN-eseD). Our results clearly demonstrate that EseD is notinvolved in autoaggregation, whereas EseC inhibits autoaggregation (Fig. 1).
EseC is homologous to enteropathogenic Escherichia coli (EPEC) EspD, Yersiniapseudotuberculosis YopB, and Salmonella Typhimurium SseC (5). We showed the loss ofEseC led to increased EseB/EseD secretion. This is quite different from the observationseen in EPEC. A ΔespD strain secretes decreased protein level of EspA, an EseBhomologue (35.2% similarity) (5, 25). Also, deletion of YopB or SseC does not affect thesecretion of EseD homologues YopD or SseD (5, 26, 27). Thus, the means by which EseCcontrols EseB/D secretion seems be unique to E. piscicida.
EseC and EseE appear to reciprocally regulate their function. While the deletion ofEseE leads to an accumulation of EseC (unable to be secreted into the supernatant) inthe bacterial cytosol (23), the deletion of eseC results in upregulated expression of eseEthat in turn augments the transcription of the escC-eseE operon. The secretion status ofEseC is also important for the production of EseE. When a 2HA tag was added at the Cterminus of EseC, EseC failed to be secreted and accumulated within the bacterial cell(Fig. 4A). This was accompanied by reduced production of EseE. Consistent with therole of EseE in promoting the transcription of the escC-eseE operon (including eseB andeseD), the loss of EseC led to increased production of intracellular EseB and EseD,whereas the secretion-incompetent EseC (i.e., provided by the ΔeseC/pACYC-eseC-2HAstrain) suppresses the production of intracellular EseB and EseD (Fig. 4A and B). Moreimportantly, the deletion of eseE in the ΔeseC mutant strain completely abolished itseffect on promoting EseB and EseD production (Fig. 4D). EseE is not only a chaperoneprotein that directly binds to EseC but also a regulator that promotes the transcriptionof the escC-eseE operon (17). These findings led us to propose that EseC suppresses thetranscription of the escC-eseE operon through sequestering EseE.
EseC and EseE also regulate biofilm formation in E. piscicida. As discussed earlier,EseB plays a key role in biofilm formation (17). The higher steady-state protein level ofEseB from the ΔeseC mutant strain helps E. piscicida cells attach to coverslips andconnect with each other, promoting biofilm formation. Therefore, biofilm formationoccurred faster and at an earlier stage (18 h postsubculture) in the ΔeseC mutant strainthan in the WT strain (Fig. 2B and 5 [left]). At 24 h postsubculture (stationary phase), theWT and the ΔeseC strains formed comparable levels of biofilms (Fig. 5, right), potentiallyresulting from the saturation of biofilm formation. Even so, we noticed much longerEseB filaments on the surface of the ΔeseC mutant strain than on the WT strain (Fig. 2B),indicating that EseC still inhibits the formation of EseB filaments at a later stage ofbiofilm formation.
In summary, our study has demonstrated that EseC suppresses EseB-mediatedbiofilm formation through sequestering EseE.
MATERIALS AND METHODSBacteria strains and culture conditions. Bacterial strains and plasmids used in this study are
described in Table 1. Edwardsiella piscicida PPD130/91 (2) and its derivative strains were grown statically
Liu et al. Applied and Environmental Microbiology
April 2019 Volume 85 Issue 8 e02133-18 aem.asm.org 8
on January 24, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

in tryptic soy broth (TSB; BD Biosciences) at 28°C. To induce the T3SS, E. piscicida strains were culturedstatically in Dulbecco’s modified Eagle medium (DMEM; Invitrogen) at 25°C under a 5% (vol/vol) CO2
atmosphere. When required, the medium was supplemented with the appropriate antibiotics at thefollowing concentrations: 12.5 �g/ml colistin (Col; Sigma), 15 �g/ml tetracycline (Tet; Amresco), 34 �g/mlchloramphenicol (Cm; Amresco), and 50 �g/ml gentamicin (Gm; Amresco).
Construction of deletion mutants and complementation plasmids. The eseD gene was deletedfrom the chromosome of E. piscicida PPD130/91 by sacB-based allelic exchange, as described previously(9, 28). The ΔeseC ΔeseE strain was constructed similarly. The eseD gene and its ribosome binding sitewere amplified with primers pJN-eseD-com-for and pJN-eseD-com-rev (Table 2) and then ligated into theEcoRI and XbaI restriction sites of pJN105 (29). The resultant plasmid, pJN-eseD, was verified by DNAsequencing and then transformed into the ΔeseD strain to create the complementation strain (ΔeseD/pJN-eseD). The ΔeseC/pJN-eseC strain was constructed similarly. The expression of EseC or EseD bycomplementation strains was induced by supplementing 10 mM L-arabinose (BioSharp, China) into theDMEM.
TABLE 1 Strains and plasmids used in this study
Strain or plasmid Description and/or genotypea Reference or source
BacteriaE. piscicida
PPD130/91 Wild type, Kms Colr Amps, LD50 � 105.0 2ΔeseB strain PPD130/91 with in-frame deletion of eseB 15ΔeseC strain PPD130/91 with in-frame deletion of eseC 23ΔeseD strain PPD130/91 with in-frame deletion of eseD This studyΔeseC/pJN-eseC strain ΔeseC mutant with pJN105-eseC This studyΔeseC/pACYC-eseC-2HA strain ΔeseC mutant with pACYC184-eseC-2HA This studyΔeseD/pJN-eseD strain ΔeseD mutant with pJN105-eseD This studyΔeseE strain PPD130/91 with in-frame deletion of eseE 15ΔeseC ΔeseE strain PPD130/91 with in-frame deletion of eseE and eseC This study
E. coliDH5� �-Complementation Stratagene
PlasmidspRE112 Suicide plasmid, pir dependent, Cmr, oriT oriV sacB 28pJN105 Arabinose-inducible gene expression vector, araC-PBAD, Gmr 29pJN-eseC pJN105 with wild-type eseC, Gmr This studypJN-eseD pJN105 with wild-type eseD, Gmr This studypACYC184 Tetr Cmr AmershampACYC-eseC-2HA pACYC184 with eseC-2HA, Tetr This studypFPV25 Vector with promoterless gfp gene 32pFPV�200–�1 bp �200 to �1 of escC gene inserted upstream of a promoterless gfp in pFPV25 This study
aKms, kanamycin sensitive; Colr colistin resistant; Amps, ampicillin sensitive; LD50, 50% lethal dose; Gmr, gentamicin resistant; Tetr, tetracycline resistant; Cmr,chloramphenicol resistant; Km: kanamycin.
TABLE 2 Oligonucleotides used in this study
Designation Nucleotide sequence
eseD-for GGGGTACCCTACATACCGCCGTGACCCAeseD-int-rev CGTCTGTTGATACTGGCGCAeseD-int-for GCCAGTATCAACAGACGCTGTCCGAATATCAGCACGGeseD-rev GGGGTACCGTGTTTCCCTCCCCTCCAGCpJN-eseD-com-for GGAATTCGCAGCCTGATGGApJN-eseD-com-rev GCTCTAGATCAGGACATGCGTACpJN-eseC-com-for GAATTCCTGGCAATACGAGTACTGGCTpJN-eseC-com-rev TCTAGATTAGGCGATATTGGCGGCGATpACYC-eseC-2HA-for CGAATTCTCTGGAGGAGCACGATCAGGpACYC-eseC-2HA-rev GAGTACTTACTAGAGGCTAGCATAATCAGGAACATCA
TACGGATAGGCGATATTGGCGGCGATGTeseB-qfor TGGATAAAGGGGAGCTCGACeseB-qrev ATTTGTTCATCTCGGCCAGCeseD-qfor CCAGTATCAACAGACGCAGCeseD-qrev TGCTAATGGTATGCGCCTCT16s-qfor ACTGAGACACGGTCCAGACTCCTAC16s-qrev TTAACGTTCACACCTTCCTCCCTACescC-200-for CGGAATTCCATGGCCCGAACACCCGAGAescC-1-rev GCTCTAGAAGTGAAACCTCCTATTGACT
EseC Inhibits Biofilm Formation through Sequestering EseE Applied and Environmental Microbiology
April 2019 Volume 85 Issue 8 e02133-18 aem.asm.org 9
on January 24, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

The eseC gene and its ribosome binding site sequences were also amplified with primers pACYC-eseC-2HA-for and pACYC-eseC-2HA-rev (Table 2) before being digested with restriction enzymes EcoRIand ScaI and inserted into pACYC184 to obtain the plasmid pACYC-eseC-2HA. After being verified by DNAsequencing, pACYC-eseC-2HA was transformed into the ΔeseC strain.
Autoaggregation assay. E. piscicida strains were inoculated into DMEM and maintained at 25°Cunder a 5% (vol/vol) CO2 atmosphere. After 24 h of culture, the bacteria were subcultured at a 1:200dilution in DMEM in transparent glass tubes. To visualize the autoaggregation of E. piscicida strains,bacterial cultures were photographed at 18 h and 24 h postsubculture.
Biofilm formation assay. E. piscicida strains were subcultured at a 1:200 dilution into a 24-well tissueculture plate embedded with coverslips. At 18 h and 24 h postsubculture, the culture supernatants wereremoved carefully, and the coverslips were gently rinsed three times with prewarmed phosphate-buffered saline (PBS) to dislodge the floating bacteria. The bacteria attached onto the coverslips werefixed with 4% paraformaldehyde (PFA) in PBS and stained with 0.1% crystal violet for 30 min before beingphotographed. Biofilms formed on the coverslips were then solubilized with 1% sodium dodecyl sulfate(SDS) buffer (200 �l per well). The OD630 values of the solubilized biofilms were read with an ELx 800microplate reader (BioTek, USA) and used as a quantitative measure of biofilm formation.
Immunofluorescent staining. E. piscicida strains were subcultured into 24-well tissue culture platesembedded with coverslips. At 18 h and 24 h postsubculture, the bacteria settled onto the coverslips werewashed and fixed in 4% PFA, followed by immunofluorescent (IF) staining with anti-EseB antibody.Briefly, mouse anti-EseB polyclonal antibody was used at a 1:200 dilution (17), and donkey anti-mouseIgG antibody (Alexa 488; Molecular Probes) was used at a 1:200 dilution. Images were taken using aconfocal laser scanning microscope (NOL-LSM 710; Carl-Zeiss).
Expression and secretion assays by immunoblotting. E. piscicida strains were subcultured at a1:200 dilution into DMEM and grown statically for 18 h at 25°C in a 5% (vol/vol) CO2 incubator. The totalbacterial proteins (TBP) and extracellular proteins (ECP) were collected, and then equal amounts of TBPand ECP were loaded onto the SDS-PAGE gel for immunoblotting. The membranes were probed withrabbit anti-EseC (1:1,000 dilution) (30), rabbit anti-EseB (1:1,000 dilution) (31), rabbit anti-EseD (1:1,000dilution) (30), rabbit anti-EseE (1:1,000 dilution) (23), rabbit anti-EseJ (1:1,000 dilution) (12), rabbitanti-EseG (1:1,000 dilution) (11), and rabbit anti-EvpC (1:5,000 dilution) (21) antibodies before incubatedwith horseradish peroxidase (HRP)-conjugated goat anti-rabbit IgG (1:2,000 dilution; Millipore, USA).Antigen-antibody complexes were detected with SuperSignal West Pico chemiluminescent substrate(Thermo) and imaged with a ChemiDoc MP imaging system (Bio-Rad, USA). The immunoblotting assaywas repeated at least three times. Densitometric analysis of immunoblotting bands was conducted usingthe Image Lab 4.1 software.
Quantitative real-time PCR. The transcript levels of eseB and eseD were quantified by real-time PCR.Total RNA was isolated from E. piscicida at 18 h postsubculture in DMEM, as described previously (23). Therelative transcript levels of different genes were determined by subtracting the cycle threshold (CT) of the16S rRNA gene, the calibrator, from that of the sample according to the following formula: ΔCT �CT(sample) – CT(calibrator). The relative transcript levels of target genes in the ΔeseC strain comparedwith those in the E. piscicida ΔeseC ΔeseE strain were calculated using the formula 2�ΔΔCT, where ΔΔCT �ΔCT(ΔeseC strain) – ΔCT(ΔeseC ΔeseE strain). The real-time PCR primers for eseB, eseD, and 16S rRNA werelisted in Table 2.
Confocal microscopy on E. piscicida. A DNA fragment (bp �200 to �1) upstream of escC wasamplified by PCR using primer pair escC-200-for/escC-1-rev. This fragment was inserted into the promot-erless gfp shuttle vector pFPV25 (32) to create pFPV�200 –�1. This construct (pFPV�200 –�1) wasintroduced into E. piscicida wild-type strain and the ΔeseC, ΔeseC ΔeseE, ΔesrB, and ΔesrC mutant strains.At 18 h postsubculture in DMEM, green fluorescent protein (GFP) signals present in each strain wereimaged using a confocal microscope. The same acquisition parameters were used for all images.
Statistical analysis. All data were analyzed using the t test in the Statistical Package for the SocialSciences (SPSS) and expressed as the mean � standard error of the mean (SEM). P values less than 0.05were considered statistically significant.
ACKNOWLEDGMENTSWe are very grateful to Richard William McLaughlin (General Studies, Gateway
Technical College, USA) for his generous help in proofreading the manuscript.This work was funded by the National Natural Science Foundation of China (NSFC;
grant 31572659), by China Agriculture Research System (grant CARS-46), and by theProject of State Key Laboratory of Freshwater Ecology and Biotechnology (grant2016FBZ04).
REFERENCES1. Ling SH, Wang XH, Xie L, Lim TM, Leung KY. 2000. Use of green
fluorescent protein (GFP) to study the invasion pathways of Edwardsiellatarda in in vivo and in vitro fish models. Microbiology 146:7–19. https://doi.org/10.1099/00221287-146-1-7.
2. Shao S, Lai QL, Liu Q, Wu HZ, Xiao JF, Shao ZZ, Wang QY, Zhang YX. 2015.Phylogenomics characterization of a highly virulent Edwardsiella strain
ET080813T encoding two distinct T3SS and three T6SS gene clusters:propose a novel species as Edwardsiella anguillarum sp. nov. Syst ApplMicrobiol 38:36 – 47. https://doi.org/10.1016/j.syapm.2014.10.008.
3. Miwa S, Mano N. 2000. Infection with Edwardsiella tarda causes hyper-throphy of liver cells in the Japanese flounder Paralichthys olivaceus. DisAquat Org 42:227–231. https://doi.org/10.3354/dao042227.
Liu et al. Applied and Environmental Microbiology
April 2019 Volume 85 Issue 8 e02133-18 aem.asm.org 10
on January 24, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

4. Padrós F, Zarza C, Dopazo L, Cuadrado M, Crespo S. 2006. Pathology ofEdwardsiella tarda infection in turbot, Scophthalmus maximus (L.). J FishDis 29:87–94. https://doi.org/10.1111/j.1365-2761.2006.00685.x.
5. Tan YP, Zheng J, Tung SL, Rosenshine I, Leung KY. 2005. Role of type IIIsecretion in Edwardsiella tarda virulence. Microbiology 151:2301–2313.https://doi.org/10.1099/mic.0.28005-0.
6. Okuda J, Takeuchi Y, Nakai T. 2014. Type III secretion system genes ofEdwardsiella tarda associated with intracellular replication and viru-lence in zebrafish. Dis Aquat Organ 111:31–39. https://doi.org/10.3354/dao02763.
7. Galán JE, Wolf-Watz H. 2006. Protein delivery into eukaryotic cells bytype III secretion machines. Nature 444:567–573. https://doi.org/10.1038/nature05272.
8. Sato H, Frank DW. 2011. Multi-functional characteristics of the Pseu-domonas aeruginosa type III needle-tip protein, PcrV; comparison toorthologs in other Gram-negative bacteria. Front Microbiol 2:142.https://doi.org/10.3389/fmicb.2011.00142.
9. Zheng J, Tung SL, Leung KY. 2005. Regulation of a type III and a putativesecretion system in Edwardsiella tarda by EsrC is under the control of atwo-component system, EsrA-EsrB. Infect Immun 73:4127– 4137. https://doi.org/10.1128/IAI.73.7.4127-4137.2005.
10. Wang B, Mo ZL, Mao YX, Zou YX, Xiao P, Li J, Yang JY, Ye XH, Leung KY,Zhang PJ. 2009. Investigation of EscA as a chaperone for the Edwardsiellatarda type III secretion system putative translocon component EseC.Microbiology 155:1260 –1271. https://doi.org/10.1099/mic.0.021865-0.
11. Xie HX, Yu HB, Zheng J, Nie P, Foster LJ, Mok YK, Finlay BB, Leung KY.2010. EseG, an effector of the type III secretion system of Edwardsiellatarda, triggers microtubule destabilization. Infect Immun 78:5011–5021.https://doi.org/10.1128/IAI.00152-10.
12. Xie HX, Lu JF, Zhou Y, Yi J, Yu XJ, Leung KY, Nie P. 2015. Identificationand functional characterization of the novel Edwardsiella tarda effectorEseJ. Infect Immun 83:1650 –1660. https://doi.org/10.1128/IAI.02566-14.
13. Cao H, Yang C, Quan S, Hu T, Zhang L, Zhang Y, Yang D, Liu Q. 2018.Novel T3SS effector EseK in Edwardsiella piscicida is chaperoned by EscHand EscS to express virulence. Cell Microbiol 20:e12790. https://doi.org/10.1111/cmi.12790.
14. Hou M, Chen R, Yang D, Núñez G, Wang Z, Wang Q, Zhang Y, Liu Q. 2017.Identification and functional characterization of EseH, a new effector ofthe type III secretion system of Edwardsiella piscicida. Cell Microbiol19:e12638. https://doi.org/10.1111/cmi.12638.
15. Zheng J, Li N, Tan YP, Sivaraman J, Mok YK, Mo ZL, Leung KY. 2007. EscCis a chaperone for the Edwardsiella tarda type III secretion systemputative translocon components EseB and EseD. Microbiology 153:1953–1962. https://doi.org/10.1099/mic.0.2006/004952-0.
16. Mikkelsen H, Bond NJ, Skindersoe ME, Givskov M, Lilley KS, Welch M.2009. Biofilms and type III secretion are not mutually exclusive inPseudomonas aeruginosa. Microbiology 155:687– 698. https://doi.org/10.1099/mic.0.025551-0.
17. Gao ZP, Nie P, Lu JF, Liu LY, Xiao TY, Liu W, Liu JS, Xie HX. 2015. Type IIIsecretion system translocon component EseB forms filaments on andmediates autoaggregation of and biofilm formation by Edwardsiellatarda. Appl Environ Microbiol 81:6078 – 6087. https://doi.org/10.1128/AEM.01254-15.
18. Tran CS, Rangel SM, Almblad H, Kierbel A, Givskov M, Tolker-Nielsen T,Hauser AR, Engel JN. 2014. The Pseudomonas aeruginosa type III translo-con is required for biofilm formation at the epithelial barrier. PLoSPathog 10:e1004479. https://doi.org/10.1371/journal.ppat.1004479.
19. Jennings ME, Quick LN, Ubol N, Shrom S, Dollahon N, Wilson JW. 2012.Characterization of Salmonella type III secretion hyper-activity whichresults in biofilm-like cell aggregation. PLoS One 7:e33080. https://doi.org/10.1371/journal.pone.0033080.
20. Zimaro T, Thomas L, Marondedze C, Sgro GG, Garofalo CG, Ficarra FA,Gehring C, Ottado J, Gottig N. 2014. The type III protein secretion systemcontributes to Xanthomonas citri subsp. citri biofilm formation. BMCMicrobiol 14:96. https://doi.org/10.1186/1471-2180-14-96.
21. Zheng J, Leung KY. 2007. Dissection of a type VI secretion system inEdwardsiella tarda. Mol Microbiol 66:1192–1206. https://doi.org/10.1111/j.1365-2958.2007.05993.x.
22. Hölzer SU, Hensel M. 2010. Functional dissection of translocon pro-teins of the Salmonella pathogenicity island 2-encoded type III se-cretion system. BMC Microbiol 10:104. https://doi.org/10.1186/1471-2180-10-104.
23. Yi J, Xiao SB, Zeng ZX, Lu JF, Liu LY, Laghari ZA, Nie P, Yu HB, Xie HX.2016. EseE of Edwardsiella tarda augments secretion of translocon pro-tein EseC and expression of the escC-eseE operon. Infect Immun 84:2336 –2344. https://doi.org/10.1128/IAI.00106-16.
24. Chakraborty S, Sivaraman J, Leung KY, Mok YK. 2011. Two-componentPhoB-PhoR regulatory system and ferric uptake regulator sense phos-phate and iron to control virulence genes in type III and VI secretionsystems of Edwardsiella tarda. J Biol Chem 286:39417–39430. https://doi.org/10.1074/jbc.M111.295188.
25. Knutton S, Rosenshine I, Pallen MJ, Nisan I, Neves BC, Bain C, Wolff C,Dougan G, Frankel G. 1998. A novel EspA-associated surface organelle ofenteropathogenic Escherichia coli involved in protein translocation intoepithelial cells. EMBO J 17:2166 –2176. https://doi.org/10.1093/emboj/17.8.2166.
26. Bröms JE, Forslund AL, Forsberg A, Francis MS. 2003. Dissection ofhomologous translocon operons reveals a distinct role for YopD intype III secretion by Yersinia pseudotuberculosis. Microbiology 149:2615–2626. https://doi.org/10.1099/mic.0.26322-0.
27. Cooper CA, Mulder DT, Allison SE, Pilar AV, Coombes BK. 2013. The SseCtranslocon component in Salmonella enterica serovar Typhimurium ischaperoned by SscA. BMC Microbiol 13:221. https://doi.org/10.1186/1471-2180-13-221.
28. Edwards RA, Keller LH, Schifferli DM. 1998. Improved allelic exchangevectors and their use to analyze 987P fimbria gene expression. Gene207:149 –157. https://doi.org/10.1016/S0378-1119(97)00619-7.
29. Khlebnikov A, Skaug T, Keasling JD. 2002. Modulation of gene expressionfrom the arabinose-inducible araBAD promoter. J Ind Microbiol Biotech-nol 29:34 –37. https://doi.org/10.1038/sj.jim.7000259.
30. Lu JF, Wang WN, Wang GL, Zhang H, Zhou Y, Gao ZP, Nie P, Xie HX. 2016.Edwardsiella tarda EscE (Orf13 protein) is a type III secretion system-secreted protein that is required for the injection of effectors, secretionof translocators, and pathogenesis in fish. Infect Immun 84:2–10. https://doi.org/10.1128/IAI.00986-15.
31. Zhou Y, Liu LY, He TT, Laghari ZA, Nie P, Gao Q, Xie HX. 2016. Edward-siella tarda EsaE (Orf19 protein) is required for the secretion of type IIIsubstrates, and pathogenesis in fish. Vet Microbiol 190:12–18. https://doi.org/10.1016/j.vetmic.2016.05.003.
32. Valdivia RH, Falkow S. 1996. Bacterial genetics by flow cytometry: rapidisolation of Salmonella Typhimurium acid-inducible promoters by differ-ential fluorescence induction. Mol Microbiol 22:367–378. https://doi.org/10.1046/j.1365-2958.1996.00120.x.
EseC Inhibits Biofilm Formation through Sequestering EseE Applied and Environmental Microbiology
April 2019 Volume 85 Issue 8 e02133-18 aem.asm.org 11
on January 24, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from