the cytoskeleton intermediary filament and beam theory · gaub,rief/ss 2005 bpm §2.2 19 - der...
TRANSCRIPT

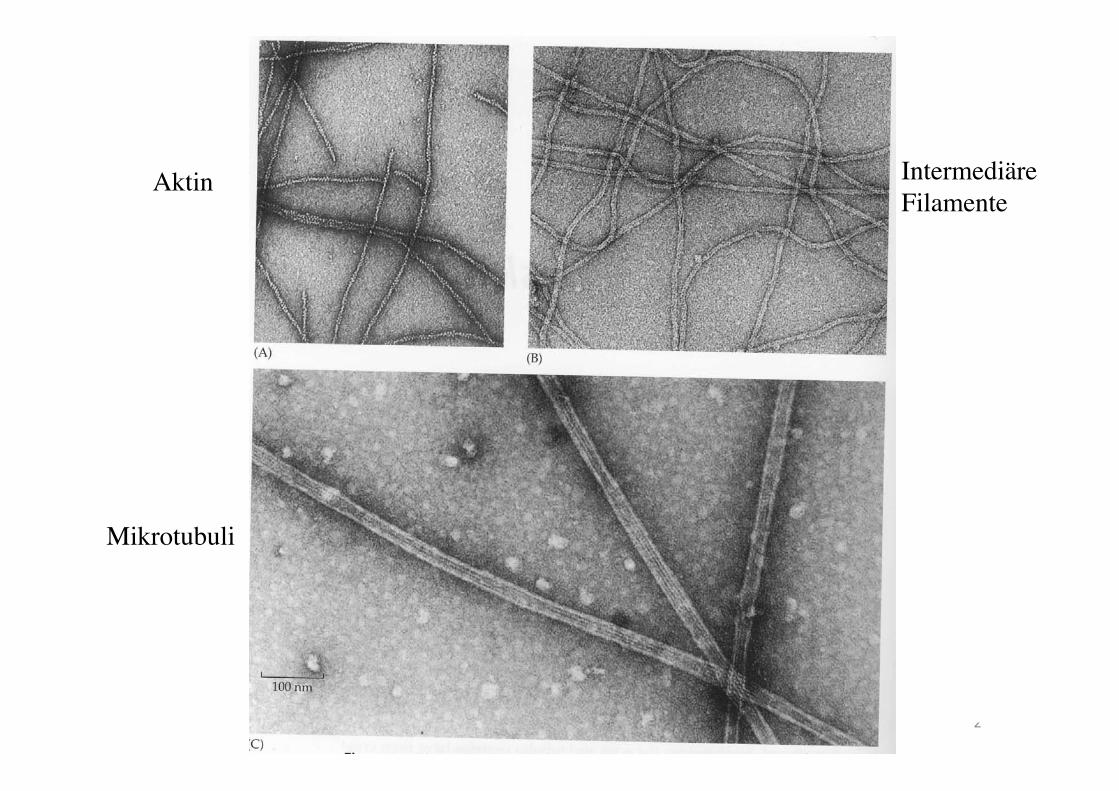
Actin
Intermediary filament
Microtubuli
The cytoskeleton and beam theory
BPM§2.2 2
Aktin Intermediäre Filamente
Mikrotubuli

Gaub,Rief/SS2005 BPM§2.2 3
actin.pdb
Aktin

BPM§2.2 4
Aktin
Gaub,Rief/SS2005 BPM§2.2 5

BPM§2.2 6
Mikrotubuli
http://genome-www.stanford.edu/group/botlab/tubulin/chime_tubulin_dimer.html

BPM§2.2 7

BPM§2.2 8
Mikrotubuli trennen die Chromosomen bei der Zellteilung
Fruchtfliege in der Mitose

BPM§2.2 9
Die coiled-coil ist die Basis-Struktur der Intermediären Filamente
http://www.bio.cmu.edu/Courses/03231/ProtStruc/2tma.htm

BPM§2.2 10

J. Käs et al. 1993

Analyse der Konturfluktuationen von Filamenten
J. Käs 1996 Nagashima 1980



Gaub,Rief/SS2005 BPM§2.2 15
Die Polymerisation einsträngiger Filamente Alle Filamente des Zytoskeletts sind erstaunlich lang. Das kann mit simplen einsträngigen Filamenten nicht erreicht werden.
+ K =A1[ ] ⋅ A1[ ]A2[ ]
+ K =A1[ ] ⋅ A2[ ]A3[ ]
...
K =A1[ ] ⋅ An[ ]An+1[ ]A1+ An↔koff
ko nAn+1
Diese Gleichung läßt sich mit folgendem Ansatz lösen:
An[ ] = K ⋅ e−nn0 K =
A1[ ] ⋅ An[ ]An+1[ ]
=Ke
−1n0 ⋅Ke
−nn0
Ke−n+1n0
= K qed
Die Längenverteilung der Filamente ist exponentiell. Es gibt immer mehr kurze Filamente als lange!
Beweis:

BPM§2.2 16
Durchschnittslänge der Filamente
Definition: an:=An[ ]K
Die mittlere Länge der Filamente, ohne die Monomere gerechnet, beträgt:
an = e−nn0
nav = n ⋅ pnn=2
∞
∑ pn ist die Wahrscheinlichkeit für das Vorkommen von Filamenten mit n Monomeren.
pn =an
ann=2
∞
∑nav =
1
ann=2
∞
∑n ⋅ an
n=2
∞
∑ =1
a1n
n=2
∞
∑n ⋅ a1
n
n=2
∞
∑
a1n
n=0
∞
∑ =1
1− a1a1n
n=2
∞
∑ =1
1− a1−1− a1 =
1− 1+ a1( ) 1− a1( )1− a1
=a12
1− a1
n ⋅ a1n
n=1
∞
∑ =a1
1− a1( )2 n ⋅ a1n
n=2
∞
∑ =a1
1− a1( )2− a1 nav = 1+
11− a1
=> an=a1n

BPM§2.2 17
atot = n ⋅ a1n
n=1
∞
∑ =a1
1− a1( )2 Gesamtzahl aller Monomereinheiten
a1= 1+12atot
−1atot
+1
4atot2
nav = 1+1
1− a1
≅1− 1atot
atot >>1
≅1+ atot
atot >>1
Wie lang wären demnach im Durchschnitt Aktinfilamente? [Atot]=200 µM, K=0,1 µM
Lösung des Problems: Vielsträngige Filamente!
nav < 50!

BPM§2.2 18
Die Polymerisation doppelsträngiger Filamente
+ K1=A1[ ] ⋅ A1[ ]A2
*[ ]
+ K2 =A1[ ] ⋅ A1[ ]A2
**[ ]
K =A1[ ] ⋅ A2[ ]A3[ ]
+
+ K =A1[ ] ⋅ An[ ]An+1[ ]
an =KK1e−nn0 a2
*=KK1a12 a2
**=KK2
a12
nav ≅K1K
AtotK
Abschätzung für Aktin mit K1=0.1 M, K= 0.1 µM, Atot = 200 µM ? nav < 1000 * 50!

Gaub,Rief/SS2005 BPM§2.2 19
- Der Trick der zweisträngigen Filamente besteht darin, daß 2 wesentlich verschiedene Gleichgewichtskonstanten existieren. Die Bindung zweier Monomere ist in der Regel sehr schwach, während die Bindung eines Monomers an ein bestehendes Filament stark ist (größere Kontaktfläche).
- Dadurch wird erreicht, daß nur sehr wenige Filamente überhaupt gestartet werden (Nukleationskeim) und die Monomere nur an schon bestehende Filamente angebaut werden. Das Abdecken freier Enden resultiert in maximalem Energiegewinn.
Polymerisation und Dimensionalität
- Die sehr breite exponentielle Längenverteilung von Filamenten hängt direkt mit ihrer Dimensionalität zusammen. Die Oberfläche (die Enden) eines eindimensionalen Objekts ändert sich nicht mit seiner Länge, deshalb ist die Anlagerung eines Monomers an ein kurzes Filament ebenso wahrscheinlich, wie an ein langes Filament.
- In 2 oder 3 Dimensionen wächst allerdings die Oberfläche eines Objekts mit dessen Volumen. Deshalb wird sich hier im Idealfall nur ein Riesenaggregat bilden.
- Aus genau derselben Ursache gibt es auch keine Phasenübergänge in 1d, weil die Oberfläche von Keimen nicht wächst.

BPM§2.2 20
a)b)
Dendritisches Wachstum von Zink bei der elektrolytischen Abscheidung http://apricot.polyu.edu.hk/~lam/dla/dla.html
Diffusionslimitierte Aggregation in 2 und 3 Dimensionen

BPM§2.2 21
Vielsträngige Filamente wachsen und schrumpfen nur über Monomeraddition oder -desorption an den Enden aber brechen und kombinieren nie.
Wachstum und Zerfall eines Filaments unter der Voraussetzung, daß nur Monomere addiert und subtrahiert werden:
dndt= kon A1[ ] − koff
Wachstum eines Filaments, wenn Multimere addiert werden.
dndt= kon A1[ ] + 2 ⋅ kon, 2 A2[ ] + ...+m ⋅ kon,m Am[ ] + ...
aber: Die Rotationsdiffusion macht die Reaktion zwischen zwei länglichen Objekten sehr viel schneller diffusionslimitiert, deshalb sind die kon,m sehr viel langsamer und nur Monomeraddition trägt bei.
Wenn kon,m gleich kon ist, dann weil [Atot] in der Regel >> [A1], wäre auch die Addition von Multimeren wichtig.
= kon Atot[ ]
€
dndt
= kon m Am[ ]m=1
∞∑

BPM§2.2 22
Dasselbe Argument gilt aus Gründen der Mikroreversibilität auch für den Bruch eines Filaments in der Mitte: Eine Bindung wird in der Mitte deswegen langsamer aufbrechen, weil durch die langsame Diffusion der Bruchstücke die Chance für ein Wiederbinden größer wird. Nicht so an den Enden.
Bruch in der Mitte Bruch am Ende
Zusätzlich wird bei vielsträngigen Filamenten die Wahrscheinlichkeit für einen Bindungsbruch in der Mitte dadurch drastisch verringert, daß mehr Bindungen für einen Bruch in der Mitte gebrochen werden müssen als für einen Bruch am Ende.

Gaub,Rief/SS2005 BPM§2.2 23
Weitere Eigenschaften vielsträngiger Filamente Es existiert eine kritische Monomerkonzentration unterhalb derer praktisch keine Polymere vorliegen. Oberhalb dieser Konzentration setzt Polymerwachstum ein.
dndt= konKc − koff =
!0Für die kritische Konzentration gilt: Kc =
koffkon
nav ≅K1K
AtotK

BPM§2.2 24
Weitere Eigenschaften vielsträngiger Filamente
K1=A1[ ] ⋅ A1[ ]A2
*[ ] K2 =A1[ ] ⋅ A1[ ]A2
**[ ]
Die Konzentration an Dimeren (Wachstumskeimen) ist sehr niedrig. Für [Atot]>Kc ist die freie Konzentration an Monomeren [A1]= Kc (siehe letzte Folie).
A2*[ ] = K 2
K1A2
**[ ] = K 2
K2für K1=K2=0,1 M und K=1 µM
A2*[ ] = A2
**[ ] = 0,01 nMDurch das Anbieten von Molekülen, die als Nukleationskeime fungieren können, in schon sehr geringen Konzentrationen kann die sonst unkontrolliert stattfindende Polymerisation gezielt an festgelegten Stellen gestartet werden. (z. B. arp2/3 Komplex)


BPM§2.2 26
Weitere Eigenschaften vielsträngiger Filamente Die Enden vielsträngiger Filamente sind immer glatt, nie “ausgefranst” . Die “ausgefranste” Konfiguration ist energetisch viel aufwendiger als die glatte.
Die mittlere Länge der Filamente steigt über der kritischen Konzentration scharf an. Somit können Proteine, die Aktinmonomere “wegfangen” die Polymerlänge regeln.