the control of the development of xenopus laevis...
TRANSCRIPT

/ . Embryol. exp. Morph. Vol. 18, 3, pp. 359-87, December 1967 3 5 9Printed in Great Britain
The control ofcell number in the lumbar ventral horns during
the development of Xenopus laevis tadpoles
By M. C. PRESTIGE 1
The developing limb has been amputated by many workers in several speciesand in each case the number of surviving motor neurones on the side of theoperation was less than normal. This may be observed among the mammals(e.g. Barron, 1945), birds (e.g. Hamburger, 1934), urodeles (e.g. Stultz, 1942),and anurans (e.g. May, 1930). The loss of motor neurones after amputation inadults appears to have been first noticed by Vulpian (1868) and Johnson &Clarke (1868). The early evidence is reviewed by Sherrington (1893) and thelater by Piatt (1948). The control that the developing leg has over proliferation,migration, maintenance and degeneration of ventral horn cells has been mostcompletely analysed in the chick, notably by Hamburger (1934, 1939, 1958),Hamburger & Keefe (1944), Bueker (1943, 1944, 1945a), Barron (1946, 1948),Mottet (1952) and Mottet & Barron (1954). Less is known about this in Anura.May (1930, 1933) has recorded that in Discoglossus motor area hyperplasia canbe produced by supernumerary limbs, and that the lateral motor horn (ventralhorn) can be suppressed by replacement of the limb area with skin grafts. Bueker(19456) found a frog {Rand) with three functional hind legs on one side; thishad a hyperplasia of 22-7 % in the ventral horn. Beaudoin (1955) ablated theearly limb-bud in Rana pipiens and found a terminal hypoplasia of 82 % inthe ventral horn. Counts at intermediate stages were interpreted by him as indi-cating that the operation had delayed differentiation and the normal loss of cellson that side. In Bufo vulgaris, Perri (1956a) removed the hind-limb area at tail-bud stages, and found that the ventral horn developed normally up to digitstages, when it regressed. The same happened when the isolated spinal cordwas transplanted (Perri, 19566). Three out of six animals with motile super-numerary limbs had hyperplastic spinal cords on the side of graft; this was notseen in any of the cases with non-motile grafts (Perri, 1957). Supernumerarylimbs only became motile when innervated by limb plexus fibres, and neverinduced formation of new ventral horns in trunk regions. Perri therefore con-cluded that for the full development of the ventral horn, both regional intrinsicand also extrinsic limb factors were essential, the former necessarily actingbefore the latter.
1 Author's address: The Department of Zoology, University of Bristol, Bristol, U.K.

360 M. C. PRESTIGE
Further evidence has been obtained from the comprehensive series of experi-ments by Hughes, using batches of Eleutherodactylus embryos to investigate theconditions for achieving innervation of grafted limbs. Amputation of the limbat early stages (5-8 days development) causes a rapid degeneration of ventralhorn cells (Hughes, 1962). This is at a time when nerve fibres have alreadyentered the limb. 'A limb grafted in place of the amputated member has littleeffect on halting the loss of cells, and . . . the main effect of the graft is in pro-moting a partial regeneration of the ventral horn through the outward migrationof new cells from the mantle layer' (Hughes, 1962). Amputation at still laterstages (Hughes, 1964) causes chromatolysis and an excess of cells to develop onthe operated side; this is succeeded by a final cell loss. The latter result was alsoobtained by Palladini & Alfei (1966) in Bufo bufo.
In Xenopus, removal of the early limb-bud before fibres enter (Hughes &Tschumi, 1958) by obstructive isolation, ablation or chemical means, does notaffect the assembly and initial differentiation of motor neuroblasts in animalsfixed before stage 53 (limb at palette stage); but after this stage the ventral hornwas absent. In individuals amputated at these later stages when the developingneuroblasts require the presence of the limb (stages 55-56), the operation causesthe immediate degeneration of some of the motor cells (Hughes, 1961). Produc-tion of an experimental hyperplasia has not been attempted in this species.
Thus in Anura it seems that the limb does not affect the initial development ofthe ventral horn, but later becomes essential for its further maintenance. Afterfibres have grown into the limb, amputation causes loss of cells; and later stillcauses chromatolysis. The leg appears to have some controlling influence overthe further differentiation of new ventral horn cells, and also their ultimatemaintenance.
One factor that has not received much attention is histogenetic degeneration(Gliicksmann, 1951)—that is, cell deaths occurring normally in the differentia-tion of functioning organs. These have been observed (Romanes, 1946) andcounted in developing motor cell columns of mouse (Harris, 1965) and thelumbar ventral horn of Xenopus (Hughes, 1961); they have also been seen in themotor columns of undisturbed chick embryos (Collin, 1906; Hamburger &Levi-Montalcini, 1949; Hughes, 1955); and counted on the contralateral sideof chick embryos after amputation of the limb-bud (Hamburger, 1958). De-generating motor cells in limb regions have also been observed in other Anura,among them Bufo (Dr A. Hughes, personal communication), Hyla (Hughes,1963), Eleutherodactylus (Hughes, 1962), and 'few' in Rana (Race & Terry,1965); also in the lizard, Lacerta (Dr A. Bellairs & Dr A. Hughes, personalcommunication). In Xenopus, the species for which we have most information,Hughes (1961) calculated that the number of degenerating cells was greatly inexcess of that required to account for the decline in cell numbers and thereforeconcluded turnover was taking place in the developing ventral horn. The samemay be true of the chick, for Hamburger (1958) observed degenerating cells (on

Cell number in ventral horns 361
the unoperated side in his experimental material) at a time when the number ofcells was not declining; thus new cells must have been entering.
Another factor that has been neglected is the later influence of the peripheryupon the ventral horn. Nearly all the anuran experiments that have been donewere performed on the early limb-bud before the fibres have grown in. Apartfrom isolated observations, only in Eleutherodactylus and Xenopus has the legbeen amputated at later stages (Hughes, 1961, 1962, 1964). In the formerspecies, however, it is very difficult to count degenerating cells, as sites arenumerous and close together. In slowly developing larvae, such as Xenopus,the sites are almost always separate and distinguishable.
At the present, we do not know what causes ventral horn cells to degenerateduring normal development, nor is it known how this process is controlled. It isthe purpose of this paper to investigate the control of the periphery over thenumber of lumbar ventral horn cells in Xenopus by amputating the hind legat representative stages throughout development, by observing the consequentchanges in numbers of living and degenerating cells, and by calculating thechanges in rate of 'production' of new cells by immigration. In this way, theresults of acute and chronic decrease of the periphery can be compared at eachstage. This has already been done for the dorsal root ganglia in a previous paper,to which this is a companion (Prestige, 1967).
MATERIAL AND METHODS
Larvae of Xenopus were reared from eggs obtained from adults injected withgonadotrophins (Nieuwkoop & Faber, 1956). They were kept at temperaturesbetween 20 and 23 °C. One leg of tadpoles and juveniles was amputated as highin the thigh as possible, using scissors or forceps under M.S. 222 anaesthesia(1:4000). The limb-bud of younger tadpoles was ablated with forceps as closeto the body wall as possible. The other side of the animal served as a control.Provided the water was kept clean, the tadpoles survived well until metamorphicclimax; at this stage, they normally begin to swim with their legs, and ceaseswimming with their tails. With only one leg, they drown in deep water. If thelevel is kept so low that they can expose their mouths above water withoutswimming, they survive. The water level can be raised again over the next weekor two as they learn to swim with one leg. This procedure must also be followedafter amputating the leg of a juvenile for the same reason. The animals werekept in tap water at all times, without antibiotics.
The lumbar region of selected specimens was fixed in half-strength Bouin;the body length and hind-limb length were measured; serial paraffin sectionswere cut at 8 fi or 10 fi transversely and stained with haematoxylin and eosin.Some were silver-impregnated (Palmgren, 1948). The lumbar ventral hornextended through 130-250 sections. The motor neurones therein were identifiedboth by position and appearance. While the largest of these are clearly distinct

362 M. C. PRESTIGE
from mantle layer cells, at the other extreme are some neuroblasts which aresimilar to them in size and appearance, especially at either end of the column.Only those cells were reckoned as ventral horn cells in which the nucleus wasconsidered to be larger and paler than those of the adjacent mantle layer neuro-blasts. These criteria are the same as those adopted and illustrated by Hughes(1961). In each specimen, the neurones of every third section throughout theseries were enumerated; only those cells were counted in which the nucleolus waspresent. Ventral horn neurones have only one nucleolus and this body is smallcompared with the section thickness. Moreover, as Jones (1937) has pointed out,nucleoli can be seen on occasion to have been dislodged from the nucleus in thedirection that the microtome knife passes through the tissue, suggesting thatnucleoli are less frequently bisected by sectioning than might be expected on thebasis of their size and numbers, and that instead they may be displaced wholeinto one or other of the adjacent sections. Counts of Palmgren and haema-toxylin-stained material did not differ. These arguments reduce the magnitudeof a correction factor to an uncertain value, and the counts therefore have notbeen corrected.
Degenerating neurones were identified by the presence of a pycnotic nucleuswith basophilic cytoplasm, as described by Hughes (1961). The degeneratingsite was counted if the nucleolar mass, or its residue, was contained in thesection; in cases of doubt, reference to the adjacent section could be made. Ithas been assumed that changes in the degenerating cell count reflect changes inthe rate of degeneration.
Neurones in chromatolysis were classed as living cells, not as degeneratingcells, though they were counted separately as well. Whereas in the dorsal rootganglia the nuclei are already eccentric, in the ventral horn the nucleus isnormally centrally placed in the cytoplasm, and its lateral protrusion forms anexcellent criterion for the identification of chromatolytic cells. The othercriterion used was the homogeneous character of the cytoplasm, like 'groundglass', and the absence of the fine lines of basophilic material that in Xenopusform the Nissl substance. Larger, wider streaks of basophilia at the cell surfaceremain or become intensified, and have to be ignored. The swelling of the cyto-plasm per se is not a good criterion as the cells are of different sizes anyway.Later, during the recovery phase, the characteristic dense ring of basophiliaaround the nucleus which may extend towards the poles of the cells is seen;in Palmgren preparations, this area is not argyrophilic. Thus the number ofcells that react can be counted at two stages of the process: in the chroma-tolytic phase and in the perinuclear basophilic phase. Such cells are not usuallyseen in unoperated tadpoles, nor on the contralateral side in the amputatedones.
Counts of chromatolytic cells are subject to two errors: first, there are cellswithin the ventral horn which are ranked in the total counts with the more mature

Cell number in ventral horns 363
ones, but do not possess sufficient cytoplasm to determine whether or not theyare undergoing chromatolysis; secondly, there are some large neurones whichdisplay some of the characteristics of chromatolysis, but not sufficiently to beidentified with certainty. Identification is easiest in haematoxylin preparationsin which no differentiation with acid alcohol has been attempted. A furthersource of variation stems from ignorance of the number of ventral horn cellsin the tadpoles at the time of the operation; only the number at fixation can becounted.
The counts of living cells in the Tables have been rounded to the nearest 10and of degenerating cells to the nearest 5.
The sampling procedure adopted was that every third section throughoutthe series was counted. Total sample counts of between 100 and 2000 living cellswere recorded: for these, the 95 % confidence limits due to random samplingare ± 20 % and ± 4-5 % respectively; the size of experimental changes inducedwas greater than this. For degenerating cells, sample counts of between 0 and80 were recorded: as estimates of the population mean, the lowest of these, iftaken from only one individual, may be as much as 100 % in error. However,the error may be decreased by pooling results from more than one animal, andthis procedure has been followed. In addition to the usual significance tests,rank and sign tests have been used because of their great power and simplicity(Siegel, 1956).
In selecting animals for an experiment, it was found that great care in thematching of the tadpoles made subsequent analysis of the results easier. It wasbest to choose examples from the same batch of eggs, reared in the same bathand with similar sized and shaped legs. Body length is not a good stage criterion.The animals were staged according to Nieuwkoop & Faber (1956). The relevantstages are here summarized: stage 49, limb bud round; stage 53, limb at palettestage—degenerating cells are first seen in ventral horn; stage 54, movementbegins in the limbs; stage 58, forelimbs emerge from pouch; stage 61, taildegeneration starts; stage 66, juvenile—no more degenerating cells in ventralhorn.
OBSERVATIONS
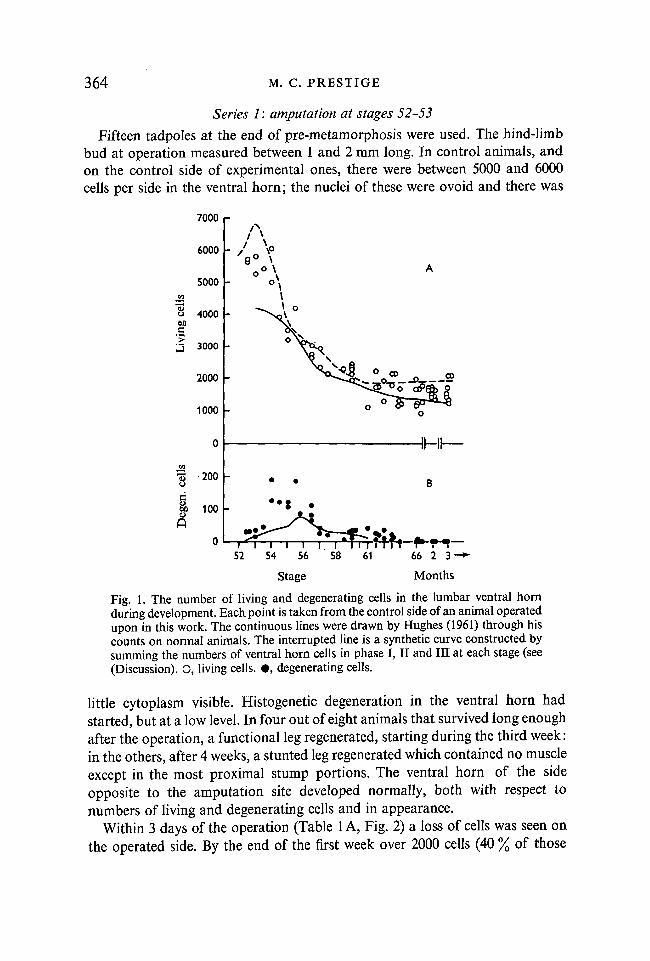
Limbs were amputated at stages 52-53, 54, 57, 61 or 66. The observations ineach series are separately described. The ventral horns of control animals and ofthe unoperated sides in experimental animals contained up to 50 % more cells thancomparable ones at the same stage in Hughes' (1961) study. The data from thepresent work are plotted in Fig. 1A (open circles) against the line drawn byHughes through his data (continuous line). In Fig. 1B (filled circles), the corres-ponding data on the number of degenerating cells are similarly plotted. Thepeak in number of degenerating cells occurs earlier and to a greater extent thanin Hughes' work.
23 JEEM l8
364 M. C. PRESTIGE
Series 1: amputation at stages 52-53
Fifteen tadpoles at the end of pre-metamorphosis were used. The hind-limbbud at operation measured between 1 and 2 mm long. In control animals, andon the control side of experimental ones, there were between 5000 and 6000cells per side in the ventral horn; the nuclei of these were ovoid and there was
7000
6000
5000
8 400000
3 3000
2000
1000
8 200
-IHI—
52 61
Stage
66 2 3 "
Months
Fig. 1. The number of living and degenerating cells in the lumbar ventral hornduring development. Each point is taken from the control side of an animal operatedupon in this work. The continuous lines were drawn by Hughes (1961) through hiscounts on normal animals. The interrupted line is a synthetic curve constructed bysumming the numbers of ventral horn cells in phase I, II and III at each stage (see(Discussion). O, living cells. • , degenerating cells.
little cytoplasm visible. Histogenetic degeneration in the ventral horn hadstarted, but at a low level. In four out of eight animals that survived long enoughafter the operation, a functional leg regenerated, starting during the third week:in the others, after 4 weeks, a stunted leg regenerated which contained no muscleexcept in the most proximal stump portions. The ventral horn of the sideopposite to the amputation site developed normally, both with respect tonumbers of living and degenerating cells and in appearance.
Within 3 days of the operation (Table 1A, Fig. 2) a loss of cells was seen onthe operated side. By the end of the first week over 2000 cells (40 % of those

Cell number in ventral horns 365
Table 1. The effect of amputation of the limb-bud at stages 52-53on the development of the ventral horn
(C = control; A = amputated.)
Dateof
fixation
17.x.
20.x.
24.x.
29.x.
9. xi.
16. xi.
25. xi.
20. xii.
Bodylength(mm)
Limblength(mm)
A. Operation
38
36
46
55
73
25
—.
•—
Operation4. xi.
10. xi.
17. xi.
26. xi.
15. xi.
16. xi.
17. xi.
61
68
32
—
39
32
41
1-2
1-6
3-4
5
14
22
26
25
Daysafter r
Stage amputation
performed 1
No. of cells
living
17. x. 66: those inunsuccessful
52-53
53
54
55-56
58-59-60
64
66
66
0
3
7
12
23
30
39
64
or
CA
CA
CA
CA
CA
CA
CA
CA
i performed 17. x. 66: those in7
16
23
23
56-7
58-59-60
64
66
B. Preliminary <2
2
2-2
53-54
53
54
18
24
31
40
CA
CA
CA
CA
experimeni3
4
5
CA
CA
CA
degen.Remarks on amount
of regeneration
which regeneration wasnot present564057205270467060503700418015002250630
1730320
1970350
1670110
which26201280230016902110
870
17601330
3030
1570
19055
185100
3520
105
50
00
—
—
—
Ilium not present
Pelvic girdle complete butfemur absent
Ilium + femur present:only muscles over hip-joint.
Foot regenerated close tobody: only reduced iliacmuscles present
Complete leg skeletonregenerated: some re-duced hip-joint musclespresent
regeneration was successful6540
3520
2010
00
: performed 12.541033805790494050102720
4520
3510
12555
Regenerate 2-6 mm:= stage 54
Regenerate 8 mm:= stage 56-57
Regenerate 17 mm:= stage 61, thigh flexorsreduced
Regenerate 20 mm:= stage 63
xi. 65—
—
—
23-2

366 M. C. PRESTIGE
initially present) had degenerated. In the animals in which a functional leg didnot regenerate, this large relative deficit was not maintained after 12 days; thenumber of cells in the ventral horn fell smoothly but less steeply on the operatedthan on the control side, so that the deficit at metamorphosis was only about1500 cells (Fig. 2B). This, however, represents a mean cell loss of 86 % in the
I
2 *3 CO
CO CO B
S -8° OCO Q>
3
XW
6000 \-
60•S 4000
2000 -
I<•
-
p
A
c1 .
I 1
<j> (
'1
DaysI I
10 20 30i I I l I I
40 64
52 54 56 58 63 66 Stage
1000
2000 L-
+0
100
Fig. 2. Amputation of the leg at stage 53. A, Numbers of living cells in the lumbarventral horn on control side (open circles) and on operated side (attached horizontalbar); those animals in which regeneration was successful are indicated by filledcircles. B, The differences in the number of living cells between the two sides.C, The differences in the number of degenerating cells between the two sides.
three juveniles. In the chick, Hamburger (1934) found a linear relation betweenthe amount of muscle lost after amputation of the limb, and the deficit in thenumber of motor neurones. If this holds also for Xenopus, then it seems thatthe operation removed around 86 % of the limb tissue.
The number of degenerating cells (Fig. 2C) observed on the operated sidewas between one-third and two-thirds that of the control side at all times after

Cell number in ventral horns 367
3 days (sign test, six pairs, P = 0-008), but at 3 days there was a large excessof degenerating cells. It seems that amputation at this stage, as Hughes (1961)found at the later stages 55-56, causes an early cell loss within the ventral hornand an excess of degenerating cells; this lasts less than a week, and in the pre-liminary experiment (Table 1B) must have lasted less than 3 days. Hughes &Tschumi (1958) showed that removal of the early limb bud did not affect thedifferentiation of the ventral horn prior to this stage, but caused its delayed dis-appearance at about this time. It is possible that in the experiment in which theexcess degeneration had ceased by the third day, the tadpoles were a few daysolder, while in the main experiment there was a delay of the same type as Hughes& Tschumi (1958) observed.
This explanation is supported by examination of the longitudinal pattern ofdegeneration and cell loss in the ventral horn. Dr A. Hughes (personal communi-cation) has recently found evidence that the posterior portion of the ventralhorn develops about a stage behind more anterior parts. On the control side,at stages 52-54 the number of histogenetic degenerating cells is increasing(Hughes, 1961); in the tadpoles of the present work the anterior two-thirds ofthe column always had many more than twice the number in the posterior third.The extent of cell loss after the operation follows a similar pattern, in that it isseen to have occurred earlier in the anterior two-thirds than elsewhere. Forexample, in animal 15. xi of Table IB, 37 % of the cells have been lost and thetotal number of degenerating cells on the operated side (20) was depressedbelow the control side (45), as in all other later tadpoles, yet at the posteriorend on the operated side the ventral horn showed no cell deficit and a localexcess of degenerating cells, compared to the unoperated side. It was only thisregion on the control side that showed no histogenetic degeneration. Thus al-though the excess degeneration due to the operation had ceased by the thirdday over most of the ventral horn, in the posterior portion it can barely havestarted.
These observations indicate that in the ventral horn the capacity to degenerateafter amputation of the limb also develops earlier in its anterior two-thirds, andat the same time as the capacity for histogenetic degeneration, to a very closeapproximation.
Effect of successful regeneration after the operation
The earliest signs of successful regeneration were recognized at 18 days, whenthe regenerate was a stage 54 leg, three stages behind the remainder of the tad-pole. This represents a difference of about 12 days and was maintained there-after. The ventral horn on the operated side (Table 1) was apparently almosttotally unaffected by the regenerating leg before this; that is, the numbers ofliving and degenerating cells were similar to those expected if there had been noregeneration. Perri (1956a) and Hughes & Tschumi (1958) showed that the

368 M. C. PRESTIGE
normally developing limb does not affect the ventral horn before stage 53, andthis is therefore also true of the regenerating limb.
After this time the regenerating leg (Fig. 2 A) maintained the number ofventral horn cells at about the same level, so that the further decline was arrested.At metamorphosis, the number of ventral horn cells for the regenerate was about1000, compared with about 300 for the unregenerate leg and about 1700 for acontrol leg. Many of these ventral horn cells for the regenerate were as large asthose on the control side. The leg in these circumstances becomes innervated(judged by sensitivity and motility) with a reduced set of ventral horn cells. Theregenerating limb, however, does not cause the ventral horn cell number todecrease, as in normal development.
Since the number of ventral horn cells did not decline after 18 days in thesecircumstances, and since cell degeneration continued, turnover of the ventralhorn cells must have been taking place while the regenerating leg was beinginnervated. Hughes (1961) calculated that a similar process of ventral horn cellturnover takes place during normal innervation of the leg.
Since the rate of degeneration on the operated side was similar in both re-generating and non-regenerating tadpoles (Table 1), the absence of a decline incell numbers after day 18 due to the regenerate must represent an increasedimmigration of cells into the ventral horn. Thus a regenerating leg inducesa further influx of neuroblasts from the mantle layer. This is similar to the effectof a replacing graft of a limb after its amputation at comparable stages inEleutherodactylus (Hughes, 1962).
Series II: amputation at stage 54
Tadpoles at these stages have legs between 2-5 and 4 mm long, which perform'flare' movements (Hughes & Prestige, 1967). They are in the first half of pro-metamorphosis. Cell degeneration and turnover are at their peak level withinthe ventral horn (Hughes, 1961). It is important to emphasize that numbers ofcells in the ventral horn are declining steeply at these stages, and thus that obser-vations on relative differences between the sides can only be made against ashifting base-line. In the above work, Hughes amputated the legs of tadpoleshigh in the thigh at stages 55-56. He found that within 3 days, many cells werelost from the ipsilateral ventral horn, and this was accompanied by an excessof degenerating cells. Equilibrium in number of ventral horn cells was restoredsome time after the first week, and peristed for a varying time (up to two furtherweeks) during which there was on some occasions an actual excess of cells onthe operated side. Ultimately there was a second and final loss of cells, so thatthe ventral horn at metamorphosis was reduced to about a third of its normalsize (38 %).
Hughes (1961) found that no regeneration took place at these stages. Thegreatersize of the residual ventral horn, when compared with those from series I,

Cell number in ventral horns 369
reflects the sparing of the iliac musculature at the operation. In series I, it waspossible to remove nearly all the tissue of the pelvic girdle as well.
In the present work, the legs of five tadpoles were amputated at stage 54(Table 2). The results show that during the first 7 days the ventral horn behavesas at stages 55-56, though the degree of initial cell loss is much greater. The tad-poles after 3, 5 and 7 days also show the same depression of the rate of degenera-tion that was seen in series I (Mann-Whitney rank test, P = 005). No chromato-lytic cells were seen. The major qualitative difference between the events follow-ing amputation at stages 52-53, and those following amputation at stages 55-56,
Table 2. The effect of amputation of the leg below the hip-joint at stage 54on the development of the ventral horn {operation performed on 18. xi. 65)
(C = control; A = amputated)
Date offixation
19. xi:
20. xi.
21. xi.
23. xi.
25. xi.
Bodylength(mm)
—
45
43
50
50
Leglength(mm)
—
4
3-5
5
6
Stage
55
55
54-55
56
56-57
Daysafter r
amputation
1
2
3
5
7
CA
CA
CA
CA
CA
No.
Living
31802600
35302720
39002190
31101670
30101890
of cells
Degen.
105235
115115
11555
8555
11055
is that in the former there is only the early period of relative cell loss, while inthe latter series, a second loss later ensues. After amputation at stages 52-53there is no evidence of renewed or second loss of cells during later development,neither as a late increase in the deficit of living cells (Fig. 2B), nor as a lateexcess of degenerating cells (Fig. 2C).
Series III: amputation at stage 57
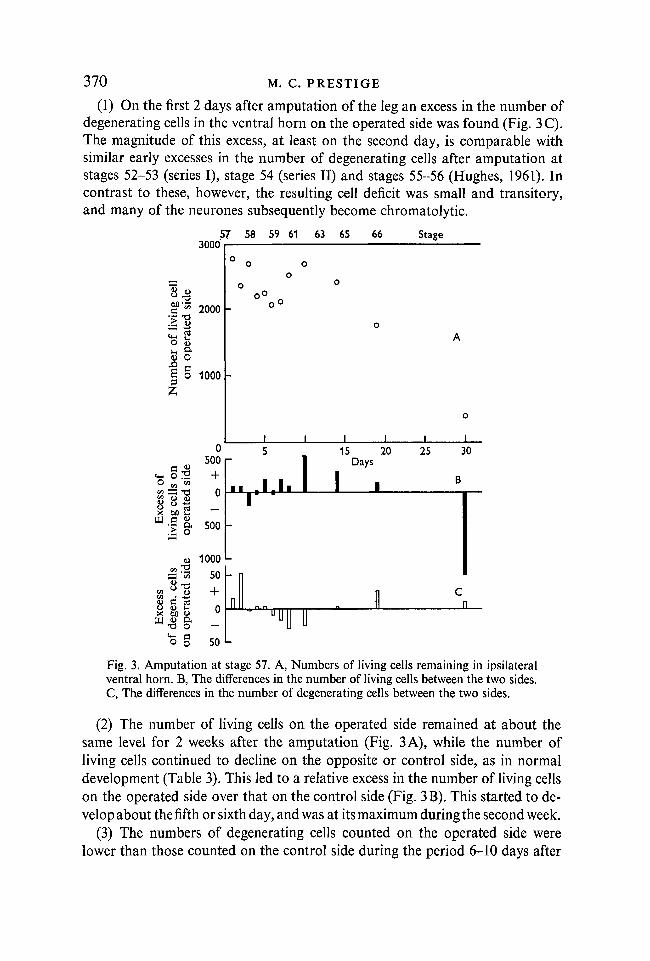
Twelve tadpoles were used. These animals were at the stage just before theforelimb emerges from the pouch. The hind legs were between 7 and 9 mm long,and at rest, the thighs were drawn up to protrude laterally; the tadpole swamby using its tail, while the legs were held extended together against the fin(Hughes & Prestige, 1967). In the ventral horn, the period of steep decline andits associated peak in counts of degenerating cells are normally just over: therewere between 2000 and 3000 cells on each side. The animals of Fig. 3 and Table 3show the following features:
370 M. C. PRESTIGE
(1) On the first 2 days after amputation of the leg an excess in the number ofdegenerating cells in the ventral horn on the operated side was found (Fig. 3C).The magnitude of this excess, at least on the second day, is comparable withsimilar early excesses in the number of degenerating cells after amputation atstages 52-53 (series I), stage 54 (series II) and stages 55-56 (Hughes, 1961). Incontrast to these, however, the resulting cell deficit was small and transitory,and many of the neurones subsequently become chromatolytic.
300057 58 59 61 63 65 66 Stage
8.§
u ft<a O
z
2000
1000
0500
o oo
x toa 500
8-a
1000
50
o —
o § 50
I I
15 20Days
25 30
Fig. 3. Amputation at stage 57. A, Numbers of living cells remaining in ipsilateralventral horn. B, The differences in the number of living cells between the two sides.C, The differences in the number of degenerating cells between the two sides.
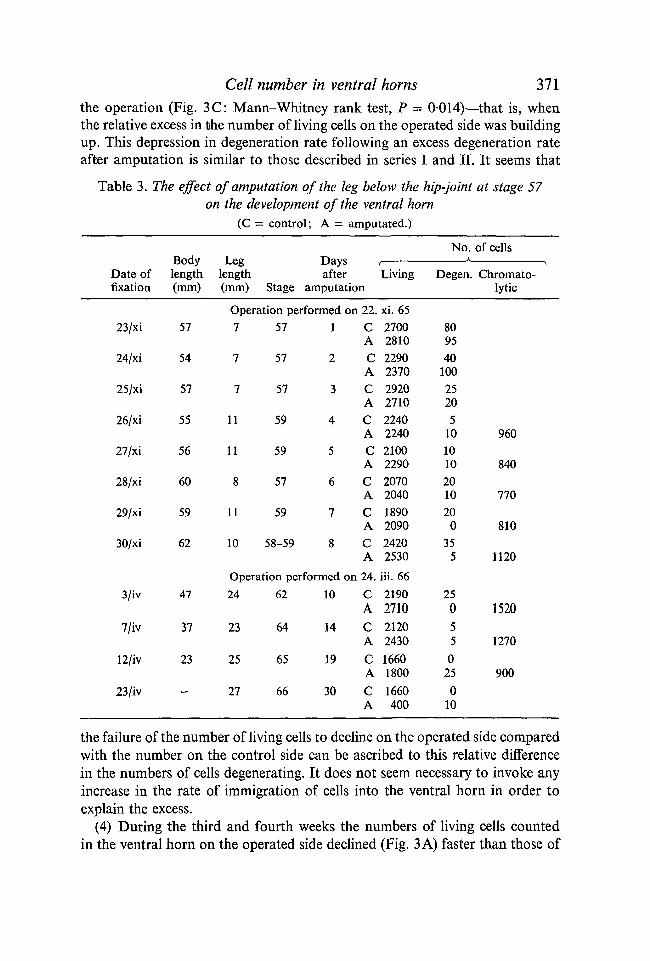
(2) The number of living cells on the operated side remained at about thesame level for 2 weeks after the amputation (Fig. 3 A), while the number ofliving cells continued to decline on the opposite or control side, as in normaldevelopment (Table 3). This led to a relative excess in the number of living cellson the operated side over that on the control side (Fig. 3B). This started to de-velop about the fifth or sixth day, and was at its maximum during the second week.
(3) The numbers of degenerating cells counted on the operated side werelower than those counted on the control side during the period 6-10 days after
Cell number in ventral horns 371
the operation (Fig. 3C: Mann-Whitney rank test, P = 0014)—that is, whenthe relative excess in the number of living cells on the operated side was buildingup. This depression in degeneration rate following an excess degeneration rateafter amputation is similar to those described in series I and II. It seems that
Table 3. The effect of amputation of the leg below the hip-joint at stage 57on the development of the ventral horn
(C = control; A = amputated.)
Date offixation
23/xi
24/xi
25/xi
26/xi
27/xi
28/xi
29/xi
30/xi
3/iv
7/iv
12/iv
23/iv
Bodylength(mm)
57
54
57
55
56
60
59
62
47
37
23
—
Leglength(mm)
Daysafter
Stage amputation
Operation performed on7
7
7
11
11
8
11
10
57
57
57
59
59
57
59
58-59
1
2
3
4
5
6
7
8
Operation performed on24
23
25
27
62
64
65
66
10
14
19
30
22.CACACACACACACACA24.
CACACACA
(Living
xi. 652700281022902370292027102240224021002290207020401890209024202530
iii. 662190271021202430166018001660400
No.
Degen.
809540
10025205
101010201020
035
5
250550
250
10
of cells
Chromato-lytic
960
840
770
810
1120
1520
1270
900
the failure of the number of living cells to decline on the operated side comparedwith the number on the control side can be ascribed to this relative differencein the numbers of cells degenerating. It does not seem necessary to invoke anyincrease in the rate of immigration of cells into the ventral horn in order toexplain the excess.
(4) During the third and fourth weeks the numbers of living cells countedin the ventral horn on the operated side declined (Fig. 3 A) faster than those of

372 M. C. PRESTIGE
the control side; as a result, the relative excess was converted to a relativedeficit (Fig. 3B). This was accompanied by the presence of cell degeneration onthe operated side throughout this period, while on the control side cell degenera-tion had ceased (Fig. 3 C), as is usual at the end of metamorphosis. This lateloss of ventral horn cells after amputation was also observed by Hughes (1961)after amputation at stages 55-56; it was not observed in series I of the presentwork after amputation at stages 52-53.
(5) At 30 days, less than 25 % of the ventral horn cells remained. It seemstherefore that the original leg amputation was approximately 75 % complete inthis series (Table 3).
(6) The number of cells in chromatolysis (mean 1024) is somewhat less thanthe excess number of cells lost in the final loss period (1500). This latter figureis the difference between the excess of cells seen in the second week and thefinal deficit.
Series IV: amputation at stage 61
A more extensive series was done with 21 tadpoles at this stage: they wereabout to enter metamorphic climax, the head had shrunk noticeably, the ankleswere flexed at rest and they swam using repeated bilateral kicks as well as thetail (Hughes & Prestige, 1967). The legs were 10-15 mm long at operation. Atthis stage in the ventral horn there are 1100-1800 cells a side; turnover ispresent, but at a much lower level than at stages 54-56 (Hughes, 1961), and themajority of the cells are large and appear mature. During the next 8 days thetadpole will change into a juvenile toad.
The animals of Table 4 shows- the following features:(1) There is no evidence of early cell loss or of an excess of degenerating cells
on the operated side, such as was seen between stages 53 and 56. Many cellsundergo chromatolysis.
(2) The operation depresses the rate of degeneration ipsilaterally after 2 days(Fig. 4B) for the remainder of the period during which histogenetic degenerationnormally takes place. This is most marked on days 2-4 (Mann-Whitney ranktest, P = 0-036). This depression is similar to that observed in series I—III.
(3) An excess of 150-200 living cells on the operated side builds up at theend of the first week (Fig. 4 A). This excess appears against a base-line of thenumber of cells in the contralateral ventral horn, which is still declining slowlyduring this week. The excess appears similar to that observed in series III.
(4) After the first week, the excess in cell numbers is maintained for 2 months;there are no changes in total cell number and hardly any degenerating cells areseen on either side (Fig. 4).
(5) A slow decline in the number of ventral horn cells on the operated sidetakes place during the third and fourth month. At 62 days there is some evidenceof excess cell degeneration which may be associated with this. The total numberof cells remaining after 6 months is 650 (43 %), most of which are in the anterior
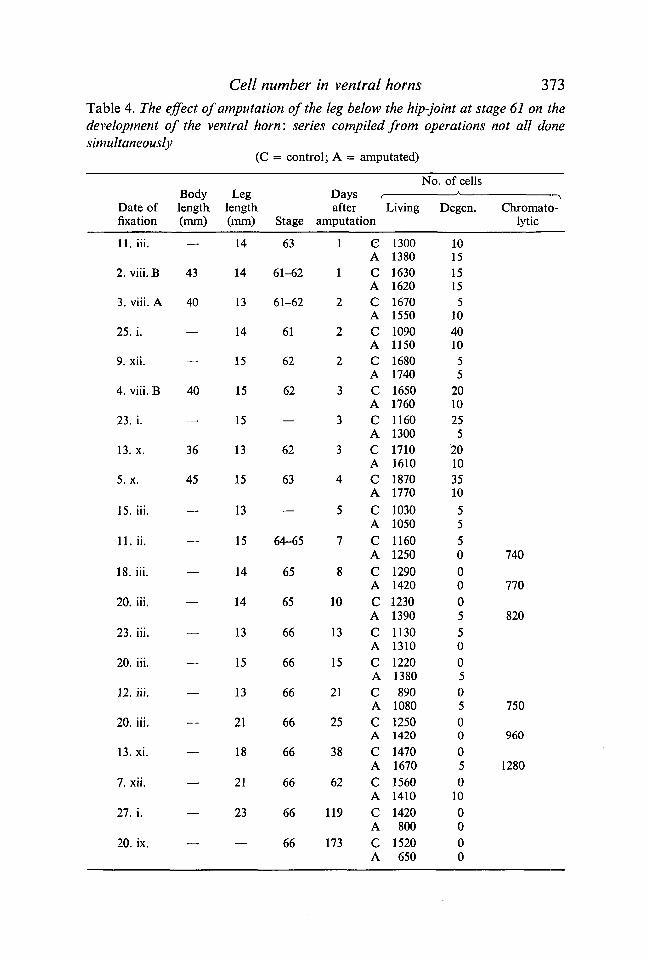
Cell number in ventral horns 373Table 4. The effect of amputation of the leg below the hip-joint at stage 61 on thedevelopment of the ventral horn: series compiled from operations not all donesimultaneously
(C = control; A = amputated)
Date offixation
11. iii.
2. viii. B
3. viii. A
25. i.
9. xii.
4. viii. B
23. i.
13.x.
5.x.
15. iii.
11. ii.
18. iii.
20. iii.
23. iii.
20. iii.
12. iii.
20. iii.
13. xi.
7. xii.
27. i.
20. ix.
Bodylength(mm)
—
43
40
—
—
40
—
36
45
—
—
—
—
—
—
—
—
—
—
—
—
Leglength(mm)
14
14
13
14
15
15
15
13
15
13
15
14
14
13
15
13
21
18
21
23
—
Stage
63
61-62
61-62
61
62
62
—
62
63
—
64-65
65
65
66
66
66
66
66
66
66
66
Days rafter
amputation
1
1
2
2
2
3
3
3
4
5
7
8
10
13
15
21
25
38
62
119
173
eA
cAcAcACACACACACACACACACACACACACACACACACA
Living
130013801630162016701550109011501680174016501760116013001710161018701770
10301050
11601250
12901420
123013901130131012201380890
108012501420
14701670
156014101420800
1520650
No. of cells
Degen.
101515155
10401055
201025
520103510
5550000550050500050
100000
Chromato-lytic
740
770
820
750
960
1280

374 M. C. PRESTIGE
part of the ventral horn. Delayed loss in this series is thus similar to that ob-served in series III and in Hughes (1961), though later and more protracted.
(6) The number of cells in chromatolysis (mean 886) is slightly less than theextra number of cells lost during the final decline (about 1000).
CO
•> 2
O Q,
W
0200 r
+
Days
5 10 15 20 25. 38 62119 173
8f. 0>
a o/55
200
400
600 L
50
+0
50
-D
Fig. 4. Amputation at stage 61. A, The differences in the number of living cells be-tween the two sides. B, The differences in the number of degenerating cells betweenthe two sides.
The question may be raised whether second loss after amputation at stages55-56 took place in cells that, though normally present at the time of theoperation, were insensitive to the absence of the limb, but that as developmentprogressed entered a phase in which they became sensitive, found the limb absentand so died. This hypothesis predicts that amputation in the present series wouldresult in immediate loss of the sensitive cells. Such behaviour is not observed,and so second loss must be due to a delayed reaction to the original operation.
A similar argument and conclusion may be drawn for the final loss of cellsafter amputation at stages 57 and 61. In this case the evidence comes from theresults of amputating 1-2 months after metamorphosis (below). Final loss istherefore due to a delayed reaction in ventral horn cells.
The conclusion from this argument stands in contrast to that obtained fromthe delayed degeneration or regression of ventral horn cells after limb-budremoval at stage 49-50 (Hughes & Tschumi, 1958). In that case, amputation atstages 53-54, the stages at which delayed regression occurs, causes immediateloss of cells. Thus the later death of these cells, after amputation at stages 49-50,is an example of delayed sensitivity to removal of the periphery.

Cell number in ventral horns 375
Series V: amputation early in juvenile life
Thirteen toads were amputated at the hip in the first few months after meta-morphosis. The data are shown in Table 5. No early cell loss took place, norwas there any renewal of cell degeneration. Many cells underwent chromato-lysis. Final delayed loss of cells took place mostly between the twelfth andthirtieth weeks, though some indications of cell degeneration were still present
Table 5. The effect of amputation of the leg below the hip-joint on the ventralhorn of the juvenile: series compiled from operations not all done simultaneously
(C = control; A = amputated)
Date offixation
3. iii. A
4. iii. A
23. iii.
3. iv.
7. iv.
7. xi.
15.x.
5. ix.
20.x.
1. viii.
30. xi.
27. i.
21. iii.
Leglength(mm)
15-18
_
17
—.
.—
30
—
26
33
_
50
45
60
Days afteramputation
1
2
4
7
11
18
25
77
122
127
163
221
274
CACACACACACACACACACACACACA
t
Living
131013301620164016401660131013701430143014801600134013801170910
16701680
12601320
206016902010132018001090
No. of cells
Degen.
5000000050000005——00——
0005
Chromato-lytic
800
750
660
750
0
after 9 months. The number of cells remaining was 61 % and 66 % in the lasttwo animals fixed. The last neurones to recover from chromatolysis and to diewere found in the centre portion of the ventral horn. The number of neuronesthat underwent chromatolysis (mean 740) was equal to the number of cells that

376 M. C. PRESTIGE
died in final loss (mean 700); they were present in similar regions of the ventralhorn.
DISCUSSION
It has been recognized for a century that developing neurones are moresusceptible to injury than mature ones. This has been most clearly documentedby Romanes (1946) and LaVelle & LaVelle (1958). These authors described acritical change in the reactivity of mammalian foetal motor neurones: in thehamster, for example, after axotomy at birth, the first changes were seen in thenucleus, and were followed in 4-6 days by cell death, while after the sameoperation 7 days post partum, the cells showed only cytoplasmic chromatolysisand 26 days later one third of the cells were still surviving.
In the anuran Xenopus amputation at stages before 52-53 has no effect on theventral horn until stages 52-54 (Perri, 1956a; Hughes & Tschumi, 1958).Amputation at stages 53-54 provokes a large early loss of cells (series I, II);at stages 55-56 a smaller early loss and also a later second loss (Hughes, 1961);at stage 57 a very small early loss and a larger late loss (series III); at stage 61only a late loss (series IV); and as juveniles only the late loss (series V). Theseexcess cell losses are accompanied by excess cell degeneration. In the periodsin which excess cell losses are not taking place the rate of histogenetic celldegeneration on the operated side is significantly depressed below that of thecontrol side.
During development three phases in the reaction of the ventral horn cells toamputation can thus be recognized:
(1) Phase I, in which ventral horn cells are unaffected by the removal of theperiphery. These cells are neuroblasts and possess little cytoplasm.
(2) Phase II, in which the ventral horn cell dies within 3-4 days of amputation.(3) Phase III, in which the ventral horn cell does not die immediately after
amputation but does so only later, usually after an extended period of chromato-lysis and recovery. These cells are mature in appearance.
Estimates of the number of cells in each phase at each stage of developmentmay be made as follows:
Phase I. The ventral horn before stages 52-53 consists entirely of phase I cells,since it is unaffected by removal of the periphery (Hughes & Tschumi, 1958).Following removal of the limb-bud at stages 52-53 (series I), any phase II cellspresent degenerate, no phase III cells can mature, and therefore the number ofremaining cells at any time on the operated side is equal to the number of phaseI cells present (after making a linear correction for 86 % limb-bud removal).The graph of this has been plotted in Fig. 5 A as curve I.
Phase II. The cell deficit that develops within 3-4 days of amputation definesthe number of phase II cells within the ventral horn at the operation aftercorrection for incomplete amputation. This has been applied in Table 6, andthe corrected estimates plotted as solid circles in Fig. 5B. The curve through

Cell number in ventral horns 311
the points is labelled curve II. (It is possible that the number of cells that de-generate after amputation is not equal to the deficit that develops; this wouldhappen if there was a differential change in immigration rates as well.)
Phase III. The number of cells in phase III is defined as the number of cellsthat die in the second or final loss in excess of those normally dying contra-aterally; this is equal to the difference between the inter-side differences beforeand after final loss, multiplied by the correction factor for incomplete amputa-tion. These have been calculated in Table 7 and plotted as open circles in Fig. 5C,curve III.
8000
6000
4000
2000
'\f \
- / \
\ \
1 1 1 1 152 54 56
A
s v I + II + IM
58 62
4000
3000
2000
1000
n
3000
2000
1000
-
-
ri " l ^
66 JUV. 52
' - . II B
•
in / * *
y*j i i i i 1111111
54 56 58 62
*
C
i i66 JUV.
Stage
Fig. 5. The numbers of ventral horn cells in each phase during development. A,Phase I (continuous line). B, Phase II (filled circles). C, Phase III—estimated by size offinal loss (open circles); by numbers of chromatolytic cells (filled circles). The sum ofthe three curves phase I+11+III is plotted in A (interrupted line) and in Fig. 1(see text).
It might be expected that the number of cells that undergo chromatolysis onamputation should be related to the number of cells involved in final loss. Thisview is supported by the close correspondence in the position of the chromato-lytic cells and those finally lost. Both are mainly in the posterior two-thirds.The numbers of cells either in chromatolysis or in the dense perinuclearbasophilic ring stage of recovery are also plotted as filled circles in Fig. 5Cafter correction for incomplete leg removal. The figures for stages 55-56 havebeen taken from the same material that Hughes (1961) used. The remainingdata have already been presented in the Tables. Comparison of the two methodsof estimation shows that counts of chromatolytic neurones are unreliablemarkers of phase III cells at early stages; that is, when there is little cytoplasm.

378 M. C. PRESTIGE
Table 6
Cell deficit Final cellDate of fixation at 3-4 days deficit (%)
—
20. xi.15. xi.16. xi.
21. xi.
—
25. xi.26. xi.
23. i.13.x.5.x.
23. iii.
* The final cell
Hughes & Tschumi (1958): before stage0 —
Series I: stages 52-53600 )
2030 \ 86850 J
Series II: stage 541710 (80)*
Hughes (1961): stages 55-56c. 600 62
Series 111: stage 57210 \
0 J 76
Series IV: stage 61-140 ]
100 [ 57100 J
Series V: juvenile-20 37
deficit was not determined for the animals
Correctedcell deficitat 3-4 days
520
6802320970
(2140)
970
2800
-250180180
-60
of series II.
Table 7
Stage and source
53 (series I)55-56 (Dr A.
No. of cells ipsilaterallyminus numbercontralaterally
i ^
(a) Before (b) Afterfinal loss final loss
No final loss —
Hughes pers. comm.) 200 -81057 (series III)61 (series IV)Juveniles(series V)
250 -1250200 - 800
0 -700
(a)-(b)
0
101015001000
700
deficit(%)
—
627657
37
Estimated
Phase IIIcells
0
163019701750
1900

Cell number in ventral horns 379
This has already been commented on in the section on Materials and Methods.At later stages, the two methods approximate.
A synthetic total ventral horn cell count curve can then be constructed byaddition of curves I, II and III. This has been plotted in Fig. 1, and 5 A (dashedline) against the observational data on the ventral horn cell totals on the controlside; considering the paucity of points and the assumption of the linear correc-tion factor, the fit is satisfactory, despite the peak at stage 53. The followingpoints can be noted:
(1) Phase I cells decrease in number from being the sole constituents of theventral horn at stage 52 to almost zero by stage 61.
(2) Phase II cells are only present in observable numbers between stages 52and 58: they account for half the total numbers around stage 54.
(3) Phase III cells increase in number from zero at stage 53 until at stage 57they are almost the only type of cell in the ventral horn.
(4) All these maturation processes occur most rapidly at stages 53-56—thatis, when the total ventral horn cell number is decreasing most rapidly and thereis a peak in the number of histogenetically degenerating cells (Hughes, 1961;Fig. 1).
(5) Small numbers (less than 200) of phase 1, II and III cells could be presentat times other than those of Fig. 5, but not resolved by the analysis.
(6) The counts are made on the state of the ventral horn. They do notdescribe the flux of neurones through the phases. To do this would requireknowledge of the pattern of immigration of cells into phase I, and whether thiswas also controlled by the periphery, as suggested by the successfully regenerat-ing animals of series I.
How does a ventral horn cell know that the leg has been amputated!Why does it subsequently die ?
The effects of amputation are ipsilateral only, which excludes any hypothesisof a systemically borne substance which may cause motor neurones todegenerate. A direct 'trauma' substance is ruled out for three reasons: first,because cell death may be postponed for many months after the operation.Secondly, in the CSL series of Hughes & Tschumi (1958), in which innervationof the growing leg was hindered by a slit before fibres had grown out, and inwhich no subsequent trauma or amputation took place, the only ventral horncells which survived were those few in which successful contact with the peri-phery was established by means of a spinal nerve; the remainder, though theyall completed their initial differentiation, degenerated or regressed. This showsthat it is the absence or interruption of the spinal nerve that is signalled andleads to cell death (ventral roots and limb nerves are present at all times whenamputation causes degeneration in the ventral horn—unpublished observationson Palmgren preparations). Thirdly, the cells that die as a result of amputatingthe leg below the hip-joint are related to the amputation stump not so much by
24 JEEM l8

380 M. C. PRESTIGE
proximity as by the pattern of spinal nerves; they lie in the posterior two-thirdsof the ventral horn, which supplies spinal nerves 9 and 10. These two spinalnerves make up the sciatic nerve in the normal plexus with only a small contri-bution from other sources. Cells of spinal nerve 8, which supply the iliacmusculature by the crural nerve, are largely unaffected.
A number of effects follow the cutting of a peripheral nerve, which involve thepassage of information from the site of the lesion to the cell body. We are hereconcerned only with the information that leads to the ultimate death of the cell.The evidence that it is not due to the mechanical effects of changing the resis-tance to axoplasmic flow by opening or squashing the axon comes from Weiss &Hiscoe (1948), who showed that the pressure changes within the fibre whichfollow squashing are only local, and so no mechanical information can be passedback to the cell body; this will be true also of cutting the axon. The injurydischarge that is set up on cutting the axon probably lasts only for a few seconds,and so this form of information transfer cannot be responsible, nor can it be dueto the injury current, for this would at most only spread electrotonically a fewhundred micra. The information must therefore be sensed either by reduction ofthe impulse traffic, or by the absence of a chemical normally transported centri-petally, or by central accumulation of a metabolite due to a failure to remove itcentrifugally. It is difficult to see how the last alternative can be correct, forcutting the axon does not halt growth and is unlikely to stop the removal of themetabolite. Moreover, axoplasmic damming (Weiss & Hiscoe, 1948) is onlytemporary and passes off long before the cell dies in mature cases. Centrifugalmetabolites of this kind would be 'lethal'; centripetal ones would be 'trophic'.It is possible that trophic information is carried by the impulse traffic; thiswould then have to be in sensory nerve fibres, as the direction of travel is wrongfor motor fibres. However, Taylor's (1944) experiment in Rana, in which hefound that motor nerves were not lost, and became functional in the absence ofsensory nerves, suggests that the maintenance information is carried centri-petally along motor nerve fibres. The concept of a trophic chemical, transportedintra-axonally in motor nerve fibres, is therefore preferred. Interruption of thenerve at amputation leads to cell death, due to absence of this 'maintenancefactor'.
The relationship between the maturity of a cell and its abilityto postpone death after cutting of its axon
Phase II cells (by definition) are not able to postpone death after amputationof the leg. Phase III cells (again by definition) can. The degree to which this isso depends on the maturity of the cell at the time of the operation. When thisis performed at stages 55-56, final loss takes place after about 3 weeks (Hughes,1961); after amputation at stage 57, during the fourth week (Table 2); afteramputation at stage 61, during the third and fourth month (Table 4); and afteramputation as juveniles, not until the fourth to seventh months (Table 5).

Cell number in ventral horns 381
Young neurones therefore die after amputation very quickly, while older onestake longer. Thus the times at which cells die after amputation plot out a maturityspectrum for the cells in the ventral horn at the time of the operation.
It has already been argued that neurones die because they are no longergetting something from the leg. Dr A. Hughes has pointed out to me that thefact that they may take a long time to die suggests that the cells are using up astore of the limb factor or its product. It is proposed therefore that the main-taining factor from the limb (or a product of it) is stored within the ventralhorn cell; it is constantly used and the cell dies if the store runs out; and moremature cells have bigger stores and so can survive longer without replenishment.Maturity can thus be measured in terms of store size. This mechanism has adap-tive significance, for it allows nerve fibres to be broken during natural lifewithout loss of the parent cell. This also provides a reason, though not anexplanation, why the store is only large enough to allow protein synthesis andaxon growth to proceed far enough to re-establish connexions.
The maintenance factor proposed comes into a general category of celluli-petal neurotrophic substances. It is known that, following section of a peri-pheral muscle nerve in adults, reinnervation of the muscle can to some extentreverse the effects of the lesion on the injured nerve cell, and that this mustinvolve passage of information from the axon tip to the perikaryon (Weiss,Edds & Cavanaugh, 1945; Aitken, Sharman & Young, 1947; and many others).Similarly, Evans & Yizoso (1951) showed that the characteristic maturation ofmotor nerve fibres in post-natal life is completely dependent on the establish-ment of peripheral connexions. Recently, Kerkut, Shapira & Walker (1967)have shown that in an isolated CNS—nerve trunk—muscle preparation,[14C]labelled material placed in the muscle compartment is carried antidromi-cally along the motor nerves to the CNS.
The maintenance factor hypothesis leaves unanswered any questions as tothe origin of the substance, and as to the nature or location of the store. It maybe asked whether the same mechanism can be extended from these results ondeveloping neurones to those of axotomy in adults. It may be that some of thegreat variety in the timing and severity of reactions to axonal lesions in differingcell groups comes from the particular quantitative combination of the size ofthe store and the change in the supply of maintenance factor. In this connexion,it is interesting to recall that adult dorsal root ganglion cells survive followingsection of the appropriate dorsal root for 2-3 years (Hinsey, Krupp & Lhamon,1937), whereas after section of the spinal nerve there is substantial cell loss(Ranson, 1906). For these cells, it apears that it is only the absence of peripheralconnexions that leads to their death, rather than axonal trauma.
24-2

382 M. C. PRESTIGE
The presence of degenerating cells in the ventral horn during normaldevelopment {histogenetic degeneration)
The most plausible explanation is that, at least from stages 53-56, theyrepresent phase II cells which have failed to develop due to a lack of a main-tenance factor from the leg. The evidence supporting this view is:
(1) There is a close correspondence between the shapes of the graphs of thenumber of phase II cells (Fig. 5B, curve II) and of the observed numbers ofdegenerating cells (Fig. IB; Hughes, 1961).
(2) In series I it was pointed out that this approximation of the capacity todegenerate after amputation and the capacity for histogenetic degenerationheld even within different zones of the same ventral horn. This suggeststhat the two types of cell death occur for the same reason. It has already beenargued that cell death after amputation is due to lack of a maintenancefactor.
(3) It is improbable that they are phase I cells, because the amount of cyto-plasm present around the pycnotic nuclei is greater than that associated withventral horn cells at stage 52, when all the cells are in phase I. Also the rate ofdegeneration can be altered by amputation, while phase I cells are independentof the leg.
(4) Following incomplete leg amputation, such as that performed in theseexperiments, the early loss of cells only reduces the number of phase II cells;the numbers of phase I and phase III cells remain unaltered by definition.When the number of phase II cells is thus reduced, the number of cells de-generating is similarly reduced: this was observed in series I-IV between theexcess periods of cell loss.
(5) If for any reason the supply of maintenance factor from the limbs wasliable to fluctuate, phase II cells (when present) would be more likely to diethan phase III cells, being intrinsically more sensitive to such a deficiency.
An alternative hypothesis is that the leg instructs the ventral horn cells to dieby an 'execution warrant' substance travelling centripetally. This would explainthe decrease in number of degenerating cells in the intermediate period afterpartial amputation. But it does not explain why any excess cell loss occurs inthe first place. Neither can it account for the cell loss in the CSL series of Hughes& Tschumi (1958) in which there was no contact between the leg and the ventralhorn. This hypothesis is therefore untenable without the additional hypothesisof a maintenance factor to explain the effects of amputation, whereas the lattermakes the former unnecessary.
For the later stages of development there are two exceptions to the concept ofhistogenetically degenerating cells all being in phase II. First, after stage 57,no phase II cells are present in the ventral horn (Fig. 5B), yet histogeneticdegeneration continues at a low rate until stage 65. This situation is due to alimitation in the definition of phase II (the observed cell loss): cells may be

Cell number in ventral horns 383
prevented from degenerating that otherwise would have, or there may beassociated changes in the immigration of cells into the ventral horn. Either ofthese events if present, would cause false estimates to be made of the realnumber of cells that die after amputation. These would be most acute when theobserved numbers are low, as after stage 57.
The second objection is that the concept does not explain the relative build-upof cells ipsilaterally after amputation at stages 57 and 61. In each case this isaccompanied by a depression in the ipsilateral rate of degeneration. If thislatter is due only to the decrease in the number of phase II cells followingpartial amputation, the continuing decline of numbers on the control side (alsoby loss of phase II cells) cannot do more than restore the balance finally. Forthe relative pile-up to take place, there must be either prevention of the degenera-tion of some cells that otherwise would have died, or there must be changes inthe immigration rate. While it is not yet possible to estimate the relative impor-tance of these, there is some evidence that phase III cells do degenerate duringthe latter half of normal development. Some of the cells seen with pycnoticnuclei at these times have as large a quantity of cytoplasm as any others. It maybe that the chromatolytic reaction holds up cell death that otherwise might haveoccurred. It is concluded that the cells which die in the ventral horn duringnormal development are phase II cells, and that phase III cells may also die inthe later stages.
The decline in cell number during development
The high sensitivity of phase II cells to loss of maintenance factor and thesteadily increasing independence of the phase III cell as it matures means thatin competition between cells of different ages, the older cell has the advantage.The design of the system is such that achievement by a neurone of even a slightlylarger peripheral connexion than its fellows confers a virtual guarantee that itsown supply of maintenance factor will not dry up. Furthermore, it could causethe disparity between itself and its fellows to grow greater. This means that thesystem produces a small number of neurones, each with a large, stable peripheralfield; the remaining neurones, those which have failed to achieve peripheralconnexions or those whose peripheral field was not large enough, becomeredundant and die. In this respect, neurones behave like industrial concerns ina laissez faire economy.
This arrangement ensures that neurones have an efficient, though not neces-sarily specific, connexion with the periphery. The establishment of specific, co-ordinated contacts must require a subsidiary mechanism.
The development of the neuromotor system
In the period between stages 53 and 57, some 2̂ —3 weeks, and especially theweek of stages 54-55:
(1) Cell number in the ventral horn falls to half its starting figure, accom-

384 M. C. PRESTIGE
panied by the appearance of large numbers of pycnotic nuclei (Hughes,1961).
(2) At stage 53 ventral horn neurones are small and neuroblastic; at stage 57they are large cells with dendrites and Nissl substance, cholinesterase-positive(Hughes, 1961) and acid phosphatase-positive (Palkama & Prestige, 1964).
(3) The leg muscles differentiate (Nieuwkoop & Faber, 1956).(4) At stage 53 ventral horn cells are still all in phase I; by stage 57 practi-
cally all are in phase III.(5) The neuronal circuitry for the 'flare' and 'stepping' movements of the
leg is established (Hughes & Prestige, 1967).(6) The spinal cord becomes extremely sensitive to strychnine, indicating the
genesis of post-synaptic inhibition (Hughes & Prestige, 1967).
SUMMARY
1. The development of the lumbar ventral horn cells in Xenopus tadpolesafter amputation of the hind leg at the hip has been studied by counting thenumbers of living, degenerating and chromatolytic cells present at intervals afterthe operation.
2. During development, each cell passes through three phases, defined by itsreaction after amputation: the operation either has no effect on the cell (phase I)or it causes it to degenerate within 3 days (phase II), or it may cause it to chrom-atolyse first and only degenerate weeks or months later (phase III). Cell loss,relative to the opposite side, takes place in steps—either early or late or both—and is accompanied by an excess of degenerating cells. At other times, celldegeneration, if normally present, is reduced in rate.
3. Removal of the limb-bud has no effect until the palette stage. At the palettestage, amputation causes a large early but no late loss; at the stepping stage(hind-leg about 4-5 mm), a small early and a small late loss; at the beginningof metamorphic climax, a negligible early and a large late loss; at tail loss, noearly and a large late loss; and as juveniles, no early and a large late loss.
4. The numbers of cells in phases I, II and III throughout development havebeen independently estimated. The algebraic sum of these three is the same as thenumber of cells on the control side for each stage.
5. The period for which cells in phase III can survive after amputation in-creases with maturity.
6. It is argued that cells in phases II and III die after amputation because theyare no longer getting from the leg an essential 'maintenance factor' which isnormally carried in the motor axons. It is suggested that phase III cells are ableto store this substance, and thus put off death temporarily; and that moremature cells have larger stores and so can survive longer without replenishment.Early maturing cells thus have a selective advantage over their fellows.

Cell number in ventral horns 385
RESUME
Regulation du nombre de cellules dans les comes medullaires lombairesventrales, au cours du developpement de tetards de Xenopus laevis.
1. On a etudie le developpement des cellules de la corne ventrale lombaire detetards de Xenopus apres amputation du membre posterieur, en comptant lenombre de cellules vivantes, en degenerescence et chromatolytiques presentes adivers intervalles de temps apres l'operation.
2. Au cours du developpement, chaque cellule passe par trois phases, definiespar sa reaction apres l'amputation: l'operation, ou bien n'a aucun effet sur lacellule (phase I), ou la fait degenerer dans les 3 jours (phase II), ou peut d'abordentrainer sa chromatolyse et ensuite sa degenerescence, des semaines ou desmois plus tard (phase III). La perte en cellules, par rapport au cote oppose, alieu par etapes, soit precocement, soit tardivement, ou les deux, et s'accompagned'un exces de cellules en degenerescence. A d'autres moments, la degenerescencecellulaire, si elle est normalement presente, voit son taux reduit.
3. L'ablation du bourgeon de membre n'a pas d'effet jusqu'au stade palette.Au stade palette, l'amputation provoque une forte perte precoce mais nontardive; au stade du membre posterieur de 4 a 5 mm, une faible perte precoceet une faible perte tardive; au debut du climax metamorphotique, une perteprecoce negligeable et une forte perte tardive; a la regression de la queue, pasde perte precoce et une forte perte tardive, de meme sur des jeunes meta-morphoses.
4. On a evalue de maniere independante le nombre de cellules des phases I,II et III tout au long du developpement. La somme algebrique de ces troisvaleurs est la meme que la nombre de cellules du cote temoin pour chaquestade.
5. La periode pendant laquelle des cellules en phase III peuvent survivreapres l'amputation augmente avec la maturite.
6. On suppose que les cellules en phase II et III meurent apres l'amputationparce qu'elles ne recoivent plus de la patte un 'facteur de maintien' essentiel quise trouve normalement transports dans les axones moteurs. On suggere que lescellules en phase III sont aptes a stocker cette substance et echappent ainsitemporairement a la mort; et que les cellules plus mures ont des reserves plusimportantes et peuvent ainsi survivre plus longtemps sans 'refaire le plein'. Lescellules a maturation precoce ont ainsi un avantage selectif sur leurs soeurs.
I am very grateful to Dr A. Hughes for encouragement and advice throughout this work;to Miss Judith Garraway who ably assisted at all times; and to the Medical Research Councilfor support.
REFERENCES
ATTKEN, J. T., SHARMAN, M. & YOUNG, J. Z. (1947). Maturation of regenerating nerve fibreswith various peripheral connections. / . Anat. 81, 1-22.
BARRON, D. H. (1945). The role of the sensory fibres in the differentiation of the spinal cordin sheep. / . exp. Zool. 100, 431-44.

386 M. C. PRESTIGE
BARRON, D. H. (1946). Observations on the early differentiation of the motor neuroblasts inthe spinal cord of the chick. / . comp. Neurol. 85, 149-69.
BARRON, D. H. (1948). Some effects of amputation of the chick wing bud on the early differ-entiation of the motor neuroblasts in the associated segments of the spinal cord. / . comp.Neurol. 88, 93-127.
BEAUDorN, A. R. (1955). The development of lateral motor column cells in the lumbo-sacral cord in Rana pipiens. I. Normal development and development following unilaterallimb ablation. Anat. Rec. 121, 81-96.
BUEKER, E. D. (1943). Intracentral and peripheral factors in the differentiation of motorneurons in transplanted lumbo-sacral spinal cords of chick embryos. / . exp. Zool. 93,99-129.
BUEKER, E. D. (1944). Differentiation of the lateral motor column in the avian spinal cord.Science, N. Y. 100, 169.
BUEKER, E. D. (1945 a). The influence of a growing limb on the differentiation of somaticmotor neurons in transplanted avian spinal cord segments. / . comp. Neurol. 82, 335-61.
BUEKER, E. D. (19456). Hyperplastic changes in the nervous system of a frog (Rana) asassociated with multiple functional limbs. Anat. Rec. 93, 323-9.
COLLIN, R. (1906). Recherches cytologiques sur le developpement de la cellule nerveuse.Nevraxe 8, 181-308.
EVANS, D. H. L. & Vizoso, A. D. (1951). Observations on the mode of growth of motornerve fibers in rabbits during postnatal development. /. comp. Neurol. 95, 429-61.
GLUCKSMANN, A. (1951). Cell deaths in normal vertebrate ontogeny. Biol. Rev. 26, 59-86.HAMBURGER, V. (1934). The effects of wing bud extirpation on the development of the central
nervous sytem in chick embryos. /. exp. Zool. 68, 449-94.HAMBURGER, V. (1939). Motor and sensory hyperplasia following limb-bud transplantation
in chick embryos. Physiol. Zool. 12, 268-84.HAMBURGER, V. (1958). Regression versus peripheral control of differentiation in motor
hypoplasia. Am. J. Anat. 102, 365-410.HAMBURGER, V. & KEEFE, E. L. (1944). The effects of peripheral factors on the proliferation
and differentiation in the spinal cord of chick embryos. J. exp. Zool. 96, 223-42.HAMBURGER, V. & LEVI-MONTALCINI, R. (1949). Proliferation, differentiation, and degenera-
tion of the spinal ganglia of the chick embryo under normal and experimental conditions.J. exp. Zool. 111,457-501.
HARRIS, A. E. (1965). Differentiation and degeneration in the motor horn of the foetal mouse.Ph.D. thesis, University of Cambridge.
HINSEY, J. C, KRUPP, M. A. & LHAMON, W. T. (1937). Reaction of spinal ganglion cells tosection of dorsal roots. / . comp. Neurol. 67, 205-14.
HUGHES, A. (1955). The development of the neural tube of the chick embryo. A study withthe ultraviolet microscope. / . Embryol. exp. Morph. 3, 305-25.
HUGHES, A. (1961). Cell degeneration in the larval ventral horn of Xenopus laevis (Daudin)./ . Embryol. exp. Morph. 9, 269-84.
HUGHES, A. (1962). An experimental study on the relationships between limb and spinal cordin the embryo of Eleutherodactylus martinicensis. J. Embryol. exp. Morph. 10, 575-601.
HUGHES, A. (1963). On the labelling of larval neurones by melanin of ovarian origin incertain Anura. / . Anat. 97, 217-24.
HUGHES, A. (1964). Further experiments on the innervation and function of grafted super-numerary limbs in the embryo of Eleutherodactylus martinicensis. J. Embryol. exp. Morph.12, 229-45.
HUGHES, A. & PRESTIGE, M. C. (1967). The development of behaviour in the hind limb ofXenopus laevis. J. Zool. 152, 347-59.
HUGHES, A. & TSCHUMI, P. A. (1958). The factors controlling the development of the dorsalroot ganglia and ventral horn in Xenopus laevis (Daud.). / . Anat. 92, 498-527.
JOHNSON, Z. & CLARKE, J. L. (1868). On a remarkable case of extreme muscular atrophy withextensive disease of the spinal cord. Medico-Chir. Trans, 51, 249-64.
JONES, R. L. (1937). Split nucleoli as a source of error in nerve cell counts. Stain Technol. 12,91-5.

Cell number in ventral horns 387
KERKUT, G. A., SHAPIRA, A. & WALKER, R. J. (1967). The transport of labelled material fromCNS ^ Muscle along a nerve trunk. Comp. Biochem. Physiol. (in the Press).
LAVELLE, A. & LAVELLE, F. W. (1958). The nucleolar apparatus and neuronal reactivity toinjury during development. / . exp. Zool. 137, 285-316.
MAY, R. M. (1930) Repercussions de la greffe de moelle sur le systeme nerveux chez l'embryonde l'Anoure, Discoglossus pictus Otth. Bull. biol. Fr. Belg. 64, 355-87.
MAY, R. M. (1933). Reactions neurogeniques de la moelle a la greffe en surnombre, ou aTablation d'une ebauche de patte posterieure chez l'embryon de l'Anoure, Discoglossuspictus Otth. Bull. biol. Fr. Belg. 67, 327-49.
MOTTET, K. (1952). The effect of removal of somatopleur on the development of motor andsensory neurons in the spinal cord and ganglia. J. comp. Neurol. 96, 519-53.
MOTTET, K. & BARRON, D. H. (1954). Some effects of the peripheral field on the cytochemicaldifferentiation of neurons. Yale J. Biol. Med. 26, 275-84.
NIEUWKOOP, P. D. & FABER, J. (1956). Normal Table o/Xenopus laevis (Daudiri). Amsterdam:North Holland Publishing Co.
PALKAMA, A. & PRESTIGE, M. C. (1964). Acid phosphatase in the developing central nervoussystem. /. Physiol. Lond., 175, 7-9 P.
PALLADINI, G. & ALFEI, L. (1966). Risposta dei centri motori spinali all' asportazione dellaperiferia durante il differenziamento in larve di Bufo bufo (L.). Atti Accad. naz. Lincei Re.40, 296-304.
PALMGREN, A. (1948). A rapid method for selective silver staining of nerve fibres and nerveendings in mounted paraffin sections. Acta Zool. Stockh. 29, 378-92.
PERRI, T. (1956a). Ricerche sulle correlazioni tra midollo spinale, gangli spinali ed arti negliAnfibi anuri. Esperienze d'asportazione di un abbozzo d'arto in Bufo vulgaris. Archo. zool.ital. 41, 369^10.
PERRI, T. (19566). Trapianti embrionali di midollo spinale e genesi dei corni motori in Bufovulgaris. Atti Accad. naz. Lincei. Re. 20, 666-670.
PERRI, T. (1957). Sul trapianto di abbozzi di arte negli Anfibi anuri con particolare reguardoalle consequenze sul sistema nervoso centrale e periferico. Riv. Biol. 49, 361-417.
PIATT, J. (1948). Form and causality in neurogenesis. Biol. Rev. 23, 1-45.PRESTIGE, M. C. (1967). The control of cell number in the lumbar spinal ganglia during the
development of Xenopus laevis tadpoles. / . Embryol. exp. Morph. 17, 453-71.RACE, J. & TERRY, R. J. (1965). Further studies on the development of the lateral motor
column in Anuran larvae. I. Normal development in Rana temporaria. Anat. Rec. 152,99-106.
RANSON, S. W. (1906). Retrograde degeneration in the spinal nerves. 7. comp. Neurol. 16,265-93.
ROMANES, G. J. (1946). Motor localization and the effects of nerve injury on the ventralhorn cells of the spinal cord. / . Anat. 80, 117-31.
SHERRINGTON, C. S. (1893). Notes on the arrangement of some motor fibres in the lumbo-sacral plexus. /. Physiol, Lond. 13. 621-772.
SIEGEL, S. (1956). Nonparametric Statistics for the Behavioral Sciences. New York: McGraw-Hill Book Company Inc.
STULTZ, W. A. (1942). Alterations in the spinal cord of Amblystoma following changes in theperipheral field. Anat. Rec. 82, 450.
TAYLOR, A. C. (1944). Selectivity of nerve fibers from the dorsal and ventral roots in thedevelopment of the frog limb. J. exp. Zool. 96, 159-85.
VULPIAN, A. (1868). Influence de l'abolition des fonctions des nerfs sur la region de la moelleepiniere qui leur donne origine. Examen de la moelle epiniere dans des cas d'amputationd'ancienne date. Arch. Physiol. norm. path. 3, 443.
WEISS, P., EDDS, M. V. & CAVANAUGH, M. (1945). The effect of terminal connections on thecaliber of nerve fibres. Anat. Rec. 92, 215-33.
WEISS, P. & HISCOE, H. B. (1948). Experiments on the mechanism of nerve growth. /. exp.Zool. 107, 315-96.
{Manuscript received 24 May 1967, revised 20 July 1967)